
Submitted:
05 May 2023
Posted:
08 May 2023
You are already at the latest version
Abstract
Feline infectious peritonitis (FIP), caused by the feline coronavirus (FCoV), is a devastating dis-ease in cats. Based on its antigenicity, FCoV can be divided into two serotypes: FCoV-I and FCoV-II. Furthermore, according to its pathogenicity, FCoV can be divided into feline enteric coronavirus (FECV) and feline infectious peritonitis virus (FIPV). There are numerous factors that affect the pathogenesis of FIP, among which host immunity and viral genetics can play an es-sential role in the development of FIP. Owing to the lack of specific symptoms, existing individual diagnostic methods can only support the suspicion of FIP, and multiple diagnostic methods and test data need to be combined to make a diagnosis. Although there are still no effective FIPV vaccines or commercial drugs available in the market, various studies have shown that some compounds can be used for treatment. Therefore, FIP is no longer incurable in cats
Keywords:
FIP
; FCoV
; genotype
; pathogenesis
; diagnosis
; treatment
; vaccine
1. Introduction
Feline infectious peritonitis (FIP) is a coronaviral disease with an extremely high mortality rate caused by a genetic variant of feline coronavirus (FCoV), referred to as the FIP virus (FIPV) [1]. This fatal disease can affect cats of any age but is most prevalent among cats under 3 years of age, especially those between the ages of 4 to 16 months. FIP often occurs in domestic cats, but in recent years, it has also been reported in non-domestic felids [2], including Panthera leo, Puma concolor, Panthera tigers, Leptailurus serval, and Panthera onca [3]. Infected cats with clinical signs of FIPV typically have an extremely high mortality rate, although some cats can live with the disease for varying periods ranging from weeks to months or even years [4].
2. Serotype study of the etiologic agent of FIP
Based on their antigenicity, FCoVs are generally divided into two serotypes: FCoV-I and FCoV-II. These two serotypes are classified according to humoral response changes caused by significant sequence differences between spike proteins [5]. Studies have shown that FCoV-II emerged through independent recombination events with canine coronavirus (CCV) [6]. Interestingly, serological studies have suggested that FCoV-II is more closely related to CCV than to FCoV-I [6] (Figure 1), but comparative sequence analysis of the ORF7a-ORF7b region revealed that type I and II FCoVs are more closely related to each other than to CCV [7]. Recombination of CoVs may generate new variants that can infect species other than their natural hosts. Thus, recombination is a common behavior of CoVs owing to their low-fidelity polymerase and unique replication selection, which will accelerate the variation speed of FCoV and increase the risk of cross-species transmission [8,9]. In terms of natural infections worldwide, FCoV-I is the most widespread in cats, whereas FCoV-II infections are relatively uncommon [10,11,12] (Figure 1). However, there have been relatively few studies on FCoV-I owing to the difficulty in culturing and isolating the virus in cells. FCoV-I is difficult to isolate in cell culture because it only grows well in macrophage [13]. Cultivation technology for FCoV-II is relatively mature. Studies have shown that FCoV-II strains adapt well to most available cell lines such as CRFK, F81, and FCWF-4 during routine operation [14,15,16,17]. Therefore, currently, most pathogenicity studies on FCoV focus on FCoV-II.
FCoV is sub-classified into two biotypes based on its pathogenicity: FECV and FIPV, and, FECV and FIPV include both serotypes I and II. FECV infection is usually asymptomatic or causes mild transient enteritis, whereas FIP is fatal if not treated timely [18]. As factors that increase the affinity of FIPV to macrophages seem to limit fecal-oral transmission, FIPV is generally considered non-transferable. It is reported that the pressure of overcrowding and the high level of virus in the environment may be conducive to the transition from FECV to FIPV [19]. Moreover, some studies have found that some FCoV strains may be more prone to this transformation than other strains [20,21,22]. The widely accepted “in vivo mutation” theory states that FIPV is caused by a mutation of FECV in the gastrointestinal tract of infected cats, which is transmitted systematically and leads to FIP [13,22,23]. The mutation sites responsible for FIP are not fully understood; however, it has been reported that some accessory genes (3c and 7b) may be the sites of critical mutations [7,24]. Another hypothesis is the "circulating virulent/avirulent" theory, which shows that both virulent and avirulent strains circulate in the cat population, and susceptible individuals exposed to the virulent strains are susceptible to the disease. Phylogenetic analyses have shown that M and 7b genes in viruses obtained from healthy cats were distinct from those obtained from sick cats and suggest the coexistence of both biotypes in cats. This hypothesis was proposed to explain the susceptibility of individuals exposed to virulent strains to the disease. [25]. Hitherto, none of the identified mutations appear to be in accordance with the pathotype from less virulent FCoV to FIP-associated FCoV [26,27,28]. Interventional studies involving animals or humans and other studies that require ethical approval must list the authority that provided the approval and the corresponding ethical approval code.
According to the clinical symptoms, FIP can be divided into two pathological forms: dry and wet. In the “wet” form, the most characteristic sign is a considerable amount of intracavitary effusion [29]. For a long time, the two forms were considered distinct viral species, but current molecular studies have demonstrated that they are two variants of the same virus with different virulence [30]. The clinical symptoms of the dry form mainly focus on the eyes and nervous system, and the typical characteristic of granulomatous lesions of dry FIP is the Focal accumulation of inflammatory cells and necrotic-proliferative lesions [31]. Studies have shown that in the terminal stages of the disease, when the immune system collapses, the dry form of FIP may become effusive [32,33]. One of the main differential diagnoses of ocular FIP is idiopathic uveitis [34]. In both clinical forms, systemic symptoms caused by severe acute systemic inflammatory reaction syndrome (SIRS), such as fever, fatigue, and emaciation, and laboratory changes, such as anemia, elevated concentrations of serum globulin components, and acute phase proteins, are commonly found [24]. A sign of FIP is disseminated vasculitis caused by complex immune reactions among viruses, antiviral antibodies, and complement [35].
Interestingly, the classification method by which FCoV can be divided into types I and II has recently been questioned, and based on the differences in spike proteins and the expected biological and clinical results, it has been suggested that these two "serotypes" should be considered as different viruses [36]. Regardless of the definition of the two serotypes, both can cause FIP and FECV [37]. Furthermore, FIP has been found to be the primary cause of death of kittens [38].
3. Pathogenesis
As a member of the genus Alphacoronavirus, FCoV is an enveloped, positive-sense, single-stranded, highly contagious virus [39,40]. The FCoV genome is as large as that of other coronaviruses, approximately 29 kb, encoding 11 proteins and four structural proteins, namely spike (S), matrix (M), nucleocapsid (N), and envelope (E), and five non-structural proteins, namely replicase 1a and 1b multi-proteins, as well as accessory proteins-3a, 3b, 3c, 7a, and 7b [41,42,43] (Figure 2). In addition to protein S, studies have revealed that the 7a-b ORFs and 3a-c are the main candidates for mutations that lead to the FIP phenotype [34,42]. FCoV-infected cats without FIP may also develop viremia, and FCoV can even be detected in the organs of healthy cats [44,45,46,47]. This coronavirus is pervasive in cats worldwide, and when it is an FECV, it is not in itself an important pathogen that causes mild and transient diarrhea [48,49]. However, once FIPV arises from specific mutations in FECV, it is devastating to cats [24,50]. There are rare occasions when a cat may recover miraculously but still relapse months or even years later [51].
The most critical step in the viral life cycle is viral entry into cells. In FIPV, viral cell entry occurs in two ways. The first pathway is cytosolic entry through early endosomes (FCoV-II) and the other is through late endosomes (FCoV-I) [52]. Studies have revealed that the cell receptor of FCoV-II is aminopeptidase N (APN), which binds to the spike (S) protein and mediates the internalization of the virus into the cell [53,54]. However, there have been no relevant studies on FCoV-I cell receptors. Studies showing the entry mechanisms of FCoV-II have been conducted using the 79-1146 strain, which is internalized through a novel clathrin- and caveolae-independent pathway that depends on dynamin [55,56].
FCoV binds to its cellular receptors and requires cytoplasmic access for replication. The cleaved S protein by cathepsin B fuses with the endosomal membrane after the complex between the viral receptor and viral S protein is endocytosed [57]. The S protein fusion peptide is located in the S2 domain. FCoV-I has two specific activation sites: S1/S2 cleaved by furin-like protease, and S2’ cleaved by cathepsin B. FCoV-II only has the S2’ site, cleaved by cathepsin B [57]. Cathepsin B may be the most important protease mediating FCoV entry, with cathepsin L possibly playing a secondary role [58]. Replication of FCoV is rapid, with the replicative cycle completed in less than 24 h. Notably, mutations in the FCoV S gene have been suggested to play an important role in the switch between cell tropism and pathogenicity [59,60,61]. It has been reported that only in the FCoV of cats with FIP, two single-nucleotide polymorphisms (SNP) in close proximity were found at nucleotide positions 23,531 and 23,537 of the S gene, which could lead to amino acid substitutions at positions 1,058 (M1,058L) and 1,060 (S1,060A), respectively, within the spike protein. Furthermore, a study found that one of the aforementioned SNPs was present in 96% of the FCoV S genes in cats with FIP [26]. Although this amino acid substitution can also be detected in tissues from cats without FIP, another study revealed that it may be more likely related to the systemic spread of FCoV rather than to the FIP phenotype [62]. Consequently, the role of these S gene mutations in FIP pathogenesis remains unclear.
After oral fecal transmission, FCoV experiences an initial intestinal infection, and the virus can spread outside the intestinal tract, leading to monocyte-related viremia, which can mutate to FIPV [25]. Productive monocyte/macrophage infections caused by FIPV, anatomical dissemination with diverse distributions, and immune-mediated perivasculitis lead to FIP [63]. In addition, due to the spread of virus in various parts of the body, FIP may exhibit clinical symptoms reflecting inflammation of various anatomical sites, including the abdominal cavity and related organs, thoracic cavity, central nervous system and/or eyes [34,64,65,66].
Feces are the main source of FCoV, and cat litter boxes and litter trays are the main sources of oral infection in cats [67,68] (Figure 3). In contrast, transplacental transmission through a shared feeding bowl plays a minor role [69,70]. Moreover, it is revealed that the wet type of FIPV may be the result of weak cell-mediated immune response [71].
To date, data regarding the risk factors for FIP remain controversial. Several studies have shown that breed and sex are not associated with FIP [72,73], whereas other studies have shown that the male sex is significantly correlated with FIP [74,75]. Additionally, there is evidence that certain breeds are more susceptible to FIP [76,77,78,79].
4. Innate Immune Response and Immune Escape Mechanisms
It has been reported that the viral load and the cat’s immune response determine whether FIP will develop, and both host immunity and viral genetics can play significant roles in the development of FIP [63,80]. The progression of FIP infection may be a consequence of severe immunosuppression caused by T cell depletion [81]. Several studies have reported on the role of antibodies in the pathogenesis of FIP. After infection with FIPV, the immune complex formed by FCoV and the anti-FCoV antibody precipitates around the blood vessel wall and induces the recruitment of macrophage production factors. The resulting cytokines damage the tissue, induce the release of neutrophils, exacerbate the inflammatory process, and cause tissue damage [82,83], resulting in the occurrence of the cavity effusion observed in wet FIP.
Feline Toll-Like Receptors (TLR) 1-9 have been isolated and exhibit varying distributions, expression, and functions in different tissues, among which TLR3 has been found to effectively prevent the invasion of FIPV [84]. Compared to the mesenteric lymph nodes of healthy cats, the transcription level of TLR3 in mesenteric lymph nodes showing typical FIP lesions was slightly lower, suggesting a general systemic stimulus to upregulate TLR3 in FIP. Moreover, in vitro studies have shown that prior stimulation of TLR3 contributes to defense against murine coronaviruses [85].
During viral infection, interferons (IFNs) are the main cytokines involved in the antiviral response [86] and are key elements in both innate and adaptive immune responses to viral infections [87]. However, there is much evidence that coronaviruses can escape the host IFN response via accessory proteins [88,89]. Recently an accessory protein 7a able to counteract IFN-α-induced antiviral response, has been identified in FIPV [90,91]. Before infection, the presence of protein 7a did not reduce the IFN produced by FIPV, indicating that protein 7a may resist downstream virus-producing IFN. Furthermore, it has been found that only in the presence of proteins encoded by ORF3 (3a or 3b), protein 7a could interfere with the IFN-α antiviral response [91]. The synergism between these two proteins allows the effective replication of wild-type FlPV, which may produce the same antiviral effect by interfering with different IFN-induced pathways. Thus, blocking the two IFN-induced pathways may be the key to overcoming the overall negative effects of IFN [88]. However, studies have found that the deletion of the ORF from wild-type FIPV does not make the virus completely sensitive to IFN, indicating that other viral proteins (Nsps and/or structural proteins) may also play a role in IFN antagonism [88,92]. Nsp5 of FIPV has recently been shown to inhibit the production of type I IFN by cutting multiple NEMO sites [90].
Natural Killer (NK) cells are specific cytotoxic lymphocytes lacking an antigen-specific receptor that can kill virus-infected cells as well as tumor- and stressed- cells [93]. Regulatory T cells (Tregs) is a CD4+ T cell group that mainly regulates the immune response caused by immunosuppression [34]. Previous research has shown that NK cells and Tregs in the peripheral blood, mesenteric lymph nodes, and spleen of FIP-infected cats were significantly reduced compared with uninfected control cats, which indicates that FIP infection might be associated with severe depletion of both NK cells and Tregs and the reduction of NK cell function [94].
5. Diagnostic Methods
Ante-mortem diagnosis of FIP is difficult and frustrating. Difficulties in definitively diagnosing FIP are due to the lack of specific clinical signs, pathognomonic biochemical abnormalities, and the low sensitivity and specificity of tests routinely used in practice [29]. The common clinical signs of FIPV, such as anorexia, lethargy, and weight loss, and neurological signs, such as gait abnormalities or abnormal mentation, are non-specific [95,96,97]. The diagnosis of FIP is usually made by weighing the clinical findings, abnormalities present in common diagnostic procedures, and, if possible, postmortem examination and histopathology weigh [29]. Currently, there is no perfect method for diagnosing FIPV. It is well known that lymphopaenia and neutrophilia are typical manifestations of FIP, but this change can be explained as a typical “stress leukogram” that occurs in many severe systemic diseases in cats [98]. Although the immunological staining of viral antigens within tissue lesions is considered the reference standard for diagnosing FIP, invasive sampling is inconvenient [29,99,100].
5.1. Reverse Transcriptase Polymerase Chain Reaction (RT-PCR) Assays
There have been many reports on the detection of FCoV using RT-PCR. RT-PCR assays may be valuable tools for screening viruses in cats because most FCoV strains can be detected [101,102,103,104,105]. Owing to differences in the S gene sequence between FCoV serotypes I and II, certain RT-PCR assays have been developed to distinguish between these serotypes based on the S gene [11,31]. In addition, some used primers targeting the other conserved regions of the virus genome, such as 7b gene [106,107,108], and 3’ untranslated regions (3’ UTRs) [109,110]. However, because viral RNA also circulates within asymptomatic FCoV-infected cats without FIP [44,111,112], RT-PCR detection of FCoV is only partially useful [109,111,113,114]. Negative FCoV RT-PCR results are common in cats with FIP. Therefore, RT-PCR is not the only detection method for diagnosing FIP, and its results should be interpreted in combination with the clinical status of infected cats. However, because cats with FIP usually have significantly higher FCoV RNA loads, as measured by quantitative RT-PCR, than cats without FIP [62,101,115,116,117,118], quantification of FCoV RNA loads is a reasonable option to narrow the diagnosis of FIP [67].
5.2. Histopathology and Immunohistochemistry
Currently, the only accurate diagnostic method is immunohistochemical (IHC) staining of FCoV antigens in macrophages within tissues with characteristic changes [29,99,100]. Although immunostaining cannot differentiate between FECV and FIPV, FIPV replicates more actively, and higher concentrations of viral antigens are found in FIP cases [35]. Viral antigen concentrations are lower in lesions of cats with dry FIP than in those with wet FIP [119].
5.3. Serology
Hematological abnormalities are common and nonspecific in cats with FIP [105]. It should be noted that lymphopenia observed in approximately 50% of cats with FIP is significantly more common in cats with effusion, but rarely reported in cats without effusion [105]. Most cats with FIP also have abnormal serum biochemistry, particularly hyperglobulinemia, which has been reported in 80% of cats with FIP [74]. Microcytosis and band neutrophilia are the common features of FIP. Cats with FIP usually show a significant increase in the concentration of acute phase protein (APP), however, they are not specific symptoms of FIP [95,120].
The measurement of serum antibodies is a useful and relatively accurate diagnostic tool for the detection of FCoV infection. However, there is still controversy regarding the use and interpretation of anti-FCoV antibody titers in serum or plasma [1]. Therefore, as a large proportion of healthy cats have anti-FCoV antibodies, antibody detection is more helpful for the management of FCoV infections than for the diagnosis of FIP. Furthermore, some cats with the wet form of FIP have low titers or even no anti-FCoV antibodies, which may be due to the large amounts of virus in the cat’s body that bind to antibodies and render them unavailable to antigens in the test or because the antibodies are lost in effusions [35].
A study reported that sensitivity and specificity of a commercial test kit for the detection of FCoV antibodies (ImmunoComb FCoV Antibody Test Kit, Biogal, Israel) were 95% and 83%, respectively [121]. Moreover, the use of anti-7b protein in serology does not seem to provide any significant advantage, which was developed on the faulty premise that FECVs lack the ORF 7b gene and therefore do not produce the ORF7b protein product, while FIPVs have the ORF7b gene, produce the protein, and therefore trigger an antibody response to the ORF7b protein [106]. Now it is well known that almost all field isolates of FIPV and FECV have complete ORF7b gene, so both can trigger anti-ORF7b antibodies.
5.4. Tests on Effusion Fluid
Compared to serological tests, the hydrops test has a higher diagnostic value in clinical tests of effusions. However, the presence of fluid accumulation in the clinic did not indicate FIPV infection. Approximately half of the cats with fluid accumulation have FIP [122]. The cytology of the effusion in cats with FIP usually consists predominantly of macrophages and neutrophils, and the protein content is very high (> 35 g/l), while the cellular content is low (< 5,000 nucleated cells/ml) and approaches that of a modified or pure transudate. Electrophoresis is a useful tool to assist in the diagnosis by determining the albumin/globulin ratio. If the albumin/globulin ratio was < 0.4, it indicated a high positive predictive value; if the ratio was > 0.8, it indicated a high negative predictive value [67]. Moreover, Rivalta’s test is useful in differentiating between FIP effusions and effusions caused by other diseases, with a positive predictive value of 86% [99].
5.5. Other Diagnosis Method
Ultrasonography, magnetic resonance imaging (MRI) and radiology are very helpful auxiliary tools for identifying the presence of fluid caused by FIPV [123]. In some cats, mesenteric lymphadenopathy can result in abdominal lesions. Radiographs can reveal pleural, pericardial, or peritoneal effusions, hepatomegaly, or renomegaly. Ultrasonography can be used to confirm the presence of abdominal fluid in cats with minimal fluid volume and to evaluate the pancreas, liver, lymph nodes, and kidneys. MRI can reveal periventricular contrast enhancement, ventricular dilation, and hydrocephalus in cats with neurological FIP [124].
6. Therapy and Prevention
As FIP is very common in cats and can cause extremely high mortality, there is an increasing demand for therapies and prevention from veterinarians and cat owners. Although no treatment has been approved at present, several studies have been conducted to develop effective, safe, and mass-produced therapeutic compounds [125].
6.1. Antivirals
In recent years, the nucleotide analog GS-441524 and the 3C-like protease inhibitor GC376 have been shown to have good efficacy in the treatment of FIPV, although the two drugs have completely different effects [126,127,128,129,130].
As a small-molecule competitive inhibitor of nucleoside triphosphates (NTPs) and developed from the precursor drugs GS-5734 and GS-5734, GS-441524 has been reported to effectively prevent the experimental Ebola virus in rhesus monkeys and inhibit Middle East Respiratory Syndrome in a mouse infection model [131]. GS-441524 is required for intracellular phosphorylation of cell kinases to nucleoside monophosphate, which is then phosphorylated to the active triphosphate metabolite (NTP). The active NTP analog acts as a competitor to the natural nucleoside triphosphate in viral RNA synthesis; when it is inserted into a transcription product, transcription is terminated prematurely, inhibiting the viral RNA transcription process.[128]. GS-441524 acts primarily during the early stages of viral replication.
Based on this point, Pedersen et al. revealed the efficacy of GS-441524 administered for 12 weeks in the treatment of 32 cats naturally infected with FCoV [127]; Peter et al. showed that GS-441524 can be used to treat FIP with neurological presentation with similar efficacy, and determined the optimal dose range for GS-441524 in cats with neurological FIP (5–10 mg/kg) [132]. In addition, treating cats with dry or neurological forms of FIP may require higher doses than those for the wet form because the blood-ocular and blood-brain barriers inhibit the diffusion of GS-441524 [127,132]. It has been reported that GS-441524 shows effective oral bioavailability [133,134] and can be used as an oral antiviral drug against coronaviruses.
The 3C-like protease inhibitor (3CLpro) binds to the active site of the viral 3C protease, blocking its catalytic activity, and thus inhibiting coronavirus replication in vivo [130]. More accurately, its breakdown product, GC373, effectively inhibits Mpro of FCoV. Kim et al. revealed that GC376 effectively inhibits coronavirus replication in naturally infected cats [135]. Pedersen et al. further showed that when GC376 is used for 12–17 weeks, it is safe and effective for the treatment of cats with various forms of natural clinical FCoV infection [126]. Other studies, including those on ferret and mink coronavirus Mpro, demonstrated the broad specificity of this protease inhibitor [135,136]. However, GC376 appears to be less effective in treating and eradicating the more chronic neurological or ocular forms of FIPV [137].
Chang et al. showed that GS-441524 was more effective than GC376 in treating cats with FIP. The remission rate with GC376 was approximately 50%, whereas that of GS-441524 was 70% [138]. However, regardless of GC376 or GS-441524 is treated as independent drugs for FIPV, they must be administered for a long time (12-17 weeks). Thus, Lv et al. tested a combination of the two drugs and found that GS-441524 combined with GC376 can be safely and effectively used to treat FIP and reduce the treatment period to 4 weeks, with an excellent cure rate [139]. However, Cook et al. found that, although evidence of compound synergy was identified for several combinations of antiviral agents, monotherapies were ultimately determined to be the most effective in inhibiting viral transcription [140].
In addition to the two most common drugs, other drugs have been discovered in recent years and have the potential to be used for the treatment of FIPV [140]. Studies suggest that ICZ strongly inhibited type I FIPV replication [141,142], by inhibiting pathways downstream of the late endosome [52]. Regan et al. found that cathepsin B inhibitors might be effective therapeutics to treat incurable and lethal afflictions in cats [58]. Tanaka et al. showed that ionophore antibiotics, including valinomycin, salinomycin, and nigericin, can act as broad-spectrum agents that inhibit FCoV proliferation in vitro in a dose-dependent manner by strongly binding to cellular potassium ions and altering the ion concentrations and pH within the cell [143,144]. It has reported that salinomycin inhibited FCoV proliferation in Fcwf-4 cell, especially exhibited its antiviral activities at a concentration of 10 µM. Therefore, PPIase inhibitors may act as alternative drugs for treating FIP [145]. Though the effect of recombinant feline IFN-ω (rfeIFN-ω) has not been reliably confirmed in FIP, topical administration of IFN-ω still has the potential for the treatment of FIP [16,99,146].
Overall, the most effective antiviral compounds were determined to mechanistically feature either a nucleoside analog or a protease inhibitor, but other antiviral compounds can also play a role in the adjuvant therapy of FIP. Despite recent clinical success, these antiviral compounds are currently unavailable for legal veterinary use in cats with FIP.
6.2. Vaccine
As a highly virulent monocyte/macrophage pathogen, FIPV variant could cause systemic immune complex mediated infection [130,147]. Thus, owing to the role of antibody-dependent enhancement (ADE) in FIP disease pathogenesis, the development of an effective vaccine against FIP has been challenging. Several studies have shown that the presence of non-neutralizing anti-coronavirus antibodies aggravates FIP disease [137,148,149].
A modified live intranasal vaccine (Felocell FIP, Zoetis), containing a temperature-sensitive mutant of the FCoV strain DF2-FIPV, can only replicate in the upper respiratory tract; therefore, it can induce a local IgA reaction in the mucosa, which has been licensed in the United States [150]. However, further studies have shown that its efficacy remains uncertain [151]. This type of vaccine is only effective if inoculated before contact with FIPV, which is not suitable for highly endemic situations [152]. Therefore, this vaccine has significant limitations, and is not recommended.
Attempts have also been made to immunize cats with an unattenuated field isolate of canine coronavirus to prevent FIP; however, these have not been successful [153]. To date, the most promising results were that in recombinant FIPV mutants lacking the ORF3abc or ORF7ab region, after lethal homologous attack, these mutants provided 100% and 80% protection, respectively [154]. Interestingly, recombinant feline coronaviruses, which differ only in the truncation (PBFIPV-DF-2) or completion (PBFIPV-DF-2-R3i) of their ORF3abc regions, are vaccine candidates that can play a protective role in SPF cats, but not in conventional cats [155].
7. Conclusions
Regardless of the disease to be treated, a study of its etiology and pathogenesis is necessary. The research on FCoV, the pathogen that causes FIP, has been going on for decades, but just the classification methods for FCoV are still not clear enough so far. And the receptor of FCoV-I has not yet been determined, which is also related to the difficulty in cultivating FCoV-I. Thus, basic research on FCoV is still in progress, and perhaps the train of thought of FCoV can refer to that of COVID-19.
Owing to some cats with FIP still look healthy at the time of diagnosis and can usually live for several days with only symptomatic treatment, their owners might not be willing to accept the diagnosis and the fact that there is no effective treatment, which has a negative impact on communication between veterinarians and owners [34]. Timely diagnosis of FIP remains challenging. Once the typical symptoms of FIP appear, cat treatment is likely to be ineffective. Therefore, regular physical checkups of cats and stress reduction are important to prevent FIP. Although commercial drugs for FIPV have not yet appeared in the market, there are certain effective drugs available for treatment. FIP is no longer an absolutely incurable disease in cats. However, the price of these drugs is high and the treatment may last for several months, which is a heavy burden for the owner. Further research is necessary to develop effective prevention and treatment methods. But all in all, what is certain is that FIP is no longer an incurable disease, the efforts made for the study of FIP brings us a ray of hope to defeat this devastating disease of cat.
Author Contributions
Conceptualization, S.L. and J.L.; methodology, Y.C.; software, S.L.; validation, S.L.; investigation, L.H., J.Z., and X.F. data curation, writing—original draft preparation, Y.C.; writing—review and editing, S.L. and J.L.; visualization, S.L.; supervision, J.L.; project administration, J.L.; funding acquisition, J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pedersen, N.C.; Liu, H.; Dodd, K.A.; Pesavento, P.A. Significance of Coronavirus Mutants in Feces and Diseased Tissues of Cats Suffering from Feline Infectious Peritonitis. Viruses 2009, 1, 166–184, . [CrossRef]
- Ratti, G.; Stranieri, A.; Giordano, A.; Oltolina, M.; Bonacina, E.; Magnone, W.; Morici, M.; Ravasio, G.; Paltrinieri, S.; Lauzi, S. Molecular Detection of Feline Coronavirus in Captive Non-Domestic Felids from Zoological Facilities. Animals 2022, 12, 1864, . [CrossRef]
- Stout, A.E.; André, N.M.; Whittaker, G.R. Feline coronavirus and feline infectious peritonitis in nondomestic felid species. J. Zoo Wildl. Med. 2021, 52, 14–27, . [CrossRef]
- Drechsler, Y.; Alcaraz, A.; Bossong, F.J.; Collisson, E.W.; Diniz, P.P.V. Feline Coronavirus in Multicat Environments. Veter- Clin. North Am. Small Anim. Pr. 2011, 41, 1133–1169, . [CrossRef]
- A Fiscus, S.; A Teramoto, Y. Antigenic comparison of feline coronavirus isolates: evidence for markedly different peplomer glycoproteins. J. Virol. 1987, 61, 2607–2613, . [CrossRef]
- Herrewegh AA, Smeenk I, Horzinek MC, Rottier PJ, de Groot RJ. Feline coronavirus type II strains 79-1683 and 79-1146 orig inate from a double recombination between feline coronavirus type I and canine coronavirus. J. Virol. 1998, 72(5), 4508-4514.
- Herrewegh AA, Vennema H, Horzinek MC, Rottier PJ, de Groot RJ. The molecular genetics of feline coronaviruses: compar- ative sequence analysis of the ORF7a/7b transcription unit of different biotypes. Virology 1995, 212(2), 6226-6231.
- Holmes, E.C.; Rambaut, A. Viral evolution and the emergence of SARS coronavirus. Philos. Trans. R. Soc. Lond B: Biol. Sci. 2004, 359, 1059–1065, . [CrossRef]
- Smith EC, Denison MR. Denison. Implications of altered replication fidelity on the evolution and pathogenesis of coronaviruses. Curr Opin Virol, 2012, 2(5), 519-524.
- Gao YY, Liang XY, Wang Q, Zhang S, Zhao H, Wang K, Hu GX, Liu WJ, Gao FS. Mind the feline coronavirus: comparison with SARS-CoV-2. Gene, 2022, 825, 146443.
- Addie, D.D.; Schaap, I.A.T.; Nicolson, L.; Jarrett, O. Persistence and transmission of natural type I feline coronavirus infection. J. Gen. Virol. 2003, 84, 2735–2744, . [CrossRef]
- Kummrow, M.; Meli, M.L.; Haessig, M.; Goenczi, E.; Poland, A.; Pedersen, N.C.; Hofmann-Lehmann, R.; Lutz, H. Feline Coronavirus Serotypes 1 and 2: Seroprevalence and Association with Disease in Switzerland. Clin Diagn Lab Immunol 2005, 12, 1209–1215, . [CrossRef]
- Pedersen, N.C.; Boyle, J.F.; Floyd, K.; Fudge, A.; Barker, J. An enteric coronavirus infection of cats and its relationship to feline infectious peritonitis. Am J Vet Res. 1981, 42, 368–377.
- Pedersen, N.C.; Boyle, J.F.; Floyd, K. Infection studies in kittens, using feline infectious peritonitis virus propagated in cell culture. Am J Vet Res. 1981, 42, 363–7.
- Black, J.W. Recovery and in vitro cultivation of a coronavirus from laboratory-induced cases of feline infectious peritonitis (FIP). Vet Med Small Anim Clin. 1980, 75, 811–4.
- Ritz S, Egberink H, Hartmann K. Effect of feline interferon-omega on the survival time and quality of life of cats with feline infectious peritonitis. J Vet Intern Med, 2007, 21(6), 1193-1197.
- McKeirnan, A.J.; Evermann, J.F.; Davis, E.V.; Ott, R.L. Comparative properties of feline coronaviruses in vitro. Can J Vet Res,. 1987, 51, 212–216.
- Jähne, S.; Felten, S.; Bergmann, M.; Erber, K.; Matiasek, K.; Meli, M.L.; Hofmann-Lehmann, R.; Hartmann, K. Detection of Feline Coronavirus Variants in Cats without Feline Infectious Peritonitis. Viruses 2022, 14, 1671, . [CrossRef]
- Pedersen, N.C.; Liu, H.; Scarlett, J.; Leutenegger, C.M.; Golovko, L.; Kennedy, H.; Kamal, F.M. Feline infectious peritonitis: Role of the feline coronavirus 3c gene in intestinal tropism and pathogenicity based upon isolates from resident and adopted shelter cats. Virus Res. 2012, 165, 17–28, . [CrossRef]
- Brown, M.A. Genetic determinants of pathogenesis by feline infectious peritonitis virus. Veter- Immunol. Immunopathol. 2011, 143, 265–268, . [CrossRef]
- Healey EA, Andre NM, Miller AD, Whittaker GR, Berliner EA. Outbreak of feline infectious peritonitis (FIP) in shelter-housed cats: molecular analysis of the feline coronavirus S1/S2 cleavage site consistent with a ’circulating virulent-avirulent theory’ of FIP pathogenesis. JFMS Open Rep, 2022, 8(1), 20551169221074226.
- Vennema, H.; Polanda, A.; Foleya, J.; Pedersen, N.C. Feline Infectious Peritonitis Viruses Arise by Mutation from Endemic Feline Enteric Coronaviruses. Virology 1998, 243, 150–157, . [CrossRef]
- Poland, A.M.; Vennema, H.; E Foley, J.; Pedersen, N.C. Two related strains of feline infectious peritonitis virus isolated from immunocompromised cats infected with a feline enteric coronavirus. J. Clin. Microbiol. 1996, 34, 3180–3184, . [CrossRef]
- Pedersen, N.C. A review of feline infectious peritonitis virus infection: 1963–2008. J. Feline Med. Surg. 2009, 11, 225–258, . [CrossRef]
- Brown, M.A.; Troyer, J.L.; Pecon-Slattery, J.; Roelke, M.E.; O’brien, S.J. Genetics and Pathogenesis of Feline Infectious Peritonitis Virus. Emerg. Infect. Dis. 2009, 15, 1445–1452, . [CrossRef]
- Chang, H.-W.; Egberink, H.F.; Halpin, R.; Spiro, D.J.; Rottier, P.J. Spike Protein Fusion Peptide and Feline Coronavirus Virulence. Emerg. Infect. Dis. 2012, 18, 1089–1095, . [CrossRef]
- Bank-Wolf, B.R.; Stallkamp, I.; Wiese, S.; Moritz, A.; Tekes, G.; Thiel, H.-J. Mutations of 3c and spike protein genes correlate with the occurrence of feline infectious peritonitis. Veter- Microbiol. 2014, 173, 177–188, . [CrossRef]
- Borschensky, C.; Reinacher, M. Mutations in the 3c and 7b genes of feline coronavirus in spontaneously affected FIP cats. Res. Veter- Sci. 2014, 97, 333–340, . [CrossRef]
- Sharif, S.; Arshad, S.S.; Hair-Bejo, M.; Omar, A.R.; Zeenathul, N.A.; Alazawy, A. Diagnostic Methods for Feline Coronavirus: A Review. Veter- Med. Int. 2010, 2010, 809480, . [CrossRef]
- Pedersen, N.C. An update on feline infectious peritonitis: Virology and immunopathogenesis. Veter- J. 2014, 201, 123–132, . [CrossRef]
- Benetka, V.; Kübber-Heiss, A.; Kolodziejek, J.; Nowotny, N.; Hofmann-Parisot, M.; Möstl, K. Prevalence of feline coronavirus types I and II in cats with histopathologically verified feline infectious peritonitis. Veter- Microbiol. 2004, 99, 31–42, . [CrossRef]
- Patel JR, Heldens JG. Review of companion animal viral diseases and immunoprophylaxis. Vaccine, 2009, 27(4), 491-504.
- Goodson T, Randell S, Moore L. Feline infectious peritonitis. Compend Contin Educ Vet. 2009, 31(10), E1-8.
- Pedersen, N.C. An update on feline infectious peritonitis: Diagnostics and therapeutics. Veter- J. 2014, 201, 133–141, . [CrossRef]
- Hartmann K. Feline infectious peritonitis. Vet Clin North Am Small Anim Pract. 2005, 35(1), 39-79.
- Jaimes, J.A.; Millet, J.K.; Stout, A.E.; André, N.M.; Whittaker, G.R. A Tale of Two Viruses: The Distinct Spike Glycoproteins of Feline Coronaviruses. Viruses 2020, 12, 83, . [CrossRef]
- Hohdatsu, T.; Okada, S.; Ishizuka, Y.; Yamada, H.; Koyama, H. The Prevalence of Types I and II Feline Coronavirus Infections in Cats.. J. Veter- Med Sci. 1992, 54, 557–562, . [CrossRef]
- Fehr, D.; Bolla, S.; A Herrewegh, A.; Horzinek, M.C.; Lutz, H. [Detection of feline coronavirus using RT-PCR: basis for the study of the pathogenesis of feline infectious peritonitis (FIP)]. Schweiz Arch Tierheilkd. 1996, 138, 74–9.
- Decaro N, Lorusso A. Novel human coronavirus (SARS-CoV-2): A lesson from animal coronaviruses. Vet Microbiol. 2020, 244, 108693.
- Gatherer, D.; Depledge, D.P.; Hartley, C.A.; Szpara, M.L.; Vaz, P.K.; Benkő, M.; Brandt, C.R.; Bryant, N.A.; Dastjerdi, A.; Doszpoly, A.; et al. ICTV Virus Taxonomy Profile: Herpesviridae 2021. J. Gen. Virol. 2021, 102, 001673, . [CrossRef]
- Tekes G, Thiel HJ. Feline coronaviruses: pathogenesis of feline infectious peritonitis. Adv Virus Res. 2016, 96, 193-218.
- Kennedy MA. Feline infectious peritonitis: update on pathogenesis, diagnostics, and treatment. Vet Clin North Am Small Anim Pract, 2020, 50(5), 1001-1011.
- Dye C, Siddell SG. Genomic RNA sequence of Feline coronavirus strain FIPV WSU-79/1146. J Gen Virol. 2005, 86(Pt 8), 2249-2253.
- Kipar, A.; Meli, M.L.; Baptiste, K.E.; Bowker, L.J.; Lutz, H. Sites of feline coronavirus persistence in healthy cats. J. Gen. Virol. 2010, 91, 1698–1707, . [CrossRef]
- Desmarets, L.M.B.; Vermeulen, B.L.; Theuns, S.; Conceição-Neto, N.; Zeller, M.; Roukaerts, I.D.M.; Acar, D.D.; Olyslaegers, D.A.J.; Van Ranst, M.; Matthijnssens, J.; et al. Experimental feline enteric coronavirus infection reveals an aberrant infection pattern and shedding of mutants with impaired infectivity in enterocyte cultures. Sci. Rep. 2016, 6, 20022–20022, . [CrossRef]
- Meli, M.; Kipar, A.; Müller, C.; Jenal, K.; Gönczi, E.; Borel, N.; Gunn-Moore, D.; Chalmers, S.; Lin, F.; Reinacher, M.; et al. High viral loads despite absence of clinical and pathological findings in cats experimentally infected with feline coronavirus (FCoV) type I and in naturally FCoV-infected cats. J. Feline Med. Surg. 2004, 6, 69–81, . [CrossRef]
- Kipar, A.; Meli, M.L.; Failing, K.; Euler, T.; Gomes-Keller, M.A.; Schwartz, D.; Lutz, H.; Reinacher, M. Natural feline coronavirus infection: Differences in cytokine patterns in association with the outcome of infection. Veter- Immunol. Immunopathol. 2006, 112, 141–155, . [CrossRef]
- Pedersen, N.C.; Allen, C.E.; Lyons, L.A. Pathogenesis of feline enteric coronavirus infection. J. Feline Med. Surg. 2008, 10, 529–541, . [CrossRef]
- Vogel, L.; Van der Lubben, M.; Lintelo, E.G.T.; Bekker, C.P.; Geerts, T.; Schuijff, L.S.; Grinwis, G.C.; Egberink, H.F.; Rottier, P.J. Pathogenic characteristics of persistent feline enteric coronavirus infection in cats. Veter- Res. 2010, 41, 71–71, . [CrossRef]
- Pedersen, N.; Sato, R.; Foley, J.; Poland, A. Common virus infections in cats, before and after being placed in shelters, with emphasis on feline enteric coronavirus. J. Feline Med. Surg. 2004, 6, 83–88, . [CrossRef]
- Legendre, A.M.; Bartges, J.W. Effect of Polyprenyl Immunostimulant on the survival times of three cats with the dry form of feline infectious peritonitis. J. Feline Med. Surg. 2009, 11, 624–626, . [CrossRef]
- Takano, T.; Wakayama, Y.; Doki, T. Endocytic Pathway of Feline Coronavirus for Cell Entry: Differences in Serotype-Dependent Viral Entry Pathway. Pathogens 2019, 8, 300.40300. [CrossRef]
- Hohdatsu, T.; Izumiya, Y.; Yokoyama, Y.; Kida, K.; Koyama, H. Differences in virus receptor for type I and type II feline infectious peritonitis virus. Arch. Virol. 1998, 143, 839–850, . [CrossRef]
- Tekes, G.; Hofmann-Lehmann, R.; Bank-Wolf, B.; Maier, R.; Thiel, H.-J.; Thiel, V. Chimeric Feline Coronaviruses That Encode Type II Spike Protein on Type I Genetic Background Display Accelerated Viral Growth and Altered Receptor Usage. J. Virol. 2010, 84, 1326–1333, . [CrossRef]
- Van Hamme, E.; Dewerchin, H.L.; Cornelissen, E.; Verhasselt, B.; Nauwynck, H.J. Clathrin- and caveolae-independent entry of feline infectious peritonitis virus in monocytes depends on dynamin. J. Gen. Virol. 2008, 89, 2147–2156, . [CrossRef]
- Van Hamme, E.; Dewerchin, H.L.; Cornelissen, E.; Nauwynck, H.J. Attachment and internalization of feline infectious peritonitis virus in feline blood monocytes and Crandell feline kidney cells. J. Gen. Virol. 2007, 88, 2527–2532, . [CrossRef]
- Burkard C, Verheije MH, Wicht O, van Kasteren SI, van Kuppeveld FJ, Haagmans BL, Pelkmans L, Rottier PJ, Bosch BJ, de Haan CA. Coronavirus cell entry occurs through the endo-/lysosomal pathway in a proteolysis-dependent manner. PLoS Pathog. 2014, 10(11), e1004502.
- Regan, A.D.; Shraybman, R.; Cohen, R.D.; Whittaker, G.R. Differential role for low pH and cathepsin-mediated cleavage of the viral spike protein during entry of serotype II feline coronaviruses. Veter- Microbiol. 2008, 132, 235–248, . [CrossRef]
- Lewis, C.S.; Porter, E.; Matthews, D.; Kipar, A.; Tasker, S.; Helps, C.R.; Siddell, S.G. Genotyping coronaviruses associated with feline infectious peritonitis. J. Gen. Virol. 2015, 96, 1358–1368, . [CrossRef]
- Licitra, B.N.; Millet, J.K.; Regan, A.D.; Hamilton, B.S.; Rinaldi, V.D.; Duhamel, G.E.; Whittaker, G.R. Mutation in Spike Protein Cleavage Site and Pathogenesis of Feline Coronavirus. Emerg. Infect. Dis. 2013, 19, 1066–1073, . [CrossRef]
- Shirato, K.; Chang, H.-W.; Rottier, P.J. Differential susceptibility of macrophages to serotype II feline coronaviruses correlates with differences in the viral spike protein. Virus Res. 2018, 255, 14–23, . [CrossRef]
- Porter, E.; Tasker, S.; Day, M.J.; Harley, R.; Kipar, A.; Siddell, S.G.; Helps, C.R. Amino acid changes in the spike protein of feline coronavirus correlate with systemic spread of virus from the intestine and not with feline infectious peritonitis. Veter- Res. 2014, 45, 49–49, . [CrossRef]
- Rottier, P.J.M.; Nakamura, K.; Schellen, P.; Volders, H.; Haijema, B.J. Acquisition of Macrophage Tropism during the Pathogenesis of Feline Infectious Peritonitis Is Determined by Mutations in the Feline Coronavirus Spike Protein. J. Virol. 2005, 79, 14122–14130, . [CrossRef]
- Diaz, J.V.; Poma, R. Diagnosis and clinical signs of feline infectious peritonitis in the central nervous system.. 2009, 50, 1091–1093.
- Foley, J.E.; Lapointe, J.; Koblik, P.; Poland, A.; Pedersen, N.C. Diagnostic Features of Clinical Neurologic Feline Infectious Peritonitis. J. Veter- Intern. Med. 1998, 12, 415–423, . [CrossRef]
- Stiles, J. Ocular manifestations of feline viral diseases. Veter- J. 2014, 201, 166–173, . [CrossRef]
- Addie, D.; Belák, S.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; Lloret, A.; Lutz, H.; et al. Feline infectious peritonitis. ABCD guidelines on prevention and management. J. Feline Med. Surg. 2009, 11, 594–604, . [CrossRef]
- Foley, J.; Rand, C.; Leutenegger, C. Inflammation and changes in cytokine levels in neurological feline infectious peritonitis. J. Feline Med. Surg. 2003, 5, 313–322, . [CrossRef]
- Addie, D.D.; Jarrett, O. Use of a reverse-transcriptase polymerase chain reaction for monitoring the shedding of feline coronavirus by healthy cats. Veter- Rec. 2001, 148, 649–653, . [CrossRef]
- Addie, D.D.; Jarrett, O. Control of feline coronavirus infection in kittens. Vet Rec. 1990, 126, 164.
- Pedersen NC. Virologic and immunologic aspects of feline infectious peritonitis virus infection. Adv Exp Med Biol. 1987, 218, 529-550.
- Rissi, D.R. A retrospective study of the neuropathology and diagnosis of naturally occurring feline infectious peritonitis. J. Veter- Diagn. Investig. 2018, 30, 392–399, . [CrossRef]
- Lin, L.; Yao, D.; Wu, L.; Fan, R.; Liu, Y.; Zhou, Z. Molecular epidemiology of type I and II feline coronavirus from cats with suspected feline infectious peritonitis in China between 2019 and 2021. Arch. Virol. 2022, 167, 189–194, . [CrossRef]
- Riemer, F.; A Kuehner, K.; Ritz, S.; Sauter-Louis, C.; Hartmann, K. Clinical and laboratory features of cats with feline infectious peritonitis – a retrospective study of 231 confirmed cases (2000–2010). J. Feline Med. Surg. 2016, 18, 348–356, . [CrossRef]
- Soma, T.; Wada, M.; Taharaguchi, S.; Tajima, T. Detection of Ascitic Feline Coronavirus RNA from Cats with Clinically Suspected Feline Infectious Peritonitis. J. Veter- Med Sci. 2013, 75, 1389–1392, . [CrossRef]
- A Worthing, K.; I Wigney, D.; Dhand, N.K.; Fawcett, A.; McDonagh, P.; Malik, R.; Norris, J.M. Risk factors for feline infectious peritonitis in Australian cats. J. Feline Med. Surg. 2012, 14, 405–412, . [CrossRef]
- Pesteanu-Somogyi, L.D.; Radzai, C.; Pressler, B.M. Prevalence of feline infectious peritonitis in specific cat breeds. J. Feline Med. Surg. 2006, 8, 1–5, . [CrossRef]
- Rohrbach, B.W.; Legendre, A.M.; Baldwin, C.A.; Lein, D.H.; Reed, W.M.; Wilson, R.B. Epidemiology of feline infectious peritonitis among cats examined at veterinary medical teaching hospitals. J. Am. Veter- Med Assoc. 2001, 218, 1111–1115, . [CrossRef]
- Foley JE, Pedersen NC. The inheritance of susceptibility to feline infectious peritonitis in purebred catteries. Feline Practice. 1996, 24(1), 14-22.
- Dewerchin, H.L.; Cornelissen, E.; Nauwynck, H.J. Replication of feline coronaviruses in peripheral blood monocytes. Arch. Virol. 2005, 150, 2483–2500, . [CrossRef]
- de Groot-Mijnes JD, van Dun JM, van der Most RG, de Groot RJ. Natural history of a recurrent feline coronavirus infection and the role of cellular immunity in survival and disease. J Virol. 2005, 79(2),1036-1044.
- Acar, D.D.; Olyslaegers, D.A.J.; Dedeurwaerder, A.; Roukaerts, I.D.M.; Baetens, W.; Van Bockstael, S.; De Gryse, G.M.A.; Desmarets, L.M.B.; Nauwynck, H.J. Upregulation of endothelial cell adhesion molecules characterizes veins close to granulomatous infiltrates in the renal cortex of cats with feline infectious peritonitis and is indirectly triggered by feline infectious peritonitis virus-infected monocytes in vitro. J. Gen. Virol. 2016, 97, 2633–2642, . [CrossRef]
- Berg AL, Ekman K, Belák S, Berg M. Cellular composition and interferon-gamma expression of the local inflammatory response in feline infectious peritonitis (FIP). Vet Microbiol. 2005, 111(1-2), 15-23.
- Turin, L.; Riva, F. Toll-Like Receptor Family in Domestic Animal Species. Crit. Rev. Immunol. 2008, 28, 513–538, . [CrossRef]
- Mazaleuskaya, L.; Veltrop, R.; Ikpeze, N.; Martin-Garcia, J.; Navas-Martin, S. Protective Role of Toll-like Receptor 3-Induced Type I Interferon in Murine Coronavirus Infection of Macrophages. Viruses 2012, 4, 901–923, . [CrossRef]
- Koyama, S.; Ishii, K.J.; Coban, C.; Akira, S. Innate immune response to viral infection. Cytokine 2008, 43, 336–341, doi:10.1016/j.cyto.2008.07.009.
- Xu, Q.; Tang, Y.; Huang, G. Innate immune responses in RNA viral infection. Front. Med. 2020, 15, 333–346, . [CrossRef]
- Dedeurwaerder, A.; Olyslaegers, D.A.J.; Desmarets, L.M.B.; Roukaerts, I.D.M.; Theuns, S.; Nauwynck, H.J. ORF7-encoded accessory protein 7a of feline infectious peritonitis virus as a counteragent against IFN-α-induced antiviral response. J. Gen. Virol. 2014, 95, 393–402, . [CrossRef]
- Hosseini, A.; Hashemi, V.; Shomali, N.; Asghari, F.; Gharibi, T.; Akbari, M.; Gholizadeh, S.; Jafari, A. Innate and adaptive immune responses against coronavirus. Biomed. Pharmacother. 2020, 132, 110859–110859, . [CrossRef]
- Chen, S.; Tian, J.; Li, Z.; Kang, H.; Zhang, J.; Huang, J.; Yin, H.; Hu, X.; Qu, L. Feline Infectious Peritonitis Virus Nsp5 Inhibits Type I Interferon Production by Cleaving NEMO at Multiple Sites. Viruses 2019, 12, 43, . [CrossRef]
- Dedeurwaerder, A.; Desmarets, L.M.; Olyslaegers, D.A.; Vermeulen, B.L.; Dewerchin, H.L.; Nauwynck, H.J. The role of accessory proteins in the replication of feline infectious peritonitis virus in peripheral blood monocytes. Veter- Microbiol. 2013, 162, 447–455, . [CrossRef]
- Capozza, P.; Pratelli, A.; Camero, M.; Lanave, G.; Greco, G.; Pellegrini, F.; Tempesta, M. Feline Coronavirus and Alpha-Herpesvirus Infections: Innate Immune Response and Immune Escape Mechanisms. Animals 2021, 11, 3548, . [CrossRef]
- Zuo, W.; Zhao, X. Natural killer cells play an important role in virus infection control: Antiviral mechanism, subset expansion and clinical application. Clin. Immunol. 2021, 227, 108727–108727, . [CrossRef]
- Vermeulen, B.L.; Devriendt, B.; Olyslaegers, D.A.; Dedeurwaerder, A.; Desmarets, L.M.; Favoreel, H.W.; Dewerchin, H.L.; Nauwynck, H.J. Suppression of NK cells and regulatory T lymphocytes in cats naturally infected with feline infectious peritonitis virus. Veter- Microbiol. 2013, 164, 46–59, . [CrossRef]
- Giori, L.; Giordano, A.; Giudice, C.; Grieco, V.; Paltrinieri, S. Performances of different diagnostic tests for feline infectious peritonitis in challenging clinical cases. J. Small Anim. Pr. 2011, 52, 152–157, . [CrossRef]
- Kipar A, Meli ML. Feline infectious peritonitis: still an enigma? Vet Pathol. 2014, 51(2), 505-526.
- Singh, M.; Foster, D.; Child, G.; Lamb, W. Inflammatory cerebrospinal fluid analysis in cats: clinical diagnosis and outcome. J. Feline Med. Surg. 2005, 7, 77–93, . [CrossRef]
- Paltrinieri, S.; Comazzi, S.; Spagnolo, V.; Giordano, A. Laboratory Changes Consistent with Feline Infectious Peritonitis in Cats from Multicat Environments. J. Veter- Med. Ser. A 2002, 49, 503–510, . [CrossRef]
- Hartmann K, Binder C, Hirschberger J, Cole D, Reinacher M, Schroo S, Frost J, Egberink H, Lutz H, Hermanns W. Comparison of different tests to diagnose feline infectious peritonitis. J Vet Intern Med. 2003, 17(6), 781-790.
- Kipar, A.; Bellmann, S.; Kremendahl, J.; Köhler, K.; Reinacher, M. Cellular composition, coronavirus antigen expression and production of specific antibodies in lesions in feline infectious peritonitis. Veter- Immunol. Immunopathol. 1998, 65, 243–257, . [CrossRef]
- Felten, S.; Leutenegger, C.M.; Balzer, H.-J.; Pantchev, N.; Matiasek, K.; Wess, G.; Egberink, H.; Hartmann, K. Sensitivity and specificity of a real-time reverse transcriptase polymerase chain reaction detecting feline coronavirus mutations in effusion and serum/plasma of cats to diagnose feline infectious peritonitis. BMC Veter- Res. 2017, 13, 228, . [CrossRef]
- Sangl, L.; Matiasek, K.; Felten, S.; Gründl, S.; Bergmann, M.; Balzer, H.-J.; Pantchev, N.; Leutenegger, C.M.; Hartmann, K. Detection of feline coronavirus mutations in paraffin-embedded tissues in cats with feline infectious peritonitis and controls. J. Feline Med. Surg. 2018, 21, 133–142, . [CrossRef]
- Emmler, L.; Felten, S.; Matiasek, K.; Balzer, H.-J.; Pantchev, N.; Leutenegger, C.; Hartmann, K. Feline coronavirus with and without spike gene mutations detected by real-time RT-PCRs in cats with feline infectious peritonitis. J. Feline Med. Surg. 2020, 22, 791–799, . [CrossRef]
- Felten, S.; Matiasek, K.; Leutenegger, C.M.; Sangl, L.; Herre, S.; Dörfelt, S.; Fischer, A.; Hartmann, K. Diagnostic Value of Detecting Feline Coronavirus RNA and Spike Gene Mutations in Cerebrospinal Fluid to Confirm Feline Infectious Peritonitis. Viruses 2021, 13, 186, . [CrossRef]
- Felten, S.; Hartmann, K. Diagnosis of Feline Infectious Peritonitis: A Review of the Current Literature. Viruses 2019, 11, 1068, . [CrossRef]
- Kennedy, M.A.; Abd-Eldaim, M.; Zika, S.E.; Mankin, J.M.; Kania, S.A. Evaluation of antibodies against feline coronavirus 7b protein for diagnosis of feline infectious peritonitis in cats. Am. J. Veter- Res. 2008, 69, 1179–1182, . [CrossRef]
- Gut, M.; Leutenegger, C.M.; Huder, J.B.; Pedersen, N.C.; Lutz, H. One-tube fluorogenic reverse transcription-polymerase chain reaction for the quantitation of feline coronaviruses. J. Virol. Methods 1999, 77, 37–46, . [CrossRef]
- Kiss I, Kecskeméti S, Tanyi J, Klingeborn B, Belák S. Preliminary studies on feline coronavirus distribution in naturally and experimentally infected cats. Res Vet Sci. 2000, 68(3), 237-242.
- A Herrewegh, A.; de Groot, R.J.; Cepica, A.; Egberink, H.F.; Horzinek, M.C.; Rottier, P.J. Detection of feline coronavirus RNA in feces, tissues, and body fluids of naturally infected cats by reverse transcriptase PCR. J. Clin. Microbiol. 1995, 33, 684–689, . [CrossRef]
- Duarte, A.; Veiga, I.; Tavares, L. Genetic diversity and phylogenetic analysis of Feline Coronavirus sequences from Portugal. Veter- Microbiol. 2009, 138, 163–168, . [CrossRef]
- A Gunn-Moore, D.; Gruffydd-Jones, T.J.; A Harbour, D. Detection of feline coronaviruses by culture and reverse transcriptase-polymerase chain reaction of blood samples from healthy cats and cats with clinical feline infectious peritonitis. Veter- Microbiol. 1998, 62, 193–205, . [CrossRef]
- Fish, E.J.; Diniz, P.P.V.; Juan, Y.-C.; Bossong, F.; Collisson, E.W.; Drechsler, Y.; Kaltenboeck, B. Cross-sectional quantitative RT-PCR study of feline coronavirus viremia and replication in peripheral blood of healthy shelter cats in Southern California. J. Feline Med. Surg. 2017, 20, 295–301, . [CrossRef]
- Li, X.; Scott, F.W. Detection of feline coronaviruses in cell cultures and in fresh and fixed feline tissues using polymerase chain reaction. Veter- Microbiol. 1994, 42, 65–77, . [CrossRef]
- Herrewegh, A.; Mähler, M.; Hedrich, H.; Haagmans, B.; Egberink, H.; Horzinek, M.; Rottier, P.; de Groot, R. Persistence and Evolution of Feline Coronavirus in a Closed Cat-Breeding Colony. Virology 1997, 234, 349–363, . [CrossRef]
- Bosch BJ, van der Zee R, de Haan CA, Rottier PJ. The coronavirus spike protein is a class I virus fusion protein: structural and functional characterization of the fusion core complex. J Virol. 2003, 77(16), 8801-8811.
- Barker, E.N.; Stranieri, A.; Helps, C.R.; Porter, E.L.; Davidson, A.D.; Day, M.J.; Knowles, T.; Kipar, A.; Tasker, S. Limitations of using feline coronavirus spike protein gene mutations to diagnose feline infectious peritonitis. Veter- Res. 2017, 48, 1–14, . [CrossRef]
- Dunbar, D.; Kwok, W.; Graham, E.; Armitage, A.; Irvine, R.; Johnston, P.; McDonald, M.; Montgomery, D.; Nicolson, L.; Robertson, E.; et al. Diagnosis of non-effusive feline infectious peritonitis by reverse transcriptase quantitative PCR from mesenteric lymph node fine-needle aspirates. J. Feline Med. Surg. 2018, 21, 910–921, . [CrossRef]
- Longstaff, L.; Porter, E.; Crossley, V.J.; E Hayhow, S.; Helps, C.R.; Tasker, S. Feline coronavirus quantitative reverse transcriptase polymerase chain reaction on effusion samples in cats with and without feline infectious peritonitis. J. Feline Med. Surg. 2016, 19, 240–245, . [CrossRef]
- Pedersen, NC. An overview of feline enteric coronavirus and infectious peritonitis virus infections. Feline Practice. 1995, 23, 7-20.
- Hazuchova, K.; Held, S.; Neiger, R. Usefulness of acute phase proteins in differentiating between feline infectious peritonitis and other diseases in cats with body cavity effusions. J. Feline Med. Surg. 2017, 19, 809–816, . [CrossRef]
- Addie, D.D.; McLachlan, S.A.; Golder, M.; Ramsey, I.; Jarrett, O. Evaluation of an in-practice test for feline coronavirus antibodies. J. Feline Med. Surg. 2004, 6, 63–67, . [CrossRef]
- Hirschberger, J.; Hartmann, K.; Wilhelm, N.; Frost, J.; Lutz, H.; Kraft, W. [Clinical symptoms and diagnosis of feline infectious peritonitis].. 1995, 23, 92–9.
- Lewis, K.M.; O’brien, R.T. Abdominal Ultrasonographic Findings Associated With Feline Infectious Peritonitis: A Retrospective Review of 16 Cases. J. Am. Anim. Hosp. Assoc. 2010, 46, 152–160, . [CrossRef]
- Lamb CR. Small animal radiology and ultrasonography. A diagnostic atlas and text. J Feline Med Surg. 2004, 6(1), VI–VII . [CrossRef]
- Delaplace, M.; Huet, H.; Gambino, A.; Le Poder, S. Feline Coronavirus Antivirals: A Review. Pathogens 2021, 10, 1150, . [CrossRef]
- Pedersen, N.C.; Kim, Y.; Liu, H.; Kankanamalage, A.C.G.; Eckstrand, C.; Groutas, W.C.; Bannasch, M.; Meadows, J.M.; Chang, K.-O. Efficacy of a 3C-like protease inhibitor in treating various forms of acquired feline infectious peritonitis. J. Feline Med. Surg. 2017, 20, 378–392, . [CrossRef]
- Pedersen, N.C.; Perron, M.; Bannasch, M.; Montgomery, E.; Murakami, E.; Liepnieks, M.; Liu, H. Efficacy and safety of the nucleoside analog GS-441524 for treatment of cats with naturally occurring feline infectious peritonitis. J. Feline Med. Surg. 2019, 21, 271–281, . [CrossRef]
- Murphy, B.; Perron, M.; Murakami, E.; Bauer, K.; Park, Y.; Eckstrand, C.; Liepnieks, M.; Pedersen, N. The nucleoside analog GS-441524 strongly inhibits feline infectious peritonitis (FIP) virus in tissue culture and experimental cat infection studies. Veter- Microbiol. 2018, 219, 226–233, . [CrossRef]
- Cho A, Saunders OL, Butler T, Zhang L, Xu J, Vela JE, Feng JY, Ray AS, Kim CU. Synthesis and antiviral activity of a series of 1’-substituted 4-aza-7,9-dideazaadenosine C-nucleosides. Bioorg Med Chem Lett. 2012, 22(8), 2705-2507.
- Chang, H.-W.; de Groot, R.J.; Egberink, H.F.; Rottier, P.J.M. Feline infectious peritonitis: insights into feline coronavirus pathobiogenesis and epidemiology based on genetic analysis of the viral 3c gene. J. Gen. Virol. 2010, 91, 415–420, . [CrossRef]
- Sheahan, T.P.; Sims, A.C.; Graham, R.L.; Menachery, V.D.; Gralinski, L.E.; Case, J.B.; Leist, S.R.; Pyrc, K.; Feng, J.Y.; Trantcheva, I.; et al. Broad-spectrum antiviral GS-5734 inhibits both epidemic and zoonotic coronaviruses. Sci. Transl. Med. 2017, 9, . [CrossRef]
- Dickinson, P.J.; Bannasch, M.; Thomasy, S.M.; Murthy, V.D.; Vernau, K.M.; Liepnieks, M.; Montgomery, E.; Knickelbein, K.E.; Murphy, B.; Pedersen, N.C. Antiviral treatment using the adenosine nucleoside analogue GS-441524 in cats with clinically diagnosed neurological feline infectious peritonitis. J. Veter- Intern. Med. 2020, 34, 1587–1593, . [CrossRef]
- Yan VC, Pham CD, Yan MJ, Yan AJ, Khadka S, Arthur K, Ackroyd JJ, Georgiou DK, Roon LE, Bushman LR, et al. Pharmacokinetics of orally administered GS-441524 in dogs. bioRxiv, 2021, 02.04, 429674.
- Xie, J.; Wang, Z. Can remdesivir and its parent nucleoside GS-441524 be potential oral drugs? An in vitro and in vivo DMPK assessment. Acta Pharm. Sin. B 2021, 11, 1607–1616, . [CrossRef]
- Kim Y, Lovell S, Tiew KC, Mandadapu SR, Alliston KR, Battaile KP, Groutas WC, Chang KO. Broad-spectrum antivirals against 3C or 3C-like proteases of picornaviruses, noroviruses, and coronaviruses. J Virol. 2012, 86(21), 11754-11762.
- Perera, K.D.; Kankanamalage, A.C.G.; Rathnayake, A.D.; Honeyfield, A.; Groutas, W.; Chang, K.-O.; Kim, Y. Protease inhibitors broadly effective against feline, ferret and mink coronaviruses. Antivir. Res. 2018, 160, 79–86, . [CrossRef]
- Vennema, H.; de Groot, R.J.; A Harbour, D.; Dalderup, M.; Gruffydd-Jones, T.; Horzinek, M.C.; Spaan, W.J. Early death after feline infectious peritonitis virus challenge due to recombinant vaccinia virus immunization. J. Virol. 1990, 64, 1407–1409, . [CrossRef]
- Kim Y, Liu H, Galasiti Kankanamalage AC, Weerasekara S, Hua DH, Groutas WC, Chang KO, Pedersen NC. Reversal of the progression of fatal coronavirus infection in cats by a broad-spectrum coronavirus protease inhibitor. PLoS Pathog, 2016. 12(3), e1005531.
- Lv, J.; Bai, Y.; Wang, Y.; Yang, L.; Jin, Y.; Dong, J. Effect of GS-441524 in combination with the 3C-like protease inhibitor GC376 on the treatment of naturally transmitted feline infectious peritonitis. Front. Veter- Sci. 2022, 9, 1002488, . [CrossRef]
- E Cook, S.; Vogel, H.; Castillo, D.; Olsen, M.; Pedersen, N.; Murphy, B.G. Investigation of monotherapy and combined anticoronaviral therapies against feline coronavirus serotype II in vitro. J. Feline Med. Surg. 2022, 24, 943–953, . [CrossRef]
- Doki, T.; Toda, M.; Hasegawa, N.; Hohdatsu, T.; Takano, T. Therapeutic effect of an anti-human-TNF-alpha antibody and itraconazole on feline infectious peritonitis. Arch. Virol. 2020, 165, 1197–1206, . [CrossRef]
- Takano, T.; Akiyama, M.; Doki, T.; Hohdatsu, T. Antiviral activity of itraconazole against type I feline coronavirus infection. Veter- Res. 2019, 50, 5, . [CrossRef]
- Tanaka, Y.; Tanabe, E.; Nonaka, Y.; Uemura, M.; Tajima, T.; Ochiai, K. Ionophore Antibiotics Inhibit Type II Feline Coronavirus Proliferation In Vitro. Viruses 2022, 14, 1734, . [CrossRef]
- Ii, D.A.K.; Meujo, D.A.; Hamann, M.T. Polyether ionophores: broad-spectrum and promising biologically active molecules for the control of drug-resistant bacteria and parasites. Expert Opin. Drug Discov. 2009, 4, 109–146, . [CrossRef]
- Tanaka, Y.; Sato, Y.; Sasaki, T. Feline coronavirus replication is affected by both cyclophilin A and cyclophilin B. J. Gen. Virol. 2017, 98, 190–200, . [CrossRef]
- Ballin, A.C.; Schulz, B.; Helps, C.; Sauter-Louis, C.; Mueller, R.S.; Hartmann, K. Limited efficacy of topical recombinant feline interferon-omega for treatment of cats with acute upper respiratory viral disease. Veter- J. 2014, 202, 466–470, . [CrossRef]
- Takano, T.; Kawakami, C.; Yamada, S.; Satoh, R.; Hohdatsu, T. Antibody-Dependent Enhancement Occurs Upon Re-Infection with the Identical Serotype Virus in Feline Infectious Peritonitis Virus Infection. J. Veter- Med Sci. 2008, 70, 1315–1321, . [CrossRef]
- Hohdatsu, T.; Yamada, M.; Tominaga, R.; Makino, K.; Kida, K.; Koyama, H. Antibody-Dependent Enhancement of Feline Infectious Peritonitis Virus Infection in Feline Alveolar Macrophages and Human Monocyte Cell Line U937 by Serum of Cats Experimentally or Naturally Infected with Feline Coronavirus.. J. Veter- Med Sci. 1998, 60, 49–55, . [CrossRef]
- Takano, T.; Yamada, S.; Doki, T.; Hohdatsu, T. Pathogenesis of oral type I feline infectious peritonitis virus (FIPV) infection: Antibody-dependent enhancement infection of cats with type I FIPV via the oral route. J. Veter- Med Sci. 2019, 81, 911–915, . [CrossRef]
- Gerber, J.; Ingersoll, J.; Gast, A.; Christianson, K.; Selzer, N.; Landon, R.; Pfeiffer, N.; Sharpee, R.; Beckenhauer, W. Protection against feline infectious peritonitis by intranasal inoculation of a temperature-sensitive FIPV vaccine. Vaccine 1990, 8, 536–542, . [CrossRef]
- Fehr, D.; Holznagel, E.; Bolla, S.; Hauser, B.; Herrewegh, A.A.; Horzinek, M.C.; Lutz, H. Placebo-controlled evaluation of a modified life virus vaccine against feline infectious peritonitis: safety and efficacy under field conditions. Vaccine 1997, 15, 1101–1109, . [CrossRef]
- A Scherk, M.; Ford, R.B.; Gaskell, R.M.; Hartmann, K.; Hurley, K.F.; Lappin, M.R.; Levy, J.K.; E Little, S.; Nordone, S.K.; Sparkes, A.H. 2013 AAFP Feline Vaccination Advisory Panel Report. J. Feline Med. Surg. 2013, 15, 785–808, . [CrossRef]
- Stoddart, C.; Barlough, J.; Baldwin, C.; Scott, F. Attempted immunisation of cats against feline infectious peritonitis using canine coronavirus. Res. Veter- Sci. 1988, 45, 383–388, . [CrossRef]
- Haijema, B.J.; Volders, H.; Rottier, P.J.M. Live, Attenuated Coronavirus Vaccines through the Directed Deletion of Group-Specific Genes Provide Protection against Feline Infectious Peritonitis. J. Virol. 2004, 78, 3863–3871, . [CrossRef]
- Bálint, .; Farsang, A.; Szeredi, L.; Zádori, Z.; Belák, S. Recombinant feline coronaviruses as vaccine candidates confer protection in SPF but not in conventional cats. Veter- Microbiol. 2014, 169, 154–162, . [CrossRef]
Figure 1.
ML tree of complete S gene is constructed using IQ-Tree under the GTR+F+G4 model.
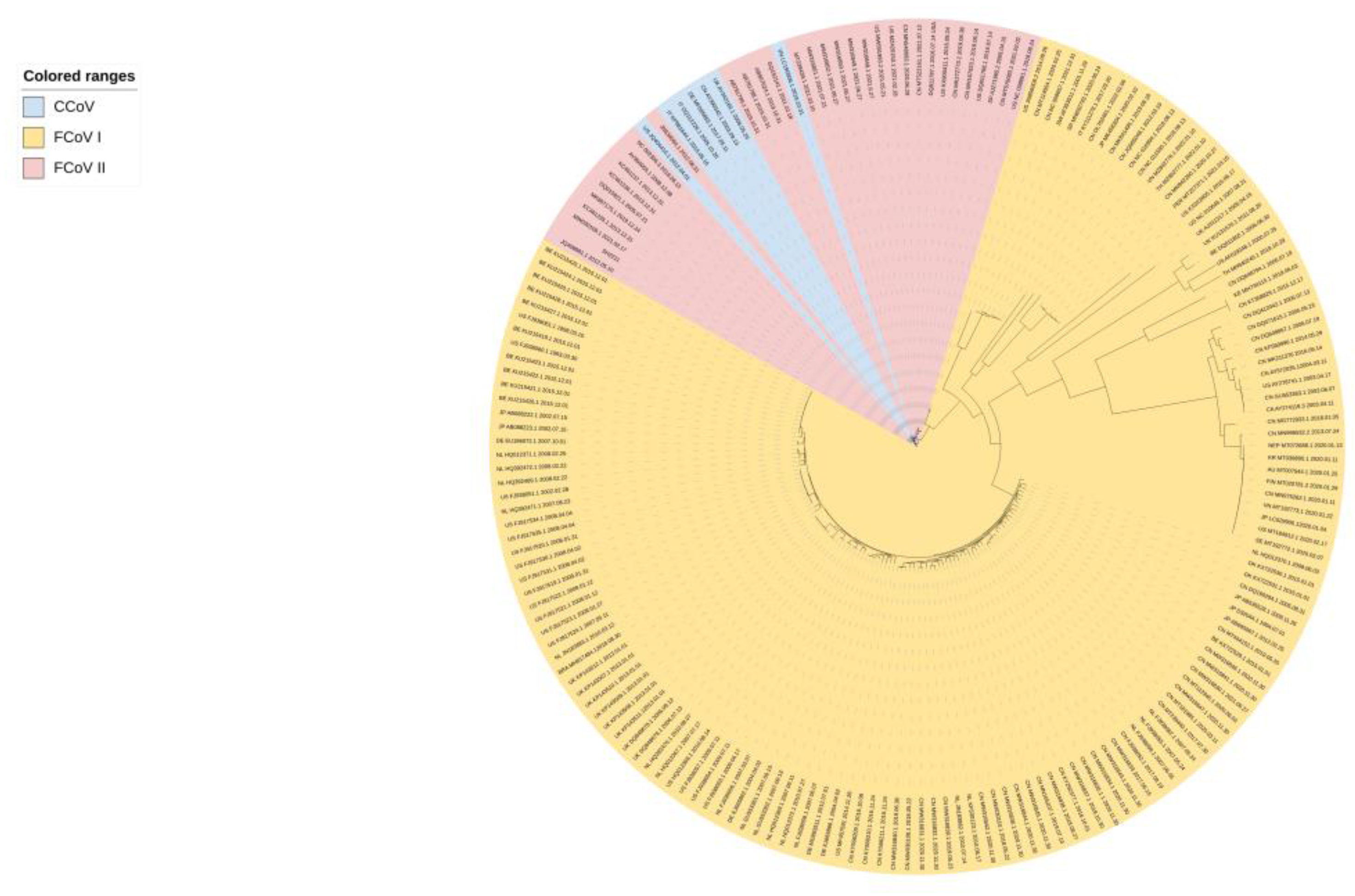
Figure 2.
Schematic representation of the genome of FCoV. The FCoV genome is approximately 29 kb in size and contains 11 ORFs encoding four structural proteins (S, M, E, and N) and seven nonstructural proteins (replicase proteins 1a and 1b and accessory proteins 3a, 3b, 3c, 7a, and 7b). The S protein consists of two subunits: S1 (the receptor binding domain, RBD) and S2 (the fusion domain, FD).
Figure 2.
Schematic representation of the genome of FCoV. The FCoV genome is approximately 29 kb in size and contains 11 ORFs encoding four structural proteins (S, M, E, and N) and seven nonstructural proteins (replicase proteins 1a and 1b and accessory proteins 3a, 3b, 3c, 7a, and 7b). The S protein consists of two subunits: S1 (the receptor binding domain, RBD) and S2 (the fusion domain, FD).
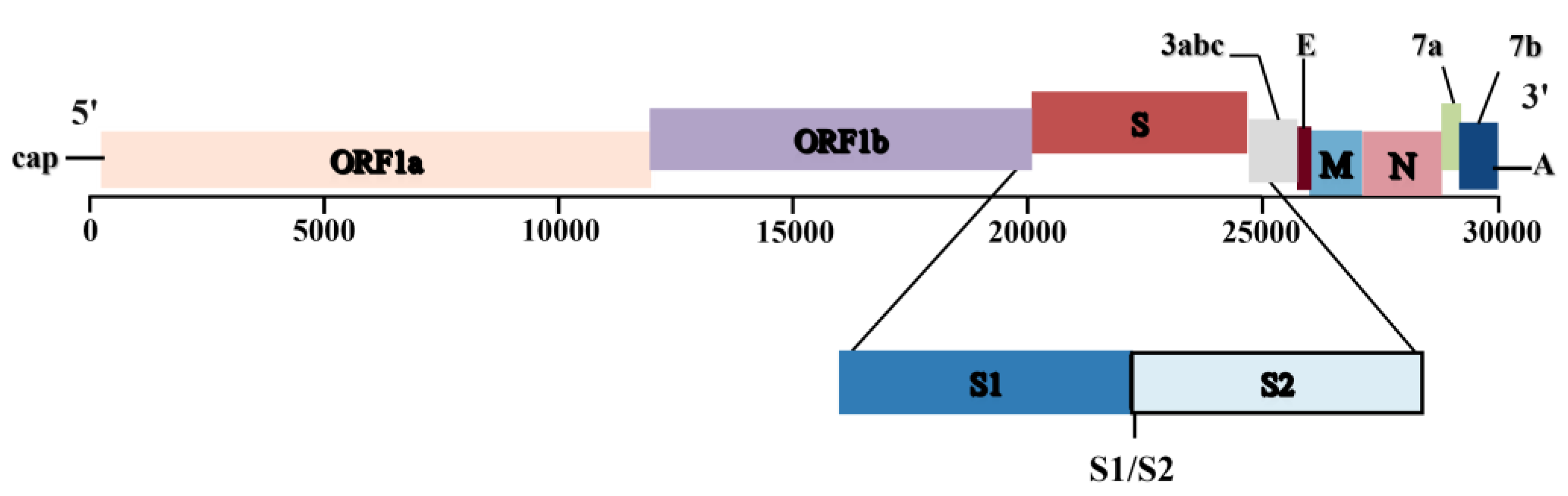
Figure 3.
Schematic diagram of the main routes of FCoV transmission between cats.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



