
Submitted:
09 May 2023
Posted:
09 May 2023
You are already at the latest version
Abstract
Multiresistant pathogens pose a serious threat to human health. The genus candida is one class of human pathogenic yeasts responsible for infections affecting healthy and immunocompromised patients. In this context, plant essential oils emerged as a future natural alternative to control the diseases caused by these pathogens. Here, it highlighted antimicrobial activity and the mechanisms of action of the essential oil extracted from Croton pluriglandulosus Carn.-Torres & Riina (CpEO) leaves on human pathogenic microorganisms in planktonic and biofilm lifestyles. In addition, for the first time, the oil composition was revealed by GC-MS analysis and the toxicity to human red blood cells (HRBC). Twenty-six chemical compounds were identified in EOCp, Elemicin, Bicyclogermacrene, caryophyllene, brevifolin and 2,4,6-trimethoxy-styrene. Through hemolytic assay, it was shown that CpEO has no toxicity to human RBCs. At the concentration of 50 μg mL-1, CpEO did not show great antibacterial potential. However, promising data were found for C. krusei and C. parapsilosis inhibiting by 89.3% and 80.7% of planktonic cell growth and 83.5% and 77.9% the biofilm formation, respectively. Furthermore, the mechanisms of action CpEO were elucidated by fluorescence. Scanning electron microscopy revealed damage to the cell membrane and pore formation, ROS overproduction, and Induction of apoptosis in candida cells. Our results reinforce the potential of CpEO as an effective alternative molecule of pharmaceutical interest.
Keywords:
Croton
; yeast
; inhibition
; chemical composition and action mechanisms
1. Introduction
Pathogens seriously threaten human health and generate global concern due to drug resistance. The genus Candida holds many species, including Candida albicans, the most common and prevalent human pathogenic yeast [1]. However, recent studies emphasize increasing clinical cases by non-albicans yeasts, such as C. glabrata, C. krusei, and C. lusitaniae, affecting healthy and immunocompromised patients [2,3]. The infection establishment can be non-invasive or invasive, causing local and systemic manifestations [4].
The treatment of such infections is made with antifungal agents that directly interfere with the cell structure and metabolism of the fungus. However, due to the misuse, these fungi acquired resistance and became less susceptible to conventional drugs [5]. One form of resistance is the formation of biofilms, which are aggregates of microbial cells embedded in an extracellular polymer matrix that adheres to surfaces and exhibits drug resistance [6].
Plants are a source of promising alternative molecules to control infections. Among these molecules are the secondary metabolites with antimicrobial action [7,8]. One of the natural products with antifungal action is essential oils (EOs). Hydrophobic and volatile substances are usually obtained through plants' plant organs (Leaves, Stem, Root and fruits) [9].
The genus Croton, from the Euphorbiaceae family, adapted to the Caatinga biome, is recognized for grouping diverse species with the potential for producing essential oils and active ingredients. These plants are widely used in folk medicine as teas to reduce pain, intestinal problems, and as anti-inflammatory [10].
In addition, some species are aromatic and produce essential oils with antifungal, insecticidal and antimicrobial properties [11]. Among them C. zehntneri; C. pulegioides Baill; C. jacobinensis Baill; C. nepetaefolius and C. blanchetianus show activity against human pathogens [3,10,12,13].
Despite this large amount of data on Croton species, works with essential oils and their biological applications are still scarce. An example is the C. pluriglandulosus Carn.-Torres & Riina, for which the only information reported in the literature is about the characteristics of the plant, such as being classified as a shrub and the morphology of its leaves as ovate. It has many trichomes on the leaves, latex, and acropeculiar glands. In Brazil, it is predominantly found in regions with the caatinga biome [14]
Given the phytotherapeutic and microbial potential of species belonging to the Croton genus, the present work sought to understand and determine the activity and mechanisms of action of the essential oil extracted from the leaves of Croton pluriglandulosus (CpOE) Carn.-Torres & Riina, which is poorly studied regarding the control of microorganisms in planktonic and biofilm form.
2. Methodology
2.1. Biological material
C. albicans (ATCC 10231), C. krusei (ATCC 6258), C. parapsilosis (ATCC22019), and the bacteria B. subtilis (ATCC 6633), Enterobacter aerogenes (ATCC 13048), Escherichia coli (ATCC 8739), Klebsiella pneumoniae (ATCC 10031), Pseudomonas aeruginosa (ATCC 25619), Staphylococcus aureus (ATCC 25923), and Salmonella enterica (ATCC 14028) were all obtained from the Laboratory of Plant Toxins, Federal University of Ceará, Brazil.
2.2. Plant material and extraction of essential oil
The harvest of the leaves of C. pluriglandulosus Carn. (CpEO) were performed in Serra do Lima, in the city of Patu in Rio Grande do Norte (6º 06' 36'' S, 37º 38' 12''O) state. A voucher specimen was deposited in the Dáardano de Andrade Lima Herbarium of the Federal University of Rural Semiarid (UFERSA), with volume number MOSS 15058. The technique used for oil extraction was based on Oliveira et al. [15], with modifications established by Malveira et al. [16]. Fresh leaves of CpEO (approximately 630 g) were subjected to hydrodistillation in a clevenger apparatus for 2 h. The fresh leaf biomass was weighed and placed in a 5 L volumetric flask containing 2 L of distilled water. The extracted oil was dehydrated with ether and anhydrous sodium sulfate and stored in an amber glass bottle under refrigeration until analysis.
2.3. Characterization of CpEO by GC/MS Analysis
Identification of the major chemical compounds of CpEO was performed by gas chromatography coupled to a mass spectrometer (GC/MS) analysis (Shimadzu GCMS-QP2010 SE, Kyoto, Japan), equipped with an Rtx®-5MS capillary column (30 m × 0.25 mm × 0.25 μm. The CpEO (55 µL) was placed in a vial (2 mL vial with 250 µL inserts) and injected for data acquisition. The operating conditions of the GC-MS/MS were optimized as follows: 70 eV, carrier gas (He), a flow rate of 1.7 mL.min-1 and a pressure of 53.5 KPa. Injector and detector interface temperatures were 25 °C and 230 °C, respectively. The oven temperature program was 100 °C for 3 min-1 and then 310 °C at a heating rate of 3.5 °C min-1 and maintained at 310 °C for 5 min-1. The identification of the constituents of the essential oils was investigated by comparing the mass spectra and Kovats index (IK) values with those of the research reference library.
2.4. Antimicrobial activity
The anticandida assay with adaptations was performed using the microdilution test based on the methodology described by the Clinical and Laboratory Standards Institute (CLST, 2008). Colonies were suspended in Sabouraud Dextrose medium (twice concentrated) and diluted to (2.5 x 106 CFU mL-1). For antimicrobial evaluation, aliquots of 50 µL of medium and yeast cells were added to wells of 96-well plates along with 50 µL of CpEO (ranging from 50 to 0.008 µg mL-1). Itraconazole antifungal and DMSO 5% negative control was used as the positive control. The plates were incubated for 24 h at 30 °C. The yeast development was monitored using a microplate reader (Epoch, Bio-Tek Instruments) at an absorbance of 600 nm. The antibacterial susceptibility assay was based on the method determined by Oliveira et al. [17], with modifications. The Mueller-Hinton Broth medium was used to grow the strains. The experimental setup followed the same principles as the test mentioned above, with only the positive control using ciprofloxacin.
2.5. Antibiofilm assay
For the evaluation of the inhibition of biofilm formation, the assay was based method described by Dias et al. [18]. The experimental strategy, CpEO concentration and assay controls with the planktonic cells. At 24 h, the supernatant was removed and washed three times with 0.15 M NaCl. Next, 200 µL of 100% methyl alcohol for 15 min at 37 °C was added to fix the cells. A 0.1% crystal violet solution (200 µL) was added and incubated for 30 min. Then, the wells were washed with distilled water and 200 µL of 33% acetic acid was added for 15 min. Finally, it was read at an absorbance of 590 nm using a plate reader.
2.6. Mechanisms of Action
2.6.1. Cell membrane integrity
The minimum inhibitory concentration was selected to be evaluated for mechanisms of action. According to Oliveira et al. [19], cell membrane integrity was analyzed with modifications. For the experiment, propidium iodide (PI) was implemented to evaluate CpEO-induced pore formation. First, 50 µL of C. krusei and C. parapsilosis (2.5 x 106 CFU mL-1, in Saboraud liquid medium) were added in the presence of 50 µL of CpEO (50 µg mL-1) and incubated for 24 h at a temperature of 30 °C. Subsequently, each treatment was washed with 100 µL of 0.15 M NaCl and centrifuged (5000 xg, 5 min at 4 °C) three times. The samples were resuspended in 50 µL of 0.15 M NaCl and 3 µL of 1 mM propidium iodide (PI) and incubated for 30 min in the dark at 37 °C. Finally, all treated were washed twice with 0.15 M NaCl, centrifuged to remove excess PI, and observed under a fluorescence microscope (Olympus System BX 60) with an excitation wavelength of 585 nm and emission wavelength 617 nm.
2.6.2. Detection of ROS Overproduction
To evaluate CpEO-induced ROS generation, the method described by Dikalov and Harrison [20], with modifications, was used. All samples were prepared as previously described, with samples incorporated 9 µL of 0.2 mM DCFH-DA (2′,7′-Dichlorofluorescein diacetate) were observed under a microscope (Olympus System BX60) using an excitation wavelength of 485 nm and an emission wavelength of 538 nm.
2.6.3. Induction of apoptosis
The Induction of apoptosis in the species studied was based on using CellEvent (ThermoFisher), following the manufacturer's restrictions for detecting caspase action. Samples were prepared following the same pattern described in the previous tests and 9 µL of CellEvent® reagent (ThermoFisher, São Paulo, SP, Brazil) was added. Finally, washes and centrifugations were performed as before. Finally, the treatments were observed with a microscope (Olympus System BX60) using an excitation wavelength of 342 nm and an emission wavelength of 441 nm.
2.7. Scanning Electron Microscopy (SEM)
SEM (Billerica, MA, USA) analysis was performed to evaluate cell morphological changes based on the method described by Staniszewska et al. [21]. The cells were treated under the same conditions described in the previous sections, then fixed with 1% (v/v) glutaraldehyde in 0.15 M sodium phosphate buffer at pH 7.2 for 16 h at room temperature. This was followed by three washes with sodium phosphate buffer pH 7.2 and centrifuged (5000g for 5 min at 4 °C). To dehydrate, the samples were dried with ethanol (30%, 50%, 70%, 100%, 100% (v/v) for 10 min each and centrifuged as above. Finally, 50% hexamethyldisilane (HMDS, Sigma, St. Louis, MI, USA) was diluted in ethanol, left for 10 min, followed by centrifugation, and 1000% HMDS was added. 15 µL was transferred to a coverslip to dry at room temperature. The cells were coated with a gold layer with an aluminum surface using a positron emission tomography (PET) coating machine (Emitech-Q150TES, Quorum Technologies, Lewes, England) and the images observed on a scanning electron microscope (Quanta 450 FEG, FEI, Waltham, MA).
2.8. Hemolytic analysis
Hemolytic analysis of CpEO was evaluated with human erythrocyte blood types A, B, and O, as Oliveira et al. [19] described. Three concentrations were tested, including the one used in all the assays mentioned above. Blood was centrifuged at 5000 g for 5 min at 10 °C, washed with 0.15 M NaCl and diluted to a concentration of 2.5%. Each blood type was incubated (300 µL) with the CpEO solution (300 µL) at concentrations of 1, 3, and 5 mg mL-1 for 30 min at 37 °C. DMSO-NaCl (5%) and 0.1% Triton X-100 (100 %) were used as negative and positive controls for hemolysis. The samples were centrifuged (5000 g for 5 min at 4 °C, Eppendorf 5810 centrifuge, Hannover, Germany). Then the supernatants were collected and directed into 96-well plates, and the hemolysis (%) was calculated by an absorbance of 414 nm using a microplate reader.
2.9. Statistical Analysis
Three independent replicates were performed for all analyses performed. Data were subjected to the application of the ANOVA test, followed by Tukey's method using GraphPad Prism software version 5.01 (GraphPad Software company, Santa Clara, CA, USA). A p < 0.05 was considered statistically significant.
3. Results and Discussion
3.1. GC-MS/MS analysis CpEO by
The GC-MS/MS analysis identified 26 metabolites in CpEO. Elemicin (25.77%), Bicyclogermacrene (9.37%), caryophyllene (8.99%), brevifolin (4.60%) and 2,4,6-trimethoxy-styrene (3.96%) represent the compounds in the highest amount (Table 1). Many metabolites identified in CpEO belong to the terpenoid family, classified as monoterpenes and sesquiterpenes and present several biological activities, including antimicrobial activity [22]. Elemicin has only a few reports in the literature about its potential against human pathogens; however, Rossi et al. [23] revealed the antimicrobial activity against the bacterium Campylobacter jejuni. Bicyclogermacrene and 2,4,6-trimethoxy-styrene, both compounds found in CpEO, have antifungal, antioxidant, larvicidal and anti-inflammatory activities [24,25,26].
Some compounds exhibit anticandidal activity. One example is eucalyptol and α-terpineol (Table 1). According to the literature, eucalyptol exhibits anti-inflammatory, antioxidant, antimicrobial and anticandidal action [27,28]. Studies show that α-terpineol has activity against yeasts and filamentous fungi, including Candida species (PINTO et al., 2014). Besides, β-elemene, 4-terpineol and δ-elemene were associated with antifungal, antibacterial, and anticancer activities [29,30]. Many of the components identified in CpEO are present in other species of the genus Croton, however, in different quantities [16]. This difference is related to factors such as location, climate, stage of plant development and the organ used for the study [31,32,33].
3.2. Antimicrobial Activity of CpEO
The inhibitory activity of CpEO (50 μg mL-1) against fungal and bacterial strains is shown in Table 2. The CpEO was not effective against bacterial pathogens (Table 1). In a recent study, Malveira et al. [16] reported that the essential oil from C. blanchetianus is ineffective against bacteria. Nevertheless, a significant reduction in the growth of two candida species tested was observed, 89.3% for C. krusei and 80.7% for C. parapsilosis at a concentration of 50 μg mL-1. The inhibition of biofilm formation for the same microorganisms was 83.5 for C. krusei and 77.9% for C. parapsilosis, respectively. The search for alternatives to treating infectious diseases caused by yeasts from the Candida genus has a significant impact, mainly because of the antifungal resistance, which is rapidly developing due to the indiscriminate use of antifungal agents [34]. In this context, plant essential oils emerged as a promising alternative for treating these pathogens.
Malveira et al. [16] reported that the essential oil of C. blanchetianus possesses activity against C. albicans and C. parapsilosis in planktonic cells and biofilm using the same concentration tested in this study. Asdade et al. [35] showed the antifungal action of Vitex agnus-castus L. essential oil, rich in monoterpene and sesquiterpene compounds, against clinical Candida strains. Taweechaisupapong et al. [36] also obtained satisfactory results using lemongrass (Cymbopogon flexuosus) oil aiming at biofilm inhibition of Candida isolates at 0.8 and 0.4 mg mL-1, concentrations 16 and 8 times higher the concentration presented by CpEO.
A study with C. rhamnifolioides oil revealed activity against Aeromonas hydrophila, E. coli, Listeria monocytogenes, S. Enteritidis, and S. aureus [37]. Filho et al. [38] reported that the oil of the Himatanthus obovathus flowers was efficient against C. krusei; however, for C. albicans, the oil was ineffective. Moremi et al. (2021) pointed out the Croton species from Southern Africa; C. gratissimus Burch., C. megalobotrys Müll. Arg., C. menyhartii Gȕrke, C. pseudopulchellus Pax, C. steenkampianus Gerstner and C. sylvaticus Schltdl., showed low inhibitory capacity against Escherichia coli. These results suggest the selectivity of essential oils against distinct species of bacteria and fungi.
Furthermore, the antimicrobial activity of CpEO may come from compounds such as Elimicin, Eucalyptol, Bicyclogermacrene, α-terpineol β-elemene, 4-terpineol and δ-elemene, already mentioned in the topic above. These compounds induce cell instability, causing membrane damage, interference with ATP synthesis, and causing extravasation of intracellular contents [39]. Additionally, it has been reported that genes such as ALS1, ALS3 and HWP1, involved in the biofilm formation process, can be silenced by the action of some plant chemical compounds [40]. This shows the effectiveness of the substances found in oils, which can act as new methods of natural control against microorganisms.
3.3. Mechanisms of Action
3.3.1. Cell membrane integrity
Fluorescence microscopy was used to understand the mechanisms of action of the CpEO. The PI cell membrane fluorescent dye became helpful in the identification of damage to yeast cell membranes, and PI binds to the DNA structure and emits red fluorescence. This substance cannot cross the membrane of living cells because they are impermeable, so PI only moves throughout damaged membranes [41].
As expected, the control with DMSO-NaCl showed no fluorescence. However, red fluorescence indicated membrane damage in the planktonic cells of C. krusei and C. parapsilosis (Figure 1). The same damage on membranes was observed in the biofilm cells in the presence of the CpEO (Figure 2). An et al. [42] highlight α-terpineol and terpene-4-ol as compounds that induced damage to the membranes of Aspergillus niger, leading to cell death. These same compounds were also identified in CpEO.
It is worth noting that the PI does not reveal the pore size. For this purpose, a new analysis was performed with dextran (6 kDa) in conjunction with FITC (fluorescein isothiocyanate - green fluorescence) in planktonic and biofilm cells (Figure 1 and Figure 2, right panel). The formation of pores in the membrane is revealed by the fluorescence (green), which was stronger in the biofilm, indicating a 6-kDa-sized pore in the cell membranes. The results presented by FITC-Dextran are significant. Although largely used, sometimes PI does not reveal a pore formed. That happens because PI sometimes increases the membrane permeability, which is not necessarily a pore formation. Another problem with PI is due to the size of the pore is too low, allowing the cells to recover [43]. However, both PI and FITC-Dextran assays provided robust results suggesting membrane pore formation.
It is feasible to suggest that the damage caused by CpEO to the cell membrane of yeasts is given the presence of components in CpEO that were already mentioned in previous topics. Since these compounds act by interfering with the enzymatic mechanism that supports the structural system of the cell wall, causing morphological damage [44].
3.3.2. ROS Overproduction and Induction of Apoptosis
CpEO induced a slight ROS production in planktonic cells (Figure 3) and biofilm (Figure 4). As expected, DMSO-NaCl did not induce ROS in any treatment. For planktonic cells, CpEO induced ROS accumulation in C. krusei and C. parapsilosis (Figure 3). However, for a biofilm lifestyle, CpEO was more efficient against C. krusei (Figure 4). In the experiment with biofilm, it is trustworthy to mention that in the light field, the number of cells in the treatment with CpEO is smaller than control (Figure 4 – light field). Other reports of ROS overproduction in human pathogenic cells induced by essential oils [16,45,46]. Recently, it was shown that the lavender (Lavandula angustifolia) induced ROS accumulation in Klebsiella pneumoniae cells, followed by programmed cell death [46].
The chemical constituents of essential oils are involved in the formation of ROS by promoting the accumulation of molecules resulting from a disorder in biochemical processes [47]. Different Cymbopogon martinii essential oil concentrations stimulated ROS production in A. flavus [48]. Thymoquinone, a phytochemical compound found in essential oil, generated oxidative stress in C. glabrata [49]. It was recently demonstrated that the essential oil from C. blanchetianus leaves, rich in terpene compounds, could induce ROS in planktonic and biofilm cells of Candida species [16] hence corroborating with the data found in this research.
ROS are essential molecules for maintaining cellular balance and even establishing biofilms but only at optimal levels [50]. Nonetheless, the levels of ROS must be tightly controlled; if there is an imbalance, their accumulation can cause cell apoptosis by damaging proteins, lipids, carbohydrates, and nucleic acids [51].
Keeping in mind that ROS accumulation induced by CpEO could lead to cell death in C. krusei and C. parapsilosis, it was performed the analysis of apoptosis in Candida cells. Caspase-3/7 activity was observed in all oil-treated cells, less intense in planktonic cells (Figure 5 and Figure 6). As expected, the control (DMSO-NaCl solution) was not induced DNA degradation, as no fluorescence was detected. Thus, CpEO induced DNA damage in C. krusei and C. parapsilosis. Sun et al. [52] showed that cells treated with eucalyptol and β-cyclocitral activated caspase-3 and caspase-9, proteases involved in the apoptosis process.
It is known that caspase-3 initiates apoptotic DNA fragmentation and triggers a series of reactions involved in this process [53]. It is worth noting that few studies with essential oils show this type of test in yeast, not describing the mechanism of action of the oil. A recent study revealed by proteomics that limonene induced a higher accumulation of proteins involved in DNA damage and apoptosis [54].
3.4. Scanning Electron Microscopy (SEM)
SEM was used to evaluate the damage caused by CpEO in planktonic cells of C. krusei and C. parapsilosis after all treatments. The images depict that the control cells for both species have healthy anatomy without damage (Figure 7A and Figure 8A). In contrast, the damage is visible in CpEO-treated cells. Among them, in cell morphology, deformations in the structure, damaged cell walls, and contorted cells (Figure 7B to F; Figure 8B to F). For both Candida species is clear that CpEO induced deadly damages. It is possible to see holes in the cell structure, indicating that some parts of the membrane and cell wall are gone.
3.5. Hemolytic assay
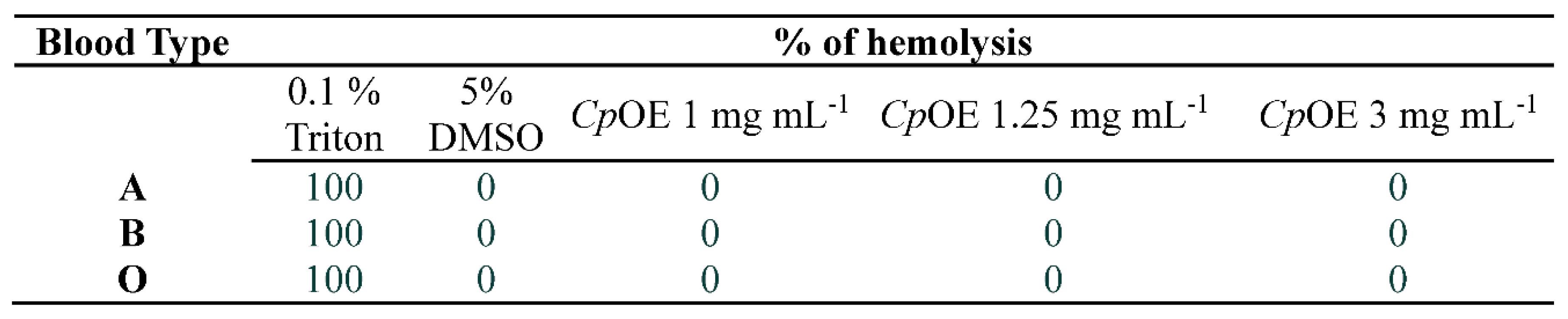
CpEO did not show hemolytic activity against any human blood type tested (Table 3), even at the concentration of 3 mg mL 1. Thus, CpEO was not toxic to human red blood cells. This result reinforces the pharmacological application of CpEO, as it does not present toxicity risks. This is one of the strong reasons for these substances to be used clinically as drugs [17]. For example, Malveira et al. [16] revealed the oil from C. blanchetianus was toxic to red blood cells type O.
Being not harmful to human red blood cells is a critical feature to molecules could be applied in treatment once blood is the vehicle for these molecules. Therefore, CpEO has a great potential to be assessed as source of new antimicrobial molecules.
4. Conclusion
Our research reveals that the essential oil from Croton pluriglandulosus Carn.-Torres & Riina leaves represents a possible alternative for the control of human pathogens that already present resistance to conventional drugs offered by the pharmaceutical industry. These molecules of natural origin, have different types of chemical compounds that interfered in the homeostasis of the tested yeast cells. It is possible to suggest and conclude that the oil was efficient against C. krusei and C. parapsilosis. It revealed membrane and cell wall damage, induced DNA fragmentation, and did not present toxicity to human red blood cells.
Data availability
Data are available under reasonable requirements.
Acknowledgments and Funding
Pedro F. N. Souza has a grant for senior researcher from CNPq at process number 305003/2022-4. We are thankful to Central Analítica at UFC for providing SEM analysis.
Conflict of interest
The authors have declared no conflict of interest, including financial, personal, or other relationships with other people or organizations.
References
- Butler, G.; Rasmussen, M.D.; Lin, M.F.; Santos, M.A.S.; Sakthikumar, S.; Munro, C.A.; Rheinbay, E.; Grabherr, M.; Forche, A.; Reedy, J.L.; et al. Evolution of Pathogenicity and Sexual Reproduction in Eight Candida Genomes. Nature 2009, 459, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Ding, Y.; Wei, B.; Wang, L.; Xu, S.; Qin, P.; Wei, L.; Jiang, L. Epidemiology of Candida Albicans and Non-C.Albicans of Neonatal Candidemia at a Tertiary Care Hospital in Western China. BMC Infect. Dis. 2017, 17, 329. [Google Scholar] [CrossRef] [PubMed]
- Lima, I. de O.; Oliveira, R. de A.G.; Lima, E. de O.; Farias, N.M.P.; de Souza, E.L. Atividade Antifúngica de Óleos Essenciais Sobre Espécies de Candida. Rev. Bras. Farmacogn. 2006, 16, 197–201. [Google Scholar] [CrossRef]
- Vieira & Nascimento, T., F. Resistência a Fármacos Antifúngicos Por Candida e Abordagem Terapêutica. Rev. Port. Farmacoter. 2017, 9, 29–36. [Google Scholar]
- Arendrup, M.C.; Patterson, T.F. Multidrug-Resistant Candida: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S445–S451. [Google Scholar] [CrossRef] [PubMed]
- do Vale, J.P.C.; Vasconcelos, M.A.; Arruda, F.V.S.; Firmino, N.C.S.; Pereira, A.L.; Andrade, A.L.; Saker-Sampaio, S.; Sampaio, A.H.; Marinho, E.S.; Teixeira, A.M.R.; et al. Evaluation of Antimicrobial and Antioxidant Potential of Essential Oil from Croton Piauhiensis Müll. Arg. Curr. Microbiol. 2021, 78, 1926–1938. [Google Scholar] [CrossRef]
- Benini, P.C.; Schwan-Estrada, K.R.F.; Klais, E.C.; Cruz, M.E.S.; Itako, A.T.; Mesquini, R.M.; Stangarlin, J.R.; Tolentino Júnior, J.B. EFEITO IN VITRO DO ÓLEO ESSENCIAL E EXTRATO AQUOSO DE OCIMUM GRATISSIMUM COLHIDO NAS QUATRO ESTAÇÕES DO ANO SOBRE FITOPATÓGENOS. Arq. Inst. Biol. (Sao. Paulo). 2010, 77, 677–683. [Google Scholar] [CrossRef]
- Souza, A.E.F.; Araújo, E.; Nascimento, L.C. Atividade Antifúngica de Extratos de Alho e Capim-Santo Sobre o Desenvolvimento de Fusarium Proliferatum Isolado de Grãos de Milho. Fitopatol. Bras. 2007, 32, 465–471. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, Function and Metabolic Engineering of Plant Volatile Organic Compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef]
- Arrais, L.G.; Lyra, H.F.S.; Batista, D.C.A.; Coutinho, F.N.; Saraiva, A.M.; Pereira, R.C.A.; Pisciottano, M.N.C.; Xavier, H.S.; Melo, S.J. Atividade Antimicrobiana Dos Extratos Metanólicos Da Raiz, Caule e Folhas de Croton Pulegioides Baill. (Zabelê). Rev. Bras. Plantas Med. 2014, 16, 316–322. [Google Scholar] [CrossRef]
- Guerra, A.M.N. de M.; Silva, D. dos S.; Santos, P.S.; dos Santos, L.B. Teste de Repelência de Óleos Essenciais Sobre Callosobruchus Maculatus. Rev. Bras. Agropecuária Sustentável 2019, 9. [Google Scholar] [CrossRef]
- da Costa, J.G.M.; Rodrigues, F.F.G.; Angélico, E.C.; Pereira, C.K.B.; de Souza, E.O.; Caldas, G.F.R.; Silva, M.R.; Santos, N.K.A.; Mota, M.L.; dos Santos, P.F. Composição Química e Avaliação Da Atividade Antibacteriana e Toxicidade Do Óleo Essencial de Croton Zehntneri (Variedade Estragol). Rev. Bras. Farmacogn. 2008, 18, 583–586. [Google Scholar] [CrossRef]
- de Vasconcelos, E.C.; Paganini, C.C.; de Figueiredo, E.A.T.; de Aragão, G.M.F. Ação Antimicrobiana Dos Compostos Voláteis Do Óleo Essencial Das Folhas Secas de Croton Blanchetianus Baill. Res. Soc. Dev. 2022, 11, e28011124785. [Google Scholar] [CrossRef]
- Riina, R.; Van Ee, B.W.; Beatriz Rossi Caruzo, M.; Santos Carneiro-Torres, D.; Freitas dos Santos, R.; Berry, P.E. THE NEOTROPICAL CROTON SECTION GEISELERIA (EUPHORBIACEAE): CLASSIFICATION UPDATE, PHYLOGENETIC FRAMEWORK, AND SEVEN NEW SPECIES FROM SOUTH AMERICA 1. [CrossRef]
- de Oliveira, A.R.M.F.; Jezler, C.N.; Oliveira, R.A.; Mielke, M.S.; Costa, L.C.B. Determinação Do Tempo de Hidrodestilação e Do Horário de Colheita No Óleo Essencial de Menta. Hortic. Bras. 2012, 30, 155–159. [Google Scholar] [CrossRef]
- Malveira, E.A.; Souza, P.F.N.; Neto, N.A.S.; Aguiar, T.K.B.; Rodrigues, N.S.; Henrique, C.W.B.; Silva, A.F.B.; Lima, L.B.; Albuquerque, C.C.; Freitas, C.D.T. Essential Oil from Croton Blanchetianus Leaves: Anticandidal Potential and Mechanisms of Action. J. Fungi 2022, 8, 1147. [Google Scholar] [CrossRef]
- Souza, P.F.N.; Marques, L.S.M.; Oliveira, J.T.A.; Lima, P.G.; Dias, L.P.; Neto, N.A.S.; Lopes, F.E.S.; Sousa, J.S.; Silva, A.F.B.; Caneiro, R.F.; et al. Synthetic Antimicrobial Peptides: From Choice of the Best Sequences to Action Mechanisms. Biochimie 2020, 175, 132–145. [Google Scholar] [CrossRef]
- Dias, L.P.; Souza, P.F.N.; Oliveira, J.T.A.; Vasconcelos, I.M.; Araújo, N.M.S.; Tilburg, M.F.V.; Guedes, M.I.F.; Carneiro, R.F.; Lopes, J.L.S.; Sousa, D.O.B. RcAlb-PepII, a Synthetic Small Peptide Bioinspired in the 2S Albumin from the Seed Cake of Ricinus Communis, Is a Potent Antimicrobial Agent against Klebsiella Pneumoniae and Candida Parapsilosis. Biochim. Biophys. Acta - Biomembr. 2020, 1862, 183092. [Google Scholar] [CrossRef]
- Oliveira, J.T.A.; Souza, P.F.N.; Vasconcelos, I.M.; Dias, L.P.; Martins, T.F.; Van Tilburg, M.F.; Guedes, M.I.F.; Sousa, D.O.B. Mo-CBP3-PepI, Mo-CBP3-PepII, and Mo-CBP3-PepIII Are Synthetic Antimicrobial Peptides Active against Human Pathogens by Stimulating ROS Generation and Increasing Plasma Membrane Permeability. Biochimie 2019, 157, 10–21. [Google Scholar] [CrossRef]
- Dikalov, S.I.; Harrison, D.G. Methods for Detection of Mitochondrial and Cellular Reactive Oxygen Species. Antioxid. Redox Signal. 2014, 20, 372–382. [Google Scholar] [CrossRef]
- Staniszewska, M.; Bondaryk, M.; Swoboda-Kopec, E.; Siennicka, K.; Sygitowicz, G.; Kurzatkowski, W. Candida Albicans Morphologies Revealed by Scanning Electron Microscopy Analysis. Brazilian J. Microbiol. 2013, 44, 813–821. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.; Ademiluyi, A.; et al. Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef] [PubMed]
- Rossi, P.-G.; Bao, L.; Luciani, A.; Panighi, J.; Desjobert, J.-M.; Costa, J.; Casanova, J.; Bolla, J.-M.; Berti, L. ( E )-Methylisoeugenol and Elemicin: Antibacterial Components of Daucus Carota L. Essential Oil against Campylobacter Jejuni. J. Agric. Food Chem. 2007, 55, 7332–7336. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, A.A.; Vieira, L.; Ribeiro, R.I.M.de A.; Thomé, R.G.; dos Santos, H.B.; Silva, D.B.; Carollo, C.A.; de Oliveira, F.M.; Lopes, D. de O.; de Siqueira, J.M.; et al. Chemical Composition and Evaluation of the Anti-Inflammatory and Antinociceptive Activities of Duguetia Furfuracea Essential Oil: Effect on Edema, Leukocyte Recruitment, Tumor Necrosis Factor Alpha Production, INOS Expression, and Adenosinergic and Opioid. J. Ethnopharmacol. 2019, 231, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Nogueira Sobrinho, A.C.; de Morais, S.M.; Marinho, M.M.; de Souza, N.V.; Lima, D.M. Antiviral Activity on the Zika Virus and Larvicidal Activity on the Aedes Spp. of Lippia Alba Essential Oil and $β$-Caryophyllene. Ind. Crops Prod. 2021, 162, 113281. [Google Scholar] [CrossRef]
- Maia, D.S.; Lopes, C.F.; Saldanha, A.A.; Silva, N.L.; Sartori, Â.L.B.; Carollo, C.A.; Sobral, M.G.; Alves, S.N.; Silva, D.B.; de Siqueira, J.M. Larvicidal Effect from Different Annonaceae Species on Culex Quinquefasciatus. Environ. Sci. Pollut. Res. 2020, 27, 36983–36993. [Google Scholar] [CrossRef]
- Seol, G.H.; Kim, K.Y. Eucalyptol and Its Role in Chronic Diseases. In; 2016; pp. 389–398.
- Mishra, P.; Gupta, P.; Srivastava, A.K.; Poluri, K.M.; Prasad, R. Eucalyptol/ $β$-Cyclodextrin Inclusion Complex Loaded Gellan/PVA Nanofibers as Antifungal Drug Delivery System. Int. J. Pharm. 2021, 609, 121163. [Google Scholar] [CrossRef]
- Sousa, O. V; Del-Vechio-Vieira, G.; Alves, M.S.; Araújo, A.A.L.; Pinto, M.A.O.; Amaral, M.P.H.; Rodarte, M.P.; Kaplan, M.A.C. Chemical Composition and Biological Activities of the Essential Oils from Duguetia Lanceolata St. Hil. Barks. Molecules 2012, 17, 11056–11066. [Google Scholar] [CrossRef]
- XIE, C.-Y.; YANG, W.; LI, M.; YING, J.; TAO, S.-J.; LI, K.; DONG, J.-H.; WANG, X.-S. Cell Apoptosis Induced by $δ$-Elemene in Colorectal Adenocarcinoma Cells via a Mitochondrial-Mediated Pathway. YAKUGAKU ZASSHI 2009, 129, 1403–1413. [Google Scholar] [CrossRef]
- Moremi, M.P.; Kamatou, G.P.; Viljoen, A.M.; Tankeu, S.Y. Croton Gratissimus - Essential Oil Composition and Chemometric Analysis of an Ethnomedicinally Important Tree from South Africa. South African J. Bot. 2021, 138, 141–147. [Google Scholar] [CrossRef]
- Moremi, M.P.; Makolo, F.; Viljoen, A.M.; Kamatou, G.P. A Review of Biological Activities and Phytochemistry of Six Ethnomedicinally Important South African Croton Species. J. Ethnopharmacol. 2021, 280, 114416. [Google Scholar] [CrossRef]
- Thompson, J.D.; Chalchat, J.C.; Michet, A.; Linhart, Y.B.; Ehlers, B. Qualitative and Quantitative Variation in Monoterpene Co-Occurrence and Composition in the Essential Oil of Thymus Vulgaris Chemotypes. J. Chem. Ecol. 2003, 29, 859–880. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and Mechanisms of Antifungal Resistance. Antibiotics 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Asdadi, A.; Hamdouch, A.; Oukacha, A.; Moutaj, R.; Gharby, S.; Harhar, H.; El Hadek, M.; Chebli, B.; Idrissi Hassani, L.M. Study on Chemical Analysis, Antioxidant and in Vitro Antifungal Activities of Essential Oil from Wild Vitex Agnus-Castus L. Seeds Growing in Area of Argan Tree of Morocco against Clinical Strains of Candida Responsible for Nosocomial Infections. J. Mycol. Med. 2015, 25, e118–e127. [Google Scholar] [CrossRef] [PubMed]
- Taweechaisupapong, S.; Aieamsaard, J.; Chitropas, P.; Khunkitti, W. Inhibitory Effect of Lemongrass Oil and Its Major Constituents on Candida Biofilm and Germ Tube Formation. South African J. Bot. 2012, 81, 95–102. [Google Scholar] [CrossRef]
- da Costa, A.C.V.; Melo, G.F.A.; Madruga, M.S.; da Costa, J.G.M.; Garino Junior, F.; Queiroga Neto, V. Chemical Composition and Antibacterial Activity of Essential Oil of a Croton Rhamnifolioides Leaves Pax & Hoffm. Semin. Ciências Agrárias 2013, 34, 2853. [Google Scholar] [CrossRef]
- Filho, A.C.; Castro, C.F.; da Silva, A.; da Cruz, R. Efeito Antifúngico Sobre Candida Pelo Óleo Essencial Da Flor de Himatanthus Obovathus (Müell. Arg.) Woodson (Apocynaceae). Rev. Arq. Científicos 2021, 4. [Google Scholar]
- Tariq, S.; Wani, S.; Rasool, W.; Shafi, K.; Bhat, M.A.; Prabhakar, A.; Shalla, A.H.; Rather, M.A. A Comprehensive Review of the Antibacterial, Antifungal and Antiviral Potential of Essential Oils and Their Chemical Constituents against Drug-Resistant Microbial Pathogens. Microb. Pathog. 2019, 134, 103580. [Google Scholar] [CrossRef]
- Raut, J.S.; Shinde, R.B.; Chauhan, N.M.; Mohan Karuppayil, S. Terpenoids of Plant Origin Inhibit Morphogenesis, Adhesion, and Biofilm Formation by Candida Albicans. Biofouling 2013, 29, 87–96. [Google Scholar] [CrossRef]
- Neto, N.A.S.; Oliveira, J.T.A.; Aguiar, T.K.B.; Bezerra, L.P.; Branco, L.A.C.; Mesquita, F.P.; Freitas, C.D.T.; Souza, P.F.N. Synergistic Antibiofilm Activity between Synthetic Peptides and Ciprofloxacin against Staphylococcus Aureus. Pathogens 2022, 11, 995. [Google Scholar] [CrossRef]
- An, P.; Yang, X.; Yu, J.; Qi, J.; Ren, X.; Kong, Q. $α$-Terpineol and Terpene-4-Ol, the Critical Components of Tea Tree Oil, Exert Antifungal Activities in Vitro and in Vivo against Aspergillus Niger in Grapes by Inducing Morphous Damage and Metabolic Changes of Fungus. Food Control 2019, 98, 42–53. [Google Scholar] [CrossRef]
- Etxaniz, A.; González-Bullón, D.; Martín, C.; Ostolaza, H. Membrane Repair Mechanisms against Permeabilization by Pore-Forming Toxins. Toxins (Basel). 2018, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- de Billerbeck, V.G.; Roques, C.G.; Bessière, J.-M.; Fonvieille, J.-L.; Dargent, R. Effects of Cymbopogon Nardus (L.) W. Watson Essential Oil on the Growth and Morphogenesis of Aspergillus Niger. Can. J. Microbiol. 2001, 47, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Zhang, K.; Yin, Y.; Wu, J. D319 Induced Antifungal Effects through ROS-Mediated Apoptosis and Inhibited Isocitrate Lyase in Candida Albicans. Biochim. Biophys. Acta - Gen. Subj. 2022, 1866, 130050. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.K.; Yusoff, K.; Thomas, W.; Akseer, R.; Alhosani, M.S.; Abushelaibi, A.; Lim, S.H.E.; Lai, K.S. Lavender Essential Oil Induces Oxidative Stress Which Modifies the Bacterial Membrane Permeability of Carbapenemase Producing Klebsiella Pneumoniae. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Tang, X.; Shao, Y.-L.; Tang, Y.-J.; Zhou, W.-W. Antifungal Activity of Essential Oil Compounds (Geraniol and Citral) and Inhibitory Mechanisms on Grain Pathogens (Aspergillus Flavus and Aspergillus Ochraceus). Molecules 2018, 23, 2108. [Google Scholar] [CrossRef]
- Lin, L.; Chen, S.; Xia, W.; Li, T.; Dong, L.; Zhang, Y.; Zhang, Y.; Li, H.; Zhao, Y.; Fu, X. A New Strategy: Inhibitory Effect and Mechanism of Cymbopogon Martinii Essential Oil on Aspergillus Flavus. Food Biosci. 2023, 51, 102315. [Google Scholar] [CrossRef]
- Almshawit, H.; Macreadie, I. Fungicidal Effect of Thymoquinone Involves Generation of Oxidative Stress in Candida Glabrata. Microbiol. Res. 2017, 195, 81–88. [Google Scholar] [CrossRef]
- Čáp, M.; Váchová, L.; Palková, Z. Reactive Oxygen Species in the Signaling and Adaptation of Multicellular Microbial Communities. Oxid. Med. Cell. Longev. 2012, 2012. [Google Scholar] [CrossRef]
- Perrone, G.G.; Tan, S.X.; Dawes, I.W. Reactive Oxygen Species and Yeast Apoptosis. Biochim. Biophys. Acta - Mol. Cell Res. 2008, 1783, 1354–1368. [Google Scholar] [CrossRef]
- Sun, Q.; Zhou, M.; Zuo, Z. Toxic Mechanism of Eucalyptol and $β$-Cyclocitral on Chlamydomonas Reinhardtii by Inducing Programmed Cell Death. J. Hazard. Mater. 2020, 389, 121910. [Google Scholar] [CrossRef]
- Janssens, S.; Tinel, A. The PIDDosome, DNA-Damage-Induced Apoptosis and Beyond. Cell Death Differ. 2012, 19, 13–20. [Google Scholar] [CrossRef]
- A, T.; G, Z.; S, K.; R, K.; S, M.; R, P.; A, S.; B, S.; M, K.; K, K.; et al. Limonene Inhibits Candida Albicans Growth by Inducing Apoptosis. Med. Mycol. 2018, 56. [Google Scholar] [CrossRef]
Figure 1.
Fluorescence images showing membrane pore formation and their respective size in planktonic cells of C. krusei and C. parapsilosis. The control was DMSO-NaCl, treated with CpEO at 50 μg mL -1. Membrane pore formation was measured by propidium iodide (PI) uptake assay, and pore size used a 6 kDa dextran-FITC.
Figure 1.
Fluorescence images showing membrane pore formation and their respective size in planktonic cells of C. krusei and C. parapsilosis. The control was DMSO-NaCl, treated with CpEO at 50 μg mL -1. Membrane pore formation was measured by propidium iodide (PI) uptake assay, and pore size used a 6 kDa dextran-FITC.
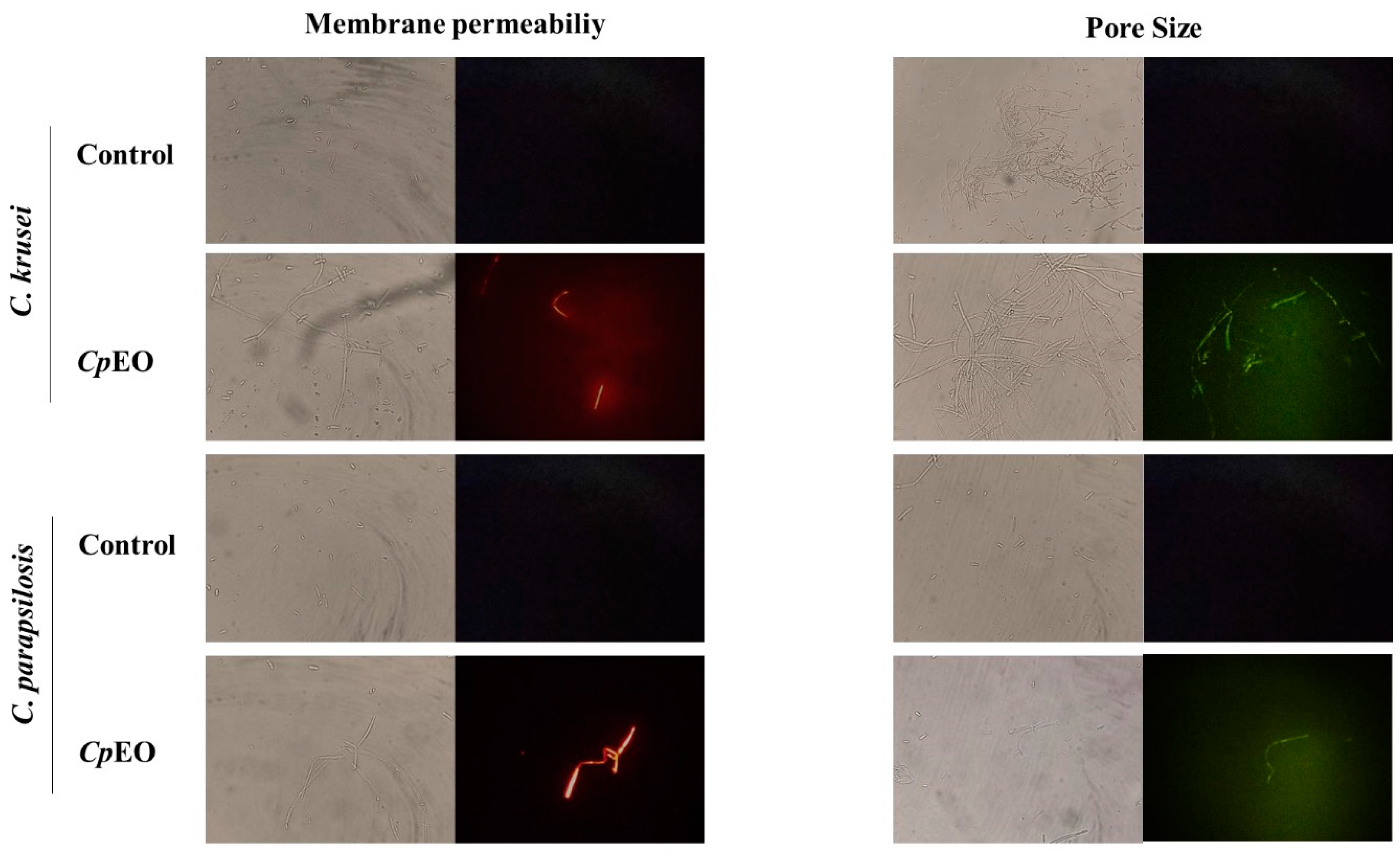
Figure 2.
Fluorescence images showing membrane pore formation and their respective size in biofilm cells of C. krusei and C. parapsilosis. Control used was DMSO-NaCl, treated with CpEO at 50 μg mL -1. Membrane pore formation was measured by propidium iodide (PI) uptake assay, and pore size used a 6 kDa dextran-FITC.
Figure 2.
Fluorescence images showing membrane pore formation and their respective size in biofilm cells of C. krusei and C. parapsilosis. Control used was DMSO-NaCl, treated with CpEO at 50 μg mL -1. Membrane pore formation was measured by propidium iodide (PI) uptake assay, and pore size used a 6 kDa dextran-FITC.
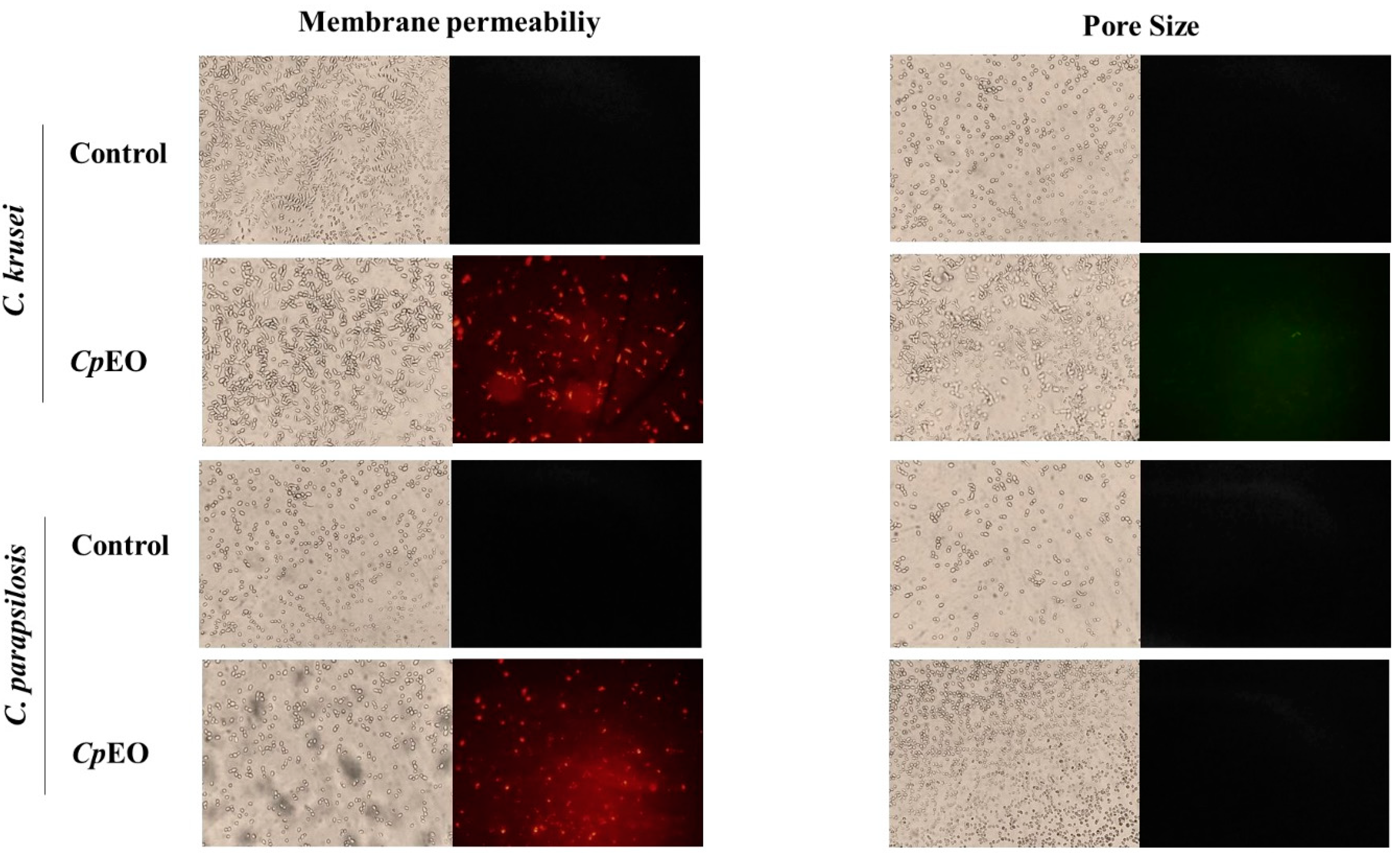
Figure 3.
fluorescence images showing overproduction of ROS in planktonic cells of C. krusei and C. parapsilosis. Control used was DMSO-NaCl, treated with CpEO at 50 μg mL-1.
Figure 3.
fluorescence images showing overproduction of ROS in planktonic cells of C. krusei and C. parapsilosis. Control used was DMSO-NaCl, treated with CpEO at 50 μg mL-1.
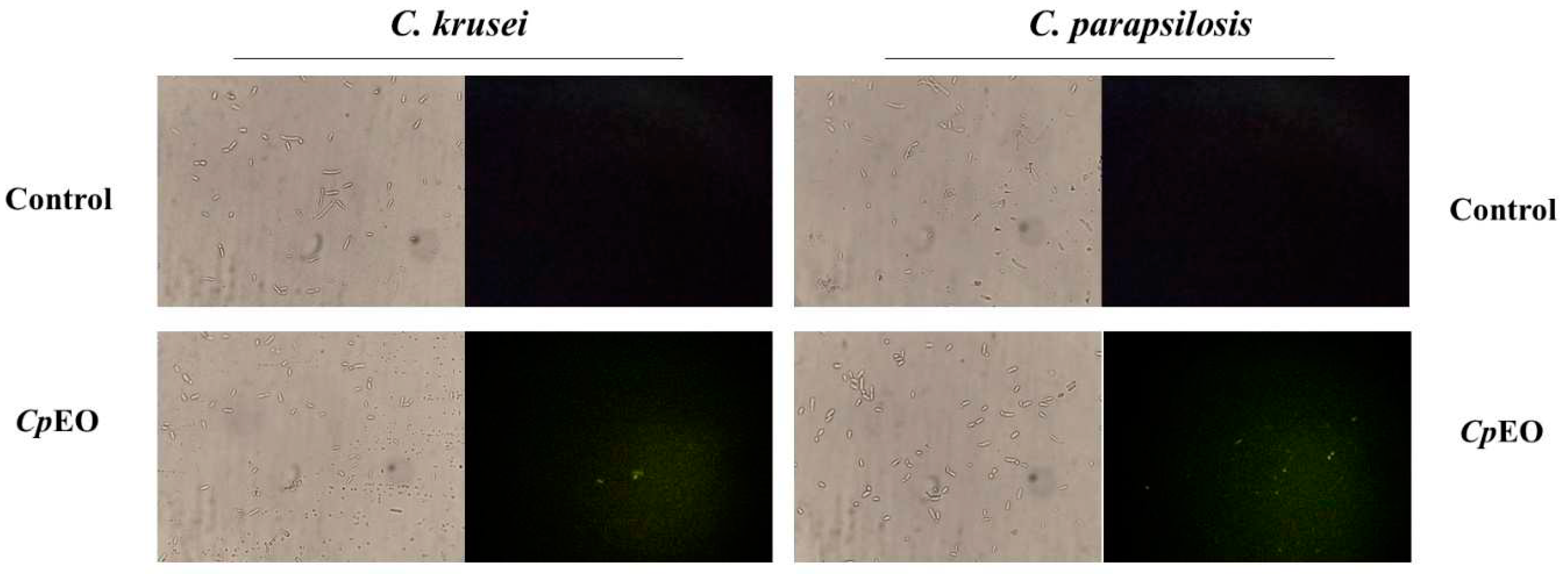
Figure 4.
Fluorescence images showing overproduction of ROS in biofilm cells of C. krusei and C. parapsilosis. Control used was DMSO-NaCl, treated with CpEO at 50 μg mL-1.
Figure 4.
Fluorescence images showing overproduction of ROS in biofilm cells of C. krusei and C. parapsilosis. Control used was DMSO-NaCl, treated with CpEO at 50 μg mL-1.
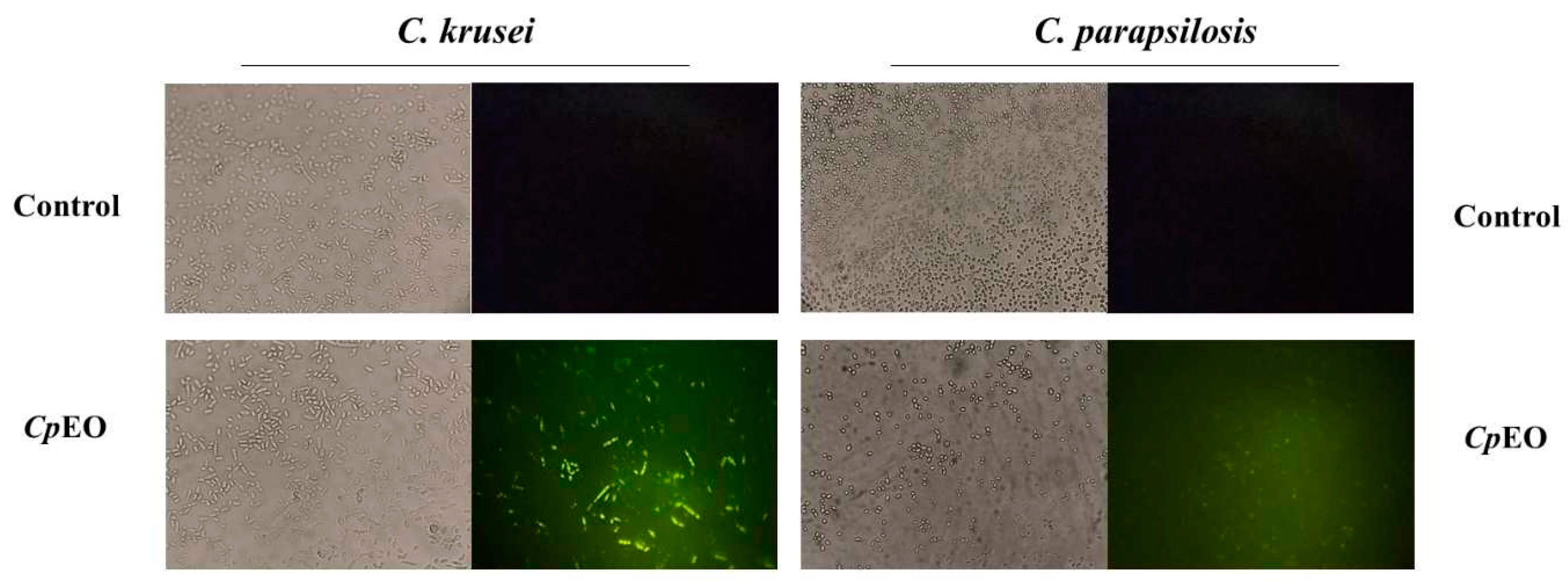
Figure 5.
Induction of apoptosis in planktonic cells of C. krusei and C. parapsilosis using CpEO at concentration 50 μg mL -1.
Figure 5.
Induction of apoptosis in planktonic cells of C. krusei and C. parapsilosis using CpEO at concentration 50 μg mL -1.

Figure 6.
Induction of apoptosis in biofilm cells of C. krusei and C. parapsilosis using CpEO at concentration 50 μg mL -1.
Figure 6.
Induction of apoptosis in biofilm cells of C. krusei and C. parapsilosis using CpEO at concentration 50 μg mL -1.

Figure 7.
Scanning electron microscopy of C. krusei cells treated with CpEO. (A) Control C. krusei cells treated with DMSO-NaCl solution. (B to F) C. krusei cells treated with CpEO. White arrows show damage to cell structure.
Figure 7.
Scanning electron microscopy of C. krusei cells treated with CpEO. (A) Control C. krusei cells treated with DMSO-NaCl solution. (B to F) C. krusei cells treated with CpEO. White arrows show damage to cell structure.
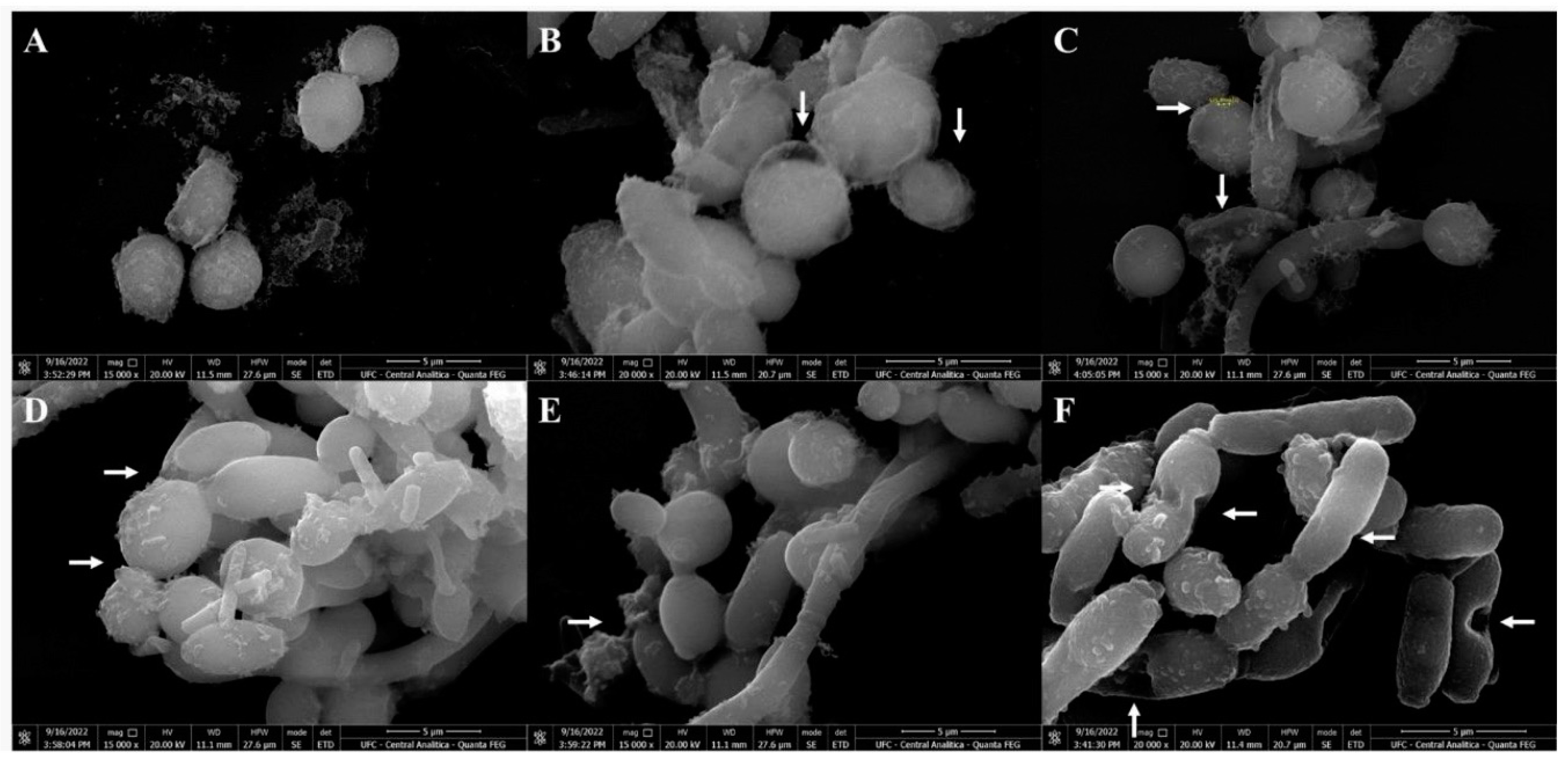
Figure 8.
Scanning electron microscopy of C. parapsilosis cells treated with CpEO. (A) Control C. parapsilosis cells treated with DMSO-NaCl solution. (B to F) C. parapsilosis cells treated with CpEO. White arrows show damage to cell structure.
Figure 8.
Scanning electron microscopy of C. parapsilosis cells treated with CpEO. (A) Control C. parapsilosis cells treated with DMSO-NaCl solution. (B to F) C. parapsilosis cells treated with CpEO. White arrows show damage to cell structure.
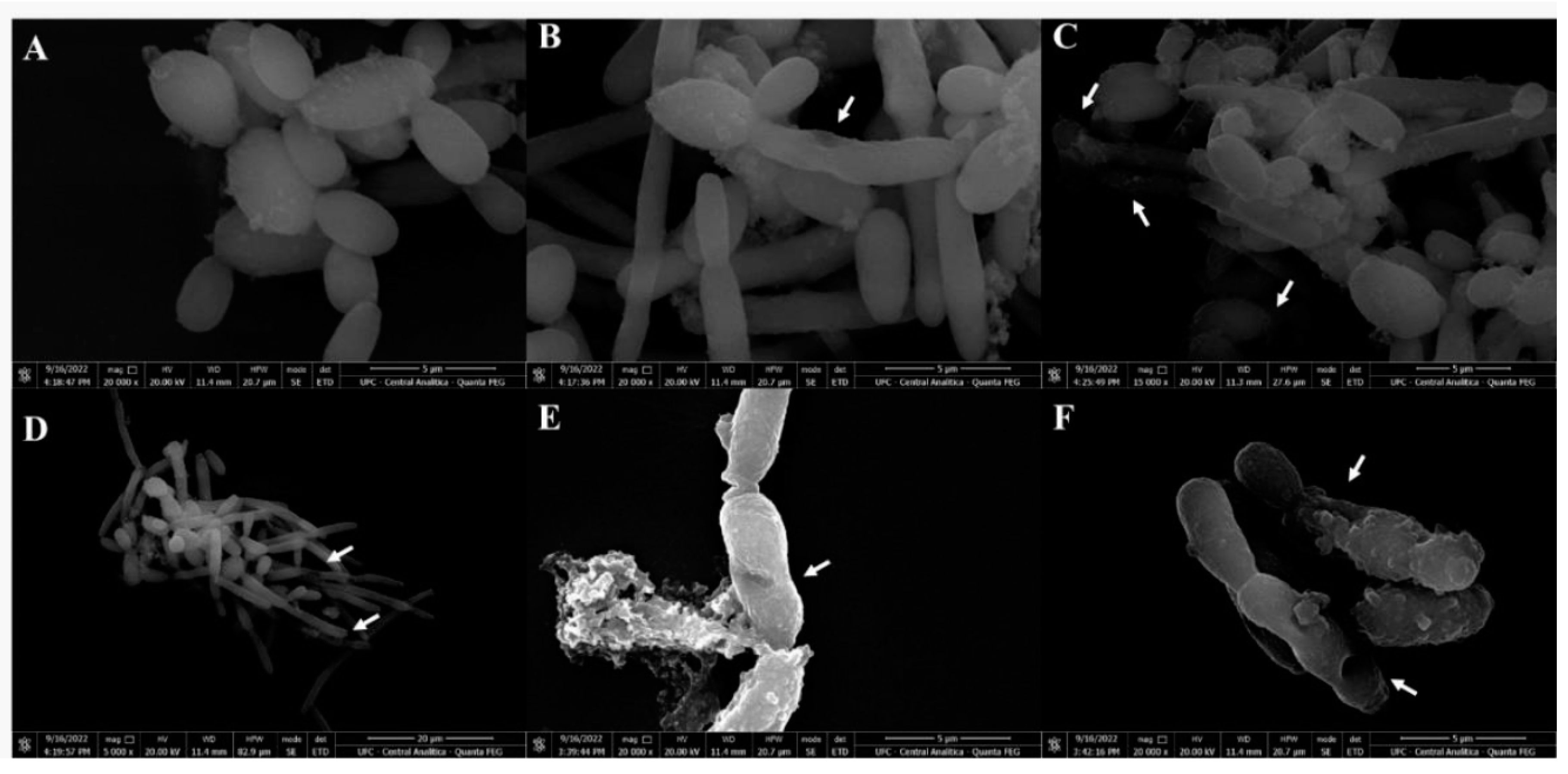

Table 1.
Main chemical components of C. pluriglandulosus essential oil by GC-MS analysis.
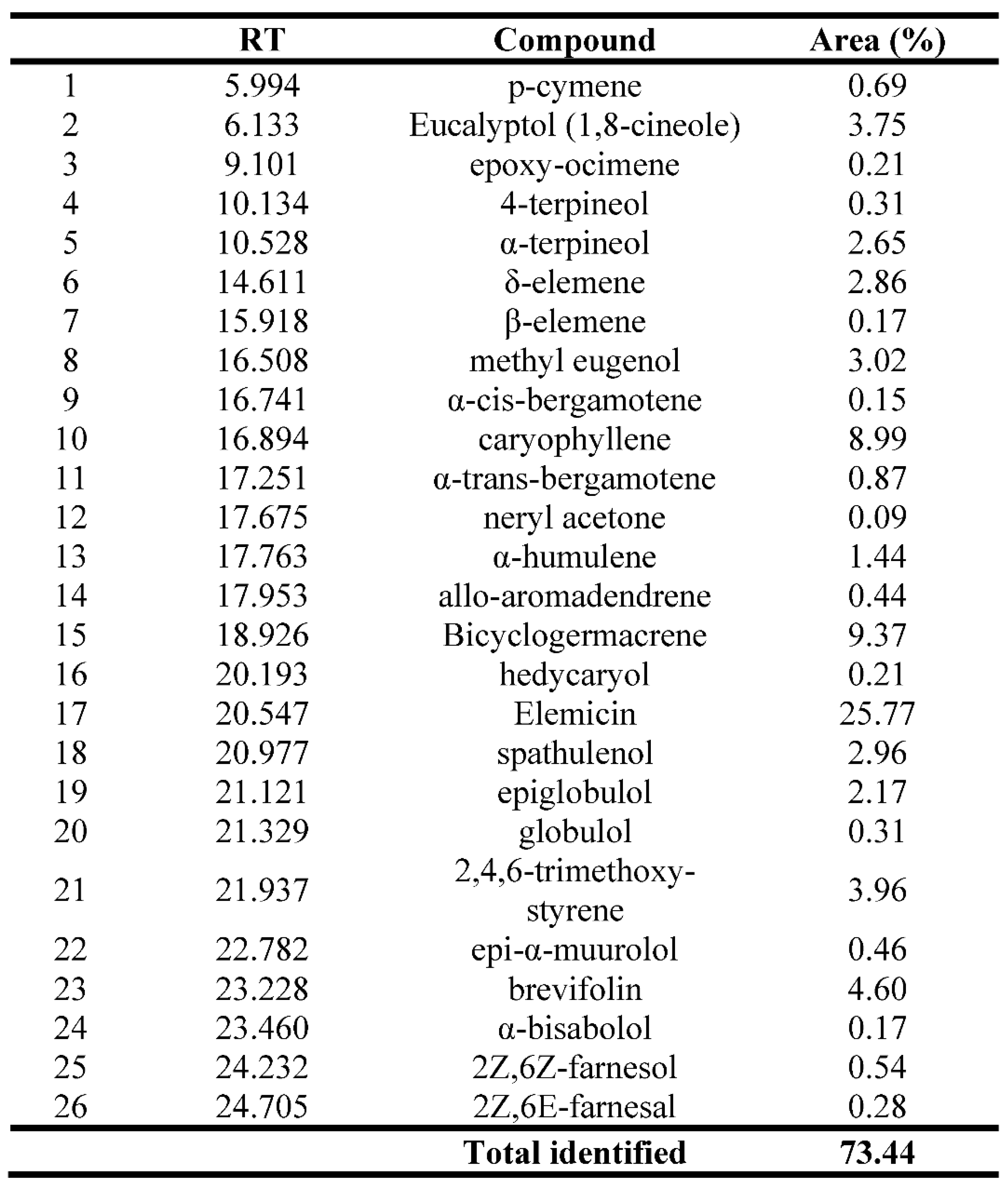
RT - Retention time (min); Area (%) - Relative area percentage.
Table 2.
Antimicrobial effects of C. pluriglandulosus Carn. essential oil against planktonic cells and
biofilm.
Table 2.
Antimicrobial effects of C. pluriglandulosus Carn. essential oil against planktonic cells and
biofilm.
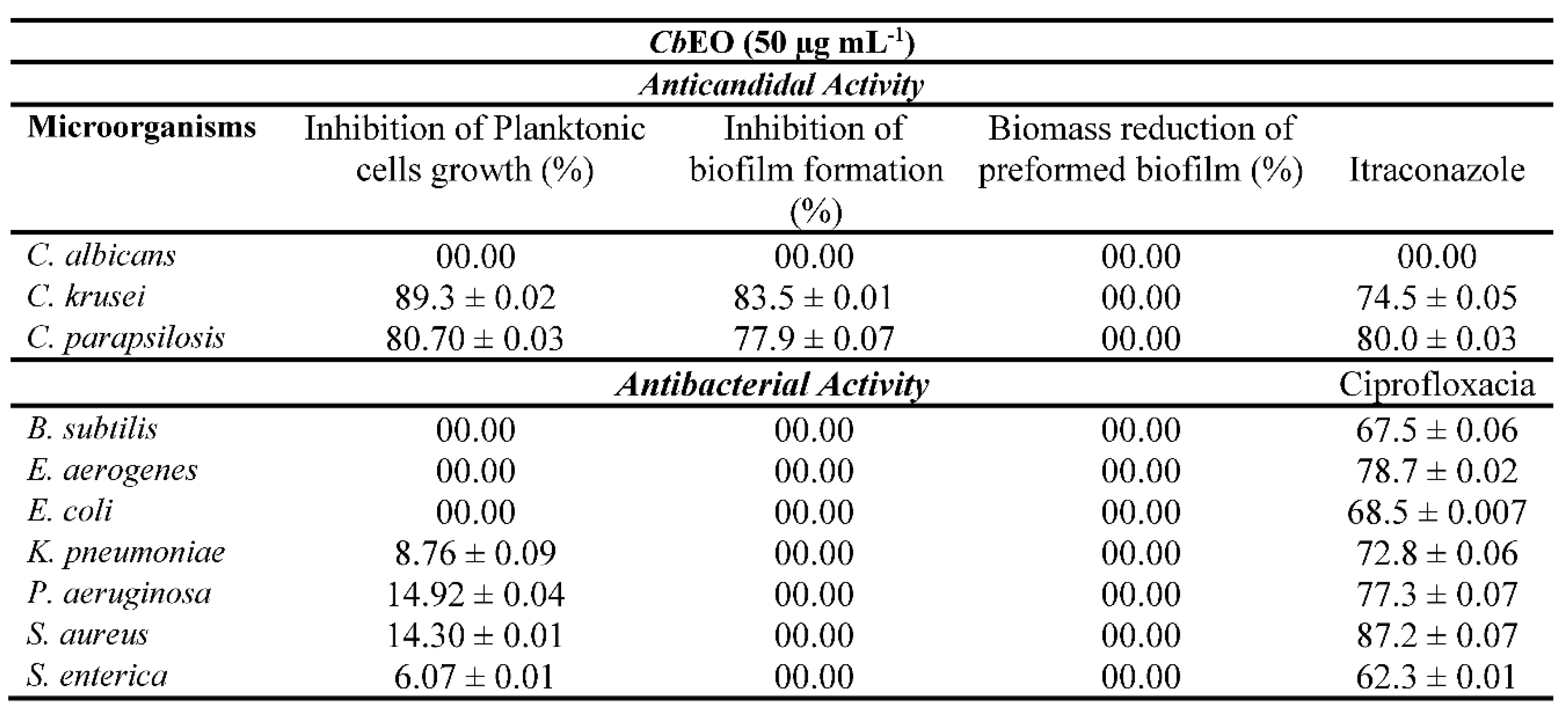
Table 3.
Hemolytic activity of CpEO Against human red Blood cells.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



