
Submitted:
08 May 2023
Posted:
09 May 2023
You are already at the latest version
Abstract
Saccharomyces cerevisiae (S. cerevisiae) ethanol fermentation has left an indelible trace in human history. It is regarded as the original beverage synthesis, and has developed into an essential energy supply for mankind today. However, the history of ethanol synthesis using S. cerevisiae, including the mechanism analysis process, has not been carefully and succinctly reported. In this paper, we review the history of ethanol synthesis using S. cerevisiae. In addition, we also review the substrates that drive S. cerevisiae ethanol synthesis, including the utilization of glucose, sucrose, starch and molasses, in order to summarize the current development status and to contribute to the further improvement of this field.
Keywords:
Saccharomyces cerevisiae
; Ethanol biosynthesis
; Material metabolism
1. Introduction
Ethanol has been used as a human beverage for thousands of years, and it has been recorded in the development history of the ancient civilizations of the world. The early use of ethanol may be attributed to its analgesic, disinfection, and spiritual effects on weddings, funerals, military expeditions, and the healing of patients. With the deepening of people's understanding of microorganisms, ethanol, as the product of microbial fermentation, has become one of the oldest forms reflecting the application of microbial biotechnology[1]. Today, the use of ethanol is no longer limited to the beverage; the high octane number of ethanol means that can be mixed with existing fuel sources and is an important part of the solution to the energy crisis and environmental problems[2].
Ethanol fermentation is a biological process performed by Saccharomyces cerevisiae (S. cerevisiae), one of the first model organisms to be studied scientifically, but the discovery of this process took generations of scientists (Figure 1). Ethanol fermentation was first studied in 1789 by the chemist Antoine Levoisier, who worked out the equation for how sucrose was converted to ethanol: grape must =carbonic acid + alcohol[3]. However, it was not until 1843 that the exact equation ‘C6H12O6→2C2H5OH+2CO2’ was calculated by Dumas. Until then, S. cerevisiae was not considered a living organism. It was only through the introduction and use of microscopic equipment that the yeast cell was described by Schwann in 1837[3], and the ‘cell theory’ that living structures originate from the formation and differentiation of units was subsequently proposed. With the development and maturation of the ethanol fermentation industry, the corresponding research became more in-depth. Between 1855 and 1875, Pasteur responded explicitly to the identity and role of yeast and proposed that fermentation was a physiological phenomenon that could be divided into aerobic and anaerobic forms. In 1857, Pasteur published his first paper on alcoholic fermentation, explaining that the breakdown of sugars into alcohol and carbonic acid was associated with life processes, and that sugars provided nutrients for yeast growth. Three years later, Pasteur reported a figure for the efficiency of ethanol fermentation, stating that only 95% of the sugar was converted to ethanol, while the remaining 5% was converted to byproducts such as glycerol, succinic acid, and ‘cellulose’[4]. In the same year, Berthelot proposed that S. cerevisiae used hydrolase to decompose sucrose into monosaccharides. The mechanism of ethanol synthesis in S. cerevisiae has been continuously explored, which has promoted people's concern for microbial biotechnology applications. Emphasis has been placed on enhancing the development of S. cerevisiae biosynthesis at the genetic manipulation level. It was not until 1996 that the whole genome sequencing of S. cerevisiae was completed, which opened the prelude to the molecular level research of S. cerevisiae[5].
S. cerevisiae is a well-studied model that has facilitated understanding of eukaryotic processes, and it was the first eukaryotic genome to be fully sequenced[5]. However, S. cerevisiae is not uniformly domesticated; strains vary in performance in different regions and have their own adaptability to each environment. With the increasing demand of people for material culture, as well as the demand for environmental protection, means there are increasing requirements on the amount of bioethanol synthesis. Therefore, the production capacity of biosynthetic ethanol will need to be greatly improved. Numerous strategies are proposed to facilitate an increase in bioethanol production, including: 1) using high-quality and cheap substrates such as starch, cellulose, and molasses; 2) improving the fermentation process and reducing energy consumption, such as through ethanol distillation; 3) obtaining robust strains to improve production performance, including speed and quality; and 4) achieving high-density fermentation. The advancement of each of these strategies is a huge boost to ethanol biosynthesis. In the U.S.A., 199 bioethanol synthesis plants have been built (up to 2022, https://www.biofuelsdigest.com/bdigest/) in 25 states, with a total capacity of 53.6 million tons per year, and the U.S. government continues to increase funding for this technology.
Despite the increasing demand and investment, ethanol fermentation still faces multiple challenges at present (Figure 2). Ethanol synthesis by S. cerevisiae can be conducted in different regions and different environments, and can be achieved through the degradation of different polysaccharides. However, the efficacy of the wild-type strain itself is often unable to meet the needs of the production design, so it faces a variety of stress factors, as follows. 1) Influence of high temperature, including biological metabolism heat production, mechanical stirring heat production, and regional high temperature climate and other factors[6,7]. 2) The effect of low temperature. For example, after the strain cryogenic preservation process, performance of the fermentation strain needs to be maintained[8,9,10]. 3) The effect of nutrient deficiency. Glucose and sucrose are the best carbon sources for fermentation and are used preferentially when other carbon sources are present. However, sudden depletion of glucose and sucrose presents a precipice of carbon metabolism and utilization (After the best carbon source is used, the carbon source that is used later will not be used immediately), and then utilization of other carbon sources commences[11]. 4) Influence of high osmotic pressure. Increasing the substrate concentration has become one of the most direct means to improve the degree of fermented wine and reduce the energy consumption of subsequent distillation[12,13]. 5) Influence of low osmotic pressure. For that need to go through the washing step in the process of seed solution cultivation, multiple infusions of water will cause cell hypoosmosis and cell swelling[10]. 6) Influence of ethanol accumulation. Studies have shown that most microbial growth is affected by 2% (v/v) ethanol concentration[6]. 7) The effect of reactive oxygen species (ROS). Various adverse factors in the fermentation process can aggravate the accumulation of ROS in cells, resulting in lipid oxidation of the cell membrane and nucleic acid damage[14,15]. 8) Other influences. Additional adverse factors in the substrate environment include the high number of ions in molasses and the influence of CO2 on culture environment.
Improving understanding of S. cerevisiae and the potential problems of the fermentation environment will enhance the efficiency of ethanol biosynthesis. The following sections focus on current ethanol synthesis processes using glucose, sucrose, starch, and molasses as substrates, and summarize the factors known to affect production.
2. Ethanol Fermentation with Saccharomyces Cerevisiae
2.1. Ethanol Fermentation Based on Glucose and Sucrose
The best carbon source for ethanol fermentation by S. cerevisiae is glucose or sucrose. Increasing the concentration of such substrates is one approach to achieve a high concentration of ethanol to reduce subsequent distillation energy. However, with the increase of substrate concentration, the osmotic pressure of the culture environment rises, and hyperosmolality can cause volume shrinkage of cells, which stresses the growth and vitality of cells[12,13]. In order to overcome the influence of high osmotic pressure and high ethanol concentration feedback inhibition, ethanol recovery with plate membrane pervaporation device[16], extraction fermentation[17], electrostatic fermentation[18], ultrasonic fermentation[19], immobilized fermentation[20], multi-stage reaction tower recycling fermentation[21,22,23] and so on have been widely studied (Table 1). But when the carbon source is used by S. cerevisiae to make ethanol, not only do these problems of high osmotic pressure and ethanol feedback inhibition occur, when multiple carbon sources are present, glucose and sucrose are always consumed first. The presence of these sugars inhibits gluconeogenesis, glyoxylic acid circulation, respiration (known as the ‘Crabtree effect’), and the intake of fewer carbohydrates. This pathway inhibition by glucose ensures that preferred sugars are metabolized before consumption of replacement carbohydrates such as maltose and galactose. However, this process can have two serious consequences. 1) Prolonged use of glucose and sucrose can result in yeast cells reducing their ability to metabolize other sugars. 2) The catabolic inhibition caused by glucose does not begin immediately with the depletion of glucose; instead, there is a lag period that causes a marked drop in carbon metabolism and affects growth[11].
2.2. Ethanol Fermentation Based on Starch
Starch is a cheap and renewable carbon source[30] composed of a linear chain of α-1, 4-linked D-glucopyranose units (amylose) and the highly branched amylopectin fraction that consists of α-1,4- and α-1,6-linked D-glucopyranose units. S. cerevisiae cannot degrade starch naturally. Starch has to be hydrolyzed through liquefaction and saccharification before use, and efficient starch hydrolysis requires both an α-amylase and a glucoamylase together with an α-1,6-debranching activity[31]. α-Amylase randomly cleaves the α-1,4-glycosidic linkages inside the starch granule and degrades the whole structure to release non-reducing ends. Subsequently, glucoamylase cleaves α-1,4- and α-1,6-glycosidic linkages from the non-reducing ends. Considering that oligosaccharides produced during decomposition can inhibit α-amylase, it is necessary to adjust the ratio of both enzymes during use; the addition of pullulanases (which hydrolyze the α-1,6-glycosidic bonds) can also greatly improve the efficiency of starch hydrolysis[32]. Industrial production of sugars from starch usually consists of four steps—jet-cooking, addition of α-amylase, cooling (regulating pH), and addition of glucoamylase—to yield sugar streams that can be used for ethanol fermentation[33]. The boiling step can consume a lot of energy—approximately 10–20% of the bioethanol price, with further damage caused by the Maillard reaction. Consequently, some factories have changed the 'boiling process' to a 'cold process' by adding raw starch-degrading enzymes, saving approximately 40–50% of the total cost[34,35]. In addition, pretreatment of starch[25] or co-culture with bacteria with starch decomposition ability[24] has also become a feasible research content (Table 1).
The demand for energy is increasing worldwide—global energy consumption is predicted to rise by more than 30% between 2018 and 2050, while bioenergy would increase by 3% per year during that time[36]. This increasing energy demand and the emergence of the global food crisis is driving the use of high-starch plant substitutes for ethanol production. Cassava (72.6–76.6% starch for dried cassava) is a very attractive raw material for bioethanol production that is not affected by food and feed shortage concerns[37]. A repeated batch fermentation of cassava yielded an ethanol concentration of more than 83.64 g/L[38]. The small aquatic plant duckweed (35.7% amylose and 64.3% amylopectin[39]) is another promising source for bioethanol production. Duckweed could be hydrolyzed with α-amylase and amyloglucosidase (2:1, by vol.), with the final ethanol concentration reaching 0.19 g ethanol/g dry biomass[36].
Starch needs to be hydrolyzed before it can be used by S. cerevisiae, and although progress has been made to make the process more efficient, adding enzymes and other reagents ultimately increases the cost of the process. Therefore, development of a recombinant S. cerevisiae that co-expresses the required glucoamylase and a-amylase has been explored. Many genes from filamentous fungi and yeasts that encode amylolytic enzymes have been expressed in S. cerevisiae[40,41,42]; However, only a limited number of fungal and bacterial strains meet the standard for commercial amylase production. A major challenge is in the simultaneous production of these amylolytic enzymes that exhibit high substrate affinity and specific enzyme activity[43]. A recombinant S. cerevisiae combined with exogenous enzymes to promote starch hydrolysis for ethanol fermentation has been engineered and its carbon source conversion rate can reach 94%[44]. The rapid development of synthetic biology may direct further developments of recombinant strains of S. cerevisiae for ethanol fermentation using starch, but this also needs to be approved for use in the corresponding part (Food regulatory agency et al).
2.3. Ethanol Fermentation Based on Molasses
No matter how volatile the world is, humans cannot live without food and energy. Consequently, the development of non-food resources for energy supply is needed for robust and sustainable global energy, and strengthening the use of molasses is an important component of this process. Molasses is a by-product of the sugar industry (3 tons of sugar produces approximately 1 ton of molasses) that is generated during sugarcane and sugar beet processing. It contains 30–60% (w/v) sugar, plus colloids and a large number of metal ions[27,45,46].
Molasses generally requires a pretreatment process before it can be used directly by S. cerevisiae. This pretreatment usually involves boiling, adding concentrated sulfuric acid to adjust the pH, cooling overnight, reheating and adding activated carbon, centrifugation, and adding hydrated lime. Despite the short pretreatment process, according to Professor Abbott's 'alert level' theory[47], there are still many factors in molasses that can stress the growth and vitality of cells. To address this issue, scholars have used semi-continuous (feed) fermentation [48,49], ceramic microfiltration[27], adsorption column[50] or dialysis ultrafiltration[51] to reduce the influence of multiple stresses during molasses fermentation. In addition, immobilization methods for yeast, such as calcium alginate[52], bagasse[53], and thin-shell silk cocoon[28](Table1), have been employed to increase the activity of yeast cells. The alcohol content in the mash made by yeast cells using molasses was improved to a certain extent by these modifications. However, the inherent genetic properties of yeast cells mean the growth and vitality of yeast cells are still limited under various stress pressures. Therefore, the research of gene editing for genetic properties to improve production has become a development, such as ethanol regulating gene replacement[29] and so on. Now, ethanol fermentation with high concentrations of molasses remains a challenge.
To overcome the genetic limitations of S. cerevisiae, the resistant strains of S. cerevisiae based on the sucrose addition model have been studied and the tolerance of these yeast cells to ethanol, osmotic pressure, and oxidative stress has been reported[54,55]. However, the research on ethanol fermentation with high concentration molasses has had little effect. A careful taste reveals that ethanol fermentation using molasses has a lower degree of fermented liquor in the limos compared with the fermentation using sucrose, which means that ethanol feedback inhibition is not the main factor affecting ethanol fermentation from molasses. Similarly, the osmotic pressure formed at high sucrose concentration did not cause lethal effects on S. cerevisiae. Therefore, identifying why S. cerevisiae struggles to use high concentration molasses for ethanol fermentation is crucial for future research. Our unpublished research indicates that the influence of K+ and Ca2+ in molasses may be a major cause of this difficulty (Date will be published in another paper). However, there is a current lack of detailed reports on the influence of ions on molasses in specific environments. Therefore, it will be an effective method to reveal the stress factors with robust strains of S. cerevisiae cultured under the environment of molasses fermentation for specific industrial production and combined with multi-omics analysis.
3. Conclusions
S. cerevisiae ethanol fermentation has made a great contribution to the development of bioenergy by humans. S. cerevisiae is a living, intelligent organism, and if we want to direct its efficient biosynthesis with human will, we must understand S. cerevisiae itself and its current production predicaments. Glucose and sucrose are the best carbon sources for S. cerevisiae. There is potential for ethanol synthesis based on starch and molasses as substrates, as well as fermentation based on cellulose. However, these substrates impose various restrictive factors on the growth and metabolism of S. cerevisiae Domestication of wild-type strains of S. cerevisiae takes a long time and there are many uncertain factors. However, the development of synthetic biology is anticipated to facilitate the directional construction of a new generation of engineered S. cerevisiae that may balance the current situation of ethanol production. The caveat is that identification of the accurate target site and target direction is needed to achieve this approach.
Author Contributions
WYW, HSB and HSS sorted out the related materials, LL revised the paper. CYL and FXN wrote the paper and collated the remaining materials. All the authors read and approved the final manuscript.
Funding
This work was funded by the central government guides the special fund for local science and technology development (No. ZY22096007), the National Natural Science Foundation of China (Grant No. 32260246), Guangxi Science and technology base and talents special project (No. AD22080011), Guangdong Basic and Applied Basic Research Foundation (No. 2019A1515110381), and the doctoral fund of Guangxi university of science and technology (No. 21Z50).
Acknowledgements
Not applicable.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Availability of data and materials
Not applicable.
Ethics approval and consent to participate
Not applicable.
References
- Samuel D. Investigation of ancient egyptian baking and brewing methods by correlative microscopy. Science 1996;273(5274):488-490. [CrossRef]
- Wang WY. Wu SH. Xie YH. Zhong M. Wei ML. Li ZY. Long XF. Niu FX. A high-throughput screening procedure (Py-Fe3+) for enhancing ethanol production by Saccharomyces cerevisiae using ARTP random mutagenesis. Processes. 2022, 10, 2186. [CrossRef]
- Barnett JA. Beginnings of microbiology and biochemistry: the contribution of yeast research. Microbiology 2003;149(3):557-567. [CrossRef]
- Lanska DJ. Encyclopedia of the neurological sciences. 2014;846-847. [CrossRef]
- Poethig RS. Life with 25000 genes. Genome Res 2001;11(3):313-316. [CrossRef]
- Marullo P, Durrens P, Peltier E, Bernard M, Dubourdieu D. Natural allelic variations of Saccharomyces cerevisiae impact stuck fermentation due to the combined effect of ethanol and temperature; a QTL-mapping study. BMC Genom 2019;20(1):680. [CrossRef]
- Khatun MM, Yu X, Kondo A, Bai F, Zhao X. Improved ethanol production at high temperature by consolidated bioprocessing using Saccharomyces cerevisiae strain engineered with artificial zinc finger protein. Bioresour Technol 2017;245:1447-1454. [CrossRef]
- Lambrechts M, Pretorius I. Yeast and its importance to wine aroma - a review. South African J. Enol Vitic 2000;21(1). [CrossRef]
- Park JI, Grant CM, Davies MJ, Dawes IW. The cytoplasmic Cu,Zn superoxide dismutase of Saccharomyces cerevisiae is required for resistance to freeze-thaw stress. Generation of free radicals during freezing and thawing. J Biol Chem 1998;273(36):22921-22928. [CrossRef]
- Diniz-Mendes L, Bernardes E, de Araujo PS, Panek AD, Paschoalin VM. Preservation of frozen yeast cells by trehalose. Biotechnol Bioeng 1999;65(5):572-578. [CrossRef]
- Verstrepen KJ, Iserentant D, Malcorps P, Derdelinckx G, Dijck PV, Winderickx J, Pretorius IS, Thevelein JM, Delvaux FR. Glucose and sucrose: hazardous fast-food for industrial yeast? Trends Biotechnol 2004;22(10):531-537. [CrossRef]
- Izawa S, Ikeda K, Miki T, Wakai Y, Inoue Y. Vacuolar morphology of Saccharomyces cerevisiae during the process of wine making and japanese sake brewing. Appl Microbio Biotechnol 2010;88(1):277-282. [CrossRef]
- Perrier-Cornet JM, Hayert M, Saurat E, Milesse C, Gervais P. Effect of osmotic stress on high pressure inactivation of Saccharomyces cerevisiae. High Press Biosci Biotechnol 1999;27-30. [CrossRef]
- Qin L, Dong S, Yu J, Ning X, Xu K, Zhang S-J, Xu L, Li B-Z, Li J, Yuan Y-J, Li C. Stress-driven dynamic regulation of multiple tolerance genes improves robustness and productive capacity of Saccharomyces cerevisiae in industrial lignocellulose fermentation. Metab Eng 2020;61:160-170. [CrossRef]
- Treu L, Campanaro S, Nadai C, Toniolo C, Nardi T, Giacomini A, Valle G, Blondin B, Corich V. Oxidative stress response and nitrogen utilization are strongly variable in Saccharomyces cerevisiae wine strains with different fermentation performances. Appl Microbiol Biotechnol 2014;98(9):4119-4135. [CrossRef]
- O'Brien D, Craig J. Ethanol production in a continuous fermentation/membrane pervaporation system. Appl Microbiol Biotechnol 1996;44:699-704. [CrossRef]
- Daugulis AJ, Axford DB, Ciszek B, Malinowski JJ. Continuous fermentation of high-strength glucose feeds to ethanol. Biotechnol Lett 1994;16:637-642. [CrossRef]
- Mathew AS, Wang J, Luo J, Yau ST. Enhanced ethanol production via electrostatically accelerated fermentation of glucose using Saccharomyces cerevisiae. Sci Rep 2015;5:15713. [CrossRef]
- Indra Neel P, Gedanken A, Schwarz R, Sendersky E. Mild sonication accelerates ethanol production by yeast fermentation. Energy Fuels 2012;26(4):2352-2356. [CrossRef]
- Kogtimas A, Gourdoupis C, Psarianos C. Kaliafas, A.; Kanellaki, M. Continuous potable alcohol production by immobilized Saccharomyces cerevisiae on mineral kissiris. Appl Biochem Biotechnol 1991;30:203-216. [CrossRef]
- Pacheco TF, Galvão de Morais Júnior W, Zanella Guidini C, Marquez LDS, Cardoso VL, Resende MM, Ribeiro EJ. Alcoholic Fermentation with Self-Flocculating Yeast in a Tower Upflow Reactor. Chem Eng Technol 2015;38(2):345-354. [CrossRef]
- Santos LD, Sousa MDB, Guidini CZ, De Resende M. M, Cardoso VL, Ribeiro EJ. Continuous ethanol fermentation in tower reactors with cell recycling using flocculent Saccharomyces cerevisiae. Process Biochem 2015;50(11):1725-1729. [CrossRef]
- Breisha, GZ. Production of 16% ethanol from 35% sucrose. Biomass Bioenergy 2010;34(8):1243-1249. [CrossRef]
- Abouzied MM, Reddy CA. Fermentation of starch to ethanol by a complementary mixture of an amylolytic yeast and Saccharomyces cerevisiae. Biotechnol Lett 1987;9:59-62. [CrossRef]
- Shin YC, Lee SY, Choe YK, Kim HS, Byunt SM. Ethanol fermentation of cassava starch pretreated with alkali. Biotechnol Bioeng. 1986;28(4):627-630. [CrossRef]
- Ping W, Singh V, Hua X, Johnston DB, Tumbleson ME. Comparison of raw starch hydrolyzing enzyme with conventional liquefaction and saccharification enzymes in dry-grind corn processing. Cereal Chem 2007;84(1):10-14. [CrossRef]
- Kaseno, Kokugan T. The effect of molasses pretreatment by ceramic microfiltration membrane on ethanol fermentation. J Biosci Bioeng 1997;83(6):577-582. [CrossRef]
- Rattanapan A, Limtong S, Phisalaphong M. Ethanol production by repeated batch and continuous fermentations of blackstrap molasses using immobilized yeast cells on thin-shell silk cocoons. Appl Energy 2011;88(12):4400-4404. [CrossRef]
- Wu R, Chen D, Cao S, Lu Z, Huang R. Enhanced ethanol production from sugarcane molasses by industrially engineered Saccharomyces cerevisiae via replacement of the PHO4 gene. RSC Adv 2020;10:2267-2276. [CrossRef]
- Javan F, Mobini-Dehkordi M. Application of alpha-amylase in biotechnology. J Biol 2012. [CrossRef]
- Knox AM, Preez J, Kilian SG. Starch fermentation characteristics of Saccharomyces cerevisiae strains transformed with amylase genes from Lipomyces kononenkoae and Saccharomycopsis fibuligera - ScienceDirect. Enzyme & Microbial Technology. 2004;34(5):453-460. [CrossRef]
- Wang X, Nie Y, Xu Y. Industrially produced pullulanases with thermostability: Discovery, engineering, and heterologous expression. Bioresour Technol 2019;278:360-371. [CrossRef]
- Bothast RJ, Schlicher MA. Biotechnological processes for conversion of corn into ethanol. Appl Microbiol Biotechnol 2005;67(1):19-25. [CrossRef]
- Robertson GH, Wong DWS, Lee CC, Wagschal K, Orts WJ. Native or raw starch digestion: a key step in energy efficient biorefining of grain. J Agric Food Chem 2006;54(2):353-365. [CrossRef]
- van Zyl WH, Bloom M, Viktor MJ. Engineering yeasts for raw starch conversion. Appl Microbiol Biotechnol 2012;95(6):1377-1388. [CrossRef]
- Af A, Aas A, Np B. Production of bioethanol from four species of duckweeds (Landoltia punctata, Lemna aequinoctialis, Spirodela polyrrhiza, and Wolffia arrhiza) through optimization of saccharification process and fermentation with Saccharomyces cerevisiae. Saudi J Biol Sci 2021;28(1):294-301. [CrossRef]
- Hu Z, Tan P, Pu G. Multi-objective optimization of cassava-based fuel ethanol used as an alternative automotive fuel in Guangxi, China. Appl Energy 2006;83(8):819-840. [CrossRef]
- Choi GW, Moon SK, Kang HW, Min J, Chung BW. Simultaneous saccharification and fermentation of sludge-containing cassava mash for batch and repeated batch production of bioethanol by Saccharomyces cerevisiae CHFY0321. J Chem Technol Biotechnol 2010;84(4):547-553. [CrossRef]
- Lee CJ, Yangcheng H, Cheng JJ, Jane J-l. Starch characterization and ethanol production of duckweed and corn kernel. Starke 2015;68(3-4):348-354. [CrossRef]
- Ülgen K Ö, Saygili B, Önsan Z.İ, Kirdar B. Bioconversion of starch into ethanol by a recombinant Saccharomyces cerevisiae strain YPG-AB. Process Biochem 2002;37(10):1157-1168. [CrossRef]
- Eksteen JM, Rensburg PV, Otero RRC, Pretorius IS. Starch fermentation by recombinant Saccharomyces cerevisiae strains expressing the alpha-amylase and glucoamylase genes from lipomyces kononenkoae and saccharomycopsis fibuligera. Biotechnol Bioeng 2003;84(6):639-646. [CrossRef]
- Nakamura Y, Kobayashi F, And MO, Sawada T. Alcohol fermentation of starch by a genetic recombinant yeast having glucoamylase activity. Biotechnol Bioeng 1997;53(1):21-25. [CrossRef]
- Haan RD, Kroukamp H, Mert M, Bloom M, Görgens J, Zyl W. Engineering Saccharomyces cerevisiae for next generation ethanol production. J Chem Technol Biotechnol 2013;88(6):983-991. [CrossRef]
- Cripwell RA, Rose SH, Favaro L, Zyl WHV. Construction of industrial Saccharomyces cerevisiae strains for the efficient consolidated bioprocessing of raw starch. Biotechnol Biofuels 2019;12:201. [CrossRef]
- Chotineeranat S, Wansuksri R, Piyachomkwan K, Chatakanonda P, Weerathaworn P, Sriroth K. Effect of calcium ions on ethanol production from molasses by Saccharomyces cerevisiae. Sugar Tech 2010;12:120-124. [CrossRef]
- Ergun, AT. Effect of zeolite nay and ca-montmorillonite on ethanol production using synthetic molasses. Appl Biochem Biotechnol 2008;144(2):161-168. [CrossRef]
- Abbott DA, Ingledew WM. Buffering capacity of whole corn mash alters concentrations of organic acids required to inhibit growth of Saccharomyces cerevisiae and ethanol production. Biotechnol Lett 2004;26(16):1313-1316. [CrossRef]
- Echegaray OF, Carvalho J, Fernandes A, Sato S, Aquarone E, Vitolo M. Fed-batch culture of Sacchoromyces cerevisiae in sugar-cane blackstrap molasses: invertase activity of intact cells in ethanol fermentation. Biomass Bioenergy 2000;19(1):39-50. [CrossRef]
- Borzani W, Hiss H, Santos T, Vairo M. Semicontinuous ethanol fermentation of sugar cane blackstrap molasses by pressed yeast. Biotechnol Lett 1992;14(10):981-984. [CrossRef]
- Hatano KI, Kikuchi S, Nakamura Y, Sakamoto H, Takigami M, Kojima Y. Novel strategy using an adsorbent-column chromatography for effective ethanol production from sugarcane or sugar beet molasses. Bioresour Technol 2009;100(20):4697-4703. [CrossRef]
- Johnson, R. Dialysis and ultrafiltration of molasses for fermentation enhancement. Sep Purif Technol 2001;22–23:239-245. [CrossRef]
- Ghorbani F, Younesi H, Sari A, Najafpour G. Cane molasses fermentation for continuous ethanol production in an immobilized cells reactor by Saccharomyces cerevisiae. Renew Energ 2011;36(2):503-509. [CrossRef]
- Babu NK, Satyanarayana B, Balakrishnan K, Rao TR, Rao GS. Study of sugarcane pieces as yeast supports for ethanol production from sugarcane juice and molasses using newly isolated yeast from toddy sap. Mycobiology 2012;40(1):35-41. [CrossRef]
- Qin L, Dong S, Yu J, Ning X, Li C. Stress-driven dynamic regulation of multiple tolerance genes improves robustness and productive capacity of Saccharomyces cerevisiae in industrial lignocellulose fermentation. Metab Eng 2020;61:160-170. [CrossRef]
- Shima J, Takagi H. Stress-tolerance of baker's-yeast (Saccharomyces cerevisiae) cells: stress-protective molecules and genes involved in stress tolerance. Biotechnol Appl Biochem 2009;53(3):155-164. [CrossRef]
Figure 1.
The cognitive history of S. cerevisiae ethanol fermentation.
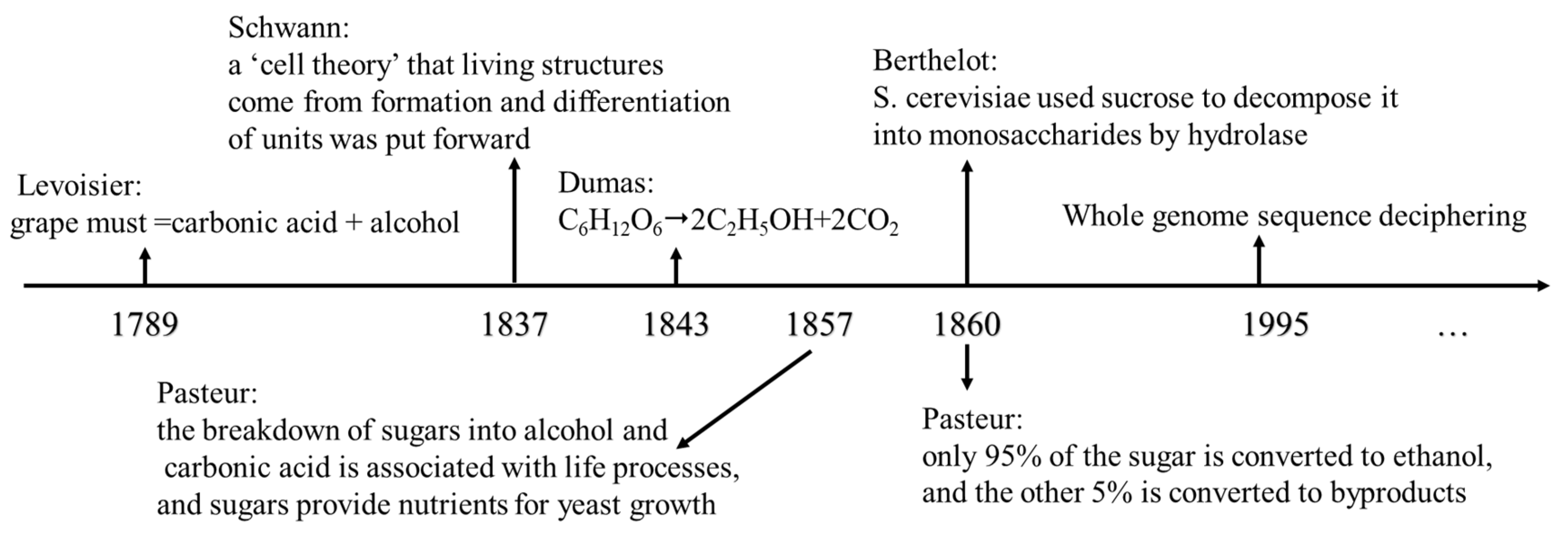
Figure 2.
Multiple challenges in ethanol synthesis of S. cerevisiae.
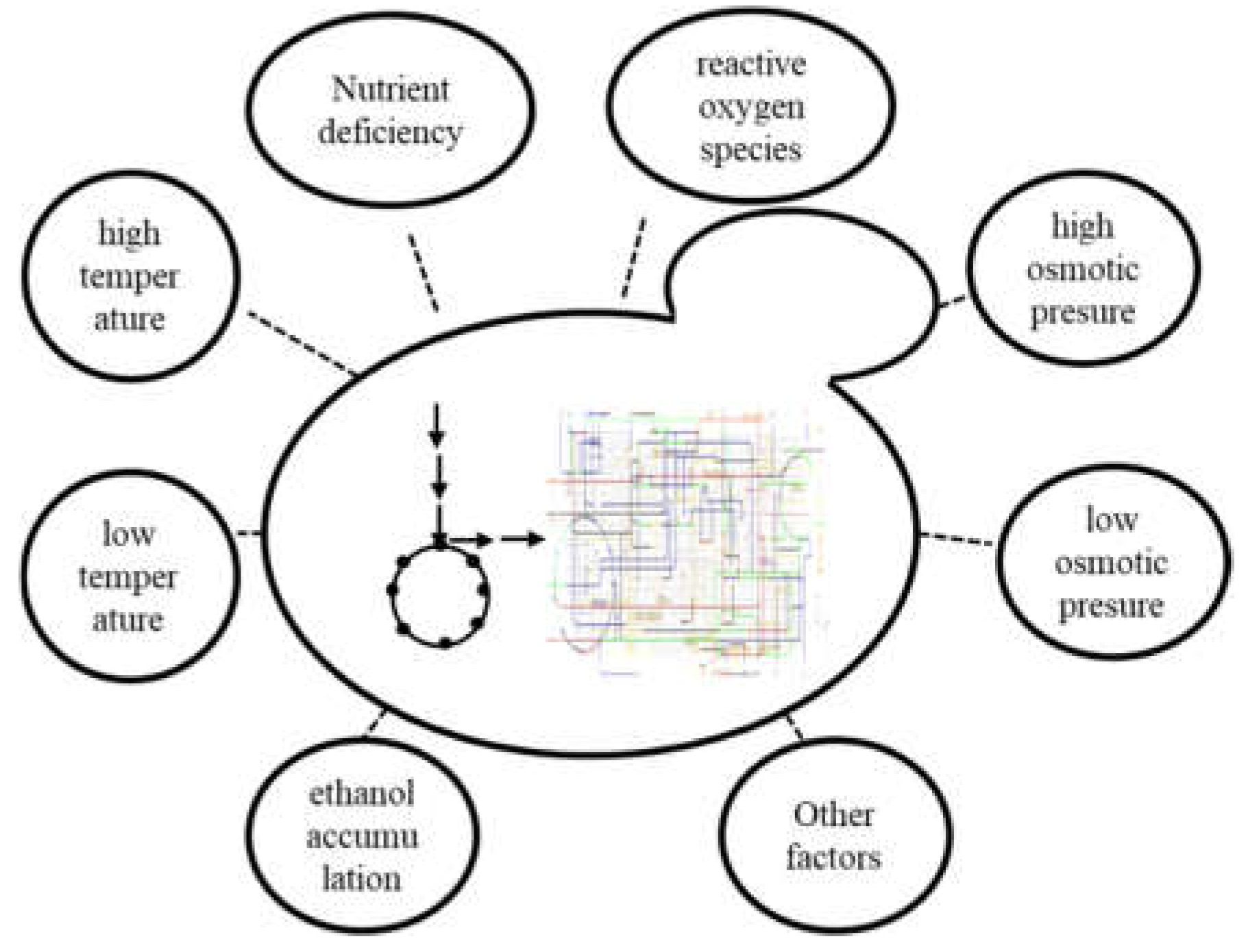

Table 1.
Ethanol fermentation based on different substrates.
| Substrate | Method | Achievement | Ref |
|---|---|---|---|
| Glucose | Coupled to a flat-plate membrane pervaporation unit to recover ethanol | ethanol productivities reached 7.8 g/L/h | [16] |
| Glucose | Extractive fermentation, aqueous feeds with 413 and 495 g/L glucose | Ethanol conversion efficiencies reached 90%-95% conversion | [17] |
| Glucose | electrostatic fermentation with 15 V | The consumption of glucose reached 98% in 20 h | [18] |
| Glucose | Fermentation with mild ultrasonication | 10 times faster than non-stirred fermentation. | [19] |
| sucrose | Continuous potable production by immobilized on mineral kissiris | ethanol productivities reached 10 g/L/h | [20] |
| sucrose | Fermentation with self-flocculating yeast in a tower upflow reactor | ethanol productivities reached 13.5 g/L/h | [21] |
| sucrose | Continuous ethanol fermentation in tower reactors with cell recycling | ethanol productivities reached 18 g/L/h | [22] |
| sucrose | Addition of air at a rate of 150 dm3 /min/m3 of reactor volume during the first 12 h | 35% sucrose is consumed and 16% ethanol is fermented | [23] |
| Starch | Co-cultured with amylolytic yeast | fermented unhydrolyzed starch to ethanol with conversion efficiencies over 90% of the theoretical maximum. | [24] |
| Starch | Starch Pretreated with 0.5 M HCl at 60℃, followed by combined actions of a-amylase and glucoamylase. | Ethanol conversion efficiencies reached 95% | [25] |
| Starch | a raw starch hydrolyzing enzyme was used that converts starch into dextrins at low temperatures, and hydrolyzes dextrins into sugars | Ethanol conversion efficiencies reached 88.4% | [26] |
| molasses | Batch and semicontinuous immobilized yeast fermentations together with a pervaporation method | Residual sugar was reduced 42%, ethanol produced increased 18.1% | [27] |
| molasses | batch and continuous fermentations of blackstrap molasses using immobilized yeast cells on thin-shell silk cocoons |
Ethanol concentration 11.5% higher than produced by free cells | [28] |
| molasses | A new candidate ethanol fermentation-related regulatory gene, PHO4, was replaced from a low ethanol yield but rapid growth strain to a high ethanol yield industrial strain. | Average high ethanol production reached 114.71 g/ L, ethanol production increased by 5.30% The fermentation time was 12.5% lower than that of the original strain. |
[29] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



