
Submitted:
08 May 2023
Posted:
09 May 2023
You are already at the latest version
Abstract
Abstract: The purpose of this review is to summarize recent knowledge on the main processes that occur during wound healing, including the responses of various cell types and the molecular mechanisms involved. Particular attention is also dedicated to new therapies often involved in the resolution of wounds, whether acute or chronic. Awareness of these dynamics is important for the various professional figures who are confronted with these kinds of problem daily.
However, the study of the healing mechanisms has yet to be defined in detail, in fact there are alterations of variously coordinated events which lead to a delayed resolution or, as in the case of keloids, to pathological states consisting in the excessive formation of scars with consequences yet to be seen. to define. Therefore, it is necessary to know the most feasible approaches and the most effective therapies.
Keywords:
Acute Wounds
; Cellular Infiltrate
; Chronic Wounds
; Keloids
; Scars
1. Introduction
The development of knowledge of WH throughout history has been remarkable. Historically, wounds were often related to infection and patients consequently died from septicemia rather than from the wound itself. With the advent of light microscopy, a greater awareness of the fine cellular mechanisms of wounds evolved, and this aided understanding of the treatment and care needed. Nevertheless, compromised WH is a major concern in public health. This is demonstrated by the fact that millions of people in the United States suffer from CW, and similar rates are found in Europe [1,2].
2. Structure of Skin
The skin (Figure 1) is the largest organ of the body. Its functions are building a protective barrier against the environment and preventing body dehydration. The layers of the skin are the epidermis, dermis, and hypodermis. The epidermis consists mainly of keratinocytes, as well as Langerhans cells or melanocytes, which provide a barrier against infection, moisture loss and solar non-ionizing ultraviolet radiation. The dermis is responsible for the elasticity and mechanical integrity of the skin. A rich cellular component (mainly keratinocytes, fibroblasts, and macrophages) is added to the vascularized extracellular matrix, which is rich in collagen, elastin and glycosaminoglycans. The hypodermis is mainly composed of adipose tissue and is also where stem cells and collagen are found. This layer serves mainly as a source of energy and is responsible for thermoregulation [3].
3. Issues Related to the Study and Management of Wound Healing
Impaired WH appears to be a major concern in the public health sector, as expensive and complex treatments are necessary for its management. Millions of patients need care for chronic wounds in the United States and USD 1 billion is estimated to be spent on this every year. This burden is increasing, mainly due to the growing presence of aging, diabetes, and other risk factors in the population. A similar scenario is observed in Europe, and in the UK around 200,000 patients have a chronic wound. It is estimated that over 1.5 million people could be affected by CW. As a result, WH has become a subspeciality, with fellowship programs offered at some academic centers for various medical professionals, including vascular surgeons, nurses, dermatologist, and general practitioners. The introduction of new technologies in WH is demand. Furthermore, problems are added to this discomfort as the understanding of the cellular mechanisms related to wound healing does not yet assume popular relevance but is more directed towards niche problems where, contrary to some disciplines, few scholars confront each other regarding the problems considered [1,2,4].
Guidelines for care of wounds are useful and represent the approach in clinical practice [5,6]. Nowadays, many research works are focused on solving the problem of WH, with most of them investigating very advanced therapies, such as cellular transplantation therapy, vascular enhancers, regenerative materials or nanoparticles in hydrogels. Despite the great number of expected novel therapies in development, these are currently very distant from being used in practice [5,6]. CW should be treated according to the TIME principles [6]: tissue debridement, infection control, moisture balance and edges of the wound. Debridement is the first step in the treatment of a CW; it must be performed weekly. Various methods for removing dead cells are used, such as surgical, autolytic, enzymatic or biological.
Biofilm presents in the extracellular matrix and is considered responsible for 80% of CW infections. Biofilm must be removed because it maintains the CW in the inflammatory stage [6]. In particular among the new techniques, it is worth mentioning those concerning the effects of the basic 3 compounds, i.e., OCT, PHMB and PVP-I on cells and the inflammatory process that act on the remotion of biofilm [6]. (See paragraph 14 for more information about recent therapies).
4. Vertebrates and Wound Healing
Wound healing is a conserved evolutionary process across species. However, the outcome of wound healing in the skin differs among species. In some lower vertebrates, including fish (zebrafish) and amphibians (axolotl and xenopus), the ability to perfectly regenerate the skin remains; in others, such as humans, it involves the formation of scar tissue. Although the latter mechanism meets the requirements of the skin’s fundamental function in preventing infections and dehydration, this process can have devastating aesthetic and psychological consequences, reducing the affected individual’s quality of life. It is, however, interesting to note that in the scientific literature, examples of scar-free healing have been described in fetal skin; and in adult skin, the regeneration of skin appendages following the infliction of large wounds has been noted [7].
5. Wound Healing Modalities
Wounds can Heal in Three Different Ways:
By first intention: this is the case for stab wounds, of which surgical wounds represent an example. These may be linear or have great curvature; they have clear edges and are sutured. This procedure, by minimizing the loss of substance by bringing the flaps together, favors their filling with granulation tissue, and leads to fast healing times and good aesthetic results.
By secondary intention: this concerns non-sutured wounds which are therefore left open, by choice or by necessity. In these cases, the granulation tissue, which forms on the bottom of the lesion, must proceed from below to the surface to fill it, through a process that takes longer and can cause serious blemishes.
By third intention: this type of healing concerns surgical wounds that have undergone partial or total dehiscence in the post-operative course. The treatment of this complication normally involves the complete reopening of the wound, its thorough cleansing, the removal of the mortified areas, and adequate plastering. Secondarily, following evaluation of the local situation and after excluding the presence of foci of infection, the flaps can be sutured again. This will favor the healing process which, in this case, will be said to take place by third intention [2,8].
6. Gross Stages of Wound Healing
The phases described represent the macroscopic evolution of the healing wound:
a) At 1 to 3 days after the injury: this stage includes blood clot formation (primary clot), activation of epidermal borders and early inflammatory response.
b) At 4 to 7 days after the injury: morphologically, this stage is marked by the formation of crusts. Histological analysis reveals migration of epidermal edges, selective proliferation of early granulation tissue and inflammatory response (lymphocytes and macrophages present in abundance).
c) At 8 to 12 days after the injury: the detachment of the crust is observed in morphologic studies. Histological findings show formation of new epidermis which differentiates by day 12. In addition, dermal closure is initiated, concomitant with granulation tissue formation. This phase is accompanied by attenuation of the inflammatory response.
d) At 12 to 30 days after the injury: characterized by matrix remodeling, terminal differentiation of the newly formed epidermis, increased elastic-fiber content and increased wound strength [8].
7. Acute Inflammatory Reaction
A feature of living organisms is reaction in the presence of an external stimulus (Figure 1). When this stimulus turns out to be an insult to the integrity of the tissue, the tissue response consists of the inflammatory reaction. This phenomenon, described by Hunter in 1793, consists of several apparently different processes, such as biochemical induction, angiogenesis and cellular response, among others. These processes aim both at the destruction of the noxious stimulus and at the repair of the tissue where it is present. Integrated within the phenomena described, but not strictly related, is the so-called vital reaction, described by Strassman in 1954 according to a previous definition by Plenk in 1786. This also includes other events not strictly considered part of the inflammatory response, such as platelet aggregation, complement activation by coagulation factors and prostaglandin metabolism [9,10].
8. Summary of Principal Events in Wound Healing
Wound healing involves a complex interaction between different cell types, which classically culminates in four overlapping phases: hemostasis, inflammation, proliferation and remodeling. This process ends with the formation of a scar.
During the hemostasis, the endothelial cells secrete von Willebrand factor, thus favoring the adhesion of platelets which, in addition to forming a clot, release numerous agents capable of stimulating or modulating inflammation. The release of these mediators ends with the formation of the fibrin clot which occludes the lesion and stops the bleeding. Injured arterial vessels rapidly narrow through smooth muscle contraction mediated by increased cytoplasmic calcium levels. Within minutes, reduced blood flow mediated by arteriole constriction leads to tissue hypoxia and acidosis. This promotes the production of vasoactive metabolites to cause reflex vasodilation and relaxation of arterial vessels. The duration of this phase is a few minutes [11].
During the inflammatory phase, the vasoconstriction is followed by vasodilation mediated principally by histamine or serotonin secreted by MCs (Figure 2B). This leads to the diapedesis of neutrophil granulocytes (Figure 2A) and monocytes (ready to transform in macrophages). This process allows the phagocytosis in the lesion against pathogens and damaged cells. In addition, leukocytes secrete cytokines and growth factors essential for initiating the next phase of that process. On the other hand, other cells such as keratinocytes also actively participate in the immune response, releasing inflammatory cytokines in response to several different biological, chemical and physical stimuli. The duration of this phase is 0-3 days [9,11].
During the proliferative phase, the fibroblasts, as well as being involved the formation of granulation tissue, regulate the migration and proliferation of keratinocytes (Figure 1) and angiogenesis. The first event of this phase is the migration of keratinocytes over the injured dermis. Fibroblasts and macrophages then replace the fibrin matrix with granulation tissue composed of hyaluronic acid, proteoglycans, glycoproteins and collagen type III, which forms a new substrate for keratinocyte migration at later stages of the repair process. The mechanism of neo-angiogenesis is operated by the endothelial cells, which, undergoing numerous mitotic cycles, generate numerous vessels able to exercise trophic function towards the granulation tissue forming at the wound. A fundamental role is also played by macrophages and MCs (Figure 2B), which provide a continuous supply of growth factors necessary to stimulate this process. In the epidermis, the keratinocytes (Figure 1) located at the edge of the lesion (Figure 1) proliferate and migrate towards the center of the injured site until the two edges are reunited, restoring the barrier function of the epithelium. The duration of this phase is 3-12 days [11].
During the maturation phase, the principal processes are collagen restoration and wound contraction, the latter occurring due to the conversion of wound fibroblasts to myofibroblasts, which express alpha-smooth muscle actin. These processes result in the formation of a scar in both children and adults. In this phase, the tensile strength increases, reaching values comparable to those of unwounded skin, thanks to the cross-linking of collagen by lisyl oxidase. The duration of this phase is 3 days to 6 months [9,10,11]. The scar formation involves a remodeling of the granulation tissue where MMPs and their inhibitors (TIMP) play a fundamental role. The synthesis of the extracellular matrix is therefore reduced, and its components are modified. Type III collagen is replaced by type I collagen, and the elastin, absent in the granulation tissue, reappears. It should also be considered that the cell death of various cell types, leading to a significant reduction in the number of cells in the granulation tissue, undoubtedly constitutes a fundamental event in wound resolution. It is debated whether the phenotype of the fibrocytes present in the dermis comes from myofibroblastic forms which progressively lose the typical morphological connotations, or from forms that differentiated late during the process described [12].

Table 1.
| Stages of wound healing | ||
|---|---|---|
| Phases | Time | Event |
| Hemostasis | A few minutes | Formation of the fibrin clot |
| Inflammatory | 3-12 days | Vasodilation, diapedesis, inflammatory response, phagocytosis. |
| Proliferation | 3 days to 12 days | Angiogenesis, granulation tissue formation, epithelialization |
| Maturation | 3 days to 6 months | Wound contraction |
9. Differences with Fetal Wound Healing
It is known from experimental animal and human studies that scarless healing occurs only during the early period of gestation. Human fetuses lose the ability to regenerate skin around 22 weeks of gestation. The main mechanisms responsible for scarless healing in fetal skin compared with adult skin are a reduced inflammatory reaction, differences in the composition and architecture of the extracellular matrix and mechanical loading in the fetal skin, differences in the proteolytic activity and secretion and sensitivity of TGFbeta, reduced angiogenesis, and morphological changes in keratinocytes during re-epithelialization. In addition, fetal fibroblasts show an “overactive morphology” producing higher amounts of extracellular matrix components. Finally, the early remodeling of fetal skin expresses lower levels of MMPs and TIMPs than adult skin [7].
10. Recent Advances about the Principal Cells Involved in Wound Healing
Phase: Hemostasis
10.1. Platelets
Platelets are non-nucleated fragments of megakaryocytes from the bone marrow. They play a crucial role in the wound healing process as they are not only essential for clot formation, but also produce multiple growth factors, cytokines and extracellular matrix involved in the various phases characteristic of this process. They sequentially promote (i) revascularization of damaged tissue through the induction of migration, proliferation, differentiation and stabilization of endothelial cells in new blood vessels; (ii) restoration of damaged connective tissue through migration, proliferation and activation of fibroblasts; and (iii) proliferation and differentiation of mesenchymal stem cells into tissue-specific cell types. Signaling molecules have been isolated from activated platelets, which influence and modulate the functions of other cell types. In addition to these factors, in response to cell membranes damaged by the stimulus of the lesion, arachidonic acid derives a series of powerful signaling molecules such as prostaglandins, leukotrienes and thromboxanes which have roles in stimulating the inflammatory response [13] (see Table 2 for details).
Phase: Inflammatory
10.2. Mast Cells
MCs (Figure 2B) contribute to wound healing by contributing to the evolution of the phases described. These cell types in fact contribute to hemostasis and clot formation by helping to increase the expression of Factor XIIIa in dermal dendrocytes through the release of TNFalpha, thus contributing to the stabilization of the clot. Furthermore, keratinocytes, secreting SCF, recruit MCs to the site. MCs also release inflammatory mediators, such as histamine, VEGF, IL6 and IL8, which contribute to the increase in endothelial permeability and vasodilation by facilitating the migration of inflammatory cells, mainly monocytes and neutrophils, towards the lesion. MC numbers and degranulation index increase at the edge of a wound within a maximum of 1-3 hours, becoming lower than baseline values after 6 hours or more. Mast cells increase in number again during the following 10 days, and then decrease, returning to control values 21 days after wounding. The late increase correlates with the upregulation of MCP1 and with the production of TGFbeta, which is also a potent chemoattractant for MC. TNFalpha, released in the first 15 minutes after wounding, and histamine, which is released even earlier, promote adhesion of leukocytes to the vessel. Leukotrienes, proteases and cytokines, released by MCs, represent chemotactic signals for the various types of granulocytes. For endothelial-leukocyte interactions, tryptase and cathepsin G play an important role. Angiogenesis is stimulated by TGFbeta, VEGF, chymase and tryptase. Conversely, heparin can inhibit angiogenesis by interacting with and inhibiting pro-angiogenic factors. In the last stages of the repair, MC-derived growth factors and cytokines influence the phenotype of fibroblasts, inducing their transformation into myofibroblasts which ensure eventual wound healing. As far as tissue remodeling is concerned, MCs can activate fibroblasts, promoting collagen synthesis: this effect is partly mediated by tryptase. MCs produce and release potent proteolytic enzymes, such as matrix metalloproteinases, thereby initiating the degradation of the extracellular matrix [14] (see Table 2 for details).
10.3. Neutrophils
Neutrophils (Figure 2A) are motile cells that infiltrate the wound in the hour following the insult, subsequently migrating to the injured site during the subsequent few hours. The complement cascade, namely the activation of various interleukins and the TGFbeta signaling, intervenes in the phenomenon related to chemotaxis that drives the same cell type towards the injured site. Infiltration is facilitated by various signaling molecules and chemotactic agents such as DAMPs released from necrotic cells, TGF-β, C3a and C5a, hydrogen peroxide, chemokines, and mediators from platelets. CXCL1, CXCL2, CXCL8 play an important role in initiating inflammatory cell recruitment. DAMPs directly activate neutrophils by binding to specific PRRs, such as TLRs, or indirectly by stimulating other cells to release neutrophil chemoattractants. At the wound site, recruited neutrophils also release some pro-inflammatory mediators, such as TNF-α, IL-1β, IL-6 and CXCL8, thereby further promoting inflammation The main function of neutrophils is to prevent wound infections by ridding the lesion of any pathogens. The achievement of this objective is linked to the generation of an oxidative burst due to ROS and through the release of antimicrobial proteins (cathepsins, defensins, lactoferrin, lysozyme) and chromatin “traps” and proteases that capture and kill bacteria in the extracellular space. Neutrophils can also promote angiogenesis and the proliferation of fibroblasts and keratinocytes by increasing the release of the cytokines IL-1β and IL-8, VEGF, and MCP-1, which promote angiogenesis and proliferation. After completing their task, the neutrophils must be cleared from the wound site. Recent evidence has also shown that neutrophils also produce chromatin and protease “traps” that capture and kill bacteria in the extracellular space. These cells then undergo apoptosis and are subsequently phagocytosed by macrophages. Uncontrolled migration of neutrophils prolongs the inflammatory process, leading to excessive generation of ROS and proteases. Toxic proteases and increased ROS levels degrade the ECM and damage cell membranes, leading to the formation of chronic wounds [15] (see Table 2 for details).
10.4. Macrophages
Macrophages are phagocytic cells that reach the lesion area 48 and 72 hours after the origin of this event attracted by the release of chemical messengers released by platelets and damaged cells. Macrophages contain numerous growth factors within their granules, such as TGFbeta and EGF, involved in the inflammatory response in angiogenesis and favoring the formation of granulation tissue due to the secretion of the extracellular matrix carried out by fibroblasts, stimulated in turn by macrophages and MCs. Two distinct activation phenotypes have been defined for macrophages: type M1 and type M2. M1 macrophages play the role of effector cells in immune responses; M2 macrophages appear to be involved in immunosuppression and tissue repair. The macrophage phenotype changes readily based on spatiotemporal cues during wound healing, and several subsets of macrophages beyond the limited boundaries of M1 and M2 have been defined based on their cell surface markers, cytokine/growth factor/chemokine production and function. Macrophages play particularly important roles in vasculature, as they position themselves around newly formed blood vessels and aid in their stabilization and fusion. At the onset of the remodeling phase, macrophages release MMPs to break down the provisional extracellular matrix, and then undergo apoptosis so that the skin can mature to resemble its original, uninjured state. In chronic wounds, pro-inflammatory macrophages persist without switching to anti-inflammatory phenotypes, which are thought to contribute to impaired tissue repair [15,16] (see Table 2 for details).
10.5. Dendritic Cells
DCs are antigen-presenting immune cells. Several studies have indicated that the recruitment of dermal DCs may be involved in the repair process of damaged tissue. DCs are rarely detected in wounds less than 1 day old, whereas DC accumulation increases over time in wounds aged 3 to 14 days. There is a specific population of DCs in the human epidermis, which express CD1a and CD207/langerin, and are called Langerhans cells. Studies have shown that CD1a+ cells rapidly increase in number within the first hour after injury. Cellular interactions between DCs and MCs are necessary for the differentiation of precursors into DCs through the release of short-acting soluble factors such as TGFbeta. It was recently shown in a mouse model that complete wound healing did not occur in cases of DC deficiency. It follows that DCs play an important role in accelerating the early events that characterize wound healing, probably by secreting factors that trigger the proliferation of the cells involved in this phenomenon [17,18] (see Table 2 for details).
10.6. Plasmacytoid Dendritic Cells
PDCs (Figure 2C) express TLR7 and TLR9 and produce large amounts of type I IFN. Recent studies now show that, although absent from normal skin, PDCs are rapidly recruited to sites of skin inflammation, acting as an early source of type I IFN which contributes to wound healing, including through interaction with Treg cells [19] (see Table 2 for details).
10.7. Lymphocytes
Lymphocytes appear in the wound after 72 hours and are believed to be important in regulating wound healing. Lymphocytes exert a specific response against microbes and other foreign material in the wound: B lymphocytes through antibodies and T lymphocytes through the production of cytokines and stimulation of cytolytic activity. The response from lymphocytes is evoked by factors such as IFNgamma and TNFalpha, as well as by antigen-presenting cells such as dendritic cells and macrophages. Recent studies indicate that Tregs facilitate cutaneous wound healing. In particular, upon wounding, Tregs induce expression of the EGF receptor. The deletion of EGFR in Tregs results in reduced Treg accumulation and activation in wounded skin, delayed wound closure, and increased proinflammatory macrophage accumulation [20] (see Table 2 for details).
Phase: Proliferation
10.8. Keratinocytes
Keratinocytes (Figures 1, 3) are the cellular component most represented in the epidermis and are directly involved in the process of wound healing. Keratinocytes are activated during the inflammatory phase of wound healing to secrete various cytokines and growth factors. Migration to the injured area occurs within 24 hours and is accompanied by morpho-functional changes of these cell types which occur a few hours after the injury, with detachment from the underlying basement membrane and migratory movement. The latter process is facilitated by MMP-1, which is expressed at high levels at the edges of the wound. Migrating basal cells are thought to express specific surface markers such as CD44, in contrast to resting basal cells. However, the mechanisms of re-epithelialization have not been fully elucidated During wound healing, keratinocytes can also modulate the functional activity of other cell types, such as fibroblasts, by inducing their proliferation to produce growth factors, which in turn will stimulate the proliferation of these cell types. In addition, it has been shown that keratinocytes can recognize PAMPs and DAMPs, leading to the activation of pro-inflammatory signaling pathways through the production of inflammasome and pro-inflammatory cytokines, such as interleukins (e.g., IL-1, IL-6, IL-8, IL-10, IL-18 or IL-20), TNF-α and chemokines (e.g., RANTES, MCP-1 or MIP-1). Furthermore, fibroblasts can differentiate into myofibroblasts under the control of keratinocytes (Figure 3). This depends on a finely tuned balance between a pro-inflammatory or TGFbeta-dominated environment [21,22,44].
10.9. Endothelial Cells
Endothelial cells play a crucial role in the regulation of inflammation during the early stages of wound healing. The loosening of cell–cell contacts and the expression of adhesion molecules allows endothelial cells to facilitate the movement of circulating inflammatory cells into the tissue at the site of injury. VEGF is a strong inducer of vascular permeability in the skin, and the correlation between VEGF expression patterns and permeable blood vessels in injured skin suggests that VEGF contributes to vascular permeability at early stages of healing. Endothelial cell proliferation and chemotaxis occurs under the action of angiogenic factors VEGF, FGF, PDGF and SDF-1, which induce the consequent modification of intracellular pathways and cytoskeleton. Heparin sulfate proteoglycans and syndecans also play a key role in regulating the angiogenic activity of VEGF and FGF. Activated endothelial cells also secrete MMPs to degrade the capillary basement membrane and allow their migration and proliferation. Endothelial cell migration appears to be a coordinated process involving changes in cell adhesion, signal transduction and dynamic reorganization of the cytoskeleton It follows that new vessels are formed and the recruitment of pericytes stabilizes the newly formed vessels in a mature conformation [23,24] (see Table 2 for details).
10.10. Pericytes
Pericytes (Figure 2D) are new cell types involved in wound healing phenomena. These cells, together with the endothelial cells, participate in neo-angiogenesis as well as contributing to the caliber of the newly formed vessel. Existing studies have shown how pericytes interact with macrophages during the inflammatory phase of wound healing. It also appears that these cells stimulate an immune response by triggering the activation of T lymphocytes through the secretion of various cytokines. There is emerging evidence indicating that pericytes may be a source of myofibroblasts, and pericyte–myofibroblast transition could occur via a PDGF-dependent mechanism [25] (see Table 2 for details).
10.11. Fibroblasts
Fibroblasts (Figure 3) are essential in all phases that characterize wound healing. These cell types play a crucial role in the secretion of the extracellular matrix, and in the remodeling of the new one. During the wound resolution process, studies have shown that the initial repair phase is mediated by fibroblasts arranged in the reticular dermis, whereas during the re-epithelialization phase fibroblasts arranged in the so-called papillary dermis are often recruited. During the inflammatory phase, activated fibroblasts engage in crosstalk that enhances the local immune response and immune cell activation in several ways: 1) by producing pro-inflammatory cytokines (TNFα, IFNγ, IL6, and IL12) and releasing a broad range of chemokines (CXCL1, CX3CL1 and CCL2); 2) by promoting cellular interactions through the expression of ICAM1 and CD40, which also activate dendritic cells; 3) by secreting MMPs necessary for a wound’s stromal remodeling. Therefore, fibroblasts modulate the recruitment of immune cells by regulating their survival in the damaged tissue. The crosstalk between fibroblasts and macrophages is crucial in regulating the transition from the inflammatory phase to the subsequent proliferative phase, thus determining the correct progression of the healing process. Fibroblast activity becomes even more important during the proliferation phase. In fact, during this phase the fibroblasts proliferate and contribute to angiogenesis by secreting pro-angiogenic molecules, including vascular VEGF, FGF, ANG-1 and TSP. Stimulated by growth factors produced by macrophages and other immune cells, fibroblasts produce MMPs, which degrade the fibrin clot promoting cell migration, and ECM molecules, including fibronectin, hyaluronic acid, proteoglycans and collagen (mainly type III), which replace the fibrin clot with a new provisional matrix that supports keratinocyte migration necessary for re-epithelialization [26] (see Table 2 for details).
Phases: Maturation and remodeling
10.12. Myofibroblasts
The contraction of the granulation tissue formed following the injury appears to be promoted by myofibroblast cells, which are differentiated from fibroblasts in situ. Myofibroblasts regulate wound contraction and tissue remodeling by combining the ability to synthesize ECM proteins and assume a contractile phenotype. Fibroblast–myofibroblast transdifferentiation is mainly regulated by TFG-β1 and by ECM stiffness. The assumption of contractile properties by myofibroblasts is linked to actin incorporation into stress fibers. Contractile activity not only causes wound contraction, but also stabilizes the ECM, which in turn induces myofibroblast differentiation and persistence. In this stage, collagen III is replaced by collagen I and the ECM undergoes remodeling towards increasing complexity, order and tensile strength [26,27].
Subsequently, after having restored the integrity of the tissue, the activities of the myofibroblasts cease and a part of their population is eliminated by apoptotic phenomena; however, the modulation of the various signals (including the timing) that trigger these phenomena is not fully understood. Nevertheless, it must be added that the persistence of the activity of these cell types will lead to the manifestation of hypertrophic scars or fibrotic lesions, as occurs in scleroderma [26,27].
10.13. Central Role of Mast Cells in Wound Healing: A Hypothesis
Reflecting on the salient data of these paragraphs, it seems fair to propose a central role for MCs in the processes regulating wound healing. In acute wounds, MC activation, by keratinocytes or microenvironmental factors, additionally leads to the release of TNFalpha for dendritic cell differentiation, the secretion of various mediators to induce angiogenesis, the release of extracellular matrix and the reorganization of the cellular infiltrate in response to the stimulus considered. These events lead to the secretion of TGFbeta from various cell types present in the microenvironment and have as their main result the differentiation of the M1 and M2 phenotype macrophages. These cells, probably in synergy with keratinocytes, activate fibroblasts, which are able to differentiate into myofibroblasts during the late stages of wound healing. The pleiotropic activity of TGFbeta also seems to be related to the differentiation of PDCs, which are able to interact with Treg cells for a mechanism that can induce tolerance through the co-expression of CD45 by the latter cell types in the healing processes of wounds. In chronic wounds, there is further evidence of the important role played by these cell types. In various articles, it has been reported that mast cells show an increased degranulation index in chronic wounds. This event leads to modifications of the inflammatory infiltrate with various responses from the various cell types involved. Among the cytokines produced here is TGFbeta, which assumes a crucial role in the various stages leading to the healing of chronic wounds. In this type of wound, as has already been observed, an interaction between mast cells and neuronal cells is described. The release of nerve mediators involved in wound healing is in fact linked to the interaction between these cell types [14].
Table 2.
Main bioactive factors secreted by cells involved in wound healing.
| Wound healing | ||||
| Phases | Time | Cells | Main bioactive factors secreted by cells involved in wound healing | Functions |
| Hemostasis | A few minutes | Platelets: |
CYTOKINES: TNFalpha GROWTH FACTORS: PDGF, TGFbeta, TGFalfa, FGF, IGF-1, VEGF CHEMOKINES: CXCL8, CXCL1, CXCL2 |
Initiation of inflammatory responses, angiogenesis |
| Inflammatory | 3-12 minutes to 3 days | Mast Cells |
BIOGENIC AMMINE: Histamine CYTOKINES: TNFalpha, IL4, IL6, IL8 GROWTH FACTORS: VEGF, FGF |
Vasodilation, Inflammatory response Production of ECM |
| As above | As above | Neutrophils |
CYTOKINES: IL1beta, IL6, IL8, TNFalpha CHEMOKINES: CXCL1, CXCL2, CXCL8 GROWTH FACTORS: IGF, VEGF |
Inflammatory response, keratinocyte proliferation, fibroblast proliferation Angiogenesis, collagen synthesis, endothelial cell activation, |
| As above | As above | Macrophages |
CYTOKINES: IFNgamma, IL1beta, IL6, IL8, IL10, TNFalpha CHEMOKINES: Rantes GROWTH FACTORS: EGF, FGF, IGF, PGDF, TGFbeta, VEGF |
Inflammatory response Fibroblast proliferation, fibroblast chemotaxis, angiogenesis, ECM deposition |
| As above | As above |
Dendritic cells, plasmacytoid dendritic cells |
GROWTH FACTORS: TGFbeta CYTOKINES: IFN gamma |
Inflammatory response |
| As above | As above | Lymphocytes |
CYTOKINES: IFNgamma, IL2, IL4, IL10 CHEMOKINES: MCP, RANTES, MIP, Lymphotactin |
Inflammatory response, decrease in collagen synthesis, synthesis of MMPS Inflammatory response |
| Proliferation | 3 days to 6 months | Keratinocytes |
CYTOKINES: IL1, IL6, IL8, IL10, IL18, IL20, TNFalpha GROWTH FACTORS: TGFbeta, VEGF, EGF, PGDF, SCF CHEMOKINES: RANTES, MCP or MIP-1 |
Proliferation of keratinocytes Angiogenesis, proliferation of keratinocytes Inflammatory response |
| As above | As above | Endothelial cells |
GROWTH FACTORS: CTGF, FGF, IGF, TGFbeta, PGDF, VEGF |
Proliferation of fibroblasts and keratinocytes, differentiation of keratinocytes, angiogenesis |
| As above | As above | Fibroblasts |
CHEMOKINES: CXCL1, CX3CL1, CCL2 CYTOKINES: IL6, IL8, IL12 GROWTH FACTORS: FGF, IGF, KGF, VEGF |
Chemotaxis of inflammatory cells, proliferation of fibroblasts, fibroblast differentiation |
Adapted from [28].
Figure 1.
Activation of the skin microenvironment in conditions of injury such as a wound. Response of keratinocytes and cellular infiltrate. HE, Light microscopy, scale bar = 10 microns (Courtesy of one of the authors of this document: Stefano Bacci).
Figure 1.
Activation of the skin microenvironment in conditions of injury such as a wound. Response of keratinocytes and cellular infiltrate. HE, Light microscopy, scale bar = 10 microns (Courtesy of one of the authors of this document: Stefano Bacci).
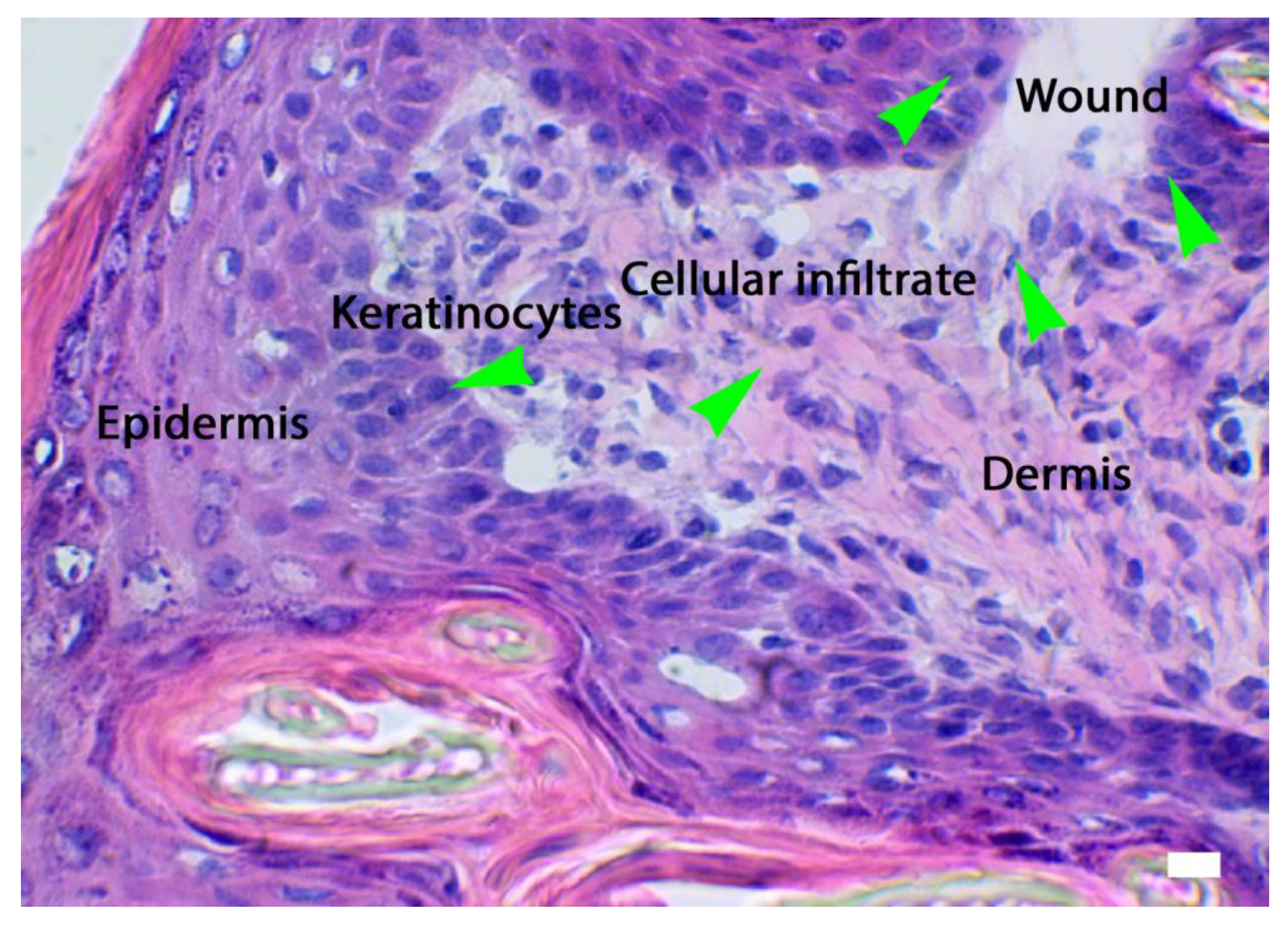
Figure 2.
Main cell types that are involved in the composition of the cellular infiltrate during wound healing: A) granulocytes stained with Ly6G B); mast cells stained with avidin; C) plasmacytoid dendritic cells stained with CD303 (BDCA-2); D) pericytes stained with PDGF. Fluorescence microscopy, scale bar =10 microns (Courtesy of one of the authors of this document: Stefano Bacci).
Figure 2.
Main cell types that are involved in the composition of the cellular infiltrate during wound healing: A) granulocytes stained with Ly6G B); mast cells stained with avidin; C) plasmacytoid dendritic cells stained with CD303 (BDCA-2); D) pericytes stained with PDGF. Fluorescence microscopy, scale bar =10 microns (Courtesy of one of the authors of this document: Stefano Bacci).
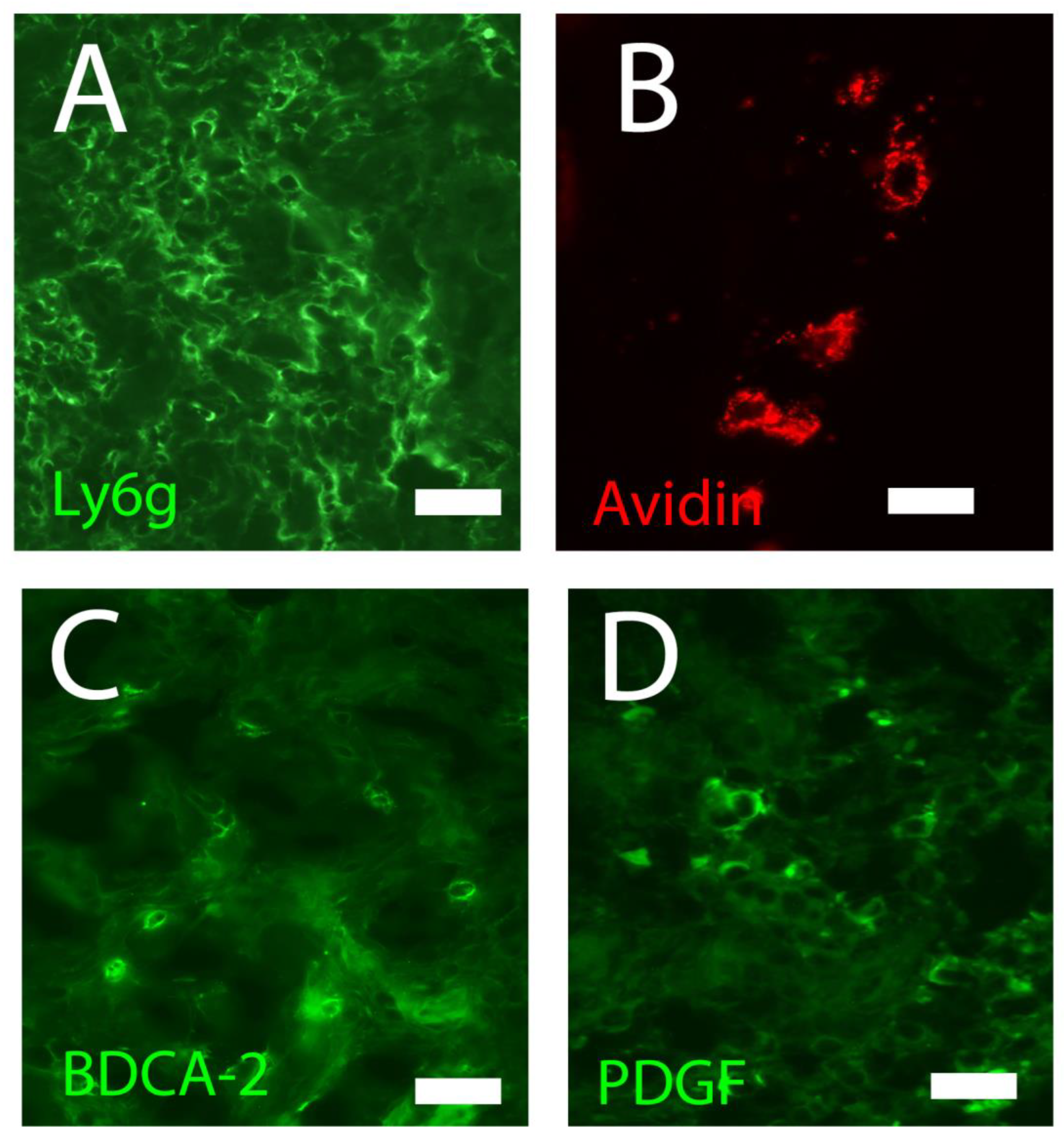
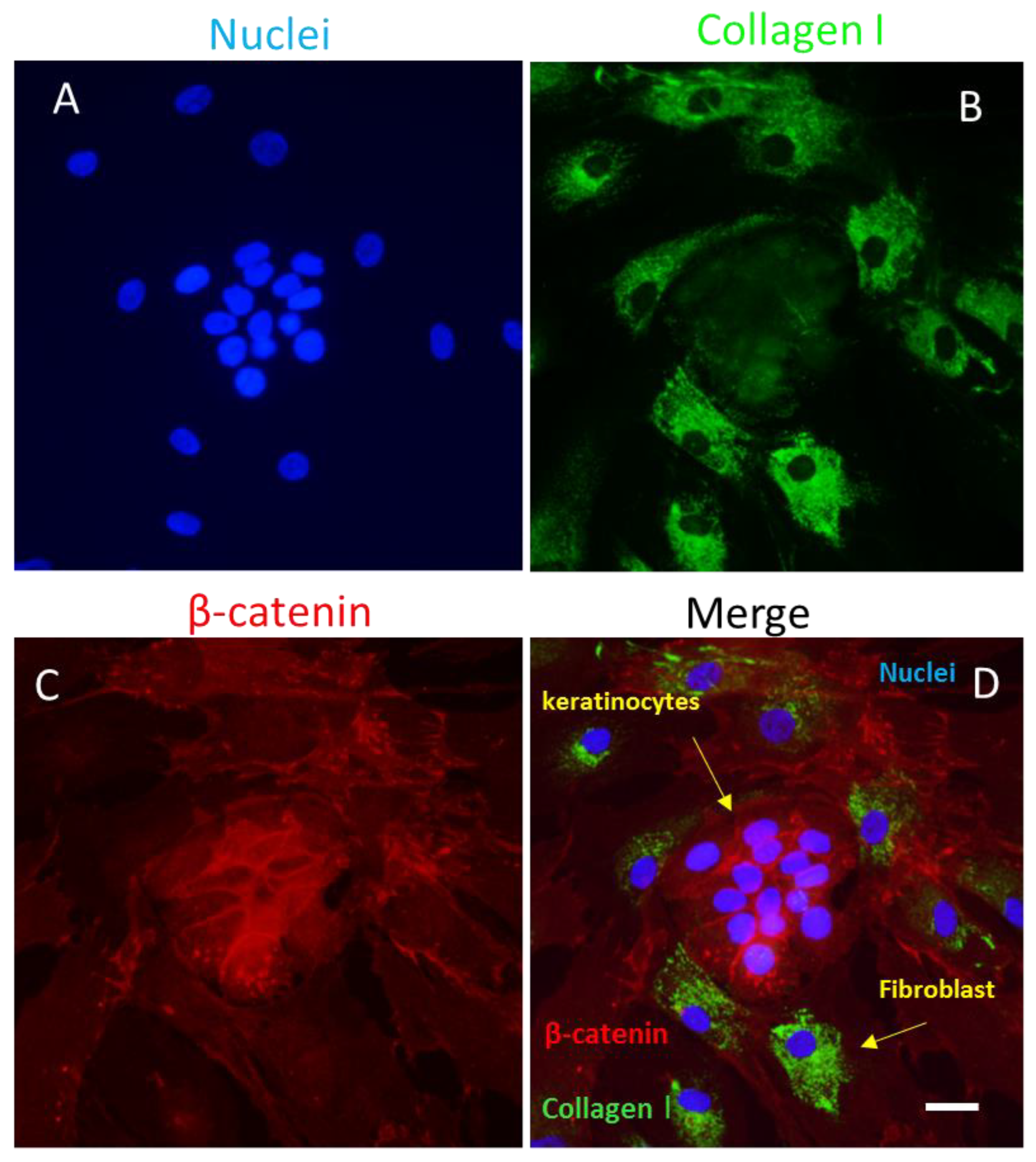
Figure 3.
Co-culture of keratinocytes and fibroblasts stained for β-catenin (Alexa red) and collagen type I (Alexa green). A. DAPI-stained nuclei, B. Fibroblasts labeled for collagen type I with Alexa green, C. Keratinocytes labeled for β-catenin with Alexa red, D. merge. Confocal microscopy, scale bar = 100 μm (Courtesy of one of the authors of this document: Maria Luisa Hernández-Bule).
Figure 3.
Co-culture of keratinocytes and fibroblasts stained for β-catenin (Alexa red) and collagen type I (Alexa green). A. DAPI-stained nuclei, B. Fibroblasts labeled for collagen type I with Alexa green, C. Keratinocytes labeled for β-catenin with Alexa red, D. merge. Confocal microscopy, scale bar = 100 μm (Courtesy of one of the authors of this document: Maria Luisa Hernández-Bule).
11. Recent Advances about the Molecular Biology of Wound Healing
Wound healing is a complex process and many of the steps in wound repair are poorly characterized at the molecular level at present. A variety of transcription factors and related molecules participate in this process, including growth factors, TGFbeta/Smad, the E2F family, STAT3, homeobox genes, hormone receptors (androgen, estrogen and glucocorticoid), PPARs, Wnt/beta catenin signaling, AP1, c-Myc and Erg1, and other factors such as proteases (including matrix metalloproteinases, MMPs), cytoskeletal proteins and enzymes that regulate the cellular redox balance. These factors are not independent but mutually associated with each other [15].
11.1. Molecular Processes in the Hemostasis Phase
During the homeostasis phase, platelets are activated and produce cytokines, as well as growth factors PDGF, TGFbeta and VEGF. These factors play a fundamental role in clot formation. The cytokines and other secreted factors stimulate the response of the target cell after binding to cell surface receptors and activation of the signaling pathway. This activation transcribes genes that stimulate angiogenesis, collagen synthesis or collagenase. Other growth factors, such as PDGF, not only stimulate neutrophils and macrophages, but are also a mitogenic and chemotactic factor for fibroblasts and smooth muscle cells [29].
11.2. Molecular Processes in the Inflammatory Phase
In the inflammatory phase, multiple molecules, such as cytokines, matrix proteins or enzymes, intervene. One type of cytokines is chemokines. Chemokines are classified into the CC, CXC, CX3C and XC families based on the spacing or presence of four conserved cysteine residues. CXC chemokines primarily attract neutrophils and lymphocytes and are believed to orchestrate the early phases of wound healing [30].
After clot formation, platelets trapped and aggregated in the clot release growth factors, such as PDGF or CTAP-III, which become proteolytically activated on NAP-2, and CXCL7, activated by neutrophils attached to the thrombus [31]. Initially, low concentrations of NAP-2 mediate neutrophil chemoattraction via CXCR2. In addition, GRO-a secretion by endothelial cells and pericytes further promotes the neutrophil migration process[32].
Neutrophil recruitment is supported by ENA-78 (CXCL5), which is expressed at lower levels compared with GROa in single mononuclear cells within the provisional wound matrix [33].
Following the neutrophils, the monocytes migrate into the wound where they become macrophages, which engulf debris and bacteria. Macrophage activation has been classified into two states, M1 and M2, depending on the stimulus. Activation of M1 results in a highly pro-inflammatory macrophage, which is mediated by TLR-4 and IFN-gamma ligands. M2 activation is mediated by IL-4 and/or IL-13, which release some growth factors such as macrophage-derived growth factors, PDGF, FGFalpha, FGFbeta, TGFalpha and TGFbeta. FGFbeta is a chemotactic and mitogenic factor for fibroblasts, endothelial cells and other mesenchymal cells, which are the stimulus for angiogenesis. In addition, FGFbeta stimulates wound contraction, epithelialization, and production of collagen, fibronectin and proteoglycans. Further, macrophages produce and direct inflammatory cytokines such as TNF, IL6 and IL1. IL-1 stimulates cell proliferation and promotes angiogenesis through the endothelium, whereas TNFalpha is a mitogen for fibroblasts. Macrophages also secrete collagenases and elastases, which lyse injured tissue and help debride the wound [29].
Macrophages, wound edge keratinocytes and endothelial cells produce MCP1, which attracts monocytes, MCs and lymphocytes during cutaneous wound healing. The attracted macrophages and lymphocytes produce growth-promoting and regulatory factors. MCs release IL4, which can stimulate fibroblast proliferation and decrease the expression of MCP1 and IL8, limiting the inflammatory reaction. In addition, MCP1 may also contribute to endothelial cell migration during angiogenesis [34].
Lymphocytes migrate to the wound during the inflammatory phase and secrete lymphokines, as well as EGF and FGFbeta, among other substances. T cells are attracted to the wound by IL1, which also contributes to the regulation of collagenase. Lymphocytes also produce chemokines, initially MCP1 and after day 4, IP10 (CXCL10), MIG (CXCL9) and MDC (CCL22).
11.3. Molecular Processes in the Proliferative Phase
In this regeneration phase, recent data suggest that fibroblasts are not only producers but also targets of chemokines. Thus, MCP1 enhances gene expression of MMP1 as well as TIMP1 in primary human dermal fibroblasts [34]. In response to growth factors synthesized by macrophages, such as PDGF, FGF, VEGF, TGFalpha, TGFbeta, KGF, etc., fibroblasts begin to migrate, proliferate and produce extracellular matrix components such as glycosaminoglycans, proteoglycans and collagen. Collagen is secreted into the extracellular space in the form of procollagen and is modified to form tropocollagen that is deposited on fibronectin and serves as a support for the myofibroblast, contributing to wound healing.
Migratory keratinocytes interact with a provisional matrix of fibrin crosslinked with fibronectin, which promotes adhesion of keratinocytes to the matrix at the base of the wound. EGF is a potent mitogen for epithelial, endothelial and fibroblast cells, and stimulates fibronectin synthesis, angiogenesis, fibroplasia and collagenase activity. Furthermore, the chemokine IL-8 stimulates the migration and proliferation of keratinocytes, which is highly expressed at the edge of the wound to promote its closure [35]. GROalpha expression is colocalized with IL-8, but GROalpha mRNA levels are significantly lower [36]. In addition, keratinocytes have been reported to express CXCR2, the IL-8 receptor and GROalpha [37].
Endothelial cells regulate the recruitment of leukocytes from the intraluminal compartment to the tissues and induce angiogenesis mediated by MCP1, RANTES, IL8, GROalpha, IP-10 and MIG, among other substances [38]. It has been shown that chemokines containing a Glu-Leu-Arg (ELR) motif [39] adjacent to their first cysteine amino acid at the NH2 terminus are potent promoters of angiogenesis. This group of ELR-containing chemokines includes IL-8, GROalpha, GRObeta (CXCL2) and GROgamma (CXCL3), as well as CTAP-III, b-thromboglobulin and NAP-2 [40]. CXC chemokines lacking the ELR motif, such as the IFNgamma-inducible cytokines IP10, MIG or ITAC, act as inhibitors of angiogenesis [39,40]. On the other hand, the expression of MCP-1 by these cells establishes a chemokine gradient, which facilitates the specific recruitment of specific types of leukocytes at sites of inflammation [41]. Therefore, chemokines not only regulate the locomotion of resident and passenger cells, but may also influence tissue remodeling.
11.4. Molecular Processes in the Maturation Phase
The remodeling phase is primarily driven by cell differentiation and mediated by various growth factors. These factors mediate mesenchymal-to-mesenchymal transition and endothelial-mesenchymal transition. Such transitions could occur through the TGFbeta signal pathway or through the Notch signal, a pathway that inhibits the expression of the endothelial cell adhesion molecule VE-cadherin [42,43]. Beta2-AR has also been described as a critical molecule mediating the EMT process [44].
Malfunctioning of processes related to wound healing
12. Chronic Wounds
Wounds that do not heal within 6 to 8 weeks are considered chronic.The difficulties or even impossibility of treating these types of wounds entail high costs in terms of care [1,45,46].
There are different types of chronic wounds, being the most frequent in clinical practice venous ulcers, pressure ulcers and diabetic foot ulcers. However, underlying common mechanism could be found. Microbial growth, also known as biofilm, is one the most important cause of delayed wound healing and consequently of the conditions leading to the evolution from acute to CW. Wound microbial communities are characterized by the presence of various species of bacteria occupying the site of infection. Among the aerobes and facultative bacteria, Staphylococcus aureus, Pseudomonas aeruginosa and β-hemolytic Streptococci remain the primary causes of chronic wound infections. Regarding the estimation of the anaerobic burden of chronic wound infections, due to the lack of isolation practices and therefore adequate culture the information remains insufficient, but it is believed that they constitute a significant percentage of the microbial population. Other bacteria founded in CW infections include Enterococcus species, Klebsiella pneumoniae, Acinetobacter baumanii and Enterobacter species (ESKAPE pathogens), coagulase-negative Staphylococci, and Proteus species. Although the clinical relevance of fungi in CW infections has been little studied, they constitute a significant component of the wound microbial burden and several endogenous fungi, including Candida, Curvularia and Malasezzia, have been implicated in CW infections [47].
In general, the processes involved in CW healing are like those in acute wound healing, but dysregulation of MMP secretion is strongly associated with CW which perpetuates the inflammatory stage. Prolonged inflammation in CW is mainly mediated by various cell types present in the cell infiltrate [45,46,47,48,49]. Neutrophils appears excessively in CW and liberate significant metalloproteinases, which not only destroys the connective tissue matrix and elastase, but also inactivate important factors involved in wound healing as PDGF and TGFbeta. However, it is necessary to consider how immune cells interact with keratinocytes. This occurs through the secretion of various signaling molecules, but the contribution of these latter cells to the formation of a CW is not fully understood. Additionally, in chronic wounds, keratinocytes express genes involved in partial proliferative activation, and this might explain the epidermal hyperproliferation presented on the ulcer edges. Besides this, the fibroblasts are unresponsive to the migratory stimulant TGFbeta. This is reflected in dramatically reduced levels of TGFbetaR, and in reduced levels of the downstream components of the TGFbetaR signaling cascade [45,46,47,48,49]. It is worth remembering how interactions of the immune system with the nervous system are important in the regulation of wound healing processes. Recent studies have demonstrated that MC interactions with neuronal cells containing neurotransmitters involved in wound healing processes, such as CGRP, NGF, NKA, NPY, SP, PGP 9.5, and VIP, are common in chronic wounds [48]. This fact can be related to the secretion of extracellular matrix by fibroblasts, as well as increases in TGFbeta levels and the response of cellular infiltrates.
13. Fibrosis: Hypertrophic Scarring and Keloids Associated with Wound Healing Phases
Hypertrophic scars and keloids are the clinical expression of an alteration in the process of wound healing and scarring, the main differences between then, is the fact that keloids could growth spontaneously and overside the limits of the injury. In both, the underlying alteration is that the myofibroblasts are not replaced by fibroblasts remaining in the proliferating phase of the wound with low rates of apoptosis [50]. These myofibroblasts generate persistent chronic inflammation, due to the continuous production of pro-fibrotic cytokines and chemokines, such as TGFbeta1, TGFbeta2, VEGF, FGF and CTGF, and synthesize a large amount of fibrotic extracellular matrix, composed of collagen types I and III and alpha-SMA, which prevents the normal functioning of the affected tissues [50].
13.1. Hypertrophic Scarring
Hypertrophic scarring is a major clinical problem, affecting some 100 million patients in developed countries. A hypertrophic scar exhibits fibrosis tissue limited to the scar, in contrast to a keloid, which overpasses the limit of the initial injury. A large body of evidence suggests that excessive inflammation generates pro-fibrotic molecules, which in turn cause excessive activation of fibroblasts. In addition, excessive angiogenesis and prolonged re-epithelialization can extend the release of pro-fibrotic growth factors. In the last few years, many biomolecules have been implicated in hypertrophic scarring; however, their exact mechanisms have yet to be fully elucidated, in part due to the complexity and overlapping nature of wound healing processes [51,52,53]. The main treatment of hypertrophic scars is prevention as possible, especially in a planned surgery. When therapies need to be added, usually the option is a combination depending on the availability or patient preference, as intralesional corticoids, pressure therapies, patching with different substances or laser, as fractionated CO2 or Erbio, are probably the most widely used [54,55].
13.2. Keloids
A keloid is an abnormal excessive proliferation of scar tissue that forms at the site of cutaneous injury that grows beyond the original margins of the scar. Keloids occur where trauma, surgery, blisters, vaccinations, acne, or body piercing have injured the skin, but can also appear in minimal trauma or be spontaneous. The treatment of keloids is challenging as they tend to recurrence and resistance to therapies, which habitually are used in combination due to this resistance. There are several treatments available as intralesional corticoids, silicone patchs, bleomycin, cryotherapy, radiotherapy or fractionated laser treatment [4, 53–55]. Recently, Aghmandy et al. proposed that signaling in both keratinocytes and fibroblasts is disrupted in keloid pathology and that the methylation status of keloids could be implicated in the mechanisms of keloid scar formation and remission [56]. In addition, some mutations have been described in the Asian population, suggesting an individual predisposition as seen in habitual clinical practice [51].
14. Impairment of Wound Healing and Recent Therapeutic Strategies
Local and systemic factors can impair wound healing. A crucial factor is tissue hypoxia, which may be caused by primary vascular diseases or secondary to metabolic diseases such as diabetes. In skin ulcers, the persistence of the inflammatory phase leads to high protease activity and degradation of growth factors and of other molecular stimuli involved in tissue repair. Imbalance between hydrolases – matrix metalloproteases, elastase and plasmin protease – and their inhibitors results in abnormal degradation of the extracellular matrix.
Various medical approaches and therapeutic interventions can affect the different processes involved in wound healing. The healing time may be shorter when there is less injured tissue, for example during minimally invasive surgery.
ESC are a convenient target to use in wound therapies because they already reside within the skin and participate in the normal healing response. They have been shown to support healing by increasing proliferation and migration of fibroblasts and keratinocytes as well as enhancing angiogenesis by human vascular endothelial cells [57].
MSC therapy is another emerging option to treat acute and chronic non-healing wounds. Beneficial effects are accomplished through structural repair via cellular differentiation, immunomodulatory responses, direct secretion of growth factors, advanced neovascularization and re epithelialization, as well as mobilization of resident staminal cells. Thereby, MSC play a pivotal role in all three healing phases. At the wound margins they stimulate the formation of granulation tissue by enhancing epidermal cell proliferation and growth of new blood capillaries. Further, endothelial cell recruitment is stimulated through the release of pro-angiogenic factors and growth factors such as vascular endothelial growth factor and angiopoietin-1. MSC modify tumor necrosis factor-α production and lower NK cell function in the inflammatory phase, thereby reducing interferon-γ activity. In the last healing phase, scar formation is reduced through secretion and lowering of TGF-β1 to TGF-β3 ratio, IL-10 up-regulation as well as IL-6 and IL-8 down-regulation. These effects are accompanied by a decline in collagen production and fibrosis. These techniques are based on the use of MSC-seeded micro-or nanostructured scaffolds with natural biomaterials, such as collagen and cellulose derivatives. Thereby, pronounced to complete regeneration of non-healing wounds (burns, decubitus ulcers, diabetic ulcers) has been reached in preclinical and clinical studies [58].
Recently Kua et al. [59] studied the potential use of human umbilical cord lining epithelial cells for treating cutaneous wounds. In vivo studies assessed their ability to promote wound healing. and sensing properties. The peptide-modified silk fibroin exhibited wound healing capacity and piezoresistive properties, and additionally demonstrated a sensitivity to humidity.
Patients with wounds often are provided pharmacologic interventions for their wounds as well as for their acute or chronic illnesses. Drugs can promote wound healing or substantively hinder some medications causing wounds or skin reactions. The health care literature includes many reviews describing the impact of pharmacologic agents on wound healing. Medications reported to delay wound healing include anticoagulants, antimicrobials (various antibiotic classes), anti-angiogenesis agents (e.g., bevacizumab, aflibercept), antineoplastic drugs, anti-rheumatoid drugs), nicotine, steroids, and vasoconstrictors. Because of their ubiquity of use, two categories of medication require special mention: steroids and NSAIDs. Several reviews support that short-term use of both categories has limited impact on wound healing. However, long-term use of steroids and NSAIDs can have marked negative impact [60]. Among the new drugs to be considered, Exe 4 can probably play an important role. Exe4 is a natural peptide sharing 53% homology with GLP-1, the insulinotropic intestinal peptide belongs to the incretin hormones. GLP-1, exerts important post prandial insulinotropic effect being responsible for approximately 60% of the insulin secretion post prandially. GLP-1 effects are secondary to activation of pancreatic G-protein coupled receptors disseminated extra pancreas too. In patients with type 2 diabetes, the insulin secreting efficiency of GLP-1 results reduced strongly. Exe4 activating GLP-1R mimics most GLP-1 effects. Experimental evidence suggests a possible role for Exe4 in promoting tissue regeneration [61,62].
Photomedicine includes both the study and treatment of diseases caused by exposure to light and on the other hand the diagnostic and therapeutic applications of light for detecting and treating disease. Light energy is capable of cause heating, mechanical effects and chemical reactions. The transfer of light energy through photon absorption can lead to many different consequences in photomedicine. Modern scientific disciplines such as biomedical optics, photochemistry, photobiology, cell biology, laser physics, and engineering have all made major contributions to the development of photomedicine as a fully-fledged division of medical science. The therapeutic uses of light are manifold, phototherapy is used for treating many skin diseases, especially those with immune components, and lasers are used in dermatology, ophthalmology, dentistry and general surgery (among other medical specialties). Lasers still been the in the first line of the treatment of asssisted scarring, most of themablative fractionated as CO2 or Erbio, but also non ablative fractionated laser or vascular lasers are habitually used. However, the role of lasers in assisted wound healing is not as clear as there are few studies [55].
The combination of harmless light in the protoporphyrin absortion spectrum, with non-toxic photosensitizing dyes is used in photodynamic therapy to kill many undesirable cells, including malignant cancer cells and infectious microorganisms [47,62]. The role of PDT in CW has been explored, been probably the main advantge of the technique the possibility of reduce all type of microorganisms inducing radial oxygen singlet (ROS) and without inducing resistence as conventional antibiotics. Besides, PDT induced tissue regeneration and decrease metalloprotenase activity and regenerate collagen [48]. However, the used of PDT as asssited CW is yet not habitual in clinical practise as there are few studies published and multiple repeated sessions are needed with the actual available lights and photosensitizers [62].
Low level laser light therapy (LLLT) consists in an exposition lo low doses of light energy to stimulate or inhibit cellular function leading to beneficial clinical effects, which is called photobiomodulation. LLLT can be applied with low fluences or laser light, but nowadays to simplify the used of the technique, light emitting diodes (LED) have been promoted. The use of low levels of visible or NIR light can reduce pain, inflammation and edema, promote healing, heal tissues and nerves, and preventing tissue damage. Despite many reports of positive findings from experiments conducted in vitro, in animal models, and in randomized controlled clinical trials, LLLT remains controversial. It has proposed that mitochondria is a likely site for the initial effects of light, specifically that the enzyme cytochrome c oxidase (unit four in the mitochondrial respiratory chain) absorbs photons and increases its activity leading to increased ATP production, modulation of reactive oxygen species and induction of transcription factors [63].
LED are revolutionizing the whole lighting industry. Their availability in almost any wavelength and with steadily increasing total output power means that light delivery applications, previously thought to require an expensive laser, can now be performed at a tiny fraction of the cost (less than 1%) by LEDS compared with the equivalent laser source. Not surprisingly, LEDs are becoming much more widely used in medical applications. LEDs have several differences from lasers, however. Firstly, the output wavelengths are much less monochromatic than lasers, with a typical LED having a Full-Width Half-Maximum of 30 nm compared to 2 nm for a laser. Secondly, LED light is non-coherent, so for LLLT so it is not as selective in delivering high energy to the target tissue. Thirdly, the light is non-collimated, and this makes it very difficult to focus it into a fiber optic cable for endoscopic and internal applications (For more details see: [64,65,66]). On the other hand, LEDs have many advantages, apart from the cost, are easy used, induce less damage, could be portable and used at home, are painless and can work in different wavelengths at the same time [64,65,66,67]. Photobiomodulation have been used as complementary therapy of CW, mostly in the initial phases, in which other physical therapies are difficult to apply, decreasing the inflammatory stage and leading to a modulate proliferation and targeting the biofilm [62]. However, further studies are necessary to develop standard protocols, as many devices and light lengths have been applied with good results.
Electrical stimulation is one of the possible adjuvant therapies more promised and in which deeper studies have been published. Some clinical trials have demonstrated the utility of electric fields stimulation in CW as pressure ulcers and leg ulcers. This technique forces the proliferation phase and regeneration mainly in the TGF beta regulation [68]. Ultrasound in different ways of delivery to the target tissue has been also used as a promising treatment [69,70].
New approaches as valuable alternatives could be useful in the treatment and management of wounds. Among the new techniques Bianchi et al. [71] purposed nano-fibrous scaffolds having anti-inflammatory, antibacterial and antioxidant activities. The scaffolds are characterized by aligned nanofibers able to mimic the tendon structure and to promote reconstruction and healing. The nanofibers were produced by electrospinning, an innovative technique, using a biodegradable and biocompatible synthetic polymer, PBCA, combined with copper oxide nanoparticles and CPP.
The resolution of infections is thus an important approach in the promotion of wound healing. However, the use of conventional antibiotics is often unsuccessful due to antibiotic resistance. Lin et al. [72], for example, developed MOFs, which are systems consisting of metal ions (copper in this case) coordinated to organic ligands to form one-, two-, or three-dimensional structures. As a result of their POD-like activity, hydroxyl radicals are produced from hydrogen peroxide. These radicals, together with Cu, possess antibacterial activity, responsible for the fast resolution of infections and thus the acceleration of wound healing. Bachor et al. [70] developed new isoxazole derivatives which showed antibiofilm activity toward Staphylococcus aureus, representing a valuable alternative to conventional broad spectrum antibiotics currently used in therapy which often suffer from antimicrobial resistance. Di Lodovico et al. [74] developed graphene oxide compounds activated by light emitting diodes, which demonstrated antimicrobial activity against Staphylococcus aureus and Pseudomonas aeruginosa-resistant strains in a dual-species biofilm.
Finally, the necessity to evaluate the efficacy of most treatment options also poses a problem in that in vivo studies are limited and, in the case of wounds, injuries to the skin must be induced in animals. Cialdai et al. [75] purposed an ex vivo model for wound healing studies based on a human skin specimen (skin biopsies) mounted in a special chamber equipped with a device able to monitor tissues changes, avoiding the unnecessary use of animals.
14. Future Perspective and Current Limitations
All that has been presented in this review takes on great significance if we consider that wound healing is a process that has yet to be fully understood, although it has been known in practice for a very long time. What is frankly surprising is that the finest cellular mechanisms are still to be resolved and that the aim of numerous authors is to identify a cellular reference point such as fibroblasts or effector cells, ignoring the fact that knowledge of the skin microenvironment is the key to fully understanding this process. Yet, seeing the scientific literature, wound healing is a process of extreme importance and that it undoubtedly carries weight in the various branches of medicine. To all this has been added the study of the cellular mechanisms involved in wound healing in space [75], which still represents an unknown field to say the least. Overall, the process might become fully understood upon gaining full knowledge of the skin microenvironment.
Author Contributions
Conceptualization, writing—review and editing; M.F.-G.; M.L.H.-L.; S.B.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are indebted to Drs K. Mazzaferro, E. Puliti and G. Paroli, for their courtesy to have provided some pictures for this review.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Denomination | Acronym |
| Actin Alpha 2 | ACTA 2 |
| Activator Protein 1 | AP1 |
| Angiopoietin 1 | ANG1 |
| Adrenergic Receptor | AR |
| Calcitonin Gene Related Peptide | CGRP |
| Caseinphosphopeptides | CPP |
| Chemokine (C-X-C motif) ligand | CXCL |
| Chemokine (C--C motif) ligand | CCL |
| Chemokine (C--C motif) receptor | CXCR |
| Chronic Wounds | CW |
| Connective tissue chemokine activating peptide | CTAP |
| Connective tissue growth factor | CTGF |
| Damage Associated Molecular Patterns | DAMPS |
| Dendritic cells | DCs |
| Exendin-4 | Exe4 |
| Extracellular matrix | ECM |
| Epithelial mesenchymal transaction | EMT |
| Epidermal Growth factor | EGF |
| Epidermal stem cellsETS-related gene | ESCERG |
| Extracellular Matrix | ECM |
| Fibroblast growth factor | FGF |
| Glucagon-Like Peptide-1 | GLP-1 |
| Growth-Related Oncogene | GRO |
| Heat shock protein | HSP |
| Hematoxylin Eosin | HE |
| IL | Interleukin |
| Insulin-like growth factor | IGF |
| Interferon | IFN |
| Interferon-g-inducible protein 10 | IP10 |
| Interferon-g-Induced Monokine | MIG |
| Interferon- inducible T cell alpha chemoattractant | ITAC |
| Keratinocyte growth factor | KGF |
| Light-emitting diodes | LED |
| Low Level Light Therapy | LLLT |
| Major Histocompatibility Complex | MHC |
| Mast Cells | MCs |
| Matrix Metalloproteinases | MMPs |
| Macrophage inflammatory protein | MIP |
| Macrophage-derived-chemokine | MDC |
| Metal–organic frameworks | MOFs |
| Mesenchymal stem cellsMonocyte chemoattractant proteins | MSCMCP |
| Monokine-induced gamma interferon | MIG |
| Natural Killer | NK |
| Nerve Growth Factor | NGF |
| Neurokinin A | NKA |
| Neuropeptide Y | NPY |
| Neutrophil-Activating Peptide 2 | NAP-2 |
| Nucleoid-Associated Protein | NAP |
| Non-steroidal anti-inflammatory drugs | NSAID |
| Pathogen-Associated Molecular Patterns | PAMPs |
| Pattern Recognition Receptors | PRR |
| Peroxidase | POD |
| Peroxisome Proliferator Activated Receptors | PPARs |
| Prostaglandine E2 | PGE2 |
| Plasmacytoid dendritic cells | PDCs |
| Platelet-Derived Growth Factor | PDGF |
| Poly(butyl cyanoacrylate) | PBCA |
| Protein Gene Product 9.5 | PGP 9.5 |
| Regulated Upon activation normal T-cell expressed and secreted | RANTES |
| Reactive oxygen species | ROS |
| Signal Transducer and Activator of Transcription 3 | STAT 3 |
| Smooth Muscle Actin | SMA |
| Stem Cell Factor | SCF |
| Stromal-derived growth factor | SDF |
| Substance P | SP |
| Tissue Inhibitor of Metalloproteinases | TIMP |
| Toll-like receptor | TLR |
| Transforming Growth Factor | TGF |
| Thrombospondin | TSP |
| Tumor Necrosis Factor | TNF |
| Ulex Europaeus Agglutinin | UEA |
| Vascular Endothelial Growth Factor | VEGF |
| Vasoactive intestinal peptide | VIP |
| Wound Healing | WH |
References
- Han, G. and Ceilley, R., Chronic wound healing: a review of current management and treatments. Adv Ther, 2017. 34(3): p. 599-610. [CrossRef]
- Sen, C.K. , Human wounds and its burden: an updated compendium of estimates. Adv Wound Care (New Rochelle), 2019. 8(2): p. 39-48. [CrossRef]
- D’Arcy, C. and Kiel, C., Cell adhesion molecules in normal skin and melanoma. Biomolecules, 2021. 11(8). p. 1213. [CrossRef]
- Bacci, S. , Cellular mechanisms and therapies in wound healing: looking toward the future. Biomedicines, 2021. 9(11). p.1611. [CrossRef]
- Gupta, S. , et al., Management of chronic wounds: diagnosis, preparation, treatment, and follow-up. Wounds, 2017. 29(9): p. S19-S36.
- Babalska, L. Z, et al., Wound antiseptics and european guidelines for antiseptic application in wound treatment. Pharmaceuticals 2021. 14(12): p. 1253. [CrossRef]
- Takeo, M. , Lee, W., and Ito, M., Wound healing and skin regeneration. Cold Spring Harb Perspect Med, 2015. 5(1): p. a023267. [CrossRef]
- Braiman-Wiksman, L. , et al., Novel insights into wound healing sequence of events. Toxicol Pathol, 2007. 35(6): p. 767-79. [CrossRef]
- Chen, L. , et al., Inflammatory responses and inflammation-associated diseases in organs. Oncotarget, 2018. 9(6): p. 7204-7218. [CrossRef]
- Nguyen, A.V. and Soulika, A.M., The dynamics of the skin’s immune system. Int J Mol Sci, 2019. 20(8). p.1611. [CrossRef]
- Canedo-Dorantes, L. and Canedo-Ayala, M., Skin acute wound healing: a comprehensive review. Int J Inflam, 2019. 2019: p. 3706315. [CrossRef]
- Gold, M.H. , et al., Updated international clinical recommendations on scar management: part 2--algorithms for scar prevention and treatment. Dermatol Surg, 2014. 40(8): p. 825-31.
- Etulain, J. , Platelets in wound healing and regenerative medicine. Platelets, 2018. 29(6): p. 556-568. [CrossRef]
- Bacci, S. , Fine regulation during wound healing by mast cells, a physiological role not yet clarified. Int J Mol Sci, 2022. 23(3): p.1820. [CrossRef]
- Tyavambiza, C. , Meyer, M., and Meyer, S., Cellular and molecular events of wound healing and the potential of silver based nanoformulations as wound healing agents. Bioengineering (Basel), 2022. 9(11). p.712. [CrossRef]
- Krzyszczyk, P. , et al., The role of macrophages in acute and chronic wound healing and interventions to promote pro-wound healing Phenotypes. Front Physiol, 2018. 9: p. 419. [CrossRef]
- Bacci, S. , et al., Immunohistochemical analysis of dendritic cells in skin lesions: correlations with survival time. Forensic Sci Int, 2014. 244: p. 179-85. [CrossRef]
- Han, Z. , et al., MiR-21/PTEN Axis Promotes Skin Wound Healing by Dendritic Cells Enhancement. J Cell Biochem, 2017. 118(10): p. 3511-3519. [CrossRef]
- Bordon, Y. , Dendritic cells: pDCs play off scratch. Nat Rev Immunol, 2011. 11(1): p. 8. [CrossRef]
- Nosbaum, A. , et al., Cutting edge: regulatory t cells facilitate cutaneous wound healing. J Immunol, 2016. 196(5): p. 2010-4. [CrossRef]
- Giantulli, S. , et al., Effect of 1-MHz ultrasound on the proinflammatory interleukin-6 secretion in human keratinocytes. Sci Rep, 2021. 11(1): p. 19033. [CrossRef]
- Piipponen, M. , Li, D., and Landen N.X., The immune functions of keratinocytes in skin wound healing. Int J Mol Sci, 2020. 21(22). [CrossRef]
- Morbidelli, L. , Genah, S., and Cialdai, F., Effect of microgravity on endothelial cell function, angiogenesis, and vessel remodeling during wound healing. Front Bioeng Biotechnol, 2021. 9: p. 720091. [CrossRef]
- Johnson, K.E. and Wilgus,T.A., Vascular endothelial growth factor and angiogenesis in the regulation of cutaneous ound repair. Adv Wound Care (New Rochelle), 2014. 3(10): p. 647-661. [CrossRef]
- Bodnar, R.J. , et al., Perycites: a newly recognized player in wound healing. Wound Repair Regent, 2016. 24(2): p.204-14. [CrossRef]
- Cialdai, F. , Risaliti, C., and Monici, M., Role of fibroblasts in wound healing and tissue remodeling on earth and in space. Front Bioeng Biotechnol, 2022. 10: p. 958381. [CrossRef]
- Monika, P. , et al., Myofibroblast progeny in wound biology and wound healing studies. Wound Repair Regen, 2021. 29(4): p. 531-547. [CrossRef]
- Schultz, G.S. , et al., Principles of wound healing, in mechanisms of vascular disease: A reference book for vascular specialists, R. Fitridge and M. Thompson, Editors. 2011: Adelaide (AU).Available from: https://www.ncbi.nlm.nih.gov/books/NBK534261.
- Qing, C. , The molecular biology in wound healing & non-healing wound. Chin J Traumatol, 2017. 20(4): p. 189-193. [CrossRef]
- Olson, T.S. and Ley, K., Chemokines and chemokine receptors in leukocyte trafficking. Am J Physiol Regul Integr Comp Physiol, 2002. 283(1): p. R7-28. [CrossRef]
- Brandt, E. , et al., The beta-thromboglobulins and platelet factor 4: blood platelet-derived CXC chemokines with divergent roles in early neutrophil regulation. J Leukoc Biol, 2000. 67(4): p. 471-8. [CrossRef]
- Gillitzer, R. and Goebeler, M., Chemokines in cutaneous wound healing. J Leukoc Biol, 2001. 69(4): p. 513-21.
- Murdoch, C. and Finn, A., Chemokine receptors and their role in inflammation and infectious diseases. Blood, 2000. 95(10): p. 3032-43.
- Yamamoto, T. , et al., Monocyte chemoattractant protein-1 enhances gene expression and synthesis of matrix metalloproteinase-1 in human fibroblasts by an autocrine IL-1 alpha loop. J Immunol, 2000. 164(12): p. 6174-9. [CrossRef]
- Rennekampff, H.O. , et al., Bioactive interleukin-8 is expressed in wounds and enhances wound healing. J Surg Res, 2000. 93(1): p. 41-54. [CrossRef]
- Engelhardt, E. , et al., Chemokines IL-8, GROalpha, MCP-1, IP-10, and Mig are sequentially and differentially expressed during phase-specific infiltration of leukocyte subsets in human wound healing. Am J Pathol, 1998. 153(6): p. 1849-60. [CrossRef]
- Kulke, R. , et al., The CXC receptor 2 is overexpressed in psoriatic epidermis. J Invest Dermatol, 1998. 110(1): p. 90-4. [CrossRef]
- Goebeler, M. , et al., The chemokine repertoire of human dermal microvascular endothelial cells and its regulation by inflammatory cytokines. J Invest Dermatol, 1997. 108(4): p. 445-51. [CrossRef]
- Strieter, R.M. , et al., The functional role of the ELR motif in CXC chemokine-mediated angiogenesis. J Biol Chem, 1995. 270(45): p. 27348-57. [CrossRef]
- Belperio, J.A. , et al., CXC chemokines in angiogenesis. J Leukoc Biol, 2000. 68(1): p. 1-8.
- Randolph, G.J. and Furie, M.B., A soluble gradient of endogenous monocyte chemoattractant protein-1 promotes the transendothelial migration of monocytes in vitro. J Immunol, 1995. 155(7): p. 3610-8.
- Yoshida, M. , et al., TGF-beta-operated growth inhibition and translineage commitment into smooth muscle cells of periodontal ligament-derived endothelial progenitor cells through Smad- and p38 MAPK-dependent signals. Int J Biol Sci, 2012. 8(7): p. 1062-74. [CrossRef]
- Li, C. , et al., Notch signal regulates corneal endothelial-to-mesenchymal transition. Am J Pathol, 2013. 183(3): p. 786-95. [CrossRef]
- Bacci, S. and Bani, D., The epidermis in microgravity and unloading conditions and their effects on wound healing. Front Bioeng Biotechnol, 2022. 10: p. 666434. [CrossRef]
- Raziyeva, K. , et al., Immunology of acute and chronic wound healing. Biomolecules, 2021. 11(5). [CrossRef]
- Falanga, K. , et al., Chronic wounds. Nat Rev Dis Primers. 2022. 8(1):p. 50. [CrossRef]
- Kadam, S. , et al., Bioengineered platforms for chronic wound infection studies: how can we make them more human-relevant? Front Bioeng Biotechnol, 2019. 7: p. 418. [CrossRef]
- Grandi, V. , et al., Cellular mechanisms in acute and chronic wounds after PDT therapy: An update. Biomedicines, 2022. 10(7):p.1624. [CrossRef]
- Tottoli, E.M. , et al., Skin wound healing process and new emerging technologies for skin wound care and regeneration. Pharmaceutics, 2020. 12(8): p.735. [CrossRef]
- Andrews, J.P. , et al., Keloids: The paradigm of skin fibrosis - Pathomechanisms and treatment. Matrix Biol, 2016. 51: p. 37-46. [CrossRef]
- Huang, J. , et al., Combined analyses of RNA-sequence and Hi-C along with GWAS loci-A novel approach to dissect keloid disorder genetic mechanism. PLoS Genet, 2022. 18(6): p. e1010168. [CrossRef]
- Lingzhi, Z. , Meirong, L., and Xiaobing, F., Biological approaches for hypertrophic scars. Int Wound J, 2020. 17(2): p. 405-418. [CrossRef]
- Jiang, D. and Rinkevich, Y., Scars or regeneration?-dermal fibroblasts as drivers of diverse skin wound responses. Int J Mol Sci, 2020. 21(2). p. 617. [CrossRef]
- Berman, B. , Maderal, A., and Raphael, B., Keloids and hypertrophic scars: pathophysiology, classification, and treatment. Dermatol Surg, 2017. 43 Suppl 1: p. S3-S18. [CrossRef]
- Leszczynski, R. , et al., Laser therapy for treating hypertrophic and keloid scars. Cochrane Database Syst Rev 2022. 26 (9):p.CD011642. [CrossRef]
- Alghamdi, M.A. , et al., Identification of differentially methylated CpG sites in fibroblasts from keloid scars. Biomedicines, 2020. 8(7). p. 181. [CrossRef]
- Yang, R. , et al., Epidermal stem cells in wound healing and regeneration. Stem cells International, 2020. p.9148310. [CrossRef]
- Isakson, M. , et al., Mesenchymal stem cells and cutaneous wound healing: current evidence and future potential. Stem cells International, 2015. p. 831095. [CrossRef]
- Kua, J. , et al., Human umbilical cord lining-derived epithelial cells: a potential source of non-native epithelial cells that accelerate healing in a porcine cutaneous wound model. Int J Mol Sci, 2022. 23(16): p. 8918. [CrossRef]
- Khalil, H. , et al., Medications affecting healing: an evidence-based analysis. Int Wound J, 2017. 14(6): p.1340-1345. [CrossRef]
- Bacci, S. , et al., The pro-healing effect of exendin-4 on wounds produced by abrasion in normoglycemic mice. Eur J Pharmacol, 2015. 764: p. 346-352. [CrossRef]
- Paroli, G. , et al., The role of mast cells in cellular modifications evoked by Exendin-4 in treated wounds: a preclinical study. J Wound Care, 2022. 31(8): p.701-708. [CrossRef]
- Fernández Guarino, M. , et al., Methyl aminolaevulinic acid versus aminolaevulinic acid photodynamic therapy of actinic keratosis with low doses of red-light LED Illumination: results of long-term follow-up. Biomedicines 2022. 10 :p 3218. [CrossRef]
- Franca, C.M. , et al., Photobiomodulation in wound healing: what are we not considering? Photomed Laser Surg 2016. 34(2):p51-52,2016. [CrossRef]
- Cicchi, R. , et al., Observation of an improved healing process in superficial skin wounds after irradiation with a blue-LED haemostatic device. J Biophotonics 2016. 9(6): p.645-655. [CrossRef]
- Magni, G. , et al., Blue LED light modulates inflammatory infiltrate and improves the healing of superficial wounds. Photodermatol Photoimmunol Photomed. 2020. 36(2):p.166-168. [CrossRef]
- Magni, G. , et al., Blue-LED-light photobiomodulation of inflammatory responses and new tissue formation in mouse-skin wounds. Life (Basel). 2022.12(10): p.1564. [CrossRef]
- Nguyen, J.K. , et al., A dose-ranging, parallel group, split-face, single-blind phase II study of light emitting diode-red light (LED-RL) for skin scarring prevention: study protocol for a randomized controlled trial. Trials 2019. 20(1):p.432. [CrossRef]
- Polak, A. , et al., A randomized, controlled clinical study to assess the effect of anodal and cathodal electrical stimulation on periwound skin blood flow and pressure ulcer size reduction in persons with neurological injuries. Ostomy Wound Manage. 2018. 64(2):p. 10-29.
- Alkahtani, S. A. , et al., Ultrasound-based techniques as alternative treatments for chronic wounds: a comprehensive review of clinical applications. Cureus. 24(12): p.e1952. [CrossRef]
- Bora, K. , et al., High-voltage electrical stimulation versus ultrasound in the treatment of pressure ulcers. Adv Skin Wound Care. 2017;30(12):p.565-570. [CrossRef]
- Bianchi, E. , et al.,Electrospun scaffolds based on poly(butyl cyanoacrylate) for tendon tissue engineering. Int J Mol Sci 2023. 24 (4): p. 3172. [CrossRef]
- Lin, C. , et al., Different dimensional copper-based metal–organic frameworks with enzyme-mimetic activity for antibacterial therapy. Int J Mol Sci 2023. 24:p. 3173. [CrossRef]
- Bachor, U., et al.,The in vitro impact of isoxazole derivatives on pathogenic biofilm and cytotoxicity of fibroblast cell line. Int J Mol Sci. 2023. 24, 2997. Int J Mol Sci. 2023, 24, 2997. [CrossRef]
- Di Lodovico, S., et al., Antimicrobial combined action of graphene oxide and light emitting diodes for chronic wound management. Int J Mol Sci. 2022, 23, 6942. [CrossRef] [PubMed]
- Cialdai, F., et al., Optimization of an ex-vivo human skin/vein model for long-term wound healing studies: ground prepa ratory activities for the ‘Suture in Space’ experiment onboard the international space station. Int J Mol Sci. 2022, 23, 14123. [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



