
Submitted:
09 May 2023
Posted:
10 May 2023
You are already at the latest version
Abstract
Genetic, developmental, biochemical, and environmental variables interact intricately to produce sex differences. The significance of sex differences in cancer susceptibility is being clarified by numerous studies. Epidemiological research and cancer registries have revealed over the past few years that there are definite sex variations in cancer incidence, progression, and survival. However, oxidative stress and mitochondrial dysfunction also have a significant impact on the response to treatment of neoplastic diseases. Young women may be more protected from cancer than men because most of the proteins implicated in the regulation of redox state and mitochondrial function are under the control of sexual hormones. In this review, we describe how sexual hormones control the activity of antioxidant enzymes and mitochondria, as well as how they affect several neoplastic diseases. The molecular pathways that underlie the gender-related discrepancies in cancer that have been identified may be better understood, which may lead to more effective precision medicine and vital information on treatment options for both males and females with neoplastic illnesses.
Keywords:
gender differences
; cancer
; oxidative stress
; oestrogens
; testosterone
; antioxidant
; reactive oxygen species
; sex hormones
; mitochondria.
1. Introduction
1.1. General considerations on gender differences in cancer susceptibility and oxidative stress
Epidemiological studies consistently demonstrate that there are gender variations in cancer incidence and mortality [1,2]. According to an analysis of the IARC's global cancer statistics, men were more likely than women to develop cancer in 32 of 35 tumour sites. The scientists concluded that the causes of the significant gender differences are unknown because in thirteen of these sites the discrepancies could not be accounted for by known risk variables [3]. Another study found that the incidence of cancer in non-reproductive organs is 1.8 times higher in men than in women [4], and animal studies have revealed gender disparities in cancer incidences, even in rodents who were not exposed to any harmful substances [5]. A study found 68 "male-specific" and 19 "female-specific" carcinogens (carcinogens that caused cancer), albeit the exact causes of this difference are still unknown [6]. However, according to a preliminary review of the literature on these gender-specific carcinogens, oxidative stress might be a key mechanism, particularly for male-specific carcinogens. Thus, it was hypothesized that oxidative stress might affect a patient's susceptibility to developing cancer from chemical carcinogens, which was supported by the literature on gender disparities [7].
The imbalance between the production of toxic reactive species (TRS) and antioxidant defence mechanisms is referred to as "oxidative stress". Reactive oxygen species (ROS) and reactive nitrogen species (RNS) are two categories of TRS. Superoxide anion, hydrogen peroxide, singlet oxygen, hydrogen superoxide, and reactive hydroxyl radical are the main components of ROS. The two RNS that are most known are nitric oxide and peroxynitrite [8]. Since ROS can damage DNA, proteins, and lipids within cells, oxidative stress is regarded as a serious condition. In cancer cells, TRS can come from a variety of sources. The chief contributors to the generation of ROS are thought to be an active metabolism and issues with the mitochondrial respiratory chain [9]. Additionally, activated macrophages infiltrate cancer tissue, increasing the inflammatory state and escalating the production of ROS and cytokines [10]. Moreover, the activation of oncogenes like RAS2 or c-Myc is one example of a disorder in cellular signaling that is thought to be a significant generator of ROS [11,12]. For cells, oxidative stress may be damaging, but intrinsic oxidative stress in cancer cells in malignant neoplasms may have dramatic effects, including cancer cell proliferation, the promotion of genetic instability, changes in cellular sensitivity to anticancer agents, and modulation of cellular redox parameters is a real possibility [13,14,15,16,17,18] (Figure 1).
In a study, the literature on twenty-six recognized human carcinogens (IARC group 1) was examined. The Tox21 data resembled those of male-specific rat carcinogens, and oxidative stress was more commonly recorded for the bulk of these carcinogens. As a result, this analysis, which was based on about 600,000 scholarly abstracts and Tox21 screening assays, suggested a connection between testosterone, oxidative stress, and male-specific cancers. This indicates that the increased cancer susceptibility seen in men may be due to a distinct cellular response to oxidative stress in men compared to women [19].
Males are thought to experience oxidative stress more frequently than females [20]. In many other species, including flies, mice, and rats [21,22,23], this notion has been confirmed. Depending on the cell type or tissue under investigation, however, a range of contentious findings have been found. It has been noted that longer-living female Drosophila melanogaster exhibit lower ROS levels and higher antioxidant enzymes than males, which may explain their reported longer lifetime [21]. In the livers of rats and mice [24], as well as the brain and heart of mice, it has been observed that female mitochondria release less superoxide radicals than those of men. According to the "Mitochondrial theory of ageing" [25], females produce less ROS than men, which is consistent with the longer lifetime that females exhibit because the rate of oxidant production by mitochondria from long-lived species is significantly lower than that of short-lived species [26,27,28].
Other experimental data seem to confirm this hypothesis. NADPH-oxidase enzymes are noteworthy for the evaluation of how gender differences affect the determinism of various oxidative stresses because they are the only oxidant enzymes that are specifically designed to produce ROS, as opposed to other oxidant enzymes that do so as a by-product of another reaction. As a result, they might have a bigger impact on the varied oxidative stress status between the sexes. NADPH-oxidase activity and function are lower in females, according to several studies [29]. This is the result of two mechanisms: first, oestrogens can directly cause reduced NADPH-oxidase activity in females, and second, females have lower levels of p47, which is necessary for the assembly of the NADPH-oxidase enzyme and lower levels of superoxide production independently of oestrogens. Females with lower levels of oxidative stress have lower levels of superoxide because of these gender differences.
Numerous researchers have reported other various explanations for the alteration of oxidative stress brought on by sex. In addition to gender differences in ROS generation, clinical and experimental studies have indicated that women have stronger antioxidant potential than men [30], and this may be because oestrogen has antioxidant qualities, making women less vulnerable to oxidative stress [31]. Since postmenopausal women do not benefit from oestrogen’s anti-inflammatory and antioxidant protective properties, they are more likely to experience increased oxidative stress [32]. Superoxide dismutase (SOD) transforms superoxide anion into hydrogen peroxide, and it has been hypothesized that different tissues may react differently to this process. Female rats were shown to have higher heart SOD activity levels than male rats, and it was noted that the levels of SOD activity in the brain and lungs were higher in females [22]. Surprisingly, castration significantly reduced the levels of SOD activity in both male and female rats compared to the corresponding controls [22], indicating that sex hormones may be related to SOD activity levels. SOD activity was reported to be higher in female erythrocytes than male erythrocytes in one human experiment [33] and in female plasma than male plasma in another [33].
Glutathione peroxidase (GPx) is another enzyme that detoxifies hydrogen peroxide into water and oxygen. Progesterone and testosterone and other sex hormones control how active the GPx enzyme is. They are tissue-dependent in terms of its sex-related differences. While male mice displayed higher GPx activity than female mice in the heart, female mice did so in the kidney and brain [34]. Additionally, previous research revealed higher GPx activity in the female brain and liver [35] and that the activity of GPx was more than twice as high in the hepatic mitochondria of female rats as compared to male rats of the same age [36]. In addition, females had more GPx activity in their livers than did men [37], and in humans, teenage girls had higher GPx activity in their blood than did men [38].
Since oestrogen replacement therapy increased erythrocyte GPx activity significantly in postmenopausal women and showed a positive correlation between GPx and serum oestrogen levels in both premenopausal and postmenopausal women, it is likely that oestrogens stimulate GPx expression. This theory is supported by the finding that total hysterectomy in pre-menopausal females reduced the mRNA expression of SOD and GPx, which was then restored by oestrogen replacement treatment, but had no effect on the expression of catalase [39].
Hepatic mitochondrial GSH levels in female rats are higher than in male rats, which is relevant to decreased glutathione (GSH) concentration. However, after ovariectomization, the levels of mitochondrial glutathione in the rats dropped to levels comparable to those in males. This decrease in mitochondrial GSH was totally reversed by oestrogen replacement therapy [24], indicating that oestrogen levels may regulate GSH levels. Adolescent girls in humans likewise had a greater blood GSH/oxidized glutathione (GSSG) ratio than did men [38] while men presented statistically significantly higher values of oxidized-reduced GSSG/GSH and GSSG in terms of concentration than did women [40]. Additionally, premenopausal women who had hysterectomies demonstrated a decrease in GSH concentration and an increase in GSSG and GSSG/GSH ratio after 30 days. It is interesting that oestrogen replacement therapy brought glutathione levels back to what they were prior to the hysterectomy, highlighting once more the critical function oestrogens play in the glutathione cycle [39].
Finally, obesity is a subject that is particularly intriguing in terms of the connection between gender differences, oxidative stress, and the development of cancer. With a significant global variation depending on the prevalence of obesity and the relative risk estimates, the burden of cancer attributable to obesity, expressed as population attributable fraction (PAF), is 11.9% in men and 13.1% in women for all obesity-related malignancies worldwide [41]. The biggest PAF is typically seen in cases of oesophageal adenocarcinoma in men and endometrial cancer in women. The overall strong relationship between obesity and gynaecologic cancer (endometrial, postmenopausal breast, and ovarian cancers) suggests that female sex hormones have a role in the aetiology of cancer. The risks for various cancers, including colon, rectal, gallbladder, kidney, and pancreatic cancers, are associated differently by gender depending on BMI and other somatometric factors [42,43]. For instance, there is high support for the association between BMI and colon cancer in males and weak support in females [17]. This discovery emphasizes the harmful impact of visceral adiposity and insulin resistance in colon cancer as well as the protective benefits of endogenous estrogenic effects against colon cancer in women [44,45,46]. Men are more likely than women to develop visceral adiposity.
Obesity, on the other hand, is a chronic inflammatory state [47] that has been implicated as a factor in the initiation and progression of cancer [48] due to the presence of numerous inflammatory components in the tumour microenvironment that support a malignant phenotype. Obese patients with metabolic abnormalities and adipose inflammation have a higher chance of developing cancer [49]. Tumour promotion is aided by ROS generation, which has been linked to obesity [50]. The formation of ROS and the release of pro-inflammatory cytokines are induced by hyperglycaemia combined with increased levels of free fatty acids, which together cause mitochondrial and DNA damage [51]. Furthermore, the folding of proteins is affected by oxidative stress. Obese people have higher amounts of free fatty acids (FFAs), which are linked to endoplasmic reticulum (ER) stress in adipocytes [52]. Particularly, FFAs cause the production of ROS, which oxidize proteins and raise the proportion of unfolded proteins in ER. An inflammatory reaction is brought on by the build-up of unfolded proteins [53]. This process' cytokines have been connected to colon cancer [54]. All the above could help explain the different incidence of obesity-induced neoplasms in the two sexes.
2. Molecular mechanisms of sex disparities
The molecular processes underlying the links between gender disparities, oxidation, and the development of cancer have been defined in certain research. The main operating factor of the cancer defence, the tumour suppressor protein p53, plays a crucial role in protecting against long-term DNA damage. As a transcription factor, p53 promotes the expression of its target genes by interacting with DNA responsive sequences in their regulatory regions [55]. Through the p53-DREAM pathway, which includes its primary transcriptional target Cyclin Dependent Kinase Inhibitor 1A, also known as P21, encoded in CDKN1A, it also can suppress other sets of genes [56]. There is growing evidence that certain cancer sex disparities are related to differences in p53 functional abilities between males and females [57]. This suggests that either innate or externally imposed effects prevent p53 from conducting its functions equally between the sexes.
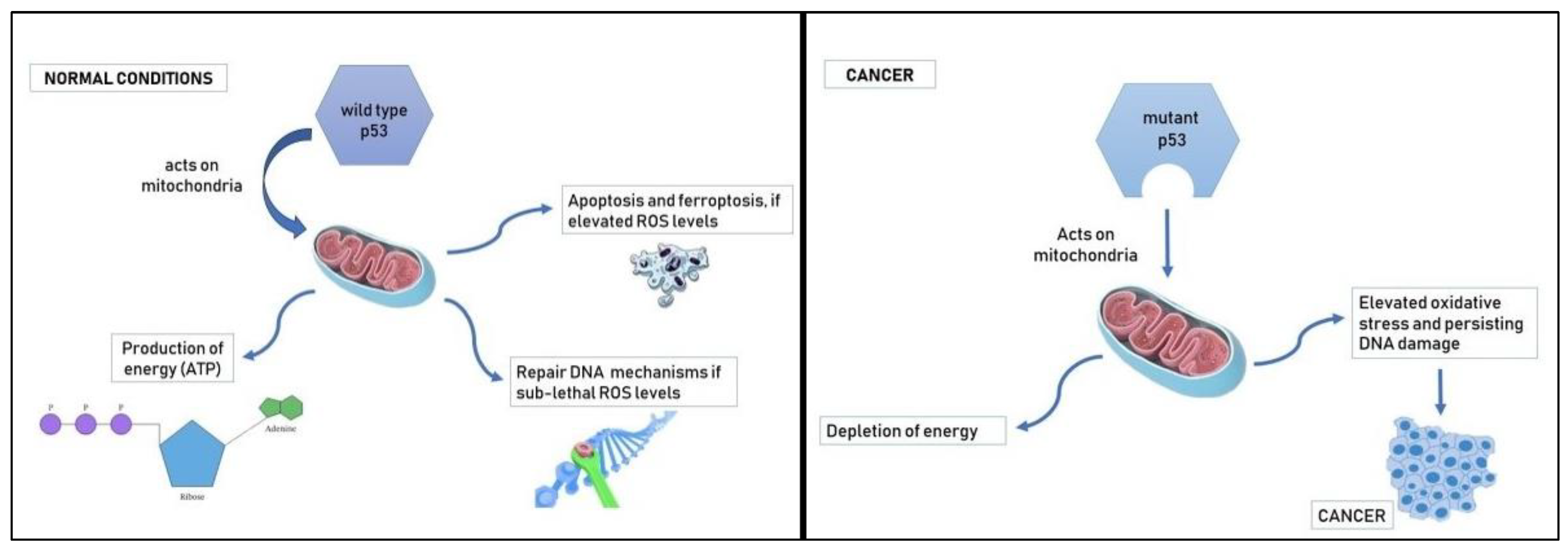
Through its exonuclease activity and transcription factor transactivation function, wild type (wt) p53 encourages mitochondrial biogenesis [58]. In the absence of wt p53, mitochondria are consistently depleted [59]. An important unanswered question regarding cancer and gender inequality is how mutant p53 affects the mitochondria. In addition to producing the cellular energy known as adenosine triphosphate (ATP), mitochondria also play a role in the pathways that lead to cell death. Normal wt p53 controls metabolism by physically interacting with and functionally altering mitochondrial components in situ in addition to controlling respiration through transcriptional transactivation of its targets [60]. Additionally, wt p53 prevents mitochondrial DNA mutation [59]. This occurs in structures with their own DNA that are inherited from the mother. Due to their solely maternal origin, this strengthens the idea that mitochondria are more adapted to female contexts than to male contexts [61]. As a result, p53 is closely related to redox reactions [62], and elevated ROS encourage the accumulation of p53. Acute ROS signaling plays a crucial role in the p53 activation pathways that lead to ferroptosis or programmed cell death (apoptosis) [63]. Sub-lethal ROS concentrations can prepare wild type p53 to activate repair pathways throughout the course of a brief cell cycle arrest. The formation of cancer is, however, at danger from sustained ROS levels at sub-lethal dosages due to DNA mutation, including TP53 mutation [64].
Figure 2.
Mutant p53 is associated with a reduction in ATP production, elevated oxidative stress and a persisting DNA damage which cause cancerogenesis.
Figure 2.
Mutant p53 is associated with a reduction in ATP production, elevated oxidative stress and a persisting DNA damage which cause cancerogenesis.
We shall attempt to assess how the differences in gender and oxidative stress can affect the start or progression of neoplastic disorders in the section that follows.
3. Sex differences in oxidative stress and neoplastic diseases
3.1. Glioma, oxidative stress and gender differences
Although it is the second most common cancer in children, brain cancer is an uncommon condition in comparison to other cancer types. 1.4% of all new cancer cases, or roughly 23700 new cases per year, are reported [65]. Men are twice as likely to develop medulloblastoma, ependymoma, and gliomas than women, according to epidemiologic research [66]. In addition, a recent study found that women outlived males and responded better to standard treatment, identifying transcriptome signatures for glioblastomas in women [67].
Since oxidative stress and inflammation are also involved in the onset and progression of brain cancer, substances able to modify oxidative stress such as phytoestrogens have been considered good candidates for brain cancer prevention due to their antioxidant and anti-inflammatory properties. In fact, consuming foods containing phytoestrogens, particularly daidzein, appears to have a protective effect against gliomagenesis, according to an epidemiologic study conducted in 2006 [68]. Additionally, new research has shown that the phytoestrogens formononetin or biochanin A and the cytotoxic drug temozolomide combined have an enhanced anticancer effect in glioblastoma multiforme cells, with greater inhibition of cell signaling and invasion pathways, and restoration of mitochondrial function [69,70].
Furthermore, long-term research has been done on the effects of gender on oxidative stress in the brain, including free radical generation, oxidative damage, and antioxidant enzyme levels and/or activity [71]. According to certain studies [72,73,74,75,76,77,78,79,80,81], male rats have greater DNA, protein, and lipid oxidative damage than female rats. The increased ROS generation in male rats [82,83] and the decreased levels and/or activities of antioxidant enzymes [84,85,86,87,88] are the causes of this oxidative damage. However, although these studies suggest that female rats have better redox homeostasis than male rats, other reports [89,90,91,92] have found no differences.
In terms of sex hormones, 17-estradiol (E2) and progesterone, which are produced by females, have neuroprotective effects in vivo and in vitro at physiological concentrations [93,94,95,96], but androgens and testosterone, which are produced by males, typically have neurotoxic effects [97]. The ability of some neurons and glial cells to create neurosteroids—sex hormones that are often produced de novo and independently of peripheral tissues—is particularly intriguing. These neurosteroids are equivalent to circulating steroids in both chemical and biological terms [98,99].
Along with oxidative stress, brain tumorigenesis has also been linked to decreased responses from nonenzyme (reduced glutathione, GSH) and enzyme antioxidant systems (SOD, catalase, and GPx) [100]. Since the central nervous system (CNS) is extremely susceptible to free-radical damage, an imbalance between the production of free radicals and the effectiveness of the antioxidant defence systems is able to initiates the neoplastic process [101]. This theory was supported by numerous research. For instance, research has shown that subcutaneous administration of hydroxytyrosol, but not oleuropein or a combination of both compounds, resulted in a significant inhibition of tumour growth through mechanisms involving endogenous enzymatic and non-enzymatic antioxidant defence systems. This was shown by the declines in oxidative stress biomarkers, such as thiobarbituric acid reactive subs, in an animal model of C6 glioma implanted at the subcutaneous region. As a result, the hydroxytyrosol therapy positively altered the enzymatic antioxidant defence systems while maintaining the non-enzymatic antioxidant defence systems at a level comparable to that of the healthy animals [102]. Oleuropein, which is a stronger antioxidant than hydroxytyrosol, on the other hand, lacked antitumor effects and even accelerated tumour growth. This suggests that hydroxytyrosol has additional effects besides altering antioxidant defence systems and may exert its antitumor effects via unidentified additional mechanisms. Similar results were confirmed by other studies [103]. Once more, this data lends support to the chemicals' alleged concurrent or alternate mechanisms of action to their antioxidant ones. In fact, none of the phenolic compounds, either alone or in combination, were able to return GSH levels to those seen in control.
In any case, the gender of the animals has a significant impact on these effects, which are caused by redox regulation mechanisms including endogenous enzymatic and non-enzymatic antioxidant defence systems [103,104]. Thus, the existence of gender differences in processes related to brain tumours, such as the management of redox status, suggested that research on brain cancer should take gender differences into account in preclinical studies, screening and prevention programs, as well as in therapeutic approaches.
3.2. Liver cancer, oxidative stress and gender differences
With 521,000 deaths per year, or 6.4% of all deaths, liver cancer is currently the second most common cancer type [105]. The 5-year survival rate for people with liver cancer only oscillates by 10% despite the use of intensive treatments [106]. 90% of liver cancer cases are caused by hepatocellular carcinoma (HCC).
Even after accounting for variations in exposure to risk factors, there is a 2- to 4-fold higher incidence of liver cancer in men than in women in humans [107,108]. Additionally, males predominate in transgenic mouse models of hepatitis virus infection and models of liver tumour induction in mice after exposure to chemical carcinogens such as AFB1, 4-aminobiphenyl (ABP), and diethylnitrosamine (DEN) [109]. Additionally, numerous human and animal studies on HCC confirmed sexual dimorphism during the onset and development of alcohol liver disease (ALD). It is likely that variations in the expression of genes that code for ethanol-metabolizing enzymes have an impact on the development and progression of ALD and liver cancer [110]. Alcohol-dehydrogenase (ADH) activity varies between sexes; it is lower in men than in women, which leads to less acetaldehyde build-up. Additionally, studies reveal that oestrogens positively affect CYP2E1 and ADH, indicating that ethanol should be metabolized more quickly in females than in males [111].
In a study, a single dose of DEN was administered to new-born male and female mice. Later, animals were given drinking water containing 10/20% (v/v) ethanol on alternate days (EtOH-DW) [112]. At the conclusion of the trial, liver pathology/function and cytokine expression were examined in liver tissue and serum. In both male and female mice, DEN consistently caused hepatic foci/tumours. Alcohol reduced liver function and exacerbated liver damage, but it did not cause the formation of hepatic foci-HCC on its own. Ethanol significantly increased tumour incidence and burden in DEN-initiated EtOH-DW animals, but only in male mice. At necropsy, male and female mice had similar blood-alcohol contents, but increased liver damage and decreased liver function/antioxidant capacity were significantly higher in males. Male mice had higher levels of SMAD3 in response to EtOH, DEN-initiation, and DEN+EtOH-DW, according to analysis of hepatic mRNA. These results show that male mice are more vulnerable than female mice to HCC incidence and progression during chronic ethanol feeding. Increased TGF-SMAD3 signaling may enhance promotion in this model of HCC progression, effects that are modified by prolonged ethanol feeding, according to differences in hepatic immune response markers in male mice [112].
It should be mentioned that lipid peroxide levels in the liver and serum are decreased by oestradiol and its derivatives, which are potent endogenous antioxidants [113,114]. The loss of SOD and glutathione peroxidase activity, as well as iron (ferric nitrilotriacetate)-induced ROS production, lipid peroxidation, activation of AP-1 and NF-B, are all suppressed by oestradiol in cultured rat hepatocytes, according to recent research [115,116]. In isolated rat liver mitochondria, oestradiol also reduces the lipid peroxidation brought on by iron [115]. These results imply that the inhibitory impact of oestradiol on AP-1 and NF-B activation may result from scavenging ROS and/or from lowering intracellular ROS generation by inducing antioxidant enzymes.
Male sex, like the viral risk factor for hepatic fibrosis, is a significant risk factor for HCC [117], while it is unknown whether males and females differ in their susceptibility to the integration of viral DNA, which causes the malignant transformation of hepatocytes. In contrast, premenopausal women are least susceptible to HCC because they lack the risk factors of older age and male sex. In a study, 901 individuals with HBV-associated HCC had their age-specific male to female ratios looked at. The younger group had a smaller percentage of females (10.5%) than the older group when the subjects were split into two age groups based on whether they were younger or older than the menopausal age of 50 years.
The differences in hepatic damage that were found are connected to alterations in cellular GSH, ROS production, and cell REDOX status brought on by the metabolism of ethanol. The imbalance between acetaldehyde and ALDH is accentuated by CYP2E1 induction, which also leads to the production of ROS, the subsequent depletion of GSH, and oxidative damage [112].
An acute increase in the Ser139 phosphorylation of histone H2AX, which results in the production of gH2AX, an early hallmark of double-stranded DNA breaks, was seen in a different study that exposed postnatally exposed mice to a tumorigenic dosage of 4-aminobiphenyl (ABP). The difference between males and females in the rise could be explained by the higher antioxidant gene expression responses in females than in males as mediated by nuclear factor erythroid 2-related factor 2 (NRF2). To combat liver oxidative stress, NRF2 nuclear accumulation upregulates a number of antioxidant proteins that can either repair or prevent oxidative damage [118], as well as guard against carcinogen-induced liver carcinogenesis in mice [119]. NRF2 is crucial for the innate immune system's ability to respond to infections by maintaining redox equilibrium and preventing abnormal inflammation signaling cascades [120]. By attaching to antioxidant response elements (AREs) located within the IL-6 promoter, NRF2 can also have a direct impact on the expression of pro-inflammatory cytokines like IL-6 [121].
In contrast, levels of the hepatotoxicity biomarker alanine aminotransferase (ALT) were 2-fold higher acutely in male adult mice exposed to ABP, DEN, or carbon tetrachloride (CCl4) than in female adult mice [122], while levels of the inflammatory biomarker interleukin-6 (IL-6) did not differ based on sex. While CCl4 produced a 40-fold ALT elevation but without sex differences, treatment of immature mice with either ABP or DEN using conventional tumour-inducing postnatal exposure protocols did not result in an increase in serum ALT or IL-6 levels in either males or females. There was no sex difference in the baseline expression of Ggt1 or Hmox1, but adult females expressed the NRF2-responsive gene Nqo1 at higher levels than adult males. Animals that were still developing sexually revealed no sex difference in the three genes' baseline expression. While CCl4 slightly increased the expression of Ggt1 in both males and females and Nqo1 only in females, postnatal DEN exposure slightly increased the expression of Ggt1 only in male mice and Nqo1 in both sexes. Together, these findings rule out the possibility that postnatal carcinogen exposure in mice results in acute hepatotoxic, inflammatory, or NRF2-activated gene responses that are responsible for the male predominance in liver tumour growth [122]. These results also imply that when extrapolating putative processes to liver carcinogenesis models that frequently employ postnatally exposed mice, acute toxicity studies conducted in adult mice should be read with caution. However, the various experimental setups used could be the cause of the disparate results found in the various studies.
AFB1 is a strong hepatotoxin and hepatocarcinogen for humans and most other mammalian species, although adult mice are remarkably resistant to it [123]. Aspergillus flavus, a mold that develops on groundnuts, grain, and maize that mice frequently consume, produces AFB1. Cytochrome P450 (CYP) transforms AFB1 in both humans and mice into a reactive AFB1- epoxide that can damage DNA by attaching to the N-7 atom of guanine [124]. Once produced, the glutathione S-transferase (GST) enzymes in the cytosol can catalyse the conjugation of the AFB1-epoxide with reduced glutathione to detoxify it. Water-soluble aflatoxin mercapturic acids (AFB1-NAC), which are eliminated in urine as glutathione conjugates of AFB1-epoxides [125]. Mice's inherent resistance to AFB1 may be due to CYP isoenzymes' poor capacity to produce reactive epoxides and/or GST isoenzymes' great capacity to produce glutathione conjugates.
The important function of GSTA3 in AFB1 resistance was confirmed by a study that produced glutathione S-transferase (GST) A3 knockout (KO) mice. GSTA3 KO mice are vulnerable to the acute cytotoxic and genotoxic effects of AFB1 [126]. The experiment revealed the effects of AFB1 treatment on histological alterations, tumour formation, biochemical changes, gender response, and the role of oxidative stress. The authors used a weekly AFB1 delivery procedure, and within 1-3 weeks they noticed substantial oval (liver stem) cell growth, which was followed by microvesicular lipidosis, megahepatocytes, nuclear inclusions, cholangiomas, and tiny nodules. All the male and female GSTA3 KO mice exhibited severely deformed livers with macro- and microscopic cysts, hepatic nodules, cholangiomas and cholangiocarcinomas, and OC proliferation. These livers were treated with 12 and 24 weekly AFB1 injections, followed by rest periods of 12 and 6 months, respectively. They proposed that long-term AFB1 therapy inhibited hepatocyte proliferation, which was then made up for by OC proliferation and finally resulted in the development of cholangiocarcinoma (CCA). Male KO mice displayed less severe acute liver damage, OC proliferation, and AFB1-DNA adducts than female KO mice at low doses of AFB1. In KO mice, there were no appreciable compensatory alterations in the levels of CYP1A2 and CYP3A11, epoxide hydrolase, GST subunits, or GST enzyme activity. Finally, a slight increase in F2 -isoprostane and isofuran in KO mice supported the putative in vivo activity of GSTA3 hydroperoxidase [126]. In contrast to the known higher incidence of liver cancer in males in humans, these findings show that initial vulnerability to AFB1 is greater in female mice, that oval cell response and GSTA3 peroxidase activity may affect susceptibility to cancer development.
Other information supports the notion that oxidative stress plays a part in the different onset of liver cancer in the two sexes. According to a study, age-related TBARS accumulation in the liver may be sex-related because it was more noticeable in old male mice compared to old female mice. Gonadotropic hormones, particularly oestrogens, may be the cause of these sex-related variations in the TBARS level [127]. The connection between oestrogens and liver oxidative damage has been shown by numerous in vitro investigations [128]. Since females at that age are in a reproductive-decline stage, hormonal changes alone cannot account for the fact that TBARS in 18-month-old females were higher than in males of the same age. The growth of tumours seen in aged male mice may be linked to gender-specific changes in TBARS. These findings are consistent with some published studies that link declining lipid peroxidation (LPO) levels to increasing tumour size [129].
Researchers looked studied the activities of total superoxide dismutase (tSOD), Gpx, and catalase (CAT). LPO, quantified in terms of TBARS, was determined by the authors to be a marker of liver oxidative damage. LPO increased with aging in both sexes. In both mouse sexes, tSOD appears to be a dormant antioxidative enzyme. The principal alterations in the liver's antioxidant capacity of aging mice were connected to sex-related increases in CAT and Gpx that were only seen in males. Surprisingly, hepatic tumours developed in more than 60% of 18-month-old men (but not girls), who first appeared at 10 months. The findings indicate that increased liver antioxidant capacity of CAT and Gpx in male mice may be an indication of oxidative stress; increase in CAT and Gpx activities in male mice are strongly correlated with incidence of hepatic tumours; and significantly increased SOD activity in tumour-bearing mice may have been caused by damage from accumulated hydrogen peroxide H2O2 [130].
The varied ways that oxidative stress behaves in the two sexes is also intriguing. An experiment revealed that during male senescence, CAT and Gpx significantly changed. In contrast to this, there was little to no change in CAT activity and no appreciable change in Gpx activity in female mice. In general, CAT and Gpx activity were 50% and 85% higher in males than in females. Tumour-bearing mice displayed elevated tSOD activity in contrast to the antioxidant enzyme status of tumour-free mice (inert tSOD activity). Antioxidant enzyme activities are typically thought to vary during or after tumour development [131]. Most past investigations have suggested that cancer has poor antioxidant enzyme activity [132]. However, most of them used cell lines, and in some of them, conclusions were reached based on blood sample activity measurements that did not accurately reflect the enzyme levels in the tumour or the affected organ. Manganese superoxide dismutase (MnSOD) expression has been shown to be high in many human cancers and in some tumors, the level of MnSOD is directly correlated with the tumour grade [133]. Additionally, Manna et al. demonstrated that MnSOD overexpression in tumors may give tumour cells a survival advantage [134]. Another author's theory that tumour cells produce a significant amount of H2O2 [135] and research showing that tSOD overexpression promotes H2O2 generation support this idea. To fulfil the demands of the increased LPO and H2O2 build-up brought on by the increased SOD activity, these facts may explain why males generally have higher CAT and Gpx activities [136].
Numerous studies have demonstrated that oxidative stress restricts the ability of cells to undergo mitosis, suggesting that oxidative stress may also condition a different proliferative capacity of cancerous cells [137]. Based on higher antioxidant enzyme levels and the oxidative stress situation prevalent in men, it is possible to infer that cell division favouring clonal growth can occur. Such a phenomenon might aid in the development of cancer. Similar findings have been published from Gonzales, where higher antioxidant levels have been linked to a faster rate of cell division [138]. Like the gender difference in the incidence of liver cancer in humans, postnatal exposure of mice to ABP causes a higher incidence of liver tumors in males than in females. ABP-DNA adducts that start tumour growth are produced because of first N-hydroxylation that is initially mediated by CYP1A2, according to a conventional theory of ABP carcinogenesis. CYP2E1 was found to be a key ABP N-hydroxylating enzyme in isozyme-selective inhibition tests employing liver microsomes from wild-type and genetically engineered mice. Oxidative stress was brought on by the N-hydroxylation of ABP by transiently expressed CYP2E1 in cultured mouse hepatoma cells. Male wild-type mice exposed postnatally to a tumour-causing dosage of ABP also experienced oxidative stress, but neither male Cyp2e1(/) mice nor female mice did. However, females showed a stronger NRF2-associated antioxidant response [139]. These results imply that CYP2E1 is a novel ABP-N-oxidizing enzyme and that sex differences in tumour incidence and cell proliferation may be related to sex differences in oxidative stress and antioxidant responses to ABP.
Finally, a particularly exciting area of research focuses on the relationships between gender differences, obesity, oxidative stress, and liver cancers. Recent population-based studies have repeatedly demonstrated that obese men are far more likely to acquire HCC. Men with a BMI of 35 kg/m2 showed a severe 4.52-fold increase in relative risk of mortality from liver cancer, although women only showed a small 1.68-fold increase, according to prospective research involving more than 900,000 persons [140]. The large gender-based variation in HCC incidence has been further validated by a cohort study of 5.24 million persons in the UK [141]. According to studies, BMI and HCC in males were correlated [141], and increased and disordered ROS production in extra adipose tissue during obesity may increase oxidative stress and the likelihood of developing HCC [142]. In contrast to subcutaneous fat accumulation, visceral fat deposition is substantially higher in males than in females [143]. In numerous datasets [144,145], men were found to have larger visceral fat and liver fat contents than women despite having similar total fat and BMI values. Liver cancer is facilitated by visceral fat, which actively secretes carcinogenic adipokines that cause persistent inflammation. High androgen receptor density may be the root cause of the differences between liver cancer and visceral fat accumulation [146]. As people get older, their visceral body fat increases while their subcutaneous body fat decreases, which is correlated with an increase in the incidence of HCC [147].
3.3. Colorectal cancer, oxidative stress and gender differences
With an incidence of over one million cases each year and more than 500,000 deaths, colorectal cancer (CRC) is the second most prevalent cause of cancer mortality among men and women globally. CRC has been rising in Asia to levels seen in North America and Europe [148]. CRC accounted for 10% of all new cancer cases and 9.4% of all cancer-related deaths in 2020, according to data on cancer from 185 countries. Drug resistance and adverse reactions continue to hinder the success of treatment, despite the fact that the overall survival rate of CRC patients has increased because to advancements in treatment methods like chemotherapy.
According to certain research, the disease affects people of various sexes at different rates, and this could be due to the oxidative stress. For instance, neutrophils and monocytes both contain the lysosomal enzyme myeloperoxidase (MPO) [149]. Hypochlorous acid, a potent oxidant produced by MPO for its microbicidal function, can target proteins, nucleic acids, and unsaturated lipids by simultaneously releasing ROS [150]. A -463 G>A transition, which is situated in the consensus binding location of the SP1 transcription factor, is a frequently occurring polymorphism in the MPO gene promoter region. In vitro, the MPO G wild-type allele confers approximately twenty-five times more transcriptional activation than the -463 A variant. According to reports, this polymorphism raises the likelihood of developing laryngeal, lung, breast, and stomach cancers [151,152,153,154,155]. According to a study, those with the genotype GA/AA were considerably less likely to get colorectal cancer than people with the GG genotype. The reduced risk was particularly significant among men according to the stratified analysis. For male individuals with the GA/AA genotype compared to GG genotype, the adjusted OR was 0.47. However, among women, the OR was not statistically significant. The possibility that oestrogen induced increased MPO -463 A promoter activity is the cause of the MPO -463 A variant's lack of protective effect in female patients is therefore plausible [156].
Oxidative stress and cancer have been linked in another research. Bilirubin is more than only the by-product of heme catabolism. It is now thought to be an essential blood component that forms endogenously and has anti-inflammatory and antioxidant activities [157]. Recent research has indicated that bilirubin, particularly unconjugated bilirubin (UCB), may provide protection against oxidative stress-related illnesses like CRC. In vitro research outcomes also demonstrated that UCB has anti-mutagenic qualities [163], which may be especially pertinent for gut health. Tetrapyrroles, a family of bile pigments that are abundant in the intestine, reduced the genotoxicity brought on by poly-/heterocyclic amines and triggered apoptosis in cancer cells [164,165,166]. Higher circulating UCB concentrations were positively linked with CRC risk in males and negatively associated with risk in women, according to a study that examined relationships between UCB and CRC risk in the European Prospective Investigation into Cancer and Nutrition (EPIC) study [167]. According to one study, every one standard deviation increase in log-UCB was associated with a lower risk of CRC in males and a higher risk in women (Pheterogeneity = 0.4 for differences between men and women) [168]. Finally, it has been demonstrated that UCB may easily cross cell membranes in vivo, infiltrate colon cancer cells to stop tumour cell growth [169], trigger death in cancer cells in vitro [170], and control gene transcription (via ERK, p53, and p27) [171]. Strogen, lower NADPH-oxidase activity, or other previously described mechanisms may make women less susceptible to oxidative stress [23].
3.4. Lung cancer, oxidative stress and gender differences
Nearly 25% of all cancer-related deaths are caused by lung cancer, which is the most common cancer in the world. The five-year survival rate is still dismal at 22% despite recent improvements in diagnosis and treatment [172,173]. There may be gender disparities in lung cancer incidence, according to epidemiologic data [174,175,176]. Agreeing to several studies, women may be more likely than men to acquire lung and colon cancer from smoking cigarettes [177,178].
The expression of genes relevant to cancer and the immune system is altered by genetic and epigenetic alterations, as well as by the abnormal expression of non-coding RNAs, which predisposes the lung epithelium to carcinogenesis. Smoking-related oxidative stress contributes to decreased genomic integrity, promoted epithelial-mesenchymal transition, and creation of a chronic inflammatory milieu. Although not all smokers develop lung cancer, this results in abnormal immune reactions that support the development of cancer. Females are more likely to accumulate oxidative stress damage due to gender differences in the metabolism of cigarette smoke, which increases their risk of developing lung cancer [179]. Additionally, ROS and RNS can activate signaling molecules like HIF1, which is a key regulator of angiogenesis and a driving force behind the development of tumors [180]. Furthermore, it has been demonstrated that the by-products of ROS and inflammation can inactivate PTEN, a tumour suppressor gene that is frequently altered in lung cancer, by creating an intramolecular disulfide bond [181,182].
Large epidemiological studies have demonstrated that for every pack-year of smoking, women are two to three times more likely to die from COPD than males [183] and are 50% more likely to develop COPD than men. One explanation is that because women's lungs are smaller than men's with comparable smoking histories, the harm from oxidative stress is more obvious in women [183]. Another is sex variations in the metabolism of tobacco: women have higher liver CYP1A1 and CYP1B1 activity levels, which activate specific tobacco smoke components to create ROS [184]. Strogen’s role in activating CYP enzyme-related pathways is a contributing factor in the enhanced CYP expression in females [185]. For instance, a study of smokers who developed lung cancer showed that females had higher levels of CYP1A1 expression and a commensurate rise in DNA adducts, even in lung tissue that was not cancerous [186]. Additionally, studies on animals showed that the injection of naphthalene—a substance found in tobacco smoke—caused more airway damage in female mice than in male mice. This was due to increased CYP enzyme expression and the production of metabolites, which led to a more severe inflammatory response in the airways and produced more ROS than in male mice [187]. Because women are more frequently exposed to biomass smoke, exposure to indoor and outdoor air pollution is also a significant risk factor for the development of COPD and lung cancer in non-smokers [188,189].
The varied ways that oxidative stress affects the incidence of pulmonary neoplasia in the two sexes could be explained by other processes. A class of pervasive environmental pollutants known as polycyclic aromatic hydrocarbons (PAHs) are mostly formed when carbonaceous materials are burned insufficiently during energy and industrial production operations. The oxidative damage to DNA brought on by the metabolic transformation of PAHs, which is also regarded to be crucial in the PAHs linked cancer initiation and can lead to a large exposure-response increase in lung cancer [190], can produce high levels of ROS. Another way that PAHs can contribute to the development of cancer is by forming bulky PAH-DNA adducts through covalent bonding with DNA [191]. Blood PAH-DNA adduct levels have been reported to be considerably higher in populations exposed to ambient air pollution [192]. It may be possible to discover gender-specific molecular modifications implicated in the lung carcinogenesis brought on by PAHs by conducting analysis on the biomarkers of PAH exposure and PAH generated early genotoxic effects, including oxidative stress and chromosome damage biomarkers.
A study identified sixteen environmental PAHs in workplaces and assessed the levels of 12 urinary PAH metabolites (OH-PAHs), plasma BPDE-Alb adducts, urinary 8-hydroxydeoxyguanosine (8-OHdG) and 8-iso-prostaglandinF2a (8-iso-PGF2a), as well as the frequency of lymphocyte micronuclei. It revealed that women who worked in the office, next to the coke oven, or on its bottom or side, respectively, had significantly higher urine 8-OHdG and 8-isoPGF2a levels and lymphocytic micronucleus frequencies than men who worked in those locations. These gender disparities persisted even after possible confounders, urinary ROH-PAHs, and plasma BPDE-Alb adducts were taken into account. Gender and BPDE-Alb adducts had a strong impact on rising micronucleus frequencies. The foregoing gender disparities were more pronounced in the median- and high-exposure groups, according to authors who further stratified all workers based on the tertiles of urinary ROH-PAHs or plasma BPDE-Alb adducts [193]. As a result, women were more vulnerable than males to the oxidative stress and chromosomal damage caused by PAHs, which could be additional evidence for gender differences in PAH exposure-related lung carcinogenesis.
3.5. Melanoma, oxidative stress and gender differences
Since the middle of the 1950s, malignant melanoma incidence has increased more quickly than that of any other cancer type in the majority of developed nations [194], and the prognosis for cases with advanced metastases is still dismal [195]. Gender has been shown to be an independent prognostic factor of melanoma survival in numerous studies, as it remains significant after adjusting for nearly all known prognostic indicators, including age, Breslow thickness, Clark level of invasion, body site, histological subtype, and even recently emerged prognostic indicators, like ulceration, sentinel node status, and mitotic rate [196,197]. Both the incidence and survival of malignant melanoma differ significantly across gender. Male patients advance more quickly to stage III [198] and maybe even stage IV melanoma [199,200]; male original melanomas appear to grow more quickly than those in females; men present with nodal and visceral metastases more frequently than women [196]. Instead, women are more likely to present with tumors that are in an earlier stage, have longer survival times, and experience better outcomes [201,202,203,204,205].
More and more evidence point to the involvement of oxidative stress, which is brought on by high amounts of ROS like superoxide anion and hydrogen peroxide, in the development of melanoma [206,207]. When compared to nearby tissues or melanocytes, melanoma cells produce a lot of ROS, which they then excrete into the extracellular space [208]. Additionally, melanoma cells have elevated intracellular ROS levels [209].
High amounts of oxidative stress are known to exist in the initial melanoma tumour environment [210,211,212], tumour related immune cells release ROS [213], and ultraviolet (UV) radiation further intensifies oxidative stress in the skin and melanocytes [214]. In contrast to surrounding non-tumour tissue, benign melanocytic nevi, and control subject skin, Sander et al. discovered a considerable upregulation of antioxidant enzymes in human melanoma biopsies, indicating that the melanoma cells were responding to increasing oxidative stress [211].
According to a different study, the advantage that females have in terms of melanoma survival is likely due to sex differences in the capacity to counteract the oxidative stress brought on by ROS [210]. In fact, it appears that the oxidative environment in the skin of male and female mice has different baseline characteristics; UV-induced oxidative stress amplifies these differences. In comparison to female hairless mice, the skin of males had a lower baseline level of antioxidant enzyme and a roughly 10-fold lower antioxidant functional capacity. In comparison to levels found in the skin of male mice exposed to UVB radiation, the skin of female mice showed a significantly higher induction of antioxidant level, greater antioxidant functional capacity, and lower levels of 8-oxo-deoxyguanosine, the most common type of DNA damage caused by ROS [215]. These findings were supported by an experiment that looked at gender differences in the development of cancer linked to UV-induced chronic inflammation [216]. According to the finding’s, photoaging damage was present in both male and female mice at the ninth week. However, only male mice in the third week developed skin tumours. Additionally, UV increased the expression of the p65, p-p65, IL-6, and TNF- proteins in skin, and these factors were more elevated in the male mouse model. The parameters of blood systemic inflammation were altered to variable degrees in the model groups, according to haematology data, whereas the internal organs of both model groups revealed varying degrees of inflammatory cell infiltration, according to pathology results. These findings suggest that UV-induced skin inflammation, carcinogenesis, and systemic damage differ between the sexes.
Additionally, it is possible that men's higher ROS levels encourage the selection of ROS-resistant melanoma cells. Consequently, ROS can promote melanoma cells' capacity for metastatic spread. Additionally, because men have weaker antioxidant defences, the ROS that melanoma cells produce damage surrounding healthy tissues more severely, which promotes metastasis. As a result, ROS could account for the reported disparities in melanoma survival between males and females [210].
After menopause, according to some researchers, the female advantage vanishes [197]. Others, however, discovered that females continue to live longer even after menopause [217]. In female rats, ovariectomies boosted peroxide generation in liver cells to levels seen in male cells, decreased antioxidant enzyme levels to those found in male cells, and restored both peroxide and antioxidant enzyme levels in female cells to the control female levels [23]. This team discovered that 17-b-estradiol decreased hydrogen peroxide production when isolated mitochondrion was incubated with it [218].
The effect of antioxidant supplementation on the incidence of melanoma has also been studied, however due to the small numbers of events in the trials, no significant effect [219] or even a negative effect [220] was discovered. More importantly, the effect of antioxidants varied by gender in each of these studies, affecting both the incidence of melanoma [220] and all cancers [221]. This strongly implies that gender has a role in the relationship between melanoma and ROS.
3.6. Non-Hodgkin Lymphoma, oxidative stress and gender differences
The ninth most common cancer killer of both sexes is Non-Hodgkin lymphoma, a lymphoid malignancy that develops in white blood cells called lymphocytes. The condition is the most prevalent haematological malignancy in the world, accounting for about 90% of all lymphomas. The Swedish Lymphoma Register was used in a population-based cohort study that looked at gender differences in the incidence of lymphoma subtypes and excess mortality among people diagnosed between 2000 and 2019 [222]. Poisson regression was used to predict the male-to-female incidence rate ratios (IRRs) and excess mortality ratios (EMRs) after adjusting for age. They discovered 36 795 instances of lymphoma, 20 738 (56.4%) of which were in men and 16 057 (43.6%), in women. Incidence rate ratios (IRRs) ranged from 1.15 in follicular lymphoma to 5.95 in hairy cell leukemia, with men being considerably more at risk for 14 of the 16 subtypes of lymphoma. Although only statistically significant for classical Hodgkin lymphoma 1.26, aggressive lymphoma not otherwise specified 1.29, and small lymphocytic lymphoma 1.52, EMRs >1 was seen in 13 out of 16 lymphoma subtypes, indicating higher mortality in men. Similar findings were obtained from a related analysis utilizing information from the Danish Lymphoma Register [222]. In conclusion, researchers found that for the majority of lymphoma subtypes, men had a significantly greater incidence and a tendency toward higher death.
The differing levels of oxidative stress experienced by the two sexes may contribute to the development and spread of lymphomas. For instance, a study [223] examined the idea that lymphomagenesis following low-dose radiation is aided by mitochondrial malfunction and elevated superoxide levels in thymocytes over expressing Bax (Lck-Bax1 and Lck-Bax38&1). Single whole-body doses of 10 or 100 cGy of 137Cs, or iron ions, or silicon ions, were administered to Lck-Bax1 single-transgenic and Lck-Bax38&1 double-transgenic mice. In female Lck-Bax1 mice, a 10 cGy dosage of 137Cs markedly increased the incidence and development of thymic lymphomas. In contrast to silicon ions, a 100 cGy dosage of high-LET iron ions significantly and dose-dependently accelerated lymphomagenesis in both male and female Lck-Bax38&1 mouse. Lck-Bax38&1 over expressing animals were bred with Sirtuin 3 knockouts, a mitochondrial protein deacetylase that controls superoxide metabolism, to ascertain the contribution of mitochondrial oxidative metabolism. Significant increases in thymocyte superoxide levels and accelerated lymphomagenesis were seen in Sirt3//Lck-Bax38&1 animals [223] (Table 1). These findings demonstrate that radiation exposure increases lymphomagenesis in Bax over-expressing animals in a manner that depends on both LET and gender. These results are consistent with the hypothesis that in Lck-Bax transgenic mice, mitochondrial dysfunction increases superoxide levels and speeds up lymphomagenesis.
4. Future perspectives
The potential that the various profiles of oxidative stress connected to sex may affect the effectiveness and toxicity of chemotherapy and radiotherapy in cancer subjects is unquestionably an exciting area for future research.
By interfering with mitochondrial function, bioenergetics, signaling pathways, and redox balance, anthracyclines can cause cell malfunction and death [224]. The majority of these targets have been shown to exhibit sexual dimorphism, including "redox features" of cells, such as altered redox-associated molecules and enzymes in relation to gender differences in terms of intracellular production and biochemical activity of reactive species, as well as expression of genes related to mitochondria [225,226]. Along with pharmacodynamics, sex-related variations in pharmacokinetics may have significant clinical ramifications, affecting drug side effects. Men have a considerably higher doxorubicin clearance than women, according to several authors [227]. This was confirmed by the observation that doxorubicinol levels were higher in men, which may be connected to higher aldoketoreductase activity [228]. A lower expression of p-glycoprotein in females may also cause doxorubicin and doxorubicinol to accumulate, increasing the risk of cardiotoxicity. The pharmacokinetics of epirubicin have been shown to differ similarly by sex [228].
Additionally, anthracyclines' overproduction of ROS and RNS causes redox stress, which causes cardiac injury [229], DNA damage, lipid peroxidation, membrane injury, and/or apoptosis, as well as changes in the enzymatic activity of the mitochondrial redox system. The respiratory complexes, Krebs cycle enzymes, oxidative phosphorylation, -oxidation, and nitric oxide synthases (NOSs) are among the changed enzymes [230,231,232]. It is possible that various oxidative stresses that influence cardiotoxicity in a gender-specific way are sustained by a complicated inter-relationship between oestrogen receptors and enzyme activity engaged in redox processes. Gonzalez and colleagues' research shows that adult male SH rats with tumours are more cardiosensitive to doxorubicin treatment than female or hormone-deficient animals, which lends credence to this theory [233]. According to these findings, doxorubicin-induced cardiotoxicity is regulated by reproductive hormones, and the selective cytotoxic mechanism probably works by increasing oxidative stress and apoptosis in male SH rats [233].
The anthracycline drug doxorubicin (Dox) is highly effective against a number of neoplastic illnesses but also causes dose-limiting cardiotoxicity [234,235]. Congestive heart failure, for instance, occurs following Dox treatment at a rate of about 4% at doses of 500–550 mg/m2, but this rate rises to 18% at doses of 55–600 mg/m2, and it reaches 36% in patients receiving >601 mg/m2 of Dox [236]. A continued follow-up of the cardiac condition of patients who received anthracyclines is recommended since cumulative and late-onset progressive cardiotoxicity might be seen years or decades after treatment [237]. In the mitochondria, doxorubicin builds up and causes an excessive amount of ROS production. Male adults (15-55 years old) are more likely than females to have cardiovascular disease overall [238,239,240,241,242]. Doxorubicin treatment for women causes cardiac failure in 6-20% of adults and 40% of pediatric patients [13]. Additionally, postmenopausal women are more susceptible to cardiac stress than men their own age following Dox treatment [243]. Finally, Dox-induced cardiotoxicity in prepubescent girls was found to be more severe than in boys of the same age [243]. Additionally, according to recent clinical reports, male adults and young girls are more cardiosensitive to Dox. Adult male SHRs with tumours are more cardiosensitive to Dox than female or hormone-deficient animals, according to a study. The selective cytotoxic mechanism is thought to work because oxidative stress and apoptosis are more strongly activated in male SHRs, and this suggests that Dox-induced cardiotoxicity inhibits or negatively regulates reproductive hormones [244].
According to some theories, oestrogen acts as a cardioprotectant by reducing left ventricular hypertrophy, preventing cardiomyocyte death, and protecting against the onset of cardiac fibrosis in females [245]. The hormones oestrogen (and possibly testosterone) may protect the heart against excessive drug-induced oxidative damage.
Radiotherapy is a topic that warrants similar thought. The idea that sex affects how the body reacts to radiation is widely established, but our understanding of the underlying molecular mechanisms is still lacking. Due to this ignorance, current risk estimates are uncertain and have low-resolution dose limits [246]. Most authorities have not yet defined dose limits with sex-specific regimens, while it is advised to use male and female reference phantoms for study [247].
By predicting cellular outcomes from proteins related with DNA damage and repair, it was possible to further highlight the qualitative variations in ionic-radiation (IR)-induced reactions between age and sex. Juvenile girls and males appeared to start separate signaling cascades as opposed to merely altering the response intensity of the same mechanism. Although both shared a suppression of cell cycle progression, males shifted towards pro-apoptosis with mitochondrial stress and reduced DNA repair while females displayed activated DNA repair and pro-survival mechanisms. Inflammatory regulators also seemed to compete for control over the activation and inhibition of immunological responses, however they were not found in females [248].
To comprehend and have a thorough understanding of the effects of ionizing radiation, radiobiology knowledge is of utmost relevance. The interaction of this physical stressor with targeted and non-targeted cells, tissues, and organs has profound implications on both the early and late development of primary and secondary malignancies. As a result, a thorough knowledge of the mechanisms underlying radiation carcinogenesis is still lacking. Some studies, however, have suggested or shown that non-targeted effects may contribute to an elevated risk of developing cancer [249].
Evidence has demonstrated that males and females experience distinct rates of radiation-induced mutation patterns, subsequent changes in gene expression and epigenetic status, and eventual malignancies [250,251,252,253]. According to research by Korturbash et al., local cerebral irradiation of mice causes DNA damage and changes in global DNA methylation that are tissue- and sex-dependent. They demonstrated that whereas nontargeted effects can result in skin hypomethylation, they have not been studied for spleen hypomethylation. Additionally, their findings suggested that males are more obviously hypomethylated than females [254]. In non-targeted tissues, similar outcomes have been seen for the control of the microRNAome and inflammatory responses [255,256].
Following exposure to X or gamma rays, oxidative DNA damage and cell death are brought on by free radical production brought on by the interaction of ionizing radiation with water molecules and redox-mediated biological pathways. In both the nucleus and the mitochondria, the interaction of free radicals with DNA results in various forms of DNA oxidation. Further DNA damage can result from oxidative stress, inflammatory reactions, and cell death due to necrosis or apoptosis. Patients with cancer and those receiving radiation for their malignancies both have higher levels of oxidized cell free DNA [257,258,259].
So-called precision radiation oncology in cancer treatment is still a long way from the bedside2. Similar to this, individualized risk evaluation for ionizing radiation (IR) exposure is required but is still in its infancy [260]. Improved medicines with lesser toxicities might be developed with a better understanding of the sex-related mechanisms of protection or harm [261].
A further field of study could be the use of natural substances for the modification of oxidative stress [262,263]. As mentioned above, several natural products have an enhanced anticancer effect via a restoration of mitochondrial function [69,70]. The use of these substances could be useful in reducing the negative effects of sex on the progression of neoplastic diseases perhaps allowing a reduction in the dosage of traditional anticancer drugs.
5. Conclusions
A vast range of systems involved in the redox characteristics of cells are affected by oxidative damage [225,264]. Because of their genetic advantages resulting from their X chromosomes, oestrogen’s antioxidant protective properties, and their lower exposure to extrinsic risk factors like drinking and smoking, women often live longer than men. Sex hormones alter the expression of several crucial transcription factors that control ROS-induced stress and in vivo responses. Compared to men, women have lower levels of ROS generation and mitochondrial damage because to oestrogen, which is also linked to increased mitochondrial function and disease resistance. Furthermore, oestrogen benefits females by influencing NRF 2 activation and the regulation of other antioxidant-related transcription factors through NRF2. Effective cancer treatment necessitates an awareness of the potential of ROS and a focus on the traits of the study target, such as the patient's gender [265]. ROS have a variety of biochemical targets in cells.
Males exhibit higher rates of oxidative damage than females [266,267,268], which is supported by a 29% higher level of urinary oxidative damage waste products in healthy males after adjusting for smoking and body mass index [269]. Males also express lower levels of antioxidants than females, such as GSH, catalase, and SOD. Given that oxidative damage can result in cancer and cardiovascular disease, this is hypothesized to explain why women generally live longer lives than men [270].
Other mechanisms, such as the genetic overexpression of antioxidant enzymes in females, which may be brought on by oestrogen receptor activation, may also be important. In fact, ovariectomy-induced menopause in mice increased oxidative damage susceptibility. Although androgens like testosterone appear to weaken these same defence mechanisms, oestrogen levels do not entirely account for these variations in antioxidant defence [271].
Regarding mitochondrial function, the traditional view of mitochondrial inheritance holds that mtDNA is only passed down through the female line, while some reports have suggested that it may also be inherited from the father [272,273,274]. The different male/female sex hormones that control mitochondrial energy, OXPHOS, and Ca2+ homeostasis may be one of the causes of the well-documented mitochondrial sexual dimorphisms [275]. Although this varies depending on the tissue in question and the age/hormonal status of the tested subject, female mitochondria are typically reported to have higher functional capacity than male mitochondria [276].
A greater knowledge of the molecular mechanisms underlying the gender-related disparities in cancer that have been discovered may be able to inform more appropriate precision medicine and important information on treatment options for both males and females with neoplastic disorders.
Author Contributions
Conceptualization, A.A. and S.G.; methodology, S.C., Sa Ge (Sara Genovese) and G.P.; software, S.C; formal analysis, S.G., Sa Ge and G.P.; data curation, S.C. Sa. Ge, and G.P.; writing—original draft preparation, S.C.; writing—review and editing, S.C. and A.A.; supervision, S.G. and A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dorak, M.T.; Karpuzoglu, E. Gender differences in cancer susceptibility: an inadequately addressed issue. Front Genet. 2012, 3, 268. [Google Scholar] [CrossRef] [PubMed]
- Ashley, D.J. A male-female differential in tumour incidence. Br. J. Cancer 1969, 23, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Edgren, G.; Liang, L.; Adami, H.O.; Chang, E.T. Enigmatic sex disparities in cancer incidence. Eur J Epidemiol. 2012, 27, 187–196. [Google Scholar] [CrossRef]
- Cook, M.B.; McGlynn, K.A.; Devesa, S.S.; Freedman, N.D.; Anderson, W.F. Sex disparities in cancer mortality and survival. Cancer Epidemiol Biomarkers Prev. 2011, 20, 1629–1637. [Google Scholar] [CrossRef] [PubMed]
- Carlus, M.; Elies, L.; Fouque, M.C.; Maliver, P.; Schorsch, F. Historical control data of neoplastic lesions in the Wistar Hannover Rat among eight 2-year carcinogenicity studies. Exp Toxicol Pathol. 2013, 65, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Kadekar, S.; Peddada, S.; Silins, I.; French, J.E.; Högberg, J.; Stenius, U. Gender differences in chemical carcinogenesis in National Toxicology Program 2-year bioassays. Toxicol Pathol. 2012, 40, 1160–1168. [Google Scholar] [CrossRef]
- Seifarth, J.E.; McGowan, C.L.; Milne, K.J. Sex and life expectancy. Gend Med. 2012, 9, 390–401. [Google Scholar] [CrossRef]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am J Physiol Lung Cell Mol Physiol 2000, 279, L1005e28. [Google Scholar] [CrossRef]
- Olinski, R.; Zastawny, T.; Budzbon, J.; Skokowski, J.; Zegarski, W. , Dizaroglu, M. DNA base modifications in chromatin of human cancerous tissues. FEBS Lett 1992, 309, 193e8. [Google Scholar] [CrossRef]
- Conner, E.M.; Grisham, M.B. Inflammation, free radicals, and antioxidants. Nutrition 1996, 12, 274e7. [Google Scholar] [CrossRef]
- Hlavata, L.; Aguilaniu, H.; Pichova, A.; Nystrom, T. The oncogenic RAS2(val19) mutation locks respiration, independently of PKA, in a mode prone to generate ROS. EMBO J 2003, 22, 3337e45. [Google Scholar] [CrossRef] [PubMed]
- Vafa, O.; Wade, M.; Kern, S.; Beeche, M.; Pandita, T.K.; Hampton, G.M.; Wahl, G.M. c-Myc can induce DNA damage, increase reactive oxygen species, and mitigate p53 function: a mechanism for oncogene-induced genetic instability. Mol Cell 2002, 9, 1031e44. [Google Scholar] [CrossRef] [PubMed]
- Ozben, T. Oxidative stress and apoptosis: impact on cancer therapy. J Pharm Sci 2007, 96, 2181e96. [Google Scholar] [CrossRef] [PubMed]
- Imbesi, S.; Musolino, C.; Allegra, A.; Saija, A.; Morabito, F.; Calapai, G.; Gangemi, S. Oxidative stress in oncohematologic diseases: an update. Expert Rev Hematol. 2013, 6, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Musolino, C.; Allegra, A.; Saija, A.; Alonci, A. , Russo, S.; Spatari, G.; Penna, G., Gerace, D.; Cristani, M.; David, A.; Saitta, S., Gangemi, S. Changes in advanced oxidation protein products, advanced glycation end products, and s-nitrosylated proteins, in patients affected by polycythemia vera and essential thrombocythemia. Clin Biochem. 1439. [Google Scholar] [CrossRef]
- Gangemi, S.; Allegra, A.; Alonci, A.; Cristani, M.; Russo, S.; Speciale, A.; Penna, G.; Spatari, G.; Cannavò, A.; Bellomo, G.; Musolino, C. Increase of novel biomarkers for oxidative stress in patients with plasma cell disorders and in multiple myeloma patients with bone lesions. Inflamm Res. 2012, 61, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- Allegra, A.; Tonacci, A.; Giordano, L.; Musolino, C.; Gangemi, S. Targeting Redox Regulation as a Therapeutic Opportunity against Acute Leukemia: Pro-Oxidant Strategy or Antioxidant Approach? Antioxidants (Basel). 2022, 11, 1696. [Google Scholar] [CrossRef] [PubMed]
- Allegra,A. ; Petrarca, C.; Di Gioacchino, M.; Casciaro, M.; Musolino, C.; Gangemi, S. Modulation of Cellular Redox Parameters for Improving Therapeutic Responses in Multiple Myeloma. Antioxidants (Basel). 2022, 11, 455. [Google Scholar] [CrossRef]
- Ali, I.; Högberg, J.; Hsieh, J.H.; Auerbach, S.; Korhonen, A.; Stenius, U.; Silins, I. Gender differences in cancer susceptibility: role of oxidative stress. Carcinogenesis. 2016, 37, 985–992. [Google Scholar] [CrossRef]
- Ide, T.; Tsutsui, H.; Ohashi, N.; Hayashidani, S.; Suematsu, N.; Tsuchihashi, M.; Tamai, H.; Takeshita, A. Greater oxidative stress in healthy young men compared with premenopausal women. ATVB 2002, 22, 438–442. [Google Scholar] [CrossRef]
- Niveditha, S.; Deepashree, S.; Ramesh, S.R.; Shivanandappa, T. Sex differences in oxidative stress resistance in relation to longevity in Drosophila melanogaster. J. Comp. Physiol. B 2017, 187, 899–909. [Google Scholar] [CrossRef]
- Barp, J.; Araújo, A.S.; Fernandes, T.R.; Rigatto, K.V.; Llesuy, S.; Bello-Klein,´A. ; Singal, P. Myocardial antioxidant and oxidative stress changes due to sex hormones. Braz. J. Med Biol. Res 2002, 35, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Kander, M.C.; Cui, Y.; Liu, Z. Gender difference in oxidative stress: a new look at the mechanisms for cardiovascular diseases. J. Cell Mol. Med. 2017, 21, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Borras, C.; Sastre, J.; García-Sala, D.; Lloret, A.; Pallardo, F.V.; Vina, J. 2003. Mitochondria from females exhibit higher antioxidant gene expression and lower oxidative damage than males. Free Radic. Biol. Med, /: 546–552. https, 1016. [Google Scholar]
- Miquel, J.; Economos, A.C.; Fleming, J.; Johnson Jr., J.E. Mitochondrial role in cell aging. Exp. Gerontol. 1980, 15, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Sohal, R.S.; Sohal, B.H.; Brunk, U.T. Relationship between antioxidant defenses and longevity in different mammalian species. Mech. Ageing Dev. 1990, 53, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Barja, G.; Cadenas, S.; Rojas, C.; Perez-Campo, R.; Lopez-Torres, M. Low mitochondrial free radical production per unit O2 consumption can explain the simultaneous presence of high longevity and high aerobic metabolic rate in birds (Oct). Free Radic. Res 1994, 21, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Barja, G. Updating the mitochondrial free radical theory of aging: an integrated view, key aspects, and confounding concepts. Antioxid. Redox Signal 2013, 19, 1420–1445. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.A.; Drummond, G.R.; Mast, A.E.; Schmidt, H.H.; Sobey, C.G. Effect of gender on NADPH-oxidase activity, expression, and function in the cerebral circulation: role of estrogen. Stroke 2007, 38, 2142–2149. [Google Scholar] [CrossRef]
- Mendoza-Núnez, V.M.; Beristain-Perez, A.; Perez-Vera, S.P.; Altamirano-Lozano, M.A. Age-related sex differences in glutathione peroxidase and oxidative DNA damage in a healthy Mexican population. J. Women’s. Health (Larchmt.). [CrossRef]
- Kendall, B.; Eston, R. Exercise-induced muscle damage and the potential protective role of estrogen. Sports Medicine 2002, 32, 103–123. [Google Scholar] [CrossRef]
- Karolkiewicz, J.; Michalak, E.; Pospieszna, B.; Deskur-Smielecka, E.; Nowak, A.; Pilaczynska-Szczesniak, L. Response of oxidative stress markers and antioxidant parameters to an 8- week aerobic physical activity program in healthy, postmenopausal women. Archives of Gerontology and Geriatrics 2009, 49, e67–e71. [Google Scholar] [CrossRef]
- Semenova, N.V.; Rychkova, L.V.; Darenskaya, M.A.; Kolesnikov, S.I.; Nikitina, O.A.; Petrova, A.G.; Vyrupaeva, E.V.; Kolesnikova, L.I. Superoxide Dismutase Activity in Male and Female Patients of Different Age with Moderate COVID-19. Bull. Exp. Biol. Med 2022, 173, 51–53. [Google Scholar] [CrossRef]
- Chen, Y.; Ji, L.L.; Liu, T.Y.; Wang, Z.T. Evaluation of gender-related differences in various oxidative stress enzymes in mice. Chin. J. Physiol. 2011, 54, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Sobocanec, S.; Balog, T.; Sverko, V.; Marotti, T. Sex-dependent antioxidant enzyme activities and lipid peroxidation in ageing mouse brain. Free Radic. Res 2003, 37, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Sheng, Y.; Jiang, P.; Ji, L.; Xia, Y.; Min, Y.; Wang, Z. , 2011. The gender- dependent difference of liver GSH antioxidant system in mice and its influence on isoline-induced liver injury. Toxicology. [CrossRef]
- Vina, J.; Borras, C.; Gambini, J.; Sastre, J.; Pallardo, F.V. Why females live longer than males: control of longevity by sex hormones. Sci. Aging Knowl. Environ. 2005, (23), 17. [Google Scholar] [CrossRef]
- Erden Inal, M.; Akgün, A.; Kahraman, A. The effects of exogenous glutathione on reduced glutathione level, glutathione peroxidase and glutathione reductase activities of rats with different ages and gender after whole-body Γ-irradiation. AGE 2003, 26, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Alkazemi, D.; Rahman, A.; Habra, B. Alterations in glutathione redox homeostasis among adolescents with obesity and anemia. Sci. Rep. /: 3034. https, 3034. [Google Scholar]
- Bellanti, F.; Matteo, M.; Rollo, T.; De Rosario, F.; Greco, P.; Vendemiale, G.; Serviddio, G. Sex hormones modulate circulating antioxidant enzymes: impact of estrogen therapy. Redox Biol. 2013, 1, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Ferri, J.; Navarro, I.; Alabadí, B.; Bosch-Sierra, N.; Benito, E.; Civera, M. : Ascaso, J.F.; Martinez-Hervas, S.; Real, J.T. Gender differences on oxidative stress markers and complement component C3 plasma values after an oral unsaturated fat load test. Clin. Invest. Arterioscler. [CrossRef]
- Arnold, M.; Pandeya, N.; Byrnes, G.; Renehan, P.A.G.; Stevens, G.A.; Ezzati, P.M.; Ferlay, J.; Miranda, J.J.; Romieu, I.; Dikshit, R.; et al. Global burden of cancer attributable to high body-mass index in 2012: a population-based study. Lancet Oncol 2015, 16, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Kyrgiou, M.; Kalliala, I.; Markozannes, G.; Gunter, M.J.; Paraskevaidis, E.; Gabra, H. , Martin-Hirsch, P.; Tsilidis, K.K. Adiposity and cancer at major anatomical sites: umbrella review of the literature. BMJ.
- Renehan, A.G.; Zwahlen, M.; Egger, M. Adiposity and cancer risk: new mechanistic insights from epidemiology. Nat Rev Cancer 2015, 15, 484–498. [Google Scholar] [CrossRef]
- Murphy, N.; Strickler, H.D.; Stanczyk, F.Z.; Xue, X.; Wassertheil-Smoller, S.; Rohan, T.E.; Ho, G.Y.; Anderson, G.L.; Potter, J.D.; et al. A prospective evaluation of endogenous sex hormone levels and colorectal cancer risk in postmenopausal women. J Natl Cancer Inst 2015, 107. [Google Scholar] [CrossRef]
- Chlebowski, R.T.; Wactawski-Wende, J.; Ritenbaugh, C.; Hubbell, F.A.; Ascensao, J.; Rodabough, R.J.; Rosenberg, C.A.; Taylor, V.M.; Harris, R.; Chen, C.; et al. Estrogen plus progestin and colorectal cancer in postmenopausal women. N Engl J Med 2004, 350, 991–1004. [Google Scholar] [CrossRef]
- Avgerinos, K.I.; Spyrou, N.; Mantzoros, C.S.; Dalamaga, M. Obesity and cancer risk: Emerging biological mechanisms and perspectives. Metabolism Clinical and Experimental 2019, 92, 121–135. [Google Scholar] [CrossRef]
- Boutari, C.; Mantzoros, C.S. Inflammation: a key player linking obesity with malignancies. Metabolism 2018, 81, A3–6. [Google Scholar] [CrossRef] [PubMed]
- 48 Deng, T.; Lyon, C.J.; Bergin, S.; Caligiuri, M.A.; Hsueh, W.A. Obesity, inflammation, and cancer. Annu Rev Pathol 2016, 11, 421–449. [Google Scholar] [CrossRef]
- Moore, L.L.; Chadid, S.; Singer, M.R.; Kreger, B.E.; Denis, G.V. Metabolic health reduces risk of obesity-related cancer in framingham study adults. Cancer Epidemiol Biomark Prev 2014, 23, 2057–2065. [Google Scholar] [CrossRef] [PubMed]
- Sabharwal, S.S.; Schumacker, P.T. Mitochondrial ROS in cancer: initiators, amplifiers or an Achilles' heel? Nat Rev Cancer 2014, 14, 709–721. [Google Scholar] [CrossRef]
- Dikalov, S.I.; Nazarewicz, R.R. Angiotensin II-induced production of mitochondrial reactive oxygen species: potential mechanisms and relevance for cardiovascular disease. Antioxid Redox Signal 2013, 19, 1085–94. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F.; Charles, K.A.; Mantovani, A. Smoldering and polarized inflammation in the initiation and promotion of malignant disease. Cancer Cell 2005, 7, 211–217. [Google Scholar] [CrossRef]
- Iyengar, N.M.; Gucalp, A. , Dannenberg, A.J.; Hudis, C.A. Obesity and cancer mechanisms: tumor microenvironment and inflammation. J Clin Oncol, 4270. [Google Scholar]
- Yehuda-Shnaidman, E.; Schwartz, B. Mechanisms linking obesity, inflammation and altered metabolism to colon carcinogenesis. Obes Rev 2012, 13, 1083–1095. [Google Scholar] [CrossRef]
- Fischer, M. Census and evaluation of p53 target genes. Oncogene 2017, 36, 3943–3956. [Google Scholar] [CrossRef]
- Engeland, K. Cell cycle arrest through indirect transcriptional repression by p53: i have a DREAM. Cell Death Differ. 2018, 25, 114–132. [Google Scholar] [CrossRef]
- Freudenstein, D.; Litchfield, C.; Caramia, F.; Wright, G.; Solomon, B.J.; Ball, D.; Keam, S.P.; Neeson, P.; Haupt, Y.; Haupt, S. TP53 Status, patient sex, and the immune response as determinants of lung cancer patient survival. Cancers (Basel) 2020, 12, 1535. [Google Scholar] [CrossRef]
- Beyfuss, K.; Hood, D. A. A systematic review of p53 regulation of oxidative stress in skeletal muscle. Redox Rep. 2018, 23, 100–117. [Google Scholar] [CrossRef] [PubMed]
- Lebedeva, M.A.; Eaton, J.S. , Shadel, G.S. Loss of p53 causes mitochondrial DNA depletion and altered mitochondrial reactive oxygen species homeostasis. Biochim. Biophys. Acta 2009, 1787, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, C.; Hu, W.; Feng, Z. Tumor suppressor p53 and metabolism. J. Mol. Cell Biol. 2019, 11, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Beekman, M.; Dowling, D.K.; Aanen, D.K. The costs of being male: are there sex-specific effects of uniparental mitochondrial inheritance? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130440. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.E.; Ceder, S.; Bykov, V.J.N. , Wiman, K.G. p53 as a hub in cellular redox regulation and therapeutic target in cancer. J. Mol. Cell Biol. 2019, 11, 330–341. [Google Scholar] [CrossRef]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G. , Castoria, G.; Migliaccio, A. ROS in cancer therapy: the bright side of the moon. Exp. Mol. Med. [CrossRef]
- Haupt, S.; Haupt, Y. Cancer and Tumour Suppressor p53 Encounters at the Juncture of Sex Disparity. Front Genet. 2021, 12, 632719. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Siegel, R.L.; Ward, E.M.; Jemal, A. Global cancer incidence and mortality rates and trends–an update. Cancer Epidemiol. Biomark. Prev. 2016, 25, 16–27. [Google Scholar] [CrossRef]
- Sun, T.; Plutynski, A.; Ward, S.; Rubin, J.B. An integrative view on sex differences in brain tumors, Cell. Mol. Life Sci. 2015, 72, 3323–3342. [Google Scholar] [CrossRef]
- Yang, W.; Warrington, N.M.; Taylor, S.J.; Whitmire, P.; Carrasco, E.; Singleton, K.W.; Wu, N. , Lathia, J.D.; Berens, M.E.; Kim, A.H.; Barnholtz-Sloan, J.S.; Swanson, K.R.; Luo, J., Rubin, J.B. Sex differences in GBM revealed by analysis of patient imaging, transcriptome, and survival data. Sci Transl Med. 2019, 11, eaao5253. [Google Scholar] [CrossRef]
- Tedeschi-Blok, N.; Lee, M.; Sison, J.D.; Miike, R.; Wrensch, M. Inverse association of antioxidant and phytoestrogen nutrient intake with adult glioma in the San Francisco Bay Area: a case-control study. BMC Cancer. 2006, 6, 148. [Google Scholar] [CrossRef]
- Desai, V.; Jain, A.; Shaghaghi, H.; Summer, R.; Lai, J.C.K.; Bhushan, A. Combination of Biochanin A and Temozolomide Impairs Tumor Growth by Modulating Cell Metabolism in Glioblastoma Multiforme. Anticancer Res. 2019, 39, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ni, Q.; Wang, Y.; Fan, H.; Li, Y. Synergistic anticancer effects of formononetin and temozolomide on glioma C6 cells. Biol. Pharm. Bull. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ruszkiewicz, J.A.; Miranda-Vizuete, A.; Tinkov, A.A.; Skalnaya, M.G.; Skalny, A.V.; Tsatsakis, A. , Aschner, M. Sex-Specific Differences in Redox Homeostasis in Brain Norm and Disease. J Mol Neurosci. 2019, 67, 312–342. [Google Scholar] [CrossRef] [PubMed]
- Candeias, E.; Duarte, A.I.; Sebastião, I.; Fernandes, M.A.; Plácido, A.I.; Carvalho, C.; Correia, S. , Santos, R.X.; Seiça, R.; Santos, M.S.; Oliveira, C.R.; Moreira, P.I. Middle-Aged Diabetic Females and Males Present Distinct Susceptibility to Alzheimer Disease-like Pathology. Mol Neurobiol. 6471. [Google Scholar] [CrossRef]
- Chakraborti, A.; Gulati, K.; Banerjee, B.D.; Ray, A. Possible involvement of free radicals in the differential neurobehavioral responses to stress in male and female rats. Behav. Brain Res. 2007, 179, 321–325. [Google Scholar] [CrossRef]
- Cole, T.B.; Coburn, J.; Dao, K.; Roqué, P.; Chang, Y.C.; Kalia, V.; Guilarte, T.R.; Dziedzic, J.; Costa, L.G. Sex and genetic differences in the effects of acute diesel exhaust exposure on inflammation and oxidative stress in mouse brain. Toxicology. 2016, 374, 1–9. [Google Scholar] [CrossRef]
- Guevara, R.; Santandreu, F.M.; Valle, A.; Gianotti, M.; Oliver, J.; Roca, P. Sex-dependent differences in aged rat brain mitochondrial function and oxidative stress. Free Radic. Biol. Med. 2009, 46, 169–175. [Google Scholar] [CrossRef]
- Katalinic, V.; Modun, D.; Music, I.; Boban, M. Gender differences in antioxidant capacity of rat tissues determined by 2,2′-azinobis (3-ethylbenzothiazoline 6-sulfonate; ABTS) and ferric reducing antioxidant power (FRAP) assays. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2005, 140, 47–52. [Google Scholar] [CrossRef]
- Silva, T.L.A.; Braz, G.R.F.; Silva, S.C.A.; Pedroza, A.A.D.S.; Freitas, C.M.; Ferreira, D.J.S.; da Silva, A.I.; Lagranha, C.J. Serotonin transporter inhibition during neonatal period induces sex-dependent effects on mitochondrial bioenergetics in the rat brainstem. Eur J Neurosci. 2018, 26. [Google Scholar] [CrossRef]
- Sobočanec, S.; Balog, T.; Kušić, B.; Šverko, V.; Šarić, A.; Marotti, T. Differential response to lipid peroxidation in male and female mice with age: correlation of antioxidant enzymes matters. Biogerontology 2008, 9, 335–343. [Google Scholar] [CrossRef]
- Guevara, R.; Gianotti, M.; Roca, P.; Oliver, J. Age and sex-related changes in rat brain mitochondrial function. Cell. Physiol. Biochem. /: 201–206. https, 1016. [Google Scholar]
- Jung, M.E.; Metzger, D.B. A sex difference in oxidative stress and behavioral suppression induced by ethanol withdrawal in rats. Behav. Brain Res. 2016, 314, 199–214. [Google Scholar] [CrossRef]
- Uzun, H.; Kayali, R.; Çakatay, U. The chance of gender dependency of oxidation of brain proteins in aged rats. Arch. Gerontol. Geriatr. /: 16–19. https, 1016. [Google Scholar]
- Khalifa, A.R.; Abdel-Rahman, E.A.; Mahmoud, A.M.; Ali, M.H.; Noureldin, M. , Saber, S.H.; Mohsen, M.; Ali, S.S. Sex-specific differences in mitochondria biogenesis, morphology, respiratory function, and ROS homeostasis in young mouse heart and brain. Physiol Rep. 2017, 5, e13125. [Google Scholar] [CrossRef] [PubMed]
- Lloret, A.; Badía, M.C.; Mora, N.J.; Ortega, A.; Pallardó, F.V.; Alonso, M.D.; Atamna, H.; Viña, J. Gender and age-dependent differences in the mitochondrial apoptogenic pathway in Alzheimer's disease. Free Radic Biol Med. 2008, 44, 2019–2025. [Google Scholar] [CrossRef] [PubMed]
- Dkhil, M.A.; Al-Shaebi, E.M.; Lubbad, M.Y.; Al-Quraishy, S. Impact of sex differences in brain response to infection with Plasmodium berghei. Parasitol. Res. 2016, 115, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Ehrenbrink, G.; Hakenhaar, F.S.; Salomon, T.B.; Petrucci, A.P.; Sandri, M.R.; Benfato, M.S. Antioxidant enzymes activities and protein damage in rat brain of both sexes. Exp. Gerontol. 2006, 41, 368–371. [Google Scholar] [CrossRef] [PubMed]
- Krolow, R.; Noschang, C.G.; Arcego, D.; Andreazza, A.C.; Peres, W.; Gonçalves, C.A.; Dalmaz, C. Consumption of a palatable diet by chronically stressed rats prevents effects on anxiety-like behavior but increases oxidative stress in a sex-specific manner. Appetite 2010, 55, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Mármol, F.; Rodríguez, C.A.; Sánchez, J.; Chamizo, V.D. Anti-oxidative effects produced by environmental enrichment in the hippocampus and cerebral cortex of male and female rats. Brain Res. 2015, 1613, 120–129. [Google Scholar] [CrossRef] [PubMed]
- 88 Noschang, C.; Krolow, R.; Arcego, D.M.; Toniazzo, A.P.; Huffell, A.P.; Dalmaz, C. Neonatal handling affects learning, reversal learning and antioxidant enzymes activities in a sex-specific manner in rats. Int. J. Dev. Neurosci. 2012, 30, 285–291. [Google Scholar] [CrossRef]
- Brocardo, P.S.; Boehme, F.; Patten, A.; Cox, A.; Gil-Mohapel, J.; Christie, B.R. , Anxiety and depression-like behaviors are accompanied by an increase in oxidative stress in a rat model of fetal alcohol spectrum disorders: protective effects of voluntary physical exercise. Neuropharmacology 2012, 62, 1607–1618. [Google Scholar] [CrossRef]
- Charradi, K.; Mahmoudi, M.; Bedhiafi, T.; Kadri, S.; Elkahoui, S.; Limam, F.; Aouani, E. Dietary supplementation of grape seed and skin flour mitigates brain oxidative damage induced by a high-fat diet in rat: Gender dependency. Biomed Pharmacother. 2017, 87, 519–526. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; García, Y.; Buccieri, K.; Revilla, S.; Suñol, C.; Cristofol, R.; Sanfeliu, C. Gender-specific neuroimmunoendocrine response to treadmill exercise in 3xTg-AD mice. Int. J. Alzheimer's Dis. 1283. [Google Scholar] [CrossRef]
- Harish, G.; Venkateshappa, C.; Mahadevan, A.; Pruthi, N. ; Srinivas, Bharath, M.M.; Shankar, S.K. Effect of premortem and postmortem factors on the distribution and preservation of antioxidant activities in the cytosol and synaptosomes of human brains. Biopreserv. Biobanking. [CrossRef]
- Engler-Chiurazzi, E.B.; Brown, C.M.; Povroznik, J.M.; Simpkins, J.W. Estrogens as neuroprotectants: estrogenic actions in the context of cognitive aging and brain injury. Prog. Neurobiol. 2017, 157, 188–211. [Google Scholar] [CrossRef]
- Liu, M.; Kelley, M.H.; Herson, P.S.; Hurn, P.D. Neuroprotection of sex steroids. Minerva Endocrinol. 2010, 127–143. [Google Scholar]
- Siddiqui, A.N.; Siddiqui, N.; Khan, R.A.; Kalam, A.; Jabir, N.R.; Kamal, M.A.; Firoz, C.K.; Tabrez, S. Neuroprotective Role of Steroidal Sex Hormones: An Overview. CNS Neurosci Ther. 2016, 22, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Spychala, M.S.; Honarpisheh, P.; McCullough, L.D. Sex differences in neuroinflammation and neuroprotection in ischemic stroke. J. Neurosci. Res. 2017, 95, 462–471. [Google Scholar] [CrossRef]
- Quillinan, N.; Deng, G.; Grewal, H.; Herson, P.S. Androgens and stroke: good, bad or indifferent? Exp. Neurol. 2014, 259, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Reddy, D.S.; Bakshi, K. Neurosteroids: biosynthesis, molecular mechanisms, and neurophysiological functions in the human brain. Horm. Signal. Biol. Med. 2020, 69–82. [Google Scholar] [CrossRef]
- Yilmaz, C.; Karali, K.; Fodelianaki, G.; Gravanis, A.; Chavakis, T.; Charalampopoulos, I.; Alexaki, V.I. Neurosteroids as regulators of neuroinflammation. Front Neuroendocrinol. 2019, 55, 100788. [Google Scholar] [CrossRef] [PubMed]
- Illan-Cabeza, N.A.; Garcia-Garcia, A.R.; Martinez-Martos, J.M.; Ramirez-Exposito, M.J.; Pena-Ruiz, T.; Moreno-Carretero, M.N. A potential antitumor agent, (6-amino-1-methyl-5-nitrosouracilato-N3)-triphenylphosphinegold(I): Structural studies and in vivo biological effects against experimental glioma. European Journal of Medical Chemistry,.
- Ramirez-Exposito, M.J.; Martinez-Martos, J.M. The Delicate Equilibrium between Oxidants and Antioxidants in Brain Glioma. Current Neuropharmacology,.
- Martínez-MartosJ.M., *!!! REPLACE !!!*; Mayas, M.D.; Carrera, P.; Arias de Saavedra, J.M.; Sánchez-Agesta, R.; Marcela Arrazola, M.; Ramírez-Expósito, M.J. 103. Martínez-MartosJ.M.; Mayas, M.D.; Carrera P.; Arias de Saavedra, J.M.; Sánchez-Agesta, R.; Marcela Arrazola, M.; Ramírez-Expósito, M.J. Phenolic compounds oleuropein and hydroxytyrosol exert differential effects on glioma development via antioxidant defense systems. J Functional Food.
- Ramírez-Expósito, M.J.; Carrera-González, M.P.; Mayas, M.D.; Martínez-Martos, J.M. Gender differences in the antioxidant response of oral administration of hydroxytyrosol and oleuropein against N-ethyl-N-nitrosourea (ENU)-induced glioma. Food Res Int. 2021, 140, 110023. [Google Scholar] [CrossRef]
- Ramírez-Expósito, M.J.; Mayas, M.D.; Carrera-González, M.P.; Martínez-Martos, J.M. Gender Differences in the Antioxidant Response to Oxidative Stress in Experimental Brain Tumors. Curr Cancer Drug Targets. 2019, 19, 641–654. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- Villanueva, A.; Llovet, J.M. Liver cancer in 2013: mutational landscape of HCC-the end of the beginning. Nat. Rev. Clin. Oncol. 2014, 11, 73–74. [Google Scholar] [CrossRef]
- El-Serag, H.B. Hepatocellular carcinoma. N Engl J Med. 2011, 365, 1118–1127. [Google Scholar] [CrossRef]
- Guy, J.; Peters, M.G. Liver disease in women: the influence of gender on epidemiology, natural history, and patient outcomes. Gastroenterol. Hepatol. 2013, 9, 633–639. [Google Scholar]
- Vesselinovitch, S.D.; Mihailovich, N.; Wogan, G.N.; Lombard, L.S.; Rao, K.V.; Sugamori, K.S.; Brenneman, D.; Sanchez, O.; Doll, M.A.; Hein, D.W.; et al. Reduced 4-aminobiphenyl-induced liver tumorigenicity but not DNA damage in arylamine N-acetyltransferase null mice. Cancer Lett. 2012, 318, 206–213. [Google Scholar] [CrossRef]
- Gramenzi, A.; Caputo, F.; Biselli, M.; Kuria, F.; Loggi, E.; Andreone, P.; Bernardi, M. Review article: alcoholic liver disease--pathophysiological aspects and risk factors. Aliment Pharmacol Ther. 2006, 24, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Harada, S. Classification of alcohol metabolizing enzymes and polymorphisms--specificity in Japanese. Nihon Arukoru Yakubutsu Igakkai Zasshi. 2001, 36, 85–106. [Google Scholar] [PubMed]
- Brandon-Warner, E.; Walling, T.L.; Schrum, L.W.; McKillop, I.H. Chronic ethanol feeding accelerates hepatocellular carcinoma progression in a sex-dependent manner in a mouse model of hepatocarcinogenesis. Alcohol Clin Exp Res. 2012, 36, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, K.; Komura, S.; Watanabe, I.; Nakagawa, Y.; Yagi, K. Effect of estrogens on serum and liver lipid peroxide levels in mice. J Clin Biochem Nutr 1987, 3, 233–239. [Google Scholar] [CrossRef]
- Lacort, M.; Leal, A.M.; Liza, M.; Martín, C.; Martínez, R.; Ruiz-Larrea, M.B. Protective effect of estrogens and catecholestrogens against peroxidative membrane damage in vitro. Lipids 1995, 30, 141–146. [Google Scholar] [CrossRef]
- Omoya, T.; Shimizu, I.; Zhou, Y.; Okamura, Y.; Inoue, H.; Lu, G.; Itonaga, M.; Honda, H.; Nomura, M.; Ito, S. Effects of idoxifene and estradiol on NF-kappaB activation in cultured rat hepatocytes undergoing oxidative stress. Liver 2001, 21, 183–191. [Google Scholar] [CrossRef]
- Inoue, H.; Shimizu, I.; Lu, G.; Itonaga, M.; Cui, X.; Okamura, Y.; Shono, M.; Honda, H.; Inoue, S.; Muramatsu, M.; Ito, S. Idoxifene and estradiol enhance antiapoptotic activity through estrogen receptor-beta in cultured rat hepatocytes. Dig Dis Sci 2003, 48, 570–580. [Google Scholar] [CrossRef]
- Tanaka, Y.; Mukaide, M.; Orito, E.; Yuen, M.F.; Ito, K.; Kurbanov, F.; Sugauchi, F.; Asahina, Y.; Izumi, N.; Kato, M.; Lai, C.L.; Ueda, R.; Mizokami, M. Specific mutations in enhancer II/core promoter of hepatitis B virus subgenotypes C1/C2 increase the risk of hepatocellular carcinoma. J Hepatol 2006, 45, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Slocum, S.L.; Kensler, T.W. Nrf2: control of sensitivity to carcinogens. Arch Toxicol. 2011, 85, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, Y.; Umemura, T.; Kanki, K.; Kodama, Y.; Kitamoto, S.; Saito, K.; Itoh, K.; Yamamoto, M.; Masegi, T.; Nishikawa, A.; Hirose, M. Increased susceptibility to hepatocarcinogenicity of Nrf2-deficient mice exposed to 2-amino-3-methylimidazo [4,5-f]quinoline. Cancer Sci. 2007, 98, 19–24. [Google Scholar] [CrossRef]
- Thimmulappa, R.K.; Lee, H.; Rangasamy, T.; Reddy, S.P.; Yamamoto, M.; Kensler, T.W.; Biswal, S. Nrf2 is a critical regulator of the innate immune response and survival during experimental sepsis. J Clin Invest. 2006, 116, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Wruck, C.J.; Streetz, K.; Pavic, G.; Götz, M.E.; Tohidnezhad, M.; Brandenburg, L.O.; Varoga, D.; Eickelberg, O.; Herdegen, T.; Trautwein, C.; Cha, K.; Kan, Y.W.; Pufe, T. Nrf2 induces interleukin-6 (IL-6) expression via an antioxidant response element within the IL-6 promoter. J Biol Chem. 2011, 286, 4493–4499. [Google Scholar] [CrossRef] [PubMed]
- Hanna, D.; Riedmaier, A.E.; Sugamori, K.S.; Grant, D.M. Influence of sex and developmental stage on acute hepatotoxic and inflammatory responses to liver procarcinogens in the mouse. Toxicology 2016, 373, 30–40. [Google Scholar] [CrossRef]
- Shupe, T.; Sell, S. Low hepatic glutathione S-transferase and increased hepatic DNA adduction contribute to increased tumorigenicity of aflatoxin B1 in newborn and partially hepatectomized mice. Toxicol Lett. [CrossRef]
- Eaton, D.L.; Gallagher, EP. Mechanisms of aflatoxin carcinogenesis. Annu Rev Pharmacol Toxicol. 1994, 34, 135–172. [Google Scholar] [CrossRef]
- Egner, P.A.; Groopman, J.D.; Wang, J.S.; Kensler, T.W.; Friesen, M.D. Quantification of aflatoxin-B1-N7-Guanine in human urine by high-performance liquid chromatography and isotope dilution tandem mass spectrometry. Chem Res Toxicol. 2006, 19, 1191–1195. [Google Scholar] [CrossRef]
- Crawford, D.R.; Ilic, Z.; Guest, I.; Milne, G.L.; Hayes, J.D.; Sell, S. Characterization of liver injury, oval cell proliferation and cholangiocarcinogenesis in glutathione S-transferase A3 knockout mice. Carcinogenesis. 2017, 38, 717–727. [Google Scholar] [CrossRef]
- Fu, D.; Hornick, C.A. Modulation of lipid metabolism at rat hepatic subcellular sites by female sex hormones. Biochim Biophys Acta 1995, 1254, 267–273. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.; Lavigne, J.A.; Trush, M.A.; Yager, J.D. (1999) Increased mitochondrial superoxide production in rat liver mitochondria, rat hepatocytes, and HepG2 cells following ethinyl estradiol treatment. Toxicol Sci 1990, 52, 224–235. [Google Scholar]
- Gerber, M.; Astre, C.; Segala, C.; Saintot, M.; Scali, J.; Simony Lafontaine, J.; Grenier, J.; Pujol, H. Tumor progression and oxidant-antioxidant status. Cancer Lett 1997, 114, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Sverko, V.; Sobocanec, S.; Balog, T.; Marotti, T. Age and gender differences in antioxidant enzyme activity: potential relationship to liver carcinogenesis in male mice. Biogerontology. 2004, 5, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Coto-Montes, A.; Boga, J.A.; Tomas-Zapico, C.; Rodriguez Colunga, M.J.; Martinez-Fraga, J.; Tolivia-Cadrecha, D.; Manendez, G.; Herdeband, R.; Tolivia, D. Physiological oxidative stress model: syrian hamster harderian gland-sex differences in antioxidant enzymes. Free Radical Bio Med 2001, 30, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Oberley, L.W.; Oberley, T.D. Role of antioxidant enzymes in cell immortalization and transformation. Mol Cell Biochem 1988, 84, 147–153. [Google Scholar] [CrossRef]
- Janssen, A.M.; Bosman, C.B.; Sier, C.F.; Griffioen, G.; Kubben, F.J.; Lamers, C.B.; van Krieken, J.H.; van de Velde, C.J.; Verspaget, H.W. Superoxide dismutase in relation to the overall survival of colorectal cancer patients. Br J Cancer 1998, 78, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.K.; Zhang, H.J.; Yan, T.; Oberley, L.W.; Aggarwal, B.B. Overexpression of manganese superoxide dismutase suppresses tumor necrosis factor-induced apoptosis and activation of nuclear transcription factor-jB and activated protein-1. J Biol Chem 1998, 273, 13245–13254. [Google Scholar] [CrossRef]
- Szatrowski, T.P.; Nathan, C.F. Production of large amounts of hydrogen peroxide by human tumor cells. Cancer Res 1991, 51, 794–798. [Google Scholar]
- Gardner, R.; Salvador, A.; Moradas-Ferreira, P. Why does SOD overexpression sometimes enhance, sometimes decrease, hydrogen peroxide production? A minimalist explanation. Free Radical Bio Med 2022, 32, 1352–1357. [Google Scholar] [CrossRef]
- Pigeolet, E.; Corbisier, P.; Houbion, A.; Lambert, D.; Michiels, C.; Raes, M.; Zachary, M.D.; Remacle, J. Glutathione peroxidase, superoxide dismutase and catalase inactivation by peroxides and oxygen-derived free radicals. Mech Ageing Dev.
- Gonzales, M.J. Lipid peroxidation and tumor growth: an inverse relationship. Med Hypotheses 1992, 38, 106–110. [Google Scholar] [CrossRef]
- Wang S, Sugamori KS, Tung A, McPherson JP, Grant DM. N-hydroxylation of 4-aminobiphenyl by CYP2E1 produces oxidative stress in a mouse model of chemically induced liver cancer. Toxicol Sci. 2015 Apr;144:393-405. [CrossRef]
- Calle, E.E.; Rodriguez, C.; Walker-Thurmond, K.; Thun, M.J. Overweight, obesity, and mortality from cancer in a prospectively studied cohort of U.S. adults. N. Engl. J. Med. 2003, 348, 1625–1638. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, K.; Douglas, I.; Forbes, H.; dos-Santos-Silva, I.; Leon, D.A.; Smeeth, L. Body-mass index and risk of 22 specific cancers: a population-based cohort study of 5.24 million UK adults. Lancet 2014, 384, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Świątkiewicz, I.; Wróblewski, M.; Nuszkiewicz, J.; Sutkowy, P.; Wróblewska, J.; Woźniak, A. The Role of Oxidative Stress Enhanced by Adiposity in Cardiometabolic Diseases. Int J Mol Sci. 2023, 24, 6382. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M. Subcutaneous and visceral adipose tissue: structural and functional differences. Obes. Rev. 2010, 11, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Setiawan, V.W.; Lim, U.; Lipworth, L.; Lu, S.C.; Shepherd, J.; Ernst, T.; Wilkens, L.R.; Henderson, B.E.; Le Marchand, L. Sex and ethnic differences in the association of obesity with risk of hepatocellular carcinoma. Clin. Gastroenterol. Hepatol. 2016, 14, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Jung, H.S.; Cho, J.; Zhang, Y.; Yun, K.E.; Lazo, M.; Pastor-Barriuso, R.; Ahn, J.; Kim, C.W.; Rampal, S.; et al. Metabolically healthy obesity and the development of nonalcoholic fatty liver disease. Am. J. Gastroenterol. 2016, 111, 1133–1140. [Google Scholar] [CrossRef]
- Freedland, E.S. Roles of critical visceral adipose tissue threshold in metabolic syndrome: implications for controlling dietary carbohydrates: a review. Nutr. Metab. 2004, 1, 12. [Google Scholar] [CrossRef]
- Björntorp, P. Endocrine abnormalities in obesity. Metabolism. 1995, 44(9 Suppl. 3), 21–23. [Google Scholar] [CrossRef]
- He, J.; Gu, D.; Wu, X.; Reynolds, K.; Duan, X.; Yao, C.; Wang, J.; Chen, C.S.; Chen, J.; Wildman, R.P.; Klag, M.J.; Whelton, PK. Major causes of death among men and women in China. N Engl J Med. 2005, 353, 1124–1134. [Google Scholar] [CrossRef]
- Riley, G.F.; Potosky, A.L.; Lubitz, J.D.; Kessler, L.G. Medicare payments from diagnosis to death for elderly cancer patients by stage at diagnosis. Med Care. 1995, 33, 828–841. [Google Scholar] [CrossRef]
- Ohnishi, S.; Murata, M.; Kawanishi, S. DNA damage induced by hypochlorite and hypobromite with reference to inflammation-associated carcinogenesis. Cancer Lett,.
- Kiyohara, C.; Yoshimasu, K.; Takayama, K. , Nakanishi, Y. NQO1, MPO, and the risk of lung cancer: a HuGE review. Genet Med. [CrossRef]
- Yang, M.; Choi, Y.; Hwangbo, B.; Lee, J.S. Combined effects of genetic polymorphisms in six selected genes on lung cancer susceptibility. Lung Cancer. 2007, 57, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ambrosone, C.B.; Hong, C.C.; Ahn, J.; Rodriguez, C.; Thun, M.J.; Calle, E.E. ; Relationships between polymorphisms in NOS3 and MPO genes, cigarette smoking and risk of post-menopausal breast cancer. Carcinogenesis. 2007, 28, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Steenport, M.; Eom, H.; Uezu, M.; Schneller, J.; Gupta, R.; Mustafa, Y.; Villanueva, R.; Straus, E.W.; Raffaniello, R.D. Association of polymorphisms in myeloperoxidase and catalase genes with precancerous changes in the gastric mucosa of patients at inner-city hospitals in New York. Oncol Rep. 2007, 18, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Cascorbi, I. ; Henning S, Brockmöller J, Gephart J, Meisel C, Müller JM, Loddenkemper R, Roots I. Substantially reduced risk of cancer of the aerodigestive tract in subjects with variant--463A of the myeloperoxidase gene. Cancer Res.
- Li, Y.; Qin, Y.; Wang, M.L.; Zhu, H.F.; Huang, X.E. The myeloperoxidase-463 G>A polymorphism influences risk of colorectal cancer in southern China: a case-control study. Asian Pac J Cancer Prev. 2011, 12, 1789–1793. [Google Scholar] [PubMed]
- Otero Regino, W.; Velasco, H.; Sandoval, H. The protective role of bilirubin in human beings. Rev. Colomb. Gastroenterol. 2009, 24, 293–301. [Google Scholar]
- Wagner, K.H.; Wallner, M.; Molzer, C.; Gazzin, S.; Bulmer, A.C.; Tiribelli, C.; Vitek, L. Looking to the horizon: The role of bilirubin in the development and prevention of age-related chronic diseases. Clin. Sci. 2015, 129, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, T.W.; Saleh, M.; Higginson, D.S.; Paul, B.D.; Juluri, K.R.; Snyder, S.H. Bilirubin and glutathione have complementary antioxidant and cytoprotective roles. Proc. Natl. Acad. Sci. USA 2009, 106, 5171–5176. [Google Scholar] [CrossRef]
- Rodrigues, C.M.; Sola, S.; Brito, M.A.; Brites, D.; Moura, J.J. Bilirubin directly disrupts membrane lipid polarity and fluidity, protein order, and redox status in rat mitochondria. J. Hepatol. 2002, 36, 335–341. [Google Scholar] [CrossRef]
- Hansen, T.W.; Mathiesen, S.B.; Walaas, S.I. Bilirubin has widespread inhibitory effects on protein phosphorylation. Pediatr. Res. 1996, 39, 1072–1077. [Google Scholar] [CrossRef]
- Fevery, J. Bilirubin in clinical practice: A review. Liver Int. Off. J. Int. Assoc. Stud Liver 2008, 28, 592–605. [Google Scholar] [CrossRef]
- Bulmer, A.C.; Ried, K.; Coombes, J.S.; Blanchfield, J.T.; Toth, I.; Wagner, K.H. The anti-mutagenic and antioxidant effects of bile pigments in the Ames Salmonella test. Mutat. Res. 2007, 629, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Molzer, C.; Huber, H.; Diem, K.; Wallner, M.; Bulmer, A.C.; Wagner, K.H. Extracellular and intracellular anti-mutagenic effects of bile pigments in the Salmonella typhimurium reverse mutation assay. Toxicol. Int. J. Public Assoc. BIBRA 2013, 27, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Molzer, C.; Huber, H.; Steyrer, A.; Ziesel, G.; Ertl, A.; Plavotic, A.; Wallner, M.; Bulmer, A.C.; Wagner, K.H. In vitro antioxidant capacity and antigenotoxic properties of protoporphyrin and structurally related tetrapyrroles. Free Radic. Res. 2012, 46, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Molzer, C.; Huber, H.; Steyrer, A.; Ziesel, G.V.; Wallner, M.; Hong, H.T.; Blanchfield, J.T.; Bulmer, A.C.; Wagner, K.H. Bilirubin and related tetrapyrroles inhibit food-borne mutagenesis: A mechanism for antigenotoxic action against a model epoxide. J. Nat. Prod. 2013, 76, 1958–1965. [Google Scholar] [CrossRef] [PubMed]
- Seyed Khoei, N.; Jenab, M.; Murphy, N.; Banbury, B.L.; Carreras-Torres, R.; Viallon, V.; Kühn, T.; Bueno-de-Mesquita, B.; Aleksandrova, K.; Cross, A.J.; et al. Circulating bilirubin levels and risk of colorectal cancer: Serological and Mendelian randomization analyses. BMC Med. 2020, 18, 229. [Google Scholar] [CrossRef] [PubMed]
- Seyed Khoei, N.; Anton, G.; Peters, A.; Freisling, H.; Wagner. K.H. The Association between Serum Bilirubin Levels and Colorectal Cancer Risk: Results from the Prospective Cooperative Health Research in the Region of Augsburg (KORA) Study in Germany. Antioxidants (Basel). 2020, 9, 908. [Google Scholar] [CrossRef]
- Ollinger, R.; Kogler, P.; Troppmair, J.; Hermann, M.; Wurm, M.; Drasche, A.; Konigsrainer, I.; Amberger, A.; Weiss, H.; Ofner, D.; et al. Bilirubin inhibits tumor cell growth via activation of ERK. Cell Cycle Georgetown Tex. 2007, 6, 3078–3085. [Google Scholar] [CrossRef]
- Keshavan, P.; Schwemberger, S.J.; Smith, D.L.; Babcock, G.F.; Zucker, S.D. Unconjugated bilirubin induces apoptosis in colon cancer cells by triggering mitochondrial depolarization. Int. J. Cancer 2004, 112, 433–445. [Google Scholar] [CrossRef]
- Grant, D.J.; Bell, D.A. Bilirubin UDP-glucuronosyltransferase 1A1 gene polymorphisms: Susceptibility to oxidative damage and cancer? Mol. Carcinogen. 2000, 29, 198–204. [Google Scholar] [CrossRef]
- Zhang, T.; Joubert, P.; Ansari-Pour, N.; Zhao, W.; Hoang, P.H.; Lokanga, R.; Moye, A.L.; Rosenbaum, J.; Gonzalez-Perez, A.; Martínez-Jiménez, F.; et al. Genomic and Evolutionary Classification of Lung Cancer in Never Smokers. Nat. Genet. 2021, 53, 1348–1359. [Google Scholar] [CrossRef]
- Adib, E.; Nassar, A.H.; Abou Alaiwi, S.; Groha, S.; Akl, E.W.; Sholl, L.M.; Michael, K.S.; Awad, M.M.; Jänne, P.A.; Gusev, A.; et al. Variation in Targetable Genomic Alterations in Non-Small Cell Lung Cancer by Genetic Ancestry, Sex, Smoking History, and Histology. Genome Med. 2022, 14, 39. [Google Scholar] [CrossRef] [PubMed]
- Gasperino, J.; Rom, W.N. Gender and lung cancer. Clin Lung Cancer 2004, 5, 353–359. [Google Scholar] [CrossRef] [PubMed]
- 176. Thomas L, Doyle LA, Edelman MJ. 2005. Lung cancer in women: Emerging differences in epidemiology, biology, and therapy. Chest.
- Kiyohara, C.; Ohno, Y. Sex differences in lung cancer susceptibility: A review. Gend Med 2010, 7, 381–401. [Google Scholar] [CrossRef] [PubMed]
- Henschke, C.I.; Yip, R.; Miettinen, O.S. Women’s susceptibility to tobacco carcinogens and survival after diagnosis of lung cancer. JAMA 2006, 296, 180–184. [Google Scholar] [PubMed]
- Parajuli R, Bjerkaas E, Tverdal A, Selmer R, Le Marchand L, Weiderpass E, Gram IT. 2013. The increased risk of colon cancer due to cigarette smoking may be greater in women than men. Cancer Epidemiol Biomarkers Prev 22:862–871.
- Forder, A.; Zhuang, R.; Souza, V.G.P.; Brockley, L.J.; Pewarchuk, M.E.; Telkar, N.; Stewart, G.L.; Benard, K.; Marshall, E.A.; Reis, P.P.; et al. Mechanisms Contributing to the Comorbidity of COPD and Lung Cancer. Int. J. Mol. Sci. 2023, 24, 2859. [Google Scholar] [CrossRef] [PubMed]
- Dewhirst, M.W.; Cao, Y.; Moeller, B. Cycling Hypoxia and Free Radicals Regulate Angiogenesis and Radiotherapy Response. Nat. Rev. Cancer 2008, 8, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Covey, T.M.; Edes, K.; Coombs, G.S.; Virshup, D.M.; Fitzpatrick, F.A. Alkylation of the Tumor Suppressor PTEN Activates Akt and β-Catenin Signaling: A Mechanism Linking Inflammation and Oxidative Stress with Cancer. PLoS ONE 2010, 5, e13545. [Google Scholar] [CrossRef]
- Cai, B.; Liu, M.; Li, J.; Xu, D.; Li, J. Cigarette Smoke Extract Amplifies NADPH Oxidase-Dependent ROS Production to Inactivate PTEN by Oxidation in BEAS-2B Cells. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2021, 150, 112050. [Google Scholar]
- Shukla, S.D.; Shastri, M.D.; Jha, N.K.; Gupta, G.; Chellappan, D.K.; Bagade, T.; Dua, K. Female Gender as a Risk Factor for Developing COPD. EXCLI J. 2021, 20, 1290–1293. [Google Scholar]
- Ben-Zaken Cohen, S.; Paré, P.D.; Man, S.F.P.; Sin, D.D. The Growing Burden of Chronic Obstructive Pulmonary Disease and Lung Cancer in Women: Examining Sex Differences in Cigarette Smoke Metabolism. Am. J. Respir. Crit. Care Med. 2007, 176, 113–120. [Google Scholar] [CrossRef]
- Meireles, S.I.; Esteves, G.H.; Hirata, R.J.; Peri, S.; Devarajan, K.; Slifker, M.; Mosier, S.L.; Peng, J.; Vadhanam, M.V.; Hurst, H.E.; et al. Early Changes in Gene Expression Induced by Tobacco Smoke: Evidence for the Importance of Estrogen within Lung Tissue. Cancer Prev. Res. 2010, 3, 707–717. [Google Scholar] [CrossRef] [PubMed]
- Mollerup, S.; Ryberg, D.; Hewer, A.; Phillips, D.H.; Haugen, A. Sex Differences in Lung CYP1A1 Expression and DNA Adduct Levels among Lung Cancer Patients. Cancer Res. 1999, 59, 3317–3320. [Google Scholar] [PubMed]
- Van Winkle, L.S.; Gunderson, A.D.; Shimizu, J.A.; Baker, G.L.; Brown, C.D. Gender Differences in Naphthalene Metabolism and Naphthalene-Induced Acute Lung Injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 282, L1122–L1134. [Google Scholar] [CrossRef] [PubMed]
- Soriano, J.B.; Kendrick, P.J.; Paulson, K.R.; Gupta, V.; Abrams, E.M.; Adedoyin, R.A.; Adhikari, T.B.; Advani, S.M.; Agrawal, A.; Ahmadian, E.; et al. Prevalence and Attributable Health Burden of Chronic Respiratory Diseases, 1990-2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet Respir. Med. 2020, 8, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Venegas, A.; Sansores, R.H.; Pérez-Padilla, R.; Regalado, J.; Velázquez, A.; Sánchez, C.; Mayar, M.E. Survival of Patients with Chronic Obstructive Pulmonary Disease Due to Biomass Smoke and Tobacco. Am. J. Respir. Crit. Care Med. 2006, 173, 393–397. [Google Scholar] [CrossRef]
- Armstrong, B.G.; Gibbs, G. Exposure-response relationship between lung cancer and polycyclic aromatic hydrocarbons (PAHs). Occup Environ Med 2009, 66, 740–746. [Google Scholar] [CrossRef]
- Veglia, F.; Matullo, G.; Vineis, P. Bulky DNA adducts and risk of cancer: A meta-analysis. Cancer Epidemiol Biomarkers Prev 2003, 12, 157–160. [Google Scholar]
- Bostrom, C.E.; Gerde, P.; Hanberg, A.; Jernstrom, B.; Johansson, C.; Kyrklund, T.; Rannug, A.; Tornqvist, M.; Victorin, K.; Westerholm, R. Cancer risk assessment, indicators, and guidelines for polycyclic aromatic hydrocarbons in the ambient air. Environ Health Perspect 2002, 110 (Suppl 3), 451–488. [Google Scholar]
- Guo, H.; Huang, K. , Zhang, X., Zhang, W.; Guan, L., Kuang, D., Deng, Q.; Deng, H., Zhang, X., He, M., Christiani, D., Wu, T. Women are more susceptible than men to oxidative stress and chromosome damage caused by polycyclic aromatic hydrocarbons exposure. Environ Mol Mutagen. 2014, 55, 472–481. [Google Scholar] [CrossRef]
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2013. CA Cancer J Clin. 2013, 63, 11–30. [Google Scholar] [CrossRef]
- Balch, C.M.; Gershenwald, J.E.; Soong, S.J.; Thompson, J.F. , Atkins, M.B.; Byrd, D.R.; Buzaid, A.C.; Cochran, A.J.; Coit, D.G.; Ding, S.; Eggermont, A.M.; et al. Final version of 2009 AJCC melanoma staging and classification. J Clin Oncol. 2009, 27, 6199–6206. [Google Scholar] [CrossRef] [PubMed]
- De Vries, E.; Nijsten, T.E.; Visser, O.; Bastiaannet, E.; Van Hattem, S.; Janssen-Heijnen, M.L.; Coebergh, J.W. Superior survival of females among 10,538 Dutch melanoma patients is independent of Breslow thickness, histologic type and tumor site. Ann. Oncol. 2008, 19, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Lasithiotakis, K.; Leiter, U.; Meier, F.; Eigentler, T.; Metzler, G.; Moehrle, M.; Breuninger, H.; Garbe, C. Age and gender are significant independent predictors of survival in primary cutaneous melanoma. Cancer 2008, 112, 1795–1804. [Google Scholar] [CrossRef] [PubMed]
- Richardson, B.; Price, A.; Wagner, M.; Williams, V.; Lorigan, P.; Browne, S.; Miller, J.G.; Mac Neil, S. Investigation of female survival benefit in metastatic melanoma. Br. J. Cancer 1999, 80, 2025–2033. [Google Scholar] [CrossRef] [PubMed]
- Kemeny, M.M.; Busch, E.; Stewart, A.K.; Menck, H.R. Superior survival of young women with malignant melanoma. Am. J. Surg.
- Daryanani, D.; Plukker, J.T.; De Jong, M.A.; Haaxma-Reiche, H.; Nap, R.; Kuiper, H.; Hoekstra, H.J. Increased incidence of brain metastases in cutaneous head and neck melanoma. Melanoma Res. 2005, 15, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Scoggins, C.R.; Ross, M.I.; Reintgen, D.S.; Noyes, R.D.; Goydos, J.S.; Beitsch, P.D.; Urist, M.M.; Ariyan, S.; Sussman, J.J.; Edwards, M.J. , Chagpar, A.B.; Martin, R.C.; Stromberg, A.J.; Hagendoorn, L.; McMasters, K.M.; Sunbelt Melanoma Trial. Gender-related differences in outcome for melanoma patients. Ann Surg. 2006, 243, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Joosse, A.; Collette, S.; Suciu, S.; Nijsten, T.; Lejeune, F.; Kleeberg, U.R.; Coebergh, J.W.; Eggermont, A.M.; de Vries, E. Superior outcome of women with stage I/II cutaneous melanoma: pooled analysis of four European Organisation for Research and Treatment of Cancer phase III trials. J Clin Oncol. 2012, 30, 2240–2247. [Google Scholar] [CrossRef]
- Joosse, A.; de Vries, E.; Eckel, R.; Nijsten, T.; Eggermont, A.M.; Hölzel, D.; Coebergh, J.W.; Engel, J. ; Munich Melanoma Group. Gender differences in melanoma survival: female patients have a decreased risk of metastasis. J Invest Dermatol. [CrossRef]
- Sondak, V.K.; Swetter, S.M.; Berwick, M.A. Gender disparities in patients with melanoma: breaking the glass ceiling. J Clin Oncol. 2012, 30, 2177–2178. [Google Scholar] [CrossRef]
- Gamba, C.S.; Clarke, C.A.; Keegan, T.H.; Tao, L.; Swetter, S.M. Melanoma survival disadvantage in young, non-Hispanic white males compared with females. JAMA Dermatol. 2013, 149, 912–920. [Google Scholar] [CrossRef]
- Fruehauf, J.P.; Trapp, V. Reactive oxygen species: an Achilles’ heel of melanoma? Expert Rev. Anticancer Ther. 2008, 8, 1751–1757. [Google Scholar] [CrossRef]
- Cannavò, S.P.; Tonacci, A.; Bertino, L.; Casciaro, M.; Borgia, F.; Gangemi, S. The role of oxidative stress in the biology of melanoma: A systematic review. Pathol Res Pract. 2019, 215, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Bittinger, F.; Gonzalez-Garcia, J.L.; Klein, C.L.; Brochhausen, C.; Offner, F.; Kirkpatrick, C.J. Production of superoxide by human malignant melanoma cells. Melanoma Res.
- Meyskens, F.L. Jr.; Chau, H.V.; Tohidian, N.; Buckmeier, J. Luminol-enhanced chemiluminescent response of human melanocytes and melanoma cells to hydrogen peroxide stress. Pigment Cell Res. 1997, 10, 184–189. [Google Scholar] [CrossRef]
- Joosse, A.; De Vries, E.; van Eijck, C.H.; Eggermont, A.M.; Nijsten, T.; Coebergh, J.W. Reactive oxygen species and melanoma: an explanation for gender differences in survival? Pigment Cell Melanoma Res. 2010, 23, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Sander, C.S.; Hamm, F.; Elsner, P.; Thiele, J.J. Oxidative stress in malignant melanoma and non-melanoma skin cancer. Br J Dermatol. 2003, 148, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Meyskens, F.L. Jr.; McNulty, S.E. , Buckmeier, J.A.; Tohidian, N.B.; Spillane, T.J.; Kahlon, R.S., Gonzalez, R.I. Aberrant redox regulation in human metastatic melanoma cells compared to normal melanocytes. Free Radic Biol Med. 2001, 31, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M. Reactive oxygen species in tumor metastasis. Cancer Lett. 2008, 266, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Trouba, K.J.; Hamadeh, H.K.; Amin, R.P.; Germolec, D.R. Oxidative stress and its role in skin disease. Antioxid Redox Signal. 2002, 4, 665–673. [Google Scholar] [CrossRef]
- Thomas-Ahner, J.M.; Wulff, B.C.; Tober, K.L.; Kusewitt, D.F.; Riggenbach, J.A.; Oberyszyn, T.M. Gender differences in UVB-induced skin carcinogenesis, inflammation, and DNA damage. Cancer Res. 2007, 67, 3468–3474. [Google Scholar] [CrossRef]
- Zhong, Q.Y.; Lin, B.; Chen, Y.T.; Huang, Y.P.; Feng, W.P.; Wu, Y.; Long, G.H. , Zou, Y.N.; Liu, Y.; Lin, B.Q.; Sang, N.L.; Zhan, J.Y. Gender differences in UV-induced skin inflammation, skin carcinogenesis and systemic damage. Environ Toxicol Pharmacol. 1035. [Google Scholar] [CrossRef]
- Masback, A. : Olsson, H.; Westerdahl, J.; Ingvar, C.; Jonsson, N. Prognostic factors in invasive cutaneous malignant melanoma: a population-based study and review. Melanoma Res.
- Borrás, C.; Gambini, J.; López-Grueso, R.; Pallardó, F.V.; Viña, J. Direct antioxidant and protective effect of estradiol on isolated mitochondria. Biochim Biophys Acta. 2010, 1802, 205–211. [Google Scholar] [CrossRef]
- Duffield-Lillico, A.J.; Reid, M.E.; Turnbull, B.W.; Combs, G.F. Jr.; Slate, E.H.; Fischbach, L.A.; Marshall, J.R.; Clark, L.C. Baseline characteristics and the effect of selenium supplementation on cancer incidence in a randomized clinical trial: a summary report of the Nutritional Prevention of Cancer Trial. Cancer Epidemiol. Biomarkers Prev. 2002, 11, 630–639. [Google Scholar]
- Hercberg, S.; Ezzedine, K.; Guinot, C.; Preziosi, P.; Galan, P.; Bertrais, S.; Estaquio, C.; Briançon, S.; Favier, A.; Latreille, J.; Malvy, D. Antioxidant supplementation increases the risk of skin cancers in women but not in men. J. Nutr. 2007, 137, 2098–2105. [Google Scholar] [CrossRef] [PubMed]
- Hercberg, S.; Galan, P.; Preziosi, P.; Bertrais, S.; Mennen, L.; Malvy, D.; Roussel, A.M.; Favier, A.; Briancon, S. The SU.VI.MAX Study: a randomized, placebo-controlled trial of the health effects of antioxidant vitamins and minerals. Arch. Intern. Med. 2004, 164, 2335–2342. [Google Scholar] [CrossRef] [PubMed]
- Radkiewicz, C.; Bruchfeld, J.B.; Weibull, C.E.; Jeppesen, M.L.; Frederiksen, H.; Lambe, M.; Jakobsen, L.; El-Galaly, T.C.; Smedby, K.E.; Wästerlid, T. Sex differences in lymphoma incidence and mortality by subtype: A population-based study. Am J Hematol. 2023, 98, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Jacobus, J.A.; Duda, C.G.; Coleman, M.C.; Martin, S.M.; Mapuskar, K.; Mao, G.; Smith, B.J.; Aykin-Burns, N.; Guida, P. , Gius, D.; Domann, F.E.; Knudson, C.M.; Spitz, D.R. Low-dose radiation-induced enhancement of thymic lymphomagenesis in Lck-Bax mice is dependent on LET and gender. Radiat Res. 2013, 180, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Štěrba, M.; Popelová, O.; Vávrová, A. , Jirkovský, E.; Kovaříková, P.; Geršl, V.; Šimůnek, T. Oxidative stress, redox signaling, and metal chelation in anthracycline cardiotoxicity and pharmacological cardioprotection. Antioxid Redox Signal, /: 899–929. https, 1089. [Google Scholar]
- Malorni, W.; Campesi, I.; Straface, E.; Vella, S.; Franconi, F. Redox features of the cell: a gender perspective. Antioxid Redox Signal 2007, 9, 1779–1801. [Google Scholar] [CrossRef] [PubMed]
- Vijay, V.; Han, T.; Moland, C.L.; Kwekel, J.C.; Fuscoe, J.C.; Desai, V.G. (2015) Sexual dimorphism in the expression of mitochondriarelated genes in rat heart at different ages. PLoS One 2015, 10, e0117047. [Google Scholar] [CrossRef]
- Dobbs, N.A.; Twelves, C.J.; Gillies, H.; James, C.A. , Harper, P.G., Rubens, R.D. (1995) Gender affects the doxorubicin pharmacokinetics in patients with normal liver biochemistry. Cancer Chemother Pharmacol 36, 473–476. [CrossRef]
- Wade, J.R.; Kelman, A.W.; Kerr, D.J.; Robert, J.; Whiting, B. Variability in the pharmacokinetics of epirubicin: a population analysis. Cancer Chemother Pharmacol 1992, 29, 391–395. [Google Scholar] [CrossRef]
- Varricchi, G.; Ameri, P.; Cadeddu, C.; Ghigo, A.; Madonna, R.; Marone, G.; Mercurio, V. , Monte, I.; Novo, G.; Parrella, P.; Pirozzi, F.; Pecoraro, A., Spallarossa, P.; Zito, C.; Mercuro, G.; Pagliaro, P.; Tocchetti, C.G. Antineoplastic drug-induced cardiotoxicity: a redox perspective. Front Physiol 2018, 9, 167. 9,. [CrossRef]
- Tocchetti, C.G.; Cadeddu, C.; Di Lisi, D.; Femminò, S.; Madonna, R.; Mele, D.; Monte, I.; Novo, G.; Penna, C.; Pepe, A.; Spallarossa, P.; Varricchi, G.; Zito, C.; Pagliaro, P.; Mercuro, G. From molecular mechanisms to clinical management of antineoplastic drug-induced cardiovascular toxicity: a translational overview. Antioxid Redox Signal 2019, 30, 2110–2153. [Google Scholar] [CrossRef]
- Pupo, M.; Pisano, A.; Abonante, S.; Maggiolini, M.; Musti, A.M. GPER activates Notch signaling in breast cancer cells and cancerassociated fibroblasts (CAFs). Int J Biochem Cell Biol 2014, 46, 56–67. [Google Scholar] [CrossRef]
- Lubecka, K.; Kurzava, L.; Flower, K.; Buvala, H.; Zhang, H.; Teegarden, D.; Camarillo, I.; Suderman, M.; Kuang, S. , Andrisani, O.; Flanagan, J.M.; Stefanska, B. (2016) Stilbenoids remodel the DNA methylation patterns in breast cancer cells and inhibit oncogenic NOTCH signaling through epigenetic regulation of MAML2 transcriptional activity. Carcinogenesis. 37, 656–668. [CrossRef] [PubMed]
- Gonzalez, Y.; Pokrzywinski, K.L.; Rosen, E.T.; Mog, S.; Aryal, B.; Chehab, L.M.; Vijay, V.; Moland, C.L.; Desai, V.G.; Dickey, J.S.; Rao, V.A. Reproductive hormone levels and differential mitochondria-related oxidative gene expression as potential mechanisms for gender differences in cardiosensitivity to doxorubicin in tumor-bearing spontaneously hypertensive rats. Cancer Chemother Pharmacol 2015, 76, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Octavia, Y.T.C.; Gabrielson, K.L.; Janssens, S.; Crijns, H.J.; Moens, A.L. Doxorubicin-induced cardiomyopathy. J Mol Cell Cardiol 2012, 52, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.S.; Burgeiro, A.; Garcia, R.; Moreno, A.J.; Carvalho, R.A.; Oliveira, P.J. Doxorubicin-induced cardiotoxicity: from bioenergetic failure and cell death to cardiomyopathy. Med Res Rev 2014, 34, 106–135. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, S.; Autschbach, R. Doxorubicin in experimental and clinical heart failure. Eur J Cardiothorac Surg. [CrossRef]
- Kumar, S.; Marfatia, R.; Tannenbaum, S.; Yang, C.; Avelar, E. Doxorubicin-induced cardiomyopathy 17 years after chemotherapy. Texas Heart Inst J 2012, 39, 424–427. [Google Scholar]
- Dimitrova, K.R.; DeGroot, K.; Myers, A.K.; Kim, Y.D. Estrogen and homocysteine. Cardiovasc Res 2002, 53, 577–588. [Google Scholar] [CrossRef]
- Ogita, H.; Node, K.; Kitakaze, M. The role of estrogen and estrogen-related drugs in cardiovascular diseases. Curr Drug Metab. 2003, 4, 497–504. [Google Scholar] [CrossRef]
- Hershman, D.L.; McBride, R.B.; Eisenberger, A.; Tsai, W.Y.; Grann, V.R.; Jacobson, J.S. Doxorubicin, cardiac risk factors, and cardiac toxicity in elderly patients with diffuse B-cell non-Hodgkin’s lymphoma. J Clin Oncol 2008, 26, 3159–3165. [Google Scholar] [CrossRef]
- Biancaniello, T.; Meyer, R.A.; Wong, K.Y.; Sager, C.; Kaplan, S. (1980) Doxorubicin cardiotoxicity in children. J Pediatr 1980, 97, 45–50. [Google Scholar] [CrossRef]
- Bell, J.R.; Bernasochi, G.B.; Varma, U.; Raaijmakers, A.J.; Delbridge, L.M. Sex and sex hormones in cardiac stress—mechanistic insights. J Steroid Biochem Mol Biol 2012, 137, 124–135. [Google Scholar] [CrossRef]
- Zhang, J.; Knapton, A.; Lipshultz, S.E.; Cochran, T.R.; Hiraragi, H.; Herman, E.H. Sex-related differences in mast cell activity and doxorubicin toxicity: a study in spontaneously hypertensive rats. Toxicol Pathol 2014, 42, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Belham, M.; Kruger, A.; Mepham, S.; Faganello, G.; Pritchard, C. Monitoring left ventricular function in adults receiving anthracycline-containing chemotherapy. Eur J Heart Fail 2007, 9, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.Y.T.F.; Tsai, C.H.; Huang, C.Y. (2011) Mechanisms governing the protective effect of 17B-estradiol and estrogen receptors against cardiomyocyte injury. BioMedicine 2011, 1, 21–28. [Google Scholar] [CrossRef]
- International Commission on Radiological Protection (ICRP), Assessing Dose of the Representative Person for the Purpose of Radiation Protection of the Public and the Optimisation of Radiological Protection: Broadening the Process. ICRP Publication 101. Ann. ICRP 36.
- International Commission on Radiological Protection (ICRP), Adult Reference Computational Phantoms. ICRP Publication 110. Ann. ICRP 39 2009.
- Langen, B.; Vorontsov, E.; Spetz, J.; Swanpalmer, J.; Sihlbom, C.; Helou, K.; Forssell-Aronsson, E. Age and sex effects across the blood proteome after ionizing radiation exposure can bias biomarker screening and risk assessment. Sci Rep. 2022, 12, 7000. [Google Scholar] [CrossRef]
- Yahyapour, R.; Motevaseli, E.; Rezaeyan, A.; Abdollahi, H.; Farhood, B.; Cheki, M.; Najafi, M.; Villa, V. Mechanisms of Radiation Bystander and Non-Targeted Effects: Implications to Radiation Carcinogenesis and Radiotherapy. Curr Radiopharm. 2018, 11, 34–45. [Google Scholar] [CrossRef]
- Guizard, A.; Boutou, O.; Pottier, D.; Troussard, X.; Pheby, D.; Launoy, G.; Slama, R.; Spira, A. The incidence of childhood leukaemia around the La Hague nuclear waste reprocessing plant (France): a survey for the years 1978-1998. J. Epidemiol. Comm. health,.
- Yoshida, K.; Nemoto, K.; Nishimura, M.; Seki, M. Exacerbating factors of radiation-induced myeloid leukemogenesis. Leukemia Res,.
- Kovalchuk, O.; Burke, P.; Besplug, J.; Slovack, M.; Filkowski, J.; Pogribny, I. Methylation changes in muscle and liver tissues of male and female mice exposed to acute and chronic low-dose Xray-irradiation. Mutat. Res.
- Koturbash, I.; Zemp, F.; Kolb, B.; Kovalchuk, O. Sex-specific radiation-induced microRNAome responses in the hippocampus, cerebellum and frontal cortex in a mouse model. Mutat. Res,.
- Koturbash, I.; Kutanzi, K.; Hendrickson, K.; Rodriguez-Juarez, R.; Kogosov, D.; Kovalchuk, O. Radiation-induced bystander effects in vivo are sex specific. Mutat. Res,.
- Koturbash, I.; Zemp, F.J.; Kutanzi, K.; Luzhna, L.; Loree, J.; Kolb, B.; Kovalchuk, O. Sex-specific microRNAome deregulation in the shielded bystander spleen of cranially exposed mice. Cell Cycle, 1658; 7. [Google Scholar]
- Chai, Y.; Calaf, G.M.; Zhou, H.; Ghandhi, S.A.; Elliston, C.D.; Wen, G.; Nohmi, T.; Amundson, S.A.; Hei, T.K. Radiation induced COX-2 expression and mutagenesis at non-targeted lung tissues of gpt delta transgenic mice. Br. J. Cancer,.
- Cheng, C.; Omura Minamisawa, M.; Kang, Y.; Hara, T.; Koike, I.; Inoue, T. Quantification of circulating cellfree DNA in the plasma of cancer patients during radiation therapy. Cancer Sci. 2009, 100, 303–309. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, L.; Rong, S.; Qu, H.; Zhang, Y.; Chang, D.; Pan, H.; Wang, W. Relation between gastric cancer and protein oxidation, DNA damage, and lipid peroxidation. Oxid. Med. Cell Longev. 2013, 2013. [Google Scholar] [CrossRef]
- Deligezer, U.; Eralp, Y.; Akisik, E.Z.; Akisik, E.E.; Saip, P.; Topuz, E.; Dalay, N. Effect of adjuvant chemotherapy on integrity of free serum DNA in patients with breast cancer. Ann. N. Y. Acad Sci. 2008, 1137, 175–179. [Google Scholar] [CrossRef]
- He, K.; Zhang, S.; Shao, L.L.; Yin, J.C.; Wu, X.; Shao, Y.W.; Yuan, S.; Yu, J. Developing more sensitive genomic approaches to detect radioresponse in precision radiation oncology: From tissue DNA analysis to circulating tumor DNA. Cancer Lett. 2020, 472, 108–118. [Google Scholar] [CrossRef]
- Allegra, A.G.; Mannino, F.; Innao, V.; Musolino, C.; Allegra, A. Radioprotective Agents and Enhancers Factors. Preventive and Therapeutic Strategies for Oxidative Induced Radiotherapy Damages in Hematological Malignancies. Antioxidants (Basel). 2020, 9, 1116. [Google Scholar] [CrossRef] [PubMed]
- Allegra, A.; Tonacci, A.; Pioggia, G.; Musolino, C.; Gangemi, S. Anticancer Activity of Rosmarinus officinalis L.: Mechanisms of Action and Therapeutic Potentials. Nutrients. 2020, 12, 1739. [Google Scholar] [CrossRef] [PubMed]
- Allegra, A.; Speciale, A.; Molonia, M.S.; Guglielmo, L.; Musolino, C.; Ferlazzo, G.; Costa, G.; Saija, A.; Cimino, F. Curcumin ameliorates the in vitro efficacy of carfilzomib in human multiple myeloma U266 cells targeting p53 and NF-κB pathways. Toxicol In Vitro. 2018, 47, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Cristani, M.; Speciale, A.; Saija, A.; Gangemi, S.; Minciullo, P.L.; Cimino, F. Circulating Advanced Oxidation Protein Products as Oxidative Stress Biomarkers and Progression Mediators in Pathological Conditions Related to Inflammation and Immune Dysregulation. Curr Med Chem. 2016, 23, 3862–3882. [Google Scholar] [CrossRef]
- Kim, S.Y. Oxidative stress and gender disparity in cancer. Free Radic Res. 2022, 56, 90–105. [Google Scholar] [CrossRef]
- May, R.C. Gender, immunity and the regulation of longevity. Bioessays 2007, 29, 795–802. [Google Scholar] [CrossRef]
- Malorni, W.; Straface, E.; Matarrese, P.; Ascione, B.; Coinu, R.; Canu, S.; Galluzzo, P.; Marino, M.; Franconi, F. Redox state and gender differences in vascular smooth muscle cells. FEBS Lett. 2008, 582, 635–642. [Google Scholar] [CrossRef]
- Vina, J.; Sastre, J.; Pallardo, F.; Borras, C. Mitochondrial theory of aging: importance to explain why females live longer than males. Antioxid. Redox Signal. 2003, 5, 549–556. [Google Scholar] [CrossRef]
- Loft, S.; Vistisen, K.; Ewertz, M.; Tjonneland, A.; Overvad, K.; Poulsen, H.E. Oxidative DNA damage estimated by 8-hydroxydeoxyguanosine excretion in humans: influence of smoking, gender and body mass index. Carcinogenesis 1992, 13, 2241–2247. [Google Scholar] [CrossRef]
- Marnett, L.J. Oxyradicals and DNA damage. Carcinogenesis 2000, 21, 361–370. [Google Scholar] [CrossRef]
- Bokov, A.F.; Ko, D.; Richardson, A. The effect of gonadectomy and estradiol on sensitivity to oxidative stress. Endocr. Res. 2009, 34, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Valencia, C.A.; Zhang, J.; Lee, N.C.; Slone, J.; Gui, B.; Wang, X.; Li, Z.; Dell, S.; Brown, J.; Chen, S.M.; Chien, Y.H.; Hwu, W.L.; Fan, P.C.; Wong, L.J.; Atwal, P.S.; Huang, T. Biparental inheritance of mitochondrial DNA in humans. Proc Natl Acad Sci USA 2018, 115, 13039–13044, 2018. [Google Scholar] [CrossRef] [PubMed]
- Rius, R.; Cowley, M.J.; Riley, L.; Puttick, C.; Thorburn, D.R.; Christodoulou, J. Biparental inheritance of mitochondrial DNA in humans is not a common phenomenon. Genet Med 2019, 21, 2823–2826. [Google Scholar] [CrossRef] [PubMed]
- Lutz-Bonengel, S.; Parson, W. No further evidence for paternal leakage of mitochondrial DNA in humans yet. Proc Natl Acad Sci USA 2019, 116, 1821–1822. [Google Scholar] [CrossRef]
- Gaignard, P.; Liere, P.; Thérond, P.; Schumacher, M.; Slama, A.; Guennoun, R. Role of sex hormones on brain mitochondrial function, with special reference to aging and neurodegenerative diseases. Front Aging Neurosci 2017, 9, 406, 2017. [Google Scholar] [CrossRef]
- Ventura-Clapier, R.; Moulin, M.; Piquereau, J.; Lemaire, C.; Mericskay, M.; Veksler, V.; Garnier, A. Mitochondria: a central target for sex differences in pathologies. Clin Sci (Lond) 2017, 131, 803–822. [Google Scholar] [CrossRef]
Figure 1.
Cancer cells stimulate the activation of macrophages that infiltrate neoplastic tissue, with the consequent production of ROS, RNS and cytokines production and, in turn, oxidative stress. Oxidative stress, as in a cycle, promotes cancerogenesis.
Figure 1.
Cancer cells stimulate the activation of macrophages that infiltrate neoplastic tissue, with the consequent production of ROS, RNS and cytokines production and, in turn, oxidative stress. Oxidative stress, as in a cycle, promotes cancerogenesis.
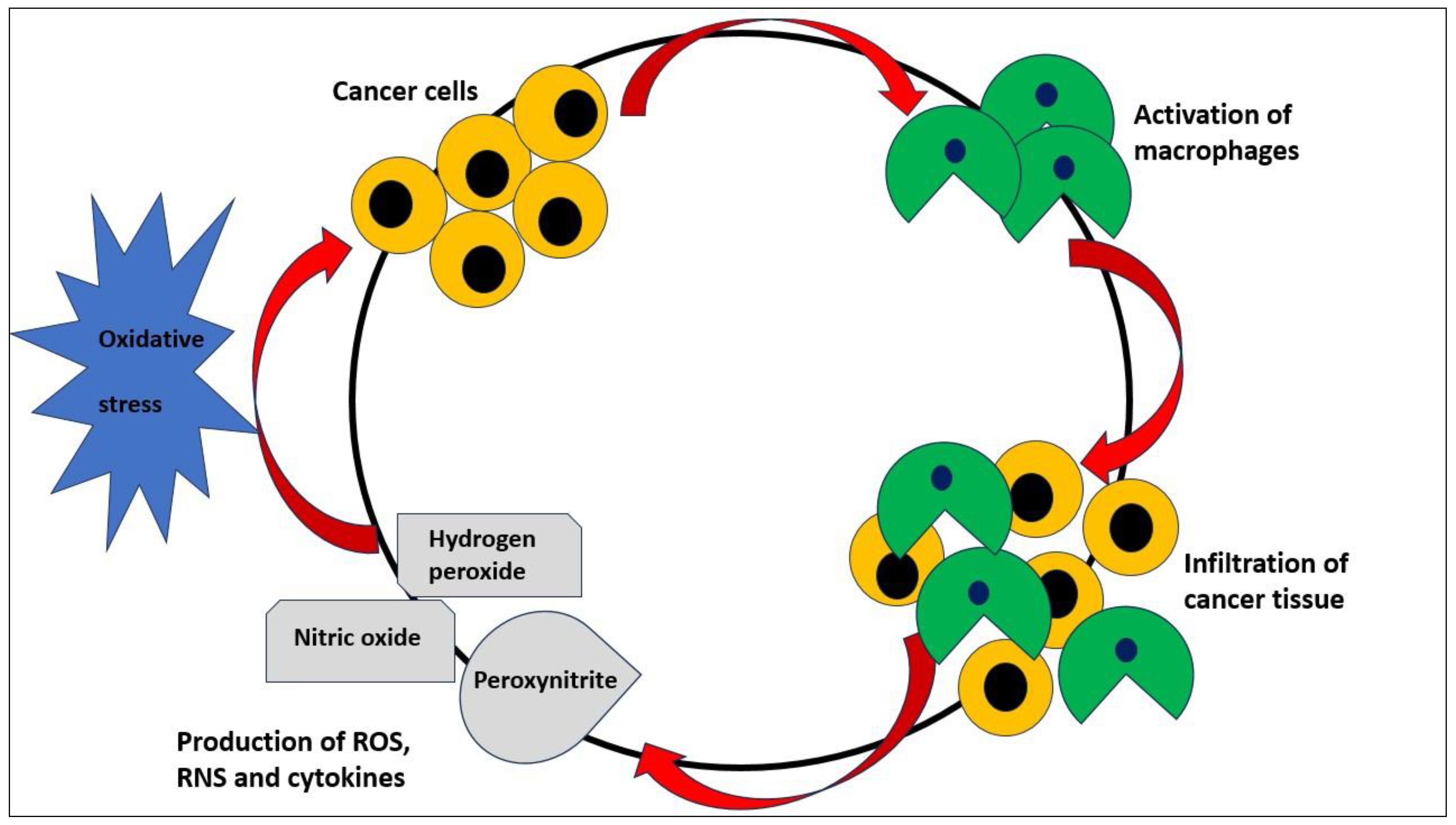

Table 1.
Different risk rates of cancer according to oxidative stress mechanisms in males and females.
Table 1.
Different risk rates of cancer according to oxidative stress mechanisms in males and females.
| Higher risk | Mechanism | References | |
|---|---|---|---|
| Glioma | men | Testosterone has neurotoxic effects | [97] |
| Liver cancer | men | Low levels of alcohol-dehydrogenase | [111] |
| Colorectal cancer | women | Low levels of unconjugated bilirubin which has antioxidant effects | [163] |
| Lung cancer | smokimg women | High levels of CYP1A1 and CYP1B1 that activate tobacco smoke components to create ROS | [184] |
| Melanoma | men | Low levels of antioxidant enzymes in the skin | [215] |
| Non-Hodgkin Lymphoma | men | High levels of superoxide in thymocytes overexpressing Bax | [223] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



