
Submitted:
08 May 2023
Posted:
10 May 2023
You are already at the latest version
Abstract
The genetic systems of Paspalum species have not been extensively studied. We analysed the ploidy, reproductive mode, mating system, and fertility of four Paspalum species - P. durifolium, P. ionan-thum, P. regnellii, and P. urvillei. An analysis of 378 individuals from 20 populations of north-eastern Argentina was conducted. All populations of the four Paspalum species were pure tetraploid and had a sexual and stable reproductive mode. However, some populations of P. durifolium and P. ionanthum showed low levels of apospory. Populations of P. durifolium and P. ionanthum had low seed sets under self-pollination but were fertile under open pollination, showing that self-incompatibility likely caused self-sterility. In contrast, populations of P. regnellii or P. urvillei showed no evidence of apospory and seed set in both self- and open pollination conditions were high, suggesting that they are self-compatible, due to the absence of pollen-pistil molecular in-compatibility mechanisms. The evolutionary origin of the four Paspalum species could explain these differences. This study supplies valuable insights into the genetic systems of Paspalum species, which could have implications for their conservation and management.
Keywords:
apospory
; cytogeography
; fertility
; mating systems
; polyploidy
; reproductive behaviour
; self-incompatibility
1. Introduction
Polyploidy is a widespread phenomenon among species that entails a considerable influence on the evolution of plants, animals, and fungi [1]. Between 30 and 80 % of angiosperms are polyploids [2]. Polyploidy acts as a primary mechanism for plant adaptation and speciation [3,4,5,6] by promoting variability through changes in chromosome numbers [7], increased allelic diversity in allopolyploids [8], and genomic restructuring [9]. In most cases, polyploidy poses an almost instantaneous and effective barrier to gene flow among cytotypes [10] allowing the coexistence of different intraspecific cytotypes [11]. Additionally, polyploid cytotypes generally differ from their diploid ancestors in morphological, ecological, physiological, reproductive, and cytological characteristics [7,12,13,14], contributing to the occupation of new ecological niches and the reinforcement of reproductive isolation mechanisms.
Paspalum L. is a large genus of Panicoid grasses, including around 310-350 species occurring mainly in the Americas [15,16]. The genus is distributed throughout tropical, subtropical, and temperate regions [17] with distinct species occupying flooded, dry, saline, or sandy soils of savannas, coastal dunes, tropical or temperate forests, and prairies [18]. The center of origin and diversification of the genus is tropical South America, with biodiversity hotspots in the Brazilian Cerrado and the Campos of Argentina, Uruguay, and southern Brazil [15,17]. The most common basic chromosome number in Paspalum is x = 10. However, a few exceptions to this number have been reported, such as species with x = 6 [16,19], x = 9 [20], and x = 16 [16].
Paspalum species show a broad range of genetic, morphological, and ecological diversity. Ortiz et al. [21] classified 72 Paspalum species into eight diverse groups based on their genetic system, which includes combinations of basic chromosome number, ploidy, reproductive modes, and mating systems. Approximately 75% of these species belong to groups that have polyploid cytotypes [21]. The most significant group includes species with sexual diploid cytotypes that are allogamous by self-incompatibility and apomictic tetraploid cytotypes from autopolyploid origin. Two other important groups lack diploid cytotypes and have an allopolyploid origin. One group is characterized by the exclusive occurrence of sexual polyploids (either 4x or 6x), and the other group has sexual tetraploids together with higher polyploid apomictic cytotypes [21].
Apomixis, the formation of asexual seeds, has been considered a “blind alley” or an evolutionary “dead end” [3,22], as apomictic plants lack mechanisms that exploit genetic variation during offspring formation, increasing developmental instability of the sexual pathway and reducing the survival ability of sexual progenies [14,23,24,25]. Nevertheless, recent population studies have proved considerable amounts of genetic diversity in natural asexual populations [26]. Similarly, genetic variability studies in several natural populations of apomictic species of Paspalum showed elevated levels of genotypic variation, which have been attributed to backcrossing with sexual parental relatives, facultative sexuality, mutations, and recurrent origin of new polyploids from divergent sexual ancestors [26,27,28,29,30,31].
Sexuality and hybridization have been significant factors in the origin and evolution of several Paspalum species. About 15% of Paspalum species are sexual tetraploids, all of which have an allopolyploid origin [21,32]. However, there is still a need for comprehensive analyses of genetic systems, cytotype composition at the population level, and reproductive variability. Four species, belonging to distinct infrageneric groups being the main morphological variants within the genus [17], are particularly relevant in this regard.
Paspalum durifolium Mez and P. ionanthum Chase are two species with multiple polyploid cytotypes that lack diploids [21]. However, the available records of ploidy and mode of reproduction are limited to single individuals [33,34,35,36,37,38]. Additionally, earlier studies of reproductive analysis relied on cytoembryology, and no studies of functional reproductive pathways using flow cytometric seed screening have been conducted.
Paspalum durifolium is a tall, robust bunchgrass with coarse leaves that grows in lowlands near rivers and streams, and sometimes in grasslands in northern Uruguay, southern Brazil, northern Argentina, and Paraguay [17,33]. It has sexual tetraploid and apomictic hexaploid cytotypes with an allopolyploid origin [33,37,38]. A rare pentaploid plant reproducing by facultative apomixis in a tetraploid population [35] suggests cytotype variability within populations exists and points to the occurrence of dynamic population-level processes.
Paspalum ionanthum is a warm-season perennial bunchgrass that grows in sandy and flooded soils in savannas of central Paraguay, Brazil, Argentina, and Uruguay [17]. This species has self-sterile sexual allotetraploid and apomictic octoploid cytotypes [34,37,39,40,41].
Paspalum regnellii Mez and P. urvillei Steud. are species in which only sexual allotetraploid individuals have been found [24,42]. Paspalum regnellii is found in southern and south-eastern Brazil, eastern Paraguay, and north-eastern Argentina, while P. urvillei is a warm-season perennial bunchgrass native to southern Brazil and northern Argentina, but it also grows from southern USA to Argentina [17]. Both species are commonly found in modified soils on the edges of forests and roads [17].
The knowledge of the geographic occurrences of cytotypes in different Paspalum species is yet fragmentary. However, in almost all cases, this information only relates to the ploidy level of a single isolated individual or germplasm collections e.g., [37,40,43,44,45,46]. Although there have been a few extensive studies on cytotype diversity and distribution, they mainly focus on apomictic species (i.e., for P. intermedium Munro ex Morong & Britton [13], for P. alcalinum Mez, P. denticulatum Trin., P. lividum Trin., P. nicorae Parodi and P. rufum Nees [29], and for P. simplex Morong [30,47]). As a result, there is a lack of information about reproductive variability and cytotype associations at local and regional scales, which is crucial for understanding the origin, dynamics, and adaptation of cytotypes in natural populations.
To address this gap, the present study aims (i) to sample a representative number of individuals from different natural populations of Paspalum durifolium, P. ionanthum, P. regnellii and P. urvillei, (ii) to analyses their cytotype composition, (iii) to assess the reproductive mode at two developmental stages (i.e. ovules and seeds) under common environmental conditions, and (iv) to determine the mating system and fertility for each species. This information will shed light on the role of different genetic systems in the establishment of new cytotypes and their stability in the evolution of Paspalum agamic complexes.
2. Materials and Methods
2.1. Plant collections
Plant materials were collected from the north-eastern region of Argentina, which lies between the parallels of 26°24.117S - 31°01.223S and parallels 54°14.845W - 59°25.005W (Table 1). Populations of P. durifolium and P. ionanthum were collected near the Iberá wetland macrosystem (Corrientes, Argentina). Populations of P. regnellii were collected in Misiones (Argentina), while the populations of P. urvillei were collected in Corrientes, Chaco, Entre Ríos, Misiones, and Santa Fe (Argentina).
Sampling was conducted during two different periods - November-December for P. durifolium and December-February for P. ionanthum, P. regnellii, and P. urvillei. The sampling sites were chosen to ensure maximum representation of the distribution range of the species in the study region, considering macro-scale (among populations) and micro-scale (within populations) trends. The populations were at least 50 km apart from each other and exceeded a linear size of 200 m. Uniform sampling was done to ensure an even representation of individuals within each population (i.e., the distance between two consecutive individuals was kept constant around 10 m). Five populations and ca. 20 plants per population were collected for each species.
Small cuttings with short rhizomes in the base were collected from each plant and cultivated in pots in a greenhouse. Once rooted, the plants were transferred to an experimental field near the city of Corrientes, Argentina. Herbarium vouchers were collected for each sampled population and deposited at the CTES and MNES Herbaria. The distribution of the ploidy levels of the analyzed populations were plotted on maps using Diva-GIS [48]. Field GPS was used to decide the collection sites.
2.2. Ploidy level estimation
To determine the ploidy level of each sample, flow cytometry (FC) was used to estimate the relative nuclear DNA content in comparison with a plant with a known ploidy level of 2n = 4x = 40. The plants used for comparison by FC were P. durifolium Q3947 [38], P. ionanthum Q3726 [41], P. regnellii V10121 and P. urvillei Q4111 [49]. The ploidy level of each plant was determined using fresh leaf tissue samples following the instructions described in Brugnoli et al. [47]. The fluorescence intensity of 4’,6-diamidino-2-phenylindole (DAPI)-stained nuclei was analyzed with a Partec PA II flow cytometer (Partec GmbH, Münster, Germany) using the CyStain UV Precise P kit from Partec. The measurements of relative fluorescence intensity of stained nuclei were performed on a linear scale using PA II’s Partec FloMax software, with at least 3,000 nuclei per sample and CV ≤8 %. The ploidy level of each sample was estimated from the histograms as a ratio between the mean value of the DNA peaks of the sample and the specific standard used.
For chromosome counting, a random sample of root tips was collected from potted plants, placed in a saturated solution of α-bromonaphthalene for 2 hours, hydrolyzed in 1.0 mol m-3 hydrochloric acid for 10 min. at 60 °C without previous fixation, and stained with Feulgen’s reagent. The root tips were macerated in a drop of 2% aceto-orcein stain, squashed under a coverslip, and observed using light transmission microscopy.
2.3. Reproductive mode
To assess the reproductive pathways at two developmental stages of the life cycle in populations of four Paspalum species, two analytical methods were employed. The first method involved cyto-embryological analysis of mature ovules, while the second employed flow cytometric seed screening (FCSS).
Cytoembryological analysis by mature embryo sac observation was performed on a random sample of five plants from each population. Spikelets at anthesis were fixed in FAA (70 % ethanol, 37 % formaldehyde, and glacial acetic acid in the ratio 18:1:1). Pistils were dissected out and clarified following the method described by Young et al. [50] with the modifications carried out by Zilli et al. [51]. Between 30 to 40 pistils were observed per plant, using a Leica microsystems Gmbh (Germany) microscope with a Nomarski differential interference contrast device. Embryo sacs were photographed with a Leica EC3 camera. Based on the structure and cell composition of the embryo sacs, four types of ovules were defined: (i) ovules carrying one meiotic embryo sac (MES) composed of an egg cell, two synergid cells, a central cell with two (rarely one or three) nuclei, and several antipodals, (ii) ovules carrying one or more aposporous embryo sacs (AES) with an egg cell, one or two synergid cells, a central cell with two (rarely one or three) nuclei, and no antipodals, (iii) ovules with mixed embryo sacs having one MES plus one (or more) AES, and (iv) ovules without embryo sacs (AbES) whereby embryo sacs were not observed or had clear signs of abortion. Plants bearing only ovules with one meiotic embryo sac (MES) were classified as sexual, while those with both ovules with MES and one or more aposporous embryo sacs (AES) were classified as facultative sexual.
The FCSS method was used to reconstruct the reproductive pathways of fully developed caryopses (seeds). Seeds from open-pollinated flowers of five plants per population were analyzed using FCSS according to protocol described by Matzk et al. [52] and following instructions described by Siena et al. [53]. Seeds were harvested from the same plants employed for cyto-embryology. Thirty seeds per individual were analyzed in bulks of five seeds each. The fluorescence intensity of 4’,6-diamidino-2-phenylindole (DAPI)-stained nuclei was determined using a Partec PA II Flow Cytometer (Partec GmbH, Münster, Germany) with the detector running at 355 nm. Ploidy levels of the endosperm and embryo tissues were estimated by comparing the different peak configurations. A flow cytometry histogram with two main peaks equivalent to values of 2C and 3C would indicate that the seed was formed sexually (embryo = n + n; endosperm = n + n + n, i.e., meiosis + syngamy). Alternatively, a histogram showing two main peaks corresponding to 2C and 5C values would reveal an apomictic origin of the seed (embryo = 2n + 0; endosperm = 2n + 2n + n, i.e., apospory + parthenogenesis + pseudogamy). Approximately 3,000 nuclei were measured per sample, and data analysis was performed using PA II’s Partec FloMax software. The mean values of relative DNA content (C-values) for embryo and endosperm tissues of single seeds were set up to infer the sexual or apomictic origin of each seed.
2.4. Reproductive pathway efficiency
The efficiency of the reproductive pathways (sexual and apomictic) was evaluated by calculating the ratio between the expected and observed frequencies of ovules undergoing the meiotic or apomictic pathway at each stage, as described by Hojsgaard et al. [23] and Reutemann et al. [62]. At the ovule stage, the proportion of embryo sacs was estimated by nm/nt for the meiotic pathway and na/nt for the apomictic pathway, where nm is the total number of ovules with a meiotic embryo sac (MES), na is the total number of ovules with apomictic embryo sacs (AES), and nt is the total number of embryo sacs. The values of nm and na include the number of observed ovules with mixed embryo sacs (MES + AES) since both reproductive pathways were expressed. At the seed stage, the observed proportions of each reproductive pathway were calculated based on the number of sexual and apomictic seeds. The expected proportions of each pathway at the seed stage were determined by using the observed proportions of the two reproductive pathways at the ovule stage.
To evaluate the differences between the observed proportions of both pathways in each stage, a paired t-test was conducted on the mean difference. Additionally, a standard Pearson’s Chi-squared test was performed to find significant differences between the expected and observed proportions of each reproductive pathway.
2.5. Mating system and seed fertility
The mating system and seed fertility were examined by analyzing the seed set during two consecutive flowering periods (2015-2016 and 2016-2017) under conditions of forced self-pollination and open pollination. Random samples of five plants per population were analyzed to determine the mating system. The degree of autogamy and the breaking of self-incompatibility were estimated by analyzing the proportions of seed set in self-pollination. To ensure self-pollination, three inflorescences per individual plant were bagged before anthesis. The degree of allogamy was estimated by comparing the seed set under open pollination and self-pollination. Additionally, three inflorescences per individual were bagged after anthesis, and spikelets with a caryopsis were separated from empty spikelets using a seed blower. After approximately one month, inflorescences were manually threshed, and seed set was calculated using the formula: Seed set = (No. of seeds / total No. of spikelets) x 100 for each pollination condition and plant.
2.6. Statistical analysis
The seed set data was analyzed using the Info-Gen software [55] in a completely randomized design where each plant stood for a replication in the population. Statistical comparisons for seed set were performed in the following sequence: (i) comparisons within populations between the two flowering periods (1st vs. 2nd) for each pollination method (self- and open pollination), (ii) comparisons among populations for each pollination method (self- and open pollination) and flowering period (1st and 2nd), (iii) comparisons within populations between the pollination methods (self- vs. open pollination), based on the overall mean values of both flowering periods, and (iv) comparisons among populations for each pollination method (self- and open pollination), based on the overall mean values of both flowering periods. Mean values, coefficients of variation, and ANOVA were computed. Tukey's test was employed ad hoc when ANOVA showed significant differences in the mean comparisons. Unless otherwise specified, all differences were considered significant at p < 0.05.
3. Results
3.1. Ploidy estimations
All individual from populations of P. durifolium resulted tetraploids (2n = 4x = 40). Ninety-two individuals analyzed showed 2C DNA content like the tetraploid standard of P. durifolium (Figure 1a). No hexaploid or pentaploid individuals were detected in the populations of this species. Similarly, all 96 plants from five populations of P. ionanthum were tetraploids (2n = 4x = 40) (Figure 1b), and no expected octoploid plants or plants with other ploidy levels were detected. The populations of P. regnellii and P. urvillei also showed tetraploid individuals, with all 96 plants from P. regnellii and 94 plants of P. urvillei being tetraploid (2n = 4x = 40) (Figure 1c,d). These results were further confirmed by chromosome counting in root tip meristems for a random sample of four plants from each population (data not shown).
3.2. Embryo sac analysis
Embryo sac analysis was conducted on one hundred plants from all populations of the four species under study. The findings, presented in Table 2, summarized the types of embryo sacs observed in each species and population. Among the populations of P. durifolium, a high proportion of ovules with well-developed embryo sacs were observed (Table 2 and Table S1). Twenty-five evaluated plants from five P. durifolium populations showed ovules mainly with MES, ranging between 79.5 and 94% (Table 2). These meiotic sacs are of Polygonum type, consisting of an egg cell, two synergids, a central cell with two polar nuclei, and three antipodal cells, which later divide into several antipodal daughters (Figure 2a). The highest percentage of ovules with meiotic embryo sacs (MESs) was observed in population PD2, and no AES or MES + AES were observed (Table 2 and Table S1). However, in the remaining four populations, some plants showed ovules with MES next to an AES carrying an egg cell, one or two synergids, and a large binucleate cell, lacking antipodal cells (Table 2, Figure 2e). The population PD4 had the highest mean percentage (6.5%) of ovules with MES + AES (Table 2). Populations PD3 and PD4 had at least one plant showing near 20% of MES + AES ovules type (Table S1). Population PD5 had the highest proportion of individuals with ovules carrying MES + AES (3 out of 5; Table S1) and a mean percentage of ovules with mixed embryo sacs of 3.2% (Table 2). No ovule carrying only AESs was observed. AbESs were also present in all populations, ranging between 6 and 17.3% (Table 2). Five populations showed significant differences with meiotic pathway dominating at the ovule stage (p < 0.001, Table 2).
In P. ionanthum, the analysis of embryo sacs among 25 individuals from all populations revealed a high percentage of ovules with viable embryo sacs (Table 2 and Table S1), with most individuals carrying one MES in a high proportion of ovules (Figure 2b). Populations PI1 and PI2 showed 2.5 and 1.3% of ovules with MES + AES, respectively (Table 2). Such ovules were observed in two individuals from PI1 and one from PI2. Ovules with a single AES were not observed (Table S1). Significant differences between both reproductive pathways were observed in the five populations (p < 0.001, Table 2), with meiotic pathway dominating at the ovule stage.
Populations of P. regnellii showed a high proportion of ovules with well-developed embryo sacs (Table 2 and Table S1), with ovules with MES (Figure 2c) ranging between 70.0 and 95.5% in all populations (Table 2). The population PR3 showed the highest percentage of ovules with MES, while PR1 showed the lowest percentage due to a high proportion of ovules having under-developed or AbESs (Table 2). No aposporous embryo sacs were observed in any of the twenty-five analyzed plants (Table 2 and Table S1). Five populations showed significant differences, with meiotic pathway as the only one expressed at the ovule stage (p < 0.001, Table 2).
Populations of P. urvillei showed the highest percentage of ovules with MES (Figure 2d), ranging between 95.8 and 100% among populations (Table 2), with all evaluated plants from population PU5 and four out of five individuals from population PU3 showing ovules with 100% MESs (Table S1). No AES were detected in the ovules of the 25 analyzed plants of P. urvillei, and the average percentage of ovules without embryo sacs was 4.3% in PU1 (Table 2). Meiotic pathway was the only one expressed at the ovule stage (p < 0.001, Table 2).
3.3. Flow cytometric seed screen
Seeds from each population of the four species were analyzed by Flow Cytometric Seed Screen (FCSS). Specifically, 150 seeds from five plants of each population were assessed (Table 2), and the FCSS analysis revealed a single type of histogram peak for all individuals in each population. This peak corresponded to the embryo: endosperm ratios of relative DNA content 2C: 3C (Figure 3). Furthermore, histograms from cell nuclei of bulked seed samples of P. durifolium were examined, and four distinct peaks were observed (Figure 3). The two highest peaks corresponded to stages G1 (4C) and G2 (8C) of tetraploid embryos, while the peaks at 150 and 300 of relative DNA content corresponded to stages G1 (6C) and G2 (12C) of the endosperms. Additionally, two smaller peaks were observed at 400 and 600 of relative DNA content, which could indicate stages G2 of an embryo (16C) and endosperm (24C) from an hexaploid seed, respectively. The peaks between 100 and 300 of relative DNA content suggested the presence of sexual reproduction, including a reductional division (meiosis) and double fertilization. However, the peaks at 400 and 600 showed double fertilization from AES (3C:5C), rather than typical sexual reproduction. Notably, the study did not observe any instances of seed originating from apospory + pseudogamy (Table 2).
3.4. Reproductive variability in ovules and seeds
The proportions of sexuality and apomixis during the reproductive stages displayed distinct patterns in P. durifolium populations. Populations PD1, PD3, PD4, and PD5 showed a low proportion of the apomictic pathway during the ovule stage, but all populations of P. durifolium produced 100% sexual seeds during the seed stage (Table 3). Furthermore, there were no significant differences between the observed and expected proportions of sexually originated seeds (p > 0.05, Table 3). The efficiency of the sexual pathway from the ovule to the seed stage ranged from 1.01 to 1.07, showing an increase in the sexual pathway (Table 3).
Similarly, in populations PI1 and PI2 of P. ionanthum, which showed a low proportion of the apomictic pathway during the ovule stage, only sexual seeds were produced (Table 3). In five populations of P. ionanthum, there was a high efficiency of the sexual pathway from the ovule to the seed stage, with values ranging from 1.0 to 1.03 (Table 3).
In P. regnellii and P. urvillei, only the sexual pathway was expressed during both reproductive stages. Both species showed a high degree of efficiency of the sexual reproductive pathway (Table 3).
3.5. Mating system and seed fertility
The intra-population seed set of P. durifolium under forced self-pollination conditions ranged from 0.4 to 0.9%, based on the overall mean values, while under open pollination conditions, it ranged from 28.8 to 51.5% (Table 4). Population PD3 exhibited the lowest and highest seed sets under self- and open pollination, respectively. The seed production under self-pollination was highly variable in both flowering periods, with a high and wide population coefficient of variation (CV). A lower CV was observed under open pollination (Table S2). There were no significant intra-population differences (p > 0.05) observed in either self- or open pollination when comparing the two flowering periods. Inter-population significant differences were only observed in open pollination (p < 0.006) when considering the overall mean values. The fertility was significantly higher under open pollination than in self-pollination (p < 0.001) (Table 4).
In P. ionanthum, the intra-population seed set under forced self-pollination conditions ranged from 0.6 to 2.8%, based on the overall mean values, while under open pollination, it ranged from 29.4 to 54.3% (Table 4). Population PI3 exhibited the highest values in both self- and open pollination, reaching up to 4.26% in self-pollination during the first flowering period (Table 4 and Table S2). The seed set in self-pollination was highly variable in both flowering periods. A lower CV was observed in open pollination (Table S2). However, regarding intra-population pollination, there were no significant differences between the two flowering periods for both pollination conditions, except for PI2 at open pollination (p < 0.002). As for inter-population pollination, significant differences were observed in open pollination during the second flowering period (p < 0.004) and when considering the overall mean values (p < 0.017). The fertility in open pollination was significantly higher than in self-pollination (p < 0.001) (Table 4).
In P. regnellii, the intra-population seed set under forced self-pollination conditions ranged from 6.4 to 18.8%, based on the overall mean values, while in open-pollination conditions, it ranged from 26.4 to 49.8% (Table 4). Population PR5 showed the highest values in both self- and open pollination. The seed set in self-pollination for the two flowering periods showed a high and wide CV, while in open pollination, it was lower (Table S2). Populations did not show significant intra-population differences between the two flowering periods for both pollination conditions, except for PR2 in self-pollination (p < 0.018). Inter-population significant differences were observed in open pollination during the first flowering period (p < 0.006). In turn, there were also significant differences among populations, both in self-pollination (p < 0.039) and open pollination (p < 0.026), when considering the overall mean values. The fertility in open pollination was significantly higher than in self-pollination (p < 0.001 to 0.031) (Table 4).
The seed set within populations of P. urvillei under forced self-pollination showed a range of 35.6 to 43.1%, based on the overall mean values. Conversely, in open pollination, the range was 70 to 80.6%, as shown in Table 4. Among the populations, PU5 and PU2 displayed the highest values in self- and open pollination, respectively. The coefficient of variation (CV) for seed set in self-pollination was higher than that in open pollination (Table S2). Notably, only population PU3 exhibited significant differences in open pollination between the two flowering periods (p = 0.007). When considering the overall mean values, there were no significant inter-population differences in either self- or open pollination during each flowering period. Fertility in open pollination was significantly higher than in self-pollination (p > 0.001 to 0.018) (Table 4).
4. Discussion
Polyploidy and apomixis are common in the Paspalum genus, with most species being tetraploid and having diploid sexual counterparts that are co-specific [21,32]. However, there is a subgroup of species within the genus that show a different genetic system, being both polyploid and sexual, and some of these also have apomictic co-specific counterparts of higher ploidy. Knowledge about the genetic system of these polyploid species has been limited until now, with most analyses being based on one or few individuals from separate collections. Population-level studies have been carried out only on apomictic species within the genus. However, a recent population analysis of four sexual polyploid species in Paspalum has allowed for a better understanding of their natural distribution and the variation in their genetic systems.
4.1. Single ploidies featured in all species
The ploidy level of the five P. durifolium natural populations were tetraploid (2n = 4x = 40) and uniform with respect to chromosome number and cytotype composition. Despite hexaploid and pentaploid individuals have been reported for the species by other authors [33,35,36,37], they were not detected in the present study. All populations were found in association with the Iberá wetland macrosystem, found in Corrientes, Argentina. Quarin [38] mentions that the tetraploid cytotype of P. durifolium is found in the central part of the Province of Corrientes, associated with the Iberá basin; while the hexaploids are found towards the eastern end of Corrientes, in the Uruguay River basin. Two tetraploid populations from this study (PD3 and PD4) were collected in Santo Tomé (Corrientes) near the Uruguay River basin, where hexaploids were previously reported by Quarin [38]. The hexaploid cytotype was also registered in Rio Grande do Sul, Brazil [36,37] and in Rivera, in the north of Uruguay [33]. We also did not find the pentaploid cytotype of P. durifolium in any of the wild populations evaluated, which could show the existence of hexaploids within the area. Natural pentaploid individuals have their origin in the hybridization between tetraploid and hexaploid cytotypes, and their persistence depends on the degree of sterility and the mode of reproduction, since both apomictic and sexual reproductive pathways are usually functional in this cytotype [35]. Our results show a wide distribution of the tetraploid cytotype of P. durifolium in its natural range at the northeast of Argentina, which may be associated with a higher chromosomic and reproductive stability.
All five populations of P. ionanthum turned out to be pure tetraploids (2n = 4x = 40) in agreement with all previous records of individual chromosome counts for the species [34,37,39,40,41,56]. Populations were found in low and humid lands, on sandy and flooding soils, which are typical environments for this species [17]. No octoploid cytotype was detected as contrary to earlier reports by Burson and Bennett [34] and Pozzobon et al. [37,40]. The octoploid cytotype of P. ionanthum may have originated through a polyploidization mechanism described for other polyploid species of Paspalum [32]. Our results show that the incidence of such mechanism in P. ionanthum and the generation of elevated level polyploids is not frequent, being the tetraploid cytotype the only one -currently- occurring in Argentina.
Ploidy analyses conducted on five populations of P. regnellii have confirmed that only the tetraploid cytotype exists in the species' natural geographic range in Argentina. Previous records of individual plants from this species have also confirmed their tetraploid nature [24,37,43,57]. All populations were found in similar environments on modified red clay soils at roadsides and edges of native or cultivated forests. The species natural distribution extends throughout the center and south of Brazil, the east and northeast of Paraguay, and the northeast of Argentina [17,24].
Populations of P. urvillei collected in northeast Argentina were exclusively found to be tetraploids, confirming earlier records that show the existence of only one cytotype [42,43,45,57,58,59,60,61]. A hexaploid reported by Nielsen [62] was represented by a plant collected in the city of Magnolia, Arkansas, United States. The age of this record and the lack of confirmation of its taxonomic identity cast doubts on the accuracy of such register. The species is native to South America, inhabiting Bolivia, Brazil, Chile, Paraguay, Argentina, and Uruguay, where it grows in low fields, estuaries, and on the edges of roads and railways, and is a common weed in crop fields [17].
4.2. Reproductive pathways and cytotype stability
The analysis of reproductive modes in Paspalum species allows for the evaluation of sexual and apomictic pathways and an understanding of the efficiency of each pathway at different developmental stages [23,62]. In our study, we measured the mode of reproduction in two stages of the plant's life cycle, the ovule, and seed, to expose variations in the expression levels of sexuality and/or apomixis at the individual and population levels. This information can help us to understand the cytotype dynamics in nature and predict possible ploidy compositions in different geographical areas.
Populations of P. durifolium showed differences in reproductive pathways at the ovule stage, particularly during the observation of the mature female gametophyte. The meiotic pathway was exclusive in population PD2, with 94% of mature meiotic embryo sacs (MESs) being fully developed. The apomictic pathway was not expressed during ovule development in this population, as no aposporous embryo sacs (AESs) were observed among the 150 ovules analyzed. Conversely, in the remaining four populations, both reproductive pathways (sexual and apomictic) were observed with distinct levels of expression. In all of them, the meiotic pathway was expressed in a higher proportion than the apomictic one. The apomictic pathway was expressed in only 2.9% of the total ovules analyzed and was always accompanied by the expression of the sexual pathway in the same ovule. All ovules with MES + AES showed a greater development of the meiotic sac than the aposporous sac. This entails a competitive advantage favouring seed development from the MES over the AESs in mixed ovules. Quarin [39] saw a similar reproductive behaviour in some tetraploid accessions of P. durifolium. Out of 79 mature ovules analyzed, 63 had a normal MES, seven had one MES plus one AES, one ovule showed two AESs, and eight had undeveloped embryo sacs. The functionality of these AESs has never been tested in P. durifolium. There is no evidence that the pentaploid or hexaploid cytotypes originated from these AESs for polyploidization. Cytological results proved that the apomictic 6x cytotype of P. durifolium was generated by allopolyploidy and it has three different genomes [33]. Moreover, the pentaploid cytotype found into a tetraploid population probably had its origin via fertilization of a reduced female gamete from a tetraploid plant (n = 20) with a reduced male gamete from a hexaploid progenitor (n = 30) [35].
A comparable reproductive behavior was observed in the mature ovule stage of P. ionanthum populations. Sexuality was the only reproductive pathway expressed in three populations (PI3, PI4, and PI5), with a potential for sexuality close to 100% due to the extremely small number of ovules without embryo sac development. Two populations (PI1 and PI2) showed both reproductive pathways (sexual and apomictic), with a low potential for apomixis since some ovules were discovered carrying both MESs and AESs. All ovules with MES + AES showed a well-developed MES and a smaller undeveloped AES. The presence of MES + AES in tetraploid P. ionanthum was also observed by Burson and Bennett [34] and Martínez et al. [40]. Nevertheless, seeds produced by parthenogenesis (apomixis) or fertilization of unreduced female gametes from AESs have never been conclusively proved in P. ionanthum. The octoploid individual of P. ionanthum (syn. P. guaraniticum Parodi) was taxonomically accurately identified and, according to meiosis analysis, originated from autopolyploidy [34]. The chromosome duplication mechanism that led to this octoploid is still unknown; however, a possible origin through the fertilization of two unreduced gametes from tetraploid plants (2n = 4x = 40), where the female gametes could be from an AES, is well-supported.
Populations of P. regnellii and P. urvillei displayed similar behavior in both reproductive stages, showing exclusively the sexual pathway. At the ovule stage, most populations of P. regnellii demonstrated a high potential for sexuality, and no evidence for vestigial apospory was detected. Population PR1 exhibited a lower potential for sexuality (70%) due to a high proportion of ovules with AbESs. Previously, the megasporogenesis and megagametogenesis of P. regnellii were evaluated in one accession by Norrmann [24] who saw only MESs of the Polygonum type. All populations of P. urvillei had at the ovule stage, a potential for sexuality over 95% because most ovules showed a well-developed MES. Evidence for apospory was not detected in our observations of P. durifolium and P. ionanthum. In Paspalum, polyploids without apomixis are extremely rare. In P. regnellii, full expression of sexuality is associated with endemism, however, in P. urvillei it is linked with potential colonization of new suitable and hospitable habitats in disturbed ecosystems.
Gametophytic apomixis is a trait often linked to polyploidy, as individuals with the potential for apomixis in tetraploid populations can give rise to new hexaploid cytotypes. In Paspalum, this can occur through fertilization of unreduced gametes from an aposporous embryo sac of a tetraploid individual by a reduced male gamete of another tetraploid individual, as described by Quarin [32]. This polyploidization mechanism was also observed in mixed populations (2x and 4x) of P. rufum, where the unreduced female gametes from the AES of a diploid plant are fertilized by reduced male gametes of tetraploid plants to create new tetraploids [63].
While sporadic AESs going with MESs have been detected in several sexual diploid Paspalum species [37,53,63,64,65,66], the functionality of these AESs has only been proved in P. rufum [53,63]. In diploid genotypes of P. rufum, variations in the functionality of AESs and the ability for apomixis have been observed in self-pollination and interspecific interploidy crosses [63]. However, seed screen analyses performed for P. durifolium, P. ionanthum, P. regnellii and P. urvillei did not detect any seeds of apomictic origin, showing that these species reproduced solely through the sexual pathway. This could be due to the competition for resources or space within the ovule, temporal development, and even to a seasonal effect, as saw in other sexual species [64,67,68,69].
The use of flow cytometry seed screen analyses revealed that none of the populations of P. durifolium, P. ionanthum, P. regnellii, and P. urvillei produced seeds of apomictic origin. The analysis of the DNA content ratios of seed embryo and endosperm tissues showed that all seeds were generated through sexual reproduction. This finding is consistent with the results obtained by Galdeano et al. [49], who also used FCSS to study these four species and found that all seeds were produced through the sexual pathway. Although the tetraploid populations of P. durifolium and P. ionanthum possess the potential for apomixis at the ovule stage, this pathway is unexpressed at the seed stage, likely due to competition for resources or space within the ovule. This behavior has also been observed in other sexual species and was attributed to the competition for resources or space within the ovule, temporal development, and even to a seasonal effect [67,68,69]. Siena et al. [53] observed functional AESs in the diploid cytotype of P. rufum, where a small proportion of seeds developed from AESs. The genetic program necessary for the development of AESs appears to be present and expressed in the tetraploid populations studied here, but factors such as the functionality of the parthenogenesis gene(s), developmental competition between embryo sacs, and the genotype background may hinder the development of apomixis in these plants. Therefore, the reproductive behavior of these plants can only be considered as obligate sexual, and the possibilities for the development of apomixis remain elusive and complex.
4.4. Two mating systems with partial breakdown of self-incompatibility
Populations of P. durifolium showed an extremely low seed set under forced self-pollination conditions, with an overall average seed set value of 0.58%. On the other hand, seed set under open pollination was 40.8% and showed significant differences between populations. Quarin [39] observed in a single tetraploid plant of P. durifolium a seed set under self- and open pollination of 0.26 and 68.5%, respectively. The lower fertility via self-pollination was attributed to the existence of a pollen-pistil self-incompatibility system, mainly supported by a lack of penetration of the pollen tube into the style, and the observation of successful cross-pollination between different tetraploid genotypes [39].
Most sexual diploid Paspalum species show self-incompatibility, which is the most common mechanism of pollen-pistil interaction in self-sterile diploids such as P. fasciculatum Willd. ex Flüggé [70], P. notatum Flüggé [71], P. procurrens Quarin [72], and P. indecorum [69]. Self-incompatible diploids generally have con-specific self-compatible apomictic tetraploids [21,32,69]. The breakdown of the self-incompatibility system in these Paspalum species is likely due to polyploidization [32,73], as has also been described in other species (revised by [74]). This mating system, in which self-sterile sexual diploids and self-fertile apomictic polyploids coexist, is seen in several Paspalum species, including P. durifolium, but at the polyploid level. Sexual tetraploids of P. durifolium are self-incompatible that exhibit allogamy [39]; while hexaploids are self-compatible pseudogamous apomictic [33].
Fertility in P. ionanthum under forced self-pollination conditions was low, with an overall population mean of 1.4%, which is like that observed by Quarin and Norrmann [37] for a single plant. Two populations (PI3 and PI5) showed higher values (4.26 and 2.85%) during the first flowering period. Seed set in the first measured period was carried out during a warmer and drier summer than in the second period (data not shown). In this species, there is a clear partial breakdown of the self-incompatibility system, and the molecular basis for it could be due to environmental stressors such as elevated temperature during pollination [75,76,77,78] or internal factors, either genetic or cytological [76,79,80]. Wilkins and Thorogood [75] showed that self-incompatibility is broken in plants of perennial ryegrass (Lolium perenne L.) after exposing maturing anthers to 34 ºC prior to anthesis, thus increasing seed set. On the other hand, Hirosaki and Niikura [76] observed in Brassica rapa L. that the level of self-incompatibility decreases when maximum temperatures are > 26 ºC and plants are in the last quarter of the flowering period. The factors influencing shifts in the level of self-incompatibility varied across genotypes, depending upon the flowering period, plant senescence, and average temperature prior to pollination [76,81]. Our results show that all populations of P. ionanthum had a higher fertility under open pollination conditions, with an overall average value of 42.4%. Quarin and Norrmann [37] obtained a seed set of 70% under open pollination conditions. Hence, we confirm that the tetraploid cytotype in P. ionanthum is mostly self-sterile, due to a system of self-incompatibility acting during the pollen-pistil interaction, and therefore it behaves as an allogamous species. Our observations also suggest that this self-incompatibility mechanism can be partially broken, depending on internal and external factors, but its influence on the seed set still is to be determined.
Populations of P. regnellii produced seeds through both self- and open pollination, although the respective percentages varied both within and between populations. In self-pollination, the overall population mean was 11.8%, with significant differences observed among populations. Conversely, in open pollination, the overall population mean was 34.9%, without inter-population differences. Norrmann [24] reported seed set percentages of 80 and 84% in self- and open pollination, respectively. The lower fertility observed in this study could be attributed to genetic and/or population-level variations, random effects caused using different sulphite paper envelopes, and/or variations in environmental conditions during data collection. Paspalum regnellii typically grows on the edge of forests or pine plantations, under semi-shaded conditions on wet lateritic red soils with high iron content [17]. However, our evaluations were conducted on plants grown in the experimental field of the Facultad de Ciencias Agrarias (FCA-UNNE) in Corrientes, Argentina, which had conditions that differed significantly from the ideal natural settings. Likewise, Norrmann [24] conducted measurements on a few plants grown in pots with lateritic red soil, kept inside a greenhouse with controlled humidity and temperature conditions. External environment and the maintenance of generous size inflorescences enveloped for several days under natural climatic conditions possibly had a greater influence on the lower seed production observed both in self- and open pollination. Nonetheless, the self-fertility values observed in this study supply evidence of the selfing behavior of P. regnellii.
Similarly, P. urvillei exhibited a similar behavior to P. regnellii regarding its fertility, producing seeds under both self- and open pollination, but with lower values when forced to self-pollinate. In self-pollination, the overall population mean was 41.4%, while in open pollination, it was 74.4%. These values were lower than those observed by Caponio and Quarin [82] in a single P. urvillei plant analysis, where they reported seed sets of 75.0% and 84.4% under self- and open pollination, respectively. The lower fertility observed in this study could be due to the same causes mentioned for P. regnellii, with a greater effect on self-pollination due to the longer enveloped period. The self-fertility values saw in this study also corroborate the selfing behavior of P. urvillei.
The correlation between polyploidy and self-fertilization is widely recognized among many flowering plants [83,84], and in some species of Paspalum, a breakdown of self-incompatibility mechanisms has been seen [21]. Selfing is often found in pioneer plants, weeds, and island endemics [85,86]. Paspalum regnellii and P. urvillei are pioneer species that typically grow in anthropic environments on the edge of forests and roadsides, where selfing is a favorable condition for their local establishment and has likely been selected by evolution. Self-fertilization systems have probably evolved early among angiosperms, requiring the breakdown of self-incompatibility systems to produce seeds [74]. Self-incompatibility causes self-sterility by preventing pollen grains from adhering to the stigma or inhibiting pollen tubes from growing down the style [74,87,88], and it is typically controlled by monogenic or multigenic control of S alleles [89,90,91]. In allotetraploid species like P. durifolium and P. ionanthum, a multiple-locus system may not eliminate the self-incompatible alleles, but it can widen allele diversity and create dosage effects. Compared to a single-allele system that acts on the interaction between haploid gametes, polyploid gametes will have at least two alleles interacting with each other, helping the breakdown of incompatibility. Alternatively, in allotetraploid species like P. regnellii and P. urvillei, the lack of self-incompatibility may be selected by ecological constraints and the evolutionary history of these two species, perhaps originating from parents with a multigenic system, making it difficult to maintain a gene-mediated self-incompatibility system.
5. Conclusions
Our study presents a comprehensive analysis of the genetic systems of four polyploid Paspalum species at the population level. Our results show a stable occurrence of a single ploidy in all species and populations, with sexual, highly fertile tetraploids showing alternative pollination syndromes. However, we saw reproductive variability among individuals and populations, and the occurrence of alternative sexual and apomictic pathways in P. durifolium and P. ionanthum. Although the apomictic pathways were non-functional in our population-level screenings, the study highlights the potential role of unreduced female gametophytes in the formation of new polyploid cytotypes not previously recorded for these species.
Our study also showed that autogamous self-compatible and allogamous self-incompatible tetraploids were equally fertile under open pollination. Nevertheless, the partial breakdown of the self-incompatibility system in P. regnellii and P. urvillei suggests a transition from allogamy to autogamy that may be influenced by the species' particular ecological niches.
These findings have significant implications for biodiversity conservation plans and the management of genetic resources, as well as for genetic improvement programs. Further genetic diversity analysis will supply insights into the evolutionary role of mating systems and polyploidy in the maintenance, dispersal, and adaptation of these species, and support the development of effective conservation strategies.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Number and percentage of ovules observed bearing different embryo sacs types in individual plants of five populations of four Paspalum species.; Table S2: Number of pollinated spikelets and seed set for two pollination conditions (self- and open-pollination) during two flowering periods (1st, 2015-2016 and 2nd, 2016-2017) in populations of four Paspalum species.
Author Contributions
Conceptualization, M.S., D.H.H., A.I.H. and E.J.M.; data curation, software, validation, visualization and formal analysis, M.S., A.V.R., E.A.B, A.L.Z. and F.G., methodology, M.S., A.V.R., A.L.Z., E.A.B., F.G. and C.A.A; writing—original draft preparation, M.S. and E.J.M.; writing—review and editing, A.V.R., D.H.H., C.A.A., A.I.H. and E.J.M.; funding acquisition, D.H.H. and E.J.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financed by grants from the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET, PIP 2012-2014-112-201101-00469), Agencia Nacional de Promoción Científica y Técnica (ANPCYT, PICT-2012-261), ANPCYT-UNNE (PICTO-OTNA 2011-080), Bilateral Cooperation Programme Level II (MINCyT-CONICET-DFG 2015-2017), and Universidad Nacional del Nordeste (UNNE, PI A003-2011). These data are part of the doctoral research of M.S. under the direction of E.J.M. and A.I.H. M.S. received a fellowship from ANPCYT and CONICET. A.V.R, A.L.Z. and E.A.B. received a fellowship from CONICET.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data presented in this study are available in the article and in Supplementary Materials.
Acknowledgments
We thank Jonathan Thompson for reviewing and correcting the English language grammar. We are grateful to Camilo L. Quarin for the technical help in the reproductive analysis.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Meirmans, P.G.; Van Tienderen, P.H. The effects of inheritance in tetraploids on genetic diversity and population divergence. Heredity 2013, 110, 131–137. [CrossRef]
- Jiao, Y.; Wickett, N.J.; Ayampalayam, S.; Chanderbali, A.S.; Landherr, L.; Ralph, P.E.; Tomsho, L.P.; Hu, Y.; Liang, H.; Soltis, P.S.; Soltis, D.E.; Clifton, S.W.; Schlarbaum, S.E.; Schuster, S.C.; Ma, H.; Leebens-Mack, J.; de Pamplilis, C.W. Ancestral polyploidy in seed plants and angiosperms. Nature 2011, 473, 97–100. [CrossRef]
- Stebbins, G.L. Variation and Evolution in plants. Columbia University Press, New York, 1950.
- Grant, V. Polyploidy. Columbia University Press. New York, 1981.
- Otto, S.P.; Whitton, J. Polyploid incidence and evolution. Ann. Rev. Genet. 2000, 34, 401–437. [CrossRef]
- Levin, D.A. The role of chromosomal change in plant evolution. Oxford University Press, New York, United States, 2002.
- Levin, D.A. Polyploidy and novelty in flowering plants. Amer. Nat. 1983, 122, 1–25. [CrossRef]
- Lewis, W.H. Polyploidy: biological relevance. Plenum Press, New York, 1980.
- Osborne, O.; Simon, S.; Collins, S. Attitudes towards science: A review of the literature and its implications. Int. J. Sci. Educ. 2003, 25, 1049–1079, . [CrossRef]
- Coyne, J.A.; Orr, H.A. Speciation. Sunderland, MA. Sinauer Associates, 2004.
- Suda, J.; Kron, P.; Husband, B.C.; Travnıcek, P. Flow cytometry and ploidy: applications in plant systematics, ecology and evolutionary biology. In J Dolezel, J Greilhuber, J Suda, eds., Flow Cytometry with Plant Cells. Analysis of Genes, Chromosomes and Genomes, Wiley–VCH Verlag GmbH & Co. KGaA, Weinheim, 2007; pp. 103–130.
- Lumaret, R. Polyploidy and the critical size of natural populations: the case of cocksfoot (Dactylis glomerata L.), a grass used as a fodder plant. Bocconea 1997, 7, 133–139.
- Karunarathne, P.; Schedler, M.; Martínez, E.J.; Honfi, A.I.; Novichkova, A.; Hojsgaard, D.H. Intraspecific ecological niche divergence and reproductive shifts foster cytotype displacement and provide ecological opportunity to polyploids. Ann. Bot. 2018, 121, 1183–1196. [CrossRef]
- Karunarathne, P.; Reutemann, A. V.; Schedler, M.; Glücksberg, A.; Martínez, E. J.; Honfi, A. I.; Hojsgaard, D. H. Sexual modulation in a polyploidy grass: a reproductive contest between environmentally inducible sexual and genetically dominant apomictic pathways. Sci. Rep. 2020, 10, 8319. [CrossRef]
- Rua, G.H.; Speranza, P.R.; Vaio, M.; Arakaki, M. A phylogenetic analysis of the genus Paspalum (Poaceae) based on cpDNA and morphology. Plant Syst. Evol. 2010, 288, 227–243. [CrossRef]
- Bonasora, M.G.; Pozzobon, M.T.; Honfi, A.I.; Rua, G.H. Paspalum schesslii (Poaceae, Paspaleae), a new species from Mato Grosso (Brazil) with an unusual base chromosome number. Plant Syst. Evol. 2015, 301, 2325–2339. [CrossRef]
- Zuloaga, F.O.; Morrone, O. Revisión de las especies de Paspalum para América del Sur austral (Argentina, Bolivia, sur del Brasil, Chile, Paraguay y Uruguay). Ann. Mo. Bot. Gard. Monogr. Syst. Bot. 2005, 102, 1–297.
- Giussani, L.M.; Zuloaga, F.O.; Quarin, C.L.; Cota-Sánchez, H.; Ubayasena, K.; Morrone, O. Phylogenetic relationships in the genus Paspalum (Poaceae: Panicoideae: Paniceae): an assessment of the Quadrifaria and Virgata informal groups. Syst. Bot. 2009, 34, 32–43. [CrossRef]
- Quarin, C.L. Relaciones citotaxonómicas entre Paspalurn almum Chase y P. hexastachyum Parodi (Gramineae). Bonplandia, 1974, 3, 115-127.
- Davidse, G.; Pohl, R.W. (1974) Chromosome numbers meiotic behaviour and notes on tropical American grasses (Gramineae). Can. J. Bot. 1974, 52, 317–328. [CrossRef]
- Ortiz, J.P.A.; Quarin, C.L., Pessino, S.C.; Acuña, C.A.; Martínez, E.J.; Espinoza, F.; Hojsgaard, D.H.; Sartor, M.E.; Cáceres, M.E.; Pupilli, F. Harnessing apomictic reproduction in grasses: what we have learned from Paspalum. Ann. Bot. 2013, 112, 767–787. [CrossRef]
- Darlington, C.D. Evolution of genetic systems. Cambridge: Cambridge University Press, 1939.
- Hojsgaard, D.H.; Martínez, E.J.; Quarin, C.L. Competition between meiotic and apomictic pathways during ovule and seed development results in clonality. New. Phytol. 2013, 197, 336–347. [CrossRef]
- Norrmann, G.A. Citología y modo de reproducción en dos especies de Paspalum (Gramineae). Bonplandia 1981, 17, 149–158.
- Quarin, C.L.; Norrmann, G.A.; Espinoza, F. Evidence for autoploidy in apomictic Paspalum rufum. Hereditas 1998, 129, 119–124. [CrossRef]
- Hörandl, E.; Paun, O. Patterns and sources of genetic diversity in apomictic plants: implications for evolutionary potentials. In Hörandl, E., Grossniklaus, U., van Dijk, P.J., Sharbel, T.F. (ed.) Apomixis: Evolutions, mechanisms and perspectives, A.R.G. Gantner Verlag, Rugell, Liechtenstein, 2007, pp. 170–194.
- Brugnoli, E.A.; Urbani, M.H., Quarin, C.L.; Zilli, A.L.; Martínez, E.J.; Acuña, C.A. Diversity in apomictic populations of Paspalum simplex Morong. Crop. Sci. 2014, 54, 1656–1664. [CrossRef]
- Daurelio, L.D.; Espinoza, F.; Quarin, C.L.; Pessino, S.C. Genetic diversity in sexual diploid and apomictic tetraploid populations of Paspalum notatum situated in sympatry or allopatry. Plant Syst. Evol. 2004, 244, 189–199. [CrossRef]
- Sartor, M.E.; Quarin, C.L.; Urbani, M.H.; Espinoza, F. Ploidy levels and reproductive behaviour in natural populations of five Paspalum species. Plant Syst. Evol. 2011, 293, 31–41. [CrossRef]
- Urbani, M.H.; Quarin, C.L.; Espinoza, F.; Penteado, M.I.O.; Rodrigues, I.F. Cytogeography and reproduction of the Paspalum simplex polyploid complex. Plant Syst. Evol. 2002, 236, 99–105. [CrossRef]
- Gornall, R.J. Population genetic structure in agamospermous plants. In: Hollingsworth, P.M.; Bateman, R.M.; Gornall, R.J., eds. Mol. Syst. Plant Evol. London: Taylor & Francis, 1999, pp. 118–138.
- Quarin, C.L. The nature of apomixis and its origin in Panicoid grasses. Apomixis Newsl. 1992, 5, 8–15.
- Burson, B.L. Cytology of Paspalum chacoense and P. durifolium and their relationship to P. dilatatum. Bot. Gaz. 1985, 146, 124–129. [CrossRef]
- Burson, B.L.; Bennett, H.W. Cytology and reproduction of three Paspalum species. J. Hered. 1970, 61, 129–132. [CrossRef]
- Honfi, A.I., Quarin, C.L. Citología y modo de reproducción de Paspalum durifolium Mez pentaploide (Poaceae: Panicoideae: Paniceae). Lilloa 2009, 45, 85.
- Moraes Fernandes, M.I.B.; Barreto, I.; Salzano, F.M.; Sacchet, A.M.O.F. Cytological and evolutionary relationships in Brazilian forms of Paspalum (Gramineae). Caryologia 1974, 27, 455–465.
- Quarin, C.L.; Norrmann, G.A. Cytology and reproductive behavior of Paspalum equitans, P. ionanthum, and their hybrids with diploid and tetraploid cytotypes of P. cromyorrhizon. Bot. Gaz. 1987, 148, 386–391. [CrossRef]
- Pozzobon, M.T.; Valls, J.F.M.; Dos Santos, S. Contagens cromossômicas em espécies brasileiras de Paspalum L. (Gramineae). Acta Bot. Bras. 2000, 14, 151–162. [CrossRef]
- Quarin, C.L. A tetraploid cytotype of Paspalum durifolium: Cytology, reproductive behavior and its relationship to diploid P. intermedium. Hereditas 1994, 121, 115–118. [CrossRef]
- Martínez, E.J., Quarin, C.L., Hayward, M.D. Genetic control of apospory in apomictic Paspalum species. Cytologia 1999, 64, 425–433. [CrossRef]
- Pozzobon, M.T.; Carvalho Machado, A.C.; Vaio, M.; Valls, J.F.M.; de Souza Peñaloza, A.D.; dos Santos, S.; Côrtes, A.L.; Rua, G.H. Cytogenetic analyses in Paspalum L. reveal new diploid species and accessions. Ciência Rural 2008, 38, 1292-1298.
- Bashaw, E.C.; Hovin, A.W.; Holt, E.C. Apomixis, its evolutionary significance and utilization in plant breeding. In: Norman MJT ed. Proceed. 11th Intl Grassl Congr. Surfers Paradise, Queensland. University of Queensland Press, St. Lucia, 1970, pp. 245-248.
- Honfi, A.I.; Quarin, C.L.; Valls, J.F.M. Estudios cariológicos en gramíneas sudamericanas. Darwiniana 1990, 30, 87–94.
- Pagliarini, M.S.; Carraro, L.R.; Freitas, P.M.; Adamowsky, E.V.; Rocha Batista, L.A.; Valls, J.F.M. Cytogenetic characterization of Brazilian Paspalum accessions. Hereditas 2002, 135, 27–34. [CrossRef]
- Hojsgaard, D.H.; Honfi, A.I.; Rua, G.H.; Daviña, J.R. Chromosome numbers and ploidy levels of Paspalum species from subtropical South America (Poaceae). Genet. Resour. Crop Evol. 2009, 56, 533–545. [CrossRef]
- Pozzobon, M.T.; Valls, J.F.M. Chromosome number in germplasm accessions of Paspalum notatum (Gramineae). Braz. J. Genet. 1997, 20, 29–34. [CrossRef]
- Brugnoli, A.E.; Urbani, M.H.; Quarin, C.L.; Martínez. E.J.; Acuña, C.A. Diversity in diploid, tetraploid and mixed diploid-tetraploid populations of Paspalum simplex. Crop Sci. 2013, 53, 1509–1516. [CrossRef]
- Hijmans, R.J.; Guarino, L.; Mathur, P.; Jarvis, A. Diva Gis. Version 4. Lizard Tech, Inc and University of California, Patente U.S. No. 5,710,835, 2004.
- Galdeano, F.; Urbani, M.H.; Sartor, M.E.; Honfi, A.I.; Espinoza, F.; Quarin, C.L. Relative DNA content in diploid, polyploid, and multiploid species of Paspalum (Poaceae) with relation to reproductive mode and taxonomy. J. Plant Res. 2016, 129, 697–710. [CrossRef]
- Young, B.A.; Sherwood, R.T.; Bashaw, E.C. Cleared-pistil and thick-sectioning techniques for detecting aposporous apomixis in grasses. Canad. J. Bot. 1979, 57, 1668–1672. [CrossRef]
- Zilli, A.L.; Brugnoli, E.A.; Marcón. F.; Billa, M.B.; Rios, E.F.; Martínez, E.J.; Acuña. C.A. Heterosis and expressivity of apospory in tetraploid Bahiagrass hybrids. Crop Sci. 2015, 55, 1189–1201. [CrossRef]
- Matzk, F.; Meister, A.; Schubert, I. An efficient screen for reproductive pathways using mature seeds of monocots and dicots. Plant J. 2000, 21, 97–108. [CrossRef]
- Siena, L.A.; Sartor, M.E.; Espinoza, F.; Quarin, C.L.; Ortiz, J.P.A. Genetic and embryological evidences of apomixis at the diploid level in Paspalum rufum support recurrent auto-polyploidization in the species. Sex. Plant Reprod. 2008, 21, 205–215. [CrossRef]
- Balzarini, M.; Di Rienzo, J. Info-Gen: Software para análisis estadístico de datos genéticos. Facultad de Ciencias Agropecuarias. Universidad Nacional de Córdoba. Argentina, 2003.
- Saura, F. Cariología de gramíneas. Revista de la Facultad de Agronomía y Veterinaria, Universidad de Buenos Aires, 1943, 2, 63–74.
- Pagliarini, M.S.; De Freitas, P.M.; Takayama, S.Y.; Batista, L.A.R. An original meiotic mutation in Paspalum regnellii. Sex. Plant Reprod. 1998, 11, 17–21. [CrossRef]
- Burton, G.W. A cytological study of some species in the genus Paspalum. J. Agric. Res. 1940, 60, 193–198.
- Burton, G.W. A cytological study of some species in the tribe Paniceae. Am. J. Bot. 1942, 19, 355–359. [CrossRef]
- Brown, W.V. A cytological study of some Texas gramineae. Bull. Torrey Bot. Club 1950, 77, 63–76. [CrossRef]
- Brown, W.V.; Emery, H.P. Apomixis in the Gramineae: Panicoideae. Am. J. Bot. 1958, 45, 253–263. [CrossRef]
- Nielsen, E.L. Grass studies III. Additional somatic chromosome complements. Am. J. Bot. 1939, 26, 366–372. [CrossRef]
- Reutemann, A. V.; Honfi, A. I.; Karunarathne, P.; Eckers, F.; Hojsgaard, D. H.; Martínez, E. J. Variation of residual sexuality rates along reproductive development in apomictic tetraploids of Paspalum. Plants 2022 11(13): 1639. https://. [CrossRef]
- Delgado, L.; Galdeano, F.; Sartor, M.E.; Quarin, C.L.; Espinoza, F.; Ortiz, J.P. Analysis of variation for apomictic reproduction in diploid Paspalum rufum. Ann. Bot. 2014, 113, 1211–1218. [CrossRef]
- Quarin, C.L. Seasonal changes in the incidences of apomixis of diploid, triploid, and tetraploid plants of Paspalum cromyorrhizon. Euphytica 1986, 35, 515–522. [CrossRef]
- Norrmann, G.A.; Quarin, C.L.; Burson, B.L. Cytogenetics and reproductive behavior of different chromosome races in six Paspalum species. J. Hered. 1989, 80, 24–28. [CrossRef]
- Martínez, E.J.; Urbani, M.H.; Quarin, C.L.; Ortiz, J.P.A. Inheritance of apospory in Bahiagrass Paspalum notatum. Hereditas 2001, 135, 19–25. [CrossRef]
- Naumova, T.N.; Hayward, M.D.; Wagenvoort, M. Apomixis and sexuality in diploid and tetraploid accessions of Brachiaria decumbens. Sex. Plant Rep. 1999, 12, 43–52. [CrossRef]
- Rebozzio, R.N.; Sartor, M.E.; Quarin, C.L.; Espinoza, F. Residual sexuality and its seasonal variation in natural apomictic Paspalum notatum accessions. Biol. Plant. 2011, 55, 391–395. [CrossRef]
- Reutemann, A. V.; Martínez, E. J.; Schedler, M.;Daviña, J. R.; Hojsgaard, D. H.; Honfi, A. I. Uniparentality: Advantages for range expansion in diploid and diploid-autopolyploid species. Bot. J. Linn. Soc. 2022, 200(4): 563-585. https://. [CrossRef]
- Urbani, M.H. Estudios sobre citología, sistema reproductivo y compatibilidad polen–pistilo de Panicum dichotomiflorum y Paspalum fasciculatum (Gramineae:Paniceae). Darwiniana 1996, 34, 193–198.
- Burton, G.W.; Forbes, I. Jr.; Jackson, J. Effect of ploidy on fertility and heterosis in Pensacola Bahiagrass. Crop Sci. 1970, 10, 63–66. [CrossRef]
- Quarin, C.L. Morfología, citología y sistema reproductivo de una nueva especie de gramínea: Paspalum procurrens. Bol. Soc. Arg. Bot. 1993, 29, 73–76.
- Quarin, C.L. Effect of pollen source and pollen ploidy on endosperm formation and seed set in pseudogamous apomictic Paspalum notatum. Sex. Plant Reprod. 1999, 11, 331–335.
- Hörandl, E. The evolution of self-fertility in apomictic plants. Sex. Plant Rep. 2010, 23, 73–86. [CrossRef]
- Wilkins, P.W.; Thorogood, D. Breakdown of self-incompatibility in perennial ryegrass at high temperature and its uses in breeding. Euphytica 1992, 64, 65–69. [CrossRef]
- Hirosaki, A.; Niikura, S. Developmental and environmental factors affecting level of self-incompatibility response in Brassica rapa L. Sex. Plant Rep. 2008, 21, 123–132. [CrossRef]
- Zilli, A.L.; Acuña, C.A.; Schulz, R.R.; Brugnoli. E.A.; Guidalevich, V.; Quarin, C.L.; Martínez, E.J. Widening the gene pool of sexual tetraploid Bahiagrass: Generation and reproductive characterization of a sexual synthetic tetraploid population. Crop Sci 2018, 58, 762–772. [CrossRef]
- Yamamoto, M.; Nishimura, K.; Kitashiba, H.; Sakamoto, W.; Takeshi, N. High temperature causes breakdown of S haplotype-dependent stigmatic self-incompatibility in self-incompatible Arabidopsis thaliana. J. Exp. Bot. 2019, 70, 5745–5751. [CrossRef]
- Carter, A.L.; McNeilly, T. Effects of increased seed humidity on pollen tube growth and seed set following self-pollination in Brussels sprout (Brassica oleracea var. gemmifera). Euphytica 1975, 24, 805–813. [CrossRef]
- Ockendon, D.J. Effect of hexane and humidity on self-incompatibility in Brassica oleracea. Theor. Appl. Genet. 1978, 52, 113–117. [CrossRef]
- Barrett, S.C.H.; Harder, L.D. The ecology of mating and its evolutionary consequences in seed plants. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 135–57. [CrossRef]
- Caponio, I.; Quarin, C.L. Intra- and interspecific hybridization between Dallisgrass and Vaseygrass. Crop Sci. 1990, 30, 362–364. [CrossRef]
- Mable, B.K. Polyploidy and self-compatibility: is there an association? New. Phytol. 2004, 162, 803–811. [CrossRef]
- Barringer, B.C. Polyploidy and self-fertilization in flowering plants. Amer. J. Bot. 2007, 94, 1527–1533. [CrossRef]
- Barrett, S.C.H.; Harder, L.D.; Worley, A.C. The comparative biology of pollination and mating in flowering plants. Phil. Trans. Roy. Soc. London B Biol. Sci. 1996, 351, 1271–1280. [CrossRef]
- Bernardello, G.; Anderson, G.J.; Stuessy, T.F.; Crawford, D.J. A survey of floral traits, breeding systems, floral visitors, and pollination systems of the angiosperms of the Juan Fernández Islands (Chile). Bot. Rev. 2001, 67, 255–308. [CrossRef]
- Kao, T.H.; Tsukamoto, T. The molecular and genetic bases of S-RNase-based self-incompatibility. Plant Cell 2004, 16, S72–S83. [CrossRef]
- Charlesworth, D.; Vekemans, X.; Castric, V.; Glemin, S. Plant self-incompatibility systems: a molecular evolutionary perspective. New Phytol. 2005, 168, 61–69. [CrossRef]
- Richards, A.J. Plant breeding systems, 2nd Edn. London: Chapman & Hall. 1997.
- de Nettancourt, D. Incompatibility and incongruity in wild and cultivated plants. 2nd ed. Springer, Berlin. 2001. [CrossRef]
- Stone, J.L. Molecular mechanisms underlying the breakdown of gametophytic self-incompatibility. Quart. Rev. Biol. 2002, 77, 17–32. [CrossRef]
Figure 1.
Histograms of cell nuclei from leaves of a sample of unknown ploidy and the standard sample for P. durifolium (a), P. ionanthum (b), P. regnellii (c), P. urvillei (d). In all cases, the ploidy of the unknown sample is the same as the tetraploid (2n = 4x = 40) standard used for each species.
Figure 1.
Histograms of cell nuclei from leaves of a sample of unknown ploidy and the standard sample for P. durifolium (a), P. ionanthum (b), P. regnellii (c), P. urvillei (d). In all cases, the ploidy of the unknown sample is the same as the tetraploid (2n = 4x = 40) standard used for each species.
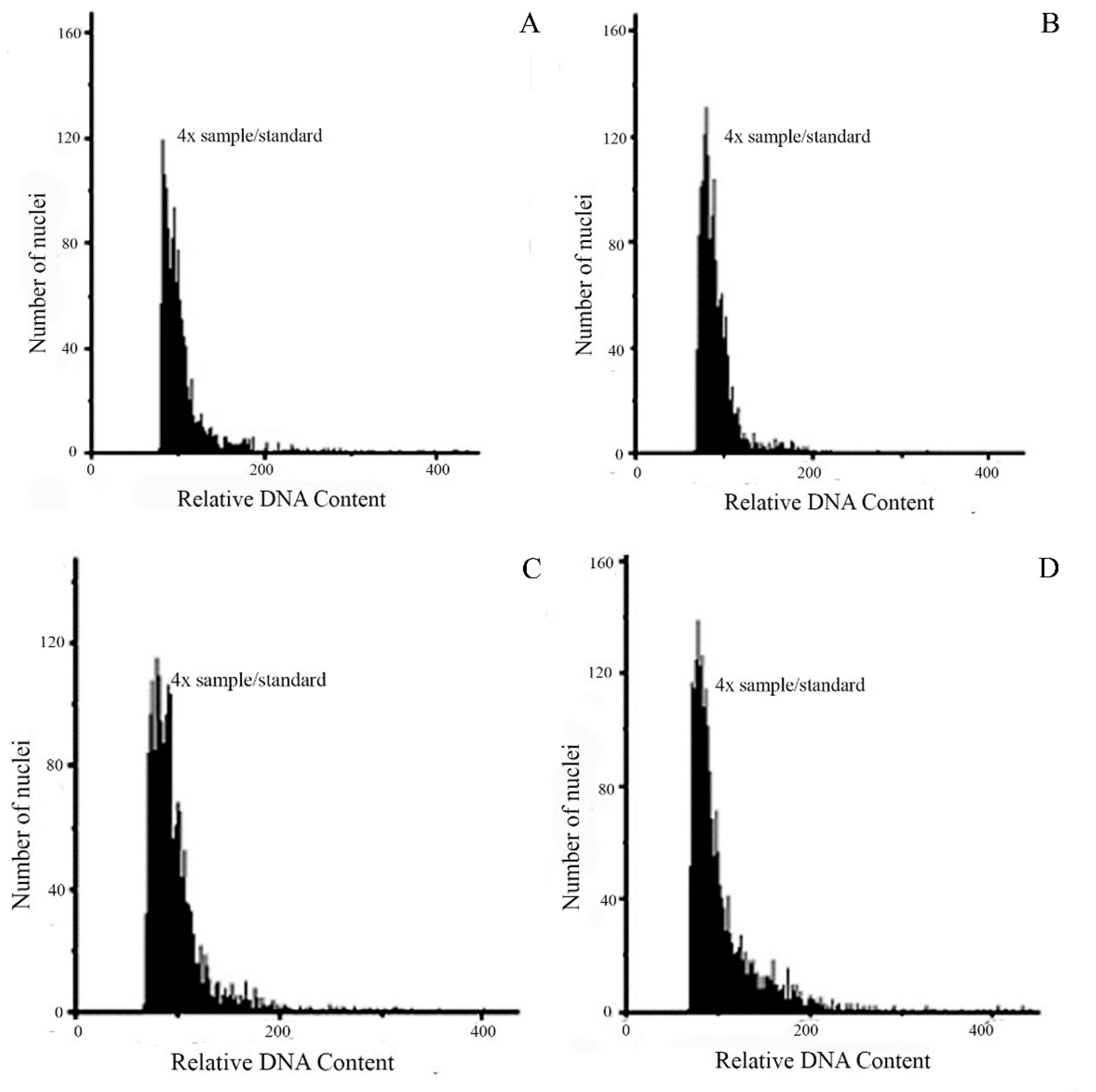
Figure 2.
Ovule images showing meiotic embryo sacs having two polar nuclei (PN) at the micropylar end (bottom), and a mass of antipodes (An) at the chalazal end (top). Egg cells and synergids were visible but in different photographic focus (not shown). Paspalum durifolium (a, e); P. ionanthum (b); P. regnellii (c); P. urvillei (d). Image (e) shows an ovule from P. durifolium carrying a typical meiotic embryo sac plus an aposporous embryo sac with two polar nuclei and lacking antipodes. Bar = 100 µm.
Figure 2.
Ovule images showing meiotic embryo sacs having two polar nuclei (PN) at the micropylar end (bottom), and a mass of antipodes (An) at the chalazal end (top). Egg cells and synergids were visible but in different photographic focus (not shown). Paspalum durifolium (a, e); P. ionanthum (b); P. regnellii (c); P. urvillei (d). Image (e) shows an ovule from P. durifolium carrying a typical meiotic embryo sac plus an aposporous embryo sac with two polar nuclei and lacking antipodes. Bar = 100 µm.
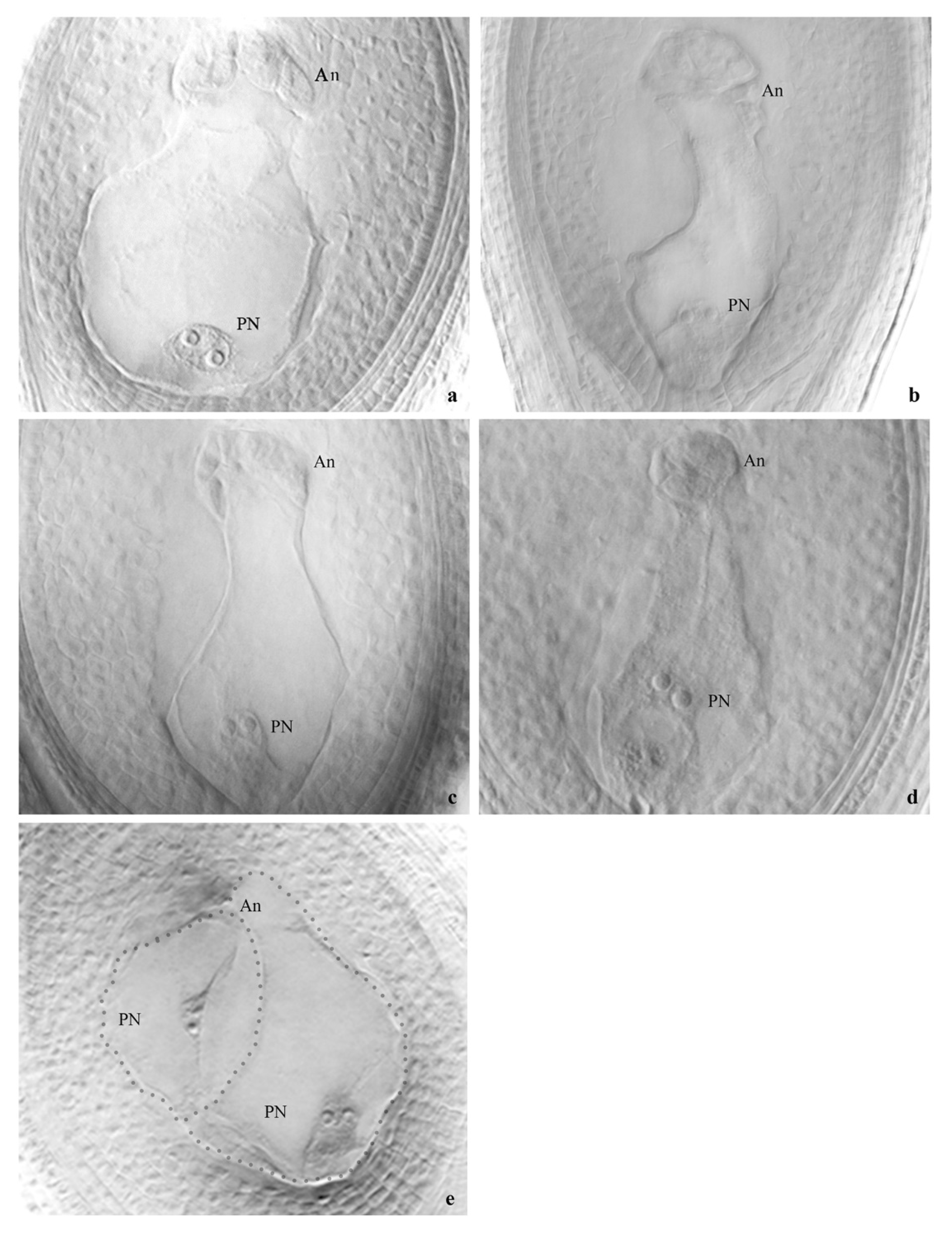
Figure 3.
Histogram of cell nuclei from five bulked seeds of a tetraploid plant from the PD3 population of Paspalum durifolium. Peaks of relative DNA contents at 100 and 200 correspond to G1 and G2 of tetraploid embryos; while at 150 and 300 to endosperm of tetraploid seeds. These peaks show the occurrence of sexual seeds originated by fertilization of meiotically reduced egg cells and reduced male gametes (Em/En ratio: 2C/3C). Two small peaks of relative DNA content at 400 and 600 correspond to embryo and endosperm, respectively, from an hexaploid seed originated by fertilization of a non-reduced egg cell and one reduced male gamete (BIII hybrid, 2n + n). Embryo (Em), Endosperm (En).
Figure 3.
Histogram of cell nuclei from five bulked seeds of a tetraploid plant from the PD3 population of Paspalum durifolium. Peaks of relative DNA contents at 100 and 200 correspond to G1 and G2 of tetraploid embryos; while at 150 and 300 to endosperm of tetraploid seeds. These peaks show the occurrence of sexual seeds originated by fertilization of meiotically reduced egg cells and reduced male gametes (Em/En ratio: 2C/3C). Two small peaks of relative DNA content at 400 and 600 correspond to embryo and endosperm, respectively, from an hexaploid seed originated by fertilization of a non-reduced egg cell and one reduced male gamete (BIII hybrid, 2n + n). Embryo (Em), Endosperm (En).
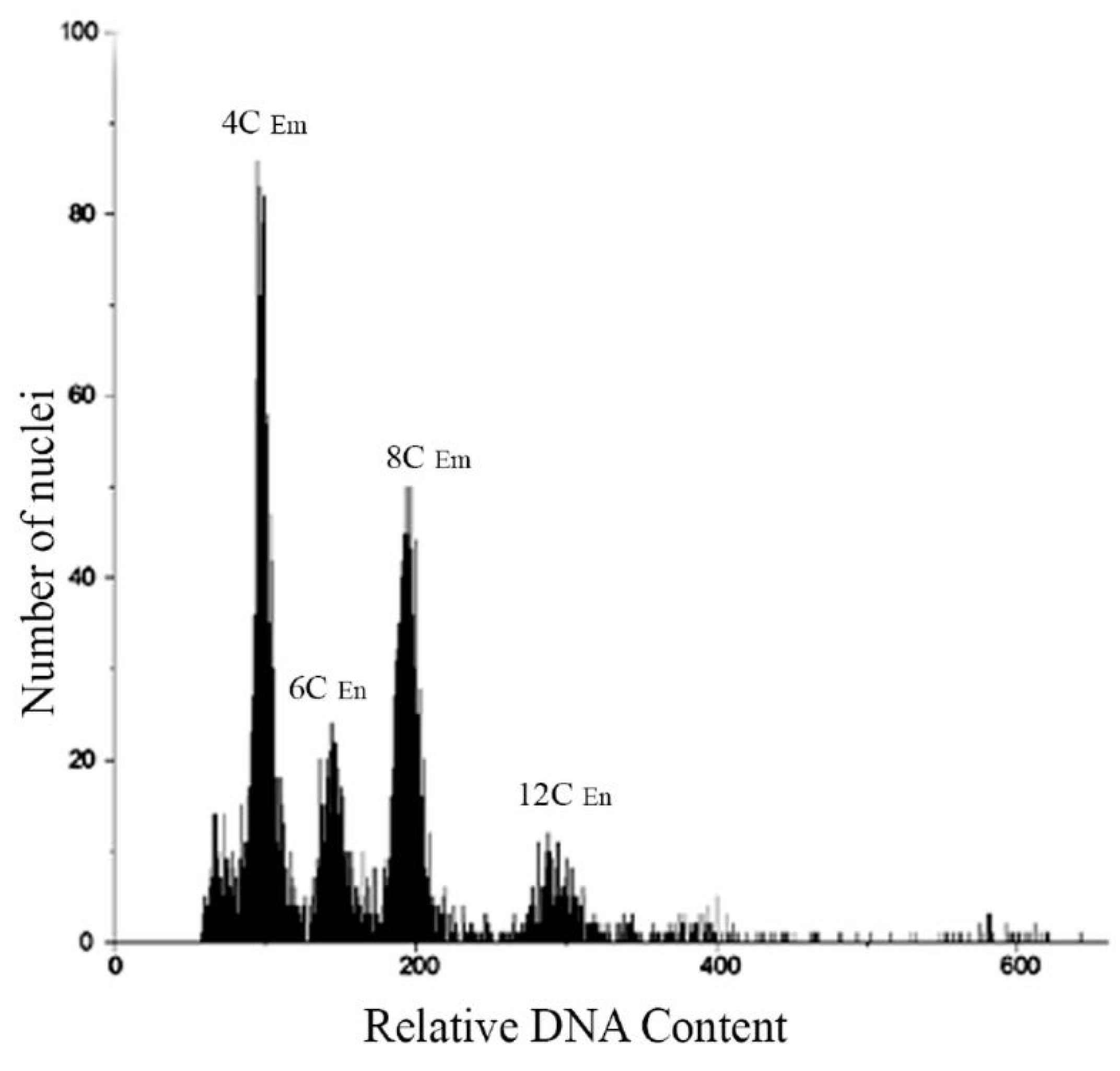

Table 1.
Identification, collection sites, and the number of plants analyzed of natural populations of Paspalum species from north-eastern Argentina.
Table 1.
Identification, collection sites, and the number of plants analyzed of natural populations of Paspalum species from north-eastern Argentina.
| Species | Population code (Voucher) | Collection sites | Geographical coordinates | No. Plants |
|---|---|---|---|---|
| P. durifolium | PD1 (M6) | Corrientes, Dpto. Ituzaingó, NR 12 | 27°37.622S, 56°44.873W | 19 |
| PD2 (M7) | Corrientes, Dpto. San Miguel, NR 118, and PR 5 | 27°55.901S, 57°30.996W | 18 | |
| PD3 (M16) | Corrientes, Dpto. Santo Tomé, Esteros del Iberá, PR 40 | 28°17.435S, 56°46.219W | 17 | |
| PD4 (M17) | Corrientes, Dpto. Santo Tomé, NR 120 | 27°49.513S, 56°15.294W | 18 | |
| PD5 (M18) | Corrientes, Dpto. Ituzaingó, NR 118 | 27°33.646S, 57°09.184W | 20 | |
| P. ionanthum | PI1 (M4) | Corrientes, Dpto. Ituzaingó, NR 120 | 27°49.265S, 56°16.328W | 20 |
| PI2 (M8) | Corrientes, Dpto. Concepción, Santa Rosa, NR 118 | 28°14.245S, 58°03.252W | 20 | |
| PI3 (M10) | Corrientes, Dpto. General Paz, Paso Florentín, PR 5 | 27°45.205S, 57°45.658W | 16 | |
| PI4 (M11) | Corrientes, Dpto. Mburucuyá, Parque Nacional Mburucuyá. | 28°02.024S, 58°02.172W | 20 | |
| PI5 (M12) | Corrientes, Dpto. Goya, Paraje Marucha, NR 12 | 29°09.906S, 59°05.795W | 20 | |
| P. regnellii | PR1 (M2) | Misiones, Dpto. Montecarlo, Puerto Laharrague, NR 12 | 26°32.492S, 54°43.506W | 20 |
| PR2 (M19) | Misiones, Dpto. San Ignacio, Jardín América, NR 12 | 27°03.170S, 55°15.040W | 19 | |
| PR3 (M20) | Misiones, Dpto. San Pedro, NR 14 | 26°43.626S, 54°14.845W | 20 | |
| PR4 (M21) | Misiones, Dpto. 25 de Mayo, PR 2 | 27°22.441S, 54°25.503W | 18 | |
| PR5 (M22) | Misiones, Dpto. 25 de Mayo, PR 4 | 27°46.628S, 55°14.998W | 19 | |
| P. urvillei | PU1 (M1) | Misiones, Dpto. San Ignacio, 2,2 Km East PR 210 | 27°16.613S, 55°27.735W | 17 |
| PU2 (M5) | Corrientes, Dpto. Ituzaingó, NR 120 | 27°49.265S, 56°16.328W | 20 | |
| PU3 (M14) | Entre Ríos, Dpto. La Paz, PR 6 | 31°01.223S, 59°25.222W | 20 | |
| PU4 (M23) | Santa Fe, Dpto. Gral. Obligado, NR 11 | 28°35.733S, 59°25.005W | 20 | |
| PU5 (M27) | Chaco, Dpto. Gral. San Martin, PR 90 | 26°24.117S, 59°22.726W | 17 |
PD = P. durifolium; PI = P. ionanthum; PR = P. regnellii; PU = P. urvillei; M = Martínez E. J. et al.; RN = National route; PR = Provincial route.
Table 2.
Observed number and percentage (%) of embryo sac types, and the observed proportions of the sexual (SP) and apomictic (AP) pathways at ovule stage in populations of four Paspalum species.
Table 2.
Observed number and percentage (%) of embryo sac types, and the observed proportions of the sexual (SP) and apomictic (AP) pathways at ovule stage in populations of four Paspalum species.
| Species | Pop | n | Number of ovules bearing (%) | Proportions | |||||
|---|---|---|---|---|---|---|---|---|---|
| MES | AES | MES + AES | AbES | SP | AP | p | |||
| P. durifolium | PD1 | 152 | 127 (83.5) | - | 1 (0.7) | 24 (15.8) | 0.99 | 0.01 | <0.001 |
| PD2 | 150 | 141 (94.0) | - | - | 9 (6.0) | 1.00 | 0.00 | <0.001 | |
| PD3 | 152 | 134 (88.2) | - | 6 (3.9) | 12 (7.9) | 0.96 | 0.04 | <0.001 | |
| PD4 | 153 | 129 (84.3) | - | 10 (6.5) | 14 (9.2) | 0.93 | 0.07 | <0.001 | |
| PD5 | 156 | 124 (79.5) | - | 5 (3.2) | 27 (17.3) | 0.96 | 0.04 | <0.001 | |
| P. ionanthum | PI1 | 158 | 150 (95.0) | - | 4 (2.5) | 4 (2.5) | 0.97 | 0.03 | <0.001 |
| PI2 | 151 | 147 (97.4) | - | 2 (1.3) | 2 (1.3) | 0.99 | 0.01 | <0.001 | |
| PI3 | 150 | 149 (99.3) | - | - | 1 (0.7) | 1.00 | 0.00 | <0.001 | |
| PI4 | 165 | 160 (97.0) | - | - | 5 (3.0) | 1.00 | 0.00 | <0.001 | |
| PI5 | 158 | 156 (98.7) | - | - | 2 (1.3) | 1.00 | 0.00 | <0.001 | |
| P. regnellii | PR1 | 160 | 112 (70.0) | - | - | 48 (30.0) | 1.00 | 0.00 | <0.001 |
| PR2 | 151 | 132 (87.4) | - | - | 19 (12.6) | 1.00 | 0.00 | <0.001 | |
| PR3 | 155 | 148 (95.5) | - | - | 7 (4.5) | 1.00 | 0.00 | <0.001 | |
| PR4 | 170 | 162 (95.3) | - | - | 8 (4.7) | 1.00 | 0.00 | <0.001 | |
| PR5 | 164 | 153 (93.3) | - | - | 11 (6.7) | 1.00 | 0.00 | <0.001 | |
| P. urvillei | PU1 | 167 | 160 (95.8) | - | - | 7 (4.2) | 1.00 | 0.00 | <0.001 |
| PU2 | 158 | 153 (96.8) | - | - | 5 (3.2) | 1.00 | 0.00 | <0.001 | |
| PU3 | 156 | 155 (99.4) | - | - | 1 (0.6) | 1.00 | 0.00 | <0.001 | |
| PU4 | 156 | 151 (96.8) | - | - | 5 (3.2) | 1.00 | 0.00 | <0.001 | |
| PU5 | 155 | 155 (100) | - | - | - | 1.00 | 0.00 | <0.001 | |
MES, meiotic embryo sac; AES, aposporous embryo sac; MES + AES, meiotic embryo sac plus aposporous embryo sac; AbES, aborted embryo sac. Significant differences (p < 0.05).
Table 3.
Expected (Ei) and observed (Oi) proportions of sexuality and apomixis at the seed stage and the reproductive efficiency of each pathway in populations of four Paspalum species.
Table 3.
Expected (Ei) and observed (Oi) proportions of sexuality and apomixis at the seed stage and the reproductive efficiency of each pathway in populations of four Paspalum species.
| Species | Pop | Sexual pathways | Apomictic pathways | Statistical analysis | Reproductive efficiency | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ei | Oi | Ei | Oi | χ2 | p | Sex | Apo | ||
| P. durifolium | PD1 | 0.99 | 1.00 | 0.01 | 0.00 | 4.67e-31 | 1.00 | 1.01 | - |
| PD2 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.06 | - | |
| PD3 | 0.96 | 1.00 | 0.04 | 0.00 | 0.333 | 0.564 | 1.04 | - | |
| PD4 | 0.93 | 1.00 | 0.07 | 0.00 | 1.049 | 0.3055 | 1.07 | - | |
| PD5 | 0.96 | 1.00 | 0.04 | 0.00 | 0.237 | 0.626 | 1.04 | - | |
| P. ionanthum | PI1 | 0.97 | 1.00 | 0.03 | 0.00 | - | - | 1.03 | - |
| PI2 | 0.99 | 1.00 | 0.01 | 0.00 | - | - | 1.01 | - | |
| PI3 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PI4 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PI5 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| P. regnellii | PR1 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - |
| PR2 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PR3 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PR4 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PR5 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| P. urvillei | PU1 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - |
| PU2 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PU3 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PU4 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PU5 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
χ2, Chi-square statistics, significant differences (p < 0.05)
Table 4.
Mean comparative analysis of seed set within and among populations for two pollination methods (self- and open pollination) during two flowering periods (1st and 2nd).
Table 4.
Mean comparative analysis of seed set within and among populations for two pollination methods (self- and open pollination) during two flowering periods (1st and 2nd).
| Species | Pop | Seed set (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Self-pollination | Open pollination | Self-pollination | Open pollination | |||||||
| 1st period | 2nd period | p valueα | 1st period | 2nd period | p valueα | Total† | Total† | p valueβ | ||
| P. durifolium | PD1 | 0.27a | 0.61a | 0.281 | 28.09a | 29.58a | 0.839 | 0.44a | 28.83b | <0.001 |
| PD2 | 0.35a | 0.65a | 0.333 | 46.30a | 51.54a | 0.326 | 0.50a | 48.92a | <0.001 | |
| PD3 | 0.16a | 0.65a | 0.262 | 57.90a | 45.16a | 0.242 | 0.41a | 51.53a | <0.001 | |
| PD4 | 1.24a | 0.47a | 0.409 | 45.52a | 42.70a | 0.881 | 0.86a | 44.11ab | <0.001 | |
| PD5 | 0.17a | 1.18a | 0.119 | 33.17a | 27.97a | 0.507 | 0.68a | 30.57b | <0.001 | |
| p valueγ | 0.284 | 0.701 | 0.173 | 0.058 | 0.74 | 0.006 | ||||
| P. ionanthum | PI1 | 1.00a | 1.00a | 0.995 | 35.60a | 23.20a | 0.168 | 1.04a | 29.41c | <0.001 |
| PI2 | 0.26a | 1.22a | 0.418 | 60.25a | 31.27b | 0.002 | 0.75a | 45.80ab | <0.001 | |
| PI3 | 4.26a | 1.36a | 0.337 | 49.66a | 58.88a | 0.661 | 2.79a | 54.27a | <0.001 | |
| PI4 | 0.16a | 1.10a | 0.164 | 52.80a | 39.22a | 0.092 | 0.61a | 46.02ab | <0.001 | |
| PI5 | 2.85a | 0.71a | 0.089 | 45.53a | 27.82a | 0.036 | 1.80a | 36.69bc | <0.001 | |
| p valueγ | 0.565 | 0.972 | 0.191 | 0.004 | 0.574 | 0.017 | ||||
| P. regnellii | PR1 | 8.51a | 4.96a | 0.44 | 23.47a | 34.19a | 0.360 | 6.73b | 28.83b | <0.001 |
| PR2 | 25.93a | 6.19b | 0.018 | 38.14a | 44.17a | 0.602 | 16.06ab | 41.16ab | 0.001 | |
| PR3 | 12.44a | 9.74a | 0.704 | 24.06a | 28.70a | 0.682 | 11.09ab | 26.38b | 0.031 | |
| PR4 | 10.48a | 2.34a | 0.16 | 35.83a | 20.77a | 0.144 | 6.41b | 28.30b | <0.001 | |
| PR5 | 25.16a | 12.47a | 0.055 | 62.20a | 37.35a | 0.103 | 18.81a | 49.77a | 0.001 | |
| p valueγ | 0.059 | 0.142 | 0.006 | 0.426 | 0.039 | 0.026 | ||||
| P. urvillei | PU1 | 44.61a | 40.75a | 0.828 | 80.15a | 59.76a | 0.12 | 42.68a | 69.96a | 0.018 |
| PU2 | 30.76a | 52.99a | 0.071 | 86.87a | 74.41a | 0.269 | 41.88a | 80.64a | <0.001 | |
| PU3 | 38.47a | 32.75a | 0.671 | 92.01a | 64.01b | 0.007 | 35.61a | 78.01a | <0.001 | |
| PU4 | 33.34a | 52.22a | 0.224 | 76.71a | 69.56a | 0.428 | 42.78a | 73.32a | 0.004 | |
| PU5 | 31.80a | 54.44a | 0.057 | 74.00a | 66.05a | 0.391 | 43.12a | 70.03a | 0.002 | |
| p valueγ | 0.906 | 0.200 | 0.091 | 0.767 | 0.893 | 0.483 | ||||
Tukey´s test significant values are in bold (p < 0.05). †Overall mean values for either self- or open-pollination condition. αWithin populations significance level between flowering periods (1st vs. 2nd) for either self- or open-pollination condition. βWithin populations significance level between pollination conditions (self- vs. open pollination), considering the two flowering periods together. γ Among populations significance level for either self- or open-pollination condition. Bold values show significant differences.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



