Submitted:
08 May 2023
Posted:
10 May 2023
You are already at the latest version
Abstract
Increasing clinical and preclinical evidences implicate gut microbiome (GM) dysbiosis as a key susceptibility factor for neurodegenerative disorders, including Alzheimer’s disease (AD) and Parkinson’s diseases (PD). In recent years, neurodegenerative diseases have not been viewed as driven solely by defects in brain, and the role of GM in modulating the central nervous system function via gut-brain axis has attracted considerable interest. Encouraged by GM researches, the next-generation probiotics may lead to tangible impacts on the treatment of neurodegenerative disorders. This review summarizes the current understandings of GM composition and characteristics associated with neurodegenerative diseases, and the demonstrations of molecules from GM affecting neurodegeneration. Particularly, applications of next-generation probiotics such as Clostridium butyricum, Akkermansia muciniphila, Faecalibacterium prausnitzii, and Bacteroides fragilis for remediation of neurodegenerative diseases are discussed.
Keywords:
Gut microbiome
; neurodegenerative diseases
; next-generation probiotics
; Clostridium butyricum
; Akkermansia muciniphila
; Faecalibacterium prausnitzii
; Bacteroides fragilis
1. Introduction
Age-dependent neurodegenerative diseases, including Alzheimer’s disease (AD) and Parkinson’s disease (PD), are chronic and progressive neurological disorders with a range of causes and clinical presentations [1,2]. The prevalence of neurodegenerative diseases has increased worldwide in parallel with the rise in life expectancy, and effective treatments for neurodegenerative diseases are highly desired. Of note, the primary prodromal symptom of AD and PD, gastrointestinal dysfunction, is detected prior to the clinical diagnosis, suggesting that the gastrointestinal tract and its connection to the central nervous system (CNS) are involved in the disease aetiology [3]. Among other causes (genetic, immune system, etc.) attributing to AD and PD pathogenesis [4], gut dysbiosis is an emerging factor and receiving more and more attentions. Clinic treatments that target gut microbiota provide a new and promising approach for reducing the risk, modulating the symptoms, and delaying the neurodegenerative progression.
In this communication, we review recently progresses in the understanding of gut dysbiosis and neurodegeneration, particularly emphasize on the potential application of next-generation probiotics for AD and PD therapies.
2. Pathological Features Of Neurodegenerative Diseases
Neurodegenerative diseases are relatively common, progressive, and devastating neurological disorders. As the population aged, the incidence and prevalence of neurodegenerative diseases have risen rapidly in the past two decades. AD is the most prevalent neurodegenerative disease and the leading cause of dementia worldwide. Pathologically, AD is characterized by β-amyloid (Aβ)-containing extracellular plaques and tau-containing intracellular neurofibrillary tangles [2]. The presentation of AD with memory impairment is most common, but the difficulties in expressive speech, visuospatial processing and executive functions co-occur, ultimately with the classic clinical signs of dementia. PD apparently results from the complex interplay of α-synuclein aggregation, neuroinflammation, mitochondrial dysfunction, and abnormal synaptic transmission, leading to the gradual, irreversible loss of dopaminergic neurons in the substantia nigra and the resulting striatal dopamine depletion [5]. In addition to the cardinal motor symptoms such as resting tremor, rigidity, bradykinesia, PD patients also exhibit non-motor symptoms, including hyposmia, sleep disorders (rapid eye movement sleep behavior disorder), psychiatric symptoms (anxiety, depression), cognitive impairment, and gastrointestinal disturbances (constipation, delayed gastric emptying, dysphagia, sialorrhoea) [6].
In general, approximately 10-30 years before the onset of dementia, Aβ peptides begin to accumulate, which occurs in the early stages of AD and is detectable in the basal temporal and medial frontal regions [7]. The early stage of PD is difficult to recognize, and by the time when patients notice the motor symptoms, the disease has usually advanced with a long period. It has been shown that non-motor symptoms, particularly gastrointestinal dysfunction, frequently occur approximately 20 years before the neurodegeneration appear. Considering the high prevalence of gastrointestinal symptoms, it has been considered as the prodromal phase of neurodegenerative diseases [6]. Specifically, the pathology in the gastrointestinal tract shows the similarity with brain. The misfolded α-synuclein and Aβ confer prion-like properties and spread gradually from the enteric nervous system (ENS) to specific brain regions, subsequently seeding the toxic aggregates in recipient neurons [8,9]. More studies are needed to understand the relationship between gastrointestinal symptoms and disease progression, with the aim discovering new biomarkers for diagnosis. Furthermore, in light of the many recent developments in gastrointestinal dysfunction, the gut might be a gateway for the development of an urgently needed disease-modifying therapy.
3. Gut microbiome alterations in neurodegenerative diseases
A healthy human gut harbors a microbiome of 200-400 species and trillions of microbial cells. This gut microbiome (GM) is dynamic, and the microbial composition and the abundances of species are affected by environment, diet, age, feeding mode, application of antibiotics, etc. [10]. Notably, the GMs of AD and PD patients displayed different features from healthy GMs, which might be involved in the pathogenesis of neurodegeneration by regulating gut barrier integrity, neuroinflammation, immune response, and neurotransmitter activity [11,12,13]. The GMs of fecal samples from AD patients showed increased abundances of Ruminococcaceae [13], Enterococcus [14], Streptococcus [15], Alistipes [16], Dorea [15], Collinsella [13,14], and Eggerthella [14], while decreased abundances of Faecalibacterium [14], Lachnospira [13,17], Roseburia [14], and Coprococcus [14]. In addition to AD, more and more evidences showed that the GM dysbiosis was implicated in PD-related pathology. Profiling of the GMs of PD patients reported many alterations compared to healthy controls, including the increased abundances of Alistipes [18], Streptococcus [19], Ruminococcus [20,21], Enterobacter [19,22], Enterococcus [19,23], Verrucomicrobium [20,24], Desulfovibrio [25], and Anaetroncus [26], whereas decreased in Faecalibacterium [19,24], Prevotella [11,20,22,27], Blautia [19,24,28], Lachnospira [24,25], and Roseburia [11,24,28]. Summarizing the current studies indicated that AD and PD shared common GM dynamics. i.e., increased abundances of Streptococcus, Ruminococcaceae, and Alistipes, and decreased abundances of Faecalibacterium, Lachnospira, and Roseburia (Table 1). It was speculated that AD and PD might be correlated with GM-induced inflammation and neurodegeneration. While an increasing number of studies revealed the association of GM dysbiosis with neurodegenerative diseases, further causative studies are still needed to reveal the mechanisms and the potential relevance to clinical manifestations.
4. Microbiota-gut-brain axis
The microbiota-gut-brain axis is well accepted that the GM exerts considerable influence on brain function. The GM communicates with the brain via the activation of vagus nerve, stimulation of enterochromaffin cells, immune system and direct transport of metabolites from the circulation into the brain [29]. In regard to the neuronal pathways for gut-brain connections, vagus nerve is the most direct and well-studied pathway. In autism spectrum disorder (ASD) mice, Lactobacillus reuteri has been reported to rescue social dysfunction in a vagus nerve-dependent manner [30]. It was found that microbial production of indole from tryptophan was more likely to develop host anxiety and depression, because bacterial indole would activate vagal neurons and negatively impact emotional behaviors [31]. The propagation of Aβ and α-synuclein in the gastrointestinal tract were transmitted via vagus nerve to brain [32,33]. Colonic enterochromaffin cells express receptors for various GM-derived metabolites, such as short-chain fatty acids (SCFAs), aromatic amino acids and secondary bile acids [34,35]. Furthermore, enterochromaffin cell production of serotonin has the potential to influence brain function directly or indirectly [36]. For immune-mediated routes, SCFAs interact closely with the immune system through activation of G protein-coupled receptor (GPCR) and inhibition of histone deacetylase (HDAC) activity, leading to the decreased neuroinflammation [37,38]. On the other direction, bacterial endotoxin lipopolysaccharide (LPS) has been shown to aggravate the neuroinflammation via directly entering the brain or activating immune response [39].
5. Linking gut microbiome dysbiosis and neurodegenerative diseases
5.1. Gut microbiome interacts with hosts Subsection
GM and host interaction is an important direction to understand the regulation of health and disease. As previously reported, the GMs from patients with bipolar disorder depression were sufficient to induce depression-like behavior in mice, which was attributed to the elevated expression of tetratricopeptide repeat and ankyrin repeat containing 1 (TRANK1), a robust risk gene of bipolar disorder [40]. Inspiringly, the interplay between GM and host was investigated in PD. Intracellular protein aggregates primarily composed of α-synuclein in Lewy bodies serve as the neuropathological hallmark of PD. α-Synuclein is encoded by SNCA gene, of which mutations lead to the drastic overexpression of α-synuclein and cause Mendelian autosomal dominant PD [41]. Considering an overabundance of opportunistic pathogens in PD, whether these pathogens are triggers of the neurodegeneration is being investigated and there is likely a connection to SNCA variants. Recently, Wallen et al. [42] reported the association of three opportunistic pathogens with PD, which is dependent on SNCA genetic variations. The candidate interacting SNCA genetic variants for Corynebacterium, Porphyromonas, and Prevotella are rs356229, rs10029694, and rs6856813, respectively. Of which, the Porphyromonas interacting genetic variant is also associated with PD risk. These findings indicate that the increased abundance of opportunistic pathogens in PD gut might be modulated by host genotype. In this sense, the genetic susceptibility to disease and the GM dysbiosis are interacted (Figure 1). Nonetheless, the power of single gene in explaining host gene and GM interaction is limited, and the conclusions may be partial and misleading [43]. Further studies to integrate multiple genetic variations in experimental models and humans will be needed to tease out these interactions.
5.2. Gut microbiome-mitochondria connection
As the endosymbiosis hypothesis demonstrated, the mitochondria are descendants of primordial aerobic pleomorphic bacteria (likely Rickettsia), which develops into a mutualistic partnership with ancient anaerobic microbes (likely Archaea) [44]. As a consequence, a stable symbiosis is established to provide energy for the host. Bacterial peptidoglycan muropeptides, the unique component of bacterial cell walls in both Gram-positive and Gram-negative species, accumulate in host intestinal mitochondria, which can maintain the mitochondrial homeostasis and suppress host mitochondrial oxidative stress [45]. Moreover, intestinal infection with Gram-negative bacteria Citrobacter rodentium can trigger mitochondrial antigen presentation and elicit the mitochondria-specific autoreactive CD8+ T cells to kill dopaminergic neurons, thereby causing the transient motor dysfunction resembling in PD (Figure 1) [46]. β-N-methylamino-L-alanine (BMAA), a natural neurotoxin produced by cyanobacteria or other microbes, has been shown to be involved in the neurodegeneration. BMAA can cross the blood-brain barrier (BBB), which was previously detected in the brains of patients with neurodegenerative disease [47]. Mechanistically, BMAA elicits mitochondrial dysfunction and AD features in cortical neurons with increased tau phosphorylation and Aβ peptides deposition [48]. Recently, Esteves et al. [49] reported that BMAA triggered a chain of events including mitochondrial dysfunction and innate immunity activation, which recapitulates the PD pathology from the gut to the brain. Although there is still no evidence that gut microbiota can produce BMAA, chronic exposure to environmental microbial neurotoxins is sufficient to induce mitochondrial dysfunction, which has been advanced as a potential cause for the neurodegeneration [50].
5.3. Defective autophagy
Autophagy, in a broad sense, refers to a cellular homeostatic mechanism delivering cytoplasmic constituents to lysosomes for degradation. Initially described as a “self-eating” survival pathway that enables nutrient recycling during starvation, autophagy can also respond to a range of inputs, including microbial products commonly as pathogen-associated molecular patterns (PAMPs) [51]. One of the best-appreciated manifestations of autophagy is to defend against microbial invasion through direct elimination of intracellular pathogens [52]. Autophagy degrades invading pathogens (e.g., Salmonella and Escherichia/Shigella), modulates the release of proinflammatory cytokines, and participates in the antigen presentation. In the intestinal epithelial cells, autophagy enhances tight junction barrier function owing to the reduced permeability of ions and small molecules by lysosomal degradation of claudin-2 [53]. Furthermore, autophagy in colonic epithelial cells has been reported to protect against colitis by the maintenance of antimicrobial peptides and secretion of mucins that act as a mucosal barrier against bacterial invasion [54,55]. The disruption of autophagy in the intestinal epithelium cells induces the alterations in the composition of gut microbiota and reduces α-diversity. In autophagy deficient mice, the abundance of Candidatus Arthromitus and Pasteurellaceae is increased, whereas Akkermansia muciniphila and Lachnospiraceae are found to be reduced (Figure 1) [55]. Indeed, both AD and PD are accompanied by the defective autophagy, leading to the failure of eliminating protein aggregates or damaged mitochondria. Given the impact of autophagy dysfunction in gastrointestinal homeostasis, there is therapeutic interest in activating autophagy to eliminate pathogenic bacteria and the spread of toxic misfolded protein aggregates, thus halting the progression of neurodegenerative diseases.
6. Next-generation of probiotics in neurodegenerative diseases
6.1. Clostridium butyricum
Clostridium butyricum, a butyrate-producing, spore-forming anaerobic bacterium, is found in a wide variety of environments, including soil, milk, and vegetables. C. butyricum was detected in 10-20% of the human gastrointestinal tract, and is one of the earliest colonizers in infants [56]. Traditionally, C. butyricum have been used as a potent probiotic owing to the beneficial effects on host health. Because of the increased butyrate production, C. butyricum is able to enhance the thickness of the mucosal layer and strengthen the gut barrier integrity via increasing the expression of tight junction proteins (e.g., occludin and ZO-1). In addition, C. butyricum plays a protective role in gastrointestinal infections and regulate the host immune system [57]. C. butyricum is effective against Clostoridioides difficile, the causative pathogen of nosocomial infections; Helicobacter pylori, the causative pathogen of gastric cancer; antibiotic-resistant Escherichia coli, as well as Staphylococcus aureus and Vibrio cholerae infection [58,59,60]. It has been shown that C. butyricum can also upregulate protectin D1, an anti-inflammatory lipid metabolite, in colon tissue under antibiotic therapy to alleviate systemic inflammation [61].
C. butyricum exerts neuroprotective effects in various neurodegenerative diseases. In PD mice model, oral administration of C. butyricum can improve motor deficits, dopaminergic neuron loss, synaptic dysfunction and microglia activation. These neuroprotective effects may be related to the increased levels of colonic glucagon-like peptide-1 (GLP-1) and cerebral GLP-1 receptor, eventually restoring gut microbiota homeostasis [62]. Moreover, the anti-depressive effects of C. butyricum in chronic unpredictable mild stress-induced depressive-like behavior may be resulted from the stimulation of intestinal GLP-1 secretion [63]. In AD models, administration of C. butyricum for 4 weeks prevents cognitive impairment, Aβ deposits, and neuroinflammation, which is mediated by the restoration of gut microbiota and butyrate (Figure 2) [64]. In vascular dementia mice, C. butyricum significantly alleviated the cognitive dysfunction and histopathological changes via anti-apoptotic properties and the consequent activation of PI3K/Akt pathway [65]. Treatment with C. butyricum defends against cerebral ischemia/reperfusion injury through anti-oxidant and anti-apoptotic mechanisms, which may be partially attributed to the increased butyrate contents in the brain [66]. Consistently, C. butyricum treatment has been shown to improve the neurological dysfunction and neurodegeneration in a mouse model of traumatic brain injury [67]. Although the neuroprotective effects of probiotic C. butyricum appears well-established, additional human randomized controlled trials would further provide valuable clinical data related to the strains’ utility as an intervention in the neurodegenerative diseases.
6.2. Akkermansia muciniphila
Akkermansia muciniphila, a gram-negative, anaerobic bacterium first identified in 2004 [68], is considered as a promising candidate as a “next-generation probiotics” [69]. The benefits of A. muciniphila are not limited to protecting the mucosal barrier integrity, improving the host metabolic functions and immune responses, A. muciniphila also possesses a value in modulating brain function. In this case, the critical role of A. muciniphila has been demonstrated in neurodegenerative diseases. Several studies have consistently reported that Akkermansia is highly effective in distinguishing PD or serves as a potential early biomarker for PD diagnosis [24,27,70]. It is noteworthy that the abundance of A. muciniphila is also found to be increased in PD patients [71]. One of the possible explanations is that the compensation of increased A. muciniphila in PD aims to fight and prevent disease progression. Although treatment with A. muciniphila has been reported to improve cognitive deficits and reduce Aβ levels in AD mice model [72], whether A. muciniphila can alleviate the neurodegeneration in PD patients remains unknown (Figure 2). There are also opposite results demonstrating that A. muciniphila-conditioned medium can initiate α-synuclein aggregation in enteroendocrine cells [73]. To date, few studies have explored the direct impact of A. muciniphila on the nervous system, the likelihood of beneficial effects exerted by A. muciniphila should be addressed. Further characterization of the relevance for the neurodegeneration will be fundamental to unveil the consequences of A. muciniphila dysbiosis.
6.3. Faecalibacterium prausnitzii
Faecalibacterium prausnitzii, an anaerobic Gram-positive bacterium, belongs to the Firmicutes phylum and the Ruminococcaceae family, also known as Clostridium cluster IV. The interest in F. prausnitzii is related to its capacity to produce beneficial metabolites, such as fructose, formic acid, and d-lactate and it is the most important butyrate-producing bacteria [74]. In addition, a 15 kDa protein with anti-inflammatory properties produced by F. prausnitzii can inhibit the nuclear factor-κB (NF-κB) pathway in intestinal epithelial cells and prevent colitis in animal models [75]. Consistently, several other studies in mice also clarify a protective role of F. prausnitzii in experimentally induced colitis. Intragastric administration of either F. prausnitzii or its culture supernatant can significantly decrease the severity of colitis by down-regulating pro-inflammatory cytokines [76]. Butyrate produced by F. prausnitzii modulates Th17/Treg balance and exerts anti-inflammatory effects in colorectal colitis rat model [77]. In terms of neurodegenerative diseases, the abundance of F. prausnitzii is decreased in the mild cognitive impairment (MCI) group compared with the healthy control, which is correlates with cognitive scores [78]. Two isolated F. prausnitzii strains from the healthy group have been shown to improve cognitive impairment in AD mice model [78]. So far, studies on the potential effects of F. prausnitzii on PD have not been reported (Figure 2). Additional researches are needed further prove the beneficial role of F. prausnitzii in the remediation of neurodegenerative diseases.
6.4. Bacteroides fragilis
Bacteroides fragilis is another promising probiotic, which is a commensal, Gram negative, non-spore forming obligatory anaerobic bacterium abundant in the human gastrointestinal tract. Traditionally, B. fragilis can interfere with other microbes via inhibiting their growth or translocation. As previously reported, B. fragilis treatment prevents Clostrioides difficile infection possibly by resisting pathogen colonization, enhancing the relative abundance of A. muciniphila and improving gut barrier integrity [79]. Indeed, B. fragilis can be classified into two subgroups: non-enterotoxigenic and enterotoxigenic B. fragilis [80]. Enterotoxin-containing B. fragilis secretes an unusually complex mixture of neurotoxins including the pro-inflammatory LPS. In this sense, enterotoxigenic B. fragilis exposure to human primary brain cells is an exceptionally potent inducer of the inflammatory pathway, driving pro-inflammatory degenerative neuropathology in the AD brain [81]. Conversely, non-enterotoxigenic B. fragilis strains exert the beneficial effects owing to an anti-inflammatory and immunomodulatory activity. Oral administration of B. fragilis has been found to increase gut microbiota diversity and beneficial commensal bacteria, thereby improving gut tight junction integrity and reducing inflammatory cytokines [82]. The fecal bacterial assessment by 16S rRNA amplicon sequencing shows that the abundance of B. fragilis is lower in PD patients than that of healthy control [83]. Considering that B. fragilis is one of the main hydrogen-producing intestinal bacteria, the decreased B. fragilis may be accounted for the lower amount of intestinal hydrogen level in PD (Figure 2) [84]. Although B. fragilis are potentially interesting as the next-generation probiotic, the role of B. fragilis in relation to the neuropathology is contrasting, thus the specific strains should be carefully evaluated for the safety and efficacy in the neurodegenerative diseases.
7. Perspectives and conclusions
Mounting information presents compelling evidence that gastrointestinal dysfunction serves as a prodromal symptom preceding the clinical manifestations of neurodegenerative diseases. Lack of understanding the mechanisms and pathophysiology of gastrointestinal manifestations hampers the diagnosis and clinical treatment. The GM, gastrointestinal tract and brain have historically been studied independently, but a growing appreciation for their interaction via gut-brain axis advocates the idea of targeting GM as a strategy for clinical intervention. Currently, there is a growing interest toward next-generation probiotics as the potential therapeutic agents by altering GM. Therefore, the safety and tolerability of these novel probiotics need to be validated in both animal models and human trials, so as to develop a personalized application. Another challenge is the storage due to the strict anaerobic conditions during microbial collection and freeze-drying. Even so, the current state of the next-generation probiotics remains largely promising in the context of neurodegenerative diseases, for the purpose of slowing down or preventing the neurodegeneration, as well as developing effective therapeutic interventions.
Author Contributions
Concepted and wrote the manuscript, M.B.; conducted the data analysis, C.L. and Y.W.; created and edited the figures, M.B.; X conceived and edited the final work, M.B. All authors have read and approved the final version of the manuscript.
Funding
This work was supported by the National Key Research and Development Program of China (2022YFA1304200, 2021YFA0717002), National Natural Science Foundation of China (32270113), and Shandong Province Natural Science Foundation (ZR2020QH125).
Conflicts of Interest
All authors declare that they have no conflict of interest.
Abbreviations
Aβ: β-amyloid; AD: Alzheimer’s disease; ASD: autism spectrum disorder; BBB: blood-brain barrier; BMAA: β-N-methylamino-L-alanine; CNS: central nervous system; ENS: enteric nervous system; GLP-1: glucagon-like peptide-1; GM: gut microbiome; GPCR: G protein-coupled receptor; HDAC: histone deacetylase; LPS: lipopolysaccharide; MCI: mild cognitive impairment; NF-κB: nuclear factor-κB; PAMP: pathogen-associated molecular pattern; PD: Parkinson’s disease; SCFAs: short-chain fatty acids; TRANK1: tetratricopeptide repeat and ankyrin repeat containing 1.
References
- Bloem, B.R.; Okun, M.S. , Klein, C., Parkinson’s disease. Lancet 2021, 397, 2284–2303. [Google Scholar] [CrossRef] [PubMed]
- Knopman, D.S.; Amieva, H.; Petersen, R.C.; Chetelat, G.; Holtzman, D.M.; Hyman, B.T.; Nixon, R.A. , Jones, D.T., Alzheimer disease. Nat Rev Dis Primers 2021, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, C.; Antonioli, L.; Colucci, R.; Blandizzi, C. , Fornai, M., Interplay among gut microbiota, intestinal mucosal barrier and enteric neuro-immune system: a common path to neurodegenerative diseases? Acta Neuropathol 2018, 136, 345–361. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, U. and Kayed, R., Amyloid beta, Tau, and alpha-Synuclein aggregates in the pathogenesis, prognosis, and therapeutics for neurodegenerative diseases. Prog Neurobiol 2022, 214, 102270. [Google Scholar] [CrossRef]
- Anis, E.; Xie, A.; Brundin, L. , Brundin, P., Digesting recent findings: gut alpha-synuclein, microbiome changes in Parkinson’s disease. Trends Endocrinol Metab 2022, 33, 147–157. [Google Scholar] [CrossRef]
- Travagli, R.A.; Browning, K.N. , Camilleri, M., Parkinson disease and the gut: new insights into pathogenesis and clinical relevance. Nat Rev Gastroenterol Hepatol 2020, 17, 673–685. [Google Scholar] [CrossRef]
- Jeremic, D.; Jimenez-Diaz, L. , Navarro-Lopez, J.D., Past, present and future of therapeutic strategies against amyloid-beta peptides in Alzheimer’s disease: a systematic review. Ageing Res Rev 2021, 72, 101496. [Google Scholar] [CrossRef]
- Holmqvist, S.; Chutna, O.; Bousset, L.; Aldrin-Kirk, P.; Li, W.; Bjorklund, T.; Wang, Z.Y.; Roybon, L.; Melki, R. , Li, J.Y., Direct evidence of Parkinson pathology spread from the gastrointestinal tract to the brain in rats. Acta Neuropathol 2014, 128, 805–20. [Google Scholar] [CrossRef]
- Ahn, E.H.; Kang, S.S.; Liu, X.; Chen, G.; Zhang, Z.; Chandrasekharan, B.; Alam, A.M.; Neish, A.S.; Cao, X. , Ye, K., Initiation of Parkinson’s disease from gut to brain by delta-secretase. Cell Res 2020, 30, 70–87. [Google Scholar] [CrossRef]
- Jones, L.; Kumar, J.; Mistry, A.; Sankar Chittoor Mana, T.; Perry, G.; Reddy, V.P. , Obrenovich, M., The Transformative Possibilities of the Microbiota and Mycobiota for Health, Disease, Aging, and Technological Innovation. Biomedicines 2019, 7. [Google Scholar] [CrossRef]
- Aho, V.T.E.; Pereira, P.A.B.; Voutilainen, S.; Paulin, L.; Pekkonen, E.; Auvinen, P. , Scheperjans, F., Gut microbiota in Parkinson’s disease: Temporal stability and relations to disease progression. EBioMedicine 2019, 44, 691–707. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Yang, X.; Xu, S.; Huang, P.; Li, B.; Du, J.; He, Y.; Su, B.; Xu, L.M.; Wang, L. , et al., Gut metagenomics-derived genes as potential biomarkers of Parkinson’s disease. Brain 2020, 143, 2474–2489. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.Q.; Shen, L.L.; Li, W.W.; Fu, X.; Zeng, F.; Gui, L.; Lu, Y.; Cai, M.; Zhu, C.; Tan, Y.L. , et al., Gut Microbiota is Altered in Patients with Alzheimer’s Disease. J Alzheimers Dis 2018, 63, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.; Zhu, M.; Yan, X.; Cheng, Y.; Shao, L.; Liu, X.; Jiang, R. , Wu, S., Structural and Functional Dysbiosis of Fecal Microbiota in Chinese Patients With Alzheimer’s Disease. Front Cell Dev Biol 2020, 8, 634069. [Google Scholar]
- Li, B.; He, Y.; Ma, J.; Huang, P.; Du, J.; Cao, L.; Wang, Y.; Xiao, Q.; Tang, H. , Chen, S., Mild cognitive impairment has similar alterations as Alzheimer’s disease in gut microbiota. Alzheimers Dement 2019, 15, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Vogt, N.M.; Kerby, R.L.; Dill-McFarland, K.A.; Harding, S.J.; Merluzzi, A.P.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Zetterberg, H.; Blennow, K. , et al., Gut microbiome alterations in Alzheimer’s disease. Sci Rep 2017, 7, 13537. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wu, L.; Peng, G.; Han, Y.; Tang, R.; Ge, J.; Zhang, L.; Jia, L.; Yue, S.; Zhou, K. , et al., Altered microbiomes distinguish Alzheimer’s disease from amnestic mild cognitive impairment and health in a Chinese cohort. Brain Behav Immun 2019, 80, 633–643. [Google Scholar] [CrossRef]
- Ren, T.; Gao, Y.; Qiu, Y.; Jiang, S.; Zhang, Q.; Zhang, J.; Wang, L.; Zhang, Y.; Wang, L. , Nie, K., Gut Microbiota Altered in Mild Cognitive Impairment Compared With Normal Cognition in Sporadic Parkinson’s Disease. Front Neurol 2020, 11, 137. [Google Scholar] [CrossRef]
- Li, W.; Wu, X.; Hu, X.; Wang, T.; Liang, S.; Duan, Y.; Jin, F. , Qin, B., Structural changes of gut microbiota in Parkinson’s disease and its correlation with clinical features. Sci China Life Sci 2017, 60, 1223–1233. [Google Scholar] [CrossRef]
- Li, F.; Wang, P.; Chen, Z.; Sui, X.; Xie, X. , Zhang, J., Alteration of the fecal microbiota in North-Eastern Han Chinese population with sporadic Parkinson’s disease. Neurosci Lett 2019, 707, 134297. [Google Scholar] [CrossRef]
- Petrov, V.A.; Saltykova, I.V.; Zhukova, I.A.; Alifirova, V.M.; Zhukova, N.G.; Dorofeeva, Y.B.; Tyakht, A.V.; Kovarsky, B.A.; Alekseev, D.G.; Kostryukova, E.S. , et al., Analysis of Gut Microbiota in Patients with Parkinson’s Disease. Bull Exp Biol Med 2017, 162, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Unger, M.M.; Spiegel, J.; Dillmann, K.U.; Grundmann, D.; Philippeit, H.; Burmann, J.; Fassbender, K.; Schwiertz, A. , Schafer, K.H., Short chain fatty acids and gut microbiota differ between patients with Parkinson’s disease and age-matched controls. Parkinsonism Relat Disord 2016, 32, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Hopfner, F.; Kunstner, A.; Muller, S.H.; Kunzel, S.; Zeuner, K.E.; Margraf, N.G.; Deuschl, G.; Baines, J.F. , Kuhlenbaumer, G., Gut microbiota in Parkinson disease in a northern German cohort. Brain Res 2017, 1667, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Hill-Burns, E.M.; Debelius, J.W.; Morton, J.T.; Wissemann, W.T.; Lewis, M.R.; Wallen, Z.D.; Peddada, S.D.; Factor, S.A.; Molho, E.; Zabetian, C.P. , et al., Parkinson’s disease and Parkinson’s disease medications have distinct signatures of the gut microbiome. Mov Disord 2017, 32, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Zheng, W.; He, Y.; Tang, W.; Wei, X.; He, R.; Huang, W.; Su, Y.; Huang, Y.; Zhou, H. , et al., Gut microbiota in patients with Parkinson’s disease in southern China. Parkinsonism Relat Disord 2018, 53, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Yang, X.; Xu, S.; Wu, C.; Song, Y.; Qin, N.; Chen, S.D. , Xiao, Q., Alteration of the fecal microbiota in Chinese patients with Parkinson’s disease. Brain Behav Immun 2018, 70, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Martinez, G.; Chin, B.; Camarillo, C.; Herrera, G.V.; Yang, B.; Sarosiek, I. , Perez, R.G., A Pilot Microbiota Study in Parkinson’s Disease Patients versus Control Subjects, and Effects of FTY720 and FTY720-Mitoxy Therapies in Parkinsonian and Multiple System Atrophy Mouse Models. J Parkinsons Dis 2020, 10, 185–192. [Google Scholar] [CrossRef]
- Keshavarzian, A.; Green, S.J.; Engen, P.A.; Voigt, R.M.; Naqib, A.; Forsyth, C.B.; Mutlu, E. , Shannon, K.M., Colonic bacterial composition in Parkinson’s disease. Mov Disord 2015, 30, 1351–60. [Google Scholar] [CrossRef]
- Needham, B.D.; Kaddurah-Daouk, R. , Mazmanian, S.K., Gut microbial molecules in behavioural and neurodegenerative conditions. Nat Rev Neurosci 2020, 21, 717–731. [Google Scholar] [CrossRef]
- Sgritta, M.; Dooling, S.W.; Buffington, S.A.; Momin, E.N.; Francis, M.B.; Britton, R.A. , Costa-Mattioli, M., Mechanisms Underlying Microbial-Mediated Changes in Social Behavior in Mouse Models of Autism Spectrum Disorder. Neuron 2019, 101, 246–259. [Google Scholar] [CrossRef]
- Jaglin, M.; Rhimi, M.; Philippe, C.; Pons, N.; Bruneau, A.; Goustard, B.; Dauge, V.; Maguin, E.; Naudon, L. , Rabot, S., Indole, a Signaling Molecule Produced by the Gut Microbiota, Negatively Impacts Emotional Behaviors in Rats. Front Neurosci 2018, 12, 216. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.J.; Kim, J.; Lee, H.J.; Ryu, H.S.; Kim, K.; Lee, J.H.; Jung, K.W.; Kim, M.J.; Kim, M.J.; Kim, Y.J. , et al., Alpha-synuclein in gastric and colonic mucosa in Parkinson’s disease: Limited role as a biomarker. Mov Disord 2016, 31, 241–9. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, E.; Murphy, S. , Martinson, H.A., Alpha-Synuclein Pathology and the Role of the Microbiota in Parkinson’s Disease. Front Neurosci 2019, 13, 369. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K. , Hsiao, E.Y., Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–76. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Grasset, E.; Manneras Holm, L.; Karsenty, G.; Macpherson, A.J.; Olofsson, L.E. , Backhed, F., Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proc Natl Acad Sci U S A 2018, 115, 6458–6463. [Google Scholar] [CrossRef] [PubMed]
- Israelyan, N.; Del Colle, A.; Li, Z.; Park, Y.; Xing, A.; Jacobsen, J.P.R.; Luna, R.A.; Jensen, D.D.; Madra, M.; Saurman, V. , et al., Effects of Serotonin and Slow-Release 5-Hydroxytryptophan on Gastrointestinal Motility in a Mouse Model of Depression. Gastroenterology 2019, 157, 507–521. [Google Scholar] [CrossRef] [PubMed]
- Kibbie, J.J.; Dillon, S.M.; Thompson, T.A.; Purba, C.M.; McCarter, M.D. , Wilson, C.C., Butyrate directly decreases human gut lamina propria CD4 T cell function through histone deacetylase (HDAC) inhibition and GPR43 signaling. Immunobiology 2021, 226, 152126. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Xu, B.; Wang, X.; Wan, W.H.; Lu, J.; Kong, D.; Jin, Y.; You, W.; Sun, H.; Mu, X. , et al., Gut microbiota-derived short-chain fatty acids regulate group 3 innate lymphoid cells in HCC. Hepatology 2022. [Google Scholar]
- Romero, E.; Ali, C.; Molina-Holgado, E.; Castellano, B.; Guaza, C. , Borrell, J., Neurobehavioral and immunological consequences of prenatal immune activation in rats. Influence of antipsychotics. Neuropsychopharmacology 2007, 32, 1791–804. [Google Scholar] [CrossRef]
- Lai, J.; Zhang, P.; Jiang, J.; Mou, T.; Li, Y.; Xi, C.; Wu, L.; Gao, X.; Zhang, D.; Chen, Y. , et al., New Evidence of Gut Microbiota Involvement in the Neuropathogenesis of Bipolar Depression by TRANK1 Modulation: Joint Clinical and Animal Data. Front Immunol 2021, 12, 789647. [Google Scholar] [CrossRef]
- Bi, M.; Kang, S.; Du, X.; Jiao, Q. , Jiang, H., Association between SNCA rs356220 polymorphism and Parkinson’s disease: A meta-analysis. Neurosci Lett 2020, 717, 134703. [Google Scholar] [CrossRef] [PubMed]
- Wallen, Z.D.; Stone, W.J.; Factor, S.A.; Molho, E.; Zabetian, C.P.; Standaert, D.G. , Payami, H., Exploring human-genome gut-microbiome interaction in Parkinson’s disease. NPJ Parkinsons Dis 2021, 7, 74. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, L.; Zhao, N.; Xu, X.; Xu, Y. , Zhu, B., Of genes and microbes: solving the intricacies in host genomes. Protein Cell 2018, 9, 446–461. [Google Scholar] [CrossRef] [PubMed]
- Franco-Obregon, A. and Gilbert, J.A., The Microbiome-Mitochondrion Connection: Common Ancestries, Common Mechanisms, Common Goals. mSystems 2017, 2. [Google Scholar]
- Tian, D. and Han, M., Bacterial peptidoglycan muropeptides benefit mitochondrial homeostasis and animal physiology by acting as ATP synthase agonists. Dev Cell 2022, 57, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Matheoud, D.; Cannon, T.; Voisin, A.; Penttinen, A.M.; Ramet, L.; Fahmy, A.M.; Ducrot, C.; Laplante, A.; Bourque, M.J.; Zhu, L. , et al., Intestinal infection triggers Parkinson’s disease-like symptoms in Pink1(-/-) mice. Nature 2019, 571, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Cox, P.A.; Banack, S.A.; Steele, J.C. , Sacks, O.W., Occurrence of beta-methylamino-l-alanine (BMAA) in ALS/PDC patients from Guam. Acta Neurol Scand 2004, 110, 267–9. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.F.; Candeias, E.; Esteves, A.R.; Magalhaes, J.D.; Ferreira, I.L.; Nunes-Costa, D.; Rego, A.C.; Empadinhas, N. , Cardoso, S.M., Microbial BMAA elicits mitochondrial dysfunction, innate immunity activation, and Alzheimer’s disease features in cortical neurons. J Neuroinflammation 2020, 17, 332. [Google Scholar] [CrossRef]
- Esteves, A.R.; Munoz-Pinto, M.F.; Nunes-Costa, D.; Candeias, E.; Silva, D.F.; Magalhaes, J.D.; Pereira-Santos, A.R.; Ferreira, I.L.; Alarico, S.; Tiago, I. , et al., Footprints of a microbial toxin from the gut microbiome to mesencephalic mitochondria. Gut 2021. [Google Scholar]
- Nunes-Costa, D.; Magalhaes, J.D.; M, G.F.; Cardoso, S.M. , Empadinhas, N., Microbial BMAA and the Pathway for Parkinson’s Disease Neurodegeneration. Front Aging Neurosci 2020, 12, 26. [Google Scholar] [CrossRef]
- Kabat, A.M.; Pott, J. , Maloy, K.J., The Mucosal Immune System and Its Regulation by Autophagy. Front Immunol 2016, 7, 240. [Google Scholar] [CrossRef] [PubMed]
- Deretic, V.; Kimura, T.; Timmins, G.; Moseley, P.; Chauhan, S. , Mandell, M., Immunologic manifestations of autophagy. J Clin Invest 2015, 125, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Nighot, P.K.; Hu, C.A. , Ma, T.Y., Autophagy enhances intestinal epithelial tight junction barrier function by targeting claudin-2 protein degradation. J Biol Chem 2015, 290, 7234–46. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, K.; Nishitani, M.; Takakura, A.; Imai, Y.; Komatsu, M. , Kawashima, H., Autophagy Protects against Colitis by the Maintenance of Normal Gut Microflora and Secretion of Mucus. J Biol Chem 2015, 290, 20511–26. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, C.; Zhao, W.; He, C.; Ding, J.; Dai, R.; Xu, K.; Xiao, L.; Luo, L.; Liu, S. , et al., Impaired Autophagy in Intestinal Epithelial Cells Alters Gut Microbiota and Host Immune Responses. Appl Environ Microbiol 2018, 84. [Google Scholar] [CrossRef] [PubMed]
- Stoeva, M.K.; Garcia-So, J.; Justice, N.; Myers, J.; Tyagi, S.; Nemchek, M.; McMurdie, P.J.; Kolterman, O. , Eid, J., Butyrate-producing human gut symbiont, Clostridium butyricum, and its role in health and disease. Gut Microbes 2021, 13, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Ariyoshi, T.; Hagihara, M.; Takahashi, M. , Mikamo, H., Effect of Clostridium butyricum on Gastrointestinal Infections. Biomedicines 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Hagihara, M.; Ariyoshi, T.; Kuroki, Y.; Eguchi, S.; Higashi, S.; Mori, T.; Nonogaki, T.; Iwasaki, K.; Yamashita, M.; Asai, N. , et al., Clostridium butyricum enhances colonization resistance against Clostridioides difficile by metabolic and immune modulation. Sci Rep 2021, 11, 15007. [Google Scholar] [CrossRef]
- Imase, K.; Takahashi, M.; Tanaka, A.; Tokunaga, K.; Sugano, H.; Tanaka, M.; Ishida, H.; Kamiya, S. , Takahashi, S., Efficacy of Clostridium butyricum preparation concomitantly with Helicobacter pylori eradication therapy in relation to changes in the intestinal microbiota. Microbiol Immunol 2008, 52, 156–61. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, J.; Ju, Z.; Wu, J.; Wang, L.; Lin, H. , Sun, S., Clostridium butyricum Ameliorates Salmonella Enteritis Induced Inflammation by Enhancing and Improving Immunity of the Intestinal Epithelial Barrier at the Intestinal Mucosal Level. Front Microbiol 2020, 11, 299. [Google Scholar]
- Ariyoshi, T.; Hagihara, M.; Eguchi, S.; Fukuda, A.; Iwasaki, K.; Oka, K.; Takahashi, M.; Yamagishi, Y. , Mikamo, H., Clostridium butyricum MIYAIRI 588-Induced Protectin D1 Has an Anti-inflammatory Effect on Antibiotic-Induced Intestinal Disorder. Front Microbiol 2020, 11, 587725. [Google Scholar] [CrossRef]
- Sun, J.; Li, H.; Jin, Y.; Yu, J.; Mao, S.; Su, K.P.; Ling, Z. , Liu, J., Probiotic Clostridium butyricum ameliorated motor deficits in a mouse model of Parkinson’s disease via gut microbiota-GLP-1 pathway. Brain Behav Immun 2021, 91, 703–715. [Google Scholar] [CrossRef]
- Sun, J.; Wang, F.; Hu, X.; Yang, C.; Xu, H.; Yao, Y. , Liu, J., Clostridium butyricum Attenuates Chronic Unpredictable Mild Stress-Induced Depressive-Like Behavior in Mice via the Gut-Brain Axis. J Agric Food Chem 2018, 66, 8415–8421. [Google Scholar] [CrossRef]
- Sun, J.; Xu, J.; Yang, B.; Chen, K.; Kong, Y.; Fang, N.; Gong, T.; Wang, F.; Ling, Z. , Liu, J., Effect of Clostridium butyricum against Microglia-Mediated Neuroinflammation in Alzheimer’s Disease via Regulating Gut Microbiota and Metabolites Butyrate. Mol Nutr Food Res 2020, 64, e1900636. [Google Scholar] [CrossRef]
- Liu, J.; Sun, J.; Wang, F.; Yu, X.; Ling, Z.; Li, H.; Zhang, H.; Jin, J.; Chen, W.; Pang, M. , et al., Neuroprotective Effects of Clostridium butyricum against Vascular Dementia in Mice via Metabolic Butyrate. Biomed Res Int 2015, 2015, 412946. [Google Scholar] [CrossRef]
- Sun, J.; Ling, Z.; Wang, F.; Chen, W.; Li, H.; Jin, J.; Zhang, H.; Pang, M.; Yu, J. , Liu, J., Clostridium butyricum pretreatment attenuates cerebral ischemia/reperfusion injury in mice via anti-oxidation and anti-apoptosis. Neurosci Lett 2016, 613, 30–5. [Google Scholar] [CrossRef]
- Li, H.; Sun, J.; Du, J.; Wang, F.; Fang, R.; Yu, C.; Xiong, J.; Chen, W.; Lu, Z. , Liu, J., Clostridium butyricum exerts a neuroprotective effect in a mouse model of traumatic brain injury via the gut-brain axis. Neurogastroenterol Motil 2018, 30, e13260. [Google Scholar] [CrossRef]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M. , de Vos, W.M., Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int J Syst Evol Microbiol 2004, 54, 1469–1476. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Q.; Cheng, L.; Buch, H. , Zhang, F., Akkermansia muciniphila is a promising probiotic. Microb Biotechnol 2019, 12, 1109–1125. [Google Scholar] [CrossRef]
- Heintz-Buschart, A.; Pandey, U.; Wicke, T.; Sixel-Doring, F.; Janzen, A.; Sittig-Wiegand, E.; Trenkwalder, C.; Oertel, W.H.; Mollenhauer, B. , Wilmes, P., The nasal and gut microbiome in Parkinson’s disease and idiopathic rapid eye movement sleep behavior disorder. Mov Disord 2018, 33, 88–98. [Google Scholar] [CrossRef]
- Bedarf, J.R.; Hildebrand, F.; Coelho, L.P.; Sunagawa, S.; Bahram, M.; Goeser, F.; Bork, P. , Wullner, U., Functional implications of microbial and viral gut metagenome changes in early stage L-DOPA-naive Parkinson’s disease patients. Genome Med 2017, 9, 39. [Google Scholar] [CrossRef]
- Ou, Z.; Deng, L.; Lu, Z.; Wu, F.; Liu, W.; Huang, D. , Peng, Y., Protective effects of Akkermansia muciniphila on cognitive deficits and amyloid pathology in a mouse model of Alzheimer’s disease. Nutr Diabetes 2020, 10, 12. [Google Scholar] [CrossRef]
- Amorim Neto, D.P.; Bosque, B.P.; Pereira de Godoy, J.V.; Rodrigues, P.V.; Meneses, D.D.; Tostes, K.; Costa Tonoli, C.C.; Faustino de Carvalho, H.; Gonzalez-Billault, C. , de Castro Fonseca, M., Akkermansia muciniphila induces mitochondrial calcium overload and alpha -synuclein aggregation in an enteroendocrine cell line. iScience 2022, 25, 103908. [Google Scholar] [CrossRef]
- Fagundes, R.R.; Bourgonje, A.R.; Saeed, A.; Vich Vila, A.; Plomp, N.; Blokzijl, T.; Sadaghian Sadabad, M.; von Martels, J.Z.H.; van Leeuwen, S.S.; Weersma, R.K. , et al., Inulin-grown Faecalibacterium prausnitzii cross-feeds fructose to the human intestinal epithelium. Gut Microbes 2021, 13, 1993582. [Google Scholar] [CrossRef]
- Quevrain, E.; Maubert, M.A.; Michon, C.; Chain, F.; Marquant, R.; Tailhades, J.; Miquel, S.; Carlier, L.; Bermudez-Humaran, L.G.; Pigneur, B. , et al., Identification of an anti-inflammatory protein from Faecalibacterium prausnitzii, a commensal bacterium deficient in Crohn’s disease. Gut 2016, 65, 415–425. [Google Scholar] [CrossRef]
- Martin, R.; Chain, F.; Miquel, S.; Lu, J.; Gratadoux, J.J.; Sokol, H.; Verdu, E.F.; Bercik, P.; Bermudez-Humaran, L.G. , Langella, P., The commensal bacterium Faecalibacterium prausnitzii is protective in DNBS-induced chronic moderate and severe colitis models. Inflamm Bowel Dis 2014, 20, 417–30. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, M.; Wang, Y.; Dorfman, R.G.; Liu, H.; Yu, T.; Chen, X.; Tang, D.; Xu, L.; Yin, Y. , et al., Faecalibacterium prausnitzii Produces Butyrate to Maintain Th17/Treg Balance and to Ameliorate Colorectal Colitis by Inhibiting Histone Deacetylase 1. Inflamm Bowel Dis 2018, 24, 1926–1940. [Google Scholar] [CrossRef]
- Ueda, A.; Shinkai, S.; Shiroma, H.; Taniguchi, Y.; Tsuchida, S.; Kariya, T.; Kawahara, T.; Kobayashi, Y.; Kohda, N.; Ushida, K. , et al., Identification of Faecalibacterium prausnitzii strains for gut microbiome-based intervention in Alzheimer’s-type dementia. Cell Rep Med 2021, 2, 100398. [Google Scholar] [CrossRef]
- Deng, H.; Yang, S.; Zhang, Y.; Qian, K.; Zhang, Z.; Liu, Y.; Wang, Y.; Bai, Y.; Fan, H.; Zhao, X. , et al., Bacteroides fragilis Prevents Clostridium difficile Infection in a Mouse Model by Restoring Gut Barrier and Microbiome Regulation. Front Microbiol 2018, 9, 2976. [Google Scholar] [CrossRef]
- De Filippis, F.; Esposito, A. , Ercolini, D., Outlook on next-generation probiotics from the human gut. Cell Mol Life Sci 2022, 79, 76. [Google Scholar] [CrossRef]
- Lukiw, W.J. , Bacteroides fragilis Lipopolysaccharide and Inflammatory Signaling in Alzheimer’s Disease. Front Microbiol 2016, 7, 1544. [Google Scholar] [CrossRef] [PubMed]
- Sofi, M.H.; Wu, Y.; Ticer, T.; Schutt, S.; Bastian, D.; Choi, H.J.; Tian, L.; Mealer, C.; Liu, C.; Westwater, C. , et al., A single strain of Bacteroides fragilis protects gut integrity and reduces GVHD. JCI Insight 2021, 6. [Google Scholar]
- Hasegawa, S.; Goto, S.; Tsuji, H.; Okuno, T.; Asahara, T.; Nomoto, K.; Shibata, A.; Fujisawa, Y.; Minato, T.; Okamoto, A. , et al., Intestinal Dysbiosis and Lowered Serum Lipopolysaccharide-Binding Protein in Parkinson’s Disease. PLoS One 2015, 10, e0142164. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Ito, M.; Hamaguchi, T.; Mori, H.; Takeda, Y.; Baba, R.; Watanabe, T.; Kurokawa, K.; Asakawa, S.; Hirayama, M. , et al., Quantification of hydrogen production by intestinal bacteria that are specifically dysregulated in Parkinson’s disease. PLoS One 2018, 13, e0208313. [Google Scholar] [CrossRef]
Figure 1.
Potential mechanisms of gut microbiome dysbiosis involvement in neurodegenerative diseases. AD is characterized by Aβ-containing extracellular plaques and PD is featured by intracellular α-synuclein accumulation to form Lewy bodies. The propagation of Aβ and α-synuclein in the gastrointestinal tract can be transmitted via vagus nerve to brain. In addition, host genetic susceptibility to the neurodegeneration and the GM dysbiosis are interacted, that is, Corynebacterium, Porphyromonas, and Prevotella interact with SNCA genetic variants rs356229, rs10029694, and rs6856813, respectively. Citrobacter rodentium and environmental microbial neurotoxin BMAA trigger the mitochondrial dysfunction, ultimately leading to the neurodegeneration. Furthermore, defective autophagy fails to eliminate the intracellular pathogens, and induces the alterations in the composition of gut microbiome.
Figure 1.
Potential mechanisms of gut microbiome dysbiosis involvement in neurodegenerative diseases. AD is characterized by Aβ-containing extracellular plaques and PD is featured by intracellular α-synuclein accumulation to form Lewy bodies. The propagation of Aβ and α-synuclein in the gastrointestinal tract can be transmitted via vagus nerve to brain. In addition, host genetic susceptibility to the neurodegeneration and the GM dysbiosis are interacted, that is, Corynebacterium, Porphyromonas, and Prevotella interact with SNCA genetic variants rs356229, rs10029694, and rs6856813, respectively. Citrobacter rodentium and environmental microbial neurotoxin BMAA trigger the mitochondrial dysfunction, ultimately leading to the neurodegeneration. Furthermore, defective autophagy fails to eliminate the intracellular pathogens, and induces the alterations in the composition of gut microbiome.

Figure 2.
The implications of next-generation of probiotics in neurodegenerative diseases.Clostridium butyricum, a butyrate-producing anaerobic bacterium, plays a protective role in the neurodegenerative diseases, which can prevent cognitive impairment and Aβ deposits in AD, and improve motor deficits, dopaminergic neuron loss, synaptic dysfunction and microglia activation in PD. Akkermansia muciniphila and Faecalibacterium prausnitzii are effective in alleviating the cognitive deficits and reducing Aβ levels in AD, however, the potential role in PD is unclear. Additionally, enterotoxigenic Bacteroides fragilis drives LPS-induced inflammation and degenerative neuropathology in AD, while non-enterotoxigenic B. fragilis exerts anti- inflammatory properties. Although B. fragilis is accounted for the amount of intestinal hydrogen level in PD, the exact role in the neurodegeneration of PD has not yet been elucidated.
Figure 2.
The implications of next-generation of probiotics in neurodegenerative diseases.Clostridium butyricum, a butyrate-producing anaerobic bacterium, plays a protective role in the neurodegenerative diseases, which can prevent cognitive impairment and Aβ deposits in AD, and improve motor deficits, dopaminergic neuron loss, synaptic dysfunction and microglia activation in PD. Akkermansia muciniphila and Faecalibacterium prausnitzii are effective in alleviating the cognitive deficits and reducing Aβ levels in AD, however, the potential role in PD is unclear. Additionally, enterotoxigenic Bacteroides fragilis drives LPS-induced inflammation and degenerative neuropathology in AD, while non-enterotoxigenic B. fragilis exerts anti- inflammatory properties. Although B. fragilis is accounted for the amount of intestinal hydrogen level in PD, the exact role in the neurodegeneration of PD has not yet been elucidated.
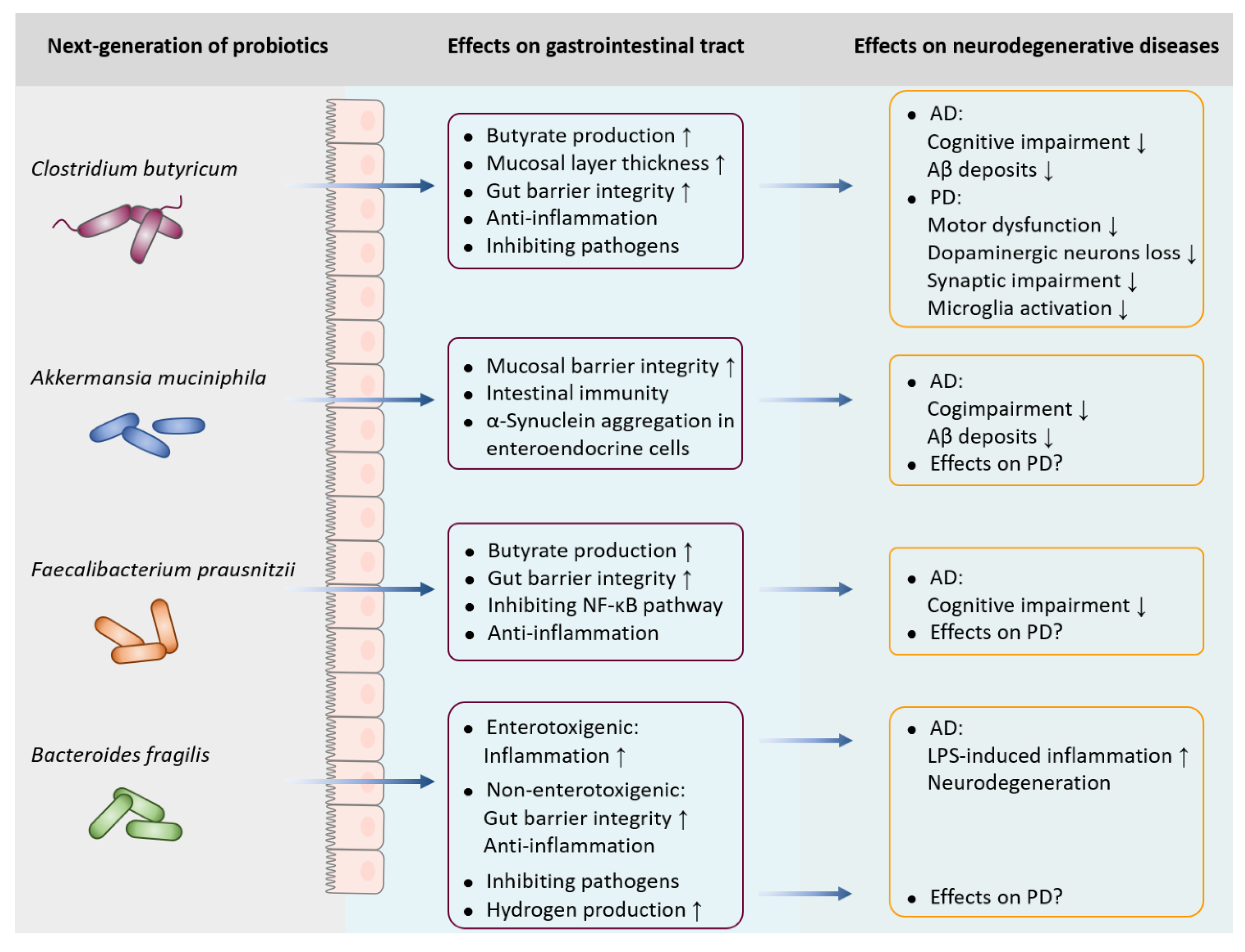

Table 1.
Gut microbiome alterations in the neurodegenerative diseases.
| Subjects | Gut microbiome alterations | |
|---|---|---|
| Increased | Decreased | |
| AD patients |
Ruminococcaceae, Enterococcus, Streptococcus, Alistipes, Dorea, Collinsella, Eggerthella |
Faecalibacterium, Lachnospira, Roseburia, Coprococcus |
| PD patients |
Alistipes, Streptococcus, Ruminococcus, Enterobacter, Enterococcus, Verrucomicrobium, Desulfovibrio, Anaetroncus |
Faecalibacterium, Prevotella, Blautia, Lachnospira, Roseburia |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



