
Submitted:
10 May 2023
Posted:
10 May 2023
You are already at the latest version
Abstract
This study aimed to investigate the ability of lactic acid bacteria to remove cholesterol in simulated gastric and intestinal fluids. The findings showed that the amount of cholesterol removed was dependent on the biomass, viability, and bacterial strain. Additionally, some cholesterol binding was stable and not released during gastrointestinal transit. The presence of cholesterol affected the fatty acid profile of bacterial cells, which could potentially influence their metabolism and functioning. However, adding cholesterol did not significantly impact the survival of lactic acid bacteria during gastrointestinal transit. Moreover, the study did not find significant effects of storage time, passage, or bacterial culture type on the cholesterol content of fermented dairy products. The researchers observed differences in cell survival among lactic acid bacteria strains in simulated gastric and intestinal fluids, depending on the environment. Furthermore, dry matter content, such as high milk protein, was more protective of bacterial cells during gastrointestinal transit than fat content. Future research should aim to better understand the impact of cholesterol on lactic acid bacteria metabolism and identify potential health benefits or risks.
Keywords:
lactic acid bacteria
; gastric juice
; intestinal juice
; fatty acid profile
; cell survival
; health benefits
1. Introduction
Changes in lifestyle and diet, as well as the development of civilization, have contributed to an increase in so-called “civilization diseases,” also referred to as chronic noncommunicable diseases. Ischemic heart disease, which is also known as coronary artery disease, is an example of a lifestyle disease, with the main cause being arteriosclerosis of the coronary arteries leading to their narrowing. Hypercholesterolemia, or elevated LDL cholesterol levels, is one of the risk factors for atherosclerosis. Elevated cholesterol has been identified as a crucial cardiovascular disease risk factor, and even a small reduction in cholesterol levels can reduce the risk of coronary artery disease [1,2,3,4].
Studies have shown that consuming fermented milk products, such as yogurt, can lower cholesterol levels in humans [5,6]. As early as 1974, Mann and Spoerry [7,8] found reduced serum cholesterol levels in African Masai tribe men, attributed to the consumption of large amounts of fermented milk containing wild strains of Lactobacillus acidophilus. Lactic acid bacteria (LAB) present in fermented milk drinks are responsible for lowering serum cholesterol levels. Some laboratory studies have also shown that traditional LAB used in the production of cheese, cream, butter, and sour milk can lower cholesterol. Much research is available in the literature on cholesterol-lowering in laboratory conditions using model media, with most studies involving lactobacilli [9,10,11,12,13,14,15,16]. Other types of bacteria that have shown similar properties are Bifidobacterium, Streptococcus, Enterococcus, Lactococcus, and Leuconostoc [9,11,17,18,19,20]. This type of research has been conducted since the 1970s [7,16,21,22,23,24,25,26]. However, it is important to note that despite the results of in vitro and in vivo animal and human studies, due to possible methodological and technical errors and lack of reproducibility, the ability of LAB to lower serum cholesterol levels cannot be confirmed or negated unequivocally [27,28,29,30,31,32]. Additionally, the serum cholesterol level not only correlates positively with the amount of cholesterol ingested with food but also depends on the intake of saturated fatty acids and refined carbohydrates. Therefore, there is still no clear confirmation of how LAB have a positive effect on cholesterol levels in humans [33,34,35].
Several hypotheses explain how LAB lower cholesterol levels. Information from the literature suggests that these mechanisms primarily include cholesterol binding, enzymatic deconjugation of bile salts, production of exopolysaccharides, and synthesis of short-chain fatty acids [11,17,19,20,33,34,36,37,38,39,40,41,42,43,44]. It has been found that LAB can bind cholesterol either through adhesion or assimilation [12,17,19,20,36,45,46,47]. Studies have also shown that LAB can hydrolyze (deconjugate) bile salts and bind to cholesterol molecules, thereby lowering cholesterol levels in the system [6,11,14,35,41,43,48,49,50,51,52,53,54]. Although the exopolysaccharides produced by many species of LAB can bind cholesterol molecules, this mechanism is still among the studied hypotheses [33,34,39,44,55,56].
Adhesion and assimilation of cholesterol by LAB can occur simultaneously when the bacterial cells are alive and biologically active. However, it is also possible that cholesterol only undergoes adhesion when the bacterial cells are dead. Understanding the factors that affect cholesterol binding across the cell wall or membrane will help determine which cholesterol-binding mechanism dominates under specific environmental conditions. The removal of cholesterol from the culture medium may indicate the possibility of cholesterol binding in the food product during fermentation or refrigerated storage. However, bound cholesterol may still be bioavailable for the human body. Therefore, separate experiments on binding, removal, and persistence of cholesterol binding in the human gastrointestinal tract should be performed to determine if the release of bound cholesterol occurs under these conditions. Such research can be easily conducted in vitro using classical culture media or conditions that simulate the human digestive system. These findings suggest that the positive effects of LAB on cholesterol levels can also occur under in vivo conditions [28,29,30,31,32].
The aim of this study was to investigate the ability of LAB cells to remove cholesterol from different ex vivo conditions and to examine the impact of selected factors on the quantity of cholesterol removed and the release of previously bound cholesterol.
2. Materials and Methods
2.1. Materials
The biological material used in this study consisted of four lactobacilli strains, namely Lactobacillus delbrueckii subsp. bulgaricus ATCC 11842, Lactobacillus helveticus LH-B01, Lactobacillus delbrueckii subsp. lactis ATCC 4797, and L. acidophilus La-5, which were deposited in the institute's collection. Before use, the strains were revived and propagated in MRS broth aliquots (at 37 ± 1°C for 18 h). Their biomass was then prepared at different cell concentrations: 10× concentrated biomass (10×), nonconcentrated biomass (1×), and 10× diluted biomass (0.1×). To obtain 10-fold concentrated biomass, a 17 h broth culture was centrifuged in an ultracentrifuge (model type 317a, Precision Mechanics Warsaw, Poland) for 7 min at 12,000×g at 4 ± 1°C. In contrast, 10-fold diluted biomass was obtained by diluting the initial 17 h broth culture (at 1-fold biomass concentration) 10-fold, followed by mixing on a micro shaker. The number of viable bacterial cells in the culture broth was determined using the deep plate method (MRS agar, 37 ± 1°C, 72 h, anaerobic conditions). Dead cell biomass was obtained by sterilizing (121°C/15 min) suspensions of live bacterial cells in the culture broth.
2.2. Media and Other Reagents
The MRS broth (basic) was prepared in the appropriate amount according to the manufacturer’s instructions (Merck, Darmstadt, Germany). A model MRS broth with added cholesterol was also prepared by measuring a suitable amount of dissolved sterile cholesterol solution into the basic sterile MRS broth to achieve the desired final concentration of cholesterol in the broth. MRS agar, used to determine lactobacilli cell counts, was prepared by dissolving powdered MRS agar medium (Merck, Darmstadt, Germany) in distilled water following the manufacturer's instructions.
To prepare the cholesterol solution, powdered cholesterol with >99% chemical purity (Sigma-Aldrich) was dissolved in a mixture of 99% ethanol and Tween 80 (Merck, Darmstadt, Germany) in a 3:1 ratio. The dissolution was performed by placing the bottle containing the prepared mixture of ethanol, Tween 80, and cholesterol in a hot water bath (approximately 95 ± 1°C). The cholesterol concentration in the resulting solution was about 40 g/L. The Cholésterol RTU® enzymatic cholesterol kit (BioMérieux) was used to determine the cholesterol concentrations in the culture samples.
The model gastric juice used in this study was taken from the publication by Clavel et al. [57]. To prepare the model gastric juice, 4.8 g NaCl (POCH), 1.56 g NaHCO3 (POCH), 2.2 g KCl (POCH), and 0.22 g CaCl2 (POCH) were dissolved in 1000 mL distilled water. The resulting solution was then adjusted to pH 2.4 ± 0.2 using a pH meter (model LPH330T, Grosseron, Tacussel Elektronique) and a 1 M HCl (POCH) solution. The model gastric juice was sterilized in portions of 50 mL in an autoclave at 121 ± 1°C for 15 min. In the experiments, a freeze-dried preparation of pepsin (Sigma-Aldrich) with a potency of 3200–4500 units/1 mg protein was used. Pepsin was added to the model gastric juice immediately before the experiments (2 mg crystalline pepsin to 50 mL model gastric juice).
The model intestinal juice used in this study was based on the publication by Marteau et al. [58], with some modifications. First, 2.5 g NaCl (POCH), 0.3 g KCl (POCH), 0.015 g CaCl2 (POCH), and 8.5 g bovine bile (Merck) were dissolved in 500 mL of the previously prepared 1 M NaHCO3 solution. The pH of the solution after sterilization was determined to be 7.0 ± 0.2 using a pH meter (model pHT 003, Eon Trading LLG, USA) and 1 M NaOH (p.a., POCH). The solution was then autoclaved at 121 ± 1°C for 15 min. A sterile solution of intestinal enzymes (750 mg pancreatin from bovine pancreas with an activity of 50,000 FIP units lipase, 40,000 FIP units amylase, and 3000 FIP units protease) was added to 160 mL portions of the model intestinal juice immediately before use.
2.3. Methods
2.3.1. Testing the Ability of Lactobacilli Cells to Remove Cholesterol from Model Gastric Juice or Model Intestinal Juice
To determine the degree of cholesterol removal, the biomasses of the strains were incubated in model juices containing added cholesterol solution. First, the biomasses were incubated in gastric juice (1:1) at 37°C for 3 h, followed by incubation in intestinal juice (1:1) at 37°C for 5 h. The cholesterol concentration in the culture liquid was determined using an enzymatic cholesterol determination kit at the beginning and end of each incubation. Separate experiments were performed using biomass from live and thermally inactivated bacterial cells. All chemicals used in this study were purchased from Merck or Sigma-Aldrich Co. (St. Louis, MO, USA) with analytical grades.
2.3.2. Testing the Ability of Lactobacilli Cells to Release Previously Bound Cholesterol under Conditions of Model Gastric Juice and Model Intestinal Juice
To allow for initial cholesterol binding, culture cell biomasses were cultured in MRS broth containing added cholesterol at 37°C for 24 h. After culture, the cholesterol concentration in the postculture liquid was determined using an enzymatic cholesterol determination kit, and the biomasses were centrifuged in an ultracentrifuge (model type 317a, Mechanika Precyzyjna, Warsaw, Poland) for 7 min at 12,000×g at 4 ± 1°C. The resulting biomasses were then incubated in model juices containing no added cholesterol solution. First, the biomasses were incubated in gastric juice (1:1) at 37°C for 3 h, followed by incubation in intestinal juice (1:1) at 37°C for 5 h. The cholesterol concentration in the culture liquid was determined using an enzymatic cholesterol determination kit at the beginning and end of each incubation. All reagents used in the study were of analytical purity. Separate experiments were performed using biomass from live and thermally inactivated bacterial cells. All chemicals used in this study were purchased from Merck or Sigma-Aldrich Co. (St. Louis, MO, USA) with analytical grades.
2.3.3. Testing the Ability of Lactobacilli Cells to Remove Cholesterol from Model Dairy Products under Conditions of Model Gastric Juice and Model Intestinal Juice
Model dairy products were prepared using fresh skim milk (0.05% fat) with the addition of cream (12% fat) or skimmed milk powder to adjust the fat and dry matter content (DM). Five variants of model dairy products were prepared: (1) fresh milk with 0.05% fat and 12% DM; (2) fermented milk with 0.05% fat and 12% DM; (3) fermented milk with 0.05% fat and 20% DM; (4) fermented milk with 12% fat and 12% DM; and (5) fermented milk with 12% fat and 20% DM. The fermentation of samples of selected variants of model dairy products was carried out at 37 ± 1°C for 5 h. The finished products were then refrigerated at 6 ± 1°C and stored for 4 weeks. After this period, samples of the model dairy products were stored in model juices without added cholesterol solution, first in gastric juice (1:1, at 37°C for 3 h) and then in intestinal juice (1:1, 37°C for 5 h). Simultaneously, test strains were cultured in MRS broth with and without the addition of cholesterol solution. After completion of fermentation and cooling of the samples, after 1 and 4 weeks of refrigerated storage, and after the end of storage under the conditions of model juices (stomach and intestines), the cholesterol content in the samples was determined using a gas chromatograph model GC-MS QP 2010 (Shimadzu). The final cholesterol content results were converted to the starting amount of the model dairy products, considering the dilution levels of the samples at each stage of the experiments. The number of viable bacterial cells in the samples was determined using the plate depth method (MRS agar, 37 ± 1°C, 72 h, anaerobic conditions). All chemicals used in the study was of analytical purity and purchased from Merck or Sigma-Aldrich Co. (St. Louis, MO, USA).
2.3.4. Determination of Cholesterol Content in Model Dairy Products Using Gas Chromatography Technology Combined with Mass Spectrometry
Fat extraction from the samples was carried out through direct saponification [59]. In brief, 0.2 g of the sample was weighed into a tube and 5 mL of 0.5 M methanolic KOH solution was added. The tube was then capped and vortexed for 15 s. Subsequently, the tube was heated in an 80°C water bath for 15 min with vortexing every 5 min for 10 s. After heating, the sample was cooled, and 1 mL of water and 5 mL of n-hexane (Merck) were added. Finally, the sample was vigorously vortexed for 1 min and centrifuged at 2000×g for 1 min. An aliquot of the upper phase was then transferred to an autosampler vial and analyzed by GC-MS QP 2010 (Shimadzu). The gas chromatograph operated under the following conditions: a DB type column measuring 30 m/0.25 m/0.25 mm; the initial temperature of 50°C with an isotherm of 2 min, temperature increased by 25°C/min to 270°C with an isotherm of 1 min, temperature ramp of 5°C/min to an end temperature of 320°C with an isotherm of 10 min, and a total analysis duration of 31.80 min. The retention time for cholesterol was about 18.2 min, and for 5-cholestane, it was 15.7 min. The dispenser temperature was 260°C, with injection in split mode, a split ratio of 1:30, column gas flow of 0.98 mL/min, and a pressure of 52.3 kPa. The mass spectrometer operated under the following conditions: ion source temperature of 220°C, the temperature of the connecting line between GC and MS 240°C, the voltage at the detector of 1.13 kV, ionization energy of 70 eV, and quadrupole filter sweep range of 100–600 m/z. All reagents used in this study was of analytical purity. For the determination of cholesterol in samples, 5α-cholesterol (Sigma-Aldrich) was used as an internal standard. All chemicals used in this study were purchased from Merck or Sigma-Aldrich Co. (St. Louis, MO, USA) with HPLC grades.
2.3.5. Determination of the Fatty Acid Profile of Lactobacilli Cells in MRS Broth with and without the Addition of Cholesterol Solution
The fatty acids were extracted using the method described in the references [60,61,62] and the gas chromatography coupled to a mass spectrometer was used for the chromatographic separation of the fatty acid methyl esters (GC-MS QP 2010 Shimadzu) with the 007-23-30-0.2F polar column (30 m/0.25 mm/0.20 m; Quadrex). The sample was injected at a split ratio of 1:25 with a dispenser temperature of 230°C. The chromatographic separation was carried out using the following conditions: initial column temperature of 60°C with a 2 min isotherm, temperature ramp of 4°C/min to 220°C, and a 10 min isotherm. The carrier gas used was helium with a flow rate of 0.37 mL/min. The detector conditions used were: ion source temperature of 200°C, temperature of the line connecting GC to MS of 220°C, detector voltage of 1.45 kV, and quadrupole filter sweep range of 50–400 m/z.
All reagents used in the study were of analytical purity, and standards such as oleic acid (Sigma-Aldrich, USA), anteiso12-methyltetradecanoic acid (Sigma-Aldrich, USA), 2-hydroxytetradecanoic acid (Sigma-Aldrich, USA), nonadecanoic acid (Sigma-Aldrich, USA), the bacterial acid methyl esters (BAME; Sigma-Aldrich, USA), GLC-674, and GLC-617 (Nu-Chek-Prep., USA) were used to identify fatty acids. Additionally, isomers of methyl esters of linoleic acid 18:2 (cis-9,trans-11 and trans-10,cis-12; Nu-Chek-Prep., USA) were also utilized. In cases where other fatty acids were identified, comparisons were made with literature data [63,64,65]. The GCMS Solution v.2.50 software was used for result analysis. All other chemicals used in this study were purchased from Merck or Sigma-Aldrich Co. (St. Louis, MO, USA) with analytical and HPLC grades.
2.4. Statistical Analysis
Each experiment was carried out in three independent replicates, and each analysis was performed in two replicates for each experiment, resulting in an average of six replicates per result. The data were subjected to a two-way analysis of variance, and mean differences between the statistical groups were tested at a significance level of α = 0.05 using Tukey’s test in the ANOVA method. Multivariate analysis was employed to describe the relationship of multiple variables for each sample at a significance level of α = 0.05. The Statgraphics Centurion XVII program (Statgraphics Technologies, Inc., The Plains, VA, USA) was used for statistical analysis of the results.
3. Results and Discussion
3.1. Testing the Ability of Lactobacilli Cells to Remove Cholesterol from Model Gastric Juice or Model Intestinal Juice
The experiments were conducted under in vitro conditions, and the results are presented in Figure 1a,c,e,g. The experimental setup selected facilitated the simultaneous examination of cholesterol removal during the transit of lactobacilli cells through a segment of model gastric juice and model intestinal juice. It was assumed that LAB do not metabolize cholesterol, and therefore, the loss of cholesterol from the culture broth should be directly proportional to the amount of cell biomass that binds and removes cholesterol. Consequently, the impact of bacterial biomass concentration and viability on the observed phenomena of cholesterol removal and release during the passage of lactobacilli cells through a section of model gastric juice and model intestinal juice was investigated.
The initial concentration of cholesterol in the cultures of the lactobacilli strains tested in the model gastric juice and model intestinal juice was 0.702 g/dm³. The findings revealed that the removal of cholesterol by LAB cells in the model gastric juice and model intestinal juice is significantly dependent on the biomass amount and viability of the bacterial cells and also varies depending on the bacterial strain under study. Among the tested strains, viable bacterial cells of L. helveticus strain LH-B01 exhibited the highest cholesterol removal in the model gastric juice, increasing the concentration 10-fold (average 0.043 g/dm3, Figure 1g).
Liong and Shah [40] noted that the number of cells significantly impacts the differences in the amount of cholesterol bound by LAB, whereas the growth dynamics of individual strains determine the amount of cell biomass and differences in experimental results. However, the absence of sufficient literature data in this area precludes a more detailed discussion of the results obtained in this study. Nonetheless, the study indicated that the initial concentration of bacterial cell biomass had a positive effect on the amount of cholesterol removed from the culture medium. However, it was expected that as the concentration of cell biomass decreased by a factor of 10, the amount of cholesterol removed from the culture broth would also decrease proportionally, by 10-fold). Interestingly, little difference was observed in the amount of cholesterol removed by biomasses with varying concentrations of living cells used. These results could provide clues to explain the hypocholesterolemic effect of products containing LAB, an area that has been widely investigated in the literature, but with contradicting findings [7,8,21,66,67,68,69].
For bacterial cells to take up cholesterol molecules, their high biological activity is necessary since, as demonstrated by Hosono and Tono-Oka [17] this phenomenon occurs most intensely during the logarithmic growth phase in lactic acid streptococci. Physical binding of cholesterol through the cell wall does not require cellular activity but only requires a sufficiently long contact time between the cells and the cholesterol molecules. Additionally, the same researchers found that not only living but also dead (autoclaved) cells of the tested strain could bind cholesterol. The fact that cholesterol removal occurs even when the bacterial cells are dead confirms that the physical binding of cholesterol molecules through the cell wall (adhesion) is one of the mechanisms of cholesterol removal by LAB cells [1,40,70,71,72]. These observations align with the experimental results presented in this study (Figure 1a,c,e,g).
3.2. Testing the Ability of Lactobacilli Cells to Release Previously Bound Cholesterol under Conditions of Model Gastric Juice and Model Intestinal Juice
In addition to the phenomenon of cholesterol binding and removal, it is also crucial to study whether bound cholesterol remains bioavailable to the human body. The experiments carried out in this section of the publication addressed this issue. The objective of the experiments was to investigate whether cholesterol previously bound by LAB cells is released under conditions of model gastric juice and model intestinal juice. The results of the experiments are illustrated in Figure 1b,d,f,h. Initially, the living and dead cell biomasses of the tested lactobacilli were cultivated in MRS broth with added cholesterol (average concentration of 0.647 g/dm3), and after 24 h, the cell biomasses were transferred to model gastric juice for 3 h, and then to model intestinal juice for 5 h.
The results of the experiments suggest that cholesterol previously removed and bound by cells of monocultures of LAB can be released. The experiments on the release of cholesterol by lactobacilli cells during their passage through a section of model gastric juice and model intestinal juice revealed statistically significant differences between the biomass of living and dead bacterial cells. Among the living lactobacilli cells, the strain L. acidophilus La-5 removed the most cholesterol from the initial amount of cholesterol in the MRS broth (0.677 g/dm3) with an average of 0.104 g/dm3 (Figure 1g). However, during the passage through a section of model gastric juice and model intestinal juice, the same strain released an average of 0.082 g/dm³ of cholesterol, which was statistically significantly less than what was removed from the MRS broth. In contrast, the dead cell biomass of the L. acidophilus La-5 strain removed the most cholesterol from the MRS broth among the lactobacilli strains tested, with an average of 0.056 g/dm3, and released an average of 0.044 g/dm3 of cholesterol during the passage through a section of model gastric juice and model intestinal juice. Statistical analyses showed that in this case, the amount of cholesterol released also depended on the viability of the bacterial cells. The dead cell biomass of the lactobacilli strains released less bound cholesterol than the living cell biomass, but they also removed less of it from the culture medium earlier.
The results obtained suggest that a small portion of cholesterol bound by LAB cells are so firmly bound that it is not released during the passage of lactobacilli cells through the section of model gastric juice and model intestinal juice. It can be assumed that the cholesterol bound in this manner is not bioavailable to the human body [1]. This finding could be consistent with studies by Albano et al. [1], Lee et al. [73], and Miremadi et al. [74] on structural changes in the bacterial cell wall. The results obtained in this study can be related to studies on the binding of aflatoxin B1 by LAB cells [73,75]. El-Nezami et al. [75] observed that the removal of aflatoxin B1 (AFB1) from the culture medium by selected cultures of LAB depended on their population and culture temperature. Lee et al. [73] also investigated the phenomenon of binding and release of AFB1 bound by living and dead cells of L. rhamnosus GG and L. rhamnosus LC-705, and found similar relationships to those observed in this study regarding the binding and removal of cholesterol by LAB cells. Lee et al. [73] also concluded that the thermal killing of the bacteria altered the bacterial cell surface and exposed additional binding sites for AFB1
Most studies on the effect of LAB on cholesterol have been conducted using classic culture broths, sometimes with the addition of bile salts [7,14,26,74,76]. However, there is no literature available on the effect of LAB cells on cholesterol binding under the conditions of model intestinal juice. It is important to note that the model intestinal juice used in the experiments contained bovine bile with both conjugated and deconjugated bile salts, meaning that the bile salt hydrolase activity produced by most intestinal strains of LAB was not required to precipitate cholesterol with free bile acids [26,35,41,42,43,48,51,52,77]. Coprecipitates of cholesterol with bile acids are known to form at low pH, below 5.5 [13,14,30,48,78]. Although the model intestinal juice used in the present study had a pH above 7.0, it is possible that the pH dropped to a level sufficient for the coprecipitation of cholesterol with free bile acids due to the addition of viable bacterial cells. This is possible in stationary cultures and is confirmed in the literature cited [14,26,35,41,42,43,49]. Such coprecipitates would rapidly dissolve under in vivo conditions if the pH rose above 5.5 [13,15,33,79], as the bile secreted by the liver travels to the duodenum where it neutralizes the acidic chyme leaving the stomach, and the pH in the small intestine is above 6.0. Therefore, the hypocholesterolemic effect caused by coprecipitation of cholesterol with deconjugated bile acids is unlikely to occur under in vivo conditions. However, this does not mean that lactobacilli are not beneficial in modifying other physiological parameters indicative of markers of metabolic syndrome, including obesity, hyperlipidemia, hyperglycemia, and insulin resistance, under in vivo conditions [28,29,30,80]. The hypocholesterolemic effect of some probiotics characterized by high BSH enzyme activity in vitro has also been confirmed in vivo in both humans and animals [7,31,32,81].
3.3. Testing the Ability of Lactobacilli Cells to Remove Cholesterol from Model Dairy Products under Conditions of Model Gastric Juice and Model Intestinal Juice
The previous experiments conducted in this study demonstrated that lactobacilli cells can effectively remove and bind cholesterol from both culture broth and model digestive juices. This process is contingent upon the number of bacterial cells present and their viability. It is worth noting that the consumption of fermented milk products involves the ingestion of a significant amount of LAB cell biomass.
The objective of the current experiment was to assess the cholesterol-removing ability of lactobacilli in model dairy products that vary in DM, fat content (0, 12, and 20%), and acidity (fresh milk and fermented milk). The products were subjected to 4 weeks of refrigerated storage and subpassage conditions in a section of model gastric juice and model intestinal juice. The results of this experiment are presented in Table 1.
The ability of specific lactobacilli cultures to lower cholesterol in culture broth has not been shown to have a direct correlation with the reduction of cholesterol levels in fermented milk products. In this study, it was discovered that the cholesterol content of the model dairy products was only statistically significant with the original total fat content of the samples. Neither the length of cold storage nor the passage of model gastric and intestinal juice had a statistically significant effect on the reduction of cholesterol levels. It is worth noting that the type of lactobacilli culture used did not have a statistically significant impact on the cholesterol content. This is to be expected since LAB cells are not capable of metabolizing cholesterol; instead, they can only bind to it through the cell wall and/or incorporate it into the cell wall or membrane simultaneously.
Although the exact mechanism of cholesterol binding by bacterial cells remains unclear, it is expected that the process of lipid extraction from a sample would extract all the cholesterol present in the sample. However, the results obtained in this portion of the study have yet to be confirmed in the literature. Aloglu and Öner [76] conducted research on Lactobacillus cultures to assess their cholesterol-removing abilities in culture media containing added bile salts and cholesterol (at 0.150 g/dm³) as well as cream. They observed a reduction in cholesterol levels by bacterial cells in both the culture broth and cream. The percentage of cholesterol removed from the cream ranged from 20.6 to 59.8% of its initial level, whereas the same cultures removed 12.1–47.5% of the initial cholesterol content from the culture medium.
3.4. Survival of Lactic Acid Bacteria Cells under Model Conditions of the Digestive System
The experiments discussed above suggest that lactobacilli cells have the ability to remove cholesterol from the culture medium and model food juices, as well as release already-bound cholesterol. It has been demonstrated that the amount of cholesterol removed or released is dependent on the quantity and viability of bacterial cell biomass. Therefore, this study aimed to investigate whether lactobacilli cells, suspended in model dairy products with varying DM, fat content (0, 12, and 20%), and acidity (fresh milk and fermented milk), could survive in an environment that simulates the conditions of the stomach or intestines. For comparison, the same conditions were applied to biomass samples of the tested lactobacilli, suspended in MRS broth with or without the addition of cholesterol. The results of these experiments are presented in Figure 2a–d.
The data presented in this study suggests that the lactobacilli cells used in these experiments were as active as in previous studies and exhibited good survival rates under refrigerated storage conditions of the model dairy samples. Before the passage of model gastric juice and model intestinal juice, the number of viable lactobacilli cells in all model milk product samples tested ranged from 7.2 log(CFU/mL) to 8.3 log(CFU/mL), regardless of the culture used. However, a 3-h incubation in model gastric juice resulted in a statistically significant reduction in the population of viable lactobacilli cells, depending on the type of sample in which the cells were suspended and the strain tested. L. helveticus strain LH-B01 suspended in MRS broth, fresh milk samples, or 12% fat fermented milk samples exhibited the poorest survival rates under these conditions (Figure 2b). For comparison, a 3-h incubation in model gastric juice most significantly reduced the population of live L. acidophilus La-5 cells (Figure 2d). In addition, a 5-h incubation in model intestinal juice resulted in the elimination of live lactobacilli cells suspended in MRS broth to below the limit of detection (<0.1 log(CFU/mL)), regardless of the strain tested. However, it is noteworthy that there was no statistically significant effect of cholesterol supplementation on the survival of lactobacilli in model gastric juice or model intestinal juice. When lactobacilli cells were suspended in model dairy products, better survival rates were observed, particularly for L. acidophilus La-5 cells (Figure 2d).
Research suggests that nonprobiotic bacterial strains exhibit lower cell survival rates in the model gastric juice when compared to probiotic strains [38,82]. Vinderola and Reinheimer [82] conducted a study on the tolerance of probiotic strains of LAB and bifidobacteria to model gastric juice and found that L. acidophilus cells were the most resistant to low pH levels in gastric juice. The good tolerance of bacterial cells to conditions in model digestive juices can be explained by the presence of these bacteria in the digestive tracts of humans or animals. Bacteria that are not natural gut flora lack natural resistance to intestinal juice conditions [83]. However, this is not conclusive since Elli et al. [84] demonstrated under in vivo conditions that some Streptococcus thermophilus can survive in the human gastrointestinal tract, despite not being a typical intestinal flora. In their study, a group of 20 volunteers were fed yogurt containing live cells of LAB. Researchers found live cells of S. thermophilus and L. delbrueckii subsp. Bulgaricus in the terminal gastrointestinal tract and feces. Therefore, it can be assumed that the survival of lactobacilli cells in the human gastrointestinal tract is dependent on the individual properties of the strains of these bacteria and is not a property of the entire species, which has also been confirmed by several other researchers [19,20,32,81].
It is important to note that the results of this study demonstrated statistically significant differences in cell survival rates among individual lactobacilli strains in model gastric juice or model intestinal juice, depending on the environment in which the bacteria entered the tested system (Figure 3). The analysis conducted in this study on the relationship between pH, fat content, DM content, and viability of the tested lactobacilli strains (L. delbrueckii subsp. Bulgaricus ATCC 11842, L. helveticus LH-B01, L. delbrueckii subsp. Lactis ATCC 4797, and L. acidophilus La-5) revealed that, after incubation of the samples in model gastric juice for 3 h, the population of living lactobacilli cells was more strongly determined by the DM content of the samples (correlation coefficient 0.89, 0.77, 0.69 for L. delbrueckii subsp. Bulgaricus ATCC 11842, L. acidophilus La-5, and L. delbrueckii subsp. Lactis ATCC 4797, respectively; in other cases, the factor was not statistically significant) than by the fat content of the samples (correlation coefficient 0.44 for L. delbrueckii subsp. Bulgaricus ATCC 11842; in other cases, the factor was not statistically significant). In contrast, the pH of the samples was an inhibiting factor for the survival of cells of specific lactobacilli strains in the model gastric juice (correlation coefficient −0.59 for the strains L. delbrueckii subsp. Bulgaricus ATCC 11842 and L. acidophilus La-5; in other cases, the factor was not statistically significant). Similar results were obtained after statistical analysis of the results obtained after incubating the samples in model intestinal juice for 5 h. The analysis of the relationship between pH, fat content, DM content, and viability of the tested lactobacilli strains performed in this case showed that the population of all tested lactobacilli strains was more strongly determined by the DM content of the samples (correlation coefficient 0.97, 0.94, 0.93, and 0.91 for L. delbrueckii subsp. Bulgaricus ATCC 11842, L. delbrueckii subsp. Lactis ATCC 4797, L. helveticus LH-B01, and L. acidophilus La-5, respectively) compared to the fat content of the samples (correlation coefficient 0.60, 0.48, and 0.47 for L. delbrueckii subsp. Lactis ATCC 4797, L. acidophilus La-5, and L. delbrueckii subsp. Bulgaricus ATCC 11842, respectively; in other cases, the factor was not statistically significant). In addition, the pH of the samples was an inhibiting factor for the survival of cells of each lactobacilli strain in the model gastric juice (correlation coefficients −0.45, −0.50, −0.50, and −0.52 for the strains L. delbrueckii subsp. Bulgaricus ATCC 11842, L. acidophilus La-5, L. delbrueckii subsp. Lactis ATCC 4797, and L. helveticus LH-B01, respectively). This may indicate that the dry weight of the products (in this case the significant content of milk-derived proteins) has a more protective effect on lactobacilli bacterial cells than the fat content during passage through the stomach and intestines.
3.5. Determination of the Fatty Acid Profile of Lactobacilli Cells in MRS Broth without and with the Addition of Cholesterol Solution
Various factors have been identified that can protect bacterial cells from the harsh conditions of the gastrointestinal tract, including acidity, organic acids, bile salts, nutrients, and the length of time the bacteria spend in the gut, as well as their initial concentration. Interestingly, cholesterol has also been shown to enhance the survival of bacteria in the stomach and intestines [49]. This is thought to be due to cholesterol’s ability to make LAB more resistant to lysis and alter the composition and function of the cell wall and membrane, thus changing their tolerance to environmental factors [33,49]. Therefore, in this study, the effect of cholesterol on the fatty acid profile of lactobacilli cells in MRS broth, with and without the addition of cholesterol solution, was also investigated. The results of these experiments are presented in Table 2, while the composition of the external standard (BAME) and the identification parameters for each fatty acid are given in Table 3.
The fatty acid profile of the lactobacilli studied was analyzed by chromatographic separation of extracted fatty acids from the bacterial biomass, identifying a total of 29 different fatty acids. Among these, six saturated fatty acids, two single-branched fatty acids with iso and anteiso structures, one hydroxy fatty acid, nine monounsaturated fatty acids with a single double bond, two polyunsaturated fatty acids with multiple double bonds, six conjugated fatty acids, and three cyclic fatty acids were identified. The most predominant fatty acids found in the bacterial biomass were C14:0, C16:0, C16:1,cis-9, C18:0, C18:1,cis-9, C18:1,cis-11, cycC19:0,cis-9,10, and cycC19:0,cis-10,11. Significant differences were observed between the fatty acid profiles of the different lactobacilli strains studied. The largest differences in fatty acid profiles were observed for 12 fatty acids, including C10:0, 15:0,iso, C16:1,trans-9, C16:1,cis-9, cycC17:0,cis-9,10, C18:1,trans-6, C18:1,cis-6, C18:1,cis-9, C18:2,trans-9,trans-12, C18:2,cis-9,cis-12, cycC19:0,cis-9,10, and cycC19:0,cis-10,11. Interestingly, two common monounsaturated fatty acids, C16:1,cis-9 and C18:1,cis-9, were identified as substrates for cell membrane synthesis and can enhance membrane flexibility and fluidity, thereby preventing cell damage and lysis. In contrast, linoleic acid isomers such as C18:2,cis-9,cis-12 and C18:2,trans-9,trans-12 were found to affect metabolic functions of lactobacilli, such as lipid synthesis and fatty acid biosynthesis, and reduce resistance to environmental stresses such as high temperature or high salt concentrations.
The results presented here demonstrate that the presence of cholesterol in MRS broth can impact the fatty acid profile of lactobacilli cells, as previously suggested by Miremadi et al. [74]. The addition of cholesterol in MRS broth induced statistically significant changes in the fatty acid profile of cells from tested lactobacilli strains. Incubation of L. delbrueckii subsp. bulgaricus ATCC 11842 and L. delbrueckii subsp. lactis ATCC 4797 strains in MRS broth with added cholesterol resulted in an increased proportion of C18:2,cis-9,cis-12 (linoleic acid) in the fatty acid pool. In cells of L. helveticus LH-B01, a statistically significant reduction in the level of C16:1,cis-9 (palmitoleic acid) was observed, while in cells of the L. acidophilus La-5 strain, there was a significant reduction in the level of two fatty acids: C18:1,cis-9 (oleic acid) and cycC19:0,cis-9,10 (dihydrosterculic acid) in the fatty acid pool. These observed changes in the fatty acid profile can influence the structure and properties of the cell membrane, which can in turn affect the metabolism and function of these bacteria. For instance, increasing the level of linoleic acid (C18:2,cis-9,cis-12) can elevate cell membrane fluidity and thus enhance the adaptability of bacteria to changing environmental conditions. Conversely, reducing the level of oleic acid (C18:1,cis-9) can increase the acidity of the cell membrane, which can affect the enzymatic and transport properties of cells. Liong and Shah [79] also examined the effect of cholesterol on the fatty acid profile of L. acidophilus bacterial cells and discovered that strains grown without the addition of cholesterol had a higher proportion of unsaturated acids (oleic and linolenic acid) compared to samples with the addition of cholesterol solution. Changes in the fatty acid profile of bacterial cells also suggest the incorporation of cholesterol into the cell membrane [85]. Boudreau and Arul [86] also found that the presence of cholesterol increases certain saturated fatty acids compared to samples without the addition of this ingredient. However, it should be noted that the effects of individual fatty acids on cell membrane and bacterial function are complex and depend on several factors such as the type of fatty acid, its concentration, and its relationship to other fatty acids. Nevertheless, it is worth mentioning that these results are based on in vitro studies, and their relevance to the physiology of lactobacilli in the human or animal body is not clear. Moreover, the effect of cholesterol on the fatty acid profile of lactobacilli cells may depend on several factors such as incubation time, cholesterol concentration, and environmental composition.
The cytoplasmic membrane is essential for the survival of bacterial cells, and the biosynthesis of cell membrane components is a crucial aspect of bacterial physiology. The synthesis of fatty acids must be controlled as it coordinates the production of membrane lipids [87,88,89]. The basic type II biosynthetic pathway of LAB fatty acids is a repetitive cycle of condensation, reduction, dehydration, and reduction of C–C bonds. Lipid components act as a barrier, controlling the permeability of the membrane and contributing to the asymmetry of the lipid membrane, which is necessary for the survival and functioning of bacterial cells [26,84]. The fatty acid profile of LAB cell membrane depends on various factors such as temperature, pH, oxygen, growth phase, composition of medium, and concentration of salts [65,90]. Corcoran et al. [91] demonstrated that C18:1,cis-9 (oleic acid) and C18:1,cis-11 (cis-vaccenic acid) have a protective effect on L. rhamnosus GG cells suspended in artificial gastric juice. They showed that only in the case of these two fatty acids, the number of bacterial cells was greater than the number of cells in the control sample. Other tested fatty acids, including C18:0 (stearic acid), C18:1,trans-9 (elaidic acid), C18:2,cis-9,cis-12 (linoleic acid), C18:2,cis-9,trans-11 (cis-9,trans-11-octadecadienoic acid), and C18:2,trans-10,cis-12 (trans-10,cis-12-octadecadienoic acid), caused a decrease in the number of viable cells—to a level of 3.9 log(CFU/mL) in the case of trans-10,cis-12-octadecadienoic acid and 4.8 log(CFU/mL) for stearic and elaidic acids. This phenomenon can be explained by the fact that oleic and cis-vaccenic acids are the substrates for the synthesis of fatty acids required for cell survival and modification of cell membrane fluidity. LAB are equipped with mechanisms to convert these fatty acids into their cyclic, polyunsaturated, or conjugated forms. Similar observations were made by Taranto et al. [49] who studied the effects of bile acids and cholesterol on the fatty acid profile of cells of L. reuteri CRL 1098. Analysis of the fatty acid profile showed that 50% of the fatty acids present were C16:0, C18:1, C18:2, C18:0, and C19:0, present in varying proportions depending on the medium used. Taranto et al. [49] also observed a similarity between the acid profile of cells cultured in the presence of cholesterol and cells cultured in broth alone when analyzing the effects of stressors on LAB. In contrast, when bile acids were added to the culture medium, no such similarity was observed [49]. Kimoto et al. [20] also noted changes in the distribution of fatty acids by L. lactis cells growing in the presence or absence of cholesterol, which were the result of its removal from the culture medium and its uptake into the cell membrane. These results could aid in understanding the effects of cholesterol on the metabolism of LAB and their impact on the quality of dairy products such as yogurt and kefir. Further research in this area could provide a better understanding of these effects and identify possible benefits or risks to human health.
4. Conclusions
In a study on the removal of cholesterol by cells of lactobacilli in model gastric juice and model intestinal juice, it was found that this variable is dependent on the biomass and viability of the bacterial cells and the bacterial strain studied. The binding of certain amounts of cholesterol by cells of LAB appears to be so strong that it is not released during its passage through the digestive system. The presence of cholesterol can affect the fatty acid profile of lactobacilli cells, which determines the structure and properties of the cell membrane, and in turn, can impact the metabolism and function of these bacteria. These findings may aid in understanding the effect of cholesterol on the metabolism of lactobacilli. However, it was observed that there was no significant effect of cholesterol supplementation on the survival of lactobacilli in model gastric juice or model intestinal juice. Further research in this area could provide a better understanding of this effect and identify possible benefits or risks to human health.
Conversely, there was no statistically significant effect of storage time, passage, or type of lactobacilli culture on the cholesterol content of fermented dairy products. The study also revealed differences in the cell survival rates of individual lactobacilli strains in model gastric juice or model intestinal juice, depending on the environment in which the bacteria entered the tested system. During passage through the stomach and intestines, the dry weight of products, such as significant milk protein content, has a more protective effect on lactobacilli bacterial cells than the fat content.
Author Contributions
Conceptualization, M.Z., D.Z., M.K., and I.S.; formal analysis, M.Z., D.Z., M.K., and I.S.; investigation, M.Z., D.Z., M.K., and I.S.; writing—original draft preparation, M.Z. and D.Z.; writing—review and editing, M.Z., D.Z.; visualization, M.Z., D.Z., M.K., and I.S.; supervision, M.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author (M.Z.), upon reasonable request.
Acknowledgments
The Authors gratefully acknowledge the Institute of Food Sciences of Warsaw University of Life Sciences WULS–SGGW for supporting and providing necessary infrastructure and research stuff.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Albano, C.; Morandi, S.; Silvetti, T.; Casiraghi, M.C.; Manini, F.; Brasca, M. Lactic Acid Bacteria with Cholesterol-Lowering Properties for Dairy Applications: In Vitro and in Situ Activity. J. Dairy. Sci. 2018, 101, 10807–10818. [Google Scholar] [CrossRef]
- Liong, M.T.; Shah, N.P. Optimization of Cholesterol Removal by Probiotics in the Presence of Prebiotics by Using a Response Surface Method. Appl. Env. Microbiol. 2005, 71, 1745–1753. [Google Scholar] [CrossRef]
- Silverman, M.G.; Ference, B.A.; Im, K.; Wiviott, S.D.; Giugliano, R.P.; Grundy, S.M.; Braunwald, E.; Sabatine, M.S. Association Between Lowering LDL-C and Cardiovascular Risk Reduction Among Different Therapeutic Interventions: A Systematic Review and Meta-Analysis. JAMA 2016, 316, 1289–1297. [Google Scholar] [CrossRef]
- Das, T.K.; Pradhan, S.; Chakrabarti, S.; Mondal, K.C.; Ghosh, K. Current Status of Probiotic and Related Health Benefits. Appl. Food Res. 2022, 2, 100185. [Google Scholar] [CrossRef]
- Song, M.; Park, S.; Lee, H.; Min, B.; Jung, S.; Park, S.; Kim, E.; Oh, S. Effect of Lactobacillus Acidophilus NS1 on Plasma Cholesterol Levels in Diet-Induced Obese Mice. J. Dairy. Sci. 2015, 98, 1492–1501. [Google Scholar] [CrossRef]
- Buck, L.M.; Gilliland, S.E. Comparisons of Freshly Isolated Strains of Lactobacillus Acidophilus of Human Intestinal Origin for Ability to Assimilate Cholesterol During Growth. J. Dairy. Sci. 1994, 77, 2925–2933. [Google Scholar] [CrossRef]
- Gilliland, S.E.; Nelson, C.R.; Maxwell, C. Assimilation of Cholesterol by Lactobacillus Acidophilust. Appl. Env. Microbiol. 1985, 49, 377–381. [Google Scholar] [CrossRef]
- Ziarno, M.; Zając, A. Fermented Dairy Products and Cholesterol Levels. Przem. Spoz. 2007, 61, 44–46 /abstract in English/. [Google Scholar]
- Rašić, J.L.; Vujičić, I.F.; Škrinjar, M.; Vulić, M. Assimilation of Cholesterol by Some Cultures of Lactic Acid Bacteria and Bifidobacteria. Biotechnol. Lett. 1992, 14, 39–44. [Google Scholar] [CrossRef]
- Walker, D.K.; Gilliland, S.E. Relationships Among Bile Tolerance, Bile Salt Deconjugation, and Assimilation of Cholesterol by Lactobacillus Acidophilus. J. Dairy. Sci. 1993, 76, 956–961. [Google Scholar] [CrossRef]
- Tahri, K.; Grill, J.P.; Schneider, F. Bifidobacteria Strain Behavior toward Cholesterol: Coprecipitation with Bile Salts and Assimilation. Curr. Microbiol. 1996, 33, 187–193. [Google Scholar] [CrossRef]
- Noh, D.O.; Kim, S.H.; Gilliland, S.E. Incorporation of Cholesterol into the Cellular Membrane of Lactobacillus Acidophilus ATCC 43121. J Dairy Sci 1997, 80, 3107–3113. [Google Scholar] [CrossRef]
- Brashears, M.M.; Gilliland, S.E.; Buck, L.M. Bile Salt Deconjugation and Cholesterol Removal from Media by Lactobacillus Casei. J. Dairy. Sci. 1998, 81, 2103–2110. [Google Scholar] [CrossRef]
- Grill, J.P.; Cayuela, C.; Antoine, J.M.; Schneider, F. Effects of Lactobacillus Amylovorus and Bifidobacterium Breve on Cholesterol. Lett. Appl. Microbiol. 2000, 154–156. [Google Scholar] [CrossRef]
- Lin, M.-Y.; Chen, T. Reduction of Cholesterol by Lactobacillus Acidophilus in Culture Reduction of Cholesterol by Lactobacillus Acidophilus in Culture Broth Broth. J. Food Drug Anal. 2000, 8, 97–102. [Google Scholar] [CrossRef]
- Hosono, A.; Otani, H.; Yasui, H.; Watanuki, M. Impact of Fermented Milk on Human Health: Cholesterol-Lowering and Immunomodulatory Properties of Fermented Milk. Anim. Sci. J. 2002, 73, 241–256. [Google Scholar] [CrossRef]
- Hosono, A.; Tono-Oka, T. Binding of Cholesterol with Lactic Acid Bacterial Cells. Milchwissenschaft 1995, 50, 556–560. [Google Scholar]
- Taranto, M.; Llano, D.G.D.; Rodríguez, A.; Hodalgo, A.P.D.R.; Valdez, G. Bile Tolerance and Cholesterol Reduction by Enterococcus Faecium, a Candidate Microorganism for the Use as a Dietary Adjunct in Milk Products. Milchwissenschaft 1996, 51, 383–385. [Google Scholar]
- Dambekodi, P.C.; Gilliland, S.E. Incorporation of Cholesterol into the Cellular Membrane of Bifidobacterium Longum. J. Dairy. Sci. 1998, 81, 1818–1824. [Google Scholar] [CrossRef]
- Kimoto, H.; Ohmomo, S.; Okamoto, T. Cholesterol Removal from Media by Lactococci. J. Dairy. Sci. 2002, 85, 3182–3188. [Google Scholar] [CrossRef]
- Harrison, V.C.; Peat, G. Serum Cholesterol and Bowel Flora in the Newborn. Am. J. Clin. Nutr. 1975, 28, 1351–1355. [Google Scholar] [CrossRef] [PubMed]
- Grunewald, K.K. Serum Cholesterol Levels in Rats Fed Skim Milk Fermented by Lactobacillus Acidophilus. J. Food Sci. 1982, 47, 2078–2079. [Google Scholar] [CrossRef]
- De Rodas, B.Z.; Gilliland, S.E.; Maxwell, C. V. Hypocholesterolemic Action of Lactobacillus Acidophilus ATCC 43121 and Calcium in Swine with Hypercholesterolemia Induced by Diet. J Dairy Sci 1996, 79, 2121–2128. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.W.; Gilliland, S.E. Effect of Fermented Milk (Yogurt) Containing Lactobacillus Acidophilus L1 on Serum Cholesterol in Hypercholesterolemic Humans. 2013, 18, 43–50. [Google Scholar] [CrossRef]
- Kießling, G.; Schneider, J.; Jahreis, G. Long-Term Consumption of Fermented Dairy Products over 6 Months Increases HDL Cholesterol. Eur. J. Clin. Nutr. 2002, 56, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.J.; Kim, S.Y.; Lee, W.K. Isolation of Cholesterol-Lowering Lactic Acid Bacteria from Human Intestine for Probiotic Use. J. Vet. Sci. 2004, 5, 391–395. [Google Scholar] [CrossRef]
- Lourens-Hattingh, A.; Viljoen, B.C. Yogurt as Probiotic Carrier Food. Int. Dairy. J. 2001, 11, 1–17. [Google Scholar] [CrossRef]
- Guo, L.; Wang, L.; Liu, F.; Li, B.; Tang, Y.; Yu, S.; Zhang, D.; Huo, G. Effect of Bile Salt Hydrolase-Active Lactobacillus Plantarum KLDS 1.0344 on Cholesterol Metabolism in Rats Fed a High-Cholesterol Diet. J. Funct. Foods 2019, 61, 103497. [Google Scholar] [CrossRef]
- Shah, N.P. Functional Cultures and Health Benefits. Int. Dairy. J. 2007, 17, 1262–1277. [Google Scholar] [CrossRef]
- Ayyash, M.M.; Abdalla, A.K.; AlKalbani, N.S.; Baig, M.A.; Turner, M.S.; Liu, S.Q.; Shah, N.P. Invited Review: Characterization of New Probiotics from Dairy and Nondairy Products—Insights into Acid Tolerance, Bile Metabolism and Tolerance, and Adhesion Capability. J. Dairy. Sci. 2021, 104, 8363–8379. [Google Scholar] [CrossRef]
- Anandharaj, M.; Sivasankari, B. Isolation of Potential Probiotic Lactobacillus Oris HMI68 from Mother’s Milk with Cholesterol-Reducing Property. J. Biosci. Bioeng. 2014, 118, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, H.; Chen, X.; Chen, Y.; Menghebilige; Bao, Q. Selection of Potential Probiotic Lactobacilli for Cholesterol-Lowering Properties and Their Effect on Cholesterol Metabolism in Rats Fed a High-Lipid Diet. J. Dairy. Sci. 2012, 95, 1645–1654. [Google Scholar] [CrossRef] [PubMed]
- Ziarno, M. Mechanisms of Cholesterol Lowering by Lactobacillus Bacteria. Żywienie Człowieka i Metabolizm 2004, 2, 10–18, /abstract in English/. [Google Scholar]
- Ziarno, M. Health-Promoting Properties of Lactobacilli. Przegląd Mlecz. 2004, 11, 4–10, /abstract in English/. [Google Scholar]
- Ziarno, M. Importance of Bile Salt Hydrolase Activity in Lactobacilli. Postępy Mikrobiol. 2004, 43, 285–296, /abstract in English/. [Google Scholar]
- Usman, P.; Hosono, A. Binding of Cholesterol to the Cells and Peptidoglycan of Lactobacillus Gasseri. Milchwissenschaft 1999, 54(9), 495–498. [Google Scholar]
- St-Onge, M.P.; Farnworth, E.R.; Jones, P.J.H. Consumption of Fermented and Nonfermented Dairy Products: Effects on Cholesterol Concentrations and Metabolism. Am J Clin Nutr 2000, 71, 674–681. [Google Scholar] [CrossRef]
- Pereira, D.I.A.; Gibson, G.R. Cholesterol Assimilation by Lactic Acid Bacteria and Bifidobacteria Isolated from the Human Gut. Appl. Env. Microbiol. 2002, 68, 4689–4693. [Google Scholar] [CrossRef]
- Pigeon, R.M.; Cuesta, E.P.; Gilliland, S.E. Binding of Free Bile Acids by Cells of Yogurt Starter Culture Bacteria. J. Dairy. Sci. 2002, 85, 2705–2710. [Google Scholar] [CrossRef]
- Liong, M.T.; Shah, N.P. Acid and Bile Tolerance and Cholesterol Removal Ability of Lactobacilli Strains. J. Dairy. Sci. 2005, 88, 55–66. [Google Scholar] [CrossRef]
- Liong, M.T.; Shah, N.P. Bile Salt Deconjugation Ability, Bile Salt Hydrolase Activity and Cholesterol Co-Precipitation Ability of Lactobacilli Strains. Int. Dairy. J. 2005, 15, 391–398. [Google Scholar] [CrossRef]
- Liong, M.T.; Shah, N.P. Bile Salt Deconjugation and BSH Activity of Five Bifidobacterial Strains and Their Cholesterol Co-Precipitating Properties. Food Res. Int. 2005, 38, 135–142. [Google Scholar] [CrossRef]
- Ziarno, M. Significance of Bile Salt Hydrolase Activity in Bacteria of the Genus Bifidobacterium. Biotechnologia 2005, 2, 183–195, /abstract in English/. [Google Scholar]
- Anandharaj, M.; Sivasankari, B.; Santhanakaruppu, R.; Manimaran, M.; Rani, R.P.; Sivakumar, S. Determining the Probiotic Potential of Cholesterol-Reducing Lactobacillus and Weissella Strains Isolated from Gherkins (Fermented Cucumber) and South Indian Fermented Koozh. Res. Microbiol. 2015, 166, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, M.; Tamura, A.; Yamada, N.; Ishida, T.; Hosoda, M.; Hosono, A. Hypocholesterolemic Effects of Viable and Heat-Sterilized Cells of Lactobacillus GG in Rats Fed a High-Cholesterol Diet. Milchwissenschaft 2004, 59(5), 249–253. [Google Scholar]
- Ishimwe, N.; Daliri, E.B.; Lee, B.H.; Fang, F.; Du, G. The Perspective on Cholesterol-Lowering Mechanisms of Probiotics. Mol. Nutr. Food Res. 2015, 59, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Abushelaibi, A.; Al-Mahadin, S.; Enan, M.; El-Tarabily, K.; Shah, N. In-Vitro Investigation into Probiotic Characterisation of Streptococcus and Enterococcus Isolated from Camel Milk. LWT 2018, 87, 478–487. [Google Scholar] [CrossRef]
- Klaver, F.A.M.; Van der Meer, R. The Assumed Assimilation of Cholesterol by Lactobacilli and Bifidobacterium Bifidum Is Due to Their Bile Salt-Deconjugating Activity. Appl. Env. Microbiol. 1993, 59, 1120–1124. [Google Scholar] [CrossRef]
- Taranto, M.P.; Fernandez Murga, M.L.; Lorca, G.; De Valdez, G.F. Bile Salts and Cholesterol Induce Changes in the Lipid Cell Membrane of Lactobacillus Reuteri. J. Appl. Microbiol. 2003, 95, 86–91. [Google Scholar] [CrossRef]
- Choi, S.B.; Lew, L.C.; Yeo, S.K.; Parvathy, S.N.; Liong, M.T. Probiotics and the BSH-Related Cholesterol Lowering Mechanism: A Jekyll and Hyde Scenario. Crit. Rev. Biotechnol. 2014, 35, 392–401. [Google Scholar] [CrossRef]
- Maske, B.L.; de Melo Pereira, G. V.; da, S. Vale, A.; de Carvalho Neto, D.P.; Karp, S.G.; Viesser, J.A.; De Dea Lindner, J.; Pagnoncelli, M.G.; Soccol, V.T.; Soccol, C.R. A Review on Enzyme-Producing Lactobacilli Associated with the Human Digestive Process: From Metabolism to Application. Enzym. Microb. Technol. 2021, 149, 109836. [Google Scholar] [CrossRef] [PubMed]
- Sridevi, N.; Vishwe, P.; Prabhune, A. Hypocholesteremic Effect of Bile Salt Hydrolase from Lactobacillus Buchneri ATCC 4005. Food Res. Int. 2009, 42, 516–520. [Google Scholar] [CrossRef]
- Jones, M.L.; Martoni, C.J.; Parent, M.; Prakash, S. Cholesterol-Lowering Efficacy of a Microencapsulated Bile Salt Hydrolase-Active Lactobacillus Reuteri NCIMB 30242 Yoghurt Formulation in Hypercholesterolaemic Adults. British Journal of Nutrition 2012. [CrossRef]
- Pereira, D.I.A.; McCartney, A.L.; Gibson, G.R. An in Vitro Study of the Probiotic Potential of a Bile-Salt-Hydrolyzing Lactobacillus Fermentum Strain, and Determination of Its Cholesterol-Lowering Properties. Appl. Env. Microbiol. 2003, 69, 4743–4752. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Suzuki, Y.; Hirota, T. Cholesterol Lowering Activity of Ropy Fermented Milk. J. Food Sci. 1992, 57, 1327–1329. [Google Scholar] [CrossRef]
- De Vuyst, L.; De Vin, F.; Vaningelgem, F.; Degeest, B. Recent Developments in the Biosynthesis and Applications of Heteropolysaccharides from Lactic Acid Bacteria. Int. Dairy. J. 2001, 11, 687–707. [Google Scholar] [CrossRef]
- Clavel, T.; Carlin, F.; Lairon, D.; Nguyen-The, C.; Schmitt, P. Survival of Bacillus Cereus Spores and Vegetative Cells in Acid Media Simulating Human Stomach. J. Appl. Microbiol. 2004, 97, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Marteau, P.; Minekus, M.; Havenaar, R.; Huis In’t Veld, J.H.J. Survival of Lactic Acid Bacteria in a Dynamic Model of the Stomach and Small Intestine: Validation and the Effects of Bile. J. Dairy. Sci. 1997, 80, 1031–1037. [Google Scholar] [CrossRef]
- Fletouris, D.J.; Botsoglou, N.A.; Psomas, I.E.; Mantis, A.I. Rapid Determination of Cholesterol in Milk and Milk Products by Direct Saponification and Capillary Gas Chromatography. J. Dairy. Sci. 1998, 81, 2833–2840. [Google Scholar] [CrossRef]
- Buyer, J.S. Rapid Sample Processing and Fast Gas Chromatography for Identification of Bacteria by Fatty Acid Analysis. J. Microbiol. Methods 2002, 51, 209–215. [Google Scholar] [CrossRef]
- Kidd Haack, S.; Garchow, H.; Odelson, D.A.; Forney, L.J.; Klug, M.J. Accuracy, Reproducibility, and Interpretation of Fatty Acid Methyl Ester Profiles of Model Bacterial Communities. Appl. Env. Microbiol. 1994, 60, 2483–2493. [Google Scholar] [CrossRef] [PubMed]
- Kankaanpää, P.; Yang, B.; Kallio, H.; Isolauri, E.; Salminen, S. Effects of Polyunsaturated Fatty Acids in Growth Medium on Lipid Composition and on Physicochemical Surface Properties of Lactobacilli. Appl. Env. Microbiol. 2004, 70, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Brondz, I. Development of Fatty Acid Analysis by High-Performance Liquid Chromatography, Gas Chromatography, and Related Techniques. Anal. Chim. Acta 2002, 465, 1–37. [Google Scholar] [CrossRef]
- Christie, W.W.; Dobson, G.; Adlof, R.O. A Practical Guide to the Isolation, Analysis and Identification of Conjugated Linoleic Acid. Lipids 2007, 42, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Montanari, C.; Sado Kamdem, S.L.; Serrazanetti, D.I.; Etoa, F.X.; Guerzoni, M.E. Synthesis of Cyclopropane Fatty Acids in Lactobacillus Helveticus and Lactobacillus Sanfranciscensis and Their Cellular Fatty Acids Changes Following Short Term Acid and Cold Stresses. Food Microbiol. 2010, 27, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.U.; Jenkins, D.J.A.; Reichert, R. The Effect of Fermented and Unfermented Milks on Serum Cholesterol. Am. J. Clin. Nutr. 1982, 36, 1106–1111. [Google Scholar] [CrossRef] [PubMed]
- Jaspers, D.A.; Massey, L.K.; Luedecke, L.O. Effect of Consuming Yogurts Prepared with Three Culture Strains on Human Serum Lipoproteins. J. Food Sci. 1984, 49, 1178–1181. [Google Scholar] [CrossRef]
- Lin, S.Y.; Ayres, J.W.; Winkler, W.; Sandine, W.E. Lactobacillus Effects on Cholesterol: In Vitro and In Vivo Results. J. Dairy. Sci. 1989, 72, 2885–2899. [Google Scholar] [CrossRef]
- McNamara, D.J.; Lowell, A.E.; Sabb, J.E. Effect of Yogurt Intake on Plasma Lipid and Lipoprotein Levels in Normolipidemic Males. Atherosclerosis 1989, 79, 167–171. [Google Scholar] [CrossRef]
- Choi, E.A.; Chang, H.C. Cholesterol-Lowering Effects of a Putative Probiotic Strain Lactobacillus Plantarum EM Isolated from Kimchi. LWT - Food Sci. Technol. 2015, 62, 210–217. [Google Scholar] [CrossRef]
- Ooi, L.-G.; Liong, M.-T. Cholesterol-Lowering Effects of Probiotics and Prebiotics: A Review of in Vivo and in Vitro Findings. Int. J. Mol. Sci. 2010, 11, 2499–2522. [Google Scholar] [CrossRef] [PubMed]
- Lye, H.S.; Rusul, G.; Liong, M.T. Removal of Cholesterol by Lactobacilli via Incorporation and Conversion to Coprostanol. J. Dairy. Sci. 2010, 93, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; El-Nezami, H.; Haskard, C.A.; Gratz, S.; Puong, K.Y.; Salminen, S.; Mykkänen, H. Kinetics of Adsorption and Desorption of Aflatoxin B1 by Viable and Nonviable Bacteria. J. Food Prot. 2003, 66, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Miremadi, F.; Ayyash, M.; Sherkat, F.; Stojanovska, L. Cholesterol Reduction Mechanisms and Fatty Acid Composition of Cellular Membranes of Probiotic Lactobacilli and Bifidobacteria. J. Funct. Foods 2014, 9, 295–305. [Google Scholar] [CrossRef]
- El-Nezami, H.; Kankaanpaa, P.; Salminen, S.; Ahokas, J. Ability of Dairy Strains of Lactic Acid Bacteria to Bind a Common Food Carcinogen, Aflatoxin B1. Food and Chemical Toxicology 1998, 36, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Aloǧlu, H.; Öner, Z. Assimilation of Cholesterol in Broth, Cream, and Butter by Probiotic Bacteria. Eur. J. Lipid Sci. Technol. 2006, 108, 709–713. [Google Scholar] [CrossRef]
- Tanaka, H.; Doesburg, K.; Iwasaki, T.; Mierau, I. Screening of Lactic Acid Bacteria for Bile Salt Hydrolase Activity. J. Dairy. Sci. 1999, 82, 2530–2535. [Google Scholar] [CrossRef]
- Taranto, M.P.; Sesma, F.; Pesce De Ruiz Holgado, A.; De Valdez, G.F. Bile Salts Hydrolase Plays a Key Role on Cholesterol Removal by Lactobacillus Reuteri. Biotechnol. Lett. 1997, 19, 845–847. [Google Scholar] [CrossRef]
- Liong, M.T.; Shah, N.P. Bile Salt Deconjugation Ability, Bile Salt Hydrolase Activity and Cholesterol Co-Precipitation Ability of Lactobacilli Strains. Int. Dairy. J. 2005, 15, 391–398. [Google Scholar] [CrossRef]
- Zhang, F.; Qiu, L.; Xu, X.; Liu, Z.; Zhan, H.; Tao, X.; Shah, N.P.; Wei, H. Beneficial Effects of Probiotic Cholesterol-Lowering Strain of Enterococcus Faecium WEFA23 from Infants on Diet-Induced Metabolic Syndrome in Rats. J. Dairy. Sci. 2017, 100, 1618–1628. [Google Scholar] [CrossRef]
- Roobab, U.; Batool, Z.; Manzoor, M.F.; Shabbir, M.A.; Khan, M.R.; Aadil, R.M. Sources, Formulations, Advanced Delivery and Health Benefits of Probiotics. Curr. Opin. Food Sci. 2020, 32, 17–28. [Google Scholar] [CrossRef]
- Vinderola, C.G.; Reinheimer, J.A. Lactic Acid Starter and Probiotic Bacteria: A Comparative “in Vitro” Study of Probiotic Characteristics and Biological Barrier Resistance. Food Res. Int. 2003, 36, 895–904. [Google Scholar] [CrossRef]
- Kailasapathy, K.; Chin, J. Survival and Therapeutic Potential of Probiotic Organisms with Reference to Lactobacillus Acidophilus and Bifidobacterium Spp. Immunol. Cell Biol. 2000, 78, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Elli, M.; Callegari, M.L.; Ferrari, S.; Bessi, E.; Cattivelli, D.; Soldi, S.; Morelli, L.; Feuillerat, N.G.; Antoine, J.M. Survival of Yogurt Bacteria in the Human Gut. Appl. Env. Microbiol. 2006, 72, 5113–5117. [Google Scholar] [CrossRef] [PubMed]
- Lye, H.S.; Rusul, G.; Liong, M.T. Removal of Cholesterol by Lactobacilli via Incorporation and Conversion to Coprostanol. J. Dairy. Sci. 2010, 93, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, A.; Arul, J. Cholesterol Reduction and Fat Fractionation Technologies for Milk Fat: An Overview. J. Dairy. Sci. 1993, 76, 1772–1781. [Google Scholar] [CrossRef]
- Benito, M.J.; Serradilla, M.J.; Ruiz-Moyano, S.; Martín, A.; Pérez-Nevado, F.; Córdoba, M.G. Rapid Differentiation of Lactic Acid Bacteria from Autochthonous Fermentation of Iberian Dry-Fermented Sausages. Meat Sci. 2008, 80, 656–661. [Google Scholar] [CrossRef]
- Thirabunyanon, M.; Hongwittayakorn, P. Potential Probiotic Lactic Acid Bacteria of Human Origin Induce Antiproliferation of Colon Cancer Cells via Synergic Actions in Adhesion to Cancer Cells and Short-Chain Fatty Acid Bioproduction. Appl. Biochem. Biotechnol. 2013, 169, 511–525. [Google Scholar] [CrossRef]
- Glatz, J.F.C.; Luiken, J.J.F.P. Fatty Acids in Cell Signaling: Historical Perspective and Future Outlook. Prostaglandins Leukot. Essent. Fat. Acids 2015, 92, 57–62. [Google Scholar] [CrossRef]
- Partanen, L.; Marttinen, N.; Alatossava, T. Fats and Fatty Acids as Growth Factors for Lactobacillus Delbrueckii. Syst. Appl. Microbiol. 2001, 24, 500–506. [Google Scholar] [CrossRef]
- Corcoran, B.M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Growth of Probiotic Lactobacilli in the Presence of Oleic Acid Enhances Subsequent Survival in Gastric Juice. Microbiology 2007, 153, 291–299. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a,c,e,g) Cholesterol removal by biomasses with different concentrations of live and dead cells of lactobacilli strains after 3 or 5 h holding in model gastric or intestinal juices, respectively. (b,d,f,h) Cholesterol removal or release by biomasses with different concentrations of live and dead cells of lactic acid bacterial strains after 24 or 8 h holding in MRS broth or model digestive juice, respectively (mean values and SD values).
Figure 1.
(a,c,e,g) Cholesterol removal by biomasses with different concentrations of live and dead cells of lactobacilli strains after 3 or 5 h holding in model gastric or intestinal juices, respectively. (b,d,f,h) Cholesterol removal or release by biomasses with different concentrations of live and dead cells of lactic acid bacterial strains after 24 or 8 h holding in MRS broth or model digestive juice, respectively (mean values and SD values).
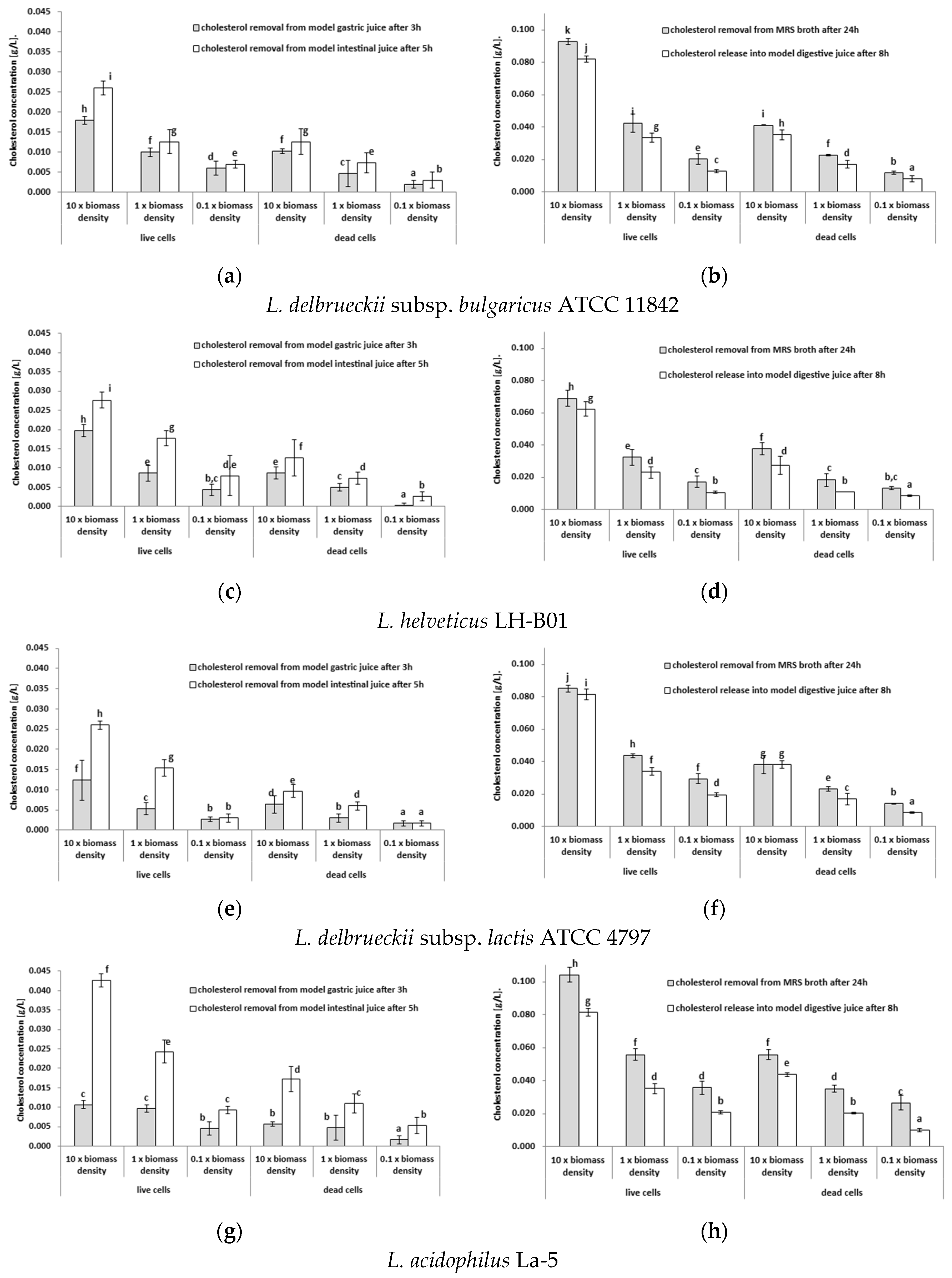
Figure 2.
Number of viable cells of lactobacilli strains when kept in model digestive juices [log(CFU/mL)]; nd, not detected.
Figure 2.
Number of viable cells of lactobacilli strains when kept in model digestive juices [log(CFU/mL)]; nd, not detected.
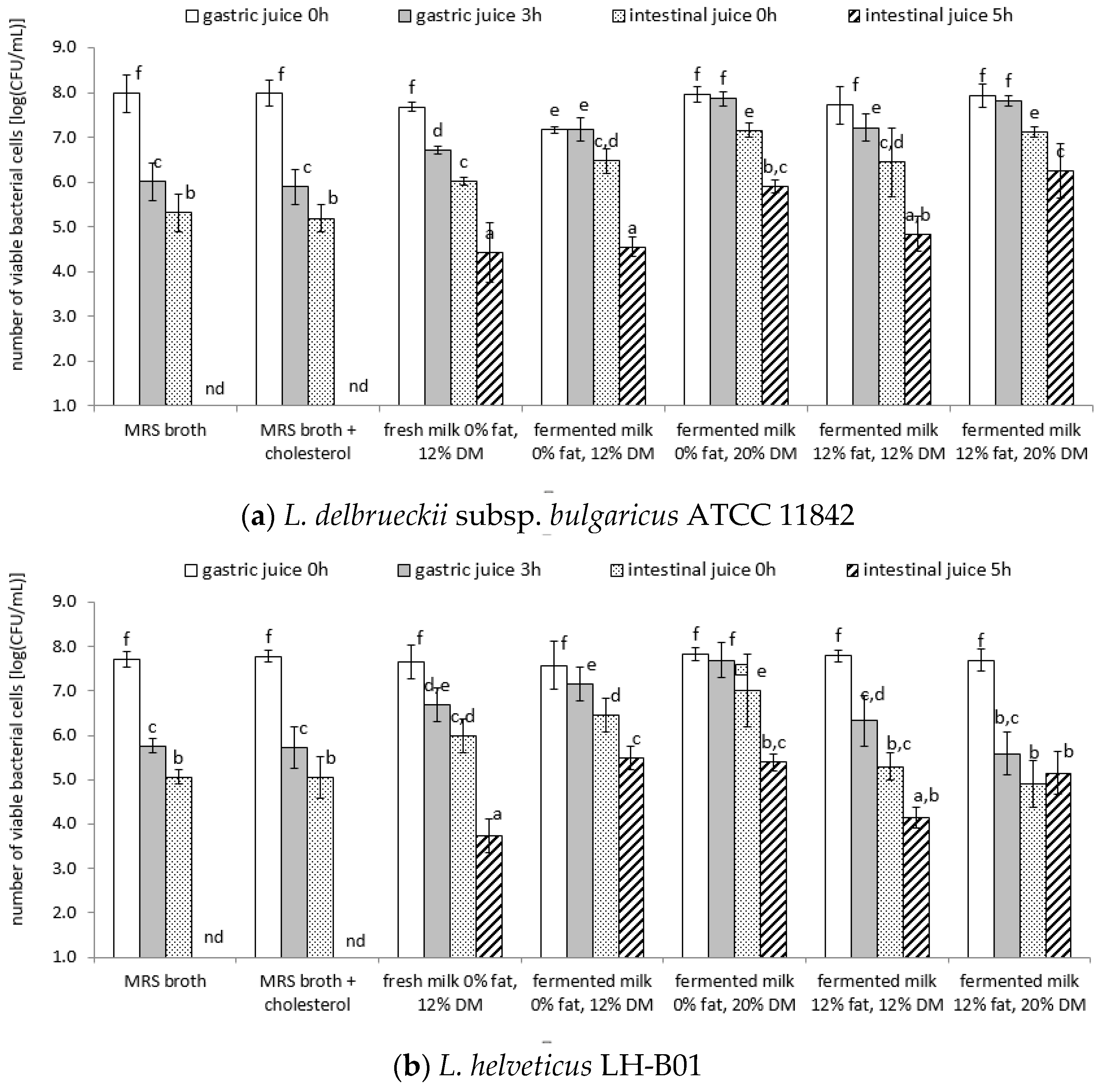
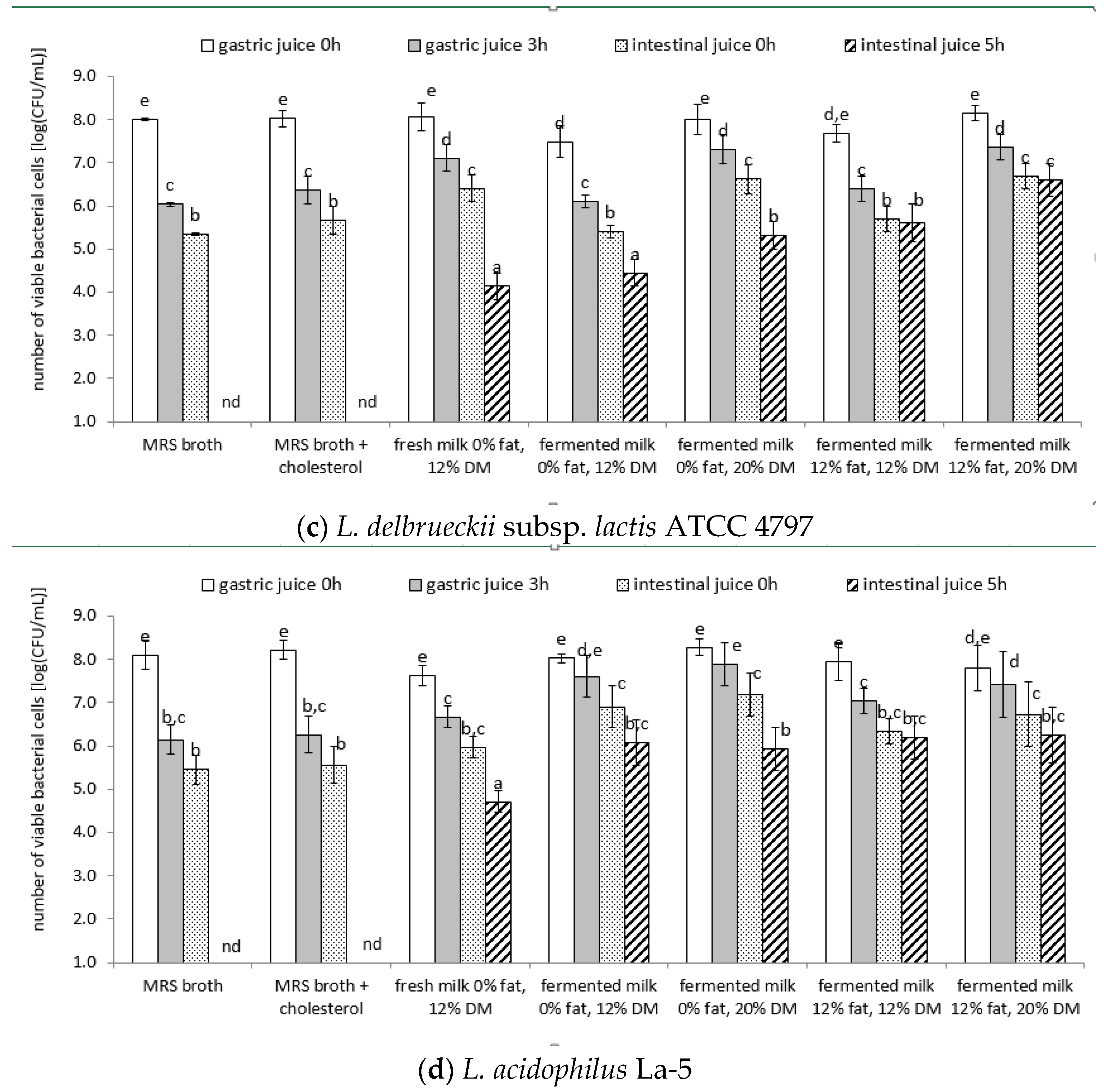
Figure 3.
Graphs of correlation matrix showing the relationship between pH, fat content, dry matter content, and the viability of tested lactobacilli strains (L. delbrueckii subsp. bulgaricus ATCC 11842, L. helveticus LH-B01, L. delbrueckii subsp. lactis ATCC 4797, and L. acidophilus La-5) (a) after 3 h of incubation in the model gastric juice (b) after 5 h of incubation in the model intestinal juice (with a confidence level of 95.0%); x, not significant at 0.05.
Figure 3.
Graphs of correlation matrix showing the relationship between pH, fat content, dry matter content, and the viability of tested lactobacilli strains (L. delbrueckii subsp. bulgaricus ATCC 11842, L. helveticus LH-B01, L. delbrueckii subsp. lactis ATCC 4797, and L. acidophilus La-5) (a) after 3 h of incubation in the model gastric juice (b) after 5 h of incubation in the model intestinal juice (with a confidence level of 95.0%); x, not significant at 0.05.
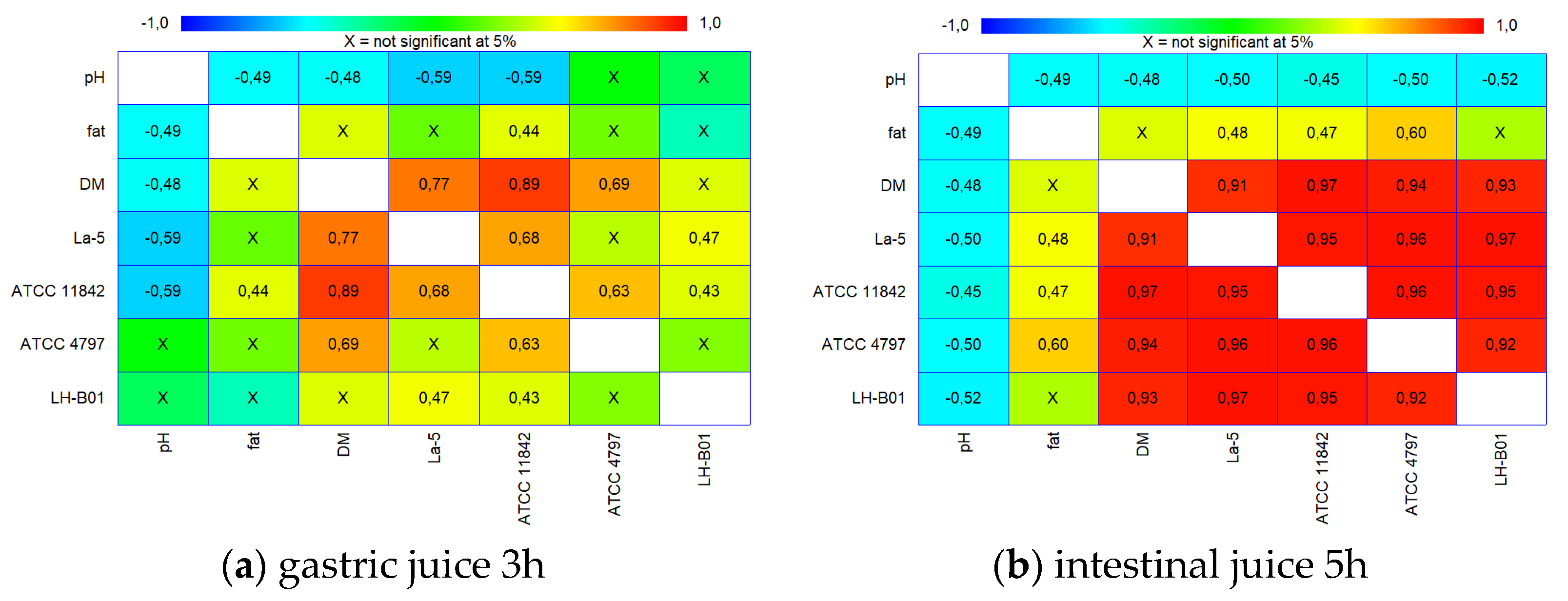

Table 1.
Changes in cholesterol content of different model product systems during cold storage and digestion in model digestive juices [calculated as mg of cholesterol in 100 g of the primary product samples].
Table 1.
Changes in cholesterol content of different model product systems during cold storage and digestion in model digestive juices [calculated as mg of cholesterol in 100 g of the primary product samples].
| Strain | Product | Directly after Preparation | After 1 Week of Cold Storage |
After 4 Weeks of Cold Storage |
Model Gastric Juice after 3 Hours | Model Duodenal Fluid after 5h |
|---|---|---|---|---|---|---|
| L. delbrueckii subsp. Bulgaricus ATCC 11842 | MRS broth | 0.0 a ±0.00 | 0.0 a ±0.00 | 0.0 a ±0.00 | 0.0 a ±0.00 | 0.0 a ±0.00 |
| MRS broth + cholesterol | 0.7 b ±0.01 | 0.7 b ±0.04 | 0.7 b ±0.02 | 0.7 b ±0.03 | 0.7 b ±0.03 | |
| fresh milk 0,05% fat, 12% DM | 1.7 c ±0.04 | 1.6 c ±0.02 | 1.6 c ±0.04 | 1.7 c ±0.05 | 1.6 c ±0.05 | |
| fermented milk 0,05% fat, 12% DM | 1.7 c ±0.04 | 1.6 c ±0.02 | 1.6 c ±0.04 | 1.7 c ±0.05 | 1.6 c ±0.05 | |
| fermented milk 0,05% fat, 20% DM | 1.6 c ±0.04 | 1.5 c ±0.02 | 1.5 c ±0.04 | 1.6 c ±0.05 | 1.5 c ±0.05 | |
| fermented milk 12% fat, 12% DM | 40.6 e ±1.07 | 39.4 e ±0.41 | 38.5 e ±1.02 | 40.5 e ±1.13 | 39.0 e ±1.13 | |
| fermented milk 12% fat, 20% DM | 37.5 d ±0.99 | 36.5 d ±0.38 | 35.6 d ±0.95 | 37.4 d ±0.97 | 36.0 d ±0.97 | |
| L. helveticus LH-B01 | MRS broth | 0.0 a ±0.00 | 0.0 a ±0.00 | 0.0 a ±0.00 | 0.0 a ±0.00 | 0.0 a ±0.00 |
| MRS broth + cholesterol | 0.7 b ±0.02 | 0.7 c ±0.02 | 0.7 c ±0.02 | 0.6 b ±0.02 | 0.6 b ±0.02 | |
| fresh milk 0,05% fat, 12% DM | 1.6 c ±0.04 | 1.6 c ±0.06 | 1.6 c ±0.01 | 1.6 c ±0.05 | 1.6 c ±0.05 | |
| fermented milk 0,05% fat, 12% DM | 1.6 c ±0.04 | 1.6 c ±0.06 | 1.6 c ±0.01 | 1.6 c ±0.05 | 1.5 c ±0.05 | |
| fermented milk 0,05% fat, 20% DM | 1.5 c ±0.04 | 1.5 c ±0.05 | 1.5 c ±0.01 | 1.5 c ±0.04 | 1.4 c ±0.04 | |
| fermented milk 12% fat, 12% DM | 39.3 e ±0.93 | 38.1 e ±1.39 | 39.1 e ±0.23 | 39.3 e ±1.15 | 37.2 e ±1.15 | |
| fermented milk 12% fat, 20% DM | 36.4 d ±0.85 | 35.3 d ±1.28 | 36.2 d ±0.22 | 36.4 d ±1.09 | 34.5 d ±1.09 | |
| L. delbrueckii subsp. Lactis ATCC 4797 | MRS broth | 0.0 a ±0.00 | 0.0 a ±0.00 | 0.0 a ±0.00 | 0.0 a ±0.00 | 0.0 a ±0.00 |
| MRS broth + cholesterol | 0.6 b ±0.03 | 0.7 b ±0.01 | 0.7 b ±0.02 | 0.6 b ±0.01 | 0.6 b ±0.01 | |
| fresh milk 0,05% fat, 12% DM | 1.6 c ±0.01 | 1.6 c ±0.02 | 1.6 c ±0.01 | 1.6 c ±0.05 | 1.5 c ±0.05 | |
| fermented milk 0,05% fat, 12% DM | 1.6 c ±0.01 | 1.6 c ±0.02 | 1.6 c ±0.01 | 1.6 c ±0.05 | 1.5 c ±0.04 | |
| fermented milk 0,05% fat, 20% DM | 1.5 c ±0.01 | 1.5 c ±0.02 | 1.5 c ±0.01 | 1.5 c ±0.04 | 1.4 c ±0.04 | |
| fermented milk 12% fat, 12% DM | 37.9 e ±0.29 | 38.6 e ±0.56 | 38.0 e ±0.25 | 37.9 e ±1.01 | 36.5 e ±1.01 | |
| fermented milk 12% fat, 20% DM | 35.1 d ±0.27 | 35.8 d ±0.51 | 35.1 d ±0.24 | 35.1 d ±1.02 | 33.8 d ±1.02 | |
| L. acidophilus La-5 | MRS broth | 0.0 a ±0.00 | 0.0 a ±0.00 | 0.0 a ±0.00 | 0.0 a ±0.00 | 0.0 a ±0.00 |
| MRS broth + cholesterol | 0.6 b ±0.03 | 0.7 b ±0.01 | 0.7 b ±0.03 | 0.6 b ±0.02 | 0.6 b ±0.02 | |
| fresh milk 0,05% fat, 12% DM | 1.6 c ±0.06 | 1.6 c ±0.03 | 1.6 c ±0.02 | 1.6 c ±0.04 | 1.5 c ±0.05 | |
| fermented milk 0,05% fat, 12% DM | 1.6 c ±0.06 | 1.6 c ±0.03 | 1.6 c ±0.02 | 1.6 c ±0.05 | 1.5 c ±0.06 | |
| fermented milk 0,05% fat, 20% DM | 1.5 c ±0.05 | 1.5 c ±0.02 | 1.5 c ±0.02 | 1.5 c ±0.05 | 1.4 c ±0.05 | |
| fermented milk 12% fat, 12% DM | 38.6 e ±1.32 | 38.8 e ±0.64 | 38.5 e ±0.51 | 38.6 e ±1.05 | 36.6 e ±1.05 | |
| fermented milk 12% fat, 20% DM | 35.8 d ±1.23 | 35.9 d ±0.59 | 35.7 d ±0.48 | 35.7 d ±1.07 | 33.9 d ±1.07 |
a,b,c,d —Means with different uppercase letters across the table are significantly different (p < 0.05).
Table 2.
Fatty acids composition (%) in the lactobacilli strains profiles obtained from the cell cultured in an MRS broth with and without cholesterol (means ± SD).
Table 2.
Fatty acids composition (%) in the lactobacilli strains profiles obtained from the cell cultured in an MRS broth with and without cholesterol (means ± SD).
| Fatty acid | L. delbrueckii subsp. bulgaricus ATCC 11842 | L. helveticus LH-B01 | L. delbrueckii subsp. lactis ATCC 4797 | L. acidophilus La-5 | |||||
|---|---|---|---|---|---|---|---|---|---|
| MRS broth | MRS broth + chol |
MRS broth | MRS broth + chol |
MRS broth | MRS broth + chol |
MRS broth | MRS broth + chol |
||
| C10:0 | caproic / decanoic | 0.0009 a,b ±0.0006 | 0.0002 a,b ±0.0002 | 0.0003 a ±0.0001 | 0.0005 a ±0.0002 | 0.0005 a,b ±0.0002 | 0.0015 a,b ±0.0009 | 0,0017 b ±0,0006 | 0,0008 b ±0,0002 |
| C12:0 | lauric / dodecanoic | 0.0047 ±0.0019 | 0.0012 ±0.0000 | 0.0118 ±0.0025 | 0.0269 ±0.0011 | 0.0044 ±0.0003 | 0.0131 ±0.0021 | 0,0100 ±0,0018 | 0,0051 ±0,0001 |
| C14:0 | myristic / tetradecanoic | 0.0081 ±0.0020 | 0.0041 ±0.0002 | 0.0038 ±0.0017 | 0.0032 ±0.0005 | 0.0026 ±0.0006 | 0.0076 ±0.0019 | 0,0285 ±0,0029 | 0,0156 ±0,0025 |
| 15:0,iso | iso-13-methyltetradecanoic | 0.0036 a,b ±0.0012 | 0.0009 a,b ±0.0001 | 0.0118 b ±0.0024 | 0.0267 b ±0.0040 | 0.0001 a ±0.0000 | 0.0002 a ±0.0001 | 0,0002 a ±0,0001 | 0,0001 a ±0,0000 |
| 15:0,anteiso | anteiso-12-methyltetradecanoic | 0.0003 ±0.0002 | 0.0001 ±0.0001 | 0.0006 ±0.0009 | 0.0014 ±0.0012 | 0.0002 ±0.0001 | 0.0007 ±0.0004 | 0,0008 ±0,0003 | 0,0004 ±0,0001 |
| C15:0 | pentadecanoic | 0.0001 ±0.0001 | 0.0000 ±0.0000 | 0.0005 ±0.0007 | 0.0011 ±0.0010 | 0.0001 ±0.0000 | 0.0004 ±0.0002 | 0,0004 ±0,0002 | 0,0002 ±0,0000 |
| C16:0 | palmitic / hexadecanoic | 0.0243 ±0.0049 | 0.0210 ±0.0068 | 0.0148 ±0.0038 | 0.0182 ±0.0027 | 0.0142 ±0.0013 | 0.0360 ±0.0016 | 0,0913 ±0,0037 | 0,0483 ±0,0050 |
| C16:1,trans-9 | palmitelaidic / trans-9-hexadecenoic | 0.0200 b ±0.0023 | 0.0194 b ±0.0086 | 0.0112 b ±0.0025 | 0.0253 b ±0.0018 | 0.0002 a ±0.0000 | 0.0005 a ±0.0003 | 0,0005 a ±0,0002 | 0,0003 a ±0,0001 |
| C16:1,cis-9 | palmitoleic / cis-9-hexadecenoic | 0.0954 b ±0.0185 | 0.0031 b ±0.0000 | 0.0031 b ±0.0013 | 0.0000 a ±0.0000 | 0.0006 a ±0.0001 | 0.0000 a ±0.0000 | 0,0065 b ±0,0012 | 0,0044 b ±0,0004 |
| C12:0,2OH | 2-hydroxydodecanoic | 0.0003 ±0.0002 | 0.0001 ±0.0001 | 0.0008 ±0.0002 | 0.0018 ±0.0006 | 0.0001 ±0.0000 | 0.0004 ±0.0002 | 0,0005 ±0,0002 | 0,0003 ±0,0001 |
| cycC17:0,cis-9,10 | cis-9,10-methylenehexadecanoic | 0.0049 b ±0.0010 | 0.0048 b ±0.0016 | 0.0000 a ±0.0000 | 0.0000 a ±0.0000 | 0.0001 a ±0.0000 | 0.0003 a ±0.0002 | 0,0000 a ±0,0000 | 0,0000 a ±0,0000 |
| C18:0 | stearic / octadecanoic | 0.0030 ±0.0008 | 0.0054 ±0.0010 | 0.0029 ±0.0005 | 0.0067 ±0.0014 | 0.0024 ±0.0004 | 0.0082 ±0.0027 | 0,0296 ±0,0025 | 0,0138 ±0,0014 |
| C18:1 | octadecenoic | 0.0010 ±0.0006 | 0.0003 ±0.0002 | 0.0009 ±0.0004 | 0.0021 ±0.0009 | 0.0000 ±0.0000 | 0.0000 ±0.0000 | 0,0013 ±0,0005 | 0,0006 ±0,0001 |
| C18:1,trans-6 | petroselaidic / trans-6-octadecenoic | 0.0000 a ±0.0000 | 0.0000 a ±0.0000 | 0.0007 b ±0.0010 | 0.0015 b ±0.0004 | 0.0000 a ±0.0000 | 0.0000 a ±0.0000 | 0,0001 a ±0,0000 | 0,0000 a ±0,0000 |
| C18:1,trans-9 | elaidic / trans-9-octa-decenoic | 0.0002 ±0.0001 | 0.0005 ±0.0004 | 0.0000 ±0.0001 | 0.0018 ±0.0008 | 0.0000 ±0.0000 | 0.0000 ±0.0000 | 0,0000 ±0,0000 | 0,0000 ±0,0000 |
| C18:1,trans-11 | trans-vaccenic / trans-11-octadecenoic | 0.0012 ±0.0007 | 0.0003 ±0.0003 | 0.0020 ±0.0019 | 0.0044 ±0.0010 | 0.0007 ±0.0002 | 0.0020 ±0.0003 | 0,0024 ±0,0009 | 0,0012 ±0,0002 |
| C18:1,cis-6 | petroselinic / cis-6-octadecenoic | 0.0004 a ±0.0003 | 0.0005 a ±0.0005 | 0.0063 b ±0.0014 | 0.0144 b ±0.0029 | 0.0000 a ±0.0000 | 0.0000 a ±0.0000 | 0,0004 a ±0,0002 | 0,0002 a ±0,0000 |
| C18:1,cis-9 | oleic / cis-9-octadecenoic | 0.1324 a,b ±0.0011 | 0.0624 a ±0.0000 | 0.0327 a ±0.0022 | 0.0584 a ±0.0016 | 0.0545 a ±0.0007 | 0.1142 a,b ±0.0072 | 0,1270 b ±0,0047 | 0,0625 a ±0,0059 |
| C18:1,cis-11 | cis-vaccenic / cis-11-octadecenoic | 0.0179 ±0.0010 | 0.1600 ±0.0047 | 0.0574 ±0.0049 | 0.1302 ±0.0071 | 0.0025 ±0.0000 | 0.0075 ±0.0002 | 0,0059 ±0,0013 | 0,0030 ±0,0006 |
| C18:2,trans-9,trans-12 | linoelaidic / trans-9,trans-12-octadecadienoic |
0.0008 b ±0.0005 | 0.0002 a,b ±0.0002 | 0.0000 a ±0.0000 | 0.0000 a ±0.0000 | 0.0000 a ±0.0000 | 0.0001 a ±0.0001 | 0,0000 a ±0,0000 | 0,0000 a ±0,0000 |
| C18:2,cis-9,cis-12 | linoleic / cis-9,cis12-octadecadienoic | 0.0005 a ±0.0003 | 0.0184 b ±0.0000 | 0.0000 a ±0.0000 | 0.0038 a,b ±0.0019 | 0.0000 a ±0.0000 | 0.0080 b ±0.0026 | 0,0043 a,b ±0,0010 | 0,0000 a ±0,0000 |
| cycC19:0,cis-9,10 | dihydrosterculic / cis-9,10-methyleneoctadecanoic |
0.0632 b ±0.0087 | 0.0108 b ±0.0000 | 0.0070 a ±0.0015 | 0.0000 a ±0.0000 | 0.0078 a ±0.0012 | 0.0068 a ±0.0007 | 0,0413 b ±0,0020 | 0,0053 a ±0,0015 |
| cycC19:0,cis-10,11 | lactobacillic / cis-11,12-methyleneoctadecanoic |
0.0087 a ±0.0013 | 0.0022 a ±0.0009 | 0.0600 b ±0.0017 | 0.1361 b ±0.0023 | 0.0000 a ±0.0000 | 0.0000 a ±0.0000 | 0,0018 a7 ±0,0007 | 0,0009 a ±0,0002 |
| 18:2,cis-9,trans-11 | conjugated octadecadienoic | 0.0058 ±0.0015 | 0.0015 ±0.0003 | 0.0069 ±0.0002 | 0.0156 ±0.0021 | 0.0015 ±0.0005 | 0.0045 ±0.0018 | 0,0057 ±0,0012 | 0,0029 ±0,0006 |
| C18:2, CLA_1 | conjugated octadecadienoic | 0.0005 ±0.0003 | 0.0001 ±0.0001 | 0.0007 ±0.0010 | 0.0015 ±0.0004 | 0.0002 ±0.0001 | 0.0007 ±0.0004 | 0,0005 ±0,0002 | 0,0003 ±0,0000 |
| 18:2,trans-10,cis-12 | conjugated octadecadienoic | 0.0031 ±0.0019 | 0.0008 ±0.0007 | 0.0074 ±0.0009 | 0.0167 ±0.0011 | 0.0023 ±0.0007 | 0.0070 ±0.0023 | 0,0062 ±0,0014 | 0,0032 ±0,0006 |
| C18:2, CLA_2 | conjugated octadecadienoic | 0.0004 ±0.0002 | 0.0001 ±0.0000 | 0.0007 ±0.0001 | 0.0015 ±0.0004 | 0.0002 ±0.0001 | 0.0006 ±0.0002 | 0,0006 ±0,0002 | 0,0003 ±0,0001 |
| C18:2, CLA_3 | conjugated octadecadienoic | 0.0001 ±0.0000 | 0.0000 ±0.0000 | 0.0006 ±0.0009 | 0.0014 ±0.0002 | 0.0002 ±0.0001 | 0.0005 ±0.0003 | 0,0006 ±0,0002 | 0,0003 ±0,0001 |
| C18:2, CLA_4 | conjugated octadecadienoic | 0.0034 ±0.0011 | 0.0009 ±0.0008 | 0.0067 ±0.0018 | 0.0151 ±0.0036 | 0.0016 ±0.0005 | 0.0049 ±0.0010 | 0,0049 ±0,0009 | 0,0025 ±0,0005 |
a,b — Means with different uppercase letters in the same row are significantly different (p < 0.05). For other fatty acids no statistically significant changes.
Table 3.
Composition of the external standard (BAME) and identification parameters for each fatty acid.
Table 3.
Composition of the external standard (BAME) and identification parameters for each fatty acid.
| fatty acid | acid name | RT [min] * | ECL | EI * |
|---|---|---|---|---|
| C11:0 | undecanoic | 15.466 | 11.000 | 74,87,143,157,200 |
| C12:0 | lauric / dodecanoic | 17.655 | 12.000 | 74,87,143,214 |
| C13:0 | tridecanoic | 19.751 | 13.000 | 74,87,143,185,228 |
| C14:0 | myristic / tetradecanoic | 21.757 | 14.000 | 74,87,143,199,242 |
| C10:0,2OH | 2-hydroxydecanoic | 22.658 | 14.481 | 69,83,143,228 |
| C15:0,iso | iso-13-methyltetradecanoic | 22.777 | 14.542 | 74,87,143,213,256 |
| C15:0,anteiso | anteiso-12-methyltetradecanoic | 23.084 | 14.701 | 74,87,143,213,256 |
| C15:0 | pentadecanoic | 23.673 | 15.000 | 74,87,143,213,256 |
| C16:0,iso | iso-14-methylpentadecanoic | 24.647 | 15.541 | 74,87,143,227,270 |
| C16:0 | palmitic / hexadecanoic | 25.505 | 16.000 | 74,87,143,227,270 |
| C17:0,iso | iso-15-methylhexadecanoic | 26.442 | 16.541 | 74,87,143,241,284 |
| C16:1,cis-9 | palmitoleic / hexadecenoic | 26.485 | 16.565 | 69,83,96,152,236 |
| C12:0,2OH | 2-hydroxydodecanoic | 26.636 | 16.653 | 69,83,97,171,230 |
| C17:0 | heptadecanoic | 27.255 | 17.000 | 74,87,143,241,284 |
| cycC17:0,cis-9,10 | cis-9,10-methylenehexadecanoic | 27.965 | 17.432 | 69,74, 83,97,250 |
| C18:0 | stearic / octadecanoic | 28.934 | 18.000 | 74,87,143,255,298 |
| C12:0,3OH | 3-hydroxydodecanoic | 29.175 | 18.159 | 71,74,83,103 |
| C18:1,trans-9 | elaidic / octadecenoic | 29.492 | 18.351 | 69,74,83,97,123,264 |
| C18:1,cis-9 | oleic / octadecenoic | 29.705 | 18.486 | 69,74,83,97,123,264 |
| C14:0,2OH | 2-hydroxytetradecanoic | 30.221 | 18.797 | 69,83,97,199 |
| C19:0 | nonadecanoic | 30.545 | 19.000 | 74,87,143,312 |
| C18:2,cis-9,cis-12 | linoleic / cis-9,cis12-octadecadienoic | 31.015 | 19.307 | 97,81,95,123,294 |
| cycC19:0,cis-9,10 | dihydrosterculic / cis-9,10-methylene-octadecanoic | 31.122 | 19.376 | 69,74,83,97,123,278 |
| C20:0 | eicosanic | 32.086 | 20.000 | 74,87,143,326 |
| C14:0,3-OH | 3-hydroxytetradecanoic | 32.592 | 20.318 | 71,74,103 |
| C16:0,2-OH | 2-hydroxyhexadecanoic | 33.475 | 20.864 | 69,83,97,227 |
* EI, electron ionization; RT, retention time.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



