
Submitted:
07 May 2023
Posted:
11 May 2023
You are already at the latest version
Abstract
Sperm morphology can predict the reproductive male fertilizing potential. This study aimed to determine the morphological and morphometric spermatozoa characteristics from guinea pigs subjected to different photoperiodic stimulation. Thirty F1 guinea pigs were randomly assigned to three photoperiodic treatments: FT1 (photoperiod with 10L/14D LED light), FT2 (photoperiod with 10L/14D sunlight), and FT0 (room without direct light source). At 107 ± 9.8 days of age, sperm concentration and motility were higher in FT0 and FT1 (p<0.05); furthermore, there were no differences in nucleus length and ellipticity between FT0 and FT1, but FT1 was higher in perimeter and nuclear area, while FT0 was higher in roughness, regularity, midpiece length and tail (p<0.01). Expanding acrosome (Type 2) was more frequent in FT2, but there was variation in head measurements between all morphological categories. Pregnancy rate, calving age and mating age were higher in FT0, meanwhile FT1 initiated successful matings earlier (p<0.01). FT0 had a higher fertility rate, and FT1 age of mating and first calving were earlier than FT0 but no pregnancies were reported for FT2. The photoperiodic stimulation can increase the morphometric dimensions of guinea pig spermatozoa, favoring the reproductive characteristics, but sunlight could reduce their size due to heat stress.
Keywords:
Cavia porcellus L.
; illumination
; morphometry
; acrosome
; sperm nucleus
; precocity
1. Introduction
Photoperiod regulates key physiological processes in animals, including reproduction, especially in seasonally reproducing species, which is regulated by an endogenous hormonal rhythm [1,2]. In mammals, this regulation may involve the activity of melatonin secreted by the pineal gland only at night [3]. Melatonin receptors are found in many structures and organs, such as the central nervous system, pituitary gland, granulosa cells of pre-ovulatory follicles, spermatozoa, etc. Melatonin concentration is related to the intensity and wavelength of light incident on the retina (high at night and low during the day) [4]. Light incidence on the eye's retina could trigger a response of the pituitary gland via the pars tuberalis for synchronisation with secretory organs involved in the reproductive activity [5,6].
One of the photoperiodic effects is observed in a marked reproductive seasonality of some species, which are found in regions with latitudes further away from the equator. For instance, variations in plasma testosterone concentration, testicular size, seminal production and sperm concentration have been reported in goats [2] and rams [7]. In higher latitude regions, species limit the birth and rearing to the less rugged season with higher food availability; for this purpose, photoperiod is a valuable signal indicating better environmental conditions, initiating ovulation and oestrus. It was reported that in non-seasonal breeding species, e.g. male guinea pigs, the sensitivity to photoperiod has been demonstrated, achieving higher body weight and testosterone concentration in a photoperiod of 16 hours of light and 8 hours of darkness at 25 °C compared to a photoperiod of 8 hours of light and 16 hours of darkness at 15 °C, showing greater precocity for sexual maturity by early peaks of testosterone in the blood of Cavia aperea f. porcellus [8] and Cavia aperea [9]. Similarly, in Cavia aperea females, increasing photoperiod may stimulate the onset of female reproduction, probably due to its role as a signal of more favorable environmental conditions [10].
Within sperm quality assessment, morphology is a characteristic able to predict the fertilizing capacity of a reproductive male, and the high rate of abnormalities could determine infertility [11,12]. In guinea pigs, the shape of the sperm head has already been classified according to the state of exocytosis of the acrosome [13], and Cabeza et al. [14] determined the dimensions of the head and tail by manual measurements, associating them with reproductive problems. Therefore, this work aims to determine guinea pig spermatozoa's morphological and morphometric characteristics subjected to different photoperiod treatments.
2. Materials and Methods
The study was conducted in Chachapoyas province, Amazonas region in the southern hemisphere, during the spring season, from September to December 2020. The area was located at an altitude of 2 339 meters above sea level, between the coordinates latitude/longitude 6°14'03''S / 77°51'07''W, in a temperate to cold climate zone, with an annual average temperature of 15 °C and 1 578 mm of rainfall (UNTRM-Amazonas Meteorological Station, 2016).
2.1. Experimental Design
The experiment was carried out under a split-plot complete randomized design, with three treatments (photoperiod-FT) and five experimental units (EU) each (EU=2 animals). The EU were F1 guinea pigs from a crossbred of four closely related males with three females, each. The mating and the F1 pups-lactation were carried out under homogeneous lighting (39.3 to 60.8 lux), under photoperiod conditions of 12 hours of light and 12 hours of darkness (12L/12D). An initial weighing was performed prior to treatment allocation to verify group homogeneity.
2.2. Animal Housing
A 6.0 x 4.5 x 2.5 breeding module made of fiber cement walls, a polypropylene roof and a curtain-based ventilation system was installed. In four cages of dimensions 1.0 x 1.0 x 0.45 meters, four five-month-old male guinea pigs were raised and bred with three females of the same age. All the animals were bought from a nearby commercial farm. Litters of six to eleven pups were born in each cage (mean of 2.9 pups per dam), which were weaned at 21 days. At the age of 26 ± 4 days and an average weight of 262 ± 60 g, thirty guinea pigs (17 males and 13 females) were randomly and paired distributed into fifteen cages of 1.0 x 0.5 x 0.45 m (13 mixed cages and 2 with only males). The cages were made of polyethene-coated wire mesh, and the floor of compacted soil. Each cage had a plate feeder and an automatic drinker.
2.3. Photoperiods
Three independent rooms of 2.8 x 1.5 x 2.3 m height, each containing five cages, were set up. Each room corresponded to each photoperiod (FT). The first FT had 10 hours of LED light (08:00 am to 06:00 pm) and 14 hours of darkness (10L/14D). The light was generated with a 62-light LED lamp of 3.1 W power and an average light intensity of 175.67 lux at 1.5 m from the source (CN-L862Y, CAFINI, China) (FT1). The second room had sunlight at an average light intensity of 1254.67 lux at 08:30 am and 476.17 lux at 04:30 pm (10L/14D). The entry of the light was managed by curtains opening from 08:00 am to 06:00 pm. Curtains were located on the eastern and north-eastern sides of the shed and 1.2 m above the floor (FT2). The third environment had no direct light source and no photoperiodic schedule (FT0), recording a mean light intensity of 2.36 lux at 08:30 am and 1.17 lux at 04:30 pm. The study area's day length during October, November, and December 2020 was 12:10 to 12:30 hours of light and 11:50 to 11:30 hours of darkness (Meteocast©). In all rooms, ambient temperature and relative humidity were recorded every 10 minutes with two Thermo hygrometers installed 10 cm above the ground (datalogger; HT71N, PCE Instruments, Germany).
Guinea pigs were fed a commercial concentrate based on yellow corn, soybean cake, wheat by-product, molasses, rice flour, rice by-product, micronutrients, vitamins, minerals, and B complex (18% crude protein and 2.8 Mcal/kg); alfalfa forage (Medicago sativa) and Guatemala grass (Tripsacum laxum). The daily ration of concentrate was 10% of live weight (recalculated weekly) together with green fodder, fed twice daily (08:00 to 09:00 am and 04:00 to 05:00 pm). For diarrhea prevention, Daimeton®T (Sulfamonomethoxine + Trimethoprim) was added to the morning feed (2 g per 1 kg) and B complex in the drinking water (1 g per liter) for the first three days. Drinking water was provided ad libitum by an automatic waterer.
2.4. Sperm Recovery and Analysis
At 107 ± 9.8 days of age, sixteen guinea pigs were slaughtered by transection of the carotid arteries and jugular vein, recording an approximate bleeding time of 30 seconds. This process was carried out following ARRIVE 2.0 (Animal Research: Reporting of In Vivo Experiments) and the American Veterinary Medical Association (AVMA) euthanasia guidelines [15,16]. Following exsanguination, the testes were removed by a cut on the left side of the scrotal pouch. The organs were placed in polyethylene bags and transported at 37 °C to the Laboratory of the Sperm Collection Centre of the Institute of Livestock and Biotechnology (IGBI - UNTRM). The caudal epididymis side was sectioned into a Petri dish on a 37 °C heat plate. The Petri dish contained 0.4 mL of tempered Tris medium (3.028 g molecular grade Tris base, 1.7 g citric acid, and 1.25 g D-fructose in 100 mL of distilled water) [17]. Slight cuts were made to promote the outflow of spermatozoa into the medium [18]. Then, 0.6 mL of Tris medium was added, recovered in 1.5 mL microtubes, and kept at 37 °C. The concentration and sperm motility was analyzed with a Makler chamber under a phase contrast microscope (CX31, OLYMPUS).
2.5. Sperm Morphology
For morphology analysis, a smear was made with a Hemacolor® blood cell differentiation stain (Merck, Germany) according to the manufacturer's protocol. Briefly, the smear was immersed in Fixation Solution I (methanol) five times for one second each, then in Solution II (eosin) three times for one second each, then in Solution III (Azure) six times for one second each, and finally in Wash Solution (pH 7.2) with two immersions of 10 seconds each, and then dried vertically at room temperature. For analysis, 50 spermatozoa per sample were captured at 100x with immersion oil, with a microscope (OLYMPUS BX53, Tokyo, Japan) equipped with a camera (OLYMPUS U-TV0.63XC, Tokyo, Japan). Cells were morphologically classified into four categories according to the state of acrosome exocytosis [13]; intact acrosome (1), expansion by acrosomal reaction (2), the onset of decomposition (3), and loss of acrosomal contents and matrix (4) (Figure 1). In addition, atypical shapes such as micronucleus, macronucleus, pyriform, elongated, and lanceolate were counted (Figure 2). Subsequently, the nucleus (length L, width W, area A and perimeter P), acrosome area and perimeter, head length, midpiece length and width, and tail length were measured (CellSens, OLYMPUS). In addition, dimensionless parameters of ellipticity (L/W), roughness (4πA/P2), elongation ([L-W]/[L+W]) and regularity (πLW/4A) of the sperm nucleus were calculated.
2.6. Fertility
Twelve EU consisted of one male and one female each. At the time of first calving, age at calving, probable age at mating (considering a mean of 68 days of gestation), pregnancy percentage, and number of offspring per calving in each FT room, were determined.
2.7. Statistical analysis
Normal distribution and homogeneity of variances were verified for live weight, testicular weight, sperm concentration, progressive motility, non-progressive motility, and total motility. Morphometric parameters did not meet the assumptions of normality (Kolmogorov-Smirnov test) and homogeneity of variances (Levene's test), which is why they were analyzed with the non-parametric Kruskal-Wallis test (p<0.05) for more than two groups and Mann-Whitney U test (p<0.05) for two groups. Age at mating, age at calving, and the number of offspring were evaluated with ANOVA and pregnancy rate with Chi-square test. Correlations were analyzed with Pearson's coefficients in SPSS v.15.0.
3. Results
Analysis of sixteen adult male guinea pigs of 107 ± 9.8 days of age showed non-significant differences in live and testicular weight between FT treatments (p>0.05), but FT0 and FT1 sperm concentrations were significantly higher than FT2 (p<0.05). Similarly, FT0 and FT1 sperm showed higher non-progressive motility and total motility compared to FT2 (p<0.05) (Table 1).
Morphometric variables of the nucleus, acrosome, midpiece, and tail were measured in 938 spermatozoa from 16 guinea pigs. Differences (p<0.01) were found in most parameters between FTs, except nucleus width (p>0.05) (Table 2). The length, ellipticity, and elongation of the nucleus of FT0 and FT1 spermatozoa did not differ from each other but were greater than those of FT2. FT1 spermatozoa had a larger nuclear perimeter and area than the other groups, but the midpiece length, and tail length of FT0 spermatozoa were significantly larger than in the other groups. The roughness and regularity of the nucleus differed between groups, being greater in FT0 and the midpiece width, which was significantly larger in FT2.
In addition, 6950 spermatozoa were morphologically classified according to the acrosome status and the nucleus abnormalities (micronucleus, macronucleus, pyriform, elongated, and lanceolate) (Table 3). The rate of nucleus abnormalities ranged from 0 to 8.95%, giving a high coefficient of variation; however, we found no significant differences between groups (p>0.05), and means did not reach 2% of abnormalities. We did not find significant differences (p>0.05) between Type 1, Type 3, and Type 4 spermatozoa among the FTs, but we did find a higher frequency of Type 2 morphology in FT2 guinea pigs (p<0.05).
Significant differences were found in most morphometric parameters, except nucleus roughness, midpiece width, and tail length (p>0.05). Larger nucleus dimensions were observed in Type 4 spermatozoa and smaller in Type 1 spermatozoa. The arrangement of the acrosome is the basis of the morphological classification; therefore, larger perimeters and acrosomal areas, and head length were observed in Type 2 spermatozoa (expanded acrosome), followed by Type 1, Type 3, and finally Type 4 (no acrosomal structure and therefore shorter head length) (p<0.01) (Table 4).
According to Pearson's coefficient, sperm classification did not correlate with live weight, testicular weight, concentration, and motility. Type 1 correlated negatively with Type 3 and Type 4, and Type 3 correlated positively with the highest frequency of sperm nucleus abnormalities (Table 5).
During the FT treatment period, each male guinea pig was housed with a female of the same age to homogenize sexual stimulation. At the end of the study, no FT2 females (0/5), all FT0 females (4/4), and half of the FT1 females (2/4) were pregnant. The chi-square test showed a significant association of pregnancy rate with FT (p<0.05), being higher in FT0, followed by FT1. However, age at parturition and probable age at mating in FT0 was significantly later than in FT1 (p<0.01), showing a higher reproductive earliness in FT1 guinea pigs, with a difference of approximately 23 days (Table 6).
Fertility variables were correlated with sperm characteristics, reported in Table 7. The pregnancy rate was correlated with progressive, non-progressive, and total motility. On the other hand, the number of offspring was positively correlated with live weight and non-progressive motility; probable age at mating was negatively correlated with testicular weight, non-progressive motility, and type 4 spermatozoa (without acrosome).This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
4. Discussion
This study evaluated the effect of three different light exposures (photoperiod with LED light or FT1, photoperiod with sunlight or FT2, and no direct light exposure or FT0) on sperm parameters in guinea pigs, where sperm parameters were lower in FT2. Reports showed that although there was no effect of FTs on live and testicular weights, male guinea pigs subjected to an 8L/16D photoperiod versus a 16L/8D photoperiod achieved lower weights, as well as inhibition of somatic and testicular growth in rats under the same photoperiod conditions [8,19]. The increased testicular growth could be related to the higher blood testosterone concentrations in seasonally and non-seasonally reproducing species influenced by photoperiod [2,7,8,9,20].
The FT0 sperm concentration and total motility are higher than those reported by Ayala Guanga et al. [21] (418.0 ± 57.0 x 106/mL and 58 ± 5.39 %), the total motility is lower than Rodriguez et al. [22] (95 %) and the concentration is higher than Ferdinand et al. [23] (149.85 ± 5.07 x 106/mL) in epididymal spermatozoa. Our findings are higher than those reported by Meza et al. [24] (43.87 ± 19.68 x 106/mL and 49.49 ± 10.54 %), Cabeza et al. [14] (47.33 x 106/mL and 69.40 %) and Benavides et al. [25] (36.7 ± 28.4 x 106/mL) for spermatozoa obtained by electroejaculation, although the latter reported higher motility (90.86 ± 6.64 %) than our study. FT1 live weight, testicular weight, and sperm concentration were higher than in FT0, although not significantly. In seasonally breeding species (sheep or goats), a photoperiodic effect on the testosterone levels, testicular volume, and sperm concentration has been demonstrated [2,7], attributed to the increase of the melatonin secretion due to reduced day length and light intensity in the retina of the eye [4,26]. Melatonin is a hormonal messenger in the environment-animal relationship for mammalian reproduction, as receptors were found in reproductive organs and spermatozoa [1]. Guinea pigs may be sensitive to photoperiod, as documented by Bauer et al. [8] and Trillmich et al. [10]. In these studies, the effect of light stimulation with long photoperiods of 14 and 16 hours was successful in anticipating male puberty. Bauer et al. [8] found earlier puberty and earlier serum testosterone peaks in a 16L/8D regime compared to an 8L/16D regime. In this study, the onset of sexual activity and successful pregnancies occurred earlier in FT1 (10L/14D) than in FT0 (no direct light stimulation). However, in FT2, sperm concentration and sperm motility were lower. In FT2, a 10L/14D photoperiodic regime was also used, similar to FT1, but with sunlight as the source.
In FT2, the direct incidence of solar radiation was from 8:00 to 10:00 am and only on 20 % of the surface of each cage. The rest of the day, the windows remained open (until 18:00) but without direct incidence of solar radiation, reaching higher temperature peaks in FT2 (33.8 °C) compared to FT1 (32.75 °C) and FT0 (32.65 °C). FT2 guinea pigs probably experienced frequent heat stress episodes caused by sunlight-generated temperature, which would explain the reduction in sperm concentration and motility. High temperatures in guinea pigs can alter thermoregulation, generating heat stress and, consequently, negative impacts on spermatogenesis and sperm quality, such as lower sperm concentration, lower individual motility, higher rate of abnormal spermatozoa and even DNA damage [27,28]. In rabbit spermatozoa, in vitro temperatures of 42°C reduced total motility values [29]. In this study, the temperature peaks at midday (33.8 °C in FT2 compared to the optimal scrotal temperature of 32.5 °C) probably compromised sperm functionality.
Morphometric analysis of guinea pig spermatozoa is currently performed manually, as their morphology makes automatic analysis in computerized analysis systems (CASA) difficult. In this study, the average values of length, width, perimeter and nucleus area, roughness, and regularity are higher than those reported by Cabeza et al. [14] in guinea pig spermatozoa obtained by electroejaculation and Yucra [30] in epididymal spermatozoa. However, they are lower in ellipticity, elongation, midpiece length, and tail length. The differences may be due to the staining method used, the measuring equipment, the technician, and even the presence of seminal plasma because Cabeza et al. [14] used Spermac® for staining and Motic Image Plus 2.0 software, while Yucra [30] used Diff-Quick and ISAS® CASA equipment for measurement and we used Hemacolor® and CellSens software from OLYMPUS on epididymal spermatozoa in this study. Staining can influence the actual sperm morphometry, either by osmosis or dehydration (20 % reduction in length and 13 % reduction in width) [31,32], but its use is vital to improve the contrast and definition of fixed spermatozoa for morphometric analysis [33,34].
In most of the morphometric parameters evaluated, FT2 presented smaller spermatozoa, except for the width of the midpiece. The incidence of higher temperature in FT2 probably produced heat stress and altered guinea pig spermatogenesis, increasing the frequency of microcephalic spermatozoa. Regarding to a larger midpiece, Malo et al. [35] argue that a bulky midpiece could hinder sperm motility, as well as a lower elongation could increase the degree of drag, making the sperm slower, even more so considering that the nucleus and head of guinea pig sperm are less elongated than in other species. On the other hand, a greater perimeter and nuclear area in FT1 could be related to the stimulation of spermatogenesis by increased melatonin and testosterone activity in males subjected to photoperiodic stimulation [4], similar to the improvement of other seminal variables seen in other mammalian species. FT0 guinea pigs had sperm with greater midpiece length and tail length. Although these animals may have had a later development than FT1 guinea pigs, greater elongation of these structures could generate more energy from more mitochondria and be positively associated with motility and swimming speed [34,35].
Morphological classification of guinea pig spermatozoa is based on the state of exocytosis of the acrosome and classified as: Type 1 or intact acrosome, Type 2 or expansion due to acrosomal reaction, Type 3 or the initiation of breakdown and Type 4 or loss of acrosomal content [13]. This study found a higher frequency of Type 2 spermatozoa in FT2. Similarly, in FT1 and FT0, the highest percentages corresponded to Type 1. All morphometric parameters differed among the classifications except for nucleus roughness, midpiece width, and tail length. The largest acrosome dimensions correspond to Type 2 as they are spermatozoa with reacted and expanding acrosome, and in Type 3, they are in decomposition. As for the larger dimensions of the nucleus in Type 4 spermatozoa, it is possible that the presence of the acrosome or acrosome residues could have influenced the staining of the head and the measurement of the nucleus; similarly, Hemacolor® could stain the overlapping nucleus and acrosome together, underestimating the nuclear dimensions. Therefore, differential staining between the two structures would be necessary for future morphological studies. Sperm morphology studies classify species as homomorphic or heteromorphic [36]. In the case of guinea pigs, Cabeza et al. [14] argue that the guinea pig is a homomorphic species due to the morphological homogeneity of the spermatozoa, especially at the level of the nucleus.
Pregnancy rate, calving age, and age at mating (considering a mean of 68 days of gestation) varied significantly among FTs. The absence of pregnancies in FT2 may be associated with low sperm motility values, possibly due to heat stress generated by the higher environmental temperature peaks recorded. On the other hand, although the pregnancy rate was higher in FT0, calving age and probable age at mating were lower in FT1. This difference of approximately 23 days between the two groups is a finding compatible with those of Bauer et al. [8] and Guenther et al. [9], who also found more remarkable sexual precocity in guinea pigs subjected to a photoperiod of more daylight hours, as evidenced by precocial puberty and early peaks of testosterone in the blood. The mean number of offspring was higher in FT1 than in FT0, although not significantly. In productive terms, this advance in precocity by photoperiodic stimulation could constitute an economic gain since, typically, the recommended mating age for females is three months and males four months; however, in this study, we experimentally achieved successful early matings with 2 to 4 pups at approximately two months of age using an artificial 10L/14D photoperiod. This photoperiodic stimulation could be included in small and medium-sized breedings to reduce intergenerational time and increase the number of births per year.
5. Conclusions
Stimulation with an LED light photoperiod may increase guinea pig sperm morphometric dimensions, but a sunlight photoperiod may reduce them, probably due to higher ambient temperature peaks that generate heat stress. A photoperiod with 10L/14D sunlight could generate heat stress, low spermiogram values, and absence of pregnancy. In contrast, a photoperiod with 10L/14D LED light improved some sperm characteristics, although not significantly compared to a compartment without any direct light stimulus. However, although the fertility rate was not improved in a photoperiod with LED light, pregnancies were earlier compared to guinea pigs housed in a compartment without direct light stimulation. An environment without direct light stimulation had the highest fertility rate, but animals under an artificial photoperiod with LED light showed earlier precocity and first parturition by up to 23 days. However, in a photoperiod with sunlight, no pregnancies were obtained.
Author Contributions
Conceptualization, H.A.Q.C. and I.S.C.C.; methodology, H.A.Q.C.; software, H.A.Q.C.; validation, I.S.C.C., Y.M.B.M.; formal analysis, H.A.Q.C.; investigation, H.A.Q.C.; resources, H.A.Q.C.; data curation, Y.M.B.M.; writing—original draft preparation, H.A.Q.C.; writing—review and editing, I.S.C.C. and Y.M.B.M.; visualization, Y.M.B.M.; supervision, I.S.C.C.; project administration, H.A.Q.C.; funding acquisition, H.A.Q.C.
Funding
The APC was funded by Vicerrectorado de Investigación of Universidad Nacional Toribio Rodríguez de Mendoza of Amazonas.
Institutional Review Board Statement
Ethical review and approval were waived for this study due to the institution is still in the process of implementing an ethics committee. However, all experiments were carried out according to the ARRIVE 2.0 guidelines and the AVMA manual for animal experimentation.
Informed Consent Statement
Not applicable.
Acknowledgments
We want to thank CONCYTEC-PROCIENCIA in the framework of the call "Proyecto de Investigación Básica 2019-01" [Contract No. 357-2019-FONDECYT] and to the Institute of Livestock and Biotechnology (IGBI - UNTRM Amazonas).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Correa, L.M.; Fernández, J.L. Influence of Melatonin on the physiology and the conduct of ungulates. Rev. Investig. Altoandin. 2017, 19(3), 337-350. [CrossRef]
- Zarazaga, L.A.; Gatica, M.C.; Celi, I.; Guzmán, J.L.; Malpaux, B. Effect of artificial long days and/or melatonin treatment on the sexual activity of Mediterranean bucks. Small Rumin. Res. 2010, 93, 110–8. [CrossRef]
- Chemineau, P. Medio Ambiente y Reproducción Animal. World Animal Review 1993, 77(1), 2–14. Retrieved from http://www.fao.org/ag/Aga/AGAP/FRG/FEEDback/War/v1650b/v1650b04.htm.
- Lincoln, G.A. Photoperiod-pineal-hypothalamic relay in sheep. Anim. Reprod. Sci. 1992, 28(1–4), 203–217. [CrossRef]
- Prendergast, B.J.; Nelson, R.J.; Zucker, I. Mammalian Seasonal Rhythms. In Hormones, Brain and Behavior. Pfaff, D.W., Arnold, A.P., Etgen, A.M., Fahrbach, S.E., Rubin, R.T. Eds., 2002, pp. 93–156. [CrossRef]
- Nakane, Y.; Yoshimura, T. 2019. Photoperiodic regulation of reproduction in vertebrates. Annu. Rev. Anim. Biosci. 2019, 7, 173-194. [CrossRef]
- Santiago-Moreno, J.; Gómez-Brunet, A.; González-Bulnes, A.; Toledano-Díaz, A.; Malpaux, B.; López-Sebastián, A. Differences in reproductive pattern between wild and domestic rams are not associated with inter-specific annual variations in plasma prolactin and melatonin concentrations. Domest. Anim. Endocrinol. 2005, 28(4), 416–29. [CrossRef]
- Bauer, B.; Womastek, I.; Dittami, J.; Huber, S. The effects of early environmental conditions on the reproductive and somatic development of juvenile guinea pigs (Cavia aperea f. porcellus). Gen. Comp. Endocrinol. 2008, 155, 680–5. [CrossRef]
- Guenther, A.; Palme, R.; Dersen, M.; Kaiser, S.; Trillmich, F. Photoperiodic effects on reproductive development in male cavies (Cavia aperea). Physiol. Behav. 2014, 123, 142-147. [CrossRef]
- Trillmich, F.; Mueller, B.; Kaiser, S.; Krause, J. Puberty in female cavies (Cavia aperea) is affected by photoperiod and social conditions. Physiol. Behav. 2009, 96(3), 476–80. [CrossRef]
- Maroto-Morales, A.; Ramón, M.; García-Álvarez, O.; Soler, A.J.; Esteso, M.C.; Martínez-Pastor, F.; Pérez-Guzmán, M.D.; Garde, J.J. Characterization of ram (Ovis aries) sperm head morphometry using the Sperm-Class Analyzer. Theriogenology 2010, 73(4), 437-448. [CrossRef]
- Yániz, J.L.; Capistrós, S.; Vicente-Fiel, S.; Soler, C.; de Murga, J.N.; Santolaria, P. Study of nuclear and acrosomal sperm morphometry in ram using a computer-assisted sperm morphometry analysis fluorescence (CASMA-F) method. Theriogenology 2014, 82(6), 921-924. [CrossRef]
- Kim, K.S.; Foster, J.A.; Gerton, G.L. Differential release of guinea pig sperm acrosomal components during exocytosis. Biol. Reprod. 2001, 64(1), 148-56. [CrossRef]
- Cabeza, U.; Ordoñez, C.; Meza, A.; Cucho, H. Morphological and morphometric characterization of the Guinea pig (Cavia porcellus). Spermova 2021, 10(2), 94-101. [CrossRef]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal research: reporting in vivo experiments: the ARRIVE guidelines. Br. J. Pharmacol. 2010, 160(7), 1577-79. [CrossRef]
- Everitt, J.I.; Gross, E.A. Euthanasia and Necropsy. In The Laboratory Rat. 2nd Ed., Suckow, M.A., Weisbroth, S.H., Franklin, C.L. Eds., Elsevier Academic Press, 2006 665–678. [CrossRef]
- Quispe, H.A.; Ciprián, R.; Ordoñez, C.; Ampuero, E.; Cucho, H. Hypoosmotic swelling test in alpaca (Vicugna pacos) spermatozoa recovered the vas deferens. Spermova 2015, 5(1), 10-14. [CrossRef]
- Ugarelli, A.; Evangelista-Vargas, S.; Santiani, A. Evaluation of acrosome integrity in epididimal alpaca sperm by flow cytometry. Rev. de Investig. Vet. del Perú 2017, 28(1), 130-40. [CrossRef]
- Heideman, P.D.; Sylvester, C.J. Reproductive photoresponsiveness in unmanipulated male Fisher 344 laboratory rats. Biol. Reprod. 1997, 57, 134–138. [CrossRef]
- Sancho, S.; Pinart, E.; Briz, M.; Garcia-Gil, N.; Badia, E.; Bassols, J.; Kádár, E.; Pruneda, A.; Bussalleu, E.; Yeste, M.; Coll, M.G.; Bonet, S. Semen quality of postpubertal boars during increasing and decreasing natural photoperiods. Theriogenology 2004, 62(7), 1271-1282. [CrossRef]
- Ayala Guanga, L.E.; Rodas Carpio, R.; Almeida, A.; Torres Inga, C.S.; Nieto Escandón, P.E. Influence of penis spicule of cavy (Cavia porcellus) on their sexual behavior, Fertility and sperm quality. Rev. Prod. Anim. 2017, 29(3), 36-42. http://scielo.sld.cu/pdf/rpa/v29n3/rpa06317.pdf.
- Rodríguez, G.J.; Barrios-Arpi, M.; Huanca, L.W.; Rodríguez, G.A.; Revuelta, L. Levels of glucose and triglycerides in seminal plasma ans spermatic motility in guinea pig fed with 10% more in digestible energy. Rev. Complut. Cienc. Vet. 2016, 10(1), 75-82. [CrossRef]
- Ferdinand, N.; Christiane, N.S.; Augustave, K.; D´Alex, T.C.; Raphael K.J.; … Joseph, T. Effect of Guava (Psidium guajava) leaves essential oil on some reproductive parameters in male guinea pig (Cavia porcellus). Biol. Syst. Open Access 2014, 3, 25. [CrossRef]
- Meza, A.; Huanca, N.; Aragón, S.; Cucho, H. Domestic guinea pig semen collection protocol (Cavia porcellus) by electroejaculation method. Spermova 2018, 8(1), 94.
- Benavides, F.; Sutovsky, P.; López, V.; Kennedy, C.; Echevarría, L. Semen parameters of fertile guinea pigs (Cavia porcellus) collected by transrectal electroejaculation. Animals 2020, 10, 767. [CrossRef]
- Kozioł, K.; Broda, D.; Romerowicz-Misielak, M.; Nowak, S.; Koziorowski, M. Melatonin concentration in peripheral blood and melatonin receptors (MT1 and MT2) in the testis and epididymis of male roe deer during active spermatogenesis. Theriogenology 2020, 149, 25-37. [CrossRef]
- Ngoula, F.; Tekam, M.G.; Kenfack, A.; Tchingo, C.D.A.T.; Nouboudem, S.; Ngoumtsop, H., ...Tchoumboue, J. Effects of heat stress on some reproductive parameters of male cavie (Cavia porcellus) and mitigation strategies using guava (Psidium guajava) leaves essential oil. J. Therm. Biol. 2017, 64, 67-72. [CrossRef]
- Durairajanayagam, D.; Sharma, R.K.; du Plessis, S.S.; Agarwal, A. Testicular heat stress and sperm quality. In Male Infertility, 1st ed.; du Plessis, S., Agarwal, A., Sabanegh, Jr. E.S., Eds.; Springer New York, NY, USA, 2014, 105-125. [CrossRef]
- Sabés-Alsina, M.; Tallo-Parra, O.; Mogas, M.T.; Morrell, J.M.; Lopez-Bejar, M. Heat stress has an effect on motility and metabolic activity of rabbit spermatozoa. Anim. Reprod. Sci. 2016, 173, 18-23. [CrossRef]
- Yucra, A. 2013. Características microscópicas del semen y morfometría del espermatozoide del cuy doméstico (Cavia porcellus) y silvestre (Cavia tschudii). Thesis of bachelor, Universidad Nacional de San Antonio Abad del Cusco, Cusco-Peru, 2013.
- Ball, B.A.; Mohammed, H.O. Morphometry of stallion spermatozoa by computer-assisted image analysis. Theriogenology 1995, 44, 367–377. [CrossRef]
- Hidalgo, M.; Rodríguez, I.; Dorado, J. Influence of staining and sampling procedures on goat sperm morphometry using the Sperm Class Analyzer. Theriogenology 2006, 66, 996-1003. [CrossRef]
- Verstegen, J.; Iguer-Ouada, M.; Onclin, K. Computer assisted semen analyzers in andrology research and veterinary practice. Theriogenology 2022, 57, 149–179. [CrossRef]
- Yániz, J.L.; Soler, C.; Santolaria, P. Computer assisted sperm morphometry in mammals: a review. Anim. Reprod. Sci. 2015, 156, 1-12. [CrossRef]
- Malo, A.F.; Gomendio, M.; Garde, J.; Lang-Lenton, B.; Soler, A.J.; Roldan, E.R. Sperm design and sperm function. Biol. Lett. 2006, 2(2), 246-249. [CrossRef]
- Soler, C.; Sancho, M.; García, A.; Fuentes, M.C.; Núñez, J.; Cucho, H. Ejaculate fractioning effect on llama sperm head morphometry as assessed by the ISAS® CASA system. Reprod. Domest. Anim. 2014, 49(1), 71-78. [CrossRef]
Figure 1.
Morphology of guinea pig spermatozoa with Hemacolor® staining. Sperm morphology was classified as: acrosome intact, dark purple and compact in appearance (a), acrosome expanded and light purple (b), acrosome in the process of decomposition (c), and no acrosomal content or matrix (d). Bar represents 10 μm, measure was taken under 100x and oil immersion.
Figure 1.
Morphology of guinea pig spermatozoa with Hemacolor® staining. Sperm morphology was classified as: acrosome intact, dark purple and compact in appearance (a), acrosome expanded and light purple (b), acrosome in the process of decomposition (c), and no acrosomal content or matrix (d). Bar represents 10 μm, measure was taken under 100x and oil immersion.
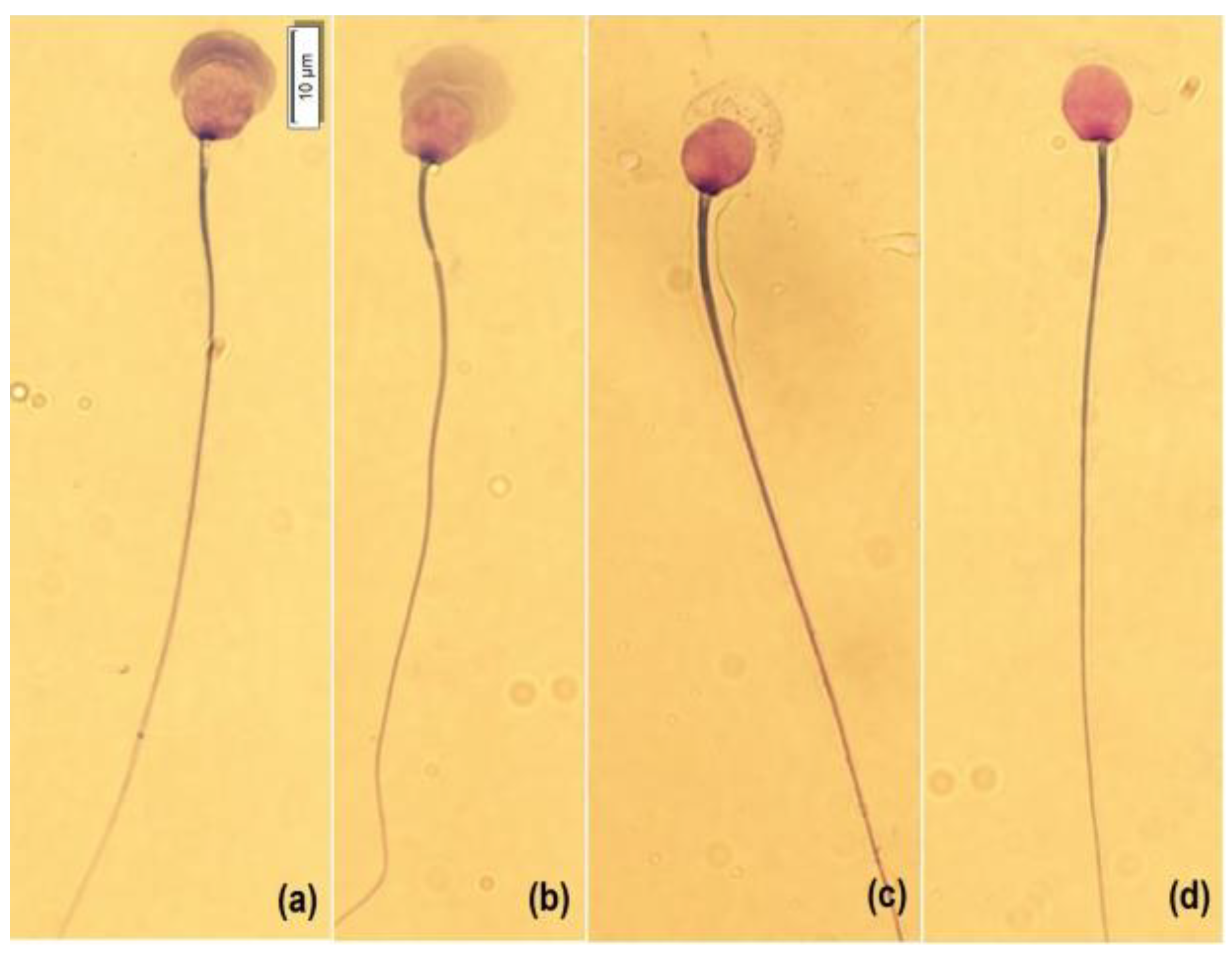
Figure 2.
Atypical forms in guinea pig spermatozoa with Hemacolor® staining. Normal (a), spermatozoa with macronucleus (b), micronucleus (c), pyriform (d), elongated (e) and lanceolate (f). Bar represents 10 μm, images was taken under 100x and oil immersion.
Figure 2.
Atypical forms in guinea pig spermatozoa with Hemacolor® staining. Normal (a), spermatozoa with macronucleus (b), micronucleus (c), pyriform (d), elongated (e) and lanceolate (f). Bar represents 10 μm, images was taken under 100x and oil immersion.


Table 1.
Sperm analysis of guinea pigs (107 ± 9.8 days of age) subjected to photoperiod treatment.
| Variable | FT0 | FT1 | FT2 | p-valor | SEM | CV |
|---|---|---|---|---|---|---|
| N | 6 | 5 | 5 | |||
| Live weight (g) | 1163.18 | 1264.38 | 1036.82 | 0.14 | 43.15 | 14.94 |
| Testicle weight (g) | 12.24 | 14.72 | 13.70 | 0.60 | 0.87 | 25.88 |
| Concentration (M/mL) | 915.23ab | 1151.10a | 456.19b | 0.04* | 108.40 | 51.28 |
| Progressive motility (%) | 29.44 | 22.88 | 15.61 | 0.27 | 3.32 | 57.61 |
| Non progressive motility (%) | 50.19a | 48.49a | 13.89b | 0.02* | 6.10 | 63.65 |
| Total motility (%) | 79.63a | 71.37a | 29.50b | 0.01* | 8.07 | 52.56 |
FT0: No direct light stimulation, FT1: Photoperiod with LED light 10L/14D, FT2: Sunlight Photoperiod 10L/14D. SEM: standard error of mean, CV: Variation coefficient. M/mL: Millions of sperm per milliliter. Different superscript letters in rows (a, b) indicate significant differences. *: Significant differences at level p<0.05.
Table 2.
Morphometric analysis of guinea pig spermatozoa subjected to photoperiod treatment.
| Traits | FT0 | FT1 | FT2 | Test K-W |
SEM | CV |
|---|---|---|---|---|---|---|
| N | 314 | 280 | 344 | |||
| Core length (µm) | 8.25a | 8.25a | 8.16b | <0.01** | 0.013 | 4.72 |
| Core width (µm) | 7.34 | 7.37 | 7.36 | 0.83 | 0.011 | 4.49 |
| Core perimeter (µm) | 26.10b | 26.40a | 26.21b | <0.01** | 0.033 | 3.86 |
| Core area (µm2) | 48.93b | 49.94a | 49.16b | <0.01** | 0.123 | 7.64 |
| Ellipticity | 1.125a | 1.121a | 1.110b | <0.01** | 0.002 | 5.28 |
| Rugosity | 0.901a | 0.899b | 0.898c | <0.01** | 0.000 | 1.05 |
| Elongation | 0.058a | 0.056a | 0.051b | <0.01** | 0.000 | 47.83 |
| Regularity | 0.972a | 0.957c | 0.961b | <0.01** | 0.000 | 2.48 |
| Acrosome perimeter (µm) | 33.83 | 33.98 | 33.56 | 0.05 | 0.113 | 8.35 |
| Acrosome area (µm2) | 79.11 | 79.92 | 77.98 | 0.05 | 0.569 | 18.04 |
| Head length (µm) | 12.25 | 12.31 | 12.31 | 0.68 | 0.035 | 6.41 |
| Midpiece length (µm) | 11.45a | 11.17b | 11.14b | <0.01** | 0.024 | 6.64 |
| Midpiece width (µm) | 0.63c | 0.65b | 0.72a | <0.01** | 0.003 | 17.97 |
| Tail length (µm) | 89.66a | 88.72b | 89.43b | <0.01** | 0.289 | 8.05 |
FT0: No direct light stimulation, FT1: Photoperiod with LED light 10L/14D, FT2: Sunlight Photoperiod 10L/14D. SEM: standard error of mean, CV: Variation coefficient. Different superscript letters in rows (a, b, c) indicate significant differences. **: Significant difference at level p<0.01.
Table 3.
Morphological classification of guinea pig spermatozoa according to acrosomal status, subjected to photoperiod treatment.
Table 3.
Morphological classification of guinea pig spermatozoa according to acrosomal status, subjected to photoperiod treatment.
| Category | FT0 | FT1 | FT2 | p-valor | SEM | CV |
|---|---|---|---|---|---|---|
| N | 2903 | 2043 | 2004 | - | - | - |
| Type 1 (%) | 62.22 | 47.67 | 38.87 | 0.06 | 5.12 | 40.66 |
| Type 2 (%) | 7.27a | 11.53ab | 26.16b | 0.04* | 3.93 | 108.49 |
| Type 3 (%) | 11.81 | 11.97 | 9.67 | 0.62 | 1.54 | 54.94 |
| Type 4 (%) | 18.70 | 28.83 | 25.30 | 0.50 | 3.88 | 64.79 |
| Abnormal (%) | 1.84 | 1.64 | 1.50 | 0.81 | 0.55 | 131.56 |
FT0: No direct light stimulation, FT1: Photoperiod with LED light 10L/14D, FT2: Sunlight Photoperiod 10L/14D. SEM: standard error of mean, CV: Variation coefficient. *: Significant difference at level p<0.05.
Table 4.
Morphometric traits of guinea pig spermatozoa according to morphological classification.
| Traits | Type 1 | Type 2 | Type 3 | Type 4 | Test K-W | SEM | CV |
|---|---|---|---|---|---|---|---|
| N | 447 | 194 | 101 | 196 | |||
| Core length (µm) | 8.211ab | 8.170b | 8.199ab | 8.279a | 0.03* | 0.013 | 4.72 |
| Core width (µm) | 7.301b | 7.424a | 7.375a | 7.406a | <0.01** | 0.011 | 4.49 |
| Core perimeter (µm) | 26.081b | 26.342a | 26.313ab | 26.426a | <0.01** | 0.033 | 3.86 |
| Core area (µm2) | 48.730b | 49.709a | 49.578ab | 50.131a | <0.01** | 0.123 | 7.64 |
| Ellipticity | 1.126a | 1.102b | 1.113ab | 1.119a | <0.01** | 0.002 | 5.28 |
| Rugosity | 0.899 | 0.899 | 0.899 | 0.901 | 0.15 | 0.000 | 1.05 |
| Elongation | 0.059a | 0.048b | 0.053ab | 0.056a | <0.01** | 0.001 | 47.83 |
| Regularity | 0.967a | 0.959b | 0.959b | 0.962b | <0.01** | 0.001 | 2.48 |
| Acrosome perimeter (µm) | 33.283b | 35.209a | 29.629c | - | <0.01** | 0.113 | 8.35 |
| Acrosome area (µm2) | 76.446b | 86.124a | 58.584c | - | <0.01** | 0.569 | 18.04 |
| Head length (µm) | 12.105b | 12.766a | 11.212c | 8.279d | <0.01** | 0.035 | 6.41 |
| Midpiece length (µm) | 11.297a | 11.318a | 11.254ab | 11.086b | <0.01** | 0.024 | 6.64 |
| Midpiece width (µm) | 0.665 | 0.673 | 0.685 | 0.666 | 0.41 | 0.004 | 17.97 |
| Tail length (µm) | 89.201 | 88.715 | 89.342 | 90.011 | 0.06 | 0.289 | 8.05 |
SEM: standard error of mean, CV: Variation coefficient. Different superscript letters in rows (a, b, c, d) indicate significant differences. *: Significant difference at level p<0.05. **: Significant difference at level p<0.01.
Table 5.
Pearson´s correlation for morphological classification and sperm variables of guinea pigs.
| Type 1 | Type 2 | Type 3 | Type 4 | Abnormalities | |
|---|---|---|---|---|---|
| Live weight | 0.00 | 0.00 | 0.08 | -0.03 | -0.07 |
| Testicular weight | -0.24 | 0.46 | 0.10 | -0.11 | -0.19 |
| Concentration | 0.22 | -0.01 | 0.00 | -0.28 | 0.14 |
| Progressive motility | 0.29 | -0.07 | -0.16 | -0.26 | 0.06 |
| Non Progressive motility | -0.07 | 0.08 | 0.23 | -0.08 | 0.05 |
| Total motility | 0.07 | 0.03 | 0.11 | -0.17 | 0.06 |
| Type 1 | -0.61* | -0.38 | -0.56* | -0.21 | |
| Type 2 | 0.11 | -0.26 | 0.25 | ||
| Type 3 | -0.01 | 0.63* | |||
| Type 4 | -0.24 |
*: Significant correlation at level p<0.05 (two-sided).
Table 6.
Fertility of guinea pigs under different light exposure, subjected to photoperiod treatment.
Table 6.
Fertility of guinea pigs under different light exposure, subjected to photoperiod treatment.
| Variable | FT0 | FT1 | FT2 | p-valor |
|---|---|---|---|---|
| N | 4 | 4 | 5 | |
| Pregnancy rate (%) | 100.0a | 50.0b | 0.0c | 0.01* |
| N° offspring | 1.5 | 3.0 | - | 0.13 |
| Calving age (days) | 152.5±4.2b | 129.5±4.2a | - | <0.01** |
| Probable mating age (days) | 84.5±4.2b | 61.5±4.2a | - | <0.01** |
FT0: No direct light stimulation, FT1: Photoperiod with LED light 10L/14D, FT2: Sunlight Photoperiod 10L/14D. Different superscript letters in rows (a, b, c) indicate significant differences. *: Significant difference at level p<0.05; **: Significant difference at level p<0.01.
Table 7.
Pearson´s correlation for fertility and sperm variables of guinea pigs.
| Pregnancy rate | N° offspring | Mating age | |
|---|---|---|---|
| Live weight | 0.38 | 0.83* | -0.71 |
| Testicular weight | -0.12 | 0.65 | -0.99** |
| Concentration | 0.33 | 0.70 | -0.19 |
| Progressive motility | 0.65* | -0.54 | -0.01 |
| Non Progressive motility | 0.67* | 0.88* | -0.82* |
| Total motility | 0.73** | -0.10 | -0.51 |
| Type 1 | 0.25 | -0.33 | 0.63 |
| Type 2 | -0.43 | -0.21 | -0.15 |
| Type 3 | 0.06 | 0.08 | -0.12 |
| Type 4 | 0.08 | 0.57 | -0.83* |
| Abnormalities | 0.10 | -0.55 | 0.37 |
**: Significant correlation at level p<0.01 (two-sided); *: Significant correlation at level p<0.05 (two-sided).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



