
Submitted:
04 May 2023
Posted:
11 May 2023
You are already at the latest version
Abstract
Down syndrome (DS) is a leading human genomic abnormality resulting from the trisomy of chromosome 21. The genomic base of the aneuploidy behind this disease is complex, and this complexity poses formidable challenges to understanding the underlying molecular basis. In the spectrum of the classic DS risk factor associations the role of nutrients, vitamins, and in general, the foodborne associated background as part of the events leading ultimately to chromosome nondisjunction has long been recognized as a well-established clinical association. The integrity of the microbiome is a basic condition in these events, and the dysbiosis may be associated with secondary health outcomes, the possible association of DS development with maternal gut microbiota should require more attention. We have hypothesized that different classes of antibiotics might promote or inhibit the proliferation of different microbial taxa and hence, we might find associations between the use of different classes of antibiotics and the prevalence of DS through the modification of the microbiome. As antibiotics are considered major disruptors of the microbiome, it could be hypothesized that the consumption/exposure of certain classes of antibiotics might be associated with the prevalence of DS in European countries (N=30). Utilizing three different statistical methods, comparisons have been made between the average yearly antibiotic consumption (1997-2020) and the prevalence of people living with DS estimated for 2019 as a percentage of the population in European countries. We have found strong statistical correlations between the consumption of tetracycline (J01A) and the narrow-spectrum, beta-lactamase-resistant penicillin (J01CF) and the prevalence of DS.
Keywords:
Down syndrome [DS]
; microbiome
; dysbiosis
; antibiotics
; tetracycline
; penicillin
1. Introduction
Down syndrome (DS) is the most common genomic disorder of intellectual disability and is caused by trisomy of human chromosome 21 (HSA21) The presence of a supernumerary chromosome 21, results in a collection of clinical features commonly known as DS. Individuals suffering from DS are also more likely to develop different health conditions, including hypothyroidism, autoimmune diseases, obstructive sleep apnea, epilepsy, hearing and vision problems, hematological disorders including leukemia, recurrent infections, anxiety disorders, early-onset Alzheimer’s disease (AD) and congenital heart diseases, like atrioventricular septum defect, patent ductus arteriosus and tetralogy of Fallot [1]. DS occurs in all populations and countries, but differences in maternal age at conception between regions and ethnicities influence the number of live births. In Europe, DS prevalence in live births has been decreasing slightly since the 1990s, although there are substantial regional differences [2]. The signs of DS usually occur during prenatal development [3]. The average live birth with Down syndrome for 100000 delivery, estimated by the WHO [4] and calculated for 1997-2020 in 24 European countries is 99.872, but considerable differences could be observed (the lowest is Austria: 15.735, the highest is Latvia: 215.433). There are well-documented, classic events in the pathogenesis of the DS like maternal age, chromosome nondisjunction, and influence of paternal factors, all of which are results of early DS research. Unanswered questions have also been addressed by methods in the recent genomic medicine era, like robust genome-wide association studies which have been performed to identify the genomic susceptibility factors behind the mother’s predisposition. However, it is still clear that there are numerous factors outside of the human genome influencing DS susceptibility in pregnancy.
The role of the human microbiome in health and diseases had been widely documented in the literature. Several factors are capable of modifying the composition of the human microbiome, resulting in dysbiosis, and as a consequence, the development of different diseases is reported in publications [5,6].
The most effective microbiome modifiers are antibiotics, which can enter humans (animals) as therapeutic agents, or from the environment as the result of antibiotic pollution.
The “growth-promoting” effect of the antibiotics was first recorded in the late 40’-s when chicks were accidentally fed with culture media used for cultivating antibiotic-producing organisms and they started growing faster than the others. Experiments later confirmed this observation [7,8]. The growth-promoting effect of antibiotics could not have been reproduced in germ-free animals [9]. It was concluded that the “collateral” effect of antibiotics might appear through the presence of bacterial flora. This observation led to the industrial utilization of antibiotics, mixed with animal fodder to enhance the growth of food animals and later it also facilitated the emergence of polyresistant microbes. [10].
In normal circumstances, intestinal flora starts developing quickly after birth. Humans are colonized by the usual microbiota with the well-known functions that protect human health, development, and disease resistance. Several publications have indicated that Bifidobacterium and Bacteroides species are maternally transmitted to infants, but when the maternal transmission is disrupted either by cesarean section or by antibiotic exposure around birth, it is associated with a higher incidence of pathogen colonization and immune-related disorders in children, like allergies, asthma, etc. [11].
Microbes might be transmitted from the placenta, amniotic fluid, and umbilical cord blood which provides a diverse spectrum of bacterial exposures to the developing fetus [12]. Shortly after birth, or even before, the neonatal gut is rapidly colonized by facultative anaerobic bacteria typically including strains of Enterobacter, Enterococcus, Staphylococcus, and Streptococcus genera. [13,14]. Breastfeeding have long-term effects on the microbiome and its effects on the immune system and the gastrointestinal (GI) tract. [15].
The human GI tract harbors a rich and complex microbial communities in healthy individuals. Intestinal microbes possess genes that encode thousands of microbial enzymes and metabolites. Microbes colonizing the human body change in composition as humans getting older, with a gradual increase in microbial diversity during childhood and relative stabilization during adolescence and adulthood. The amount of the microbiome might be calculated as 2-3% of the body weight. The recently estimated number of bacterial cells in the microbiome in the 70 kg "reference man" is 3.8·1013 [16].
Novel technologies allowed scientists to phylogenetically determine and/or quantify the components of the gut microbiota by analyzing nucleic acids (DNA and RNA) which is directly extracted from stools. Many of these technologies are based on the extraction of DNA and the amplification of the 16S ribosomal RNA gene (rRNA) [17,18]. 16S rRNA sequencing became the most useful technique to demonstrate the diversity and abundance of the microbiome. The 16S rRNA gene sequences can be used with polymerase chain reaction (PCR) and metagenomics sequencing to characterize the microbial strains [19]. The gut microbiota encodes over 3 million genes producing thousands of metabolites, whereas the human genome includes approximately 23,000 genes [20]. Gut microbiota is composed of several species of microorganisms, including bacteria, yeast, and viruses. Taxonomically, bacteria can be classified according to phyla, classes, orders, families, genera, and species. Only a few phyla are represented, accounting for more than 160 species. The dominant gut microbial phyla are Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria, Fusobacteria, and Verrucomicrobia, with the two phyla Firmicutes and Bacteroidetes representing 90% of gut microbiota. The Firmicutes phylum is composed of more than 200 different genera such as Lactobacillus, Bacillus, Clostridium, Enterococcus, and Ruminicoccus. Clostridium genera represent 95% of the Firmicutes phyla. Bacteroidetes consist of predominant genera such as Bacteroides and Prevotella. The Actinobacteria phylum is proportionally less abundant and mainly represented by the Bifidobacterium genus [21]. Detailed analyses indicated that certain bacterial taxa, called core microbiomes, are always present in adults. The so called core microbiome is composed of Bacteroides, Eubacterium, Faecalibacterium, Alistipes, Ruminococcus, Clostridium, Roseburia, and Blautia; with Faecalibacterium prausnitzii, Oscillospira guillermondii, and Ruminococcus obeum as the top three taxa shared by all adults. [22].
The gut microbiota protects the health of the host by providing sufficient benefits in the form of immune system development, prevention of infections, nutrient acquisition, and perhaps even brain and nervous system functionality [23].
Althogh microbiome might vary among healthy individuals, microbial functions are maintained and associated with the production of microbial metabolites influencing both microbes and host. It might be considered that the modification of the microbiome results in the generation of different molecules which might promote the development of different ailments through the gut-brain axis [GBA], or other mechanisms. The imbalance of the human microbiome is called dysbiosis, which is the change in abundance and the diversity of the microbial taxa of the microbiome. This association makes dysbiosis the main concept for understanding how the human microbiota contributes to health and disease [24,25]. The gut-brain axis involves two-way interplay between the central nervous sytem and the gut including the gut-associated immune system, the enteric neuroendocrine system (ENS), and the gut microbiome. The view of gut-brain interactions has explained the pathophysiology of several brain disorders that had previously been considered exclusively to pathophysiological processes limited only to the brain. Research in the past two decades indicated disorders of altered GB interactions as well as psychiatric and neurological disorders such as depression, anxiety, AD, Parkinson’s disease (PD), and autism spectrum disorder (ASD) [26].
Bacterial metabolites produced by the microbiome can also support host processes and functionally complete host metabolic capabilities. A recent review article by Ruan et al, summarized the bacterial molecular products which are known as quorum sensors, influencing bacterial homeostasis, growth, spore formation, programmed cell death, virulence, and biofilm formation. Microbes can produce biologically active molecules including but not restricted to, gamma-aminobutyric acid (GABA), tryptophan metabolites, polyamines, and histamine. Microbial neuromodulators like GABA can communicate with the enteric and central nervous systems, and microbial-derived immunomodulators like histamine can interact with intestinal immune cells. Serpins, which are microbial-produced immunomodulators, are similar to eukaryotic serine protease inhibitors and they inhibit inflammatory responses by suppressing elastase activity. Lactobacillus-secreted lactocepins are bacterial enzymes that can downregulate pro-inflammatory signals like lymphocyte recruiting chemokine and can suppress pro-inflammatory signaling cascades. Short-chain fatty acids (SCFAs) are involved in immune regulation, pH regulation, sodium and water absorption, etc. The most abundant and well-studied SCFAs are acetate, propionate, and butyrate; however, the intestinal composition of SCFAs is contingent on microbial composition, diet, and intestinal pH. Outer membrane vesicles (OMVs) are another key immunomodulatory factor the gut microbiota produces [27].
Dysbiosis had been observed in several diseases, and the permanent question is that, whether dysbiosis is a consequence or associated with the development of the disease. Clinical observations and experiments indicated that dysbiosis could be observed long before the clinical manifestation of the disease. Autism spectrum disorder (ASD) seems to be related to unusual compositions of the gut microbiome indicating that Clostridia, Desulfovibrio, Sutterella, and Bacteroidetes are elevated in the stool of autistic children. In contrast, Firmicutes, Prevotella, and Bifidobacteria have been noted to be decreased [28,29,30]. The gut microbiota plays a role in the development and the progression of obesity also. Germ-free mice that receive fecal microbes from obese humans gain more weight than mice that received microbes from healthy-weight humans. A large study of UK twins found that the genus Christensenella was rare in overweight people and when given to germ-free mice prevented weight gain [31]. A higher ratio of Firmicutes to Bacteroides/Prevotella in obese people augments the microbial genes involved with the degradation of polysaccharides and increases the level of SCFAs [32]. Appropriate amount of knowledge suggests that a modulation in the gut microbiome (GM) composition may play an important role in the pathogenesis of PD by facilitating the characteristic ascending neurodegenerative spread of α-synuclein aggregates from the enteric nervous system to the brain [33].
The knowledge of the role of the gut microbiome in modulating brain function has rapidly increased over the past decade, but mainly in animal experiments. Increasing number of clinical and preclinical studies consider the microbiome as a possible key susceptibility factor for neurological disorders, including AD, ASD, MS, PD, and stroke [34].
Antibiotics are considered major modifier of the microbiome. The gut flora of newborn babies and young children is particularly sensitive to antibiotic exposure. Studies have indicated that preterm infants receiving prolonged antibiotic treatment had less diverse bacterial populations and reduced species abundance in their gut and more antibiotic-resistance genes [35]. A detailed review of the literature on the effects of antibiotics influencing the gut flora can be found in reference [36,37,38].
It is important to notice when we talk about the association between the use of antibiotics and the development of certain diseases that different types of antibiotics might augment, or inhibit the proliferation of different microbial taxa and hence it can support, or inhibit the development of different diseases through the modification of the gut flora via the gut-brain axis. Comparative analysis of the consumption of different types of antibiotics and the prevalence/incidence of certain metabolic, neurodegenerative diseases, and malignancies in European countries showed a significant association between antibiotic consumption patterns and the prevalence/incidence of different diseases. It has been estimated that in countries with outstanding consumption of narrow-spectrum penicillin, and tetracycline the incidence/prevalence of type I. diabetes (T1DM), PD, multiple sclerosis (MS), and ASD along with certain malignancies (melanoma, breast cancer, colorectal cancer), are higher than in other countries. Countries with high consumption of broad-spectrum antibiotics recorded a high prevalence of childhood obesity, AD, and type II. diabetes (T2DM) [39].
The modification of the gut microbiome in DS patients has also been reported [40,41,42]. Recent papers have indicated, that pregnant mothers, delivering babies with DS showed an altered microbiome compared to the control group. Clostridiaceae, Parteurellacae, and Pastereullaes were more abundant in DS pregnant, compared to the controls in this pilot study [42], and this alteration might occur in the newborn babies as well, even if this was not investigated in these studies.
2. Objectives/Hypothesis
Based on the above-detailed background, we have hypothesized that the alteration of the maternal microbiome might be a factor inducing the development of DS. Antibiotics are considered a major disruptor of the human microbiome, entering humans as therapeutic agents or from the environment, as pollutants. The effect of antibiotics on the gut flora is well-known [36]. We have suspected that different classes of antibiotics might promote or inhibit the proliferation of different microbial taxa and hence, we might find associations between the use of different classes of antibiotics and the prevalence of DS through the modification of the microbiome.
3. Methods
We calculated the average yearly antibiotic consumption in Defined Daily Dose/1000 inhabitants/ day (DID) between 1997 and 2020 of different classes of antibiotics covering 94% of the total, systemic antibiotic consumption in the community featured in the ECDC database [43] and comparison was performed with the prevalence of people living with DS estimated for 2019 in 30 European countries [44].
4. Statistics
Statistical tests were performed using IBM SPSS 26 software package. Pearson correlation was applied to estimate the correlation between antibiotic consumption and the prevalence of DS. A significant correlation (positive/negative) was considered when p values were ≤ 0.05. A non-significant correlation was estimated when the p values fall between 0.051-0.09. Positive (supportive) and negative (inhibitor) significant correlations were considered and evaluated. Ordinal logistic regression analysis was performed to determine the odds ratio and confidence intervals (OR, CI 95%) for each antibiotic class. The Kruskal-Wallis test was performed to determine statistically significant differences among the groups formed by country ranking. A significant difference was considered when p values were ≤ 0.05 and a non-significant difference was considered when p values were between 0.051-0.09.
5. Results
Statistical results are included at the bottom line of Table 1 and the significant associations are marked with yellow color. Related diagrams are shown in Figure 1 and Figure 2. We have observed a significant positive association between the consumption of tetracycline [J01A, Pearson p: 0.007, OR p: 0.005, Kruskal-Wallis p: 0.034], narrow-spectrum, beta-lactamase-resistant penicillin [J01CF, Pearson p: 0.077, OR p: 0.021, Kruskal-Wallis: 0.152 /non-significant/, and the prevalence of DS in 30 European countries. A strong, significant correlation has been detected with all three statistical methods regarding the use of tetracycline and the prevalence of inhabitants living with DS, while the use of the narrow spectrum penicillin showed a non-significant correlation with the prevalence of DS using Pearson correlation. The OR p indicated an elevated risk of DS with the use of narrow-spectrum penicillin, but the Kruskal-Wallis test did not find any association between the databases. This difference is well demonstrated in Figure 1 and Figure 2 as the diagram showing the association between the use of tetracycline and DS is steeper than the one indicating the association between the use of penicillin and DS.
6. Discussion
Almost 95 % of DS originates from a full trisomy of chromosome 21. The remaining DS cases are due to either mosaicism for chromosome 21 or the inheritance of a structural rearrangement leading to partial trisomy. Full trisomy 21 and mosaicism originate from errors in cell divisions during the development of the egg, sperm or embryo. Full trisomy for chromosome 21 should be further divided into cases of maternal origin, which is the majority, and cases of paternal origin, less than 10 % [45]. Advanced maternal age at conception is considered a major risk factor for trisomy 21, as is true for all human autosomal trisomies [46]. Females in advanced age (above 35 years) are at a much higher risk of infertility, losing the baby, or having a pregnancy affected by congenital birth defects such as DS. However, the molecular background of nondisjunction is still being investigated, and the outcome of the genomic research shows that several exogenous and endogenous factors contribute to the age-related increase in oocyte aneuploidy. There is some research that associates specific gene variations with Down syndrome. It is suspected that the shortage of folate has been shown to disrupt the way DNA and chromosomes separate and recombine. In the case of such a gene variation in one of the parents that interfere with their body’s capability to metabolize folate, that may enhance their chance of having a child with DS [47]. Researchers found folate regulator gene mutations that might be related to maternal meiosis II error. Smoking is considered a risk factor also, and in an Asian population with the habit of using smokeless chewing tobacco, the risk of delivering babies with DS was higher [48]. Analysis of the mother’s reproductive tract flora and the intestinal flora of DS cases indicated significant differences regarding the intestinal microbiome of DS patients compared to a healthy control group, while the reproductive tract flora did not show any significant difference [49]. The composition of the microbial taxa in children with DS was 16.67% Bacteroides, 37.35% Firmicutes, 26.13% Proteobacteria, 50% Verrucomicrobia, and 15.64% Actinobacteria. The normal group was mainly composed of 45.30% Bacteroides, 22.85% Firmicutes, 15.64% Proteobacteria, 28% Verrucomicrobia, and 3.02% Actinobacteria. Analysis of the fecal samples of women who delivered babies suffering from DS indicated that the microorganisms of the families Clostridiaceae and Pasteurellaceae were more numerous in the group of women having delivered DS neonates than the group of women having delivered healthy newborns. The analysis of the collateral effect of antibiotics on the gut microbiome indicated that tetracycline/doxycycline reduced the abundance of several commercial components of the microbiome isolates, most effectively E. coli, even at low concentrations [50]. Interestingly enough that the use of narrow-spectrum penicillin and tetracycline might be associated with the prevalence/incidence of certain neurodegenerative diseases also, like PD and MS [51,52,53]. Maternal microbial dysbiosis has been implicated in adverse postnatal health conditions in offspring, such as obesity, cancer, and neurological disorders, but in the case of DS, most of the publications are focused on maternal risk factors, and the only certain risk factors for the birth of a child with DS are advanced maternal age at conception and recombination errors, even though the molecular mechanisms leading to chromosome 21 nondisjunction are still uncertain. Trisomy of chromosome 21 (TS21) is the most common autosomal aneuploidy with a prevalence of 1 in 700 newborns. Its phenotype is highly complex with permanent features, such as mental retardation, congenital dysmorphic traits, and hypotonia, and variable features including heart defects, susceptibility to AD, type 2 diabetes, obesity, and immune disorders. Overexpression of genes on chromosome-21 (Hsa21) is responsible for the pathogenesis of DS phenotypic features either in a direct or indirect manner since many Hsa21 genes can affect the expression of other genes mapping to different chromosomes. Many of these genes are involved in mitochondrial function and energy conversion and play a central role in mitochondrial dysfunction and chronic oxidative stress, consistently observed in DS subjects [54]
Tetracycline is a broad-spectrum polyketide antibiotic produced by the Streptomyces genus of Actinobacteria and it specifically inhibits the 30S ribosomal subunit, blocking the binding of the aminoacyl-tRNA to the acceptor site on the mRNA-ribosome complex on the bacterial cell. When this process stops, a cell can no longer maintain proper functioning and will be unable to grow or further replicate. The tetracycline can penetrate the bacterial cell wall by passive diffusion or by an energy-dependent active transport system, which is probably mediated in a pH-dependent manner. Protein synthesis is ultimately inhibited, leading to a bacteriostatic effect. Recent publications indicate that tetracycline binds to various synthetic double-stranded RNAs (dsRNAs) of random base sequences, suggesting that the double-stranded structures may play a more important role in the binding of the tetracycline to RNA other than specific base pairs, as earlier thought. This is imperative to consider possible alternative binding modes or sites that could help explain the mechanisms of action of the tetracycline against various pathogens and disease conditions [55].
The narrow-spectrum, beta-lactamase resistant penicillin (J01CF) inhibits the proliferation of Gram-positive cocci, Bifidobacteria, Lactobcilli, Eubacteria, and Lachnospiraceae in the microbiome, and promotes the proliferation of Enterobacteria and Bacteroidaceae [36].
The association of the antibiotic-modified microbiome and the development of consecutive, non-contagious diseases, prompted researchers to suspect that the use of antibiotics, might influence the microbiome to reverse the clinical manifestations of certain diseases through the possible elimination of intestinal pathogens triggering the disease [56,57,58].
Clinical approach to modify gut microbiome generally focus on reducing the overabundant microbial taxa or overall microbial load using antibiotics or antifungal agents. Modulation through diet, or supplementation with live microbes (single or mixed species) might be a possibility, but the introduction of the fecal microbial transplantation (FMT) has been found as a better solution. Antimicrobial agents are not generally considered appropriate for the long-term management of chronic conditions, given the need for repeat dosing and concerns about the emergence of antimicrobial resistance. Early data indicate that the use of antimicrobial drugs, as a method to eliminate pathogenic microbiomes before FMT or microbial supplementation, improves the engraftment of beneficial species and enhances treatment efficacy [59].
The new antibiotic-treatment method for AD and PD has emerged as a possible novel approach to ameliorate the symptoms in those cases and other ailments. The research in animal experiments AD/PD models provided sufficient evidence of the anti-amyloidogenic, anti-inflammatory, antioxidant, and antiapoptotic activity of tetracyclines, associated with cognitive improvement. The discovery of the neuroprotective effects of minocycline and doxycycline in animals initiated an investigation of their clinical efficacy in AD and PD patients, leading to inconclusive results because long stating therapy with tetracycline and minocycline might raise safety issues. The safety issues should be considered as the long-lasting use of antibiotics might result in the emergence of resistant pathogens, and the modification of the microbiome with unforeseeable consequences. The sub-antimicrobial doxycycline doses should be carefully applied for their effectiveness and long-term safety, especially in AD/PD populations. Minocycline may have some neuroprotective activity in various experimental models such as cerebral ischemia, traumatic brain injury, amyotrophic lateral sclerosis, PD, Huntington’s disease, and MS [60,61].
Autism spectrum disorder (ASD) is a complex range of neurodevelopmental symptoms, including the impairment of social interactions and communication, together with restrictive and repetitive patterns of behaviors. According to the latest studies, autistic children have shown significant gut microbiota composition changes and the GI symptoms may represent the inflammatory processes. It has been reported that microbial interventions, such as probiotics, and antibiotics (vancomycin) can contribute to the reduction of social behavioral symptoms and the level of inflammation in individuals with ASD [62,63].
Alteration of the gut-brain axis has been reported in people with MS, suggesting a possible role in disease pathogenesis and making it a potential therapeutic target. Antibiotic treatment ameliorated experimental autoimmune encephalitis (EAE) in mice through effects mediated via the gut microbiota. Disease amelioration was not observed when antibiotics were given intraperitoneally, bypassing the gut, suggesting that modulation of the gut microbiota produces protective effects.
A recent report found that oral treatment with ampicillin decreased EAE severity, but in different other EAE models, the results have been controversial [64].
Aging is associated with dysbiosis, defined as a loss of number and diversity in gut microbiota, which has been linked with various aspects of cognitive functions. Therefore, the gut microbiome has the potential to be an important therapeutic target for symptoms of cognitive impairment. It was reported that 12 weeks of probiotic treatment with a supplement containing Lactobacillus acidophilus, Lactobacillus casei, Bifidobacterium bifidum, and Lactobacillus fermentum positively affected the cognitive function and metabolic homeostasis in AD patients [65].
7. Conclusion
Our comparative analysis using three different statistical methods, clearly indicated a strong statistical association between the average yearly consumption (1997-2020) of tetracycline (J01A) and narrow spectrum, beta-lactamase resistant penicillin (J01CF) expressed in DID and the prevalence of people living with DS in 30 European countries estimated for 2019. It is suspected that antibiotic (J01A, J01CF) modified gut microbiome could play some role in the development of DS-related dysbiosis in the mothers delivering babies with DS, and this altered flora also affects the intestinal microbiome of the newborn babies with DS. The theory could be raised that certain molecules, produced by the altered maternal microbiome, might act as a factor in enhancing 21-chromosome trisomy, which is a possible new insight in the pathomechanisms of DS. It could be theorized that probiotics/prebiotics, taken for an appropriately long time before the planned pregnancy, might stabilize the maternal microbiome preventing DS-related dysbiosis.
Limitations of our study: Our results could not be applied at the individual level, and we could not provide data on the molecular mechanism of how antibiotics act in the development of trisomy.
Strength of the study: Using international, large-scale data analysis and three different statistical methods this study provides convincing evidence on the significant involvement of certain classes of antibiotics inducing dysbiosis which might lead to the development of DS.
Author Contributions
GT: Writing - original draft, conceptualization, GM: Formal analysis, KS: Corrections, ZsB: selecting references, GB: Methodology, LM: collecting references, ASz: developing diagrams, tables: BM: conceptualization, writing, supervision
Funding
This research was supported by the National Scientific Research Program [NKFI] K 138669,
Institutional Review Board Statement
Not applicable for studies not involving humans or animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable
Conflicts of Interest
The authors declare no conflict of interest.
References
- Antonarakis, S.E.; Skotko, B.G.; Rafii, M.S.; Strydom, A.; Pape, S.E.; Bianchi, D.W.; Sherman, S.L.; Reeves, R.H. Down syndrome. Nature Reviews Disease Primers 2020, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- de Graaf G, Buckley F & Skotko BG Birth and population prevalence of Down syndrome in European countries. 2018.
- Dierssen, M. Down syndrome: the brain in trisomic mode. Nat. Rev. Neurosci. 2012, 13, 844–858. [Google Scholar] [CrossRef] [PubMed]
- https://gateway.euro.who. 7120.
- Dominguez-Bello, M.G.; Godoy-Vitorino, F.; Knight, R.; Blaser, M.J. Role of the microbiome in human development. Gut 2019, 68, 1108–1114. [Google Scholar] [CrossRef] [PubMed]
- Heintz-Buschart, A.; Wilmes, P. Human Gut Microbiome: Function Matters. Trends Microbiol. 2018, 26, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.; Evenson, A.; Luckey, T.; McCoy, E.; Elvehjem, C.; Hart, E. USE OF SULFASUXIDINE, STREPTOTHRICIN, AND STREPTOMYCIN IN NUTRITIONAL STUDIES WITH THE CHICK. J. Biol. Chem. 1946, 165, 437–441. [Google Scholar] [CrossRef] [PubMed]
- E Carpenter, L. Effect of aureomycin on the growth of weaned pigs. . 1950, 27, 469–71. [Google Scholar]
- Coates, M.E. , Fuller, R., Harrison, G.F., Lev, M. and Suffolk, S.F. A comparision of the growth of chicks in the Gustafsson germ-free apparatus and in a conventional environment, with and without dietary supplements of penicillin. British journal of nutrition 1963, 17, 141–150. [Google Scholar] [CrossRef]
- Wegener, H.C. Antibiotics in animal feed and their role in resistance development. Curr. Opin. Microbiol. 2003, 6, 439–445. [Google Scholar] [CrossRef]
- Browne, H.P.; Shao, Y.; Lawley, T.D. Mother–infant transmission of human microbiota. Curr. Opin. Microbiol. 2022, 69, 102173. [Google Scholar] [CrossRef]
- Wright, M.L.; Starkweather, A.R. Antenatal microbiome: potential contributor to fetal programming and establishment of the microbiome in offspring. Nursing Research 2015, 64, 306–319. [Google Scholar] [CrossRef]
- Grech, A.; E Collins, C.; Holmes, A.; Lal, R.; Duncanson, K.; Taylor, R.; Gordon, A. Maternal exposures and the infant gut microbiome: a systematic review with meta-analysis. Gut Microbes 2021, 13, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol Mol Biol Rev. 2017, 8;81[4]:e00036-17.
- I McBurney, M.; Davis, C.; Fraser, C.M.; O Schneeman, B.; Huttenhower, C.; Verbeke, K.; Walter, J.; E Latulippe, M. Establishing What Constitutes a Healthy Human Gut Microbiome: State of the Science, Regulatory Considerations, and Future Directions. J. Nutr. 2019, 149, 1882–1895. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Fuchs, S.; Milo, R. Revised estimates for the number of human and bacteria cells in the body. PLoS biology 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed]
- Poretsky, R.; Rodriguez-R, L.M.; Luo, C.; Tsementzi, D.; Konstantinidis, K.T. Strengths and Limitations of 16S rRNA Gene Amplicon Sequencing in Revealing Temporal Microbial Community Dynamics. PLOS ONE 2014, 9, e93827. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi-Man, O.; Davenport, E.R.; Gilad, Y. Taxonomic Classification of Bacterial 16S rRNA Genes Using Short Sequencing Reads: Evaluation of Effective Study Designs. PLOS ONE 2013, 8, e53608. [Google Scholar] [CrossRef]
- Human Microbiome Jumpstart Reference Strains Consortium; Nelson, K. E.; Weinstock, G.M.; Highlander, S.K.; Worley, K.C.; Creasy, H.H.; Wortman, J.R.; Rusch, D.B.; Mitreva, M.; Sodergren, E.; Chinwalla, A.T. A catalog of reference genomes from the human microbiome. Science 2010, 328, 994–999. [Google Scholar]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; Bertalan, M. Enterotypes of the human gut microbiome. nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Shetty, S.A.; Hugenholtz, F.; Lahti, L.; Smidt, H.; de Vos, W.M. Intestinal microbiome landscaping: insight in community assemblage and implications for microbial modulation strategies. FEMS Microbiol. Rev. 2017, 41, 182–199. [Google Scholar] [CrossRef]
- Foster, K.R.; Schluter, J.; Coyte, K.Z.; Rakoff-Nahoum, S. The evolution of the host microbiome as an ecosystem on a leash. Nature 2017, 548, 43–51. [Google Scholar] [CrossRef]
- Tiffany, C.R.; Bäumler, A.J. Dysbiosis: from fiction to function. American Journal of Physiology-Gastrointestinal and Liver Physiology 2019, 317, G602–G608. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.A.; Hennet, T. Mechanisms and consequences of intestinal dysbiosis. Cell. Mol. Life Sci. 2017, 74, 2959–2977. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Nance, K.; Chen, S. The gut–brain axis. Annual Review of Medicine 2022, 73, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.; Engevik, M.A.; Spinler, J.K.; Versalovic, J. Healthy Human Gastrointestinal Microbiome: Composition and Function After a Decade of Exploration. Dig. Dis. Sci. 2020, 65, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Pequegnat, B.; Sagermann, M.; Valliani, M.; Toh, M.; Chow, H.; Allen-Vercoe, E.; Monteiro, M.A. A vaccine and diagnostic target for Clostridium bolteae, an autism-associated bacterium. Vaccine 2013, 31, 2787–2790. [Google Scholar] [CrossRef] [PubMed]
- Finegold, S.M. Desulfovibrio species are potentially important in regressive autism. Med Hypotheses 2011, 77, 270–274. [Google Scholar] [CrossRef]
- Heberling, C.A.; Dhurjati, P.S.; Sasser, M. Hypothesis for a systems connectivity model of autism spectrum disorder pathogenesis: Links to gut bacteria, oxidative stress, and intestinal permeability. Med Hypotheses 2013, 80, 264–270. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human Genetics Shape the Gut Microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: a review. Antonie van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef]
- Lubomski, M.; Tan, A.H.; Lim, S.-Y.; Holmes, A.J.; Davis, R.L.; Sue, C.M. Parkinson’s disease and the gastrointestinal microbiome. J. Neurol. 2019, 267, 2507–2523. [Google Scholar] [CrossRef]
- Cryan, J.F.; O'Riordan, K.J.; Sandhu, K.; Peterson, V.; Dinan, T.G. The gut microbiome in neurological disorders. Lancet Neurol. 2020, 19, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Gasparrini, A.J.; Wang, B.; Sun, X.; Kennedy, E.A.; Hernandez-Leyva, A.; Ndao, I.M.; Tarr, P.I.; Warner, B.B.; Dantas, G. Persistent metagenomic signatures of early-life hospitalization and antibiotic treatment in the infant gut microbiota and resistome. Nat. Microbiol. 2019, 4, 2285–2297. [Google Scholar] [CrossRef] [PubMed]
- Patangia, D.V.; Ryan, C.A.; Dempsey, E.; Ross, R.P.; Stanton, C. Impact of antibiotics on the human microbiome and consequences for host health. Microbiologyopen 2022, 11, e1260. [Google Scholar] [CrossRef] [PubMed]
- Vandenplas, Y.; Carnielli, V.P.; Ksiazyk, J.; Luna, M.S.; Migacheva, N.; Mosselmans, J.M.; Picaud, J.C.; Possner, M.; Singhal, A.; Wabitsch, M. Factors affecting early-life intestinal microbiota development. Nutrition 2020, 78, 110812. [Google Scholar] [CrossRef] [PubMed]
- Requena, T.; Velasco, M. The human microbiome in sickness and in health. Revista Clínica Española (English Edition) 2021, 221, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Ternák, G. 1: A new discovery: The prevalence/incidence of different non-contagious diseases are associated with the antibiotic consumption patterns Paperback –2022 Lambert Academic Press, ISBN-13978-6204984452 pages, 2022.
- Biagi, E.; Candela, M.; Centanni, M.; Consolandi, C.; Rampelli, S.; Turroni, S.; Severgnini, M.; Peano, C.; Ghezzo, A.; Scurti, M.; et al. Gut Microbiome in Down Syndrome. PLOS ONE 2014, 9, e112023–e112023. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Wang, X.; Qin, J.; Mu, Q.; Ye, S.; Zhang, Y.; Yu, W.; Guo, J. Altered gut microbiota correlates with cognitive impairment in Chinese children with Down’s syndrome. Eur. Child Adolesc. Psychiatry 2021, 31, 189–202. [Google Scholar] [CrossRef]
- Hursitoglu, M.; Kural, A.; Kuras, S.; Akdeniz, E.; Sezer, S.; Caypinar, S.S.; Kazezoglu, C.; Yaprak, B.; Karandere, F.; Guven, H.Z. Maternal Gut Microbiota in Pregnancies Resulting in Down Syndrome Newborns – a Pilot Study. Acta Clin. Croat. 2021, 60., 722–730. [Google Scholar] [CrossRef]
- Quality Indicators for Antibiotic Consumption in the Community. Available online: https://www.ecdc.europa.eu/en/antimicrobial-consumption/database/quality-indicators (accessed on 10 March 2021).
- https://ourworldindata.
- Coppedè, F. Risk factors for Down syndrome. Archives of toxicology 2016, 90, 2917–2929. [Google Scholar] [CrossRef]
- Antonarakis, S.E.; Skotko, B.G.; Rafii, M.S.; Strydom, A.; Pape, S.E.; Bianchi, D.W.; Sherman, S.L.; Reeves, R.H. Down syndrome. Nature Reviews Disease Primers 2020, 6, 9. [Google Scholar] [CrossRef]
- Mikwar, M.; MacFarlane, A.J.; Marchetti, F. Mechanisms of oocyte aneuploidy associated with advanced maternal age. Mutat. Res. Mutat. Res. 2020, 785, 108320. [Google Scholar] [CrossRef] [PubMed]
- Halder, P.; Pal, U.; Ganguly, A.; Ghosh, P.; Ray, A.; Sarkar, S.; Ghosh, S. Understanding etiology of chromosome 21 nondisjunction from gene × environment models. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yannan Liu, Dongjie Hu, Chunfang Dai et al. Characteristics of Intestinal Microbiota and Mother's Reproductive Tract Flora in Children With Down Syndrome, 2021, PREPRINT [Version 1] available at Research Square, 2021. [CrossRef]
- Maier, L.; Goemans, C.V.; Wirbel, J.; Kuhn, M.; Eberl, C.; Pruteanu, M.; Müller, P.; Garcia-Santamarina, S.; Cacace, E.; Zhang, B.; et al. Unravelling the collateral damage of antibiotics on gut bacteria. Nature 2021, 599, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Ternák, G.; Németh, M.; Rozanovic, M.; Márovics, G.; Bogár, L. Antibiotic Consumption Patterns in European Countries Are Associated with the Prevalence of Parkinson’s Disease; the Possible Augmenting Role of the Narrow-Spectrum Penicillin. Antibiotics 2022, 11, 1145. [Google Scholar] [CrossRef] [PubMed]
- Nørgaard, M.; Nielsen, R.B.; Jacobsen, J.B.; Gradus, J.L.; Stenager, E.; Koch-Henriksen, N.; Lash, T.L.; Sørensen, H.T. Use of Penicillin and Other Antibiotics and Risk of Multiple Sclerosis: A Population-based Case-Control Study. Am. J. Epidemiology 2011, 174, 945–948. [Google Scholar] [CrossRef]
- Ternák, G.; Berényi, K.; Márovics, G.; Sümegi, A.; Fodor, B.; Németh, B.; Kiss, I. Dominant Antibiotic Consumption Patterns Might Be Associated With the Prevalence of Multiple Sclerosis in European Countries. Vivo 2020, 34, 3467–3472. [Google Scholar] [CrossRef]
- Izzo, A.; Mollo, N.; Nitti, M.; Paladino, S.; Calì, G.; Genesio, R.; Bonfiglio, F.; Cicatiello, R.; Barbato, M.; Sarnataro, V.; et al. Mitochondrial dysfunction in down syndrome: molecular mechanisms and therapeutic targets. Mol. Med. 2018, 24, 1–8. [Google Scholar] [CrossRef]
- Chukwudi, C.U. rRNA Binding Sites and the Molecular Mechanism of Action of the Tetracyclines. Antimicrob. Agents Chemother. 2016, 60, 4433–4441. [Google Scholar] [CrossRef]
- Schwartz, D.J.; Langdon, A.E.; Dantas, G. Understanding the impact of antibiotic perturbation on the human microbiome. Genome medicine 2020, 12, 1–12. [Google Scholar] [CrossRef]
- Manor, O.; Dai, C.L.; Kornilov, S.A.; Smith, B.; Price, N.D.; Lovejoy, J.C.; Gibbons, S.M.; Magis, A.T. Health and disease markers correlate with gut microbiome composition across thousands of people. Nat. Commun. 2020, 11, 5206. [Google Scholar] [CrossRef]
- Durack, J.; Lynch, S.V. The gut microbiome: Relationships with disease and opportunities for therapy. J. Exp. Med. 2019, 216, 20–40. [Google Scholar] [CrossRef] [PubMed]
- Keshteli, A.H.; Millan, B.; Madsen, K.L. Pretreatment with antibiotics may enhance the efficacy of fecal microbiota transplantation in ulcerative colitis: a meta-analysis. Mucosal Immunol. 2017, 10, 565–566. [Google Scholar] [CrossRef] [PubMed]
- Markulin, I.; Matasin, M.; Turk, V.E.; Salković-Petrisic, M. Challenges of repurposing tetracyclines for the treatment of Alzheimer’s and Parkinson’s disease. Journal of Neural Transmission 2022, 129, 773–804. [Google Scholar] [CrossRef] [PubMed]
- Cankaya, S.; Cankaya, B.; Kilic, U.; Kilic, E.; Yulug, B. The therapeutic role of minocycline in Parkinson’s disease. Drugs Context 2019, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sorboni, S.G.; Moghaddam, H.S.; Jafarzadeh-Esfehani, R.; Soleimanpour, S. A Comprehensive Review on the Role of the Gut Microbiome in Human Neurological Disorders. Clin. Microbiol. Rev. 2022, 35, e0033820. [Google Scholar] [CrossRef]
- Fattorusso, A.; Di Genova, L.; Dell’Isola, G.B.; Mencaroni, E.; Esposito, S. Autism Spectrum Disorders and the Gut Microbiota. Nutrients 2019, 11, 521. [Google Scholar] [CrossRef]
- Ghezzi, L.; Cantoni, C.; Pinget, G.V.; Zhou, Y.; Piccio, L. Targeting the gut to treat multiple sclerosis. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Sun, Y.; Baptista, L.C.; Roberts, L.M.; Jumbo-Lucioni, P.; McMahon, L.L.; Buford, T.W.; Carter, C.S. The Gut Microbiome as a Therapeutic Target for Cognitive Impairment. Journals Gerontol. Ser. A 2020, 75, 1242–1250. [Google Scholar] [CrossRef]
Figure 1.
Scatter diagram showing the positive association between tetracycline consumption [J01A] and the prevalence of people living with DS, estimated for 2019 [in 30 European countries].
Figure 1.
Scatter diagram showing the positive association between tetracycline consumption [J01A] and the prevalence of people living with DS, estimated for 2019 [in 30 European countries].
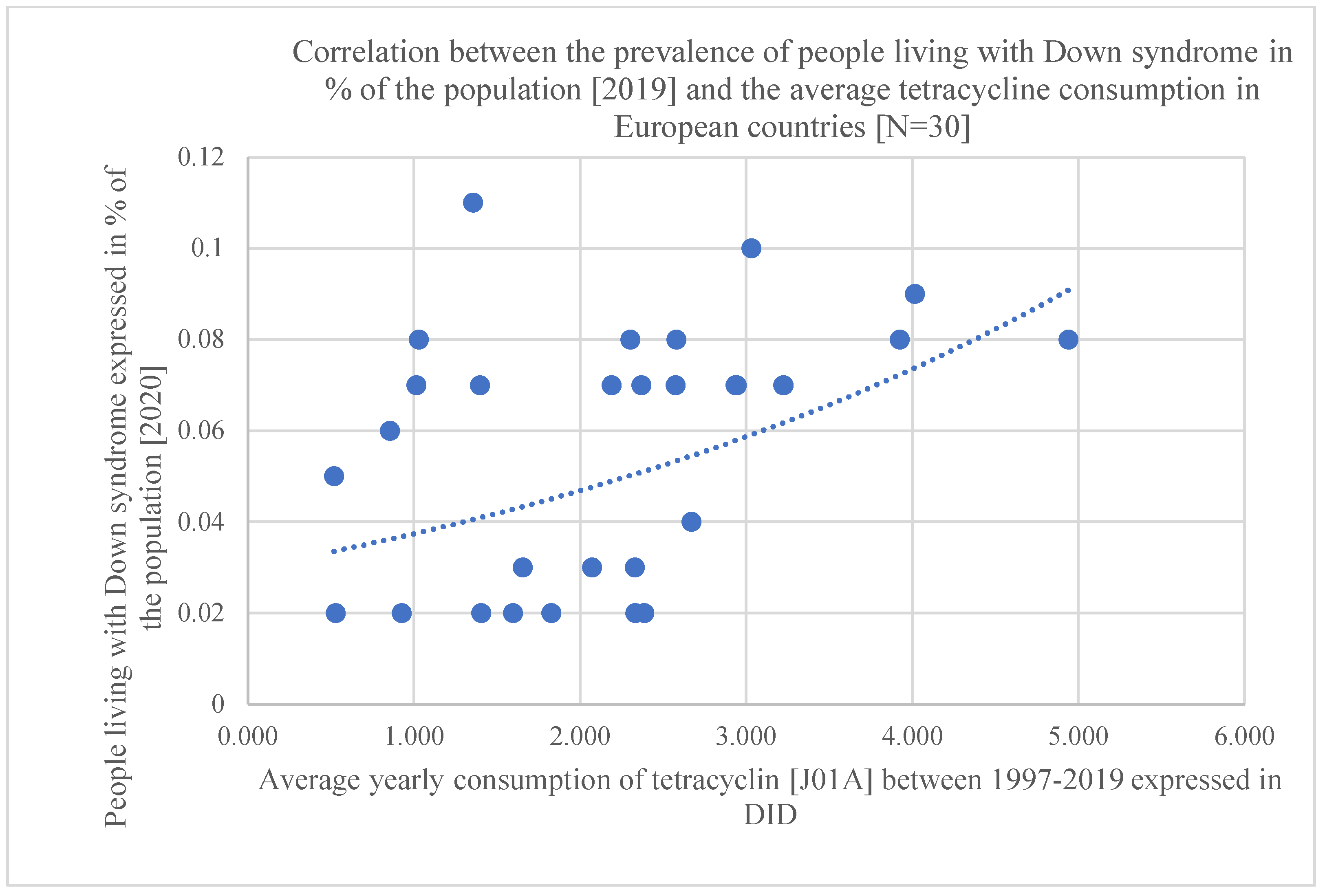
Figure 2.
Scatter diagram showing the association between the consumption of narrow-spectrum, beta-lactamase resistant penicillin [J01CF] and the prevalence of people living with DS estimated for 2019 [In 30 European countries].
Figure 2.
Scatter diagram showing the association between the consumption of narrow-spectrum, beta-lactamase resistant penicillin [J01CF] and the prevalence of people living with DS estimated for 2019 [In 30 European countries].
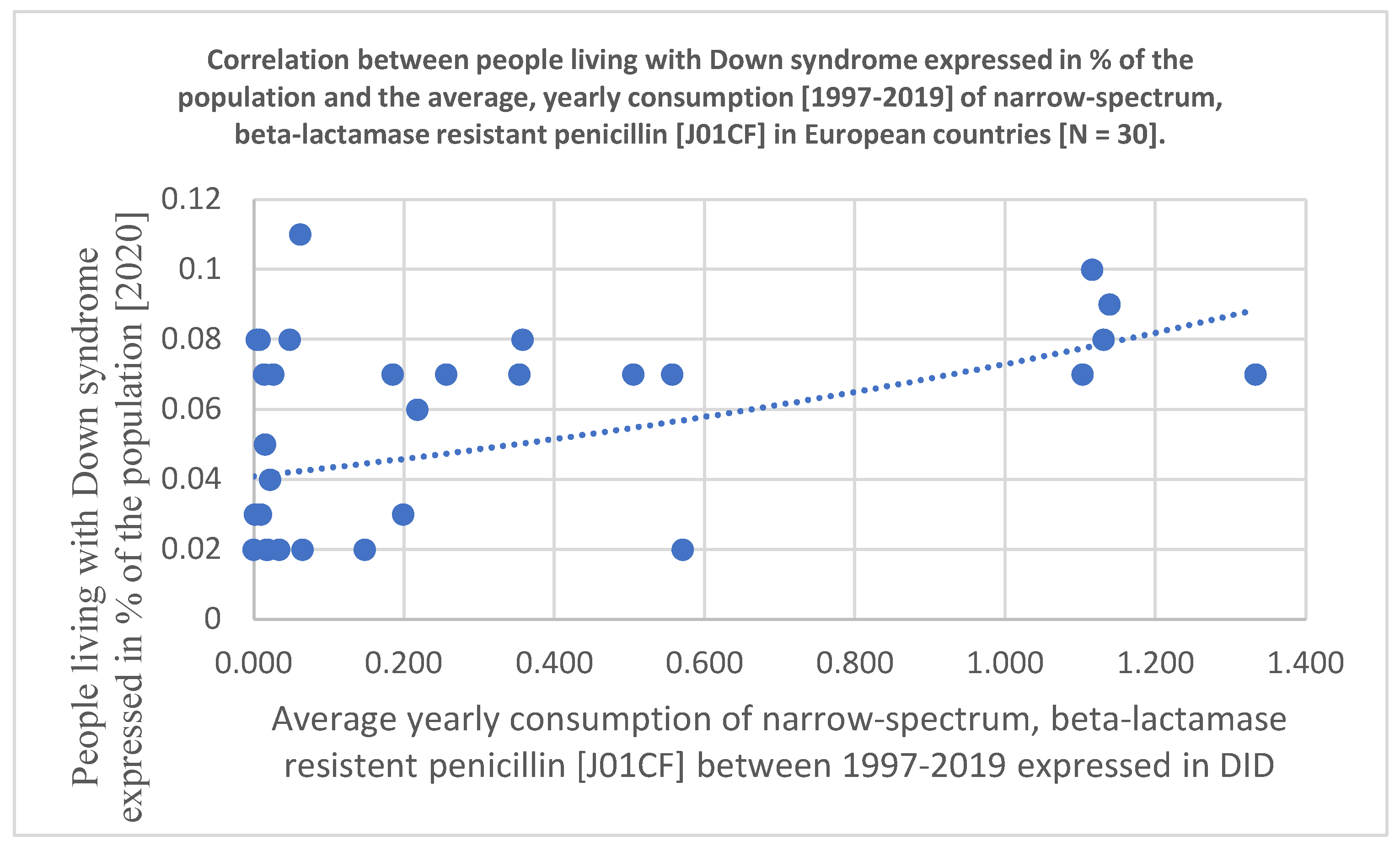

Table 1.
Average, yearly consumption of different classes of antibiotics between 1997-2020 expressed in DID compared to the prevalence of DS in the population expressed in %. Significant positive correlations are marked with yellow color, and a non-significant positive association is marked with blue.
Table 1.
Average, yearly consumption of different classes of antibiotics between 1997-2020 expressed in DID compared to the prevalence of DS in the population expressed in %. Significant positive correlations are marked with yellow color, and a non-significant positive association is marked with blue.
| Antibiotic consumption 1997-2020 DID | J01 | J01A | J01C | J01CA | J01CE | J01CF | J01CR | J01D | J01F | J01M | People living with DS in % of the population. 2019 | % |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Austria | 11.683 | 1.03 | 4.251 | 0.819 | 0.946 | 0.008 | 2.474 | 1.565 | 3.076 | 1.311 | Austria | 0.08 |
| Belgium | 21.427 | 2.368 | 8.725 | 3.796 | 0.091 | 0.256 | 4.578 | 2.387 | 3.205 | 2.206 | Belgium | 0.07 |
| Bulgaria | 17.828 | 2.386 | 6.55 | 4.039 | 0.953 | 0.019 | 1.56 | 2.672 | 2.589 | 1.991 | Bulgaria | 0.02 |
| Croatia | 17.911 | 1.404 | 7.581 | 2.467 | 1.007 | 0.034 | 4.074 | 3.172 | 2.641 | 1.496 | Croatia | 0.02 |
| Cyprus | 27.338 | 3.226 | 9.454 | 3.118 | 0.101 | 0.026 | 6.234 | 5.923 | 3.126 | 4.623 | Cyprus | 0.07 |
| Czech Rep. | 14.685 | 2.333 | 5.539 | 1.355 | 1.877 | 0.065 | 2.222 | 1.55 | 3.028 | 1.03 | Czech Rep. | 0.02 |
| Denmark | 14.068 | 1.398 | 8.782 | 2.732 | 4.575 | 1.103 | 0.378 | 0.03 | 2.031 | 0.455 | Denmark | 0.07 |
| Estonia | 10.947 | 2.072 | 3.701 | 2.422 | 0.264 | 0.01 | 1.007 | 0.892 | 1.947 | 0.83 | Estonia | 0.03 |
| Finland | 16.157 | 3.925 | 4.559 | 2.349 | 1.514 | 0.048 | 0.65 | 2.094 | 1.474 | 0.799 | Finland | 0.08 |
| France | 24.478 | 3.224 | 11.423 | 6.804 | 0.18 | 0.354 | 4.067 | 2.818 | 4.087 | 1.923 | France | 0.07 |
| Germany | 12.61 | 2.574 | 3.419 | 2.033 | 1.064 | 0.014 | 0.302 | 1.908 | 2.293 | 1.205 | Germany | 0.07 |
| Greece | 30.474 | 2.581 | 8.579 | 4.088 | 0.441 | 0.004 | 4.018 | 7.183 | 8.026 | 2.686 | Greece | 0.08 |
| Hungary | 14.711 | 1.595 | 5.269 | 1.467 | 0.634 | 0 | 3.164 | 2.162 | 3.054 | 1.843 | Hungary | 0.02 |
| Iceland | 19.148 | 4.941 | 9.219 | 3.392 | 2.446 | 1.131 | 2.232 | 0.567 | 1.56 | 0.797 | Iceland | 0.08 |
| Ireland | 18.256 | 3.032 | 8.219 | 2.816 | 0.93 | 1.116 | 3.374 | 1.57 | 3.413 | 0.858 | Ireland | 0.1 |
| Italy | 21.524 | 0.518 | 9.092 | 3.509 | 0.013 | 0.015 | 5.56 | 2.835 | 4.726 | 3.073 | Italy | 0.05 |
| Latvia | 10.653 | 2.329 | 4.04 | 2.717 | 0.096 | 0.002 | 1.226 | 0.544 | 1.435 | 0.982 | Latvia | 0.03 |
| Lithuania | 16.714 | 1.656 | 8.744 | 5.103 | 2.225 | 0.199 | 1.227 | 1.301 | 1.747 | 0.983 | Lithuania | 0.03 |
| Luxembourg | 22.41 | 2.189 | 7.974 | 3.09 | 0.088 | 0.185 | 4.618 | 4.087 | 3.998 | 2.413 | Luxembourg | 0.07 |
| Malta | 18.465 | 1.356 | 6.265 | 0.537 | 0.092 | 0.062 | 5.572 | 4.034 | 3.771 | 2.1 | Malta | 0.11 |
| Netherlands | 9.218 | 2.304 | 2.954 | 1.266 | 0.357 | 0.358 | 0.972 | 0.089 | 1.383 | 0.836 | Netherlands | 0.08 |
| Norway | 15.046 | 2.944 | 6.114 | 1.919 | 3.681 | 0.505 | 0.009 | 0.144 | 1.567 | 0.468 | Norway | 0.07 |
| Poland | 18.773 | 2.671 | 6.186 | 3.653 | 0.421 | 0.022 | 2.092 | 2.315 | 3.346 | 1.276 | Poland | 0.04 |
| Portugal | 18.094 | 1.015 | 7.836 | 2.006 | 0.028 | 0.557 | 5.248 | 2.308 | 3.179 | 2.496 | Portugal | 0.07 |
| Romania | 22.897 | 0.926 | 10.725 | 4.063 | 0.695 | 0.571 | 5.478 | 4.281 | 2.866 | 2.954 | Romania | 0.02 |
| Slovakia | 21.098 | 1.827 | 8.279 | 2.232 | 2.82 | 0.017 | 3.218 | 3.604 | 4.632 | 1.842 | Slovakia | 0.02 |
| Slovenia | 12.873 | 0.529 | 7.15 | 2.148 | 1.982 | 0.148 | 2.881 | 0.56 | 2.462 | 1.265 | Slovenia | 0.02 |
| Spain | 17.697 | 0.854 | 9.166 | 3.611 | 0.101 | 0.218 | 5.134 | 2.089 | 2.538 | 2.359 | Spain | 0.06 |
| Sweden | 13.37 | 2.936 | 6.387 | 1.047 | 3.821 | 1.333 | 0.182 | 0.28 | 0.73 | 0.825 | Sweden | 0.07 |
| UK | 15.294 | 4.016 | 5.851 | 3.257 | 0.746 | 1.139 | 0.734 | 0.546 | 2.627 | 0.534 | UK | 0.09 |
| 2019 R [Pearson] | 0.174 | 0.479 | -0.040 | -0.139 | -0.043 | 0.328 | 0.005 | 0.072 | 0.109 | -0.009 | ||
| 2019 p [Pearson] | 0.358 | 0.007 | 0.835 | 0.464 | 0.821 | 0.077 | 0.981 | 0.706 | 0.567 | 0.963 | ||
| 2019 OR | 1.074 | 3.896 | 0.861 | 0.741 | 0.497 | 14.491 | 0.942 | 0.510 | 2.031 | 2.737 | ||
| 2019 CI95% | 0.932-1.246 | 1.632-11.626 | 0.560-1.304 | 0.418-1.262 | 0.193-1.117 | 1.764-186.274 | 0.572-1.508 | 0.139-1.578 | 0.784-5.969 | 0.537-16.944 | ||
| 2019 OR p | 0.324 | 0.005 | 0.478 | 0.274 | 0.109 | 0.021 | 0.806 | 0.263 | 0.160 | 0.239 | ||
| 2019 Kruskal-Wallis p | 0.431 | 0.034 | 0.348 | 0.49 | 0.237 | 0.152 | 0.842 | 0.897 | 0.517 | 0.362 |
Glossary: J01: total antibiotics for systemic use, J01A: tetracycline, J01C: penicillin, J01CA: broad-spectrum, beta-lactamase-sensitive penicillin, J01CE: narrow-spectrum, beta-lactamase sensitive penicillin, J01CF: narrow-spectrum, beta-lactamase resistant penicillin, J01CR: broad-spectrum, beta lactamase-resistant, combination penicillin, J01D: cephalosporin, J01F: macrolides, J01M: quinolone.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



