
Submitted:
10 May 2023
Posted:
11 May 2023
You are already at the latest version
Abstract
Knowledge about ecological conditions and processes in centers of endemism (CoEs) is still limited with respect to various systematic groups of organisms, ecosystem types, ecological conditions and ecosystem functions. Endemics and CoEs play a prominent role in nature conservation monitoring and management, and in the organization of zoos, aquaria and botanical gardens. We review the meaning of different groups of organisms and indicators for the characterization of endemic-rich regions, the importance of heterogeneity in space, continuity in time, isolation and ex-situ management for the survival of endemic and threatened species. Currently, conversion of land cover and land use change are the most important causes of biodiversity decline and extinction risk of endemic and endangered species concentrated in CoEs. These are followed by climate change, including severe weather, and then natural processes such as volcanism, landslides or tsunamis. For conservation purposes the management of zoos, aquaria, botanical gardens, of regional land use, and social aspects of the diversity of endemics and CoEs have to be taken into account as well. We need better answers to questions about the relationship between ecoregions, CoEs and regional awareness of biodiversity, which is linked with human culture including the management of zoos, aquaria and botanical gardens, aesthetics, wellbeing, health and trade.
Keywords:
heterogeneity in space
; spatial scale
; land cover
; continuity in time
; isolation
1. Introduction
The start of research on endemism and endemics in a macroecological context can be dated back to the transfer of terms from medicine to biogeography by De Candolle in 1820 [1,2,3]. Interest in the topic increased along with investigations of ecological and evolutionary questions on the one hand, and of declining biodiversity, an increasing number of threatened species and Red Lists worldwide on the other [4,5,6,7,8].
Meanwhile, neo-, paleo- (palaeo-), patro-, schizo-, apo-, hyper-, micro-, narrow, stenochorous, local, regional, national, supranational, and more or less widespread endemics, biodiversity hotspots, ecoregions and CoEs have been defined and identified with respect to biome, spatial scale, habitat type, systematic level and taxonomic group e.g. [9,10,11,12,13]. As a first approximation, most of these terms represent biogeographical labels, comparable e.g. with neophyte, indigenous or alien/invasive species [14]. The evolutionary history, ecological conditions or biological traits of range-restricted organisms is secondary, and it may be that taxa with a small range represent other ecological conditions or biological traits than widespread members of the same taxonomic group [15,16,17,18]. Rare species can be common where they occur [19], and niche specialization does not necessarily explain this result. However, there are few studies comparing rare and common congeners, [20] and the measures of attributes were so varied that it was not possible to compare such studies in order to assess if there is any generalized cause of rarity [21,22].
Rarity is scale-dependent [23], a finding that has implications for the IUCN Red Listing processes relating to area of occupancy and extent of occurrence.
Niche specialization and degree of distribution have been combined to provide an assessment measure of rarity, recognizing seven possible different dimensions of rarity [24,25]. This approach has not been widely used, but it does provide a unique insight into rarity, and has been used for plants [26,27,28] and a diverse range of animals e.g. [29,30].
Rarity of individual taxa on its own is no indication of levels of endemism, nor does it allow for the identification of centers of endemism. The link between rarity and richness of a region needs to be made, but only when richness is controlled for [31]. A narrow endemic aligns with the idea of rarity [32], but endemism does not necessarily imply rarity as it is scale-dependent. These authors also consider the primary source of information on species rarity and endemism to come from taxonomists, but ecologists have perhaps done more to explore aspects such as traits, niche and other aspects of rare and endemic taxa.
The whole inhabitable area of the Earth can be divided in ecoregions with high or low concentrations of endemics [33,34]. There are now countless checklists, floras and faunas providing information about endemic species and regions, as well as an ever increasing number of publications focussing on patterns and processes, evolutionary and genetic/phylogenetic analyses, and ecological and conservation goals cf. [35,36,37,38]. Regions with high numbers of endemic taxa often show high spatial heterogeneity in combination with long temporal continuity, e.g. relatively stable climate conditions [39,40,41,42,43]. This raises several questions: is it possible to disentangle the importance of spatial heterogeneity vs. continuity through time [44,45,46]? How are species compositions and endemism influenced by spatial separation and isolation through time? And what does this mean for our understanding of biogeographical patterns of endemism, ecosystem functioning and nature conservation practices in both island regions and mainland areas [47,48,49,50,51]?
Knowledge of patterns and processes of endemism and CoEs is still limited with respect to different taxonomic groups and their evolutionary history including migration and dispersal under conditions of climate, landscape and land use change. The meaning of specific conditions for survival at different spatial and temporal scales might differ from one place and taxonomic group to the other.
In a world of increasing pressure on biodiversity and declining diversity of native species in many regions, it is necessary to find serious political, technological, legal and other solutions for the survival and wellbeing of endemic and threatened taxa on the basis of scientific knowledge, even if not all questions can be answered yet [52]. With this review we want to mirror the state of the art and future prospects on the topic, and emphasize the importance of research on CoEs in the context of nature conservation planning.
Figure 1 shows a schematic depiction of the relationship between evolutionary diversification, number of endemics and extinction risk of endemic and endangered taxa. It indicates that increasing and decreasing numbers of endemics and endangered species reflect different bundles of processes. Thus, the effects of land use and climate change on the assemblage and migration of assemblage cannot simply be compensated by restoration.
2. What are centers of endemism
If a region is characterized by high endemism it is called a center of endemism (CoE) [58,59,60,61,62]. If the original natural vegetation is additionally damaged then it can be called biodiversity hotspot [63]. However, the characteristics differ according to region, composition of habitat types, group of organisms considered and/or scientific mode of calculation and indication [64,65,66,67,68]. The key issue here is that there is no single or even subset of preferred methods or criteria for defining CoEs. In addition, sampling bias and density can skew data and counts, resulting in misidentified CoEs or inaccurate decisions regarding boundaries of CoEs. For example, after four years of field work, a new center of plant endemism in southern Africa was identified [69], a region already abundant in CoEs. However, after additional field work in a region adjacent to the new CoE, it was then noted that this recognition may have been premature [70].
There is a historical legacy linking chorology with the identification of biodiversity and CoEs. Chorology, e.g. in plants, has been applied at family, genus and species levels e.g. [71]. The concept of chorology dates back to 1866 when it was used by Haeckel in relation to the dispersal of organisms away from a center of origin [72]. While chorology was initially a manual method of plotting distributions on maps and identifying iso-chores, more recent approaches have utilized computational power to analyze distribution data and identify CoEs e.g. [73,74].
A comparison of numbers of endemic taxa or other indicators of endemism is, in principle, possible by comparing units of the same or of different range size. When comparing units of the same size such as quadrats of 100 or 10,000 square kilometers, numbers of endemic taxa can be compared directly [75]. However, even grid cells of degree latitude/longitude have different range sizes in different latitudinal belts, and numbers of endemic taxa in this case are not directly comparable without mathematical correction. Meanwhile many indicators for CoEs such as number of endemics (E), Endemics-Area Relationships (EARs), Weighted Endemism (WE, CWE), Range Size Rarity (RSR), Parsimony Analysis of Endemicity (PAE), Proportion of endemics (S/E), Bykow´s Index of Endemicity (BI), and others have been calculated by using different spatial scales and models [76,77,78]. The indication value of each method is limited and the mode of calculation must be taken into consideration [79,80,81,82,83].
If, for example, the only native terrestrial mammal of the Hawaiian Islands is an endemic species (Lasiurus semotus) then the level of endemism is 100 %. If it is a non-endemic species (Lasiurus cinereus) then there would be no species endemism in terrestrial mammals on Hawaiian Islands (0 %). With respect to its biology, this bat species has also been classified as endemic subspecies (Lasiurus cinereus subsp. semotus). This example shows that level (ratio) of endemism cannot indicate the same as for example number of endemics [84].
However, many ecoregions are labelled as CoEs because they have an obvious concentration of endemics, i.e. much more than in neighboring regions. This assumes that sampling effort is uniform or equitable across the ecoregion. However, all too often sampling is biased, favoring easy access routes etc. There is also the “Diversity Tracking” effect [85] where collectors of specimen and distribution data tend to visit places of known diversity so as to maximize their returns.
Due to methodological difficulties, and because of scientific underrepresentation, many groups of invertebrates such as insects or mollusks seem to rarely occur as endemics compared to vertebrates or vascular plants, which comprise the taxonomic groups mainly characterizing biodiversity hotspots and CoEs [86,87]. Thus, a stronger focus on these underrepresented groups might be appropriate to discover further centers of endemism.
3. Endemism in different groups of organisms
In general, the transition zone between land and sea is the most effective border for the distribution of many phyla and most animal and plant species. Oceans are much older, longer inhabited by animals and larger than e.g. freshwater ecosystems, but the richness of marine fish species is comparable with the richness of freshwater fish species. Thus, after the niche occupation in rivers and lakes the diversification rate in freshwater fish species must have been higher than in marine fish species.
Whilst there are much more animal and plant species including endemic species in non-marine habitats (> 75 % of the whole species diversity?), marine environments contain a much larger number of phyla including endemic phyla. Explaining this disparity was and still is part of a long-lasting discussion about origin and evolution, migration history, the meaning of dispersal, isolation, streams and currents, and continuity/change of ecological conditions [88,89,90]. In terms of marine biogeography, substantial leaps have been made in seeking or testing hypotheses of common causes to explain coincidental boundaries to distributions. For example, numerous phylogeographic studies on animal and algal taxa along the southern African coastline have indicated no clear congruent patterns in disjunctions, and when undertaken, dating analyses have not converged on a single historical period when such a disjunction came into being (cf. [91], and subsequent papers by this author). Similar findings have been made along the Australian coastline [86].
Various indicators such as the ratio of species/genera or genera/families, level or other indicators of endemism, indices of similarity, e.g. Sørensen-Inex, Jaccard-Index, or molecular-genetic differences in phylogenetic diversity have been established to calculate the degree of uniqueness, isolation or insularity of the species pool or assemblage [92].
Figure 2 shows current estimates of global species numbers of selected taxa with minima and maxima according to species concept, calculation mode, and expectation of further discoveries [93,94,95,96,97].
Taxonomic groups of animals, plants or other groups differ in dispersal mode and speed of migration, and thus have very different average range sizes. The degree of endemism in ecoregions and countries often increases e.g. from lichens, bryophytes, and birds with relatively small percentage values to vascular plants and other vertebrates (intermediate level) to amphibians and non-marine gastropods, which regularly show much higher or highest rates of endemism (Figure 3 and Figure 4).
Despite a larger number of vascular plant species on Earth, endemism in amphibians or non-marine gastropods at regional scales, for example, is often higher [98,99,100,101]. Even if range is one of the most curious concepts in biogeography, amphibian or non-marine gastropod species are on average more restricted than e.g. birds. This has to do with the fact that they are slow and not able to fly.
Most members of the two richest plant families, Asteraceae and Orchidaceae, have the possibility of long-distance dispersal by wind (anemochory). They nevertheless contain many endemics at regional to continental scales. Endemic orchids are numerous in the tropics, whereas in many regions at higher latitudes and altitudes endemic Asteraceae are regularly on top of the counts in checklists. Most endemics in these and many other families are pollinated by insects, which favors occupation of, and evolution in, narrow niches e.g. [102,103,104].
4. How are endemics distributed and related to environmental heterogeneity in space?
Elevation of a region, number of predefined climate zones and many other indicators have been used to calculate heterogeneity in space. Thus, various indicators of heterogeneity in space are available for biogeographical analyses e.g. [105,106,107,108,109].
Different groups of organisms show a latitudinal gradient with low numbers of species (S) and endemics (E) towards the poles and high numbers to the tropics [110,111]. This means that tropical and subtropical zones are richer in species and endemics than temperate or boreal-arctic regions, even if exceptions in several taxonomic groups have been discovered e.g. [112,113,114]. Mountain ranges harbor higher numbers of endemic taxa than lowland areas of comparable size, longitude and latitude [115,116,117,118,119]. There is also a well-documented mid-slope peak in diversity up elevational gradients in mountains termed the mid-domain effect [120,121,122,123].
Large species pools, CoEs and phylogenetic diversity often represent the same ecoregions or overlapping areas [124,125]. Despite large congruence between E, E/S, S and other values characterizing CoEs, exceptions are numerous and e.g. maxima in various taxonomic groups characterize different regions [126] (cf. Table 1).
Several islands and archipelagos distant from the mainland support high numbers of endemics and highest levels of endemism. In contrary, tropical and subtropical mainland regions often harbor much more species in total and higher phylogenetic diversity than comparable islands or archipelagos. This also has to do with dispersal, biological connectivity, spatial separation and genetic isolation effects through time [140].
Springs, lakes, rivers, fens, bogs, mountain tops, rocky outcrops, or areas with serpentine soils in terrestrial regions are ecologically more or less isolated habitat-islands and separated comparable with true islands [141,142]. However, isolation is relative. Saltwater is a barrier for many taxonomic groups, but medium of dispersal e.g. for marine animals and coconut (Cocos nucifera). Thus, these habitat islands may be only islands under current climates. They may well have once been more widespread and connected.
A preliminary list of hyperendemics [143] showed that the focus on species with an extremely small range of a single square kilometer or a maximum of 50 individuals can elucidate the importance of habitat types and landscape units that are often located outside of already identified CoEs. For example, many small springs harbor freshwater and wetland gastropods with a rather restricted range.
5. How is endemism related to continuity, change in time and isolation?
Species compositions and species richness of landscapes and regions reflect continuity and change of ecological conditions during the history, the length of time periods and evolutionary speed. Specific modes of dispersal, migration routes and the increase or decrease of numbers of species depend on the distribution of water bodies and terrestrial land, i.e. on barriers and drivers of migration/dispersal. Distances to other water bodies or land masses are seen as important components to calculate the degree of isolation. The similarity of ecological conditions, assemblages and phylogenetic diversity are used to characterize the degree of uniqueness and isolation as well.
Ecological conditions and the amount of endemic plant species, e.g. of the higher mountain belts of the Macaronesian Islands, differ much more than e.g. coastal habitats of the archipelago, and they are more distant [144,145,146,147,148].
Under stable geological, climate, soil and hydrological conditions, the number of species increases via evolution and migration/dispersal. Isolation processes also favor the evolution of endemism. Isolation in a biogeographical context can be defined as a condition reducing or preventing genetic, social and other biological aspects such as intra- and interspecific relationships and dispersal. Because of the strong isolation of a first propagule arriving at a new oceanic island, the survival of this species is highly questionable. This propagule must find adequate ecological conditions, build a founder population, may overcome a genetic bottleneck, and so on. Isolation, on the other hand, reduces dispersal with the effect that mainland regions normally have higher numbers of non-endemics than ecologically comparable island regions. In general, both large species pools and isolation, which often show opposite patterns, favor the evolution of endemics [149,150,151,152]. In plants, there is a link between successful dispersal and polyploidy e.g. [153,154]. Ploidy data is scattered (especially for endemics) but it would be an interesting line of enquiry to assess levels of ploidy in endemics and widespread congeners and taxa in general.
According to the Theories on Adaptive Dynamics and Assembly Optimization, optimization of resource use is one of the most important drivers for processes controlling ecosystem functioning, species composition, and diversity [155,156,157,158,159,160]. If processes and conditions of dispersal are reduced because an archipelago such as the Hawaiian Islands is located far distant from mainland areas and other archipelagos, then evolution of new taxa can help to optimize nutrient cycling and resource use. As a result, the total number of taxa (S) on isolated islands often is relatively small compared to mainland areas of the same range size and similar ecological conditions, the number of endemics (E) is high and the level of endemism (E/S) extremely high [161,162,163].
Because there is no other planet enabling life in our vicinity, all taxa on Earth are at the same time global endemics (S = E). The isolation is maximal and the level of endemism 100 %. In general, high percentage values indicate strong isolation. The level of endemism of an archipelago is normally higher than of each island belonging to the archipelago. This indicates stronger isolation for the whole archipelago in comparison with the islands belonging to the archipelago [164].
Meanwhile, studies on the history of climate change at regional to continental scales, on refugia and microrefugia during glaciation periods, and climate continuity as a precondition for the evolution and survival of endemics are numerous. Dynessius and Jansen’s work and theories are applicable here [165,166,167]. The relation between number of endemics, proximity to refugia and climate continuity has often been pronounced and modelled. Moreover, the relative importance of time stability for evolution and survival is often hypothesized as higher than heterogeneity in space cf. [168,169,170,171].
6. How important are zoos and botanical gardens for endemics and vice versa?
Mankind’s relationship with animals kept for a specific purpose dates back millennia, and this relationship and purpose has changed over time [174]. Zoos, aquaria and botanic gardens have been considered as “Arks” where taxa are kept and distributed between such organizations with the aim of sustainable “insurance” populations, which can then be used for augmentation of existing small populations in the wild, or reintroduction into the wild where species are extinct in nature [175,176,177]. Thus, the role and function of zoos over the last four or five decades has resulted in a much more conservation focused approach [178]. However, the global distribution of zoos is disproportionately biased towards the global west first world countries [https://en.wikipedia.org/wiki/Zoo#/media/File:World_zoo.png; downl. 20/4/2023], generally distant from areas of faunal endemism and species in need of ex situ and in situ conservation.
Zoos are considered to be “ethically contested institutions” in terms of their existence as well as their aims, policies, and practices, which are underpinned by commitments to shared values of animal welfare and species conservation [179]. The authors argue that these values may be in tension, resulting in decisions that fulfil some aims at the expense of others, or result in unsatisfactory trade-offs. Among other conclusions, they suggest that zoos should hold a higher number of threatened taxa than currently in captivity, and that species that don’t require so much physical space, including amphibians, reptiles, fish and invertebrates, could be prioritised. From an animal welfare perspective these groups also present fewer challenges.
Various studies indicate that mammals are favoured by the public ([180]; and references therein) but there is a growing greater representation of birds and fish species diversity. The analysis of certain biological and geographical parameters shows that mammal and bird species in zoos are larger than their close relatives not held in zoos, and that these captive taxa represent species with larger spatial ranges which are less likely to be endemic, and are less likely to be threatened with extinction [181].
Approximately 95% or more animals are invertebrates, and if zoos were to represent this diversity proportionally, only 5% of their collections should comprise mammals, birds, reptiles, fish and amphibians [179]. However, other systematic groups such as insects or terrestrial snails only play a subordinate role, even if there are exceptions such as botanical gardens with butterflies as very attractive group of insects [182]. Representation effort and success also differs within the main groups [183]. For example, almost all species of Cactaceae are shown in botanical gardens, while this is not the case within the two largest plant families Orchidaceae and Asteraceae [184]. The situation in vascular plants is comparable with the vertebrate species. The representation is biased. Some groups are almost fully represented, but whales, for example, cannot be adequately housed in aquariums [185,186].
Rare, endemic and endangered species are now also being kept and propagated through conservation breeding programs in zoos and botanical gardens [187]. In this way, an increasingly important contribution to nature conservation can be made. At the same time, the attractiveness of the gardens increases. So it should be a win-win situation [188]. However, the options are limited. Endemic vertebrate species, vascular plant species, and other systematic groups are unlikely to be maintained in zoos and botanical gardens beyond individual conservation programs [189].
Interestingly, the role that zoos, aquaria and botanic gardens play in endemic species conservation, versus simply anthropomorphically appealing species, is a little difficult to assess, and review studies are few in comparison to individual taxon - specific studies. We present below a brief overview of the role and success of endemic species conservation according to major taxonomic grouping by zoos, aquaria and gardens.
Amphibians are the most threatened group of vertebrates, with over a third of extant species classified as “Threatened” [190]. A global analysis of zoo and aquarium collections showed that only 2.78% of the collections comprised amphibian taxa, and consider this both a challenge and opportunity for zoos and aquaria to increase their capacity for the conservation of rare and threatened taxa [179]. Many zoos utilise amphibians for displays and education, without a conservation goal or strategy [191]. Amphibians are ideal for zoo-based research due to their small size, high fecundity and ease of husbandry, and facilities for their breeding and maintenance are relatively cheap. In addition, amphibians are considered to be an excellent opportunity for conservation research partnerships. Assessments of amphibian conservation breeding programs showed, that species in breeding programs were likely to be under greater threat, but had similar range restricted distributions as those taxa not being bred for conservation purposes [192]. Furthermore, amphibians in zoos seem to be as threatened as their close relatives not in living collections [193]. In addition, those species in zoos are generally larger bodied, more widely distributed and more likely to be habitat generalists. These findings indicate that the ex situ conservation network of zoos and other institutions are not prioritizing endemics as range-restricted habitat specialists with greater extinction risk. As an example, Madagascar has 370 native amphibian species, and amphibian endemism is extremely high, but a survey [194] found that a mere 36 of these species are kept in zoos globally. Of these 36 species, 10 are considered as “Threatened”, and the remaining species are not benefitting from any ex situ conservation actions. On a positive note, almost a third of those species in captivity have successfully reproduced [194].
Reptiles are quite well-represented in zoos and aquariums [180]. However, we were not able to find general reviews of reptiles in zoological collections. As with amphibians and fish, there has however been a detailed analysis of the highly endemic Malagasy reptile fauna [194]. This fauna is rich, with 437 reptile species of which 420 are endemic to Madagascar. Of these, only 87 are kept in zoos around the world, and the majority of these are species not considered to be threatened. However, what is perhaps encouraging for endemic Malagasy reptiles is the finding that almost 40% of their geographic range, on average, is within a protected area of some kind [195]. Reptiles, as with many other taxa, are also targeted by legal and illegal traders, and the consequences of this include escapes and subsequent invasions. Species traded are not generally endemic taxa, and tend to be larger and more colourful or patterned species e.g. [196]. Reptiles in zoos are also larger bodied species, and endangered species, which may not be visually attractive to humans, do not feature extensively in zoo collections; a fact that cannot be explained by the difficulty of obtaining rare species [197].
The preference for larger and more colourful birds is also evident in zoo collections. Eight percent of the world’s threatened bird species are kept in captivity [185], and zoos tend to favour birds that are not endemic or threatened [181]. Additionally, there is an indication that the average diversity of bird species in zoos declined between 1960 and 2018 [198]. As with other taxa, Madagascar is a hotspot of bird diversity, and 142 out of the 195 bird species on Madagascar are endemic, 28 of which are threatened [187]. A survey of 131 institutions indicated that only 15 of these endemic species were kept in their living collections, meaning that 89% of Malagasy birds are not represented in ex situ collections. It thus appears that zoos do not have a strong track record of ex situ conservation of endemic birds, but given the global plethora of conservation organisations with a strong focus on birds perhaps this role is not considered a key strategy for zoos.
Freshwater fish species comprise approximately 25% of all vertebrate diversity, and freshwater fish comprise about 50% of all fish diversity [199]. The majority of freshwater fish species lack any formal IUCN Red List status. However, holdings of fish in zoos and aquaria increased between 1960 and 2018 [198]. Aquariums and zoos only hold about 7% of all threatened fishes, and highlight the important role that hobbyists play in the conservation and breeding of endemic and threatened taxa [200]. These hobbyists make up 99% of the global ornamental fish trade, and may play a vital role in fish conservation. This role has been facilitated by the formation of the CARES [Conservation, Awareness, Recognition, Encouragement, and Support] preservation program in 2004. CARES has published a priority list which contains nearly six hundred species [24 of which are extinct in the wild] from twenty families. Notably, species from Tanzania and Mexico, which are major centres of fish diversity and endemism, had the greatest representation in this list. However, the legal (and illegal) trade in endemic freshwater fishes for the hobbyist market can also be highly detrimental to conservation efforts [201]. Another freshwater fish hotspot for endemism is Madagascar, which has 173 fish species, 79 of which are endemic. These face extinction due to deforestation, overfishing, and the introduction of exotic species. However, only 21 of these species including 19 endemics are kept in zoos, with some success of ex situ breeding [202].
Being small and generally inconspicuous, invertebrates tend not to feature highly on the agenda of zoos and aquaria [203]. In 1991, one successful international invertebrate captive breeding program was reported [204], but by 1994, a growing public interest in, and value of, invertebrate collections in zoos was being recognised [205], and zoos embarked on invertebrate conservation e.g. [206,207]. Butterflies are considered to be flagship taxa, and there has been a rapid increase in “butterfly houses” at zoos and in tropical houses of botanical gardens, but there are a number of risks to butterfly biodiversity linked to this “industry” [208].
As with zoos, humans have cultivated plants in gardens since ancient civilisations, notably for the cultivation of medicinal plants, exotic fruits and spices [211]. The focus on “physic gardens” in the 1500’s in Europe set the scene for the subsequent growth of gardens as a consequence of the era of exploration and European colonisation. These gardens facilitated the rise of taxonomy, and the economic value of selected species such as coffee, rubber, tea, cotton, opium, sugarcane etc. is linked to societal evils such as slavery and warfare e.g. [209,210]. There are over 3000 botanic gardens worldwide, but as with the global distribution of zoos, two thirds of these are located far away from the world's 36 biodiversity hotspots Figure 2 in cf. [211]. Despite this, botanic gardens and arboreta grow at least one third of all known vascular plant species. Of these, over 16,000 are tree species, of which 1700 are globally threatened [212]. More recently, the role of botanic gardens in researching and conserving plants in the face of climate change studies and food security have become increasing priorities [213,214,215]. However, the space required in gardens to grow trees means that only a limited number of species and individuals can be accommodated. There is thus a specific role for arboreta in the conservation of tree species. In South Africa, for example, there are currently 121 arboreta. Historically there were at least 172, but many have been lost. The origins of some of these is linked to the rise of the forestry industry and the need for research on suitable tree species. These arboreta house 2309 species from around the world, of which 128 species are threatened [216]. In recent times, activities and strategies of botanic gardens worldwide is guided by the Botanic Gardens Conservation International [BGCI] Manual on Planning, Developing and Managing Botanic Gardens. The BGCI was founded in 1987. It was this organisation that drove the rise of plant conservation activities at botanic gardens [217,218,219,220]. However, gardens cannot readily grow sufficient number of individuals so as to ensure the maintenance of genetic diversity, and the potential for genetic erosion exists [221,222,223]. Even the role of seed banks in maintaining genetically diverse collections is limited. However, seed banks are able to store many more species than living collections in gardens. There are over 350 botanic gardens involved in seed banking in 74 countries [224]. According to their survey, 56,987 taxa have been banked as seed, 9,000 of which are threatened. In the context of the space requirements for trees, tree seed banking is an obvious solution, and seeds of 6881 tree species [half of which are endemics] from 166 countries are stored in seed banks [224]. However, these authors make a call for even more effort to collect and store seeds of threatened species. Meaningful long term partnerships in endemic species conservation between institutions in countries in the global north and the biodiverse rich and often developing world need to be strongly encouraged, and the “parachute science” approach discouraged [225,226].
7. Conclusion and outlook: What are the perspectives for endemics and centers of endemism?
The main goal of nature conservation programs according to the aspiration of the Convention on Biological Diversity (CBD) is survival and evolution of biological diversity in its entirety, i.e. wellbeing of all species on Earth. Since endemism is a precondition of extinction, the focus on the survival of endemics and CoEs is essential.
Endemism and endangered species are closely coupled, and different kinds of threat cause different impacts and risks of extinction [227]. According to the IUCN Red List, land use, land use change and intensification of use has a much stronger impact on ecological conditions for most critically endangered species than e.g. climate change. Cities, urban habitats and arable land with prevailing unfavorable conditions for the survival of threatened endemics are continuously growing, while natural and semi-natural habitats harboring large numbers of endemics are declining in quantity and quality. Anthropogenic influences, change and intensification of land use activities, conversion of whole ecosystems, the application of agrochemicals, the growth of arable land and urban environments, and the use of biological resources have the strongest impact and cause the greatest risk for the existence of populations and survival of threatened endemics [228,229,230,231]. Climate change, including severe weather is less important, but has a stronger impact than e.g. natural processes such as tsunamis, volcanism or landslides [232,233,234,235,236].
Vascular plants and vertebrates are by far the most important groups of zoos, aquaria and botanical gardens. Nevertheless, most species on Earth are invertebrates and most of them are not scientifically described. Little is known about their ecological requirements and specific environmental problems. With the exception of certain groups of vascular plants, vertebrates and a few other groups of organisms, most taxa and their ecology have not yet been adequately studied. This knowledge deficit should be seriously considered when ecosystems, landscapes and regions are economically used and ecological conditions are impacted by anthropogenic activities [237,238,239].
If continuity of environmental conditions is the best predictor for survival of endemics, many of them threatened with extinction, then avoidance of intensification of use and change of conditions at landscape scales would simply stabilize conditions of their existence. Avoidance of land use and land cover change at landscape scales can be seen as the most important precondition for ecosystem functioning and survival of biodiversity, even if this is not guaranteed. This is even more important since humankind obviously is not able to limit climate change in the coming years and decades. Even with avoidance of land use changes, climate change is impacting rare species and increasing the risk of extinctions.
Thus, the limitation of direct anthropogenic influences at landscape scales, wherever possible, should have highest priority for maintaining ecosystem functioning, survival of endemics and provision of adequate ecological conditions. Because CoEs harbor high numbers of endemics, monitoring of these areas can seriously contribute to effective nature conservation management with a focus on biodiversity conservation.
The importance of zoos, aquaria and botanical gardens with respect to nature conservation is increasing since decades. However, the distribution of the world’s zoos and botanic gardens in general represents great distances from the world’s CoEs. Botanic gardens are more effective at conservation of endemics, but zoos face far greater challenges in this regard, having to balance the demands of the public, wishing to see iconic, larger species on the one hand with dedicated conservation projects on threatened taxa. In addition, zoos have space limitations and animal welfare including behavior considerations that limit their ability to house large numbers of endemic species. The representation of different groups of taxa is biased. E.g. amphibians compared to invertebrates have a more globally coordinated conservation approach than others. Holdings of endemic invertebrates in zoos and other facilities needs to be increased. Seed banking of plants is clearly space-efficient, but ensuring sufficient genetic diversity is obtained is essential. There is an obvious role for seed banks and botanic gardens in addressing food security challenges – especially in terms of crop wild relatives and indigenous food plants used by peoples around the world.
Compared to the benefits of zoos, aquaria and botanical gardens, little is known about the benefits of the regional and global biodiversity for cultural life and wellbeing. This aspect is also related e.g. to tourism and land use in species-rich regions and CoEs. Ecosystem functions of species-rich and -poor ecoregions and CoEs are directly linked with social aspects including human aesthetics, wellbeing, health, ecology and economy. Nevertheless, this is an open field with many questions that science has to answer in the future. As a first step it might be helpful to intensify the communication between ecologists and members of social sciences [240,241,242,243,244]. Meaningful long term partnerships in endemic species conservation between institutions in countries in the global north and the biodiverse rich and often developing world need to be strongly encouraged, and the “parachute science” approach discouraged [245,246] if there is to be a meaningful increase in the ex situ conservation of endemic and rare taxa from these regions of endemism.
Author Contributions
Conceptualization and first draft preparation C.H., discussion of all parts of the manuscript C.H. and N.B., N.B. collated the section about zoos and botanical gardens, C.H. the tables and visulization of figures. All authors have read, discussed and agreed to the published version of the manuscript.
Funding
No external funding.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Laura Sutcliffe and Merel Herdeg for helpful comments, for support with the design of illustrations, and correcting the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Candolle, A.B. Essai elementaire de geographie botanique. Dictionnaire des sciences naturelles 1820, 18, 1–64. [Google Scholar]
- Bramwell, D. (Ed.) Plants and Islands; Academic Press: London, UK, 1979; ISBN 0-12-125460-7. [Google Scholar]
- Hendrych, R. Material and notes about the geography of the highly stenochoric to monotopic endemic species of the European flora. Acta Universitatis Carolinae-Biologica 1982, 3, 335–372. [Google Scholar]
- Cardona, M.A.; Contandriopoulos, J. Endemism and evolution in the islands of the Western Mediterranean. In Plants and Islands; Bramwell, D., Ed.; Academic Press: London, UK; New York, NY, USA; Toronto, ON, Canada; 2007; pp. 133–170. [Google Scholar]
- Clark, V.R.; Barker, N.P.; Mucina, L. The Sneeuberg: A new centre of floristic endemism on the Great Escarpment. South Africa. S. Afr. J. Bot. 2008, 75, 196–238. [Google Scholar] [CrossRef]
- Bruchmann, I. Plant endemism in Europe: Spatial distribution and habitat affinities of endemic vascular plants. Doctoral Dissertation, University of Flensburg, Germany, 2011. [Google Scholar]
- Anádon-Irizarry, V.; Wege, D.C.; Upgren, A.; Young, R.; Boom, B.; Le´on, Y.M.; Arias, Y.; Koenig, K.; Morales, A.L.; Burke, W.; Perez-Leroux, A.; Levy, S.; Koenig, S.; Gape, L. Sites for priority biodiversity conservation in the Caribbean Islands Biodiversity Hotspot. J. Threatened Taxa 2012, 4, 2806–2844. [Google Scholar] [CrossRef]
- Irl, S.D.H.; Harter, D.E.V. , Steinbauer, M.J.; Puyol, D.G.; Fernandez-Palacios, J.M., Jentsch, A. & Carl Beierkuhlein. Climate vs. topography - spatial patterns of plant species diversity and endemism on a high-elevation island. J. Ecol. 2015, 103, 1621–1633. [Google Scholar] [CrossRef]
- Myers, N. The biodiversity challenge: Expended hotspots analysis. Environmentalist 1990, 10, 243–256. [Google Scholar] [CrossRef]
- Wyk, A.E. Van; Smith, G., F. Regions of floristic endemism in southern Africa: A review with emphasis on succulents; Umdaus Press: Pretoria, South Africa, 2001; ISBN 1919766197. [Google Scholar]
- Melendo, M.; Gimenez, E.; Cano, E.; Gomez-Mercado, F.; Valle, F. The endemic flora in the south of the Iberian Peninsula: Taxonomic composition, biological spectrum, pollination, reproductive mode and dispersal. Flora 2003, 198, 260–276. [Google Scholar] [CrossRef]
- Barthlott, W.; Mutke, J.; Rafiqpoor, D.; Kier, G. , Kreft, H. Global centers of vascular plant diversity. Nova Acta Leopoldina 2005, 92, 61–83. [Google Scholar]
- IUCN, *!!! REPLACE !!!* (Ed.) The IUCN Red List of Threatened Species. Version 2022-2, 2022. Available online: https://www.iucnredlist.org (accessed on 26 January 2023).
- Hobohm, C.; Tucker, C. The increasing importance of endemism, responsibility, the media and education. In Endemism in vascular plants; Hobohm, C., Ed.; Springer: Dordrecht, The Netherlands, 2014; Volume 9, pp. 3–9. [Google Scholar]
- Burgess, N.D.; Clarke, G.P.; Rodgers, W.A. Coastal forests of eastern Africa: Status, endemism patterns and their potential causes. Biol. J. Linn. Soc. 1998, 64, 337–367. [Google Scholar] [CrossRef]
- Chambers, T.C.; Drinnan, A.N.; McLoughlin, S. Some morphological features of Wollemi Pine (Wollemia nobilis: Araucariaceae) and their comparison to cretaceous plant fossils. Int. J. Plant. Sci. 1998, 159, 160–171. [Google Scholar] [CrossRef]
- Ackerman, J.D.; Trejo-Torres, J.C.; Crespo-Chuy, Y. Orchids of the West-Indies: Predictability of diversity and endemism. J. Biogeography 2007, 34, 779–786. [Google Scholar] [CrossRef]
- Clark, V.R.; Bentley, J.; Dold, A.P.; Zikishe, V.; Barker, N.P. The rediscovery of the Great Winterberg endemic Lotononis harveyi B.–E.van Wyk after 147 years, and notes on the poorly known Amathole endemic Macowania revoluta Oliv. (southern Great Escarpment, South Africa). PhytoKeys 2016, 62, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Lesica, P.; Yurkewycz, R.; Crone, E.E. Rare plants are common where you find them. American journal of Botany 2006, 93, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Bevill, R.L.; Louda, S.M. Comparisons of related rare and common species in the study of plant rarity. Conservation Biology 1999, 13, 493–498. [Google Scholar] [CrossRef]
- Lavergne, S.; Thompson, J.D.; Garnier, E.; Debussche, M. The biology and ecology of narrow endemic and widespread plants: A comparative study of trait variation in 20 congeneric pairs. Oikos 2004, 107, 505–518. [Google Scholar] [CrossRef]
- Casazza, C.; Barberis, G.; Minuto, L. Ecological characteristics and rarity of endemic plants of the Italian Maritime Alps. Biol. Conserv. 2005, 123, 361–371. [Google Scholar] [CrossRef]
- Hartley, S.; Kunin, W.E. Scale dependency of rarity, extinction risk, and conservation priority. Conservation biology 2003, 17, 1559–1570. [Google Scholar] [CrossRef]
- Rabinowitz, D. Seven forms of rarity. In The Biological aspects of rare plant conservation; Synge, H., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 1981; pp. 205–217. [Google Scholar]
- Rabinowitz, D.; Cairns, S.; Dillon, T. Seven forms of rarity and their frequency in the flora of British Isles. In Conservation Biology: Science of Scarcity and Diversity; Soulé, M.E., Ed.; Sinauer Associates: Sunderland, Mass, 1986; pp. 182–204. [Google Scholar]
- Barker, N.P.; Fish, L. Rare and infrequent southern African grasses: Assessing their conservation status and understanding their biology. Biodiversity and Conservation 2007, 16, 4051–4079. [Google Scholar] [CrossRef]
- Caiafa, A.N.; Martins, F.R. Forms of rarity of tree species in the southern Brazilian Atlantic rainforest. Biodiversity and conservation 2010, 19, 2597–2618. [Google Scholar] [CrossRef]
- Choe, H.; Thorne, J.H.; Hijmans, R.; Seo, C. Integrating the Rabinowitz rarity framework with a national plant inventory in South Korea. Ecology and evolution 2019, 9, 1353–1363. [Google Scholar] [CrossRef]
- Toledo, L.F.; Becker, C.G.; Haddad, C.F.; Zamudio, K.R. Rarity as an indicator of endangerment in neotropical frogs. Biological Conservation 2014, 179, 54–62. [Google Scholar] [CrossRef]
- McClain, C.R. The commonness of rarity in a deep-sea taxon. Oikos 2021, 130, 863–878. [Google Scholar] [CrossRef]
- Jetz, W.; Rahbek, C.; Colwell, R.K. The coincidence of rarity and richness and the potential signature of history in centres of endemism. Ecology Letters 2004, 7, 1180–1191. [Google Scholar] [CrossRef]
- Kruckeberg, A.R.; Rabinowitz, D. Biological aspects of endemism in higher plants. Annual review of ecology and systematics 1985, 16, 447–479. [Google Scholar] [CrossRef]
- Riemann, H.; Ezcurra, E. Endemic regions of the vascular flora of the peninsula of Baja California, Mexico. J. Veg. Sci. 2007, 18, 327–336. [Google Scholar] [CrossRef]
- Nowak, A.; Nobis, M. Tentative list of endemic vascular plants of the Zeravshan Mts in Tajikistan: Distribution, habitat preferences and conservation status of species. Biodivers. Res. Conserv. 2010, 19, 65–80. [Google Scholar] [CrossRef]
- Brochmann, C.; Rustan, O.H.; Lobin, W.; Kilian, N. The endemic vascular plants of the Cape Verde Islands, W. Africa. Sommerfeltia 1997, 24, 1–356. [Google Scholar] [CrossRef]
- Izquierdo, I.; Martín, J.L.; Zurita, N.; Arechavaleta, M. (Eds). Lista de especies silvestres de Canarias (hongos, plantas, y animales terrestres); Consejería de Medio Ambiente y Ordenación Territorial, Gobierno de Canarias: La Laguna, Spain, 2004; ISBN 84-89729-23-9. [Google Scholar]
- Latheef, S.A.; Prasad, B.; Bavaji, M.; Subramanyam, G. A database on endemic plants at Tirumala hills in India. Bioinformation 2008, 2, 260–262. [Google Scholar] [CrossRef]
- Clark, V.R.; Timberlake, J.R.; Hyde, M.A.; Mapaura, A.; Palgrave, M.C.; Wursten, B.T.; Ballings, P.; Burrows, J.E.; Linder, H.P.; McGregor, G.K.; Chapano, C.; Plowes, D.C.H.; Childes, S.L.; Dondeyne, S.; Müller, T.; Barker, N.P. ; Barker, N.P. A first comprehensive account of floristic diversity and endemism on the Nyanga Massif, Manica Highlands (zimbabwe-Mozambique). Kirkia 2017, 19, 1–53. [Google Scholar]
- Lowry, P.P. Diversity, endemism, and extinction in the flora of New Caledonia: A review. In Rare, threatened, and endangered floras of Asia and the Pacific Rim; Peng, C.I.; Lowry, P.P., Eds.; Academia Sinica Monograph Series; 1998; Volume 16, pp. 181–206.
- Cowling, R.M.; Lombard, A.T. Heterogeneity, speciation/extinction history and climate: Explaining regional plant diversity patterns in the Cape Floristic Region. Divers. Distrib. 2002, 8, 163–179. [Google Scholar] [CrossRef]
- Giménez, E.; Melendo, M.; Valle, F.; Gomez-Mercado, F.; Cano, E. Endemic flora biodiversity in the south of the Iberian peninsula: Altitudinal distribution, life forms and dispersal modes. Biodivers. Conserv. 2004, 13, 2641–2660. [Google Scholar] [CrossRef]
- Helme, N.A.; Trinder-Smith, T.H. The endemic flora of the Cape Peninsula, South Africa. South Afr. J. Bot. 2006, 72, 205–210. [Google Scholar] [CrossRef]
- Kier, G.; Kreft, H.; Lee, T.M.; Jetz, W.; Ibisch, P.L.; Nowicki, C.; Mutke, J.; Barthlott, W. A global assessment of endemism and species richness across island and mainland regions. PNAS 2009, 106, 9322–9327. [Google Scholar] [CrossRef] [PubMed]
- Fenu, G.; Mattana, E.; Congiu, A.; Bacchetta, G. The endemic vascular flora of Supramontes (Sardinia), a high priority plant conservation area. Candollea 2010, 65, 347–358. [Google Scholar] [CrossRef]
- Huang, J.-H.; Chen, B.; Liu, C.; Lai, J.; Zhang, J.; Ma, K. Identifying hotspots of endemic woody seed plant diversity in China. Divers. Distrib. 2011, 18, 673–688. [Google Scholar] [CrossRef]
- Anguinano, M.; Dean, E.; Starbuck, T. Rodriguez, A.; Munguía-Lino, G. Diversity, species richness distribution and centers of endemism of Lycianthes (Capsiceae, Solanaceae) in Mexicao. Phytotaxa 2021, 514, 39–60. [Google Scholar] [CrossRef]
- Svenning, J.-C.; Normand, S.; Skov, F. Plio-Pleistocene climate change and geographic heterogeneity in plant-diversity relationships. Ecography 2009, 32, 13–21. [Google Scholar] [CrossRef]
- Schuldt, A.; Assmann, T. Environmental and historical effects on richness and endemism patterns of carabid beetles in the western Palearctic. Ecography 2009, 32, 705–714. [Google Scholar] [CrossRef]
- Fritz, S.A.; Rahbek, C. Global patterns of amphibian phylogenetic diversity. J. Biogeogr. 2012, 39, 1373–1382. [Google Scholar] [CrossRef]
- Voskamp, A.; Baker, D.J.; Stephens, P.A.; Valdes, P.J.; Willis, S.G. Global patterns in the divergence between phylogenetic diversity and species richness in terrestrial birds. J. Biogeogr. 2017, 44, 709–721. [Google Scholar] [CrossRef]
- Taylor, P.J.; Kearney, T.; Dalton, D.L.; Chakona, C.; Kelly, C.M.R.; Barker, N.P. Biomes, geology and past climate drive speciation of laminate-toothed rats on South African mountains (Murinae: Otomys). Zoological Journal of the Linnean Society 2019, 20, 1–21. [Google Scholar] [CrossRef]
- Hobohm, C.; Beierkuhnlein, C.; Börtitz, C.; Clark, V.R.; El Balti, N.; Fichtner, A.; Franklin, S.; Gaens, T.; Härdtle, W.; Hansen, A.S.; Janišová, M.; Jansen, J.; Lindner, M.; Moro-Richter, M.; Müller-Benedict, V.; Ott, K.; Reinmuth, K.C.; van Rooijen, N.; Sandberg, M.; Schamineé, J.H.J.; Tang, C.Q.; Vahle, H.-C.; Vanderplank, S.E. Land Use Change and the Future of Biodiversity. In Environmental Challenges and Solutions; Springer: Cham, 2021; pp. 451–483. [Google Scholar]
- Fjeldsa, J.; Lovett, J.C. Geographical patterns of old and young species in African forest biota: The significance of specific montane areas as evolutionary centres. Biodiv. Cons. 1997, 6, 325–346. [Google Scholar] [CrossRef]
- Goldie, X.; Gillman, L.; Crisp, M.; Wright, S. Evolutionary speed limited by water in arid Australia. Proc. Biol. Sci. 2010, 277, 2645–2653. [Google Scholar] [CrossRef] [PubMed]
- Bruchmann, I.; Hobohm, C. Factors that create and increase endemism. - In, Hobohm, C., Ed. Endemism in vascular plants. Plant and Vegetation 2014, 9, 51–68. [Google Scholar]
- Alagador, D.; Cerdeira, J.O.; Araujo, M.B. Climate change, species range shifts and dispersal corridors: An evaluation of spatial conservation models. Methods in Ecology and Evolution 2016, 7, 853–866. [Google Scholar] [CrossRef]
- Zobel, M. The species pool concept as a framework for studying patterns of plant diversity. J. Veg. Sci. 2016, 27, 8–18. [Google Scholar] [CrossRef]
- Zizka, G.; Klemmer, K. Pflanzen- und Tierwelt der Galapagos-Inseln - Entstehung, Erforschung, Gefährdung und Schutz. Kleine Senckenbergreihe 1994, 20, 1–151. [Google Scholar]
- Wilmé, L.; Ravokatra, M.; Dolch, R.; Schuurman, D.; Mathieu, E.; Schuetz, H.; Waeber, P.O. Toponyms for centers of endemism in Madagascar. Madagascar Conservation & Development 2012, 7, 30–40. [Google Scholar]
- Cordier, J.M.; Lescano, J.N.; Ríos, N.E.; Leynaud, G.C.; Nori, J. Climate change threatens micro-endemic amphibians of an important South American high-altitude center of endemism. Amphibia-Reptilia 2019, 41, 233–243. [Google Scholar] [CrossRef]
- Olefeld, J.L.; Bock, C.; Jensen, M.; Vogt, J.C.; Sieber, G.; Albach, D.; Boenigk, J. ; Boenigk, J. Centers of endemism of freshwater protists deviate from pattern of taxon richness on a continental scale. Scientific Reports 2020, 10, 14431. [Google Scholar] [CrossRef]
- Suissa, J.S.; Sundue, M.A. Diversity Patterns of Neotropical Ferns: Revisiting Tryon’s Centers of Richness and Endemism. American Fern Journal 2020, 110, 211–232. [Google Scholar] [CrossRef]
- Robuchon, M.; Pavoine, S.; Véron, S.; Delli, G.; Faith, D.P.; Mandrici, A.; Pellens, R.; Dubois, G.; Leroy, B. Revisiting species and areas of interest for conserving global mammalian phylogenetic diversity. Nat. Comm. 2021, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.D.; Heywood, V.H.; Hamilton, A.C. (Eds.) Centres of plant diversity: Vol 1, Europe, Africa, South West Asia and the Middle East; IUCN Publications: Cambridge, UK, 1994; ISBN 2-8317-0197-X. [Google Scholar]
- Davis, S.D.; Heywood, V.H. , Hamilton, A.C. (Eds.) Centres of plant diversity: Vol 2, Asia, Australasia and the Pacific; IUCN Publications: Cambridge, UK, 1995; ISBN 2-8317-0198-8. [Google Scholar]
- Davis, S.D.; Heywood, V.H.; Herrera-MacBryde, O.; Villa-Lobos, J.; Hamilton, A.C. (Eds.) Centres of plant diversity: Vol 3, the Americas; IUCN Publications: Cambridge, UK, 1997; ISBN 2-8317-0199-6. [Google Scholar]
- Olefeld, J.L.; Bock, Jensen, M. ; Vogt, J.C.; Sieber, G.; Albach, D.; Boenigk, J. Centers of endemism of freshwater protists deviate from pattern of taxon richness on a continental scale. Scientific Reports 2020, 10, 14431. [Google Scholar] [CrossRef] [PubMed]
- Waeber, P. , Wilmé, L.; Mercier, J.L.; Rakotozafy, L.M.A.; Garcia, C.; Sorg, J.P. The role of lakes in the context of the centers of endemism. Akon’ny Ala 2015, 32, 34–47. [Google Scholar]
- Clark, V.R.; Barker; N. P.; Mucina, L. The Sneeuberg: A new centre of floristic endemism on the Great Escarpment, South Africa. S. Afr. J. Bot. 2009, 75, 196–238. [Google Scholar] [CrossRef]
- Clark, V.R.; Dold, A.P.; McMaster, C.; McGregor, G.; Bredenkamp, C.; Barker, N. Rich sister, poor cousin: Plant diversity and endemism in the Great Winterberg-Amatholes (Great Escarpment, Eastern Cape, South Africa). South African Journal of Botany 2014, 92, 159–174. [Google Scholar] [CrossRef]
- Clayton, W.D. Chorology of the genera of Gramineae. Kew Bulletin 1975, 30, 111–132. [Google Scholar] [CrossRef]
- Fattorini, S. A history of chorological categories. History and Philosophy of the Life Sciences 2016, 38, 1–21. [Google Scholar] [CrossRef]
- Linder, H.P. , J., Mutke, J.M., Barthlott, W., Jürgens, N., Rebelo, T.; Küper, W. A numerical re-evaluation of the sub-Saharan phytochoria of mainland Africa. In Plant diversity and complexity patterns: Local, regional and global dimensions. Proceedings of an International Symposium held at the Royal Danish Academy of Sciences and Letters in Copenhagen, Denmark, 25-28 May, 2003.; Küper, W. A numerical re-evaluation of the sub-Saharan phytochoria of mainland Africa. In Plant diversity and complexity patterns: Local, regional and global dimensions. Proceedings of an International Symposium held at the Royal Danish Academy of Sciences and Letters in Copenhagen, Denmark, 25-28 May, 2003; pp. 2005229–252. [Google Scholar]
- Bradshaw, P.L.; Colville, J.F.; Linder, H.P. Optimising regionalisation techniques: Identifying centres of endemism in the extraordinarily endemic-rich Cape Floristic Region. PLoS ONE 2015, 10, e0132538. [Google Scholar] [CrossRef]
- Hobohm, C.; Tucker, C. How to quantify endemism. Plant and Vegetation 2014, 9, 11–48. [Google Scholar]
- Rosauer, D.; Laffan, S.W.; Crisp, M.D.; Donnellan, S.C.; Cool, L.G. Phylogenetic endemism: A new approach for identifying geographical concentrations of evolutionary history. Mol. Ecol. 2009, 18, 4061–4072. [Google Scholar] [CrossRef] [PubMed]
- Chiarucci, A.; Beierkuhnlein, C.; Essl, F.; Fernández-Palacios, J.M.; Jentsch, A.; Hobohm, C.; Kreft, H.; Krestov, P.V.; Löbel, S.; Steinbauer, M.J.; Storch, D.; Triantis, K.; Weigelt, P.; Dengler, J. Global patterns of vascular plant species richness and endemicity, a new approach to identify hotspots. In Biodiversity & Vegetation: Patterns, Processes, Conservation; Mucina, L., Price, J.N., Kalwij, J.M., Eds.; Kwongan Foundation: Perth, AU, USA, 2014; p. 78. [Google Scholar]
- Herrera, J.P. Prioritizing protected areas in Madagascar for lemur diversity using a multidimensional perspective. Biol. Cons. 2017, 207, 1–8. [Google Scholar] [CrossRef]
- Cowling, R.M.; Samways, M.J. Predicting global patterns of endemic plant species richness. Biodiversity Letters 1995, 2, 127–131. [Google Scholar] [CrossRef]
- Hobohm, C.; Janišová, M.; Steinbauer, M.; Landi, S.; Field, R.; Vanderplank, S.; Beierkuhnlein, C.; Grytnes, J.-A.; Vetaas, O.R.; Fildelis, A.; De Nascimento. L.; Clark, V.R.; Fernández-Palacios, J.M.; Franklin, S.; Guarino, R.; Huang, J.; Krestov, P.; Ma, K.; Onipchenko, V.; Palmer, M.W.; Fragomeni, S.M.; Stolz, C.; Chiarucci, A. Global endemics-area relationships of vascular plants. Perspectives Ecol. Cons. 2019, 17, 41–49. [Google Scholar]
- Daru, B.H.; Farooq, H.; Antonelli, A.; Faurby, S. Endemism patterns are scale dependent, Nature Communications 2020, 11, 1–11. 11. [CrossRef]
- Shipley, B.; McGuire, J.L. Interpreting and integrating multiple endemism metrics to identify hotspots for conservation priorities. Biol. Cons. 2021, 265, 109403. [Google Scholar] [CrossRef]
- Jacobs, J.G.; Lashley, M.A.; Cove, M.V. Fawn counts and adult female site use are mismatched indicators of habitat quality in an endangered deer. Diversity 2021, 13, 92. [Google Scholar] [CrossRef]
- Pinzari, C.; Peck, R.; Zinn, T.; Gross, D.; Montoya-Aiona, K.; Brinck, K.; Gorresen, M.; Bonaccorso, F. Hawaiian Hoary Bat (Lasiurus cinereus semotus): Activity, diet and prey availability at the Waihou Mitigation Area, Maui. Technical Report 2019 HCSU-090, 1-60.
- Romo, H. , García-Barros, E.; Lobo, J.M. Identifying recorder-induced geographic bias in an Iberian butterfly database. Ecography 2006, 29, 873–885. [Google Scholar] [CrossRef]
- Grant, P.B.C.; Samways, M.J. Micro-hotspot determination and buffer zone value for Odonata in a globally significant biosphere reserve. Biol. Conserv. 2011, 144, 772–781. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2022-2. 2023. Available online: https://www.iucnredlist.org (accessed on 17 January 2023).
- Bailey, R.G. Ecoregions: The geography of oceans and continents; Springer; New York, NY, USA, 1998; ISBN 0-387-98311-2.
- Wiens, J.J. Faster diversification on land than sea helps explain global biodiversity patterns among habitats and animal phyla. Ecology Letters 2015, 18, 1234–1241. [Google Scholar] [CrossRef]
- Thode, V.A.; Inacio, C.D.; Eggers, L.; Reginato, M.; Souza-Chies, T.T. Spatial-temporal evolution and diversification in Sisyrinchium (Iridaceae) with emphasis on abiotic drivers. Bot. J. Linnean Soc. 2021, 199, 93–108. [Google Scholar] [CrossRef]
- Teske, P.R.; Sandoval-Castillo, J.; Waters, J.; Beheregaray, L.B. An overview of Australia's temperate marine phylogeography, with new evidence from high-dispersal gastropods. Journal of Biogeography 2017, 44, 217–229. [Google Scholar] [CrossRef]
- Murali, G.; Gumbs, R.; Meiri, S.; Roll, U. Global determinants and conservation of evolutionary and geographic rarity in land vertebrates. Science Advances 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Govaerts, R. How many species of seed plants are there? Taxon 2001, 50, 1085–1090. [Google Scholar] [CrossRef]
- Lydeard, C.; Cowie, R.H.; Ponder, W.F.; Bogan, A.E.; Bouchet, P.; Clark, S.A.; Cummings, K.S.; Frest, T.J.; Gargominy, O.; Herbert, D.G.; Hershler, R.; Perez, K.E.; Roth, B.; Seddon, M.; Strong, E.E.; Thompson, A.G. The global decline of nonmarine mollusks. BioScience 2004, 54, 321–330. [Google Scholar] [CrossRef]
- Paton, A.J.; Brummit, N.; Govaerts, R.; Harman, K.; Hinchcliffe, S.; Allkin, B.; Lughadha, N. . Towards target 1 of the global strategy for plant conservation: A working list of all known plant species – progress and prospects. Taxon 2008, 57, 602–611. [Google Scholar]
- Chapman, A.D. Numbers of Living Species in Australia and the World, 2nd. ed.; Canberra, Australia, 2009. ISBN 978-0-642-56860-1.
- Barrowclough, G.F.; Cracraft, J.; Klicka, J.; Zink, R.M. How many kinds of birds are there and why does it matter? PLoS ONE 2016, 11, e0166307. [Google Scholar] [CrossRef]
- Groombridge, B.; Jenkins, D. World atlas of biodiversity; University of California Press: Berkeley, 2002; ISBN 0-520-23668-8. [Google Scholar]
- Whittaker, R.J.; Fernández-Palacios, J. Island biogeography: Ecology, evolution, and conservation, 2nd ed.; Oxford University Press: Oxford, UK, 2007; ISBN 9780198566120. [Google Scholar]
- Rabitsch, W.; Essl, F. (Eds.) Endemiten – Kostbarkeiten in ¨Osterreichs Pflanzen- und Tierwelt; Umweltbundesamt: Vienna, Austria, 2009; ISBN 978-3-85328-049-2. [Google Scholar]
- Hobohm, C.; Janišová, M.; Jansen, J.; Bruchmann, I.; Deppe, U. 2014. Biogeography of endemic vascular plants - overview. In: Hobohm C (Ed). Endemism in vascular plants. Plant and Vegetation 2014, 9, 85–163. [Google Scholar]
- Walter, K.S. ; H. J.; Gillett, H.J. (Eds.) IUCN Red List of Threatened Plants, Ed.; The World Conservation Union: Gland, Switzerland; Cambridge, UK, 1998; ISBN 2-8317-0328-X. [Google Scholar]
- Vanderplank, S.E.; Moreira-Muñoz, A.; Hobohm, C.; Pils, G.; Noroozi, J.; Clark, V.R.; Barker, N.P.; Yang, W.; Huang, J.; Ma, K.; Tang, C.Q.; Werger, M.J.A.; Ohsawa, M.; Yang, Y. ,. Endemism in mainland regions: Case studies. Plant and Vegetation 2014, 9, 205–308. [Google Scholar]
- Zhang, Z.; Yan, Y.; Tian, Y.; He, J.-S.; Tang, Z. Distribution and conservation of orchid species richness in China. Biol. Cons. 2015, 181, 64–72. [Google Scholar] [CrossRef]
- Forman, R.T. Land mosaics: The ecology of landscapes and regions; Cambridge Univ. Press: Cambridge, UK, 1995; ISBN 978-1-59726-646-8. [Google Scholar]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends in Ecology & Evolution.
- Olofsson, J. , de Mazancourt, C.; Crawley, M.J. Spatial heterogeneity and plant species richness at different spatial scales under rabbit grazing. Oecologia 2008, 156, 825–834. [Google Scholar] [CrossRef]
- Megías, A.G.; Gómez, J.M.; Sánchez-Pinero, F. Spatio-temporal change in the relationship between habitat heterogeneity and species diversity. Acta Oecologica 2011, 37, 179–186. [Google Scholar] [CrossRef]
- Chen, J.; Shiyomi, M.; Wei, Z. 2015; 61. [CrossRef]
- Huston, M.A. Biological diversity: The coexistence of species on changing landscapes; Cambridge University Press: Cambridge, UK, 1994; ISBN 0-521-36093-5. [Google Scholar]
- Fortin, M.-J.; Dale, M.R.T. Spatial analysis: A guide for ecologists; Cambridge Univ. Press: Cambridge, UK, 2005; ISBN 0521143500. [Google Scholar]
- Pugh, P.J.A.; Convey, P. Surviving out in the cold: Antarctic endemic invertebrates and their refugia. J. Biogeogr. 2008, 35, 2176–2186. [Google Scholar] [CrossRef]
- Huang, J.-H.; Chen, B.; Ying, J.-S.; Ma, K. Features and distribution patterns of Chinese endemic seed plant species. J. Syst. Evol. 2011, 49, 81–94. [Google Scholar] [CrossRef]
- Loiseau, N.; Mouquet, N.; Casajus, N.; Grenié, M.; Guéguen, M.; Maitner, B.; Mouillot, D.; Ostling, A.; Renaud, J.; Tucker, C.; Velez, L.; Thuiller, W.; Violle, C. ; Violle, C. Global distribution and conservation status of ecologically rare mammal and bird species, Nature Communications 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Pawlowski, B. Der Endemismus in der Flora der Alpen, der Karpaten und der Balkanischen Gebirge im Verhältnis zu den Pflanzengesellschaften. Mitteilungen der ostalpin-dinarischen pflanzensoziologischen Arbeitsgemeinschaft 1969, 9, 167–178. [Google Scholar]
- Talbot, S.S.; Yurtsev, B.A.; Murray, D.F.; Argus, G.W.; Bay, C.; Elvebakk, A. Atlas of rare endemic vascular plants in the Arctic. Conservation of Arctic flora and fauna (CAFF). Technical Report 1999, 3, 1–73. [Google Scholar]
- Beard, J.S.; Chapman, A.R.; Gioia, P. Species richness and endemism in the Western Australian flora. J. Biogeogr. 2000, 27, 1257–1268. [Google Scholar] [CrossRef]
- Jonsell, B.; Karlsson, T. Endemic vascular plants in Norden. In Flora Nordica. General volume; Jonsell, B., Ed.; Bergius Foundation: Stockholm, Sweden, 2004; pp. 139–159. [Google Scholar]
- Wollenberg, K.C.; Vieites, D.R.; van der Meijden, A.; Glaw, F.; Cannatella, D.C.; Vences, M. Patterns of endemism and species richness in Malagasy cophyline frogs support a key role of mountainous areas for speciation. Evolution 2008, 62, 1890–1907. [Google Scholar] [CrossRef]
- Colwell, R.K.; Lees, D.C. The mid-domain effect: Geometric constraints on the geography of species richness. Trends in Ecology & Evolution 2000, 15, 70–76. [Google Scholar]
- McCain, C.M. The mid-domain effect applied to elevational gradients: Species richness of small mammals in Costa Rica. Journal of Biogeography 2004, 31, 19–31. [Google Scholar] [CrossRef]
- McCain, C.M. Elevational gradients in diversity of small mammals. Ecology 2005, 86, 366–372. [Google Scholar] [CrossRef]
- McCain, C.M.; Grytnes, J.-A. . Elevational Gradients in Species Richness. In Encyclopedia of Life Sciences (ELS); 2010. [CrossRef]
- Storch, D.; Keil, P.; Jetz, W. Universal species-area and endemics-area relationships at continental scales. Nature 2012, 488, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Varzinczak, L.H.; Zanata, T.B.; Moura, M.O.; Passos, F.C. Geographical patterns and current and short-term historical correlates of phylogenetic diversity and endemism for New World primates. J. Biogeography 2019, 47, 890–902. [Google Scholar] [CrossRef]
- Orme, C.; Davies, R.; Burgess, M.; Eigenbrod, F.; Pickup, N.; Olson, V.A.; Webster, A.J.; Ding, T.-S.; Rasmussen, P.C.; Ridgely, R.S.; Stattersfield, A.J.; Bennett, P.M. , Blackburn, T.M.; Gaston, K.J.; Owens, I.P.F. Global hotspots of species richness are not congruent with endemism or threat. Nature 2005, 436, 1016–1019. [Google Scholar] [CrossRef] [PubMed]
- Wikelski, M.; Wilcove, D.S. Endemic species in the narrowest niches. In The atlas of global conservation; Molnar, J.L., Ed.; University of California Press: Berkeley, 2010; ISBN 978-0-520-26256-0. [Google Scholar]
- Cronk, Q.C.; Percy, D.M. Endemism. In Encyclopedia of islands; Gillespie, R.G., Clague, D.A., Eds.; University of California Press: Berkeley, USA, 2009. [Google Scholar]
- Hobohm, C. Plant species diversity and endemism on islands and archipelagos - with special reference to the Macaronesian Islands. Flora 2000, 195, 9–24. [Google Scholar] [CrossRef]
- Hobohm, C.; Janišová, M.; Jansen, J.; Bruchmann, I.; Deppe, U. Biogeography of endemic vascular plants - overview. Plant and Vegetation 2014, 9, 85–163. [Google Scholar]
- Hobohm, C.; Müller-Benedict, V. Vergleich der Biodiversität insularer und kontinentaler Regionen unter besonderer Berücksichtigung der Endemitenvielfalt. Ber. RTG 2018, 30, 57–71. [Google Scholar]
- Stuessy, T.F.; Grau, J.; Zizka, G. Diversidad de plantas en las Islas Robinson Crusoe. In: Grau, J.; Zizka, G. Eds. Flora silvestre de Chile. Palmengarten Sonderheft 1992, 19, 54–66. [Google Scholar]
- Tryon, R. Biogeography of the Antillean fern flora. In Plants and islands; Bramwell, D., Ed.; 1979; pp. 55–68.
- Balslev, H.; Valencia, R.; Paz y Min ̃o, G.; Christensen, H.; Nielsen, I. Species count of vascular plants in one hectare of humid lowland forest in Amazonian Ecuador. In Forest biodiversity in North, Central and South America, and the Caribbean: Research and Monitoring; Dallmeier, F., Comiskey, J.A., Eds.; UNESCO: Paris, France, 1998; pp. 585–594. [Google Scholar]
- Rolecek, J.; Drevojan, P.; Hajkova, P.; Goia, I.; Hajek, M. Update on maxima of fine-scale vascular plant species richness in a Transsylvanian steppe meadow. Tuexenia 2021, 41, 459–466. [Google Scholar]
- Pan, Y.; Birdsey, R.A.; Phillips, O.L.; Jackson, R.B. The structure, distribution, and biomass of the world’s forests. Ann. Rev. Ecol. Evol. Syst. 2013, 593–622. [Google Scholar] [CrossRef]
- Keith, H.; Mackey, B.G.; Lindenmayer, D.B. Re-evaluation of forest biomass carbon stocks and lessons from the world’s most carbon-dense forests. Proc. Natl. Acad. Sci. 2009, 106, 11635–40. [Google Scholar] [CrossRef] [PubMed]
- Morison, J.I.L.; Piedade, M.T.F.; Müller, E.; Long, S.P.; Junk, W.J.; Jones, M.B. Very high productivity of the C4 aquatic grass Echinochloa polystachya in the Amazon floodplain confirmed by net ecosystem CO2 flux measurements. Oecologia 2000, 125, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Piedade, M.T.F.; Long, S.P.; Junk, W.J. Leaf and canopy photosysthesis CO2 uptake of a stand of Echinochloa polystachya on the Amazon floodplain. Funct. Ecol. 1994, 11, 60–65. [Google Scholar] [CrossRef]
- Stuessy, T.F.; Ono, M. (Eds.) Evolution and speciation of island plants; Cambridge University Press: Cambridge, UK, 1998; ISBN 0-521-49653-5. [Google Scholar]
- Roberts, B.A.; Proctor, J. (Eds.) The ecology of areas with serpentinized rocks: A world view; Kluwer Academic Publishers: Dordrecht, Netherlands, 1992; ISBN 978-94-011-3722-5. [Google Scholar]
- Stevanovic, V.; Tan, K.; Iatrou, G. Distribution of the endemic Balkan flora on serpentine I: Obligate serpentine endemics. Plant. Syst. Evol. 2003, 242, 149–170. [Google Scholar] [CrossRef]
- Hobohm, C. , Moro-Richter, M.; Beierkuhnlein, C. Distribution and Habitat Affinity of Endemic and Threatened Species - Global Assessment. In Environmental Challenges and Solutions; 2021; pp. 233–277.
- MacArthur, R.; Wilson, E.O. Island Biogeography; Princeton University Press: Princeton, UK, 1967; ISBN 0-691-08836-5. [Google Scholar]
- Humphries, C.J. Endemism and evolution in Macaronesia. In Plants and islands; Bramwell, D., Ed.; Academic Press: London, UK, 1979. [Google Scholar]
- Givnish, T.J. Adaptive radiation, dispersal, and diversification of the Hawaiian Lobeliads. In The biology of biodiversity; Kato, M., Ed.; Springer: Tokyo, Japan, 2000; pp. 67–90. [Google Scholar]
- Mucina, L.; Wardell-Johnson, G. Landscape age and soil fertility, climatic stability, and fire regime predictability: Beyond the OCBIL framework. Plant Soil 2011, 341, 1–23. [Google Scholar] [CrossRef]
- Rosauer, D.; Jetz, W. Phylogenetic endemism in terrestrial mammals. Global Ecol. Biogeogr. 2014, 24, 168–179. [Google Scholar] [CrossRef]
- Médail, F.; Verlaque, R. Ecological characteristics and rarity of endemic vascular plants from Southeastern France and Corsica: Implications for biodiversity conservation. Biol. Cons. 1997, 80, 269–281. [Google Scholar] [CrossRef]
- Pärtel, M. Local plant diversity patterns and evolutionary history at local scale. Ecology 2002, 83, 2361–2366. [Google Scholar] [CrossRef]
- Price, M.R.; Hadfield, M.G. Population genetics and the effects of a severe bottleneck in an ex situ population of critically endangered hawaiian tree snails. PLoS ONE 2014, 9, e114377. [Google Scholar] [CrossRef]
- Moreira-Muñoz, A.; Elórtegui Francioli, S.; Hobohm, C. , Sequeira, M. de 2014. Endemism on islands: Case Studies. Plant and Vegetation 2014, 9, 165–204. [Google Scholar]
- Linder, H.P.; Barker, N.P. Does polyploidy facilitate long-distance dispersal? Annals of Botany 2014, 113, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Meudt, H.M.; Albach, D.C.; Tanentzap, A.J.; Igea, J.; Newmarch, S.C.; Brandt, A.J.; Lee, W.G.; Tate, J.A. Polyploidy on islands: Its emergence and importance for diversification. Frontiers in plant science 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Reiss, M.J. Optimization theory in behavioural ecology. J. Biol. Educ. 1987, 21, 241–247. [Google Scholar] [CrossRef]
- Parker, G.A.; Maynard Smith, J. Optimality theory in evolutionary biology. Nature 1990, 348, 27–33. [Google Scholar] [CrossRef]
- Richardson, R. C Optimization in evolutionary ecology. In Proceedings of the Biennial Meeting of the Philosophy of Science Foundation; Cambridge University Press, 1994; Volume 1, pp. 13–21. [Google Scholar]
- Fukami, T.; Morin, P.J. Productivity-species diversity relationships depend on the history of community assembly. Nature 2003, 424, 423–426. [Google Scholar] [CrossRef]
- Goh, C.K.; Tan, K.C. Evolutionary multi-objective optimization. In uncertain environments: Issues and algorithms; Springer: Berlin, Germany, 2009; ISBN 978-3-540-95975-5. [Google Scholar]
- Reluga, T.C.; Shaw, A.K. Resource distribution drives the adaption of migratory, partially migratory, or residential strategies. Theor. Ecol. 2015, 8, 437–447. [Google Scholar] [CrossRef]
- McGlone, M.S.; Duncan, R.P.; Heenan, P.B. Endemism, species selection and the origin and distribution of the vascular plant flora of New Zealand. J. Biogeogr. 2001, 28, 199–216. [Google Scholar] [CrossRef]
- López-Aguirre, C.; Hand, S.J.; Laffan, S.W.; Archer, M. ; Archer, M. Zoogeographical regions and geospatial patterns of phylogenetic diversity and endemism of New World bats. Ecography 2019, 6, 1188–1199. [Google Scholar] [CrossRef]
- Hobohm, C.; Vanderplank, S.E. Resources for Humans, Plants and Animals: Who is the Ruler of the Driver? And: Can Resource Use Explain Everything? In Environmental Challenges and Solutions; 2021; pp. 79–106.
- Beierkuhnlein, C.; Walentowitz, A.; Welss, W. FloCan—A Revised Checklist for the Flora of the Canary Islands. Diversity 2021, 13, 480. [Google Scholar] [CrossRef]
- Dynesius, M.; Jansson, R. Evolutionary consequences of changes in species' geographical distributions driven by Milankovitch climate oscillations. Proceedings of the National Academy of Sciences 2000, 97, 9115–9120. [Google Scholar] [CrossRef]
- Jansson, R.; Dynesius, M. The fate of clades in a world of recurrent climatic change: Milankovitch oscillations and evolution. Annual review of ecology and systematics 2002, 33, 741–777. [Google Scholar] [CrossRef]
- Magellan, K.; Weyl, O.L.F.; Booth, A.J. Preference for artificial refugia over natural refugia in an endangered fish. Diversity 2021, 13, 635. [Google Scholar] [CrossRef]
- Jansson, R. Global patterns in endemism explained by past climatic change. Proc. R. Soc. Lond. B 2003, 270, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Magoulick, D.D.; Kobza, R.M. The role of refugia for fishes during drought: A review and synthesis. Freshw. Biol. 2003, 48, 1186–1198. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Tang, C.Q.; Matsui, T.; Ohashi, H.; Dong, Y.-F.; Momohara, A.; Herrando-Moraira, S.; Qian, S.; Yang, Y.; Ohsawa, M.; Luu, H.T.; Grote, P.J.; Krestov, P.V.; LePage, B.; Werger, M.; Robertson, K.; Hobohm, C.; Wang, C.-Y.; Peng, M.-C. , Chen, X.; Wang, H.-C.; Su, W.-H.; Zhou, R.; Li, S.; He, L.-Y.; Yan, K.; Zhu, M.-Y.; Hu, J.; Yang, R.-H.; Li, W.-J.; Tomita, M.; Wu, Z.-L.; Yan, H.-Z.; Zhang; G.-F.; He, H.; Yi. S.-R.; Gong, H.; Song, K.; Song, D.; Li, X.-S.; Zhang, Z.-Y.; Han, P.-B.; Shen, L.-Q.; Huang, D.-S.; Luo, K.; López-Pujol, J. Identifying long-term stable refugia for relict plant species in East Asia. Nature Communications 2018, 9, 4488. [Google Scholar] [CrossRef] [PubMed]
- Casazza, G.; Zappa, E.; Mariotti, M.G.; Medail, F.; Minuto, L. Ecological and historical factors affecting distribution pattern and richness of endemic plant species: The case of the maritime and Ligurian Alps hotspot. Divers. Distrib. 2008, 14, 47–58. [Google Scholar] [CrossRef]
- López-Aguirre, C.; Hand, S.J.; Laffan, S.W.; Archer, M. ; Archer, M. Phylogenetic diversity, types of endemism and the evolutionary history of New World bats. Ecography 2018, 41, 1955–1966. [Google Scholar] [CrossRef]
- Sampaio, M.B.; Schiel, N.; da Silva Souto, A. , 2020. From exploitation to conservation: A historical analysis of zoos and their functions in human societies. Ethnobiology and Conservation 2020, 9. [Google Scholar] [CrossRef]
- Marešová, J.; Frynta, D. Noah's Ark is full of common species attractive to humans: The case of boid snakes in zoos. Ecological Economics 2008, 64, 554–558. [Google Scholar] [CrossRef]
- Bowkett, A.E. Recent captive-breeding proposals and the return of the ark concept to global species conservation. Conservation Biology 2009, 23, 773–776. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, T.; Gardner, R.; Kraaijeveld, A.R.; Riordan, P. Contributions of zoos and aquariums to reintroductions: Historical reintroduction efforts in the context of changing conservation perspectives. International Zoo Yearbook 2017, 51, 15–31. [Google Scholar] [CrossRef]
- Fraser, J.; Wharton, D. , 2007. The future of zoos: A new model for cultural institutions. Curator: The Museum Journal 2007, 50, 41–54. [Google Scholar] [CrossRef]
- Palmer, C.; Kasperbauer, T.J.; Sandøe, P. Bears or butterflies? How should zoos make value-driven decisions about their collections. In The Ark and Beyond: The Evolution of Zoo and Aquarium Conservation; Minteer, B.A., Maienschein, J., Collins, J.P., Eds.; University of Chicago Press: Chicago, London, 2018; ISBN 978-0-226-538446-4. [Google Scholar]
- Brereton, J.; Brereton, S. Sixty years of collection planning: What species do zoos and aquariums keep? International Zoo Yearbook. 2020, 54, 131–145. [Google Scholar] [CrossRef]
- Martin, T.E.; Lurbiecki, H.; Joy, J.B.; Mooers, A.O. Mammal and bird species held in zoos are less endemic and less threatened than their close relatives not held in zoos. Animal Conservation 2014, 17, 89–96. [Google Scholar] [CrossRef]
- Fattorini, S.; Mantoni, C.; Dapporto, L.; Davini, G.; Di Biase, L. Using Botanical Gardens as Butterfly Gardens: Insights from a Pilot Project in the Gran Sasso and Monti Della Laga National Park (Italy). Conservation 2023, 3, 109–126. [Google Scholar] [CrossRef]
- Swanson, W.F.; Johnson, W.E.; Cambre, R.C.; Citino, S.B.; Quigley, K.B.; Brousset, D.M.; Morais, R.N.; Moreira, N.; O'Brien, S.J.; Wildt, D.E. Reproductive status of endemic felid species in Latin American zoos and implications for ex situ conservation. Zoo Biology: Published in affiliation with the American Zoo and Aquarium Association 2003, 22, 421–441. [Google Scholar] [CrossRef]
- Barthlott, W.; Rauer, G.; Ibisch, P.L.; von den Driesch, M.; Lobin, W. Biodiversität und Botanische Gärten. In: Bundesamt für Naturschutz (Ed.): Botanische Gärten und Biodiversität. Erhaltung Biologischer Vielfalt durch Botanische Gärten und die Rolle des Übereinkommens über die Biologische Vielfalt (Rio de Janeiro, 1992). Landwirtschaftsverlag, Münster, Germany, 1999, pp. 1–24. ISBN 978-3-89624-615-8.
- Price, M.R.S.; Maunder, M.; Soorae, P.S.; Guerrant, E.O.; Havens, K. Ex situ support to the conservation of wild populations and habitats: Lessons from zoos and opportunities for botanic gardens. Ex situ plant conservation: Supporting species survival in the wild 2004, 3, 84. [Google Scholar]
- Conde, D.; Colchero, F.; Gusset, M.; Pearce-Kelly, P.; Byers, O.; Flesness, N.; Browne, R.; Jones, O. Zoos through the lens of the IUCN Red List: A global metapopulation approach to support conservation breeding programs. PLoS ONE 2013, 8, e80311. [Google Scholar] [CrossRef]
- Lammers, R.; Scholten, C.; Marcordes, B.; Pagel, T.; Rödder, D.; Ziegler, T. Malagasy birds in zoological gardens–an analysis of zoo databases as basis for improved ex situ conservation measures. Madagassische Vögel in Zoologischen Gärten - Zoodatenbankanalysen als Grundlage für verbesserten ex- situ-Artenschutz. Der Zoologische Garten 2022, 90, 121–150. [Google Scholar] [CrossRef]
- Carr, N. Star attractions and damp squibs at the zoo: A study of visitor attention and animal attractiveness. Tourism Recreation Research 2016, 41, 1–13. [Google Scholar] [CrossRef]
- Biega, A.; Lamont, M.; Mooers, A.; Bowkett, A.; Martin, T. Guiding the prioritization of the most endangered and evolutionary distinct birds for new zoo conservation programs. Zoo Biology 2019, 38. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Hilton-Taylor, C.; Angulo, A.; Böhm, M.; Brooks, T.M.; Butchart, S.H.; Carpenter, K.E.; Chanson, J.; Collen, B.; Cox, N.A.; Darwall, W.R. The impact of conservation on the status of the world’s vertebrates. Science 2010, 330, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Browne, R.K. , Wolfram, K., García, G., Bagaturov, M.F. and Pereboom, Z.J.J.M., 2011. Zoo-based amphibian research and conservation breeding programs. Amphibian and Reptile Conservation 2011, 5, 1–14. [Google Scholar]
- Biega, A.; Martin, T.E. Do amphibian conservation breeding programmes target species of immediate and future conservation concern? Oryx 2018, 52, 723–729. [Google Scholar] [CrossRef]
- Biega, A. , Greenberg, D.A., Mooers, A.O., Jones, O.R.; Martin, T.E. Global representation of threatened amphibians ex situ is bolstered by non-traditional institutions, but gaps remain. Animal Conservation 2017, 20, 113–119. [Google Scholar] [CrossRef]
- Ziegler, T.; Kamphausen, J.; Glaw, F.; Crottini, A.; Garcia, G.; Rödder, D.; Rauhaus, A.; Stenger, L.; Wahle, A. Threatened Malagasy amphibians and reptiles in zoos–a call for enhanced implementation of the IUCN’s One Plan Approach. Der Zoologische Garten 2022, 90, 21–69. [Google Scholar]
- Jenkins, R.K.; Tognelli, M.F.; Bowles, P.; Cox, N.; Brown, J.L.; Chan, L.; Andreone, F.; Andriamazava, A.; Andriantsimanarilafy, R.R.; Anjeriniaina, M.; Bora, P. Extinction risks and the conservation of Madagascar's reptiles. PLoS ONE, 1001; 9. [Google Scholar]
- Van Wilgen, N.J.; Wilson, J.R.U.; Elith, J.; Wintle, B.A.; Richardson, D.M. Alien invaders and reptile traders: What drives the live animal trade in South Africa? Animal Conservation 2010, 13, 24–32. [Google Scholar] [CrossRef]
- Marešová, J.; Frynta, D. Noah's Ark is full of common species attractive to humans: The case of bold snakes in zoos. Ecological Economics 2008, 64, 554–558. [Google Scholar] [CrossRef]
- Brereton, J.; Brereton, S. Short Communication: Examining taxa representation in Asian zoos and aquaria using historic records. Biodiversitas Journal of Biological Diversity 2021, 22, 2870–2875. [Google Scholar] [CrossRef]
- Carrizo, S.F. , Smith, K.G.; Darwall, W.R.T. Progress towards a global assessment of the status of freshwater fishes (Pisces) for the IUCN Red List: Application to conservation programmes in zoos and aquariums. International Zoo Yearbook 2013, 47, 46–64. [Google Scholar] [CrossRef]
- Valdez, J.W.; Mandrekar, K. Assessing the species in the CARES preservation program and the role of aquarium hobbyists in freshwater fish conservation. Fishes 2019, 4, 49. [Google Scholar] [CrossRef]
- Raghavan, R.; Dahanukar, N.; Tlusty, M.F.; Rhyne, A.L.; Kumar, K.K.; Molur, S.; Rosser, A.M. . Uncovering an obscure trade: Threatened freshwater fishes and the aquarium pet markets. Biological Conservation 2013, 164, 158–169. [Google Scholar] [CrossRef]
- Leiss, L. , Rauhaus, A., Rakotoarison, A., Fusari, C., Vences, M.; Ziegler, T. Review of threatened Malagasy freshwater fishes in zoos and aquaria: The necessity of an ex situ conservation network—A call for action. Zoo Biology 2022, 41, 244–262. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.E.; Brereton, J.E.; Rowden, L.J.; de Figueiredo, R.L.; Riley, L.M. What’s new from the zoo? An analysis of ten years of zoo-themed research output. Palgrave Communications 2019, 5. [Google Scholar] [CrossRef]
- Hughes, D.G.; Bennett, P.M. Captive breeding and the conservation of invertebrates. International zoo yearbook 1991, 30, 45–51. [Google Scholar] [CrossRef]
- Pearce-Kelly, P. Invertebrate propagation and re-establishment programmes: The conservation and education potential for zoos and related institutions. Creative Conservation: Interactive management of wild and captive animals.
- Pearce-Kelly, P.; Jones, R.; Clarke, D.; Walker, C.; Atkin, P.; Cunningham, A.A. , 1998. The captive rearing of threatened Orthoptera: A comparison of the conservation potential and practical considerations of two species' breeding programmes at the Zoological Society of London. Journal of Insect Conservation 1998, 2, 201–210. [Google Scholar] [CrossRef]
- Saul-Gershenz, L. Insect Zoos. In Encyclopedia of insects; Academic Press: Cambridge, MA, USA, 2009; pp. 516–523. [Google Scholar]
- Boppré, M.; Vane-Wright, R.I. The butterfly house industry: Conservation risks and education opportunities. Conservation and Society 2012, 10, 285–303. [Google Scholar] [CrossRef]
- Hobhouse, H. Seeds of change: Six plants that transformed mankind; Papermac, an imprint of Macmillan Publishers Ltd.: New York, USA, 1999; ISBN 9780333736289. [Google Scholar]
- Fara, P. Sex, botany and empire: The story of Carl Linnaeus and Joseph Banks; Icon Books: London, UK, 2004; ISBN 9780231134262. [Google Scholar]
- Westwood, M.; Cavender, N.; Meyer, A.; Smith, P. , 2021. Botanic garden solutions to the plant extinction crisis. Plants, People, Planet.
- Oldfield, S.F. Botanic gardens and the conservation of tree species. Trends in Plant Science 2009, 14, 581–583. [Google Scholar] [CrossRef]
- Donaldson, J.S. Botanic gardens science for conservation and global change. Trends in Plant Science 2009, 14, 608–613. [Google Scholar] [CrossRef]
- Miller, A.J.; Novy, A.; Glover, J.; Kellogg, E.A.; Maul, J.E.; Raven, P.; Jackson, P.W. , 2015. Expanding the role of botanical gardens in the future of food. Nature Plants 2015, 1, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Moreau, T.; Kuehny, J.; Novy, A.; Greene, S.L.; Khoury, C.K. Resetting the table for people and plants: Botanic gardens and research organizations collaborate to address food and agricultural plant blindness. Plants, People, Planet 2019, 1, 157–163. [Google Scholar] [CrossRef]
- Cheek, D.; Procheş, S. The value of arboreta in South Africa. South African Journal of Science 2022, 118, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Sharrock, S. Botanic gardens and food security–the results of BGCI's survey. BGJournal 2013, 10, 3–7. [Google Scholar]
- Maunder, M. Botanic gardens: Future challenges and responsibilities. Biodiversity & Conservation 1994, 3, 97–103. [Google Scholar]
- Smith, P. Building a global system for the conservation of all plant diversity: A vision for botanic gardens and Botanic Gardens Conservation International. Sibbaldia: The International Journal of Botanic Garden Horticulture 2016, 14, 5–13. [Google Scholar] [CrossRef]
- Smith, P. The challenge for botanic garden science. Plants, People, Planet 2019, 1, 38–43. [Google Scholar] [CrossRef]
- Maunder, M.; Higgens, S.; Culham, A. The effectiveness of botanic garden collections in supporting plant conservation: A European case study. Biodiversity & Conservation 2001, 10, 383–401. [Google Scholar]
- Mounce, R.; Smith, P.; Brockington, S. Ex situ conservation of plant diversity in the world’s botanic gardens. Nature Plants 2017, 3, 795–802. [Google Scholar] [CrossRef]
- Volis, S. Complementarities of two existing intermediate conservation approaches. Plant Diversity 2017, 39, 379–382. [Google Scholar] [CrossRef]
- O'Donnell, K.; Sharrock, S. The contribution of botanic gardens to ex situ conservation through seed banking. Plant Diversity 2017, 39, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Sun, W. The role of botanical gardens in scientific research, conservation, and citizen science. Plant diversity 2018, 40, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Asase, A.; Mzumara-Gawa, T.I.; Owino, J.O.; Peterson, A.T.; Saupe, E. Replacing “parachute science” with “global science” in ecology and conservation biology. Conservation Science and Practice 2022, 4, e517. [Google Scholar] [CrossRef]
- Van der Werff, H.; Consiglio, T. Distribution and conservation significance of endemic species of flowering plants in Peru. Biodiv. Cons. 2004, 13, 1699–1713. [Google Scholar] [CrossRef]
- Mackay, R. The atlas of endangered species: Threatened plants and animals of the world; Earthscan: London, UK, 2002; ISBN 1853838748. [Google Scholar]
- Harding, J. Historic deforestation and the fate of endemic invertebrate species in streams. New Zealand J. Marine Freshwater Research 2003, 37. [Google Scholar] [CrossRef]
- Pearson, R. Climate change, biodiversity and extinction risk. Sterling, New York, USA, 2011.
- Hobohm, C.; Vanderplank, S.E. . Change: Risks and Predictability. Environmental Challenges and Solutions 2021, 181–193. [Google Scholar] [CrossRef]
- Fordham, D.A.; Akçakaya, H.R.; Araújo, M.B.; Keith, D.A.; Brook, B.W. Tools for integrating range change, extinction risk and climate change information into conservation management. Ecography 2001, 36, 1–9. [Google Scholar] [CrossRef]
- Gerlach, J. Climate change, species extinctions and ecosystem collaps. Phelsuma 2010, 17, 13–31. [Google Scholar]
- Crisp, M.D.; Laffan, S.; Linder, H.P.; Monro, A. Endemism in the Australian flora. J Biogeogr 2001, 28, 183–198. [Google Scholar] [CrossRef]
- Platts, P.J.; Gereau, R.E.; Burgess, N.D.; Marchant, R. Spatial heterogeneity of climate change in an Afromontane centre of endemism. Ecography 2013, 36, 518–530. [Google Scholar] [CrossRef]
- Li, T.; Luo, P.; Xiong, Q.; Yang, H.; Gu, X.; Qiu, Y.; Lin, B.; Liu, Y.; La, C. . Spatial heterogeneity of tree diversity response to climate warming in montane forests. Ecol. Evol. 2021, 11, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, J.M.; Molnar, J.L.; Jennings, M.; Revenga, C.; Spalding, M.D.; Boucher, T.M.; Robertson, J.C.; Heibel, T.J. The atlas of gobal conservation; University of California Press: Berkeley, 2010; ISBN 9780520262560. [Google Scholar]
- Hobohm, C. (Ed.) 2021. Perspectives for Biodiversity and Ecosystems. Environmental Challenges and Solutions 2021, 1–483. [Google Scholar]
- Hobohm, C.; Vanderplank, S.E. Change: Risks and Predictability. Environmental Challenges and Solutions 2021, 181–193. [Google Scholar]
- Poulin, R. Parasite biodiversity revisited: Frontiers and constraints. Int. J. Parasitol. 2014, 44, 581–9. [Google Scholar] [CrossRef] [PubMed]
- McCallum, H.I. Lose biodiversity, gain disease. Proc. Natl. Acad. Sci. USA. 2015, 112, 8523–8524. [Google Scholar] [CrossRef] [PubMed]
- Aerts, R.; Honnay, O.; Van Nieuwenhuyse, A. Biodiversity and human health: Mechanisms and evidence of the positive health effects of diversity in nature and green spaces. Br. Med. Bull. 2018, 127, 5–22. [Google Scholar] [CrossRef]
- Tribot, A.-S.; Deter, J.; Mouquet, N. Integrating the aesthetic value of landscapes and biological diversity. Proc. R. Soc. B. 2018, 285. [Google Scholar] [CrossRef]
- Giacinto, J.J.; Fricker, G.A. , Ritter, M.; Yost, J.; Doremus, J. Urban forest biodiversity and cardiovascular disease: Potential health benefits from California's street trees. PLoS ONE. 2021, 16, e0254973. [Google Scholar] [CrossRef]
- Chen, G.; Sun, W. The role of botanical gardens in scientific research, conservation, and citizen science. Plant diversity 2018, 40, 181–188. [Google Scholar] [CrossRef]
- Asase, A.; Mzumara Gawa, T.I.; Owino, J.O.; Peterson, A.T.; Saupe, E. Replacing "parachute science" with "global science" in ecology and conservation biology. Conservation Science and Practice.
Figure 1.
Endemic and endangered taxa between evolution/diversification and extinction risk due to anthropogenic and natural processes cf. [53,54,55,56,57].

Figure 2.
Global species richness of various groups of animals and plants (values rounded, blue and green colors indicate minima and maxima, for refs. see text).
Figure 2.
Global species richness of various groups of animals and plants (values rounded, blue and green colors indicate minima and maxima, for refs. see text).
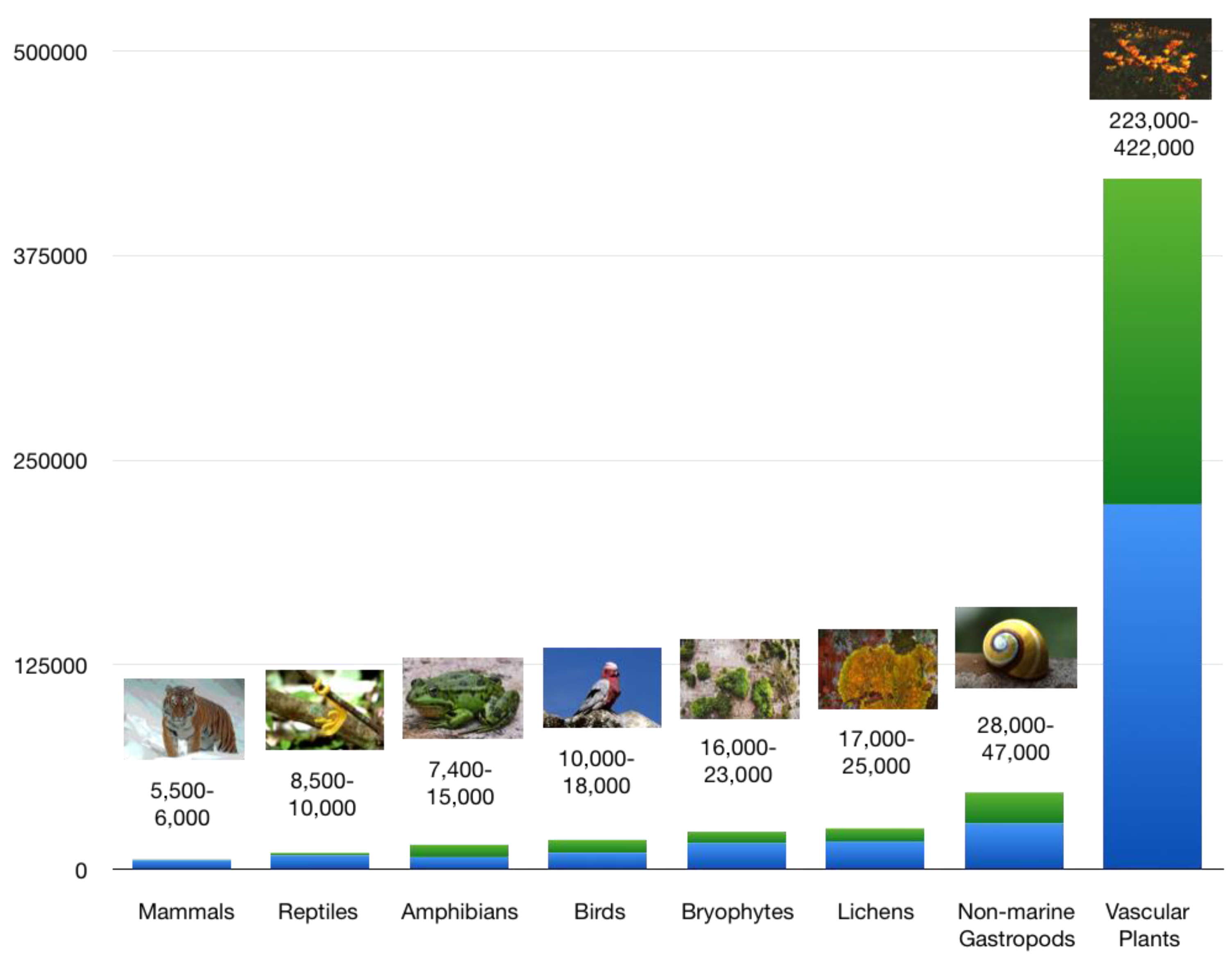
Figure 3.
Levels of endemism in species of non-marine gastropods, amphibians, vascular plants, reptiles, mammals, and birds in selected regions/countries (for refs. see text).
Figure 3.
Levels of endemism in species of non-marine gastropods, amphibians, vascular plants, reptiles, mammals, and birds in selected regions/countries (for refs. see text).

Figure 4.
Hypothetical order of levels of endemism in different groups of plants and animals at intermediate spatial scales (for refs. see text and Figure 3).
Figure 4.
Hypothetical order of levels of endemism in different groups of plants and animals at intermediate spatial scales (for refs. see text and Figure 3).


Table 1.
Putative world records of endemism in several taxonomic groups. World records of richness in vascular plant species per unit area, biomass and productivity are included for comparison.
Table 1.
Putative world records of endemism in several taxonomic groups. World records of richness in vascular plant species per unit area, biomass and productivity are included for comparison.
| Maxima | Mode of calculation | Country/region | Ecosystem (dominant) | Climate (dominant) | |
|---|---|---|---|---|---|
| Endemism in mammals, birds plus reptiles [127] | > 70 | numbers of endemics by terrestrial ecoregion | Eastern Madagascar | Rainforest | Wet tropical and subtropical |
| Endemism in birds [128] | 92 | E/S as percentage value | Hawaiian Islands | div. | Humid tropical and subtropical oceanic |
| Endemism in freshwater animals (vertebrates and invertebrates) [128] | 54 | E/S as percentage value | Lake Baikal, Russia | Freshwater lake | Temperate continental |
| Endemism in fish, freshwater turtles, crocodiles plus amphibians [127] | >150 | numbers of endemics by freshwater ecoregion | High Andes, western India, East African Rift Valley lakes | Wetlands and freshwater ecosystems | Tropical and subtropical |
| Endemism in cichlid fishes [128] | up to 99 | E/S as percentage value | Tectonic Lakes Tanganyika, Malawi, Victoria, Africa | Freshwater lake | Tropical |
| Endemism in land snails [129] | c. 100 | E/S as percentage value (rounded) | Hawaiian Islands | div. | Humid tropical and subtropical oceanic |
| Endemism in vascular plants [129,130] | > 80 | E/S as percentage value | New Caledonia, Hawaiian Islands, Madagascar, St. Helena, New Zealand | div. | Subtropical and tropical oceanic |
| Endemism in vascular plants [131,132] | 4.7-5.1 and 4-4.4 | Relative distance of residual to regression (Res. E) | Mas a Tierra, Chile, and St. Helena | Forest | Subtropical oceanic |
| Endemism in pteridophytes [133] | 37/31.7 | Percentage of endemism/index of insularity | Easter Island | Reeds and grasslands replace the original tropical forest | Tropical oceanic/subtropical humid |
| Species richness in vascular plants [134] | 942 | No. of species per 10,000 m2 | Ecuador | Lowland rainforest | Humid tropical |
| Species richness in vascular plants [135] | 115 | No. of species per 10 m2 | Romania | Steppe meadow (currently grazed) | Temperate |
| Biomass [136,137] | 1,819 or 2,844 | tC ha-1 (above-ground biomass) or tC ha-1 (total biomass) | SE Australia | Eucalyptus regnans forest | Warm temperate |
| Productivity [138,139] | 8.93-9.93 | kg m-2 year-1 (dry matter) | Amazon | Swamps dominated by C4 grass Echinochloa polystachya | Wet tropical |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



