Submitted:
13 October 2023
Posted:
16 October 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Acute respiratory viruses (ARVs) are leading cause of diseases in human worldwide. High risk individuals including children and elderly could potentially develop severe illness that could result in hospitalization or death in the worst case. Most common ARVs are Human respiratory syncytial virus, Human Metapneumovirus, Human Parainfluenza Virus, rhinovirus, coronaviruses (including SARS and MERS CoV), adenoviruses, Human Bocavirus, enterovirus (-D68 and 71), and influenza viruses. The olfactory deficits due to ARVs infection is a common symptom among patients. This mini review provides an overview of the role of SARS-CoV-2 and other common ARVs in the development of human olfactory pathophysiology. We highlight the critical need for understanding the signaling underlying the olfactory dysfunction and the development of therapeutics for this wide-ranging category of AVRs to restore the altered or loss of smell in affected patients.
Keywords:
Respiratory viruses
; Anosmia
; Olfaction Disorders
; loss of smell
; Covid-19
1. Introduction
Respiratory viral infections are very common and constitute a huge health concern around the world, with new infectious diseases continuing to emerge [1,2]. Such pathologies could lead to death particularly in elderly and increase the expenses of the health care system worldwide. Respiratory viruses have the propensity to infect and trigger diseases through the human lower and upper respiratory tracts. We will be more interested on viral upper respiratory infections (URI) as they are considered to be one of the most common causes of olfactory dysfunction, accounting for up to 45% of all cases [3,4]. Although the alteration of smell following viral URI is noticed in several cases, a little treatment is currently available. The occurrence of this alteration is termed post-viral olfactory dysfunction (PVOD) [5,6,7]. The most common viruses implicated in PVOD include parainfluenza virus (PIV) [8], rhinoviruses (RV), respiratory syncytial virus (RSV) and coronaviruses (CoV) [4,9,10,11,12]. In 1956, RSV was found and isolated from chimpanzees, and from infants suffering severe lower respiratory tract illness a year later. In older children and healthy adults, RSV causing repeated URI are common and can lead to symptomatic UR tract diseases [13]. According to Heikkinen and colleagues, 5% to 10% of URI are attributed to RSV. Studies performed in mice have shown that RSV infections are associated with the damage to olfactory receptor neurons [14]. Despite these findings, the occurrence of olfactory loss associated with RSV infections seems low and needs further investigations in more broad regions of the world and during cold and warm periods to establish clearer picture of this virus-induced olfactory dysfunction [12,15,16]. Like RSV, the PIV was discovered in 1950s and is associated with lower respiratory tract infections as well as URI in children. Young children being infected by this virus are often diagnosed with respiratory irritants, vitamin A deficiency or malnutrition [17,18]. Although PIV is found in respiratory secretions, the major diagnosis is observed from pulmonary secretions and confirmed by chest x-ray [19]. The RV that targets humans, are considered among the most infectious agents worldwide and are associated with mild upper respiratory tract infections in people [20,21,22]. The CoV were identified in the 1960s. Little attention was given to this family of viruses until both outbreaks of the severe acute respiratory syndrome (SARS)-CoV and the Middle East respiratory syndrome (MERS)-CoV that were identified respectively in 2003 and 2012 respectively [23,24,25]. The novel coronavirus disease 2019 (COVID-19) caused by SARS-CoV-2, has spread fast all over the world [1,2]. Both MERS-CoV and SARS-CoV-2 are highly pathogenic coronaviruses and have a huge spatial range of epidemics areas globally, but regarding MERS-CoV, the vast majority of cases are confined to the Middle East [25,26]. A key factor in the transmissibility of these viruses, is the active virus replication in upper respiratory tract (URT) tissues and therefore its massive excretion [27,28]. Since the beginning of the pandemic, growing reports have shown the issues of partial to complete loss of smell in patients contracted COVID-19 [29,30,31,32] and have brought new focus to PVOD. This review mainly focuses on understanding the molecular signaling underlying the olfactory pathophysiology in human patients from COVID-19 infection and common viruses induced URTI. The importance of using such mechanisms to find potential targets to overcome the loss of smell will also be discussed.
2. Sources and selection criteria
The present study intends to synthesize the current knowledge regarding the relationship of the respiratory viral pathogenesis of the olfactory system and the mechanisms underlying the loss of smell in patients infected by respiratory viruses including COVID-19. We have performed a literature search using mostly the database PubMed Central (PMC), Google scholar and ScienceDirect to parse original articles, meta-analyses and systematic reviews that have tackled the animals and human respiratory viruses having a negative impact on the olfactory system functionality. For our search, we used the combination of the following keywords: respiratory virus, coronavirus, rhinovirus, parainfluenza viruses and respiratory syncytial virus (RSV), anosmia, parosmia, hyposmia, olfactory epithelium, human, mouse, hamster, loss of smell or olfactory dysfunction were considered in this review. Only the papers that have met the keyword criteria listed above were considered in this work.
3. Olfactory receptor and odorant detection
In most animals, the functional olfactory system detects and discriminates among diverse chemical stimuli. Odours are important for behaviours such as eating, mating, and avoiding dangerous smells, such as smoke, leaking propane gas, and spoiled food [33,34,35]. The importance of the behaviours leads to a strong belief that the loss of olfactory function is indirectly life-threatening [36,37]. Two different olfactory systems have been developed in mammals such as rodents: the main olfactory epithelium (MOE or OE), also called olfactory mucosa, connected to the main olfactory bulb, and the accessory system called vomeronasal organ (VNO) connected to the accessory olfactory bulb [38,39,40,41,42]. Here, the VNO will not be discussed. The configuration of the olfactory epithelium (OE) presents unique cytological characteristics as it contains different cell types such as the ciliated olfactory receptor neurons (ORNs), the sustentacular supporting cells, and the cells of Bowman’s glands. The olfactory mucosa hosts many different types of cells including ORNs in the intermediate layer, the sustentacular cells on the apical and basal sides and the sensory cilia present at the apical pole where the dendrites of olfactory neurons are extended [43,44].
A deep understanding of the molecular signaling of the smelling recognition is required to understand the basis of the olfactory system and, consequently, the loss of olfactory function. Starting from the beginning of 1990s, pioneers have developed and studied the physiology of the olfactory system based on molecular biology, biochemistry, anatomy, and bioinformatics [33,45]. At first glance, getting insight into the molecular mechanisms of the perception of odours has emerged from several disciplines such as chemistry, biology and professional odour detectors [33,45].
The detection occurs when the odorants penetrate into the nasal cavity and reach the olfactory mucosa. The odorants then interact with specific ORN in the olfactory mucosa. Once an ORN is activated by an odorant, a nervous influx is sent to the cortex via the olfactory bulb. Readers interested in the mammalian olfactory epithelium and the perception of odour coding are invited to view an excellent review by Kurian and colleagues published in 2020 [46].
4. Viral infection causing olfactory dysfunction
The fact that the olfactory receptor neurons (ORNs) are found in the nasal cavity and expressed in the olfactory epithelium (OE) makes them directly exposed to all kinds of air-bound and air-way pathogens that make the ORNs vulnerable. Whether the cause is physiological or pathological, the lifespan of ORNs is relatively short of few weeks in the OE. Moreover, the stem cell reprogramming ensures the continuous regeneration of new ORNs from OE basal cells either in a physiological turnover of ORNs or in response to inflammation and OE severe damage mediated by neural injury [47,48,49]. Several airway pathogens, such as viruses, are causing damage to OE through particularly the sustentacular cells, triggering anosmia, hyposmia, phantosmia or parmosmia in mammals [44,50,51,52,53]. Many respiratory tract infections due to viruses like rhinovirus (RV), parainfluenza viruses (PIV) [8], respiratory syncytial virus (RSV), coronaviruses (CoV) and Epstein-Barr viruses (EBV) [4,11,12,54,55], have been involved in the development of olfactory disorders such as partial or total loss of smell. Doty and others have termed this pathology as virus-induced olfactory dysfunction as post-viral olfactory disorder (PVOD) [5,6,56,57]. Viral infection destroys many cells within the apical layer of the OE that could lead to ORNs functional impairment in the nose. Interestingly, the OE basal cells can constantly replace damaged ORNs to new olfactory neurons allowing patients to recover functional olfactory response [48,49]. In the following sections, the common viral infection of the upper respiratory tract (URT) leading to olfactory dysfunction like anosmia, hyposmia, phantosmia and parmosmia [4] in animal models and humans will be discussed.
5. Viruses impacting respiratory system
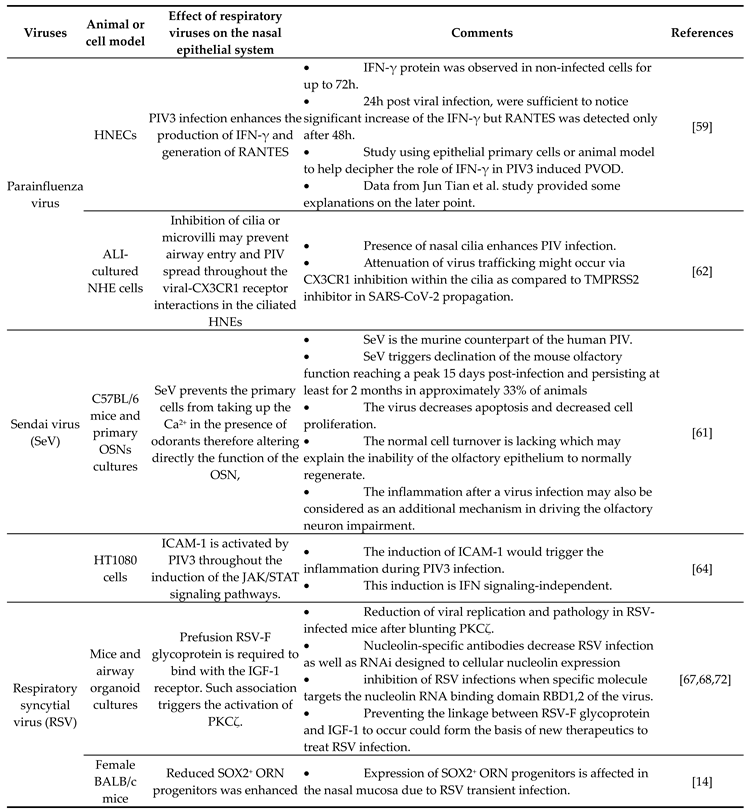
The respiratory system is exposed to the environment and is in permanent contact with air-way pathogens like viruses. A recent investigation in humans, has identified 18 viruses in patients with PVOD. Several known viruses are associated with olfactory impairment and it is crucial to investigate the mechanisms of infection as well as the specific receptors each virus targets (Table 1).
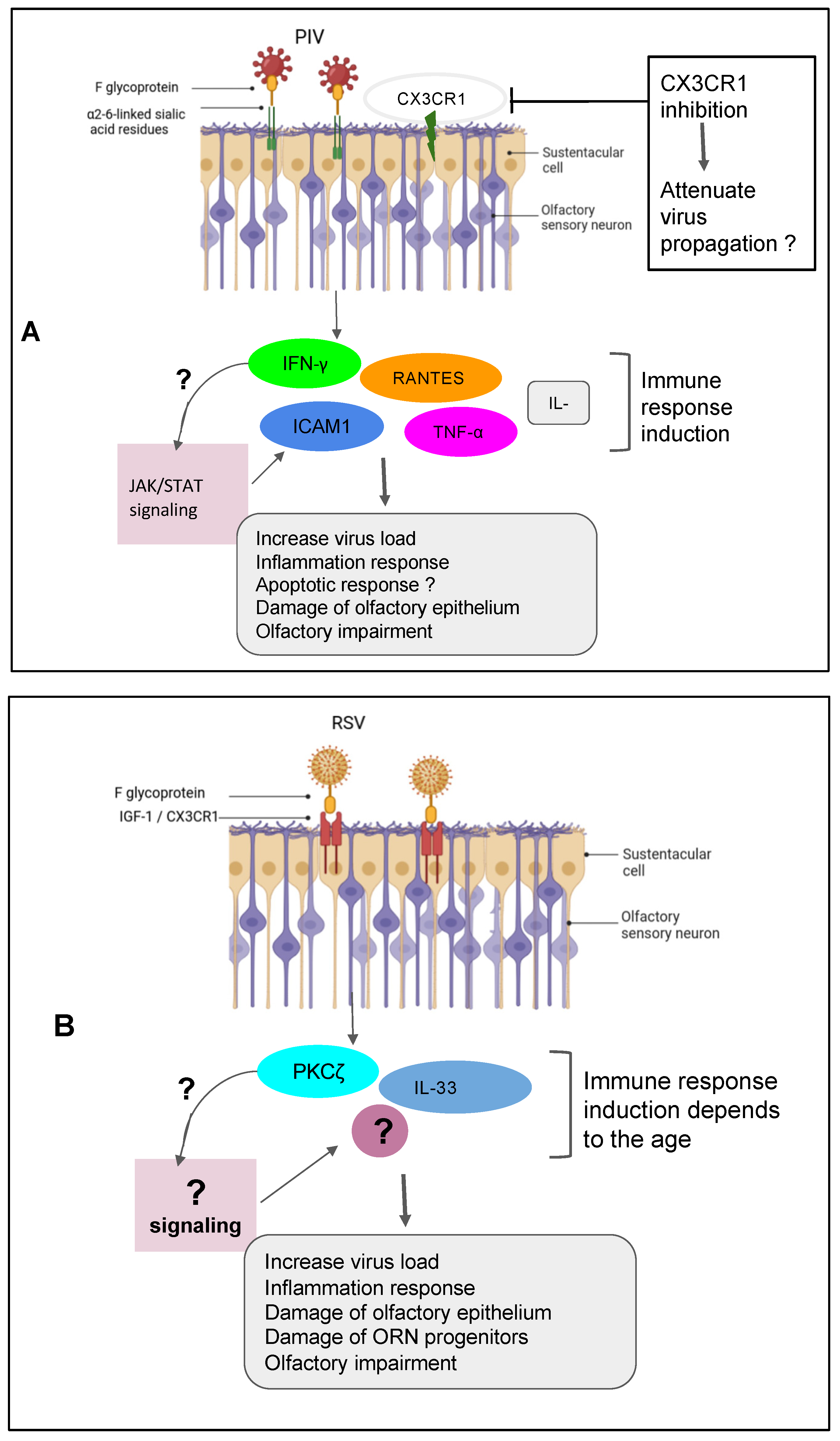
The presence of parainfluenza type 3 (PIV3) was observed in human nasal epithelial cells (HNECs) from 88% of patients with PVOD as compared to 9% of control patients [58]. This data suggests potential involvement of PIV3 infection into the upper airway pathology. PIV3 in the turbinate epithelial cells of PVOD is responsible for respectively 60% of hyposmia and 40% of anosmia in patients [58]. PIV3 has been shown to infect the HNECs and therefore exacerbates the production of IFN-γ and pro-inflammatory cytokines [59]. This data is in line with a previous study suggesting that PIV3 may cause olfactory dysfunction through mechanisms other than nasal obstruction in patients [52]. The Sendai virus (SeV), the murine counterpart of the human PIV, has been shown to directly infect the mouse brain via the olfactory neurons [60]. Another investigation demonstrated that SeV infection led to impairing mouse olfaction. Interestingly, the virus persists in OE and OB tissues for over two months, and reduces the regenerative power and the functionality of the ORNs [61]. A very recent findings have shown that the depletion of nasal cilia via the silencing of CEP83, a protein critical for motile cilia formation in all ciliated cells would impede the PIV infection. Indeed, the PIV receptor CX3CR1 colocalizes to motile cilia is a plausible viral entry mechanism into the cells [62]. An additional study showed that the intercellular adhesion molecule-1 (ICAM-1) and related cytokine molecules is induced by PIV3 and it is thought that this activation participates in the inflammation during infection by viruses [63,64] (Figure 1A). Further research using the powerful of the transcriptomic analysis is necessary to help to delineate the role of PIV and implicated mechanisms in the development of the broad range of olfactory dysfunctions in patients and to find host-response transcript signature for possible treatments.
Recent findings have demonstrated that respiratory syncytial virus RSV can infect OSN in the nasal cavity of the mouse before accessing to the central nervous system of the animal [65]. As PIV, the RSV targets the cilia of the epithelial cells in the airways by fusionning its F-glycoprotein to the cellular receptor human nucleolin (NCL). RSV uses also another mechanism that activates protein kinases like IGF1R to get into the cells [63,66,67]. Furthermore, recent studies have shown the essential role of the nucleolin RNA binding domain RBD1,2 in allowing the infection of SRV [68]. Previous works have pointed out the importance of ORN progenitors in the turnover. They showed that SRV infection is causing of SOX2+ ORN progenitors damage prior to the manifestation of ORN impairment. Unfortunately, the airway allergy seems to amplify this damage induced by the RSV infection leading to a possible loss of OMP+ ORNs [14]. Transcriptomic analysis demonstrated that olfactory signaling is among the altered pathways in patients suffering from RSV infection. This finding further supports previous work that described RSV as a causative agent of post-viral olfactory dysfunction. Interestingly, the authors highlighted that this molecular signaling could be a promising future route to investigate drug targets against RSV infection [36,56]. Sourimant et al., have recently shown that 4′-fluorouridine, a ribonucleoside analog inhibits RSV in a selective manner in cells and human airway epithelia organoids. As this orally therapeutic drug is efficient in small animal models, further investigations are needed to apply this molecule in human with the same yield [69] (Figure 1B). Efforts are underway in the phase of clinical trials when mRNA vaccines are combined to antigens to fight multiple respiratory viruses including SRV [70,71].

Table 1.
Summary of virus-implication in PVOD.
|
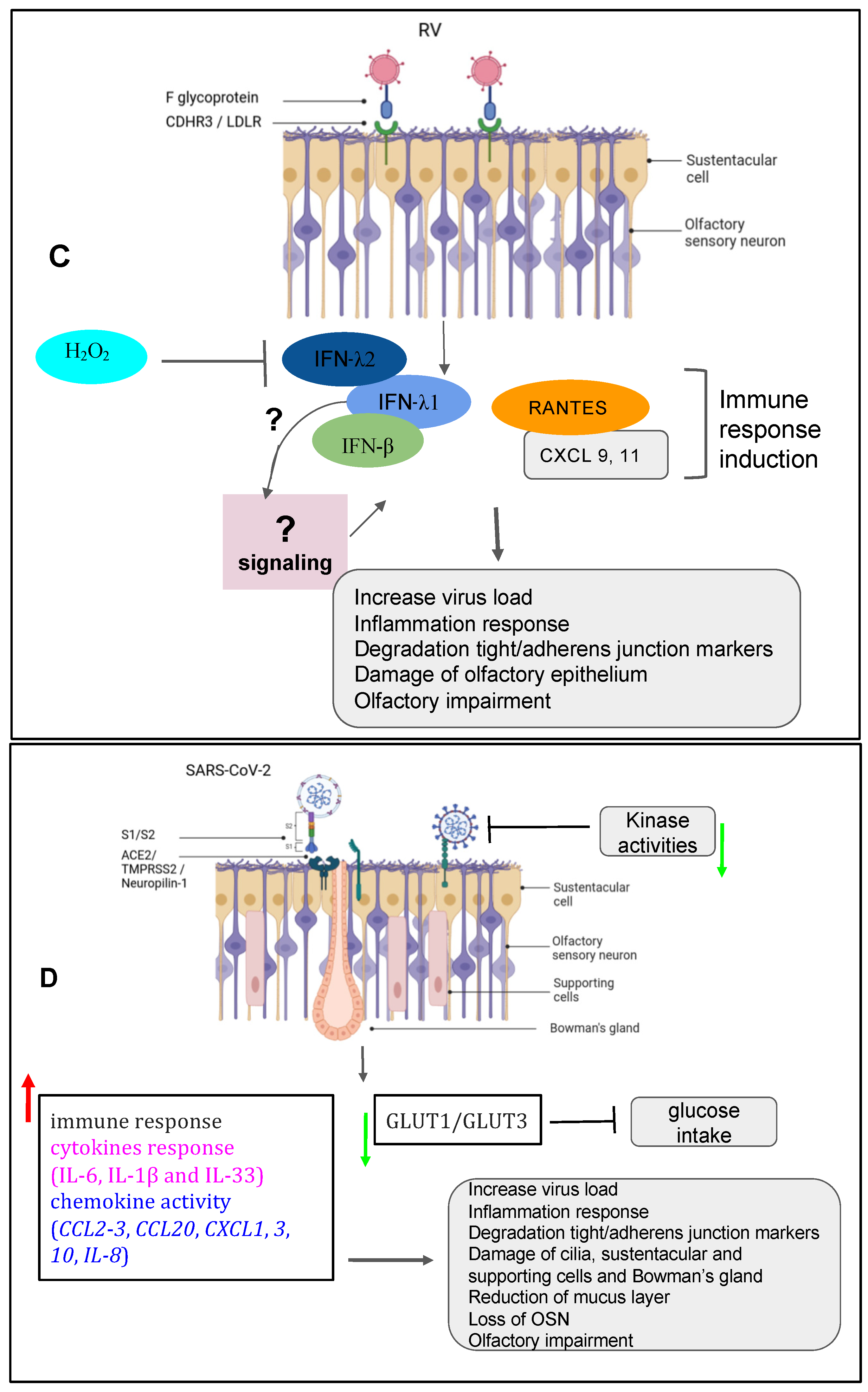
Recently, rhinoviruses (RV) were shown among the more predominant causative agents of PVOD in patients. The study showed that patients with anosmia was higher than in those with hyposmia (58.8% vs. 19.0%, p= 0.018) [54]. In line with these findings, it is tempting to suggest that the persistence of the virus could be one factor that governs a more severe injury to the olfactory system. In fact, RV primary invade the ciliated respiratory epithelial cells via the glycoprotein members such as the intercellular adhesion molecule 1 (ICAM-1), the low-density lipoprotein receptor (LDLR) family members, and the cadherin-related family member 3 (CDHR3) [73,74,76,92]. RV infection induces Toll-like receptor 7 (TLR7) and retinoic acid-inducible gene I (RIG-1) that trigger the activation of cytokine expression (type I and type III IFNs) [93,94,95]. Understanding the mechanisms of RV viral induced asthma for new therapeutic directions have gained more attention in the recent past [96,97,98]. Papi et al., have investigated the role of reducing agents such DMSO on RV infection in the nasal epithelium. They showed that rhinovirus-induced ICAM-1 mRNA expression was inhibited by reducing agents in a dose-dependent manner. Interestingly, NF-κB and TNF-α activation which is necessary for the ICAM-1 promoter, was completely abolished in those treated epithelial cells [99]. Moreover, it has been shown that CDHR3 genetic variants impact on severity of RV-related pediatric respiratory tract infections by upregulating the epithelial expression of RV receptors thus helping clinicians to predict the susceptibility and severity of RV infection [100]. Complementary recent studies have demonstrated that both vitamin D and hydrogen peroxide play a critical role in attenuating the RV mediating the ICAM-1 activation and the production of type I (IFN-β) and type III (IFN- λ1 and λ2) interferons respectively [77,101] (Figure 1C). The identified mechanisms focusing on oxidant biomarkers could be a plausible road for reducing the clinical severity or treating RV infection induced-olfactory dysfunction in patients.
6. Mechanisms of SARS-CoV-2 mediating the loss of smell
The post-COVID and the long-term-COVID have both tremendously triggered a lot of complications in different human systems. The loss or reduction of smell, among other complications of the nervous system, is an associated symptom for patients affected by different variants of COVID-19 including omicron variant [102,103,104,105,106]. Moreover, studies reported that the prevalence of olfactory dysfunction differs greatly between populations and approaches [106,107,108]. Currently, many COVID-19 vaccines are authorized to help protect and eliminate the virus. The COVID-19 pathology and the cellular mechanism by which the olfactory dysfunction occurs, have gained a lot of attention since the pandemic and researchers are still investigating underlying signaling and complications (Table 1) [62,90,91,106,109,110,111,112]. Earlier in the pandemic, reports hypothesized that five potential mechanisms were considered to get insights into the olfactory dysfunction in COVID-19 patients : (1) obstruction/congestion and rhinorrhea of the nasal airway, (2) damage and loss of ORNs, (3) Olfactory center damage in the brain, (4) damage of the olfactory supporting cells in the OE, and (5) Inflammation-related olfactory epithelium dysfunction [113,114]. Butowt et al., have recently reviewed that at least the following hypotheses (1)-(3) turned out to be implausible, for explaining the olfactory dysfunction in patients [115]. This allegation is further confirmed by very recent studies showing that SARS-CoV-2 infection significantly increased the expression of interferon-stimulated and inflammatory genes. Alteration of extracellular matrix genes were also observed in ALI and iALI infected cells [91]. Here, we will particularly review the mechanisms related to the second, the fourth and the fifth scenarios according the available findings. Healthy sensory cilia of ORNs in the olfactory epithelium are crucial in perceiving odorant molecules before sending the information to the olfactory bulbs and then to the upper parts of the brain [47]. It has been reported in humans that the SARS-CoV-2 may indirectly affects the olfactory cilia, hindering the smelling system’s efficacy [116]. Reports suggested that ORNs lack to express the entry proteins of SARS-CoV-2 in the OE. The virus seems to establish a first contact in human nasal epithelia by binding its spike S protein to specific cells in the OE [117]. These reports are confirmed by study based on in-silico data, predicting the that mature ORNs do not express the virus entry protein, the angiotensin-converting enzyme 2 (ACE2), and therefore are not likely to be infected by SARS-CoV-2 [118]. Furthermore, supporting data by Bryche et al., showed that SARS-CoV-2 was not detected in the ORNs of golden Siryan hamsters [119]. However, in few cases, authors suggested that SARS-CoV-2 could infect ORNs in hamsters [120]. Based on the fact that COVID-19-related loss of smell disappeared within 1-2 weeks, while the regeneration of dead ORNs needs more than 2 weeks, many data tend to conclude that COVID-19-related olfactory dysfunction (OD) is not directly associated with the impairment of the ORNs [113,114,117,121,122]. Consequently, studying the entry proteins expression within the cells in the OE will help to understand the sensitivity of the OE to SARS-CoV-2 infection-related to the high prevalence of ODs in patients. Many groups are now interested to the organization of the sustentacular cells in the OE and thought that they might play central role in leading to OD. A high level of expression of ACE2 and the transmembrane serine protease 2 (TMPRSS2) is particularly found on the sustentacular cells suggesting a path to the neurotropism of SARS-CoV-2 in the OE. The ACE2 and TMPRSS2 are respectively known as the SARS-CoV-2 receptor and the SARS-CoV-2 cell entry-priming protease. ACE2 is found mainly on different parts of the sustentacular cells both in human and mouse. The ACE2 and TMPRSS2 genes tend to be co-regulated [114,123,124,125,126,127]. Different approaches using tissues, cells and organ systems in human, golden Syrian hamster, and hACE2 transgenic mouse have been employed to study the pathological impact of the SARS-CoV-2. Here, we discussed findings related particularly to the OE in inducing ODs in human. The spike protein (S protein) of SARS-CoV-2 mediates the passage of the virus into the host cell by fusing the viral and host cell membranes. In fact, via his spike S, SARS-CoV-2 employs the ACE2 as host functional receptor and TMPRSS2 as the cellular priming protease facilitating viral uptake, both signaling being confirmed by Single-cell RNA sequencing (scRNA-seq) datasets from the Human Cell Atlas consortium [128,129,130]. Another study showed that SARS-CoV-2 Nucleocapsid protein (NP), was observed in human OE through the neuronal marker Tuj1, 9 hours post infection. This data further supported the enrichment of ACE2 in human olfactory sustentacular cells [124,131]. Earlier in the pandemic, the golden Syrian hamster was used as a model to document the pathology of SARS-CoV-2 in the OE post infection. Reports showed that the sustentacular cells are rapidly infected by SARS-CoV-2. This viral neurtropism is associated with a massive recruitment of immune cells in the OE and lamina propria, which could drive the disorganization of the OE structure [119]. This study is consistent with high level of Tumor Necrosis Factor α (TNF α) observed in OE samples from COVID-19 suffering patients and in ALI and iALI infected cells [78,91]. Furthermore, the inflammation induced by SARS-CoV-2 infected supporting cells may play an important role in the onset and persistence of loss of smell in patients. This SARS-CoV-2-associated inflammation status was confirmed the transcriptome of the in vitro human airway epithelium and by analyzing the expression of selected targets in the olfactory bulb using RNA-seq and RT-qPCR tools. Interestingly, this study showed that the proinflammatory markers including NFKBIA, CSF1, FOSL1, Cxcl10, Il-1β, Ccl5 and Irf7 overexpression continued up to 14 dpi, when animals had recovered from ageusia/anosmia [91,132]. These findings are in line with a very recent study showing the implication of immune cell infiltration and altered gene expression in OE in driving persistent smell loss in a subset of patients with SARS-CoV-2. Moreover, this study particularly, demonstrates that T cell–mediated inflammation lasts longer in the OE after the acute SARS-CoV-2 infection has been eliminated from the tissue, suggesting a mechanistic insights into the long-term post–COVID-19 smell loss [105]. The OE disorganization is followed by a drastic deterioration of the cilia layer of the ORNs that leads to the impairment of the olfactory capacity of the animal [119]. Investigations in humans and hamsters using respectively, Transmission Electron Microscopy (TEM) studies and Scanning Electron Microscopy (SEM) analysis showed various levels of cilia height that undergo regeneration in the course of patient recovery, including smell restoration. Data using the golden Syrian hamster showed that the regenerated cilia in the epithelium is accompanied by a decreased expression of FOXJ1+ highlighting the importance of this marker in the respiratory ciliogenesis. This later finding by Schreiner et al., could in part shed light on the inquiry of how could we regenerate cilia during patient recovery, although a lot needs to be done in the roadmap of treating loss of smell related to nasal cilia deterioration by SARS-CoV-2 [62,104,133,134] (Figure 1D; Table 1).
According to the literature, different variants of SARS-CoV-2 do not directly target particularly the ORNs in the OE, instead they are found in majority expressed in the sustentacular cells [135]. Recent study by Seehusen et al., showed that K18-hACE2 transgenic mouse expressing the human ACE2 is highly sensitive to at least five variants of SARS-CoV-2 that infected not only the supportive cells in OE and the respiratory epithelium but invaded the CNS of the animal five days post infection. Interestingly, the expression of hACE2 seems to convey higher binding affinity when compared to the wild-type mouse [136]. Using this transgenic mouse reveals to be a serious option for therapy development against loss of smell as these animals exhibited low mortality when treated with COVID-19 convalescent antisera [79,136]. For instance, the miRNA is shown to play a crucial role in the regulation of immune genes deregulated and the development of miRNA antagonists or mimics seems to be a promising new therapeutic strategies for the treatment of patients with COVID-19 on other respiratory viruses-induced PVOD [70,71,91,137,138,139,140].
It is now accepted that ACE2 is not the only obligate entry for SARS-CoV-2 as it has been suggested that molecules including PIKfyve or neuropilin-1 (NRP-1) may participate in SARS-CoV-2 entry [81,82,141]. Like ACE2, NRP-1 is highly expressed in the respiratory and olfactory epithelium which further support the infectivity and entry of SARS-CoV-2 in the human OE. NRP-1 is not only found in supportive cells but is expressed in nearly every cell type in the nasal passages including the ORN, therefore giving SARS-CoV-2 a route to access those cells and impair the olfactory response. Interestingly, Daly et al., demonstrated that the selective inhibition of the S1-NRP-1 interaction reduces SARS-CoV-2 infection [85,86,141].
Taken together, the highly expression of ACE2, TMPRSS2 and NRP-1 in supportive and other olfactory cells and their impact in olfactory neurophysiology maintenance and in the development of human olfactory pathophysiology supports them as potential targets for signaling-based therapeutics of olfactory dysfunction.
6. Conclusion and perspective
Our literature review further confirms the previous extended investigations showing that loss of smell and taste are among the key associated symptoms with most COVID-19 variants, including the omicron variant which causes runny nose, headache, fatigue, sneezing, and sore throat [142]. The last three years have been an important rush towards deciphering the underlying mechanisms the SARS-CoV-2 deploys to impair the olfaction in infected patients. Furthermore, it is interesting to delineate the similarities and the differences of the molecular mechanisms of SARS-CoV-2 and the other respiratory viruses induced olfactory dysfunction. Both SARS-CoV-2 and non- SARS-CoV-2 attach to the cilia during initial stages of infection to later enter to the nasal epithelium [62]. Interestingly, all these findings underly the importance of the immune mediated inflammatory injury to the olfactory neuroepithelium that is now accepted as a consequence of those URT virus infection. However, the molecular signature-related to the type of olfactory dysfunction according to the Table 1 seems particular for each respiratory virus. The animal or cellular models being used and the seasonal periods could add more complexity for the putative mechanisms for viral infection induced olfactory dysfunction. Investigative literature on the COVID-19 mechanistic route have made clear that this virus seems to attach to ACE2-TMPRSS2 complex and/or NRP-1 on the host cell prior to infection and later triggers intrinsic immune responses. In the nasal mucosal microenvironment, those markers play a crucial role in inflammatory response mechanisms and are confirmed by several recent studies on understanding SARS-CoV-2 invasion [79,81,82,85,136,141,143]. Currently, the mechanism on how SARS-CoV-2 causes smell loss is widely documented and more investigations are needed on the non-SARS-CoV-2 to complete the picture on comparing the particularity on each respiratory virus causing URT related PVOD. For instance, this work points the urge and necessity of finding an adequate therapeutic solution against the COVID-19 and other respiratory viral pathogens induced olfactory dysfunction. In addition, the mechanisms of taste dysfunction due to COVID-19 infection is not discussed in this review. But it would be interesting to decipher the possible pathogenesis between ageusia and anosmia and other types of PVOD in COVID-19 in other non-COVID-19 patients in the future.
Author Contributions
Writing—original draft preparation, S.F.W. and A.A.M.D.; Writing—review and editing, S.F.W., A.A.M.D., B.N., F.T., and N.D.; Conceptualization and supervision, S.F.W. and A.A.M.D. All authors contributed to the literature review, writing the article, revising the manuscript, have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wu, J.T.; Leung, K.; Leung, G.M. Nowcasting and forecasting the potential domestic and international spread of the 2019-nCoV outbreak originating in Wuhan, China: a modelling study. Lancet 2020, 395, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.S.; E, I.A.; Madani, T.A.; Ntoumi, F.; Kock, R.; Dar, O.; Ippolito, G.; McHugh, T.D.; Memish, Z.A.; Drosten, C.; et al. The continuing 2019-nCoV epidemic threat of novel coronaviruses to global health - The latest 2019 novel coronavirus outbreak in Wuhan, China. Int J Infect Dis 2020, 91, 264–266. [Google Scholar] [CrossRef]
- Seiden, A.M. Postviral olfactory loss. Otolaryngol Clin North Am 2004, 37, 1159–1166. [Google Scholar] [CrossRef]
- Zhen Yu, L.; Luigi Angelo, V.; Paolo, B.-R.; Abigail, W.; Claire, H. Post-viral olfactory loss and parosmia. BMJ Medicine 2023, 2, e000382. [Google Scholar] [CrossRef]
- Moran, D.T.; Jafek, B.W.; Eller, P.M.; Rowley, J.C. , 3rd. Ultrastructural histopathology of human olfactory dysfunction. Microsc Res Tech 1992, 23, 103–110. [Google Scholar] [CrossRef]
- Welge-Lussen, A.; Wolfensberger, M. Olfactory disorders following upper respiratory tract infections. Adv Otorhinolaryngol 2006, 63, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Dicpinigaitis, P.V. Post-viral Anosmia (Loss of Sensation of Smell) Did Not Begin with COVID-19! Lung 2021, 199, 237–238. [Google Scholar] [CrossRef]
- Khachatryan, V.; Sirunyan, A.M.; Tumasyan, A.; Adam, W.; Bergauer, T.; Dragicevic, M.; Ero, J.; Friedl, M.; Fruhwirth, R.; Ghete, V.M.; et al. Distributions of topological observables in inclusive three- and four-jet events in pp collisions at [Formula: see text][Formula: see text]. Eur Phys J C Part Fields 2015, 75, 302. [Google Scholar] [CrossRef]
- Tian, J.; Pinto, J.M.; Li, L.; Zhang, S.; Sun, Z.; Wei, Y. Identification of Viruses in Patients With Postviral Olfactory Dysfunction by Multiplex Reverse-Transcription Polymerase Chain Reaction. Laryngoscope 2021, 131, 158–164. [Google Scholar] [CrossRef]
- Suzuki, M.; Saito, K.; Min, W.P.; Vladau, C.; Toida, K.; Itoh, H.; Murakami, S. Identification of viruses in patients with postviral olfactory dysfunction. Laryngoscope 2007, 117, 272–277. [Google Scholar] [CrossRef]
- Imam, S.A.; Lao, W.P.; Reddy, P.; Nguyen, S.A.; Schlosser, R.J. Is SARS-CoV-2 (COVID-19) postviral olfactory dysfunction (PVOD) different from other PVOD? World J Otorhinolaryngol Head Neck Surg 2020, 6, S26–S32. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, M.; Aiba, T.; Mori, J.; Nakai, Y. An epidemiological study of postviral olfactory disorder. Acta Otolaryngol Suppl 1998, 538, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Chiguer, D.L.; Tirado-Mendoza, R.; Marquez-Navarro, A.; Ambrosio-Hernandez, J.R.; Ruiz-Fraga, I.; Aguilar-Vargas, R.E.; Lira-Martinez, J.M.; Lopez-Valdes, J.C. Detection and molecular characterization of respiratory viruses that cause acute respiratory infection in the adult population. Gac Med Mex 2019, 155, S7–S12. [Google Scholar] [CrossRef] [PubMed]
- Ueha, R.; Mukherjee, S.; Ueha, S.; de Almeida Nagata, D.E.; Sakamoto, T.; Kondo, K.; Yamasoba, T.; Lukacs, N.W.; Kunkel, S.L. Viral disruption of olfactory progenitors is exacerbated in allergic mice. Int Immunopharmacol 2014, 22, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Heikkinen, T.; Jarvinen, A. The common cold. Lancet 2003, 361, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Potter, M.R.; Chen, J.H.; Lobban, N.S.; Doty, R.L. Olfactory dysfunction from acute upper respiratory infections: relationship to season of onset. Int Forum Allergy Rhinol 2020, 10, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Henrickson, K.J. Parainfluenza viruses. Clin Microbiol Rev 2003, 16, 242–264. [Google Scholar] [CrossRef] [PubMed]
- Rafeek, R.A.M.; Divarathna, M.V.M.; Noordeen, F. A review on disease burden and epidemiology of childhood parainfluenza virus infections in Asian countries. Rev Med Virol 2021, 31, e2164. [Google Scholar] [CrossRef]
- Weston, S.; Frieman, M.B. Respiratory Viruses. In Encyclopedia of Microbiology (Fourth Edition); Schmidt, T.M., Ed.; Academic Press: Oxford, 2019; pp. 85–101. [Google Scholar] [CrossRef]
- van Kempen, M.; Bachert, C.; Van Cauwenberge, P. An update on the pathophysiology of rhinovirus upper respiratory tract infections. Rhinology 1999, 37, 97–103. [Google Scholar]
- Xatzipsalti, M.; Kyrana, S.; Tsolia, M.; Psarras, S.; Bossios, A.; Laza-Stanca, V.; Johnston, S.L.; Papadopoulos, N.G. Rhinovirus viremia in children with respiratory infections. Am J Respir Crit Care Med 2005, 172, 1037–1040. [Google Scholar] [CrossRef]
- Loeffelholz, M.J.; Trujillo, R.; Pyles, R.B.; Miller, A.L.; Alvarez-Fernandez, P.; Pong, D.L.; Chonmaitree, T. Duration of rhinovirus shedding in the upper respiratory tract in the first year of life. Pediatrics 2014, 134, 1144–1150. [Google Scholar] [CrossRef]
- WHO. Severe Acute Respiratory Syndrome (SARS).
- de Groot, R.J.; Baker, S.C.; Baric, R.S.; Brown, C.S.; Drosten, C.; Enjuanes, L.; Fouchier, R.A.; Galiano, M.; Gorbalenya, A.E.; Memish, Z.A.; et al. Middle East respiratory syndrome coronavirus (MERS-CoV): announcement of the Coronavirus Study Group. J Virol 2013, 87, 7790–7792. [Google Scholar] [CrossRef]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Engl J Med 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- de Groot, R.J.; Baker, S.C.; Baric, R.S.; Brown, C.S.; Drosten, C.; Enjuanes, L.; Fouchier, R.A.; Galiano, M.; Gorbalenya, A.E.; Memish, Z.A.; et al. Middle East respiratory syndrome coronavirus (MERS-CoV): announcement of the Coronavirus Study Group. J Virol 2013, 87, 7790–7792. [Google Scholar] [CrossRef]
- Khalafalla, A.I.; Lu, X.; Al-Mubarak, A.I.; Dalab, A.H.; Al-Busadah, K.A.; Erdman, D.D. MERS-CoV in Upper Respiratory Tract and Lungs of Dromedary Camels, Saudi Arabia, 2013-2014. Emerg Infect Dis 2015, 21, 1153–1158. [Google Scholar] [CrossRef]
- Wolfel, R.; Corman, V.M.; Guggemos, W.; Seilmaier, M.; Zange, S.; Muller, M.A.; Niemeyer, D.; Jones, T.C.; Vollmar, P.; Rothe, C.; et al. Virological assessment of hospitalized patients with COVID-2019. Nature 2020, 581, 465–469. [Google Scholar] [CrossRef]
- de Melo, G.D.; Lazarini, F.; Levallois, S.; Hautefort, C.; Michel, V.; Larrous, F.; Verillaud, B.; Aparicio, C.; Wagner, S.; Gheusi, G.; et al. COVID-19-related anosmia is associated with viral persistence and inflammation in human olfactory epithelium and brain infection in hamsters. Sci Transl Med 2021, 13. [Google Scholar] [CrossRef]
- Zazhytska, M.; Kodra, A.; Hoagland, D.A.; Frere, J.; Fullard, J.F.; Shayya, H.; McArthur, N.G.; Moeller, R.; Uhl, S.; Omer, A.D.; et al. Non-cell-autonomous disruption of nuclear architecture as a potential cause of COVID-19-induced anosmia. Cell 2022, 185, 1052–1064. [Google Scholar] [CrossRef]
- Bilinska, K.; Butowt, R. Anosmia in COVID-19: A Bumpy Road to Establishing a Cellular Mechanism. ACS Chem Neurosci 2020, 11, 2152–2155. [Google Scholar] [CrossRef]
- Samaranayake, L.P.; Fakhruddin, K.S.; Panduwawala, C. Sudden onset, acute loss of taste and smell in coronavirus disease 2019 (COVID-19): a systematic review. Acta Odontol Scand 2020, 78, 467–473. [Google Scholar] [CrossRef]
- Dammalli, M.; Dey, G.; Madugundu, A.K.; Kumar, M.; Rodrigues, B.; Gowda, H.; Siddaiah, B.G.; Mahadevan, A.; Shankar, S.K.; Prasad, T.S.K. Proteomic Analysis of the Human Olfactory Bulb. OMICS 2017, 21, 440–453. [Google Scholar] [CrossRef]
- Dammalli, M.; Dey, G.; Kumar, M.; Madugundu, A.K.; Gopalakrishnan, L.; Gowrishankar, B.S.; Mahadevan, A.; Shankar, S.K.; Prasad, T.S.K. Proteomics of the Human Olfactory Tract. OMICS 2018, 22, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Oboti, L.; Peretto, P.; Marchis, S.D.; Fasolo, A. From chemical neuroanatomy to an understanding of the olfactory system. Eur J Histochem 2011, 55, e35. [Google Scholar] [CrossRef] [PubMed]
- Barral-Arca, R.; Gomez-Carballa, A.; Cebey-Lopez, M.; Bello, X.; Martinon-Torres, F.; Salas, A. A Meta-Analysis of Multiple Whole Blood Gene Expression Data Unveils a Diagnostic Host-Response Transcript Signature for Respiratory Syncytial Virus. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Boesveldt, S.; Postma, E.M.; Boak, D.; Welge-Luessen, A.; Schopf, V.; Mainland, J.D.; Martens, J.; Ngai, J.; Duffy, V.B. Anosmia-A Clinical Review. Chem Senses 2017, 42, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Firestein, S. How the olfactory system makes sense of scents. Nature 2001, 413, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Brennan, P.A.; Keverne, E.B. Something in the air? New insights into mammalian pheromones. Curr Biol 2004, 14, R81–R89. [Google Scholar] [CrossRef] [PubMed]
- Buck, L.B. The molecular architecture of odor and pheromone sensing in mammals. Cell 2000, 100, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Mombaerts, P. Genes and ligands for odorant, vomeronasal and taste receptors. Nat Rev Neurosci 2004, 5, 263–278. [Google Scholar] [CrossRef]
- Restrepo, D.; Arellano, J.; Oliva, A.M.; Schaefer, M.L.; Lin, W. Emerging views on the distinct but related roles of the main and accessory olfactory systems in responsiveness to chemosensory signals in mice. Horm Behav 2004, 46, 247–256. [Google Scholar] [CrossRef]
- Lavoie, J.; Gasso Astorga, P.; Segal-Gavish, H.; Wu, Y.C.; Chung, Y.; Cascella, N.G.; Sawa, A.; Ishizuka, K. The Olfactory Neural Epithelium As a Tool in Neuroscience. Trends Mol Med 2017, 23, 100–103. [Google Scholar] [CrossRef]
- Liang, F.; Wang, Y. COVID-19 Anosmia: High Prevalence, Plural Neuropathogenic Mechanisms, and Scarce Neurotropism of SARS-CoV-2? Viruses 2021, 13. [Google Scholar] [CrossRef]
- Olender, T.; Lancet, D.; Nebert, D.W. Update on the olfactory receptor (OR) gene superfamily. Hum Genomics 2008, 3, 87–97. [Google Scholar] [CrossRef]
- Kurian, S.M.; Gordon, S.; Barrick, B.; Dadlani, M.N.; Fanelli, B.; Cornell, J.B.; Head, S.R.; Marsh, C.L.; Case, J. Feasibility and Comparison Study of Fecal Sample Collection Methods in Healthy Volunteers and Solid Organ Transplant Recipients Using 16S rRNA and Metagenomics Approaches. Biopreserv Biobank 2020, 18, 425–440. [Google Scholar] [CrossRef]
- Glezer, I.; Malnic, B. Olfactory receptor function. Handb Clin Neurol 2019, 164, 67–78. [Google Scholar] [CrossRef]
- Schwob, J.E. Neural regeneration and the peripheral olfactory system. Anat Rec 2002, 269, 33–49. [Google Scholar] [CrossRef]
- Graziadei, P.P.; Graziadei, G.A. Neurogenesis and neuron regeneration in the olfactory system of mammals. I. Morphological aspects of differentiation and structural organization of the olfactory sensory neurons. J Neurocytol 1979, 8, 1–18. [Google Scholar] [CrossRef]
- Dicpinigaitis, P.V. Post-viral Anosmia (Loss of Sensation of Smell) Did Not Begin with COVID-19! Lung 2021, 199, 237–238. [Google Scholar] [CrossRef]
- Urata, S.; Maruyama, J.; Kishimoto-Urata, M.; Sattler, R.A.; Cook, R.; Lin, N.; Yamasoba, T.; Makishima, T.; Paessler, S. Regeneration Profiles of Olfactory Epithelium after SARS-CoV-2 Infection in Golden Syrian Hamsters. ACS Chem Neurosci 2021, 12, 589–595. [Google Scholar] [CrossRef]
- Suzuki, M.; Saito, K.; Min, W.P.; Vladau, C.; Toida, K.; Itoh, H.; Murakami, S. Identification of viruses in patients with postviral olfactory dysfunction. Laryngoscope 2007, 117, 272–277. [Google Scholar] [CrossRef]
- Lee, D.Y.; Lee, W.H.; Wee, J.H.; Kim, J.W. Prognosis of postviral olfactory loss: follow-up study for longer than one year. Am J Rhinol Allergy 2014, 28, 419–422. [Google Scholar] [CrossRef]
- Tian, J.; Pinto, J.M.; Li, L.; Zhang, S.; Sun, Z.; Wei, Y. Identification of Viruses in Patients With Postviral Olfactory Dysfunction by Multiplex Reverse-Transcription Polymerase Chain Reaction. Laryngoscope 2021, 131, 158–164. [Google Scholar] [CrossRef]
- Suzuki, M.; Saito, K.; Min, W.P.; Vladau, C.; Toida, K.; Itoh, H.; Murakami, S. Identification of viruses in patients with postviral olfactory dysfunction. Laryngoscope 2007, 117, 272–277. [Google Scholar] [CrossRef]
- Seiden, A.M. Postviral olfactory loss. Otolaryngol Clin North Am 2004, 37, 1159–1166. [Google Scholar] [CrossRef]
- Doty, R.L.; Hawkes, C.H. Chemosensory dysfunction in neurodegenerative diseases. Handb Clin Neurol 2019, 164, 325–360. [Google Scholar] [CrossRef]
- Wang, J.H.; Kwon, H.J.; Jang, Y.J. Detection of parainfluenza virus 3 in turbinate epithelial cells of postviral olfactory dysfunction patients. Laryngoscope 2007, 117, 1445–1449. [Google Scholar] [CrossRef]
- Lewandowska-Polak, A.; Brauncajs, M.; Paradowska, E.; Jarzebska, M.; Kurowski, M.; Moskwa, S.; Lesnikowski, Z.J.; Kowalski, M.L. Human parainfluenza virus type 3 (HPIV3) induces production of IFNgamma and RANTES in human nasal epithelial cells (HNECs). J Inflamm (Lond) 2015, 12, 16. [Google Scholar] [CrossRef]
- Mori, I.; Komatsu, T.; Takeuchi, K.; Nakakuki, K.; Sudo, M.; Kimura, Y. Parainfluenza virus type 1 infects olfactory neurons and establishes long-term persistence in the nerve tissue. J Gen Virol 1995, 76 ( Pt 5) Pt 5, 1251–1254. [Google Scholar] [CrossRef]
- Tian, J.; Pinto, J.M.; Cui, X.; Zhang, H.; Li, L.; Liu, Y.; Wu, C.; Wei, Y. Sendai Virus Induces Persistent Olfactory Dysfunction in a Murine Model of PVOD via Effects on Apoptosis, Cell Proliferation, and Response to Odorants. PLoS One 2016, 11, e0159033. [Google Scholar] [CrossRef]
- Wu, C.T.; Lidsky, P.V.; Xiao, Y.; Cheng, R.; Lee, I.T.; Nakayama, T.; Jiang, S.; He, W.; Demeter, J.; Knight, M.G.; et al. SARS-CoV-2 replication in airway epithelia requires motile cilia and microvillar reprogramming. Cell 2023, 186, 112–130. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Volpe, S.J.; Chang, E.H. The Role of Viruses in the Inception of Chronic Rhinosinusitis. Clin Exp Otorhinolaryngol 2022, 15, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Choudhary, S.; Banerjee, A.K.; De, B.P. Human parainfluenza virus type 3 upregulates ICAM-1 (CD54) expression in a cytokine-independent manner. Gene Expr 2000, 9, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Bryche, B.; Fretaud, M.; Saint-Albin Deliot, A.; Galloux, M.; Sedano, L.; Langevin, C.; Descamps, D.; Rameix-Welti, M.A.; Eleouet, J.F.; Le Goffic, R.; et al. Respiratory syncytial virus tropism for olfactory sensory neurons in mice. J Neurochem 2020, 155, 137–153. [Google Scholar] [CrossRef]
- Griffiths, C.D.; Bilawchuk, L.M.; McDonough, J.E.; Jamieson, K.C.; Elawar, F.; Cen, Y.; Duan, W.; Lin, C.; Song, H.; Casanova, J.L.; et al. IGF1R is an entry receptor for respiratory syncytial virus. Nature 2020, 583, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Tayyari, F.; Marchant, D.; Moraes, T.J.; Duan, W.; Mastrangelo, P.; Hegele, R.G. Identification of nucleolin as a cellular receptor for human respiratory syncytial virus. Nat Med 2011, 17, 1132–1135. [Google Scholar] [CrossRef] [PubMed]
- Mastrangelo, P.; Chin, A.A.; Tan, S.; Jeon, A.H.; Ackerley, C.A.; Siu, K.K.; Lee, J.E.; Hegele, R.G. Identification of RSV Fusion Protein Interaction Domains on the Virus Receptor, Nucleolin. Viruses 2021, 13. [Google Scholar] [CrossRef]
- Sourimant, J.; Lieber, C.M.; Aggarwal, M.; Cox, R.M.; Wolf, J.D.; Yoon, J.J.; Toots, M.; Ye, C.; Sticher, Z.; Kolykhalov, A.A.; et al. 4'-Fluorouridine is an oral antiviral that blocks respiratory syncytial virus and SARS-CoV-2 replication. Science 2022, 375, 161–167. [Google Scholar] [CrossRef]
- Li, H.H.; Xu, J.; He, L.; Denny, L.I.; Rustandi, R.R.; Dornadula, G.; Fiorito, B.; Zhang, Z.Q. Development and qualification of cell-based relative potency assay for a human respiratory syncytial virus (RSV) mRNA vaccine. J Pharm Biomed Anal 2023, 234, 115523. [Google Scholar] [CrossRef]
- Whitaker, J.A.; Sahly, H.M.E.; Healy, C.M. mRNA vaccines against respiratory viruses. Curr Opin Infect Dis 2023, 36, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, C.D.; Bilawchuk, L.M.; McDonough, J.E.; Jamieson, K.C.; Elawar, F.; Cen, Y.; Duan, W.; Lin, C.; Song, H.; Casanova, J.L.; et al. IGF1R is an entry receptor for respiratory syncytial virus. Nature 2020, 583, 615–619. [Google Scholar] [CrossRef]
- Bochkov, Y.A.; Watters, K.; Ashraf, S.; Griggs, T.F.; Devries, M.K.; Jackson, D.J.; Palmenberg, A.C.; Gern, J.E. Cadherin-related family member 3, a childhood asthma susceptibility gene product, mediates rhinovirus C binding and replication. Proc Natl Acad Sci U S A 2015, 112, 5485–5490. [Google Scholar] [CrossRef]
- Staunton, D.E.; Merluzzi, V.J.; Rothlein, R.; Barton, R.; Marlin, S.D.; Springer, T.A. A cell adhesion molecule, ICAM-1, is the major surface receptor for rhinoviruses. Cell 1989, 56, 849–853. [Google Scholar] [CrossRef] [PubMed]
- Hofer, F.; Gruenberger, M.; Kowalski, H.; Machat, H.; Huettinger, M.; Kuechler, E.; Blaas, D. Members of the low density lipoprotein receptor family mediate cell entry of a minor-group common cold virus. Proc Natl Acad Sci U S A 1994, 91, 1839–1842. [Google Scholar] [CrossRef] [PubMed]
- Bochkov, Y.A.; Gern, J.E. Rhinoviruses and Their Receptors: Implications for Allergic Disease. Curr Allergy Asthma Rep 2016, 16, 30. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Han, M.S.; Lee, T.H.; Lee, D.B.; Park, J.H.; Lee, S.H.; Kim, T.H. Hydrogen peroxide attenuates rhinovirus-induced anti-viral interferon secretion in sinonasal epithelial cells. Front Immunol 2023, 14, 1086381. [Google Scholar] [CrossRef] [PubMed]
- Torabi, A.; Mohammadbagheri, E.; Akbari Dilmaghani, N.; Bayat, A.H.; Fathi, M.; Vakili, K.; Alizadeh, R.; Rezaeimirghaed, O.; Hajiesmaeili, M.; Ramezani, M.; et al. Proinflammatory Cytokines in the Olfactory Mucosa Result in COVID-19 Induced Anosmia. ACS Chem Neurosci 2020, 11, 1909–1913. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wong, L.R.; Li, K.; Verma, A.K.; Ortiz, M.E.; Wohlford-Lenane, C.; Leidinger, M.R.; Knudson, C.M.; Meyerholz, D.K.; McCray, P.B., Jr.; et al. COVID-19 treatments and pathogenesis including anosmia in K18-hACE2 mice. Nature 2021, 589, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Bilinska, K.; Jakubowska, P.; Von Bartheld, C.S.; Butowt, R. Expression of the SARS-CoV-2 Entry Proteins, ACE2 and TMPRSS2, in Cells of the Olfactory Epithelium: Identification of Cell Types and Trends with Age. ACS Chem Neurosci 2020, 11, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Randeva, H.S.; Chatha, K.; Hall, M.; Spandidos, D.A.; Karteris, E.; Kyrou, I. Neuropilin-1 as a new potential SARS-CoV-2 infection mediator implicated in the neurologic features and central nervous system involvement of COVID-19. Mol Med Rep 2020, 22, 4221–4226. [Google Scholar] [CrossRef]
- Kang, Y.L.; Chou, Y.Y.; Rothlauf, P.W.; Liu, Z.; Soh, T.K.; Cureton, D.; Case, J.B.; Chen, R.E.; Diamond, M.S.; Whelan, S.P.J.; et al. Inhibition of PIKfyve kinase prevents infection by Zaire ebolavirus and SARS-CoV-2. Proc Natl Acad Sci U S A 2020, 117, 20803–20813. [Google Scholar] [CrossRef]
- Mayi, B.S.; Leibowitz, J.A.; Woods, A.T.; Ammon, K.A.; Liu, A.E.; Raja, A. The role of Neuropilin-1 in COVID-19. PLoS Pathog 2021, 17, e1009153. [Google Scholar] [CrossRef]
- Bryche, B.; St Albin, A.; Murri, S.; Lacote, S.; Pulido, C.; Ar Gouilh, M.; Lesellier, S.; Servat, A.; Wasniewski, M.; Picard-Meyer, E.; et al. Massive transient damage of the olfactory epithelium associated with infection of sustentacular cells by SARS-CoV-2 in golden Syrian hamsters. Brain Behav Immun 2020, 89, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.L.; Simonetti, B.; Klein, K.; Chen, K.E.; Williamson, M.K.; Anton-Plagaro, C.; Shoemark, D.K.; Simon-Gracia, L.; Bauer, M.; Hollandi, R.; et al. Neuropilin-1 is a host factor for SARS-CoV-2 infection. Science 2020, 370, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, C.; Blanchard, K.; Bacigalupo, J.; Vergara, C. Possible ATP trafficking by ATP-shuttles in the olfactory cilia and glucose transfer across the olfactory mucosa. FEBS Lett 2019, 593, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Villar, P.S.; Vergara, C.; Bacigalupo, J. Energy sources that fuel metabolic processes in protruding finger-like organelles. FEBS J 2021, 288, 3799–3812. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Nordqvist, H.; Ambikan, A.T.; Gupta, S.; Sperk, M.; Svensson-Akusjarvi, S.; Mikaeloff, F.; Benfeitas, R.; Saccon, E.; Ponnan, S.M.; et al. Metabolic Perturbation Associated With COVID-19 Disease Severity and SARS-CoV-2 Replication. Mol Cell Proteomics 2021, 20, 100159. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Yoo, S.J.; Clijsters, M.; Backaert, W.; Vanstapel, A.; Speleman, K.; Lietaer, C.; Choi, S.; Hether, T.D.; Marcelis, L.; et al. Visualizing in deceased COVID-19 patients how SARS-CoV-2 attacks the respiratory and olfactory mucosae but spares the olfactory bulb. Cell 2021, 184, 5932–5949. [Google Scholar] [CrossRef] [PubMed]
- Assou, S.; Ahmed, E.; Morichon, L.; Nasri, A.; Foisset, F.; Bourdais, C.; Gros, N.; Tieo, S.; Petit, A.; Vachier, I.; et al. The Transcriptome Landscape of the In Vitro Human Airway Epithelium Response to SARS-CoV-2. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Hofer, F.; Gruenberger, M.; Kowalski, H.; Machat, H.; Huettinger, M.; Kuechler, E.; Blaas, D. Members of the low density lipoprotein receptor family mediate cell entry of a minor-group common cold virus. Proc Natl Acad Sci U S A 1994, 91, 1839–1842. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, T.B.; Kalie, E.; Crisafulli-Cabatu, S.; Abramovich, R.; DiGioia, G.; Moolchan, K.; Pestka, S.; Schreiber, G. Binding and activity of all human alpha interferon subtypes. Cytokine 2011, 56, 282–289. [Google Scholar] [CrossRef]
- Jaks, E.; Gavutis, M.; Uze, G.; Martal, J.; Piehler, J. Differential receptor subunit affinities of type I interferons govern differential signal activation. J Mol Biol 2007, 366, 525–539. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, R.P.; Sheikh, F.; Kotenko, S.V.; Dickensheets, H. The expanded family of class II cytokines that share the IL-10 receptor-2 (IL-10R2) chain. J Leukoc Biol 2004, 76, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ninaber, D.K.; van Schadewijk, A.; Hiemstra, P.S. Tiotropium and Fluticasone Inhibit Rhinovirus-Induced Mucin Production via Multiple Mechanisms in Differentiated Airway Epithelial Cells. Front Cell Infect Microbiol 2020, 10, 278. [Google Scholar] [CrossRef]
- Lo, D.; Kennedy, J.L.; Kurten, R.C.; Panettieri, R.A., Jr.; Koziol-White, C.J. Modulation of airway hyperresponsiveness by rhinovirus exposure. Respir Res 2018, 19, 208. [Google Scholar] [CrossRef]
- Loxham, M.; Smart, D.E.; Bedke, N.J.; Smithers, N.P.; Filippi, I.; Blume, C.; Swindle, E.J.; Tariq, K.; Howarth, P.H.; Holgate, S.T.; et al. Allergenic proteases cleave the chemokine CX3CL1 directly from the surface of airway epithelium and augment the effect of rhinovirus. Mucosal Immunol 2018, 11, 404–414. [Google Scholar] [CrossRef]
- Papi, A.; Papadopoulos, N.G.; Stanciu, L.A.; Bellettato, C.M.; Pinamonti, S.; Degitz, K.; Holgate, S.T.; Johnston, S.L. Reducing agents inhibit rhinovirus-induced up-regulation of the rhinovirus receptor intercellular adhesion molecule-1 (ICAM-1) in respiratory epithelial cells. FASEB J 2002, 16, 1934–1936. [Google Scholar] [CrossRef]
- Song, Y.P.; Tang, M.F.; Leung, A.S.Y.; Tao, K.P.; Chan, O.M.; Wong, G.W.K.; Chan, P.K.S.; Chan, R.W.Y.; Leung, T.F. Interactive effects between CDHR3 genotype and rhinovirus species for diagnosis and severity of respiratory tract infections in hospitalized children. Microbiol Spectr 2023, e0118123. [Google Scholar] [CrossRef]
- Greiller, C.L.; Suri, R.; Jolliffe, D.A.; Kebadze, T.; Hirsman, A.G.; Griffiths, C.J.; Johnston, S.L.; Martineau, A.R. Vitamin D attenuates rhinovirus-induced expression of intercellular adhesion molecule-1 (ICAM-1) and platelet-activating factor receptor (PAFR) in respiratory epithelial cells. J Steroid Biochem Mol Biol 2019, 187, 152–159. [Google Scholar] [CrossRef]
- Khani, E.; Khiali, S.; Beheshtirouy, S.; Entezari-Maleki, T. Potential pharmacologic treatments for COVID-19 smell and taste loss: A comprehensive review. Eur J Pharmacol 2021, 912, 174582. [Google Scholar] [CrossRef]
- Silva Andrade, B.; Siqueira, S.; de Assis Soares, W.R.; de Souza Rangel, F.; Santos, N.O.; Dos Santos Freitas, A.; Ribeiro da Silveira, P.; Tiwari, S.; Alzahrani, K.J.; Goes-Neto, A.; et al. Long-COVID and Post-COVID Health Complications: An Up-to-Date Review on Clinical Conditions and Their Possible Molecular Mechanisms. Viruses 2021, 13. [Google Scholar] [CrossRef]
- Fernandez-de-Las-Penas, C.; Cancela-Cilleruelo, I.; Rodriguez-Jimenez, J.; Gomez-Mayordomo, V.; Pellicer-Valero, O.J.; Martin-Guerrero, J.D.; Hernandez-Barrera, V.; Arendt-Nielsen, L.; Torres-Macho, J. Associated-Onset Symptoms and Post-COVID-19 Symptoms in Hospitalized COVID-19 Survivors Infected with Wuhan, Alpha or Delta SARS-CoV-2 Variant. Pathogens 2022, 11. [Google Scholar] [CrossRef]
- Rodriguez-Sevilla, J.J.; Guerri-Fernadez, R.; Bertran Recasens, B. Is There Less Alteration of Smell Sensation in Patients With Omicron SARS-CoV-2 Variant Infection? Front Med (Lausanne) 2022, 9, 852998. [Google Scholar] [CrossRef]
- Chee, J.; Chern, B.; Loh, W.S.; Mullol, J.; Wang, Y. Pathophysiology of SARS-CoV-2 Infection of Nasal Respiratory and Olfactory Epithelia and Its Clinical Impact. Curr Allergy Asthma Rep 2023, 23, 121–131. [Google Scholar] [CrossRef]
- Mutiawati, E.; Fahriani, M.; Mamada, S.S.; Fajar, J.K.; Frediansyah, A.; Maliga, H.A.; Ilmawan, M.; Emran, T.B.; Ophinni, Y.; Ichsan, I.; et al. Anosmia and dysgeusia in SARS-CoV-2 infection: incidence and effects on COVID-19 severity and mortality, and the possible pathobiology mechanisms - a systematic review and meta-analysis. F1000Res 2021, 10, 40. [Google Scholar] [CrossRef]
- Butowt, R.; von Bartheld, C.S. Anosmia in COVID-19: Underlying Mechanisms and Assessment of an Olfactory Route to Brain Infection. Neuroscientist 2021, 27, 582–603. [Google Scholar] [CrossRef]
- Kalra, R.S.; Dhanjal, J.K.; Meena, A.S.; Kalel, V.C.; Dahiya, S.; Singh, B.; Dewanjee, S.; Kandimalla, R. COVID-19, Neuropathology, and Aging: SARS-CoV-2 Neurological Infection, Mechanism, and Associated Complications. Front Aging Neurosci 2021, 13, 662786. [Google Scholar] [CrossRef] [PubMed]
- Rebholz, H.; Braun, R.J.; Ladage, D.; Knoll, W.; Kleber, C.; Hassel, A.W. Loss of Olfactory Function-Early Indicator for Covid-19, Other Viral Infections and Neurodegenerative Disorders. Front Neurol 2020, 11, 569333. [Google Scholar] [CrossRef] [PubMed]
- Reyna, R.A.; Kishimoto-Urata, M.; Urata, S.; Makishima, T.; Paessler, S.; Maruyama, J. Recovery of anosmia in hamsters infected with SARS-CoV-2 is correlated with repair of the olfactory epithelium. Sci Rep 2022, 12, 628. [Google Scholar] [CrossRef] [PubMed]
- Tanzadehpanah, H.; Lotfian, E.; Avan, A.; Saki, S.; Nobari, S.; Mahmoodian, R.; Sheykhhasan, M.; Froutagh, M.H.S.; Ghotbani, F.; Jamshidi, R.; et al. Role of SARS-COV-2 and ACE2 in the pathophysiology of peripheral vascular diseases. Biomed Pharmacother 2023, 166, 115321. [Google Scholar] [CrossRef]
- Mutiawati, E.; Fahriani, M.; Mamada, S.S.; Fajar, J.K.; Frediansyah, A.; Maliga, H.A.; Ilmawan, M.; Emran, T.B.; Ophinni, Y.; Ichsan, I.; et al. Anosmia and dysgeusia in SARS-CoV-2 infection: incidence and effects on COVID-19 severity and mortality, and the possible pathobiology mechanisms - a systematic review and meta-analysis. F1000Res 2021, 10, 40. [Google Scholar] [CrossRef]
- Butowt, R.; von Bartheld, C.S. Anosmia in COVID-19: Underlying Mechanisms and Assessment of an Olfactory Route to Brain Infection. Neuroscientist 2021, 27, 582–603. [Google Scholar] [CrossRef]
- Butowt, R.; Bilinska, K.; von Bartheld, C.S. Olfactory dysfunction in COVID-19: new insights into the underlying mechanisms. Trends Neurosci 2023, 46, 75–90. [Google Scholar] [CrossRef]
- Buqaileh, R.; Saternos, H.; Ley, S.; Aranda, A.; Forero, K.; AbouAlaiwi, W.A. Can cilia provide an entry gateway for SARS-CoV-2 to human ciliated cells? Physiol Genomics 2021, 53, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Butowt, R.; Meunier, N.; Bryche, B.; von Bartheld, C.S. The olfactory nerve is not a likely route to brain infection in COVID-19: a critical review of data from humans and animal models. Acta Neuropathol 2021, 141, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Bilinska, K.; Jakubowska, P.; Von Bartheld, C.S.; Butowt, R. Expression of the SARS-CoV-2 Entry Proteins, ACE2 and TMPRSS2, in Cells of the Olfactory Epithelium: Identification of Cell Types and Trends with Age. ACS Chem Neurosci 2020, 11, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Bryche, B.; St Albin, A.; Murri, S.; Lacote, S.; Pulido, C.; Ar Gouilh, M.; Lesellier, S.; Servat, A.; Wasniewski, M.; Picard-Meyer, E.; et al. Massive transient damage of the olfactory epithelium associated with infection of sustentacular cells by SARS-CoV-2 in golden Syrian hamsters. Brain Behav Immun 2020, 89, 579–586. [Google Scholar] [CrossRef]
- Sia, S.F.; Yan, L.M.; Chin, A.W.H.; Fung, K.; Choy, K.T.; Wong, A.Y.L.; Kaewpreedee, P.; Perera, R.; Poon, L.L.M.; Nicholls, J.M.; et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Lechien, J.R.; Chiesa-Estomba, C.M.; De Siati, D.R.; Horoi, M.; Le Bon, S.D.; Rodriguez, A.; Dequanter, D.; Blecic, S.; El Afia, F.; Distinguin, L.; et al. Olfactory and gustatory dysfunctions as a clinical presentation of mild-to-moderate forms of the coronavirus disease (COVID-19): a multicenter European study. Eur Arch Otorhinolaryngol 2020, 277, 2251–2261. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Min, P.; Lee, S.; Kim, S.W. Prevalence and Duration of Acute Loss of Smell or Taste in COVID-19 Patients. J Korean Med Sci 2020, 35, e174. [Google Scholar] [CrossRef]
- Fodoulian, L.; Tuberosa, J.; Rossier, D.; Boillat, M.; Kan, C.; Pauli, V.; Egervari, K.; Lobrinus, J.A.; Landis, B.N.; Carleton, A.; et al. SARS-CoV-2 Receptors and Entry Genes Are Expressed in the Human Olfactory Neuroepithelium and Brain. iScience 2020, 23, 101839. [Google Scholar] [CrossRef]
- Chen, M.; Shen, W.; Rowan, N.R.; Kulaga, H.; Hillel, A.; Ramanathan, M., Jr.; Lane, A.P. Elevated ACE-2 expression in the olfactory neuroepithelium: implications for anosmia and upper respiratory SARS-CoV-2 entry and replication. Eur Respir J 2020, 56. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Ziegler, C.G.K.; Allon, S.J.; Nyquist, S.K.; Mbano, I.M.; Miao, V.N.; Tzouanas, C.N.; Cao, Y.; Yousif, A.S.; Bals, J.; Hauser, B.M.; et al. SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets across Tissues. Cell 2020, 181, 1016–1035. [Google Scholar] [CrossRef]
- Gkogkou, E.; Barnasas, G.; Vougas, K.; Trougakos, I.P. Expression profiling meta-analysis of ACE2 and TMPRSS2, the putative anti-inflammatory receptor and priming protease of SARS-CoV-2 in human cells, and identification of putative modulators. Redox Biol 2020, 36, 101615. [Google Scholar] [CrossRef]
- Sungnak, W.; Huang, N.; Becavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-Lopez, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat Med 2020, 26, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, M.; Ou, G. COVID-19, cilia, and smell. FEBS J 2020, 287, 3672–3676. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, A.; Naujokat, C. Structural features of coronavirus SARS-CoV-2 spike protein: Targets for vaccination. Life Sci 2020, 257, 118056. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Pekosz, A.; Villano, J.S.; Shen, W.; Zhou, R.; Kulaga, H.; Li, Z.; Beck, S.E.; Witwer, K.W.; Mankowski, J.L.; et al. Evolution of nasal and olfactory infection characteristics of SARS-CoV-2 variants. bioRxiv 2022. [Google Scholar] [CrossRef]
- von Bartheld, C.S.; Wang, L. Prevalence of Olfactory Dysfunction with the Omicron Variant of SARS-CoV-2: A Systematic Review and Meta-Analysis. Cells 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- von Bartheld, C.S.; Hagen, M.M.; Butowt, R. The D614G Virus Mutation Enhances Anosmia in COVID-19 Patients: Evidence from a Systematic Review and Meta-analysis of Studies from South Asia. ACS Chem Neurosci 2021, 12, 3535–3549. [Google Scholar] [CrossRef]
- Schreiner, T.; Allnoch, L.; Beythien, G.; Marek, K.; Becker, K.; Schaudien, D.; Stanelle-Bertram, S.; Schaumburg, B.; Mounogou Kouassi, N.; Beck, S.; et al. SARS-CoV-2 Infection Dysregulates Cilia and Basal Cell Homeostasis in the Respiratory Epithelium of Hamsters. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Liang, F.; Wang, Y. COVID-19 Anosmia: High Prevalence, Plural Neuropathogenic Mechanisms, and Scarce Neurotropism of SARS-CoV-2? Viruses 2021, 13. [Google Scholar] [CrossRef]
- Seehusen, F.; Clark, J.J.; Sharma, P.; Bentley, E.G.; Kirby, A.; Subramaniam, K.; Wunderlin-Giuliani, S.; Hughes, G.L.; Patterson, E.I.; Michael, B.D.; et al. Neuroinvasion and Neurotropism by SARS-CoV-2 Variants in the K18-hACE2 Mouse. Viruses 2022, 14. [Google Scholar] [CrossRef]
- Trobaugh, D.W.; Klimstra, W.B. MicroRNA Regulation of RNA Virus Replication and Pathogenesis. Trends Mol Med 2017, 23, 80–93. [Google Scholar] [CrossRef]
- Hanna, J.; Hossain, G.S.; Kocerha, J. The Potential for microRNA Therapeutics and Clinical Research. Front Genet 2019, 10, 478. [Google Scholar] [CrossRef] [PubMed]
- Osan, J.K.; DeMontigny, B.A.; Mehedi, M. Immunohistochemistry for protein detection in PFA-fixed paraffin-embedded SARS-CoV-2-infected COPD airway epithelium. STAR Protoc 2021, 2, 100663. [Google Scholar] [CrossRef] [PubMed]
- Li, H.H.; Xu, J.; He, L.; Denny, L.I.; Rustandi, R.R.; Dornadula, G.; Fiorito, B.; Zhang, Z.Q. Development and qualification of cell-based relative potency assay for a human respiratory syncytial virus (RSV) mRNA vaccine. J Pharm Biomed Anal 2023, 234, 115523. [Google Scholar] [CrossRef] [PubMed]
- Mayi, B.S.; Leibowitz, J.A.; Woods, A.T.; Ammon, K.A.; Liu, A.E.; Raja, A. The role of Neuropilin-1 in COVID-19. PLoS Pathog 2021, 17, e1009153. [Google Scholar] [CrossRef] [PubMed]
- Iacobucci, G. Covid-19: Runny nose, headache, and fatigue are commonest symptoms of omicron, early data show. BMJ 2021, 375, n3103. [Google Scholar] [CrossRef] [PubMed]
- Ohkubo, K.; Lee, C.H.; Baraniuk, J.N.; Merida, M.; Hausfeld, J.N.; Kaliner, M.A. Angiotensin-converting enzyme in the human nasal mucosa. Am J Respir Cell Mol Biol 1994, 11, 173–180. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Respiratory viruses playing a role in the olfactory impairment. (A) Parainfluenza virus [8] induced the inflammation response with damage of olfactory epithelium and the impairment of olfactory response leading to more virus susceptibility. (B) Respiratory syncytial virus (RSV) induced the inflammation response related to the age group with damage of olfactory epithelium and OSN progenitors and the impairment of olfactory response leading to more virus susceptibility. (C) Rhinovirus (RV) induced the inflammation response with damage of olfactory epithelium, the degradation of the tight junction and adherent junction markers and the impairment of olfactory response leading to more virus susceptibility. (D) SARS-CoV-2 induced disruption of the nasal epithelium, with loss/damage of olfactory sensory neurons, sustentacular cells, Bowman’s gland and supporting cells. The figure was created with BioRender.com.
Figure 1.
Respiratory viruses playing a role in the olfactory impairment. (A) Parainfluenza virus [8] induced the inflammation response with damage of olfactory epithelium and the impairment of olfactory response leading to more virus susceptibility. (B) Respiratory syncytial virus (RSV) induced the inflammation response related to the age group with damage of olfactory epithelium and OSN progenitors and the impairment of olfactory response leading to more virus susceptibility. (C) Rhinovirus (RV) induced the inflammation response with damage of olfactory epithelium, the degradation of the tight junction and adherent junction markers and the impairment of olfactory response leading to more virus susceptibility. (D) SARS-CoV-2 induced disruption of the nasal epithelium, with loss/damage of olfactory sensory neurons, sustentacular cells, Bowman’s gland and supporting cells. The figure was created with BioRender.com.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



