
Submitted:
11 May 2023
Posted:
12 May 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Since 2020 COVID-19 has caused serious mortality around the world. Given the ambiguity in es-tablishing COVID-19 as the direct cause of death we first investigate the effects of age and sex upon all-cause mortality during 2020 and 2021 in England and Wales. Since infectious agents have their own unique age profile for death, we explore several methods to adjust single year of age deaths in England and Wales during 2019 (the pre-COVID-19 base year) to a pathogen-neutral single year of age baseline. This adjusted base year is then used to confirm the widely reported higher deaths in males for most ages above 43 years in both 2020 and 2021. During 2020 (+ COVID-19 but no vac-cination) both male and female population-adjusted deaths were significantly increased above age 35. A significant reduction in all-cause mortality among both males and females aged 75+ could be demonstrated in 2021 during widespread COVID–19 vaccination, however, below age 75 deaths progressively increased. This finding arises from a mix of vaccination and year of age profiles of deaths for the different SARS-CoV-2 variants. In addition, specific effects for age around puberty were demonstrated where females had higher deaths rather than males. There is evidence that year-of-birth cohorts may also be involved, indicating that immune priming to specific pathogen outbreaks in the past may lead to lower deaths for some birth cohorts. To specifically identify the age profile for the COVID-19 variants in 2020 to 2023, we employ the proportion of total deaths at each age which are potentially due to or ‘with’ COVID-19. The original Wuhan strain, and the Alpha variant show somewhat limited divergence in the age profile with the Alpha variant shifted to a moderately higher proportion of deaths below age 84. The Delta variant specifically targeted persons below age 65. The Omicron variants showed significantly lower proportion of overall mortality, with markedly higher relative proportion of deaths above age 65 steeply increasing with age to a maximum around 100 years of age. A similar age profile for the variants can be seen in the age-banded deaths in US states – although slightly obscured by using age bands rather than single year of age. However, the US data shows that higher male deaths are greatly dependent on age and the COVID-variant. Deaths determined as ‘due to’ COVID-19 (as opposed to ‘involving’ COVID-19) in England and Wales were especially over-estimated in 2021 relative to the change in all-cause mortality. This arose as a by-product of an increase in COVID-19 testing capacity in late 2020. Potential structure-function mechanisms for the year of age specificity of SARS-CoV-2 var-iants are discussed. The question is posed as to whether vaccines based on different variants carry the specific age profile through into the vaccine.
Keywords:
COVID–19
; all-cause mortality
; single year of age
; gender
; complex system
; pathogen interference
; seasonality
; small RNAs
1. Introduction
The SARS-CoV-2 pandemic has caused considerable human morbidity and mortality, necessitating studies to develop clinical protocols, repurpose existing drugs and develop vaccines to counter these adverse effects [1]. The pandemic has seen a progression of new variants emerging in different parts of the world which appear to have different age profiles for death [1,2] – although the precise nature of the age profile for death is obscured by the fact that different studies use a wide range of age bands ending at age 75+ through to 90+ [3,4,5,6,7].
Influenza and SARS-CoV-2 are both examples of the class of RNA viruses showing high mutation rates [8,9,10,11,12]. In the UK, the COVID–19 pandemic commenced somewhere in early 2020 with the first laboratory-confirmed death occurring on 2 March 2020 [13]. However, COVID–19 testing capacity in the UK was very low at that time and earlier deaths due to COVID-19 were possible [14].
Research in the USA suggests that COVID–19 deaths may have started in early January 2020 [15]. A general paucity of testing capacity in the UK during the first year of the pandemic [14] implies that all-cause mortality becomes a good proxy for the incremental changes in real COVID-19 mortality [16].
In both the UK and USA, the original strain predominates during 2020. The Alpha strain appears around October/November 2020 and predominates from January to June 2021, the Delta strain commences around May 2021 and predominates from July to December 2021. While Omicron first emerges in November 2021 but begins to spread in December 2021 and dominates in 2022 from around March onwards [17,18]. The Beta strain did not appear to gain a major foothold in these two countries [16,17]. Hence, we have the pre-COVID-19 era which ends in December 2019 through to the ongoing surges as new variants come to the fore from the end of 2020 onward [17,18].
It is important to recall that deaths are a lagging indicator since infection, illness and hospitalization precedes recovery or death. The gap between infection and death is highly likely to be a probability distribution which depends on age and risk factors [19]. The Alpha variant caused slightly higher mortality than the original strain and primarily affected mortality in the winter of 2020/21. The Delta variant mainly affected the winter of 2021/22 with higher transmission and a slightly lower or equal mortality risk [20,21,22].
In the UK, COVID-19 vaccines were approved and deployed in the following order: Pfizer/BioNTech (2 December 2020 - deployed 8 December 2020), AstraZeneca (30 December 2020 - deployed 4 January 2021), Moderna (8 January 2021 - deployed 7 April 2021) [23,24,25,26]. Data regarding the proportions of persons vaccinated by age and time with the different manufacturers (Pfizer/AstraZeneca/Moderna) are not publicly available.
COVID–19 vaccination began on 8 Dec 2020 for care home residents, persons aged 80+, and some health care workers, by 18 January 2021 this included age 70+ and persons with very high clinical risk, by 15 February age 65+ and persons with high risk, and by 22 May age 32+ and age 18+ by 18 June 2021 [23,24,25,26]. Following reports of a rare type of blood clot in late March 2021 for the AstraZeneca vaccine, persons under 30 years were all given the mRNA vaccine from 7 April 2021 onward, and those aged under 40 from 7 May 2021 onward [26]. Vaccination of persons aged 16 ̶ 17 years was from July 2021 onward, 12 ̶ 15 years from September 2021 onward and 5 ̶ 11 years from February 2022 onwards. The majority aged 12+ were vaccinated during late 2021. All with mRNA as per the age under-40 rule from above. From around spring 2022 onward all persons were vaccinated (including booster) mainly with the mRNA vaccine [23,24,25,26].
As mentioned above, studies on the effects of age generally use wide age bands which can conceal the underlying year of age behavior. A world bank analysis of the COVID-19-related mortality rate during the first wave of COVID-19 in 2020 used 10-year age bands commencing at 30-39 through to age 80+ to demonstrate typical logarithmic decay in the mortality rate as age reduces. The World Bank study notes that the mortality rate in the low-income countries is flatter than for high-income countries with up to 10-times higher mortality rate at age 30-39 [3].
Another study using 2020 data shows a similar log-decay and estimates that the COVID-19 mortality rate, relative to the average for influenza and pneumonia (1999-2018), is 5-times higher at age 25-34, 8-times higher at 55-64, but only 3-times higher at 85+ [4]. Adjei et al [5] compare in-hospital mortality rates during Delta and Omicron variants using 15-year age bands, 0-17, through to 80+. Mortality rates at age band 0-17 is Delta/Omicron 0.7/0.9 through to 21.6/6.5 at age 80+. Stepanova et al [6] used 10-year age bands commencing at age <55 years, through to age 75+ when studying the effects of vaccination upon hospitalization and death during Alpha, Delta, and Omicron variants [6]. Leiner et al [7] use age bands <59 years, 60-69, 70-79, 80+ to adjust hospital outcomes during Delta and Omicron [7]. Without detailing all studies there is gross variation between age bands and hidden assumptions regarding behavior within age bands. Arbitrary age bands can cut across, and so conceal, underlying trends.
An excellent example of the potential for age bands to conceal relevant behavior was a large maximum in deaths during the 1918-19 Spanish flu pandemic at the precise age of 28, due to exposure of this birth cohort to the previous 1889-90 Russian flu pandemic. Seemingly this birth cohort were ‘vaccinated’ due to natural exposure against Russian flu which had adverse outcomes when later exposed to the antigenically dissimilar Spanish flu [27].
As far as we are aware there are no detailed year of age studies for SARS-CoV-2 variants, and this study therefore seeks to establish the year of age profiles for the deaths arising from different variants which predominated in England and Wales between 2020 to 2022. These will be compared to age-banded results for the USA and US states. A method based on the proportion of total all-cause deaths either ‘with’ or ‘due to’ COVID-19 is used to separate out the specific effects against age per se.
We first use year of age calendar year data for 2019 to 2022 for England and Wales covering all-cause mortality which has the advantage of containing more than 530,000 deaths per annum. We then turn to monthly analysis to refine the age profiles for deaths arising from different COVID-19 variants.
2. Materials and Methods
2.1. Data Sources
Year of age all-cause and ‘caused by’ COVID–19 deaths in England and Wales is from the Office for National Statistics (ONS) [28,29,30,31]. Monthly deaths in England and Wales are from the ONS [32]. Year of age population estimates for 2011 ̶ 2020 are from the ONS [33]. Weekly deaths in the UK are from the ONS [34].
The year of age male and female population for 2021 was estimated using the method described in Section 2.3. Deaths by vaccination status are also from the ONS [35]. Annual births in England and Wales are also from the ONS [36]. Monthly data covering ‘with’ COVID-19 deaths and all-cause mortality is from the USA is from the US Centers for Disease Control and Prevention (CDC) [37]. The CDC masks all state-level deaths between 1 and 9, hence, all blanks were replaced with 1 and the likely value interpolated by the ratio of total COVID-19 deaths in the peak variant month to the month with missing values. Any value of this ratio >9 is limited to a value of 9. National deaths for the USA are not masked in this way. Analysis was mainly conducted for the larger states where very few low value data needed to be masked.
Finally, Population forecasts (including deaths) in England are from the ONS [38].
2.2. Date at which A Death Is Reported versus Date of Occurrence
As with all countries, deaths in England and Wales are the date at which the death is registered, as opposed to occurred. Depending on the disposition of public holidays at the end/beginning of calendar years this can lead to a 0.3% to 0.5% difference in the number of registered deaths. The difference between years from deaths occurred carried forward at the beginning and end of the year is usually reasonably well matched [39]. As long as the method remains unchanged such differences do not have a material impact on this study. The effect on monthly data is likewise minimized due to the data being summed over the multiple months during which each SARS-CoV-2 variant predominated.
2.3. Estimation of Year of Age Population in 2021
The estimates of population between the decennial census’s represents a potential for bias. The census-based 2021 population was 1.1% lower than the 2018-based forecast. However, this occurs mostly among working age younger adults due to inward/outward immigration between England and Wales and the European Union’s Eastern European countries [38]. Once again this does not have a material impact on this study due to the very low COVID-19 deaths in these age groups.
Due to the 2021 Census, the release of the 2021 population estimates for England and Wales have been delayed to 2023 and were not available for this study. However, estimation based on 2020 forecast population data and 2021 deaths gives an estimate with good accuracy. The method is straightforward. Living persons of any age move forward by one year of age each year. Take this number and then subtract the number of deaths. This was performed between 2011 and 2020.
This can be called a population aging model without net migration. This will be an underestimate due to net inward immigration. Inward immigration can be inferred from the difference between the result of this method and the ONS estimates. Net inward immigration is highest in the younger age groups, hence in 2020 the population of 20-year-old males/females was estimated to be 2.7% and 2.0% higher than the simple population aging model, while that for 1-year-old males/females was 1.1% higher for both genders, etc. In 2020, the net inward migration component is less than 0.3% per annum for all ages above 32 years. The 2021 population was calculated using this net migration adjusted method. This is very similar to the ONS model.
2.4. Estimation of Year of Age Baseline for Deaths in England and Wales in 2019
Actuarial forecasts for future deaths are notoriously unreliable, See Supplementary material S1 [16,38,40,41], mainly due to high uncertainty in the future mortality rate. These annual numbers show high volatility (as shown in S1) largely due to winter infectious outbreaks [40,41], which in the Northern Hemisphere span two calendar years.
Most government statistical agencies use the average of the previous five years to estimate the baseline [31], which from S1 can be seen to be subject to bias depending on whether deaths are falling (through to 2011 in S1) or rising (after 2011 in S1). The falling/rising patterns in death depend on past demographic shifts (mainly births and immigration) and are particular to each country.
The year 2019 represents the pre-COVID-19 baseline for mortality. The raw deaths in 2019 for England and Wales are available [28,29], however, they are subject to both Poisson-based and systematic variation due largely to infectious outbreaks [40,41], and the occasional prolonged heatwave or prolonged excessive cold – note that the emphasis is on prolonged, or above and beyond a normal summer/winter [42,43]. Each source of volatility will have its own age profile. It is therefore necessary to adjust the actual 2019 deaths to give an estimate the 2019 baseline which would be the case had infectious and other age-specific events been at ‘the average’ in that year. This estimation was performed as follows.
Firstly, raw deaths between 2011 ̶ 2018 (from [28]) were converted to a 2019 equivalent by multiplying by the 2019 mid-year year of age estimated population divided by that in each of the years 2011 ̶ 2018. Given that human mortality is changing over time [44], both linear and a second order polynomial regression was applied to these adjusted deaths to give the most likely average for 2019 which will be relatively free of both Poisson and systematic variation. Visual inspection showed that a second order polynomial usually gave the best estimate for the ‘average’ or ‘baseline’ deaths in 2019 – although in most instances the actual differences are minor. This is consistent with the observation that there was a break point in international mortality improvement from 2011 onward [44]. Exceptions include infants in the first year of life where population-adjusted male and female mortality was constant between 2013 and 2019.
Due to high uncertainty in the population of persons aged 90+ the ONS reports the population for 90+ as a single group [28,29]. In 2019 this includes persons born in 1929 (age 90) and before and will include a wave in population numbers due to the World War I baby boom occurring in England and Wales [35].
We note that the WW I baby boom affects some countries far less than the UK. While most statistical agencies avoid analysis of the 100+ age group we nevertheless include such analysis given the potential for birth cohort effects which is discussed later. WW I was associated with the Spanish flu pandemic [27,45], and the surviving children and newborns represent a potential group of interest.
Year of age deaths for ages above 90 were estimated based on either a second order polynomial regression of actual deaths between 2011 and 2019 or using adjustment based on the number of births [35]. These ages are complicated by the presence of the World War I baby boom (see Supplementary material S2 [28,29,40,41,45]). For adjustment based on births the number of births is substituted for the population (as above) with all years adjusted to the 2019 equivalent. Baseline deaths in 2019 were also estimated using a second order polynomial based on raw deaths between 2011 and 2019 (ages 80 to 105+) and using the average of deaths in 2018 and 2019. The rationale for this last adjustment is given in the Supplementary material S2 and S3. A large infectious event in early 2018 with an associated age profile necessitated an additional method based on the average deaths from 2017 to 2019). The final estimate for the 2019 baseline was derived from the average of the 3 or 4 different methods, depending on age, and is shown in Supplement S4.
The Poisson-based standard deviation (STDEV) associated with deaths, as a percentage value, was calculated as the square root of the number of deaths divided by the number of deaths. The variation associated with the difference between male and female deaths was calculated as the STDEV for males plus the STDEV for females – as a percentage value.
The difference between actual deaths in 2019 and the estimated baseline or ‘average’ for 2019 is shown in Figure A1 in the Appendix where in the younger ages the difference between actual and baseline is usually within ± 1 standard deviation, however, above age 70 is consistently below -1 standard deviation. Hence, it is the older age groups in which 2019 deaths are low. See S2 for explanation. Several other ages show systematic deviation, namely, males aged 41-43, 57, 63, 66-70, 73 (all low), age 74, 98 (both high), and for females aged 99 (high).
2.5. Calculation of Proportion of Deaths ‘with’ or ‘due to’ COVID-19 Variants
Inspection of the monthly raw data for England and Wales and US states shows approximately similar timing in the occurrence of deaths from the different variants. Hence, the following time periods are used to aggregate both COVID-19 deaths (numerator) and all-cause mortality (denominator): Original Wuhan strain (March 2020 to September 2020), Alpha variant (October 2020 to June 2021), Delta variant (July 2021 to February 2022), Omicron variants (March 2022 onward).
3. Results
It is very common for government statistical agencies to present detailed analysis of deaths from one calendar year to the next. We first present analysis using annual year of age data in England and Wales for 2019 (corrected as above) versus 2020 and 2021. We then move to detailed monthly analysis of the highly specific age profiles for the various SARS-CoV-2 variants and how these may impact the results seen in various calendar years. This study has a focus on all-cause mortality simply because assessing a genuine COVID-19 induced death appears to be subject to international ambiguity [16] – see Supplementary materials S5 [46,47,48,49,50,51].
3.1. Validation of deaths ‘due to’ COVID-19 and methods for adjusting deaths to a common year
This study used two primary data sources:
- Monthly deaths for all-cause and ‘due to’ COVID-19 mortality by year of age [30] – however, not split by male and female, but also covers 2022.
These two sources were used to cross-check each other, and the full detail is given in the Supplementary Materials S2–S5 [38,39,40,41,42,43,44,45,46,47,48,49,50,51].
The main conclusions are as follows:
- Adjusting deaths to the 2019-equivalent using mid-year population estimates contains hidden assumptions regarding the monthly profile for the influx of births during the World War I and II baby booms and an alternative adjustment using annual births can be utilized.
- The matching between population/births and deaths also contains assumptions around the pattern of seasonal deaths. This is not an issue in the Southern hemisphere where winter occurs in the middle of a calendar year but creates inconsistencies in the Northern hemisphere since winter spans two calendar years.
- The age profiles for excess mortality (with various adjustments) versus COVID-19 ‘due to’ deaths are roughly similar, however, deaths ‘due to’ COVID-19 appear to have been over-estimated, especially in 2021 among elderly females (with vaccination) – reflecting the well-known difficulty of attributing cause of death in elderly persons with multi-morbidities along with other factors [16]. See Supplementary Material S5.
- No discernable impact of COVID-19 upon all-cause deaths can be observed for ages 100+, or ages 1-30 either with or without adjustment – partly due to small number variation as in S2.
Regardless of the sources of variation in monthly deaths, it is concluded that the method based on the proportion (%) of monthly all-cause deaths which are assessed as either ‘with’ or ‘due to’ COVID-19 is reliable for the estimation of year of age profiles for COVID-19 deaths. This is because both total all-cause deaths and assessed COVID-19 deaths in a month or collection of months are always matched. Since it is a proportion, this method is independent of differences in population numbers between ages.
3.2. Population-Adjusted Deaths in 2020 and 2021 versus 2019
The first waves of the COVID–19 pandemic during 2020 were especially hard and made worse by the absence of any vaccine, repurposed drugs, or clinical protocols [1,2]. Figure 1 therefore illustrates this effect for males and females during both 2020 and 2021 versus the baseline deaths in 2019 - as in the Supplementary Material S2 – with 2018 included for additional comparison.
From Figure 1 the pre-COVID-19 age profile is more peaked for females and the absolute number of deaths reaches a higher maximum in females, peaking at 89 in females compared to 84 in males. Certain ages are exaggerated during COVID-19 while the two pre-COVID-19 years (2018 and the 2019 ‘adjusted’) are very close together. It should be noted that the use of 5-year or 10-year age bands would completely obscure the unique behavior at specific ages. Two sub-peaks around ages 72 and 75 are the outcome of various baby booms around World War II [36]. The age 72 sub-peak is after the war finishes while the age 75 sub-peak arose from final visits home for soldiers departing to the D-day landings. However, by far the greatest absolute number of deaths occur above age 78 which is the outcome of a mid-WW II mini-baby boom (see Figure S2.2 in the Supplementary materials), which in theory should represent the priority target population for vaccination – see further discussion later. Such nuances will be specific to England and Wales and will vary depending on the population age distribution.
As can be seen all-cause deaths in 2020 (without vaccination) were especially higher for males aged over 50 years while in females this is not the case until above age 78. In 2021 (with vaccination) the shape of the age profile changes especially above age 75 and at younger ages male deaths in 2021 are in fact higher than for 2020. The situation is far more nuanced than could simply arise from the introduction of vaccination in 2021.
Also note than in the first two years of COVID-19 there is no appreciable change in male deaths up to age 37 and up to 42 in females. For males there is a slight increase between ages 37 and 42, peaking at age 40, dropping back to no change at age 42. From age 42 through to 64, deaths in 2021 (with vaccination) are higher than in 2020 (without vaccination), while above age 75 deaths in 2021 (with vaccination) are lower than in 2020 (without vaccination). Above age 100 there does not appear to be an appreciable excess of deaths arising from COVID-19, possibly an expression of a particular immune phenotype in the oldest old [52,53]. This behavior above age 100 was confirmed in the analysis of the trends in deaths between 2011 and 2021 shown in Supplement S4.
Other year-of-age cohort effect may lie hidden in the data and for females aged 11-13 deaths are 2.2 standard deviations higher than the expected average in 2021 or 2.3 standard deviations higher for 2020 and 2021 combined. In this same age cohort male deaths are 2.1 standard deviation lower during 2020 and 2021 combined. A potential effect due to puberty will be discussed later.
3.3. Male versus Female Deaths in England and Wales
It is widely recognized that COVID–19 causes higher male mortality [5,6,7,54,55], and the effect against all-cause mortality should be strongly influenced by the higher COVID–19 mortality. This generalization is tested in Figure 2 where the difference between male and female deaths in 2020 and 2021 compared to the 2019 baseline are shown.
From Figure 2, male deaths are generally higher in 2020 (without vaccination) than for 2021 (with vaccination), which tends to suggest that males may have benefitted slightly more from vaccination, although the difference in SARS-CoV-2 variants between the two years may also play a role. Below the age of 50 (data not shown) several ages appear to have significantly greater differences, namely, in 2020 ages 7-9 males, 12 females, 33 females, 36-38 females, while in 2021 ages 7-9 females, 12 female, 11-12 female, 33 female, 36-38 female.
Once again, this behavior would be completely obscured using 5 or 10-year age bands. The differences look to be mostly systematic rather than random and hence immune/infectious history since birth may be playing a role.
The possibility that the peaks and troughs in Figure 2 are systematic rather than random is explored in Figure A2 where the difference in the mortality rate among the unvaccinated (deaths per 100 000 person years) for various age bands is explored for the three variants Alpha, Delta, and Omicron. This data comes from an ONS study relating to vaccination status [34]. In Figure A2 it is apparent that the difference between males and females is highest for the youngest age band of 18-39 years and lowest in the age 90+ group. Up to age 59 the gap between males and females is greatest for the Delta variant, however above this age it is highest for the Alpha variant. Omicron has the lowest gender gap except for age band 60-69, while for age 90+ the gender gap during Omicron reverses to females higher than males. We are clearly dealing with a complex system where age, gender, and variant interact. Judicious use of both single year of age and age banded data is required to reveal the full complexity. It is possible that year-of-birth cohorts may lie behind some of the systematic differences.
Given the time-based cumulative effects of various global and individual influences on immunity (specific patterns and timing of pathogen exposure, vaccination with various types of vaccines, sex- and age-related hormonal patterns, distribution of numerous pollutants in form of pesticides, veterinary antibiotics, radiation, food additives, among others, there is no reason that males and females of all ages should be equally affected [56]. For example, persons born after WW I (now aged up to 100 years) had to survive without antibiotics or most modern vaccines until the widespread introduction of penicillin during WW II, which those born after WW II largely benefited from. The situation is more nuanced than first appears and year of age specificity may occur as has been noted for influenza [56].
3.4. Net ‘Real-World’ Change in All-Cause Deaths by Age and Gender
Figure 3 investigates the ‘real world’ change in all-cause mortality during 2021 (with vaccination + Alpha/Delta variants) compared with 2020 (without vaccination + mainly original Wuhan strain and some Alpha at the very end of 2020). The term ‘real world’ is used as opposed to the estimated excess mortality either ‘with’ or supposedly ‘due to’ COVID-19. Persons below the age of 19 are generally not vaccinated during 2021 – hence the age cutoff in Figure 3.
Above age 65 males look to have benefited slightly more so than females (as in Figure 2). Above age 70 all changes show high significance (due to high numbers), while below age 70 changes can be obscured by Poisson randomness. However, the overall trend which is illustrated by the dotted orange line for females is clearly toward higher deaths in the younger ages. This is also clearly seen in Figure 1. While the lower deaths in 2021 above age 65 may be due to vaccination, it is difficult to attribute vaccination as the sole cause of generally higher overall deaths below age 65. More complex issues are involved.
3.5. The Delta Variant, Second Half of 2021, Targeted Younger Ages
A complicating factor in any COVID–19 analysis is the change in variants over time. We are not aware of any studies looking at the detailed age profile of deaths for COVID-19 variants, however, Figure 4 summarizes the average/median age at death for males and females assessed as ‘due to’ COVID–19.
In Figure 4 the median is always higher than the average indicating a skewed distribution. Males die at a lower median and average age. The gap between both average and median age at death for males versus females is around 4 years during the original and the Alpha variant, around 3 years during Delta, and around 2 years during Omicron. Females have a slightly higher gap of 0.4 years (average) between the median and average. The average age at death may slightly increases at the peak of an outbreak, possibly due to additional transmission into care and nursing homes.
Given that the Delta variant originated in India with a majority younger population it is unsurprising that this variant was associated with a shift to COVID–19 deaths at younger ages. Seemingly average age at death follows profiles which are seasonal (summer minimum), strain and outbreak magnitude (peak outbreak maxima) dependent. Seasonal factors are involved in gene expression [57,58]. Figure 4 also allows the transitions between SARS-CoV-2 variants to be visualized. The transitions last several months as the deaths arising from the former variant are replaced by the newer one.
The abrupt changes in the average and median age in Figure 4 strongly suggest that it is the variant rather than vaccination or changes in clinical protocols or drugs which is the primary regulator of the age profile. Vaccination likely modifies the magnitude of the effect of each variant rather than the age profile per se.
The above suggests that variant-specific analysis is required. While changes in the average in Figure 4 highly suggest changes in the underlying age profile the detail can be explored using the proportion of total deaths at each age which are assessed as due to COVID-19. To this end Figure 5 presents a detailed analysis in which the trends have been normalized such that all have the same nominal maximum value.
The results presented in Figure 5 come from a process of evaluating the age profile of COVID-19 deaths in each month from March 2020 to October 2022. This analysis required is because there is a lag between arrival of the variant, subsequent spread and then an infectious outbreak followed by deaths. In general, the age profile shows mixed features during a 2-month transition period – see Table 1. The lines in Figure 5 are therefore restricted to data in the non-transition months, which is summed and turned into an overall proportion. Hence, March 2022 to October 2022 for Omicron, etc. The term ‘Variant arrives’ indicates the point at which infections commence to the point of being an outbreak.
In Figure 5 the trends have been scaled to the age at the maximum proportion observed for ages above 40 years. Due to variability in the Delta data this was scaled to the average of age 33 to 45 which represented the range for maximum deaths. Scaling factors to give 100% for the maximum are: 5.3 for the original strain w/o vaccine; 4.6 for Alpha; 11.1 for Delta; 17.8 for Omicron variants. The higher the scaling factor the lower the overall mortality. Due to limited data in some of the younger ages, above age 0 and below age 25, data are grouped by 5-year age band. While there was a succession of variants during Omicron there was no evidence for a fundamental change in the age profile.
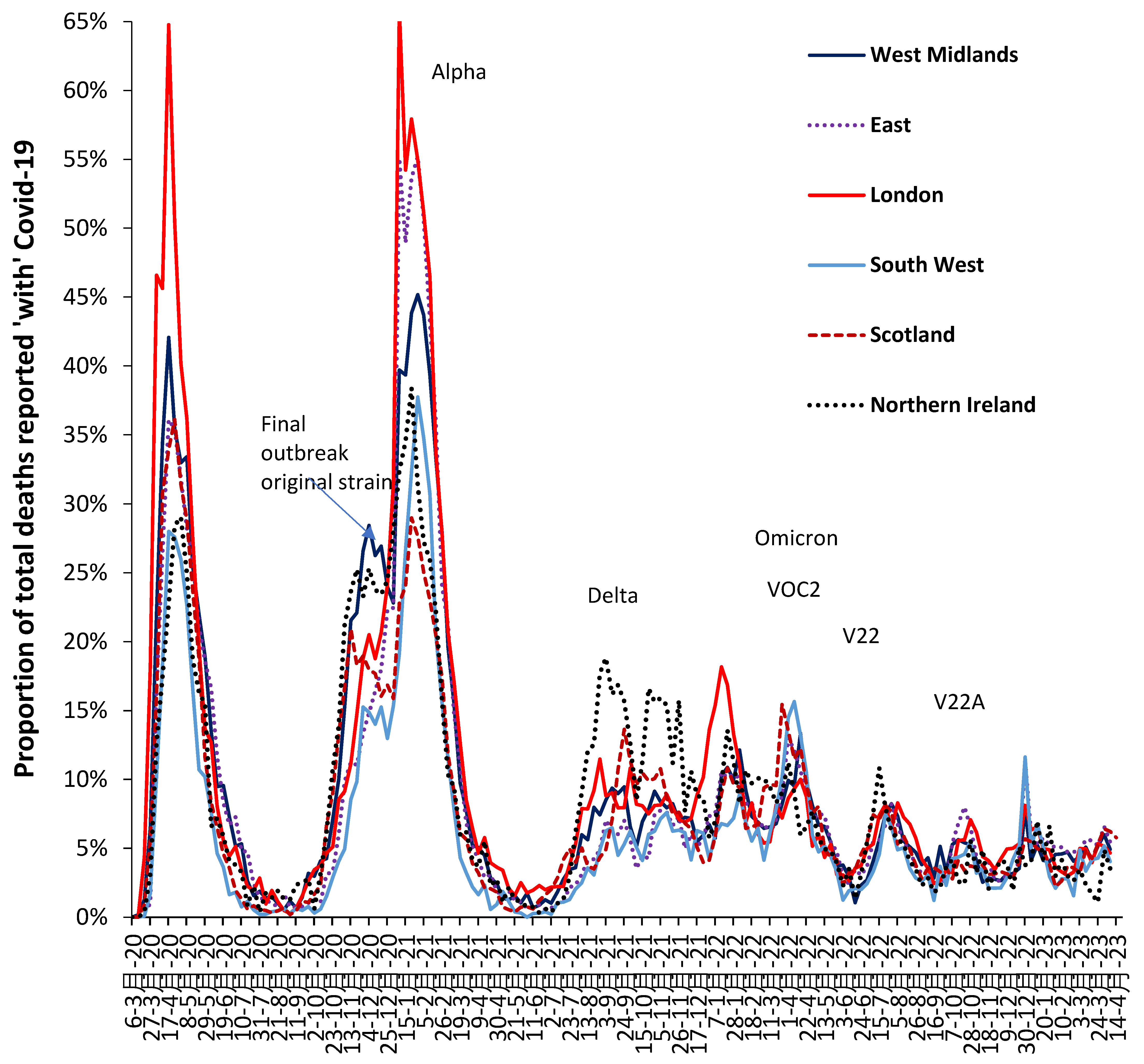
Further details regarding the relative impact of each variant upon the all-age proportion of total deaths is given in Figure A4 (appendix) along with an indication of the time-dependent spread of SARS-CoV-2 variants across the UK where London is usually worst affected, and that the Omicron variant affected London before the rest of the UK. Hence the need for a two-month window between each variant in Table 1 which also allows for the spread between the regions of England and Wales.
Figure A4 likewise shows that the Delta variant marked a transition to endemic behavior for deaths, which also continued in the Omicron variant.
The most important point is the divergent age profile for the Delta variant. The population of India contains a far higher proportion of younger people than that of an ageing UK population. Unsurprisingly Delta is optimized for younger ages with highest values of death between ages 30 ̶ 65. It also has the highest proportion of deaths for infants under the age of 1 year. The Delta profile (Aug-21 to Dec-21) is modified by month of year factors and shows a higher proportion of age 65+ deaths compared to the Omicron variant months (Mar-22 to Oct-22).
At the other extreme the age profile for the lower mortality Omicron variants (which seemingly originated in South Africa) has a broad slightly elevated lower proportion below age 55 and then peaks around age 100 – the highest for any variant. The diversity of year of age profiles between variants supports or claim that the use of wide age-bands is not helpful when attempting to compare results between variants. These findings also concur with our previous age-specific analysis showing high age variability for various influenza outbreaks [56]. Issues regarding the age profile of vaccine effectiveness will be covered in the Discussion. The result is that the period of any study (in different countries/regions) has a material effect upon its apparent outcomes.
3.6. Modification of the Age Profile in Different Countries/States/Areas
Each country/state/area has its own unique mix of risk factors for COVID-19 death in each age. Hence proportion of the population in high exposure occupations, with obesity and other clinical risk factors including genetic factors, risk of transmission due to city/rural or risk-taking behavior, etc. [19,20,21,47,48,49,50,59,60,61,62,63,64,65,66,67,68,69,70,71]. This is illustrated in Figure 6 using data from the USA rather than England and Wales and includes both males and females. Note that the data from the USA is for death ‘with’ rather than ‘due to’ COVID-19, and that only 10-year wide age banded data is available above age 1-4 [37]. The red and purple lines give the proportion of total all-cause deaths which occur in each age band.
As can be seen from Figure 6 the age profile for the USA is slightly different to that in England and Wales in Figure 5, although the differences are magnified using 10-year age bands in the US data and that 85+ is the final age band. This upper age truncation pushes the profile to the left. The US data has not been scaled as in Figure 5. By way of comparison Figure 6 shows the proportion of total deaths ‘with’ COVID-19 for both males and females. Recall that the profile of all-cause deaths in males and females is very different, as illustrated in Figure 1 and that cause of death has different profiles between the two genders. However, Figure 6 powerfully illustrates that each COVID-19 variant has an age specific profile unique to each gender as was also observed in Figure 2 and Figure 3 for England and Wales. Note that impact on total deaths (right hand axis) is not greatly shifted by the Delta or other variants since age 45-54 only accounts for 6% of total deaths, etc. It appears that the general observation of higher male deaths during COVID-19 is both age, sex and variant specific. A single year of age investigation would likely yield greater detail as was shown in Figure 1.
Hence, despite skewing due to the use of age bands in the USA, the shape of the profile is broadly similar to that seen in England and Wales for all four variants suggesting that the primary year of age profiles are determined by the variant and then modified by location-specific factors.
It must be highlighted that the evident transition to childhood COVID-19 mortality for Delta and Omicron seen in Figure 5 and Figure 6 is a strong argument for single year of age studies. The age bands in Figure 6 are likely concealing such behavior. The precise year of age detail is required to inform which public health interventions should be favored, such as acquired immunity versus vaccination. Given the very limited number of childhood deaths beyond the first year of life aggregated data from multiple countries is required. Figure 5 reverted to age bands in childhood because England and Wales had too few deaths to investigate in more detail.
Given that the USA is so large each of the states could effectively be different countries. Hence analysis at state level is warranted [71]. This is illustrated in Figure 7 using data from American states and the age profile for the original Wuhan strain which predominated for most of 2020.
Figure 7 shows the magnitude of the Wuhan strain outbreak(s) in each state with New York city having the highest magnitude of impact, followed by New Jersey, down to a far smaller impact in Puerto Rico (which in more recent times has become a US state). States such as Alaska and Hawaii have even lower magnitude Wuhan strain outbreaks, but the data is not shown. Due to the sheer size of the USA an initial outbreak at county level took at least 12-months to spread throughout the USA and even after 12-months some counties had not yet registered a COVID-19 death [71].
Figure 7 also indicates the slightly modified age profiles depending on state population risk characteristics. As can be seen Alaska is barely affected in the original outbreak and the age profile reflects small number randomness. Charts showing the shape of the age profile for the Alpha, Delta and Omicron variants are given in the Appendix Figures A3.1–A3.3.
In Figure A3.1 Puerto Rico has a divergent profile to other states for the Alpha variant, while in Figure A3.2 the Delta variant peaks in the younger ages, 35-44 in Hawaii and 45-54 in the other states. In Figure A3.3 for Omicron all states show increasing proportions of total deaths up to age 85+. This upper age band acts to hide the single year of age behavior revealed in Figure 5.
The shape has been subtly modified in each state due to the age band population risk characteristics of that state and possibly via differential spread through various communities within the state, i.e., popular retirement locations, cities, farming areas, etc [71].
Hence the overall conclusion remains that each strain has a unique age profile which is then modified by the mix of population risk factors present in each location at each age.
3.7. The All-Cause Mortality Peak for Each Variant
While Figures 5 to 7 represent a proportion of total all-cause deaths averaged over the period of each variant, the issue of the maximum value of the all-cause mortality during the period of each variant modifies the overall number of deaths at each age. To this end Figure 8 shows the maximum all-cause mortality rate for various age bands in the unvaccinated at the point of maximum monthly mortality rate during the Alpha, Delta, and Omicron variants. The Omicron outbreak mostly occurred during summer of 2022. The minimum all-cause mortality is the lowest value in the time series. Data in Figure 8 comes from an Office for National Statistics study regarding vaccination status [35].
Note that the Y-axis is a logarithmic scale. Starting from around 100 000 all-cause deaths per 100 000 person years at ages 90+ the mortality rate rapidly drops below 1000 deaths per 100 000 person years – a 99% reduction – for ages below 60 years.
The shape of the profile within each age band varies considerably. For example, a moderately even reduction across variants for age band 60-69, through to a rapid decline across the original and Alpha variants at age 18-39, etc. Likewise, the gap between male and female varies by variant and age band as noted in Section 3.3.
Also note that by 2022 during an outbreak of one of the high transmission-low mortality Omicron variants, the all-cause mortality rate is substantially reduced, especially in the age band 18 ̶ 39. The difference in the mortality gap between males and females reduces with each successive variant. The minimum value represents the point of baseline all-cause mortality rate. The Omicron outbreak is barely above the minimum.
These results confirm the profiles for age, gender and variant seen in the USA in Figure 6.
4. Discussion
4.1. Impact of SARS-CoV-2 Variants on Disease Severity and Mortality
The year of age profiles in this study confirm a trend to reducing mortality with the progression of variants. This is especially so for the Omicron variant [5,6,7]. For example, a study by the ONS in England regarding the mortality from Omicron versus Delta showed an 87% reduction at ages 15-59, an 86% reduction at ages 60-69, and a 55% reduction at ages 70+. The reduction in mortality was gender specific with a 75% reduction in males but only a 56% reduction in females [64]. Such differential effects on males/females are confirmed in this study.
These results are broadly consistent with the pattern of all-cause mortality in Figures 5 and A2 where a shift to younger ages is highlighted for the Delta variant. However, the year of age profiles in Figure 5 provide far greater detail especially below age 55. The elevated mortality for Omicron in the elderly, especially beyond age 70 years and peaking at age 100/101, shown in Figure 5 is obscured in most other studies due to the use of age 85+ or 90+ end points.
4.2. Local Risk Factors
At local level the unique age profile for each SARS-CoV-2 variant is likely to be the combined effect of a variety of mechanisms, namely:
- Age profiles for infection by the virus due to social networks or low risk aversion, i.e., nightclub and other large event attendance [65]
- Proportion of persons in each group in high exposure work categories, i.e., healthcare staff, taxi drivers, teachers, police, Bar- and restaurant staff, etc. [66].
- Family transmission risk factors such as household crowding [67].
Hence, the observed age profile should vary by country and region as the interactions between the above factors modify the actual year of age mortality. This was illustrated in Figures 7 and A3.1 to 3.3. using data for American states where the height of the line represents the magnitude, and the shape of the line illustrates the location-specific modification due to the #4 points above.
The fact that the mix of risk factors can vary by year of age has been largely overlooked and almost rarely measured at that degree of detail.
4.3. The Shift in Mortality to Younger Children Subsequent to the Wuhan Strain
As a result of the first year of circulation in the human population, and because of the virus adaptation to new host (the human), the virus receptor-binding sites have changed and will have increased their affinity (tropism) to various human receptors (cell surface proteins), including increased affinity to children's cell receptors. Virus replication machinery may also have adapted to some peculiarities of the young (children) cell metabolism and gene expression pattern. This could explain why in 2020 there was no mortality in children 0 - 10 years old (because their cells were not permissive for virus adhesion and replication) [30].
But high natural changeability of the virus plus the vaccination-accelerated virus changeability resulted in adaptation of some virus subpopulation(s) to replication in children's cells. In England and Wales, the first ‘due to’ COVID–19 deaths in infants age under 1 does not occur until August 2021, October for age 1, November for age 2, and December for age 3 which is in the wave front of the Delta variant outbreak [30]. Age 5 is not until May 2022 [30], i.e., month of death increases in order of increasing age.
4.4. Year-of-Birth Cohort Effects
Throughout the study we have hinted to the existence of year-of-birth cohort effects (Figures 1, 2, 3, etc.) which leads to specific ages (more correctly year of birth) where there is clustering of high/low behavior. We went to some lengths in Supplements S2 to S4 to establish a reliable year of age profile for 2019 (the before COVID-19 baseline) to give some assurance that such birth cohorts were real. Some variation in apparent magnitude will occur due to Poisson variation as per Figure A1. Birth cohorts will arise from the exposure of infants to a variety of pathogens plus additional environmental effects [27]. Year-of-birth cohort effects are a well-recognized phenomena in disease expression [72,73,74,75], and it would be extremely surprising if they were absent during the COVID-19 pandemic.
As far as we are aware there are no other studies investigating birth cohort effects upon infection with, or the severity of, COVID-19 disease. Once again, such studies can only be conducted with year of age data.
Identification of birth cohorts is important for two reasons:
- Which birth cohorts show enhanced resistance to COVID-19 infection and death.
- Which birth cohorts show enhanced response to COVID-19 vaccination.
We point out that the WW I birth cohort either survived the 1918-1919 Spanish flu pandemic or were born to parents who had survived the pandemic. Our analysis shows that this cohort seemed to experience little if any increase in deaths during the COVID-19 pandemic. Genetic material retained in biobanks may shed light on the issue. Further studies on the topic of birth cohort effects are highly recommended.
4.5. Puberty and Excess Female Deaths
On the other hand, some infections and conditions are specifically age related [76,77]. For example, the Japanese surveillance system for infectious diseases was used in the analysis of seven viral and four bacterial infectious diseases [76]. The male-to-female morbidity (MFM) ratios in different age groups were estimated. MFM ratios were >1 for five viral infections out of seven in childhood, i.e., male children were more frequently reported as infected than females with pharyngoconjunctival fever, herpangina, hand-foot-and-mouth disease, mumps, and varicella. More males were also reported to be infected with erythema infectiosum (parvovirus B19) and exanthema subitem (roseola), but only in children 1 year of age. By contrast, in adulthood the MFM ratios decreased to <1 for all of the viral infections above except varicella, i.e., adult women were more frequently reported to be infected than men. Sex- and age-related differences in reported morbidity were also documented for bacterial infections. Reported morbidity for enterohemorrhagic Escherichia coli infection was higher in adult females and females were reportedly more infected with mycoplasma pneumonia than males in all age groups up to 70 years [76]. Such factors will lie behind the age-gender differences seen in this study.
Another systematic analysis identified 142 datasets with information on severity of disease by age for 32 different infectious diseases, 19 viral and 13 bacterial [77]. It was commonly seen that for almost all infections, school-age children have the least severe disease, and severity starts to rise long before old age. For many infections even young adults have more severe disease than children, and dengue was the only infection that was most severe in school-age children. Together with data on vaccine response in children and young adults, the findings suggest peak immune function is reached around 5–14 years of age. Relative immune senescence may begin much earlier than assumed, before accelerating in older age groups. They suggest implications for understanding resilience to infection, optimal vaccine scheduling, and appropriate health protection policies across the life course [77].
A cluster of excess female all-cause deaths aged 10 – 13 was noted in Section 3.2. This was matched by a deficit of male deaths at this age which suggests a common link to puberty. During puberty females develop heightened immune reactogenicity [78,79,80,81], which is accompanied by a very high number of epigenetic changes (7-times more than boys) in mainly high affinity estrogen response genes involved in immune and inflammatory functions [78]. Similar hormonal effects occur during puberty in males [78,79,80,81]. On this occasion such hormonal effects during puberty appeared to protect adolescent males but disadvantaged females.
This leads to the interesting possibility that age-matched females for pre-, during-, and post-menopause may experience modified age profiles and/or different mortality rates ‘due to’ COVID-19.
We infer that human year of age susceptibility to pathogens outlined above must be interacting with the year of age profiles for death of the COVID-19 variants, and other pathogens. Such interactions are implied in a recent study relating to pathogen interference [56]. Such issues will now be discussed in greater detail.
4.6. Structure-Function and Other Aspects of Variant Age Dependence
In this section we present a framework to interpret how various factors interact to determine the final age profile and then examine several important aspects within this framework.
SARS-CoV-2 variants arise from the well-recognized virus mutation process [8,9,10,11,12,82,83,84]. Millions of random mutations are thrust out into the world population and its various people groups. Most mutations fail to thrive, but those that do can further ‘optimize’ their genetic make-up in a manner specific to the people group and/or environment where they have become entrenched. Such genetic make-up may extend beyond that regulating the surface of the virus – although wider genetic aspects are poorly studied [84].
Virus infectious process initiation and outcomes depends on multiple conditions:
- Biological properties, peculiarities of the virus species.
-
Biological properties, peculiarities of a potential host.
- strength of innate and adaptive immunity against specific pathogens
- presence and distribution (localization) of appropriate cell receptors for initial adsorption of virus particles
- presence and activity of cellular components necessary for virus’ cell entry and virus genome release, i.e., function of pinocytosis, the presence and activity of specific proteolytic enzymes in cytoplasm, etc.
- presence and activity of specific cellular RNA- or DNA-replication pathways
- Environmental conditions.
Therefore, to find reasons of age specificity of an infection, i.e., conditions when #1 and #3 are compatible, we should try to find age-related differences in #2. Hence to find age-related differences in immunity, protein composition and RNA/DNA replication machinery of host organisms. For this purpose, it is worthy to search for age-related gene expression patterns (peculiarities) in host organism and to consider possible interrelations between such differences and virus replication activity.
There are numerous research data demonstrating age-related differences in gene expression of various cellular components which can be responsible for age-related variations in virus adsorption, entry, virus RNA-release, virus RNA replication, offspring virus particles assembly and shedding [85,86]. Certainly, there are gene expression-related reasons of age-related variations in immunity [87].
For example, significant changes in the immune system during puberty period due to hormonal rearrangement [78,79,80,81] – as discussed above. This may cause enhanced susceptibility to some infections of children in puberty period and explain age-and sex-related differences in a disease incidence in population [76,77].
In modern conditions of active, massive anthropogenic biological interventions (multiple vaccinations, change of food components, etc.) there are potential reasons for age specificity of some infections. For example, extensive vaccinations with various vaccines of specific age groups may provide them increased protection against same or different pathogens – as observed in previous BCG vaccination on susceptibility to COVID-19 infection and mortality [88,89].
The situation regarding #1 and how specific mutations can confer age-specificity depends on both #2 and #3. However, this is a vastly neglected area of study, and several possibilities will now be discussed.
4.6.1. Roles for Immune Priming in Age Specificity
Both influenza(s) and α and β coronavirus(s), of which the α types are HCoV-NL63 and HCoV-229E, while β are HCoV-OC43 and HCoV- HKU1, have been infecting humans for many thousands of years [90]. However, SARS-CoV-2 is sufficiently different from previous coronaviruses to create a unique set of immune issues. Such immune issues can be illustrated from influenza.
In the case of influenza, the age specificity of different clades (variants) is greatly influenced by the phenomenon of ‘antigenic original sin’ [91,92,93], or more recently called ‘antigenic priming’ [94]. Hence previous (usually in childhood – called a birth cohort) exposure to one clade will generally invoke a response to the first clade when exposed to an antigenically different one. This response magnifies the disease in that birth cohort because the ‘wrong’ antigen is produced. However, when the new clade is sufficiently similar to the former clade the response is helpful because both B- and T-cells have already been trained and the time taken in the training process can be avoided resulting in a much faster immune response [92,93,94]. On this occasion natural immunity can be more effective than vaccination which can be unhelpful if the antigenic mix in the vaccine is dissimilar to the circulating strains.
This process was amply demonstrated during the 2009 Swine flu (H1N1) pandemic where persons aged over 65, i.e., born before 1944, had been exposed to an antigenically similar clade during the 1918-1919 Spanish flu pandemic which continued to circulate through to 1957 [95,96,97,98]. In the USA some 65% of those aged over 65 had existing immunity [95]. On this occasion age 65+ is a commonly used age band in influenza vaccine effectiveness studies and hence based on the 1957 end point for the post 1918 pandemic H1N1 variants, persons aged 52+ would have pre-existing immunity, but probably not above age 91+. Hence mortality was skewed to the younger ages who had no previous exposure [95,96,97,98]. However, severity was seemingly higher in the Americas than in Australia, New Zealand and Europe [94,95] possible due to regional differences in the prevalence of the 1A.1.1.3 and 1B.2.1 genetic clades [99].
A similar skew to younger ages occurred in the 1918-1919 Spanish flu pandemic where mortality in Canada and the USA peaked at the exact age of 28, because 28-year-olds had been previously exposed to the antigenically dissimilar 1889-1890 Russian flu pandemic [27]. The Russian flu itself had a unique timeline of international spread [100] and so an exact age of 28 will slightly vary between countries.
The situation is slightly more complex since repeat influenza infections/vaccinations leave their own antibody landscapes [101], although childhood exposure remains a dominant force [102]. Hence, immune priming is part of far wider antigenic landscapes which develop over time from repeated infection with variants of the same virus. Vaccination seemingly activates antibodies to all previous variants encountered by the individual, irrespective of antigenic distance, although the primary strain encountered in early life shows the highest antibody response [102]. Antigenic priming is a more nuanced and ongoing process.
In the case of coronaviruses none had made the huge antigenic leap as occurred in SARS-CoV-2. Among the coronaviruses HCoV-NL63 and HCoV-OC43 infections are common in infancy and look to confer some protection against subsequent HCoV-229E and HCoV-HKU1 [90].
Such birth cohorts were first observed in influenza [91] but apply to any virus which has different strains and variants [92,93,94]. Hence, exposure of infants to antigenically similar strains of other α- and β-coronaviruses – most likely those causing the common cold [90]. This directly applies to SARS-CoV-2 infection but in a more limited way [95], and to COVID-19 vaccination [92].
.It is also highly likely that the process of antigenic priming will begin to affect emerging variants following initial exposure to the Wuhan or subsequent strains. Acquired natural immunity (from any variant) has been shown to confer protection against the subsequent effects of further COVID-19 infection [103,104,105,106,107,108]. The extent of exposure to each variant is highly dependent on location as shown in Figures 7, A3.1 to A3.3.
Hence regarding the potential role of antigenic priming upon the year of age profiles of SARS-CoV-2 variants we conclude that this virus has not had the very long-term interaction with the human population as seen in influenza, and that such mechanisms are as only beginning to play a central role in the expression of the age profile – although in the sense that antigenic priming only acts as yet another factor to modify the intrinsic age profile of the particular (future) variants which would otherwise be expressed in a ‘blank sheet’ population.
4.6.2. Roles for Pathogen Interference
COVID-19 infection does not operate in supreme isolation from other pathogens [56,109,110,111] and the interaction between pathogens via the effects against small noncoding RNAs (miRNAs) may contribute to year-of-age effects – under the likelihood that each SARS-CoV-2 variant can instigate differential patterns of miRNA production [112,113,114,115,116,117,118,119]. In cancer patients both COVID-19 and other pathogens intersect at NAMPT/NAD metabolism which can direct both innate immune cell effector functions and the homeostatic robustness, in both cancer and infection [120].
In a previous study we emphasized the role of small non-coding RNAs (miRNAs) in gene expression and the processes of ‘pathogen interference’ [56]. Among a multitude of functions miRNAs are central to metabolic regulation [112,121]. Cells produce a multitude of miRNAs when infected with COVID-19 [112,113,114], and COVID-19 also has several miRNAs which are coded into its genome [115,116,117,118,119].
It is important to note that all major COVID-19 risk factors are associated with dysregulated miRNA profiles, hence in obesity [122], diabetes [122,123], heart disease [124], and cancer [120,121,125]. A recent study has suggested that miRNAs are involved in unusual all-cause mortality outcomes following COVID-19 vaccination [126]
Relatively stable extracellular miRNAs are present in most biological fluids. These circulating miRNAs are transported by membrane-derived vesicles (exosomes and microparticles), lipoproteins, and other ribonucleoprotein complexes [127]. Such miRNAs are selectively exported from cells with distinct signatures that have been found to be altered in many pathophysiology’s, as observed above. Functional miRNAs are delivered to recipient cells by various routes. Transferred miRNAs use cellular machinery to reduce target gene expression and alter cellular phenotype. miRNAs mediate cell-to-cell communication linking disparate cell types, diverse biological mechanisms, and homeostatic pathways [127]. Although significant advances have been made, miRNA intercellular communication is full of complexities and many questions remain [127]. However, such selective export of miRNAs seems to explain why different studies have detected a different range in miRNAs following COVID-19 infection.
The processes of aging are evident in human miRNA production. For example, miRNA 92a declines with age in CD3+CD8+CD62L+ cells and CD8+ T-lymphocytes. This suggests that the age-related attrition of human naïve T cells could be connected to a reduced miRNA-92a in T-lymphocytes and downregulation of the miRNA-92a level might indicate exhaustion of naïve T-cells due to alteration of the immunologic condition with aging, and hence in vaccine response [128]. How the year of age profiles for each variant will modify the range of miRNAs arising from COVID-19 infection remains an unexplored area. Sites specific to miRNA production appear to reside on the X-chromosome [129] which may explain the higher male mortality from COVID-19 and is related to wider genetic polymorphs associated with higher COVID-19 risk [130,131,132].
These factors may interact with the age specificity of SARS-CoV-2 variants, but in yet unknown ways.
4.6.3. Viral Entry as A Molecular Signal Event
The very fact that the SARS-CoV-2 variants have recognizable changes in the spike antigen [133,134], responsible for binding to the cell membrane, provides a clue to where the molecular processes behind age specificity reside. Several potential mechanisms are available, namely, hydrogen bond and hydrophobic interactions between the variants and the ACE-2 receptor sites [133,134,135,136], and previously unexplained variations in the small noncoding RNAs (miRNAs) produced in response to COVID-19 infection which then alter gene expression.
It has been recognized for many years that the production of miRNAs occurs in response to ‘environmental cues’ leading to systemic signaling [137]. Exactly how this happens has never been elucidated, i.e., we can observe miRNAs appearing in a cell, but we have no knowledge of how that pattern of miRNAs was selected.
Following infection many viruses produce fusogen proteins which then lead to further fusion of infected cells with nearby uninfected cells [138,139]. This includes members of the coronavirus family including COVID-19. This leads to the formation of giant cells called syncytia mostly from cell-cell fusion of alveolar epithelial cells and from cell-cell fusion of infected alveolar macrophages [139].
The action of fusogens leads to lipid rearrangement in the membrane [139]. We can assume that viral fusion and entry triggers a host of signaling events which lead to miRNA production. The molecular process called ’pathogen interference’ is one among many observable outcomes of the action of miRNAs [56]. In this instance the miRNAs then regulate the production of various interferons which in turn act to limit or enhance infection by a second pathogen [140,141].
We propose that it is the spectrum of miRNAs released following entry into a cell which is the primary basis for the observed age profiles of different SARS-Cov_2 variants.
4.7. Personal Risk versus Population Risk
We have previously emphasized the role of genetic risk and the substantial variation in human immunity regarding ‘personal’ risk as opposed to population risk [56]. Several loci for genetic risk due to COVID-19 have likewise been identified [142,143,144,145] and the expression of genetic risk is age-dependent [142,143,144,145]. Likewise, there is no single antibody against COVID-19, but rather humans produce a wide variety of antibodies, some of which are more effective than others [146]. Individuals experiencing sequelae after acute COVID-19 infection experience an aberrant immune response which is revealed by subsequent vaccination [147]. Once again it is unknown how such factors interact with the year of age profiles for SARS-CoV-2 variants.
4.8. Limitations of the Study
The study only covers two high income countries for variants which were common in England and the USA. It is unknown how vaccination based on the original strain affected the age profiles in 2021 and beyond. This could be resolved by comparing the proportion of total deaths due to COVID-19 in the vaccinated and unvaccinated groups. The inability to accurately assess ‘due to’ COVID-19 death remains an international problem, however, it should not unduly affect the age profile.
4.9. Wider Application
Given the fact that the age profile for COVID-19 deaths has central implications as to which ages should be vaccinated it is recommended that all countries report the year of age proportion of weekly deaths which are either ‘with’ or ‘due to’ COVID-19. This data can be interim and then updated later. The WHO may be a logical organization to conduct this analysis. In this manner emerging shifts in the age profile can be identified early rather than later. Speed rather than accuracy is of the essence. The public can then be warned to take appropriate voluntary age-specific measures and Public Health agencies can modify vaccination campaigns accordingly.
5. Conclusions
This study has established that each SARS-CoV-2 variant has an intrinsic year of age profile relating to death, which is then modified by population characteristics such as the gender, proportion of persons at each age with high occupational, clinical, or genetic risk as demonstrated by differences between US states.
Similar unique age profiles for death have been reported for influenza clades [56]. We are unaware that any studies have been directed at the question as to whether the difference in the age profile for the variants upon which vaccines are based and the prevailing variants have a major effect upon vaccine effectiveness (VE). We note that VE for influenza vaccines can occasionally go negative [56]. This has been assumed to be due to ‘antigenic distance’ between the circulating clade and the administered vaccine [56], which may also include aspects of the differences in age profiles [56]. It is highly likely that birth cohort effects are present during COVID-19 induced mortality.
The high ambiguity in establishing if a death was directly caused by COVID-19 implies that establishing a year of age baseline for 2019 is vitally important. We recommend wider studies on the topic of age specificity for COVID-19 variants with data originating from different countries having markedly different population age structures and levels of poverty, along with greater exposure to some of the other variants which were not common in the UK and USA. We have proposed that small noncoding RNAs may regulate the age profiles for COVID-19 variants, and this requires further investigation. Indeed, the sudden transition of COVID-19 outbreaks from seasonal to endemic (as per Figure A4) for both the Delta and Omicron variants requires detailed examination of potential changes in miRNA profiles using blood stored for research.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Section S1: Actual deaths versus forecast deaths in England (2001-2061); Section S2: Methods for adjusting deaths to the 2019 baseline value; Section S3: Adjustment factors based on births to give 2019-equivalent estimate; Section S4: Results for the alternative methods to calculate the 2019 ‘average’ number of deaths; Section S5: Deaths assessed as ‘due to’ COVID-19 in 2020 and 2021 compared to excess deaths relative to 2019.
Author Contributions
Conceptualization, R.P.J.; methodology, R.P.J.; validation, R.P.J.; formal analysis, R.P.J.; investigation, R.P.J.; resources, R.P.J.; data curation, R.P.J.; writing—original draft preparation, R.P.J.; writing—review and editing, R.P.J. and A.P.; visualization, R.P.J.; supervision, R.P.J.; project administration, R.P.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are publicly available, and sources are listed in the Materials and Methods section.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A: Figures A1 to A5
Figure A1.
Difference between 2019 actual and the underlying trend of deaths (the ‘average’) in 2019, by single year of age in England and Wales. STDEV = standard deviation (Poisson).
Figure A1.
Difference between 2019 actual and the underlying trend of deaths (the ‘average’) in 2019, by single year of age in England and Wales. STDEV = standard deviation (Poisson).

Figure A2.
Percentage higher male all-cause mortality rate during SARS-CoV-2 variant circulation; Alpha (January to June 2021), Delta (July to December 2021) and Omicron (January to May 2022) for persons who are unvaccinated. Analysis of data from [33]. The percent higher male mortality was calculated for each month/age range and the Y-axis shows the median value for each period. All mortality rates are age-standardized within each age band.
Figure A2.
Percentage higher male all-cause mortality rate during SARS-CoV-2 variant circulation; Alpha (January to June 2021), Delta (July to December 2021) and Omicron (January to May 2022) for persons who are unvaccinated. Analysis of data from [33]. The percent higher male mortality was calculated for each month/age range and the Y-axis shows the median value for each period. All mortality rates are age-standardized within each age band.
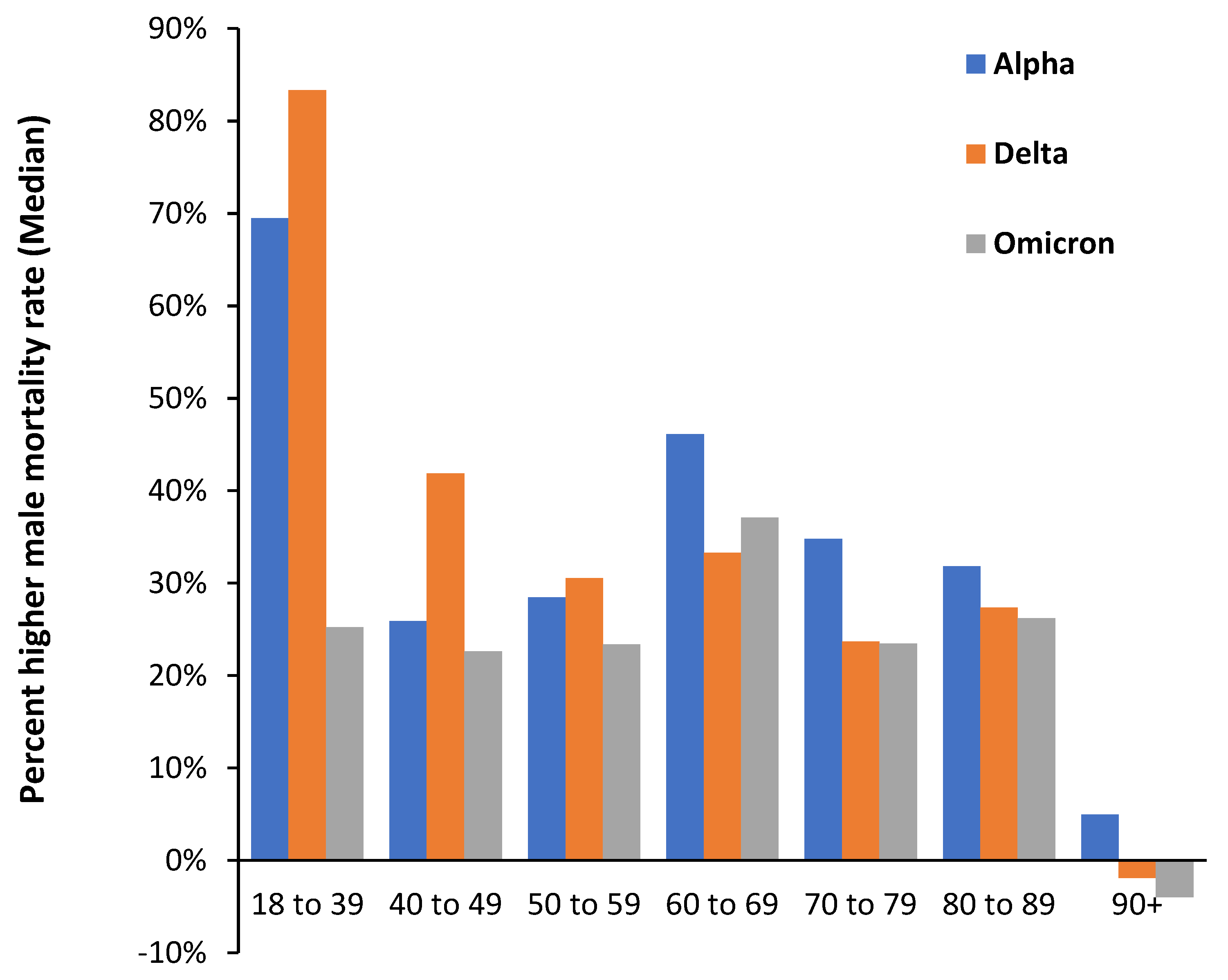
Figure A3.1.
Proportion of total deaths ‘with’ COVID-19 in a selection of American states during the period of the Alpha variant. Analysis of data from [35].
Figure A3.1.
Proportion of total deaths ‘with’ COVID-19 in a selection of American states during the period of the Alpha variant. Analysis of data from [35].
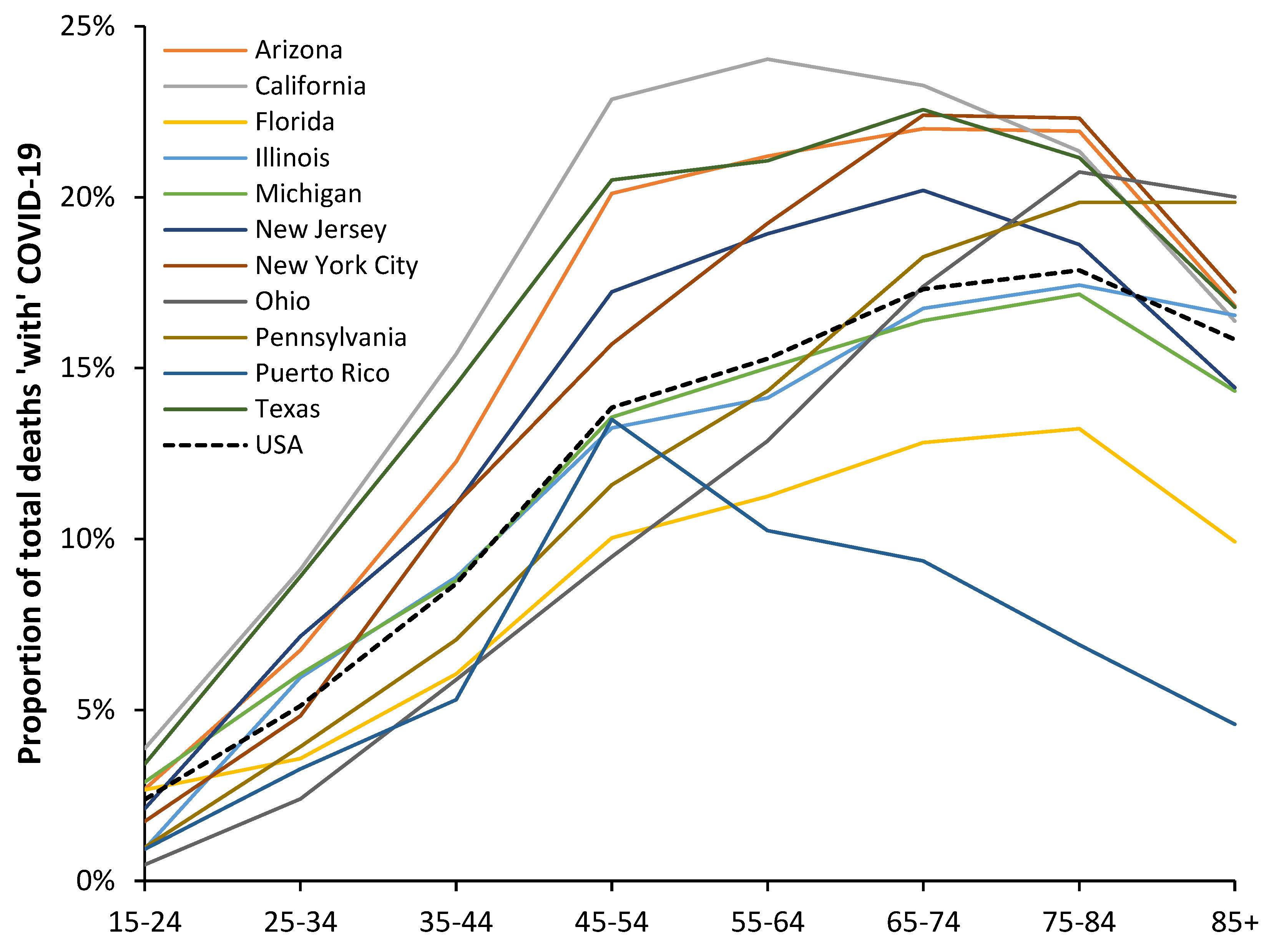
Figure A3.2.
Proportion of total deaths ‘with’ COVID-19 in a selection of American states during the period of the Delta variant. Data from [35].
Figure A3.2.
Proportion of total deaths ‘with’ COVID-19 in a selection of American states during the period of the Delta variant. Data from [35].
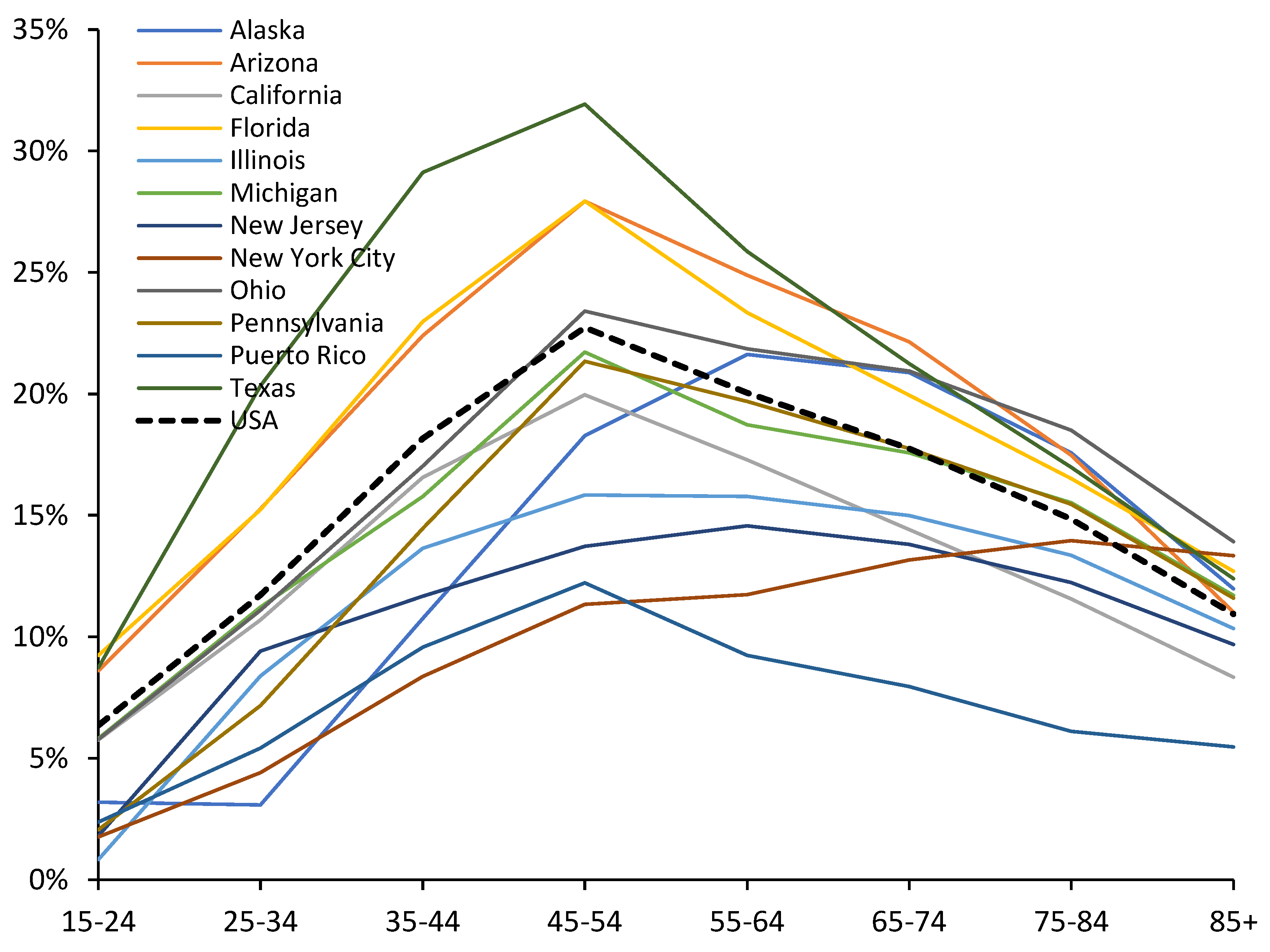
Figure A3.3.
Proportion of total deaths ‘with’ COVID-19 in a selection of American states during the period of the Omicron variant. Analysis of data from [35]. Lower deaths during Omicron imply that the data below age 55 shows higher scatter especially for the smaller states.
Figure A3.3.
Proportion of total deaths ‘with’ COVID-19 in a selection of American states during the period of the Omicron variant. Analysis of data from [35]. Lower deaths during Omicron imply that the data below age 55 shows higher scatter especially for the smaller states.
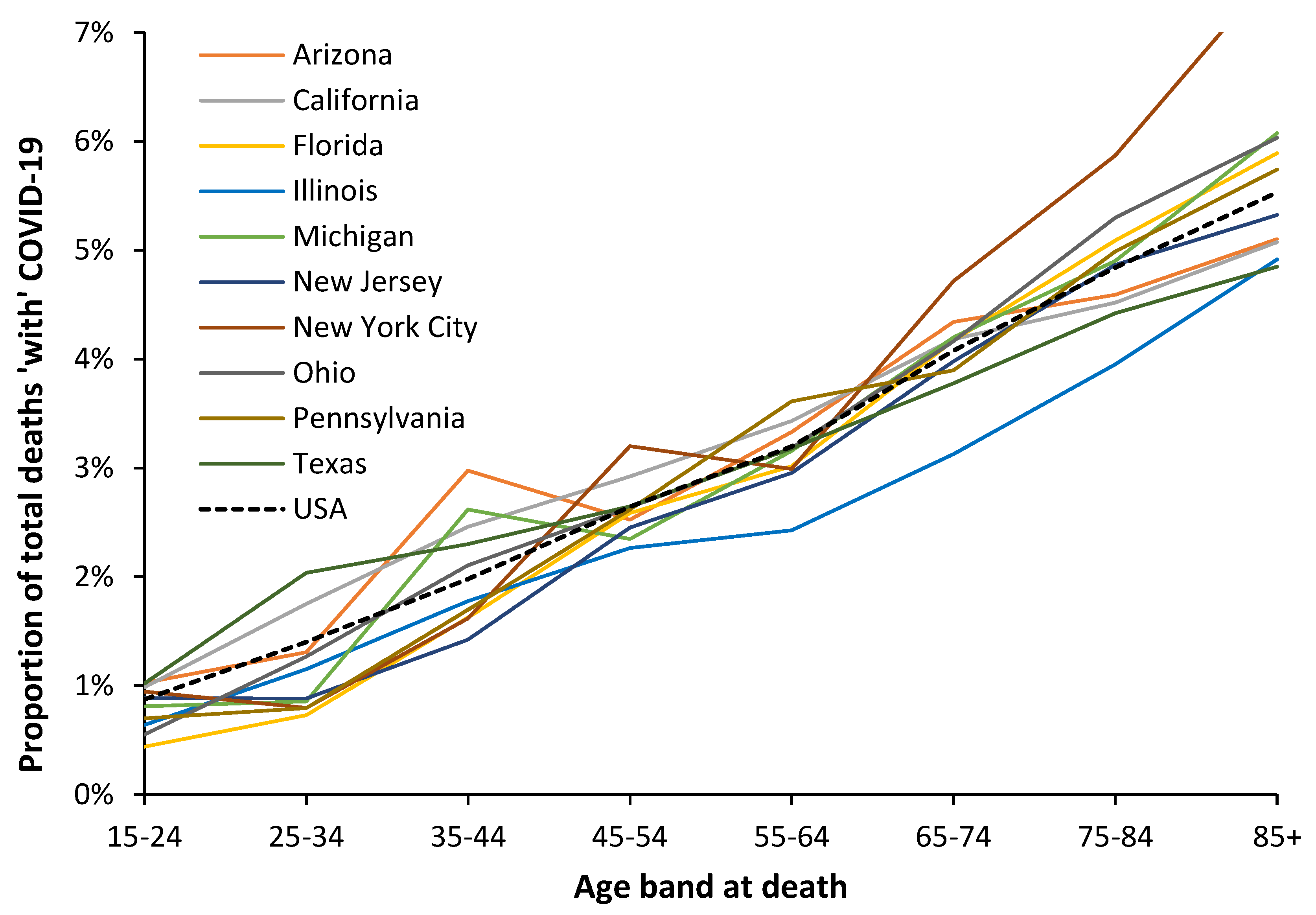
Figure A4.
The proportion of all-ages weekly deaths reported as ‘with’ COVID–19 for various parts of the UK, 6 March 2020 to 14 April 2023. Analysis of data from [32]. Due to a deficit in COVID-19 testing capacity the magnitude of the first peak may be underestimated. As per Supplement S5 the real mortality ‘caused by’ COVID-19 will be lower than the ‘with’ COVID-19 data suggests.
Figure A4.
The proportion of all-ages weekly deaths reported as ‘with’ COVID–19 for various parts of the UK, 6 March 2020 to 14 April 2023. Analysis of data from [32]. Due to a deficit in COVID-19 testing capacity the magnitude of the first peak may be underestimated. As per Supplement S5 the real mortality ‘caused by’ COVID-19 will be lower than the ‘with’ COVID-19 data suggests.
References
- Basu, D.; Chavda, V.; Mehta, A. Therapeutics for COVID-19 and post COVID-19 complications: An update. Curr. Res. Pharm. Drug Discov. 2022, 3, 100086. [Google Scholar] [CrossRef] [PubMed]
- Thakur, V.; Bhola, S.; Thakur, P.; et al. Waves and variants of SARS-CoV-2: understanding the causes and effect of the COVID-19 catastrophe. Infection 2022, 50, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Demombynes, G.; COVID-19 Age-Mortality Curves Are Flatter in Developing Countries. World Bank Group, July 2020. Available online: World Bank Document (accessed on 29 January 2023).
- Sasson, I. Age and COVID-19 mortality. Demog Res. 2021, 44, 379–396. [Google Scholar] [CrossRef]
- Adjei, S.; Hong, K.; Molinari, N.; Bull-Otterson, L.; Ajani, U.; Gundlapalli, A.; Harris, A.; Hsu, J.; Kadvi, S.; et al. Mortality Risk Among Patients Hospitalized Primarily for COVID-19 During the Omicron and Delta Variant Pandemic Periods — United States, April 2020–June 2022. MMWR. 2022, 71, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, M.; Lam, B.; Younossi, E.; Felix, S.; Ziayee, M.; Price, J.; Pham, H.; de Avila, L.; Terra, K.; Austin, P.; et al. The impact of variants and vaccination on the mortality and resource utilization of hospitalized patients with COVID–19. BMC Infect. Dis. 2022, 22, 702. [Google Scholar] [CrossRef] [PubMed]
- Leiner, J.; Pellissier, V.; Hohenstein, S.; König, S.; Schuler, E.; Möller, R.; Nachtigall, I.; Bonsignore, M.; Hindricks, G.; Kuhlen, R.; Bollmann,A. Characteristics and outcomes of COVID-19 patients during B.1.1.529 (Omicron) dominance compared to B.1.617.2 (Delta) in 89 German hospitals. BMC Infect. Dis. 2022, 22, 802. [Google Scholar] [CrossRef] [PubMed]
- Sanjuan, R.; Nebot, M.; Chirico, N.; Mansky, L.; Belshaw, R. Viral mutation rates. J. Virol. 2010, 84, 9733–9748. [Google Scholar] [CrossRef]
- Nobusawa, E.; Sato, K. Comparison of the mutation rates of human influenza A and B viruses. J. Virol. 2006, 80, 3675–8. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, H.; Wu, X.; Zhong, Y.; Zhang, K.; Zhang, Y.; Boerwinkle, E.; Fu, Y. Moderate mutation rate in the SARS coronavirus genome and its implications. BMC Evol. Biol. 2004, 4, 21. [Google Scholar] [CrossRef]
- Pathan, R.; Biswas, M.; Khandaker, M. Time series prediction of COVID-19 by mutation rate analysis using recurrent neural network-based LSTM model. Chaos Solitons Fractals. 2020, 138, 110018. [Google Scholar] [CrossRef]
- Cosar, B.; Karagulleoglu, Z.; Unal, S.; Ince, A.; Uncuoglu, D.; Tuncer, G.; Kilinc, B.; Ozkan, Y.; Ozkoc, H.; Demir, I.; et al. COVID–19 mutations and their viral variants. Cytokine Growth Factor. Rev. 2022, 63, 10–22. [Google Scholar] [CrossRef] [PubMed]
- GOV.UK. COVID–19 confirmed deaths in England (to 31 December 2020) – report. Available online: COVID–19 confirmed deaths in England (to 31 December 2020): report - GOV.UK (www.gov.uk) (accessed 29 August 2022).
- GOV.UK. Daily tests processed and testing capacity (UK): 20 March to 22 September 2020. Available online: Daily tests processed and testing capacity (UK): 20 March to 22 September 2020 - GOV.UK (www.gov.uk) (accessed 28 December 2022).
- Mueller, B.; When was the first U.S. Covid death? C.D.C. investigates 4 early cases. The New York Times, 9 September 2021. Available online: When Was the First U.S. Covid Death? CDC Investigates 4 Early Cases - The New York Times (nytimes.com) (accessed 2 September 20220).
- Ioannidis, J.; Zonta, F.; Levitt, M. Flaws and uncertainties in pandemic global excess death calculations. Eur. J. Clin. Invest. 2023, 00, e014008. [Google Scholar] [CrossRef]
- GOV.UK. Investigation of COVID–19 variants: technical briefings. Available online: Investigation of COVID–19 variants: technical briefings - GOV.UK (www.gov.uk) (accessed 26 August 2022).
- Centers for Disease Control and Prevention. COVID–19 variant classifications and definitions. Available online: COVID–19 Variant Classifications and Definitions (cdc.gov) (accessed 2 August 2022).
- Jin, R. The lag between daily reported Covid-19 cases and deaths and its relationship to age. J. Public. Health Res. 2021, 10, 2049. [Google Scholar] [CrossRef] [PubMed]
- Freeman, A.; Watson, A.; O'Regan, P.; Wysocki, O.; Burke, H.; Freitas, A.; Livingstone, R.; Dushianthan, A.; Celinski, M.; Batchelor, J.; et al. Wave comparisons of clinical characteristics and outcomes of COVID–19 admissions - Exploring the impact of treatment and strain dynamics. J. Clin. Virol. 2022, 146, 105031. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, M.; Lam, B.; Younossi, E.; Felix, S.; Ziavee, M.; Price, J.; Pham, H.; de Avila, L.; Terra, K.; Austin, P.; et al. The impact of variants and vaccination on the mortality and resource utilization of hospitalized patients with COVID-19. BMC Infect. Dis. 2022, 22, 702. [Google Scholar] [CrossRef]
- Webster, H.; Nyberg, T.; Sinnathamby, M.; Abdul Aziz, N.; Ferguson, N.; Seghezzo, G.; Blomquist, P.; Bridgen, J.; Chand, M.; Groves, N.; et al. Hospitalisation and mortality risk of SARS-COV-2 variant Omicron sub-lineage BA.2 compared to BA.1 in England. Nat. Commun. 2022, 13, 6053. [Google Scholar] [CrossRef] [PubMed]
- Wikipedia. COVID–19 vaccination in the United Kingdom. Available online: COVID–19 vaccination in the United Kingdom - Wikipedia (accessed 26 August 2022).
- GOV.UK. COVID vaccinations in United Kingdom. Available online: Vaccinations in England | Coronavirus in the UK (data.gov.uk) (accessed 26 August 2022).
- Majeed, A.; Pollock, K.; Hodes, S.; Papaluca, M. Implementation of COVID–19 vaccination in the United Kingdom. BMJ. 2022, 378, e070344378. [Google Scholar] [CrossRef]
- GOV.UK. JCVI advises on COVID–19 vaccine for people aged under 40. 7 May 2021. Available online: JCVI advises on COVID–19 vaccine for people aged under 40 - GOV.UK (www.gov.uk) (accessed 4 September 2022).
- Gagnon, A.; Miller, M.; Hallman, S.; Bourbeau, R.; Herring, D.; Earn, D.; Madrenas, J. Age-Specific Mortality During the 1918 Influenza Pandemic: Unravelling the Mystery of High Young Adult Mortality. PLOS ONE 2013, 8, e69586. [Google Scholar] [CrossRef]
- Office for National Statistics. Deaths registered by single year of age, UK. Available online: Deaths registered by single year of age, UK - Office for National Statistics (ons.gov.uk) (accessed 28 August 2022).
- Office for National Statistics. Deaths registered in England and Wales, 2021 edition. Available online: Deaths registered in England and Wales - Office for National Statistics (ons.gov.uk) (accessed 28 August 2022).
- Office for National Statistics. Single year of age and average age of death of people whose death was due to or involved COVID–19. Available online: Single year of age and average age of death of people whose death was due to or involved COVID–19 - Office for National Statistics (ons.gov.uk) (accessed 5 September 2022).
- Office for National Statistics. Deaths registered in England and Wales: 2021 (refreshed populations), January 2023. Available online: Deaths registered in England and Wales 2021 (refreshed populations) (1).pdf (accessed on 28 January 2023).
- Office for National Statistics. Deaths registered monthly in England and Wales. Available online: Deaths registered monthly in England and Wales - Office for National Statistics (ons.gov.uk) (accessed 5 September 2022).
- Office for National Statistics. Analysis of population estimates tool for UK. Available online: Deaths registered by single year of age, UK - Office for National Statistics (ons.gov.uk) (accessed 28 August 2022).
- Office for National Statistics. Deaths registered weekly in England and Wales, provisional. Available online: Deaths registered weekly in England and Wales, provisional - Office for National Statistics (ons.gov.uk) (accessed on 28 January 2023).
- Office for National Statistics. Deaths by vaccination status, England. Available online: Deaths by vaccination status, England - Office for National Statistics (accessed 28 August 2022).
- Office for National Statistics. Births in England and Wales: summary tables. Available online: Births in England and Wales: summary tables - Office for National Statistics (ons.gov.uk) (accessed on 10 February 2023).
- US Centers for Disease Control and Prevention. Provisional COVID-19 deaths by sex and age, updated 8 February 2023. Available online: Provisional COVID-19 Deaths by Sex and Age | Data | Centers for Disease Control and Prevention (cdc.gov) (accessed 10 February 2023).
- Office for National Statistics. Principal projection – England Summary. Available online: Principal projection - England summary - Office for National Statistics (ons.gov.uk) (accessed on 27 April 2023).
- Office for National Statistics. Mortality statistics in England and Wales QMI, July 2022. Available online: Mortality statistics in England and Wales QMI - Office for National Statistics (ons.gov.uk) (accessed on 28 January 2023).
- Jones, R. Excess winter mortality (EWM) as a dynamic forensic tool: Where, when, which conditions, gender, ethnicity, and age. Int. J. Environ. Res. Public. Health 2021, 18, 2161. [Google Scholar] [CrossRef]
- Jones, R. End of life and financial risk in GP commissioning. Brit J. Healthc. Manage 2012, 18, 374–381. [Google Scholar] [CrossRef]
- Xu, Z.; FitzGerald, G.; Guo, Y.; Jalaludin, B.; Tong, S. Impact of heatwave on mortality under different heatwave definitions: A systematic review and meta-analysis. Environ. Internat. 2016, 89-90, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Dear, K.; Wang, Z. Climate and health: mortality attributable to heat and cold. Lancet. 2015, 386, 320–322. [Google Scholar] [CrossRef] [PubMed]
- Djeundje, V.B.; Haberman, S.; Bajekal, M.; Lu, J. The slowdown in mortality improvement rates 2011–2017: a multi-country analysis. Eur. Actuar. J. 2022. [Google Scholar] [CrossRef]
- Jones, R.; Ponomarenko, A. Trends in excess winter mortality (EWM) from 1900/01 to 2019/20 – evidence for a complex system of multiple long-term trends. Int J Environ Res Public Health 2022, 19, 3407. [Google Scholar] [CrossRef] [PubMed]
- Ely, J.; Chris Whitty warns Britain faces 'prolonged period' of excess deaths NOT caused by Covid due to collateral effects of lockdown. Mail Online, 9 February 2023. Available online: Chris Whitty warns Britain faces 'prolonged period' of excess deaths | Daily Mail Online (accessed on 17 February 2023).
- Office for National Statistics. Deaths due to COVID-19, registered in England and Wales. Published on 1 July 2022. Available online: Deaths due to COVID-19, registered in England and Wales - Office for National Statistics (ons.gov.uk) (accessed on 17 February 2023).
- Stokel-Walker, C. What do we know about covid vaccines and preventing transmission? BMJ 2022, 376, o298. [Google Scholar] [CrossRef] [PubMed]
- Eyre, D.; Taylor, D.; Purver, M.; Chapman, D.; Fowler, T.; Pouwels, K.; Walker, A.; Peto, T. Effect of Covid-19 Vaccination on Transmission of Alpha and Delta Variants. N. Engl. J. Med. 2022, 386, 744–756. [Google Scholar] [CrossRef]
- Vitiello, A. , Ferrara, F. , Troiano, V. et al. COVID-19 vaccines and decreased transmission of SARS-CoV-2. Inflammopharmacol. 2021, 29, 1357–1360. [Google Scholar] [CrossRef]
- National Records Scotland, Available online: weekly-deaths-23.xlsx (live.com). Accessed on (28 April 2023).
- Luo, O. , Lei, W., Zhu, G.; Ren, Z.; Xu, Y.; Xiao, C.; Zhang, H.; Cai, J.; et al. Multidimensional single-cell analysis of human peripheral blood reveals characteristic features of the immune system landscape in aging and frailty. Nat. Aging 2022, 2, 348–364. [Google Scholar] [CrossRef]
- Ramasubramanian, R. , Meier, H.C.S., Vivek, S.; Klopack, E.; Crimmins, E.; Faul, J.; Nikolich-Zugich, J.; Thyagarajan, B. Evaluation of T-cell aging-related immune phenotypes in the context of biological aging and multimorbidity in the Health and Retirement Study. Immun. Ageing 2022, 19, 33. [Google Scholar] [CrossRef]
- Vasa, A.; Kini, M.; Neugarten, J.; Bellin, E.; Golestaneh, L. Does comorbidity burden explain the higher COVID–19 mortality risk among men? A retrospective cross-sectional analysis of a well-defined cohort of patients in Bronx, New York. BMJ Open 2022, 12, e063862. [Google Scholar] [CrossRef]
- Nguyen, N.; Chinn, J.; De Ferrante, M.; Kirby, K.; Hohmann, S.; Amin, A. Male gender is a predictor of higher mortality in hospitalized adults with COVID–19. PLoS One 2022, 16, e0254066. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; Ponomarenko, A. Roles for pathogen interference in influenza vaccination, with implications to vaccine effectiveness (VE) and attribution of influenza deaths. Infect. Dis. Rep. 2022, 14, 710–758. [Google Scholar] [CrossRef]
- Goldinger, A.; Shakhbazov, K.; Henders, A.; McRae, A.; Montgomery, G.; Powell, J. Seasonal effects on gene expression. PLoS One. 2015, 10, e0126995. [Google Scholar] [CrossRef] [PubMed]
- Dopico, X.; Evangelou, M.; Ferreira, R.; Guo, H.; Pekalski, M.; Smyth, D.; Cooper, N.; Burren, O.; Fulford, A.; Hennig, B.; et al. Widespread seasonal gene expression reveals annual differences in human immunity and physiology. Nat. Commun. 2015, 6, 7000. [Google Scholar] [CrossRef] [PubMed]
- Niemi, M.; Daly, M.; Ganna, A. The human genetic epidemiology of COVID-19. Nat Rev Genet 2022, 23, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Baranova, A.; Cao, H.; Teng, S.; Zhang, F. A phenome-wide investigation of risk factors for severe COVID-19. J Med Virol. 2022, 95, e28264. [Google Scholar] [CrossRef]
- Mani, A.; Ojha, V. Thromboembolism after COVID-19 Vaccination: A Systematic Review of Such Events in 286 Patients. Ann. Vasc. Surg. 2022, 84, 12–20. [Google Scholar] [CrossRef]
- Bilotta, C.; Perrone, G.; Adelfio, V.; Spatola, G.; Uzzo, M.; Argo, A.; Zerbo, S. COVID-19 Vaccine-Related Thrombosis: A Systematic Review and Exploratory Analysis. Front. Immunol. 2021, 12, 729251. [Google Scholar] [CrossRef]
- Office for National Statistics. Comparing the risk of death involving coronavirus (COVID-19) by variant, England: December 2021. Available online: Comparing the risk of death involving coronavirus (COVID-19) by variant, England - Office for National Statistics (ons.gov.uk) (accessed on 28 January 2023).
- Karaivanov, A. A social network model of COVID-19. PLOS ONE. 2020, 15, e0240878. [Google Scholar] [CrossRef]
- Office for National Statistics. Which occupations have the highest potential exposure to the coronavirus (COVID-19)? Published 11 May 2020. Available online: Which occupations have the highest potential exposure to the coronavirus (COVID-19)? - Office for National Statistics (Accessed on 25 February 20230.
- Villela, D. Household crowding hampers mitigating the transmission of SARS-CoV-2. Rev Soc Bras Med Trop. 2021, 54, e08212020. [Google Scholar] [CrossRef]
- Hu, J.; Wang, Y. The Clinical Characteristics and Risk Factors of Severe COVID-19. Gerontology 2021, 67, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Chen, X.; Wang, C.; Zhang, S.; Yu, R.; Xie, Y.; Yuan, S.; Cheng, L.; Shi, L.; Zhang, X. An integrated strategy to identify COVID-19 causal genes and characteristics represented by LRRC37A2. J Med Virol. 2023, 95, e28585. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.; Hernandez, K.; Aronow, B.; Wurtele, E. African Americans and European Americans exhibit distinct gene expression patterns across tissues and tumors associated with immunologic functions and environmental exposures. Sci. Rep. 2021, 11, 9905. [Google Scholar] [CrossRef] [PubMed]
- Benetti, E.; Tita, R.; Spiga, O.; Ciolfi, A.; Birolo, G.; Bruselles, A.; Doddato, G.; Giliberti, A.; Marconi, C.; Musacchia, F.; et al. ACE2 gene variants may underlie interindividual variability and susceptibility to COVID-19 in the Italian population. Eur. J. Hum. Genet. 2020, 28, 1602–1614. [Google Scholar] [CrossRef] [PubMed]
- Jones, R. Multidisciplinary insights into health care financial risk and hospital surge capacity, Part 1: Nearness to death, infectious outbreaks, and Covid-19. J Health Care Fin. 2021, 47, 1–20. [Google Scholar]
- Lawlor, D.; Andersen, A.-M.; Batty, G. Birth cohort studies: past, present and future. Int. J. Epidemiol. 2009, 38, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.; Chiang, C.; Chiang, Y.; Wu, C.; Lee, W.; Cheng, W. Age-specific trend and birth cohort effect on different histologic types of uterine corpus cancers. Sci. Rep. 2023, 13, 1019. [Google Scholar] [CrossRef]
- Ajdacic-Gross, V.; Schmid, M.; Tschopp, A.; Gutzwiller, F. Birth Cohort Effects in Neurological Diseases: Amyotrophic Lateral Sclerosis, Parkinson’s Disease and Multiple Sclerosis. Neuroepidemiol. 2012, 38, 56–63. [Google Scholar] [CrossRef]
- Ohuma, E.; Okiro, E.; Ochola, R.; Sande, C.; Cane, P.; Medley, G.; Bottomley, C.; Nokes, D. The Natural History of Respiratory Syncytial Virus in a Birth Cohort: The Influence of Age and Previous Infection on Reinfection and Disease, Amer J Epid. 2012, 176, 794-802. [CrossRef]
- Eshima, N.; Tokumaru, O.; Hara, S.; Bacal, K.; Korematsu, S.; Karukaya, S.; Uruma, K.; Okabe, N.; Matsuishi, T. Age-specific sex-related differences in infections: A statistical analysis of National Surveillance Data in Japan. PLOS ONE. 2012, 7, e42261. [Google Scholar] [CrossRef]
- Glynn, J.; Moss, P. Systematic analysis of infectious disease outcomes by age shows lowest severity in school-age children. Sci. Data. 2020, 7, 329. [Google Scholar] [CrossRef]
- Ucciferri, C.; Dunn, S. Effect of puberty on the immune system: Relevance to multiple sclerosis. Front. Pediatr. 2022, 10, 1059083. [Google Scholar] [CrossRef]
- Kane, L. , Ismail, N. Puberty as a vulnerable period to the effects of immune challenges: Focus on sex differences. Behav. Brain Res. 2017, 320, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, R.; Cheung, A.; Pang, K.; Saffery, R.; Novakovic, B. Sexual dimorphism in innate immunity: The role of sex hormones and epigenetics. Front. Immunol. 2021, 11, 604000. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E.; Nicodemus-Johnson, J.; Kim, K.; Gern, J.; Jackson, D.; Lemanske, R.; Ober, C. Global DNA methylation changes spanning puberty are near predicted estrogen-responsive genes and enriched for genes involved in endocrine and immune processes. Clin. Epigenet. 2018, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- Sanjuán, R.; Domingo-Calap, P. Mechanisms of viral mutation. Cell Mol. Life Sci. 2016, 73, 4433–4448. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Nazli, A.; Al-Furas, H.; Asad, M.; Ajmal, I.; Khan, D.; Shah, J.; Farooq, M.; Jiang, W. An overview of viral mutagenesis and the impact on pathogenesis of SARS-CoV-2 variants. Front. Immunol. 2022, 13, 1034444. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Basu, R.; Bashyam, M. Assessing the evolution of SARS-CoV-2 lineages and the dynamic associations between nucleotide variations. Access Microbiol. 2023, in press. [CrossRef]
- Yamamoto, R.; Chung, R.; Vazquez, J.; Sheng, H.; Steinberg, P.; Ioannidis, N.; Sudmant, P. Tissue-specific impacts of aging and genetics on gene expression patterns in humans. Nat. Commun. 2022, 13, 5803. [Google Scholar] [CrossRef]
- Frenk, S.; Houseley, J. Gene expression hallmarks of cellular ageing. Biogerontol. 2018, 19, 547–566. [Google Scholar] [CrossRef]
- Cao, J.; Gollapudi, S.; Sharman, E.; Jia, Z.; Gupta, S. Age-related alterations of gene expression patterns in human CD8+ T cells. Aging Cell. 2010, 9, 19–31. [Google Scholar] [CrossRef]
- Escobar, L.; Molina-Cruz, A.; Barillas-Mury, C. BCG vaccine protection from severe coronavirus disease 2019 (COVID-19). Proc Natl Acad Sci U S A. 2020, 117, 17720–17726. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Batra, G.; Kumar, M.; Mishra, A.; Singla, R.; Singh, A.; Singh, R.; Medhi, B. BCG as a game-changer to prevent the infection and severity of COVID-19 pandemic? Allergologia et Immunopathologia 2020, 48, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Coerdt, K.; Khachemoune, A. Corona viruses: reaching far beyond the common cold. Afr. Health Sci. 2021, 21, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Francis, T. On the doctrine of original antigenic sin. Proc. Amer Phil Soc. 1960, 104, 572–578. [Google Scholar]
- Rijkers, G.; van Overveld, F. The “original antigenic sin” and its relevance for COVID–19 (COVID–19) vaccination. Clin. Immunol. Comm. 2021, 1, 1,13–16. [Google Scholar] [CrossRef]
- Vatti, A.; Monsalve, D.; Pacheco, Y.; Chang, C.; Anaya, J.; Gershwin, M. Original antigenic sin: A comprehensive review. J. Autoimmun. 2017, 83, 12–21. [Google Scholar] [CrossRef]
- Lyons-Weiler, J. Pathogenic priming likely contributes to serious and critical illness and mortality in COVID–19 via autoimmunity. J. Transl. Autoimmun. 2020, 3, 100051. [Google Scholar] [CrossRef]
- Adalja, A.A. , & Henderson, D. Original Antigenic Sin and Pandemic (H1N1) 2009. Emerg Infect Dis. 2010, 16, 1028–1029. [Google Scholar] [CrossRef]
- Xu, R.; Ekiert, D.; Krause, J.; Hai, R.; Crowe, J.; Wilson, I. Structural basis of preexisting immunity to the 2009 H1N1 pandemic influenza virus. Science 2010, 328, 357–360. [Google Scholar] [CrossRef]
- Nguyen, A.; Noymer, A. Influenza Mortality in the United States, 2009 Pandemic: Burden, Timing and Age Distribution. PLOS ONE 2013, 8, e64198. [Google Scholar] [CrossRef]
- Simonsen, L.; Spreeuwenberg, P.; Lustig, R.; Taylor, R.; Fleming, D.; Kroneman, M.; Van Kerkhove, M.; Mounts, A.; Paget, W. ; GLaMOR Collaborating Teams. Global mortality estimates for the 2009 Influenza Pandemic from the GLaMOR project: a modeling study. PLoS Med. 2013, 10, e1001558. [Google Scholar] [CrossRef]
- Venkatesh, D.; Anderson, T.; Kimble, J.; Chang, J.; Lopes, S.; Souza, C.; Pekosz, A.; Shaw-Saliba, K.; Rothman, R.; et al. Antigenic Characterization and Pandemic Risk Assessment of North American H1 Influenza A Viruses Circulating in Swine. Microbiol. Spectr. 2022, 10, e0178122. [Google Scholar] [CrossRef] [PubMed]
- Mehfoud, N.; Kimmerly, V.; Shipe, M.; Mapping the 1889-1890 Russian flu. August 2014. Available online: Mapping the 1889-1890 Russian Flu – Circulating Now from NLM (nih.gov) (accessed on 26 January 2023).
- Fonville, J.; Wilks, S.; James, S.; Fox, A.; Ventresca, M.; Aban, M.; Xue, L.; Jones, T.; Le, N.; et al. Antibody landscapes after influenza virus infection or vaccination. Science. 2014, 346, 996–1000. [Google Scholar] [CrossRef] [PubMed]
- Dugan, H.; Guthmiller, J.; Arevalo, P.; Huang, M.; Chen, Y.; Neu, K.; Henry, C.; Zheng, N.; Lan, L.; et al. Preexisting immunity shapes distinct antibody landscapes after influenza virus infection and vaccination in humans. Sci. Transl. Med. 2020, 12, eabd3601. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, M.; Lam, B.; Younossi, E.; Felix, S.; Ziayee, M.; Price, J.; Pham, H.; de Avila, L.; Terra, K.; Austin, P.; et al. The impact of variants and vaccination on the mortality and resource utilization of hospitalized patients with COVID–19. BMC Infect. Dis. 2022, 22, 702. [Google Scholar] [CrossRef] [PubMed]
- Zhong, D.; Xiao, S.; Debes, A.; Egbert, E.; Caturegli, P.; Colantuoni, E.; Milstone, A. Durability of Antibody Levels After Vaccination With mRNA SARS-CoV-2 Vaccine in Individuals With or Without Prior Infection. JAMA. 2021, 326, 326,2524–2526. [Google Scholar] [CrossRef]
- Deng, L. , Li, P., Zhang, X. et al. Risk of SARS-CoV-2 reinfection: a systematic review and meta-analysis. Sci Rep. 2022; 12, 20763. [CrossRef]
- Stein, C.; Nassereldine, H.; Sorensen, R.; Amlag, J.; Bisignano, C.; Byrne, S.; Castro, E.; Coberly, K.; Collins, J.; Dalos, J.; et al. Past SARS-CoV-2 infection protection against re-infection: a systematic review and meta-analysis. The Lancet. 2023. [CrossRef]
- Diani, S.; Leonardi, E.; Cavezzi, A.; Ferrari, S.; Iacono, O.; Limoli, A.; Bouslenko, Z.; Natalini, D.; Conti, S.; Mantovani, M.; et al. SARS-CoV-2-The Role of Natural Immunity: A Narrative Review. J. Clin. Med. 2022, 11, 6272. [Google Scholar] [CrossRef]
- Shenai, M.; Rahme, R.; Noorchashm, H. Equivalency of protection from natural immunity in COVID-19 recovered versus fully vaccinated persons: A systematic review and pooled analysis. Cureus. 2021, 13, e19102. [Google Scholar] [CrossRef]
- Aghbash, P.; Eslami, N.; Shirvaliloo, M.; Baghi, H. Viral coinfections in COVID-19. J Med Virol. 2021, 93, 5310–5322. [Google Scholar] [CrossRef]
- Lansbury, L.; Lim, B.; Baskaran, V.; Lim, W. Co-infections in people with COVID-19: a systematic review and meta-analysis. J Infect. 2020, 81, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Fattorini, L.; Creti, R.; Palma, C.; Pantosti, A. Bacterial coinfections in COVID-19: an underestimated adversary. Ann. Ist. Super. Sanita. 2020, 56, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Agbu, P. , Carthew, R.W. MicroRNA-mediated regulation of glucose and lipid metabolism. Nat. Rev. Mol. Cell Biol. 2021, 22, 425–438. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Bravo Vázquez, L.; Reyes-Pérez, P.; Estrada-Meza, C.; Aponte Alburquerque, R.; Pathak, S.; Banerjee, A.; Bandyopadhyay, A.; Chakraborty, S.; Srivastava, A. The role of microRNAs in solving COVID–19 puzzle from infection to therapeutics: A mini-review. Virus Res. 2022, 308, 198631. [Google Scholar] [CrossRef] [PubMed]
- Abu-Izneid, T.; AlHajri, N.; Ibrahim, A.; Javed, M.; Salem, K.; Pottoo, F.; Kamal, M. Micro-RNAs in the regulation of immune response against SARS CoV-2 and other viral infections. J. Adv. Res. 2021, 30, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Moatar, A.; Chis, A.; Marian, C.; Sirbu, I. Gene Network Analysis of the Transcriptome Impact of COVID–19 Interacting MicroRNAs in COVID–19 Disease. Int. J. Mol. Sci. 2022, 23, 9239. [Google Scholar] [CrossRef] [PubMed]
- Grehl, C.; Schultheiß, C.; Hoffmann, K.; Binder, M.; Altmann, T.; Grosse, I.; Kuhlmann, M. Detection of COVID–19 derived small RNAs and changes in circulating small RNAs associated with COVID–19. Viruses. 2021, 13, 1593. [Google Scholar] [CrossRef] [PubMed]
- Marchi, R.; Sugita, B.; Centa, A.; Fonseca, A.; Bortoletto, S.; Fiorentin, K.; Ferreira, S.; Cavalli, L. The role of microRNAs in modulating COVID–19 infection in human cells: a systematic review. Infect. Genet. Evol. 2021, 91, 104832. [Google Scholar] [CrossRef]
- Meng, F.; Siu, G.; Mok, B.; Sun, J.; Fung, K.; Lam, J.; Wong, N.; Gedefaw, L.; Luo, S.; Le, e T.; Yip, S.; Huang, C-L. Viral microRNAs encoded by nucleocapsid gene of COVID–19 are detected during infection, and targeting metabolic pathways in host cells. CL.Cells. 2021, 10, 1762. [Google Scholar] [CrossRef]
- Singh, M.; Chazal, M.; Quarato, P.; Bourdon, L.; Malabat, C.; Vallet, T.; Vignuzzi, M.; van der Werf, S.; Behillil, S.; Donati, F.; et al. A virus-derived microRNA targets immune response genes during COVID–19 infection. EMBO Rep. 2022, 23, e54341. [Google Scholar] [CrossRef]
- Sica, A.; Colombo, M.; Trama, A.; Horn, L.; Garassino, M.; Torri, V. Immunometabolic Status of COVID-19 Cancer Patients. Physiological Rev. 2020, 100, 1839–1850. [Google Scholar] [CrossRef] [PubMed]
- Sassen, S. , Miska EA, Caldas, C. MicroRNA: implications for cancer. Virchows Arch. 2008 Jan, 452, 1–10. [Google Scholar] [CrossRef]
- Heyn GS, Corrêa LH, Magalhães KG. The Impact of Adipose Tissue-Derived miRNAs in Metabolic Syndrome, Obesity, and Cancer. Front Endocrinol (Lausanne). 2020 Oct 6;11:563816. [CrossRef]
- Kim M, Zhang, X. The Profiling and Role of miRNAs in Diabetes Mellitus. J. Diabetes Clin. Res. 2019, 1, 5–23. [CrossRef]
- Zhang GQ, Wang SQ, Chen Y, Fu LY, Xu YN, Li L, Tao, L., Shen XC. MicroRNAs Regulating Mitochondrial Function in Cardiac Diseases. Front. Pharmacol. 2021 May 28;12:663322. [CrossRef]
- Uzuner E, Ulu GT, Gürler SB, Baran, Y. The Role of MiRNA in Cancer: Pathogenesis, Diagnosis, and Treatment. Methods Mol. Biol. 2022, 2257, 375–422. [CrossRef]
- Jones, R.P.; Ponomarenko, A. Effect of Age, Sex, and COVID-19 Vaccination History on All-Cause Mortality: Unexpected Outcomes in a Complex Biological and Social System. Preprints.org 2023, 2023040248. [Google Scholar] [CrossRef]
- Boon, R.; Vickers, K. Intercellular transport of microRNAs. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 186–92. [Google Scholar] [CrossRef]
- Shin, J.; Bayry, J. An age-related decline of CD62L and vaccine response: a role of microRNA 92a? Hum Vaccin Immunother. 2014, 10, 1404–5. [Google Scholar] [CrossRef]
- Di Palo, A.; Siniscalchi, C.; Salerno, M.; Russo, A.; Gravholt, C.; Potenza, N. What microRNAs could tell us about the human X chromosome. Cell. Mol. Life Sci. 2020, 77, 4069–4080. [Google Scholar] [CrossRef]
- Nakanishi, T.; Pigazzini, S.; Degenhardt,F. ; Cordioli,M.; Butler-Laporte,G.; Maya-Miles, D.; Bujanda, L.; Bouysran,Y.; Niemi, M.; Palom,A.; Ellinghaus, D.; Khan, A.; Martínez-Bueno, M.; et al. Age-dependent impact of the major common genetic risk factor for COVID–19 on severity and mortality. J. Clin. Invest. 2021, 131, e152386. [Google Scholar] [CrossRef] [PubMed]
- Zeberg, H.; Pääbo, S. The major genetic risk factor for severe COVID–19 is inherited from Neanderthals. Nature 2020, 587, 610–612. [Google Scholar] [CrossRef] [PubMed]
- Zeberg, H. The major genetic risk factor for severe COVID–19 is associated with protection against HIV. PNAS. 2022, 119, e2116435119. [Google Scholar] [CrossRef] [PubMed]
- Tian F, Tong B, Sun L, Shi S, Zheng B, Wang Z, Dong X, Zheng, P. N501Y mutation of spike protein in SARS-CoV-2 strengthens its binding to receptor ACE2. Elife. 2021, 10, e69091. [CrossRef]
- Liu Y, Liu J, Plante KS, Plante JA, Xie X, Zhang X, Ku Z, An Z, Scharton D, Schindewolf C, Widen SG, Menachery VD, Shi PY, Weaver SC. The N501Y spike substitution enhances SARS-CoV-2 infection and transmission. Nature. 2022, 602, 294–299. [CrossRef]
- The Animation Lab. Building a visual consensus model of the SARS-CoV-2 life cycle. Available online: SARS-CoV-2 Visualization and Annotation Project – The Animation Lab (utah.edu) (accessed on 26 February 2023).
- Dubrow, A. Hidden states of the COVID-19 spike protein. Available online: Hidden States of the COVID-19 Spike Protein - Latest News - Texas Advanced Computing Center (utexas.edu) (accessed 26 February 2023).
- Luhur, A.; Chawla. G.; Sokol, N. MicroRNAs as components of systemic signaling pathways in Drosophila melanogaster. Curr. Top. Dev. Biol. 2013, 105, 97–123. [Google Scholar] [CrossRef]
- Leroy, H.; Han, M.; Woottum, M.; Bracq, L.; Bouchet, J.; Xie, M.; Benichou, S. Virus-Mediated Cell-Cell Fusion. Int. J. Mol. Sci. 2020, 21, 9644. [Google Scholar] [CrossRef]
- Hernández, J.; Podbilewicz, B. The hallmarks of cell-cell fusion. Development. 2017, 144, 4481–4495. [Google Scholar] [CrossRef]
- Sedger, L. microRNA control of interferons and interferon induced anti-viral activity. Mol. Immunol. 2013 Dec;56:781-93. [CrossRef]
- Bouvet, M.; Voigt, S.; Tagawa, T.; Albanese, M.; Chen, Y.; Chen, Y.; Fachko, D.; Pich, D.; Göbel, C.; Skalsky, R.; et al. Multiple Viral microRNAs Regulate Interferon Release and Signaling Early during Infection with Epstein-Barr Virus. mBio. 2021 Mar 30;12:e03440-20. 0344. [Google Scholar] [CrossRef]
- The Severe COVID–19 GWAS Group. Genomewide association study of severe COVID–19 with respiratory failure. N. Engl. J. Med. 2020, 383, 1522–1534. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.; Bes, T.; Velho, M.; Marques, E.; Jannes, C.; Valino, K.; Dinardo, C.; Costa, S.; Duarte, A.; Santos, A.; et al. Genetic risk factors and COVID–19 severity in Brazil: results from BRACOVID study. Human Molecular Genetics, 2022, 31, 3021–3031. [Google Scholar] [CrossRef] [PubMed]
- Downes, D.; Cross, A.; Hua, P. Identification of LZTFL1 as a candidate effector gene at a COVID–19 risk locus. Nat. Genet. 2021, 53, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Kousathanas, A.; Pairo-Castineira, E.; Rawlik, K.; Stuckey, A.; Odhams, C.; Walker, S.; Russell, C.; Malinauskas, T.; Wu, Y.; Millar,J. ; Shen, X.; et al. Whole-genome sequencing reveals host factors underlying critical COVID–19. Nature 2022, 607, 97–103. [Google Scholar] [CrossRef]
- Pushparaj, P.; Nicoletto, A.; Sheward, D.; Das, H.; Castro Dopico, X.; Perez Vidakovics, L.; Hanke, L.; Chernyshev, M.; Narang, S.; Kim, S.; et al. Immunoglobulin germline gene polymorphisms influence the function of SARS-CoV-2 neutralizing antibodies. Immunity. 2023, 56, 193–206.e7. [Google Scholar] [CrossRef]
- Joung, S.; Weber, B.; Wu, M.; Liu, Y.; Tang, A.; Driver, M.; Sternbach, S.; Wynter, T.; Hoang, A.; Barajas, D.; et al. Serological response to vaccination in post-acute sequelae of COVID. BMC Infect. Dis. 2023, 23, 97. [Google Scholar] [CrossRef]
Figure 1.
Single year of age adjusted all-cause deaths in England and Wales for males and females between 2018 and 2021. Data for 2019 is the ‘average’ expected for that year based on the trend between 2011 and 2019. Age concludes at 105+.
Figure 1.
Single year of age adjusted all-cause deaths in England and Wales for males and females between 2018 and 2021. Data for 2019 is the ‘average’ expected for that year based on the trend between 2011 and 2019. Age concludes at 105+.
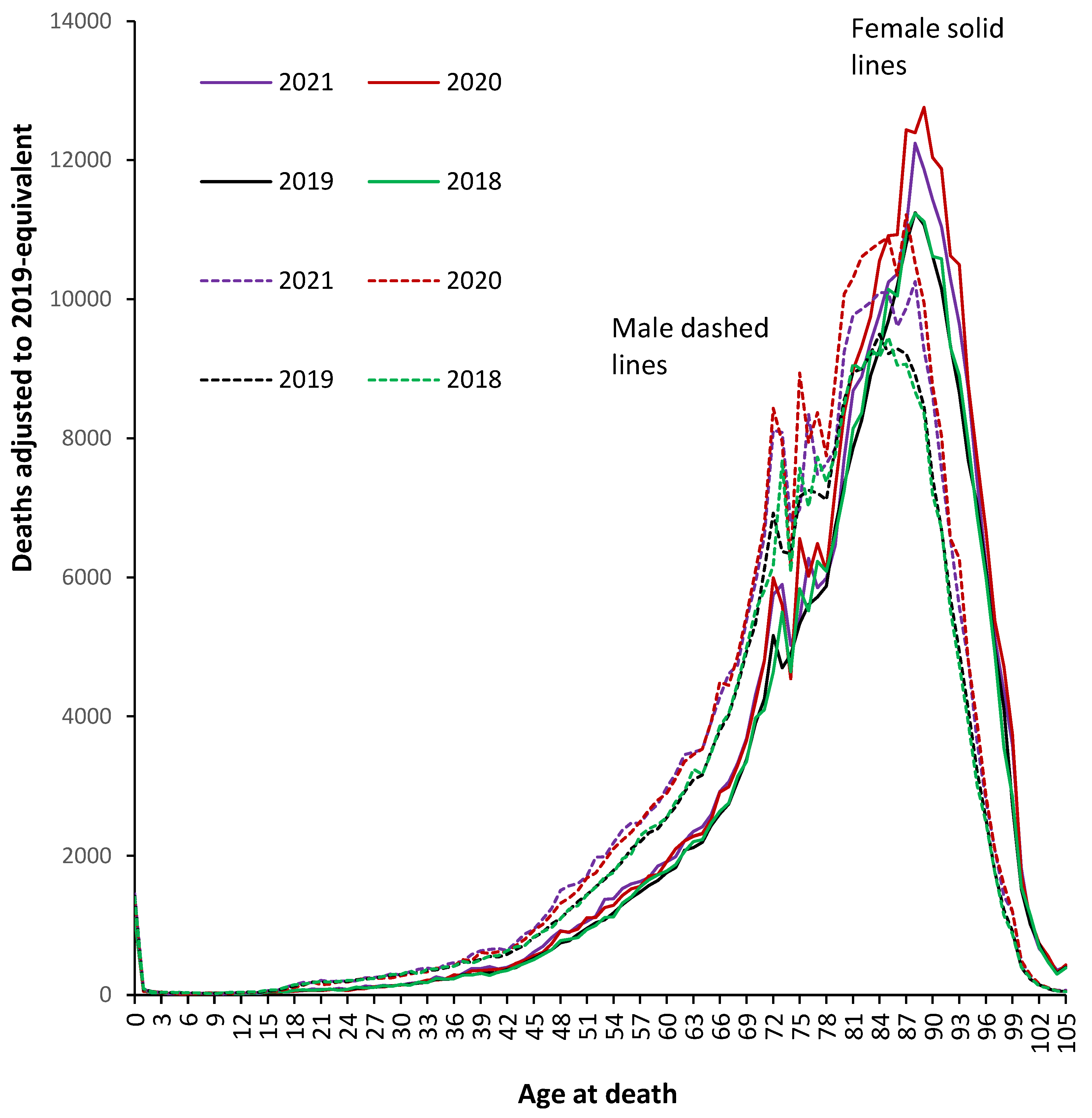
Figure 2.
Excess all-cause male deaths (as a percentage difference) during 2020 and 2021 (against the 2019 baseline) across ages 50 to 105+. The grey line shows the value of 1 standard deviation (STDEV) of Poisson variation based on the 2019 baseline.
Figure 2.
Excess all-cause male deaths (as a percentage difference) during 2020 and 2021 (against the 2019 baseline) across ages 50 to 105+. The grey line shows the value of 1 standard deviation (STDEV) of Poisson variation based on the 2019 baseline.
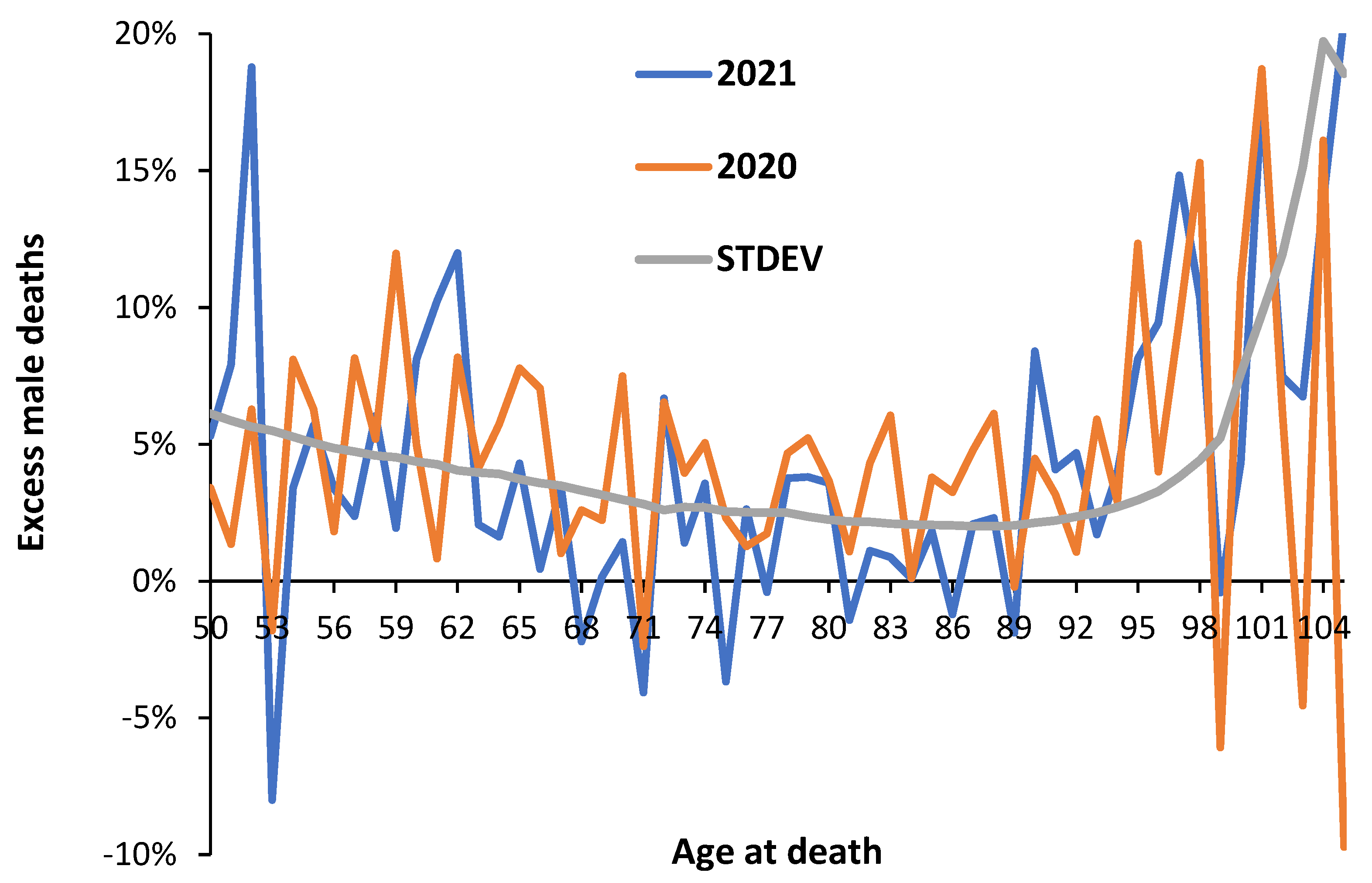
Figure 3.
Change in all-cause mortality for males and females during 2021 (with vaccination) versus 2020 (without vaccination). The difference is expressed as a percentage relative to the 2019 baseline.
Figure 3.
Change in all-cause mortality for males and females during 2021 (with vaccination) versus 2020 (without vaccination). The difference is expressed as a percentage relative to the 2019 baseline.
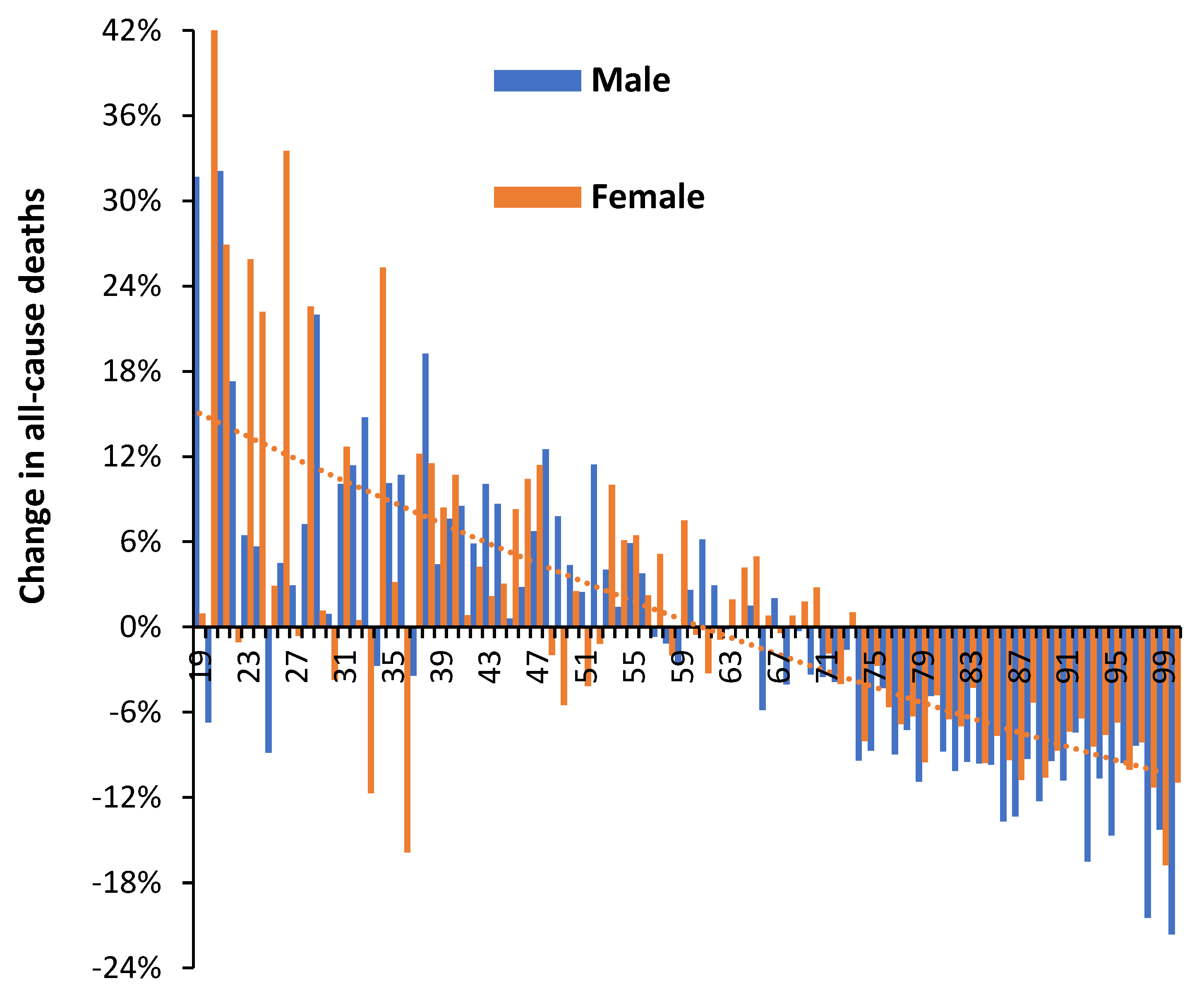
Figure 4.
Average/median age at death for males and females in England and Wales for deaths ‘due to’ COVID–19 between March 2020 and December 2022. Data is from ONS [29,30].
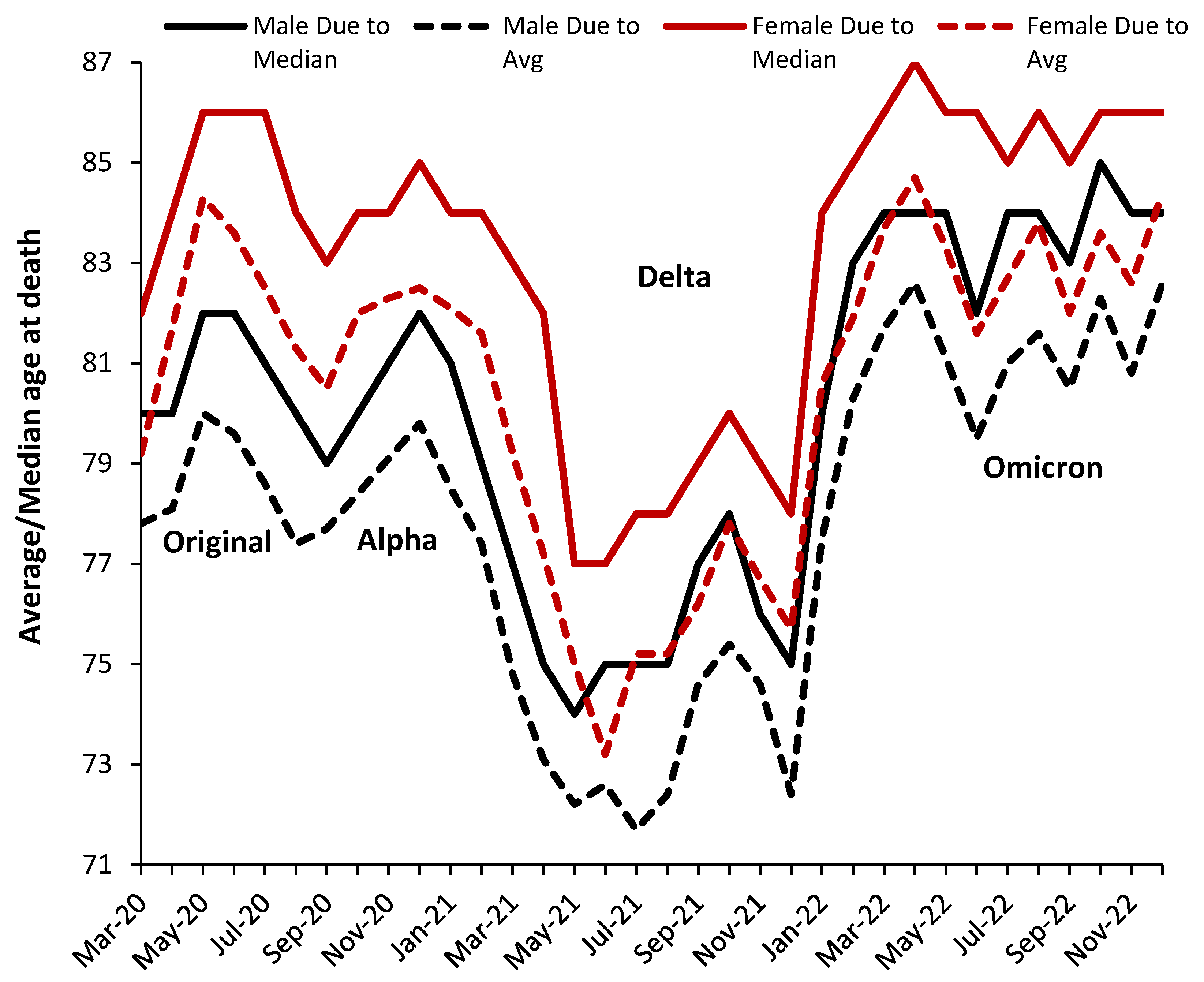
Figure 5.
Single-year-of-age (SYOA) profiles for the proportion of total deaths assessed as ‘due to’ COVID–19 in England and Wales. Below age 25, data has been aggregated to age 1 ̶ 4, 5 ̶ 9, through to 20 ̶ 24. Analysis of raw data in [30].
Figure 5.
Single-year-of-age (SYOA) profiles for the proportion of total deaths assessed as ‘due to’ COVID–19 in England and Wales. Below age 25, data has been aggregated to age 1 ̶ 4, 5 ̶ 9, through to 20 ̶ 24. Analysis of raw data in [30].
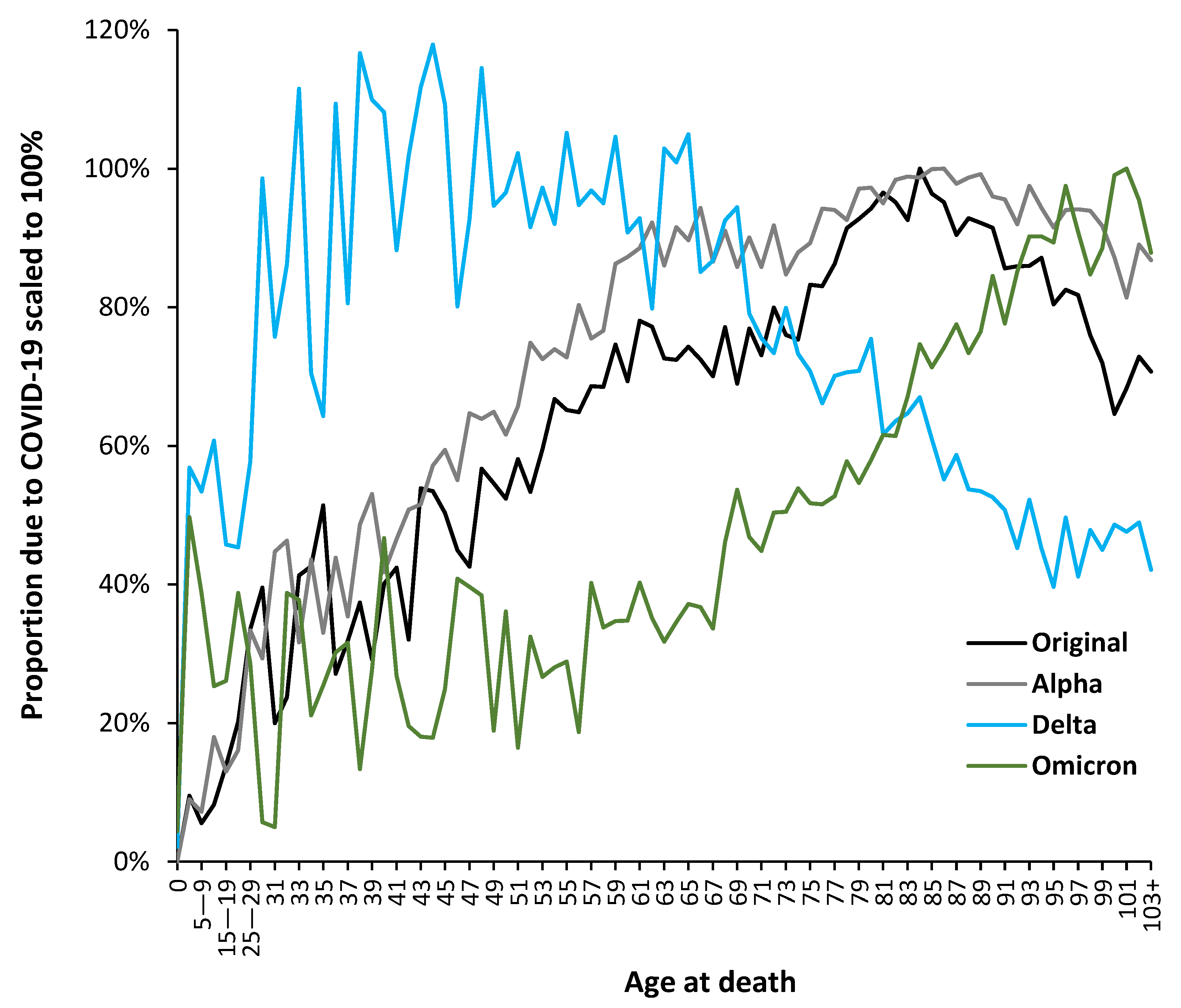
Figure 6.
Proportion of male or female total deaths ‘with’ COVID-19 during each SARS-CoV-2 variant for the USA, plus proportion of total all-cause deaths. Analysis is from data in [37].
Figure 6.
Proportion of male or female total deaths ‘with’ COVID-19 during each SARS-CoV-2 variant for the USA, plus proportion of total all-cause deaths. Analysis is from data in [37].
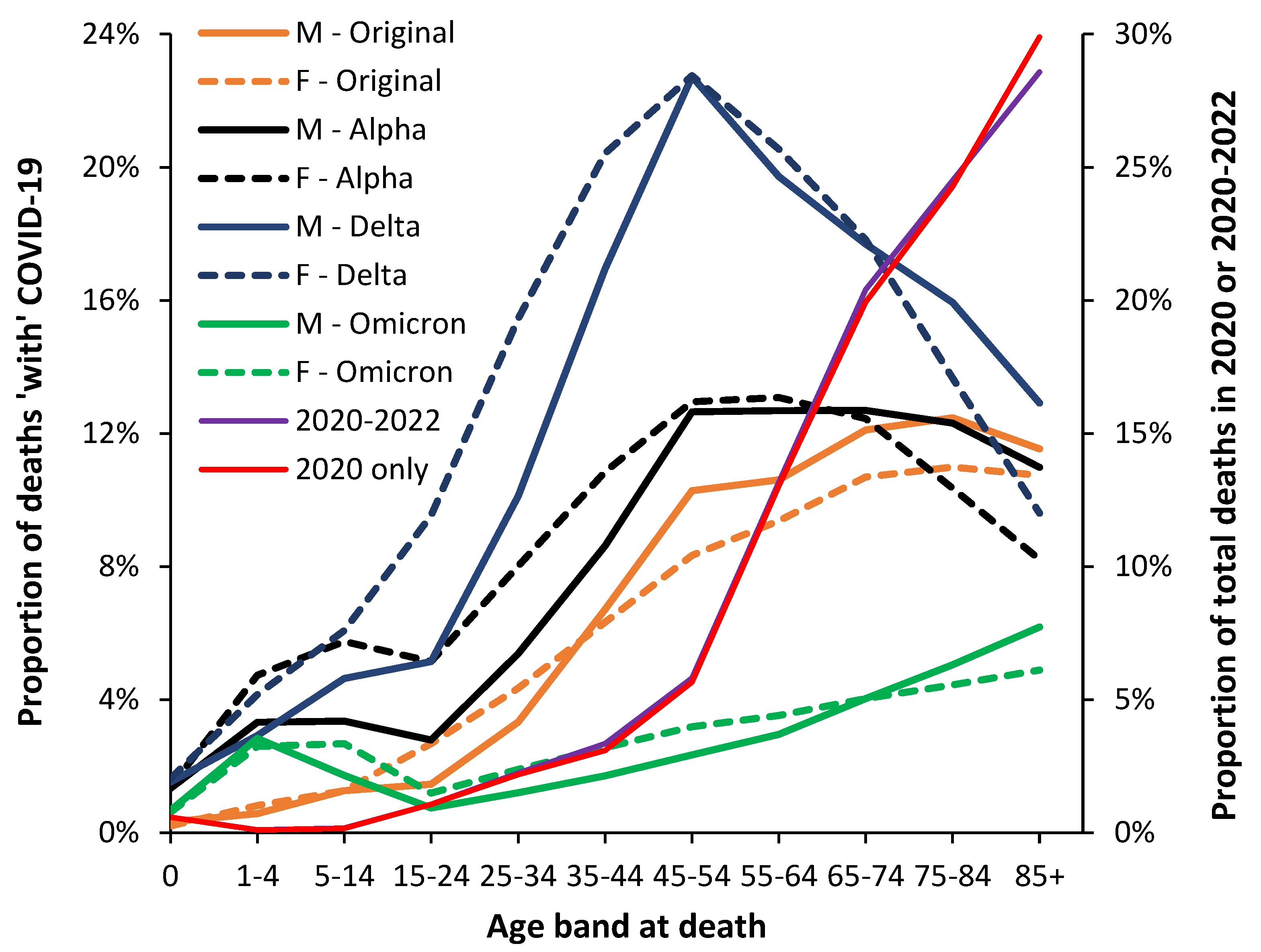
Figure 7.
Proportion of total deaths ‘with’ COVID-19 for several American states and for the USA during the period of the original Wuhan strain. Analysis of data from [37]. As mentioned in the methods Section 2.1. the CDC masks death values between 1-9 at state level and these missing values were interpolated. This mainly affects the two youngest age bands and the smallest states.
Figure 7.
Proportion of total deaths ‘with’ COVID-19 for several American states and for the USA during the period of the original Wuhan strain. Analysis of data from [37]. As mentioned in the methods Section 2.1. the CDC masks death values between 1-9 at state level and these missing values were interpolated. This mainly affects the two youngest age bands and the smallest states.
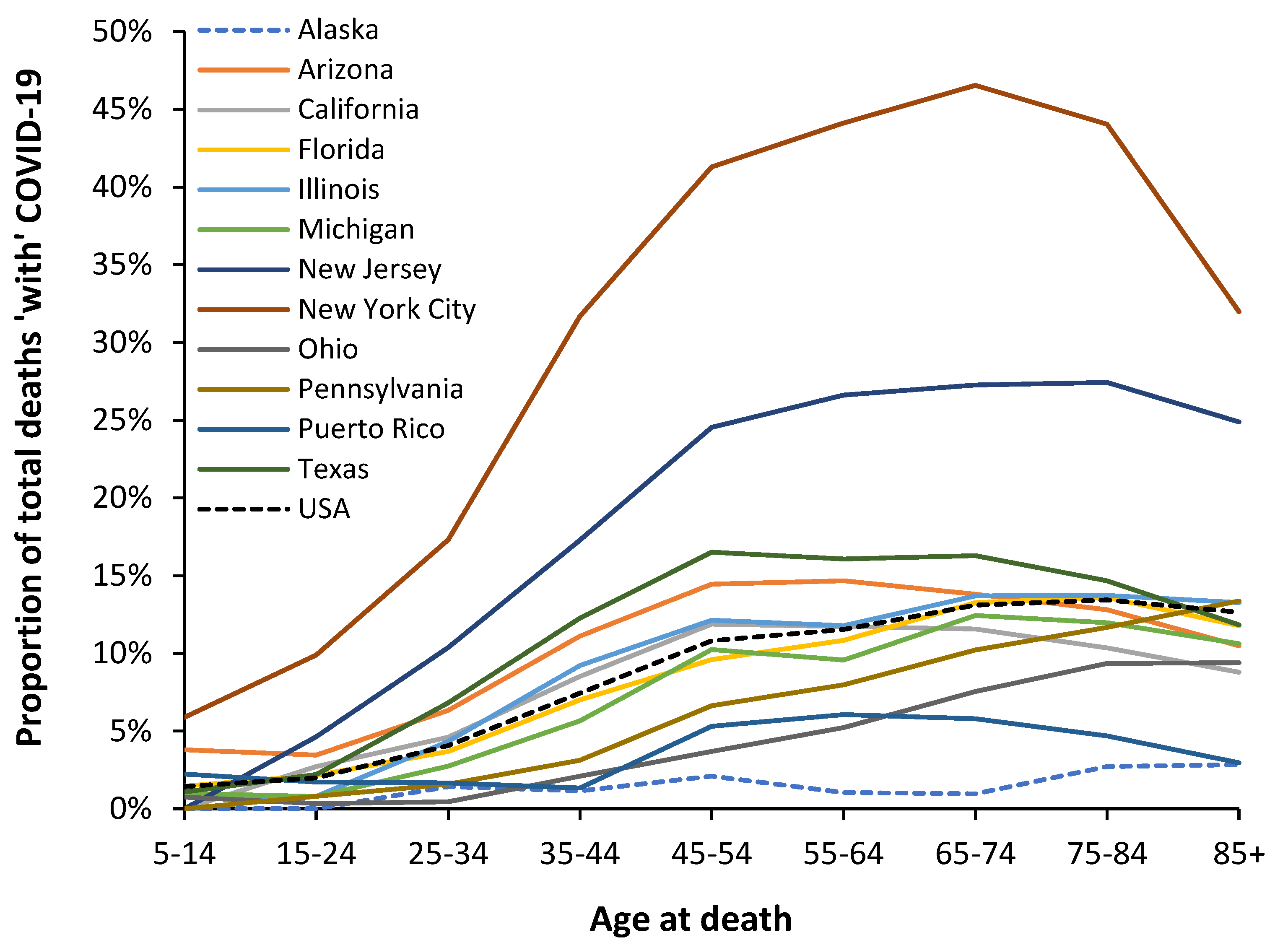
Figure 8.
Maximum all-cause mortality (ACM) rate for unvaccinated males and females for the three COVID–19 outbreaks during the Alpha, Delta, and Omicron variants. Analysis of data in [35].
Figure 8.
Maximum all-cause mortality (ACM) rate for unvaccinated males and females for the three COVID–19 outbreaks during the Alpha, Delta, and Omicron variants. Analysis of data in [35].
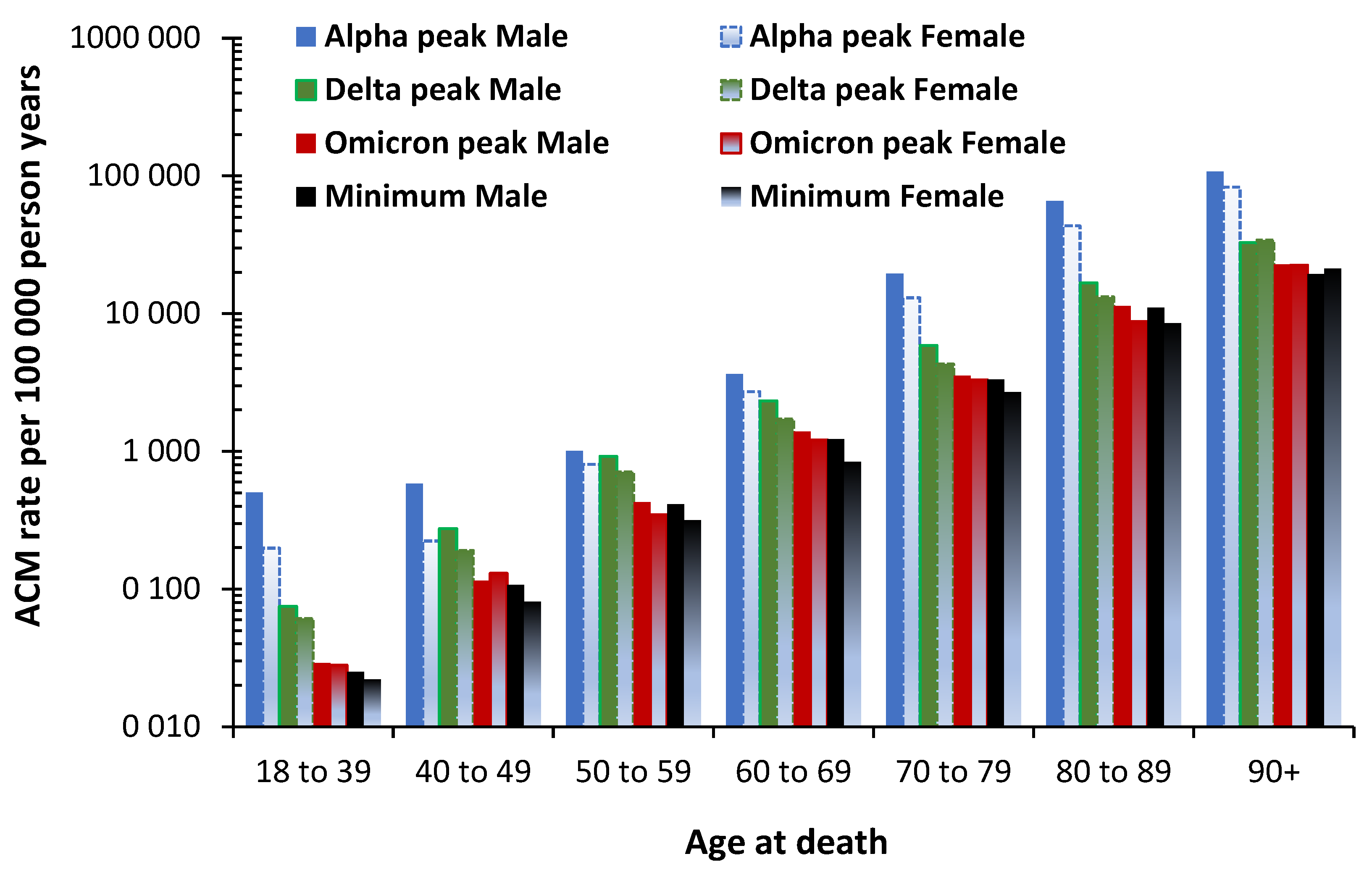

Table 1.
A time profile for months in which deaths are dominated by a single variant in England and Wales. (β) - the absolute magnitude of COVID-19 deaths may be underestimated in the early months because testing capacity was inadequate [14]. Point for maximum deaths aged 65+ is shown in bold.
Table 1.
A time profile for months in which deaths are dominated by a single variant in England and Wales. (β) - the absolute magnitude of COVID-19 deaths may be underestimated in the early months because testing capacity was inadequate [14]. Point for maximum deaths aged 65+ is shown in bold.
| Month | Proportion of total deaths due to COVID-19: average age 65+ | Variant arrives (infections) | Variant dominates deaths |
|---|---|---|---|
| October 2022 | 4% | Omicron | |
| September 2022 | 2% | Omicron | |
| August 2022 | 4% | Omicron | |
| July 2022 | 4% | Omicron | |
| June 2022 | 2% | Omicron | |
| May 2022 | 4% | Omicron | |
| April 2022 | 7% | Omicron | |
| March 2022 | 5% | Omicron | |
| February 2022 | 6% | Mixed Delta/Omicron | |
| January 2022 | 8% | Omicron starts | Mixed Delta/Omicron |
| December 2021 | 5% | Delta | |
| November 2021 | 7% | Delta | |
| October 2021 | 6% | Delta | |
| September 2021 | 6% | Delta | |
| August 2021 | 5% | Delta | |
| July 2021 | 2% | Mixed Alpha/Delta | |
| June 2021 | 1% | Delta starts | Mixed Alpha/Delta |
| May 2021 | 1% | Alpha | |
| April 2021 | 2% | Alpha | |
| March 2021 | 9% | Alpha | |
| February 2021 | 30% | Alpha | |
| January 2021 | 38% | Alpha | |
| December 2020 | 22% | Alpha | |
| November 2020 | 19% | Alpha | |
| October 2020 | 7% | Mixed Alpha/Original | |
| September 2020 | 1% | Alpha starts | Mixed Alpha/Original |
| August 2020 | 1% | Original | |
| July 2020 | 3% | Original | |
| June 2020 | 9% | Original | |
| May 2020 (β) | 23% | Original | |
| April 2020 (β) | 32% | Original | |
| March 2020 (β) | 3% | Original |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



