
Submitted:
12 May 2023
Posted:
12 May 2023
You are already at the latest version
Abstract
This study aimed to evaluate the effects of a mixture of olive, laurel and rosemary leaf powders, on performance and egg quality, the oxidative state and biochemical, immune, and intestinal morphophysiological parameters of laying hens. One hundred Lohaman Brown hens (28 weeks old) were equally assigned to two groups (n. 50) corresponding to a basal control diet (CON) or the diet supplemented with 6g/kg feed of leaf powder mixture (LPM) containing olive, laurel and rosemary leaves (1:1:1), for 8 weeks. Egg traits, oxidative status, biochemical indices, immune response, cecal short chain fatty acids (SCFAs) and intestinal morphological characteristics were evaluated at the end of the experiment. The results indicated that body weight and egg quality parameters were not influenced by LPM. However, LPM improved (P<0.01) the oxidative status and the immune system, total protein (P<0.05) and HDL cholesterol (P<0.01) and decreased total cholesterol (P<0.01) and LDL cholesterol (P<0.05), as compared to the CON. Aspartate amino transferase (AST), alkaline phosphatase (ALP) and alanine amino transpherase were significantly better in the LPM than in the CON group. A significant increase (P<0.05) of SCFA content established in the caecum and in villi height and crypt depth in both duodenum and ileum LPM treated hens was observed.
Keywords:
olive
; laurel
; rosemary
; histology
; SCFA
; immunomodulation
; liver
1. Introduction
Phytochemicals and plants rich in nutrients and bioactive substances play a pivotal role in the health and productive performance of poultry [1,2]. Their use is gaining interest in the livestock industry due to their safety and less hazardous properties compared to all feed additives [3]. Natural plants get a lot of attention as they are cheap and contain wide varieties of bioactive compounds with anti-oxidative, anti-inflammatory, anti-microbial, and antiparasitic effects in poultry [2,4,5]. In addition, their use constitutes an emerging innovative food processing technique to promote a sustainable food industry [6].
Among natural products, polyphenols have raised increasing interest in poultry nutrition [7] as promoters of growth performance [8] and egg quality [9].
Polyphenol compounds are among the most extensive layers produced by plants, as around 8,000 of these compounds have been identified [10]. Polyphenols are present in several parts of the plant, including the leaves, flowers, seeds, and fruits [11]. Phytogenic feed supplements, such as spices, intact herbs, and their extracts, have beneficial effects on the health and performance of animals, due to their secondary metabolite content, including polyphenols [7]. The beneficial impact of polyphenols in poultry nutrition [9,12] includes improving antioxidant status and intestinal health, which can promote increased production [13,14]. Phytochemicals, including aromatic plants and plant extracts, may improve feed intake, gastric secretion, and histological morphometry of the small intestine in poultry [15,16,17]. Thus, the supplementation of poultry diets with leaves, spices, and intact herbs or their extracts, can constitute a simple, suitable strategy to improve animal health and productive performance and to introduce natural antioxidants into the products improving oxidative stability and shelf life.
Olive leaves constitute one of the many waste by-products obtained during the harvesting of the olives, the pruning of olive trees (25 kg per olive tree), and the cleaning procedures before the extraction of the olive oil (10% of the total weight of collected material) [18]. Peculiar molecular and biological characteristics are attributed to olive leaves [19,20], such as antioxidant [21] and anti-inflammatory [22] properties, the inhibition of low-density lipoprotein receptors [23], and the inhibition of microbial disorders [24], deriving from the action of phytochemicals present in olive leaves, in particular polyphenols [19,25]. Olive leaves contain more than 30 phenolic compounds classified as phenolic acids, phenolic alcohols, flavonoids, secoiridoid, and lignans. The main family of compounds is made up of secoiridoids and oleuropein is the most abundant bioactive compound in the leaves [26,27], constituting 6-9% of dry olive leaves [28]. Flavonoids are present in significant quantities in leaves while simple phenols (such as hydroxytyrosol and tyrosol) and phenolic acids are less representative but contribute considerably to the overall antioxidant power of the leaves [19]. The potential of olive leaf polyphenols to reduce oxidative stress and to influence the health of farm animals has not been extensively studied. In poultry, olive leaf dietary supplementation positively affects body weight in broilers [29] and hens [30], as well as the blood profile and oxidative status in Japanese quails treated with olive leaf extract [31].
Laurus nobilis L., an aromatic and medicinal plant, belongs to the Laureacea family. It is widespread in Mediterranean countries and is cultivated in other temperate and warm parts of the world [32]. Laurus is characterized by a high content of proteins, organic acids, a favorable PUFA/SFA and n-6/n-3 ratio, and antioxidant activity [33]. Conforti et al. (2006) [32] showed that the amounts of phenolics in Laurus leaves were 210 and 219 mg/g for wild and cultivated plant extracts, respectively. Regarding antioxidant activity, such as scavenging activity, reducing power and lipid peroxidation inhibition [33,35], abundant phenolic compounds were found in laurel leaves [36,37]. The hydroxyl groups attached to the ring structure of flavonoids conferred them with antioxidant properties, acting as reducing agents, hydrogen donators, metal chelators and radical scavengers, preventing oxidative stress [35].
The bioactive components in laurel leaves have been shown to decrease serum total cholesterol and LDL cholesterol content and to increase HDL cholesterol levels [38,39]. The leaf essential oil contains predominantly oxygenated monoterpenoids (66.2%) [40]. It includes 1.8-cineol (over 50%), eugenol, acetyl and methyl eugenol, α-and β-pinene, phellandrene, linalool, geraniol and terpineol, camphene, sabinene and limonene [41]. Laurel leaves and extracts have also been reported to have anti-inflammatory and antibacterial properties [42,43].
Among the natural antioxidant additives in poultry diets, much attention has been directed to herbs and spices [1,4]. Rosemary (Rosmarinus officinalis L.), an herb of the Labiatae family, is a spice obtained by drying the leaves and flowers of the plant. The many biological activities attributed to rosemary [44] are mostly represented by flavonoids (genkwanin, cirsimaritin and homoplantaginin), phenolic diterpenes (carnosic acid, carnosol, and rosmanol), and triterpenes (ursolic acid) [45]; while the most bioactive components are represented by volatile oils that include 1.8 cineol, α-pinene and camphor [46].
The European Food Safety Authority has proposed rosemary extracts as feed additives [47] in the antioxidants class [48]. Rosemary is considered a plant with high antioxidant activity via the eradication of free radicals, the constitution of chelates with metal ions and prevention or reduction of oxidation [49]. In particular, carnosic acid is the most active antioxidant present in rosemary powder or extract [50]. Carnosic acid and carnosol account for over 90% of rosemary antioxidant activity [51]. Both compounds reduce membrane damage, inhibit lipid peroxidation, and lower DNA damage [52]. Shan et al. (2005) [53] found similar antioxidant activity for rosemary and laurel while de Falco et al. (2022) [54] reported that the radical scavenging activity of Rosmarinus officinalis is two times higher than that of L. nobilis. In terms of different pharmacological activities, rosemary has also been considered an anti-inflammatory, immunomodulatory, and antimicrobial agent [55,56,57].
Rosemary supplementation can decrease blood cholesterol and improve blood pressure and stomach function in rats [58,59]. It has also been reported that dietary rosemary supplementation increases nutrient uptake and appetite, saliva secretion, the synthesis and digestion of bile acids, and the absorption of lipids in poultry [60] and, ultimately, it enhances growth [61]. Moreover, Alagawany and Abd El-Hack (2015) [62] showed that dietary supplementation of rosemary powder was able to improve egg production and blood parameters in laying hens.
Gut health can be considered synonymous with animal health and is of crucial importance for animal performance [63]. Gut health depends on the maintenance of the delicate balance between the host, intestinal microbiota, intestinal morphology, and dietary compounds. Dietary polyphenols affect intestinal microflora and gut morphology, nutrient digestibility, and poultry performance [64].
It has been suggested that the use of more antioxidants with multifunctional properties in the diet can provide better protection with respect to mono antioxidant compounds [65]. A mixture of antioxidant sources performed better than individual antioxidants in pork [66] and dairy cows [67]. A significant improvement in body weight gain and feed efficiency were observed when broilers were given diets supplemented with a mixture of 14 herbs [68].
The dietary effects of leaves of the three plants, Olea europaea L., Laurel nobilis L., and Rosmarinus officinalis L. have been studied individually, revealing good results as nutraceutical supplements in animals. However, there is little evidence about their use as mixed supplementation and/or inclusion as a powder in the diet of laying hens.
In this study, we hypothesized that a powder supplement of a mixture of olive, laurel and rosemary leaves, due to their positive evidence, could reduce oxidative stress, improving intestinal health and the metabolic profile, leading to better performance among laying hens. Therefore, in this study we set out to evaluate the effect of dietary supplementation with a powder mixture of olive, laurel, and rosemary leaves on performance and egg quality, biochemical blood profile and oxidative status, caecal compound characteristics, and intestinal morphology in laying hens.
2. Material and Methods
The experimental protocol of the study and implemented animal care procedures were approved by the Institutional ethics committee of the Department of Emergencies and Organ Transplantation of the University of Bari (Prot n. 04/2020).
2.1. Plant material collection
The olive, rosemary, and laurel leaves were harvested during the spring on private farms with organic farming practices in the agricultural area of Bari. The collected leaves were then properly dried and ground to pass a 2mm screen. The single plant leaves were evaluated for proximate composition [69], and total phenol content [70] using the Folin-Ciocalteau reagent (Merck- Darmstadt, Germany) (Table 1).
2.2. Animal, Diets, and Experimental Design
The trial was performed in a private farm located in Apulia, South Italy (latitude 41 04′N, 17 05′E, 5 m s.l.m.). The trial was conducted in autumn and lasted 60 days on a total of one hundred Lohman Brown laying hens, 28 weeks old with an initial body weight of 2140 ± 140g. All hens were vaccinated according to the vaccination schedule required by commercial systems. The experimental period extended from 28 to 36 weeks of age.
The hens were assigned on the basis of their weight to two equal groups corresponding to the dietary treatments. The dietary treatments included a commercial basal diet, the control (CON), and the basal diet supplemented with 6g/kg feed of the experimental leaf powder mixture (LPM), which contained 2g/kg olives leaves + 2g/kg laurel leaves + 2g/kg rosemary leaves. The feed for laying hens was prepared according to the nutritional requirements reported by [71] and the nutrition levels were shown in Table 2.
2.3. Animal housing
The hens were reared using an indoor-outdoor rearing system with two pens (4 m2/hen), one for each experimental group, separated by a 2 m high net, to prevent their mixing.
Each enclosure contained a poultry house at a low density (6 hens/m2) with nests (1/6 hens) and perches, allowing the hens free access to the outdoor enclosure (outside option from 08:00 h to 18:00). Feed and water were offered ad libitum. A photoperiod of 16h light and 8h dark was maintained with artificial light.
2.4. Recorded egg quality
All hens were weighed at the beginning and at the end of the experiment. At the end of the experiment (36th week), a total of 30 eggs from each experimental group were randomly collected over 3 days for the analysis of egg quality.
The collected eggs were weighed individually using an electronic balance (0.1 g sensitivity). When broken, the egg contents were split up using an egg separator and were curled on wet paper to remove the white albumen; subsequently, the egg white, yolk, and shell were weighed using an electronic balance (0.1 g sensitivity). Eggshell thickness was measured from the middle part of the egg using a micrometer (25M-5, Ames).
The egg yolk coloration was measured visually according to the RYCF (Roche Yolk Colour Fan; Hoffmann-La Roche Ltd., Basel, Switzerland), based on a scale of colors from 1 (light pale) to 15 (dark orange).
The following egg quality indexes were calculated:
Yolk ratio = yolk weight (g)/egg weight (g)×100;
Albumen ratio = albumen weight (g)/egg weight (g)× 100;
Thickness shell ratio = shell weight (g)/egg weight (g)×100
2.5. Blood sampling and analyses
At the end of the experimental period, approximately 2 mL of blood samples were taken from the brachial vein of 30 hens, randomly chosen from each experimental group, and were collected in plastic vacuum tubes (BD Vacutainer Advance, Becton Dickinson, NJ, USA).
The blood samples were centrifuged at 3000 rpm (1814g) for 15 min and the serum aliquots were stored at −20°C until the analysis of biochemical, immunological, and antioxidant parameters.
Blood biochemical characteristics were determined using an automated biochemical analyzer (TC-220 TECOM, Jiangxi, China) using commercial kits according to the colorimetric method. The concentration of total protein (TP, monitored at 540 nm), triglicerides (TG, monitored at 505 nm), total cholesterol (TC, monitored at 505 nm), high-density lipoprotein-cholesterol (HDL-C, monitored at 570 nm), low-density lipoprotein (LDL), alkaline phosphatase (ALP, monitored at 405 nm), total bilirubin (monitored at 555 nm) and aspartate aminotransferase (AST) were estimated using diagnostic kits produced by SPINREACT (Sant Esteve de Bas, Girona, Spain). Non esterified fatty acids (NEFA, monitored at 550 nm) were determined in fresh serum immediately after the blood sampling, using a commercial kit produced by Randoz Laboratories Ltd. (Crumlin, County Antrim, UK). Alanine aminotransferase (ALT) was determined (at 340 nm) using a commercial kit produced by PRO-EKO (Petacciato, Campobasso, Italy).
2.6. Determination of cytokines
The level of inflammatory cytokines IL-1β, TNFα, and IL-6 were measured with corresponding assay kits provided by Immunological Sciences (Rome, Italy) following the manufacturer’s instructions, using Microplate reader TECAN infinite M1000Pro plate reader (Tecan, Mannedorf, Switzerland).
2.7. Determination of oxidative status
The serum oxidant/antioxidant markers were assessed spectrophotometrically. Total oxidative status (TOS) was measured according to Erel (2004) [72]; results were given as micromolar H2O2 equivalent/L. Measurements of total antioxidant status (TAS) were done according to Erel (2005) [73]; Trolox equivalent/L units were used. ROM values were determined spectrophotometrically (wavelength 505 nm), using a commercial kit (Diacron, Grosseto, Italy), according to the manufacturer’s instruction. Results were expressed in Carr units (1 U/Carr corresponds to 0.024 mmol/L of H2O2).
2.8. Intestinal Tissue Sampling and Analyses
At the end of the experiment, a total of 8 animals from each experimental group were slaughtered. The intestinal tract was removed for SCFA analyses and histologic measurements.
2.8.1. Short Chain Fatty Acid (SCFA) Analyses
The contents of the caecum of 8 animals were squeezed out by finger pressure, collected in Eppendorf tubes, and stored at −20℃ until analyzing the short-chain fatty acids (SCFAs).
The SCFA concentrations were determined by gas chromatography. The method reported by Lashkari et al. (2014) [74] was used, and slightly modified. Briefly, samples were blended with deionized water for 15min. The homogenate was filtered through filter paper (Whatman grade 1; Sigma Aldrich, Darmstadt, Germany); the filtrate (5 mL) had meta-phosphoric acid (25%, wt/vol; 1 mL) added. After shaking (10 min), volatile fatty acids were extracted with toluene and quantified by flame ionization detection gas chromatography using the Agilent 6890A GC system equipped with an FID detector (Agilent 7890A GC, Agilent Technology Italia Spa, Roma, Italia) and capillary column (SACtm-5 column 300 cm × 0.25 mm, Supelco, USA). Fatty acids were identified and quantified on the basis of standard elution times (Volatile Free Acid Mix Supelco, Bellefonte, PA, USA).
2.9. Histological Morphometry
2.9.1. Sampling and tissue preparation
Segments of approximately 3 cm were collected from the duodenum and ileum of 6 animals and fixed in 4% (v/v) phosphate-buffered-saline paraformaldehyde for 24 h at 4°C. After dehydration through a graded series of ethanol, the samples were embedded in paraffin wax. Serial sections (5μm thick) were cut and, after being de-waxed with xylene and hydrated in an ethanol series of descending concentrations, they were stained with Haematoxylin-eosin for morphological and morphometric studies.
2.9.2. Morphometry analysis
Hematoxylin-eosin-stained sections of 10 well-oriented villi and crypts of the duodenum and ileum from each animal were photographed with a 4 × lens using a light microscope (Eclipse Ni-U; Nikon, Japan) and used to measure the villus height (VH) and crypt depth (CD). Then, the ratio of the villus height to crypt depth (VH:CD) was calculated.
2.10. Statistical analysis
Data collected from biochemical, immunological and antioxidant assays, the body weight of the animals and egg quality parameters were analyzed using statistical software (SPSS software for Windows, release 23.0., Inc., Chicago, IL, USA). The difference between the means of the experimental groups was calculated using a one-way analysis of variance (ANOVA). Differences were considered statistically significant at P<0.05. Data on morphometric measurements were evaluated for statistical significance by Student’s t-test. P values were two-tailed and a P value <0.05 was significant. The values were expressed as mean ± standard deviation (S.D.).
3. Results and Discussion
3.1. Egg quality
The effect of olive, rosemary, and laurel leaf powder mixture on selected egg quality traits of laying hens during the last days of the experimentation are reported in Table 3.
The dietary supplementation of leaf powder mixture did not affect (P>0.05) the final body weight of the animals (2108 ± 180 g vs. 2182 ± 220 g, for the CON and LPM groups respectively). There were no significant differences between groups (P>0.05) in egg weight, albumen weight, yolk weight, shell weight, shell thickness, yolk color, yolk ratio, albumen ratio, and shell ratio.
To our knowledge, this research is the first study of the effects of dietary supplementation with olive, rosemary, and laurel leaf mixtures in laying hens. Therefore, the discussion of our findings will be made considering similar studies that investigated dietary supplementation with single leaves. Similar results to this study have been reported in previous studies on egg quality traits related to feed supplemented with only olive leaves, at 3% of the diet [30] or 5g/kg diet [75] in laying hens and in Japanese quails (10-20 g/kg diet [76]). Ahmed et al. (2017) [77] reported that no outer and inner egg quality traits were significantly affected by diet supplementation with different levels of oleuropein. As for laurel leaves, their supplementation (2 or 4 g/kg feed) in quails had no effect on body weight, feed intake, or external and internal egg quality traits [78]. Moreover, the addition of rosemary essential oil to the diet did not affect the performance of laying hens [79], and rosemary leaf powder supplementation (5g/kg diet) did non alter egg quality characteristics [80]. In contrast, other studies found beneficial effects of dietary supplementation with leaves or their extracts. The use of olive leaves resulted in an improvement in yolk color [30], and olive leaf extract increased body weight in broilers [81]. The addition of laurel leaves (3g/kg diet) resulted in weight gain in chicken broilers [82]. Feed supplementation with essential oils of rosemary (300 mg/kg diet) improved performance in laying hens [83].
The mode of action of phytochemical additives in the diet of poultry is not yet fully understood. The effectiveness of phytochemical supplementation in diets is linked to certain active compounds, at a well-defined dose, and the change in their concentration may be the cause of a non-response. Ferdous et al. (2019) [84] reported that the efficacy of phytogenics can depend on numerous factors, including composition, inclusion level in the feed, bird genetics, and feed composition. Lipinski et al. (2017) [85] showed that in monogastric animals the dietary strategy does not always have the same effect on body weight and productive performance. Depending on the specific phytochemical supplemented in the diet, it may result in an improvement, deterioration or no change. In our study, one reason for the nonpositive effect of the inclusion of mixed powder in some productive indicators would be the uncontrolled use of mixed powder leaf supplements. Martínez et al. (2013) [86] indicated that the secondary metabolite effect of plants depends on the concentration and relationship of these compounds and their inclusion or supplementation in animal diets.
3.2. Histology of the Small Intestine
The intestinal mucosa consisted of villi, finger-like projections, and basal crypts (Figure 1A,B). Villi were covered by a simple columnar epithelium in which goblet cells were scattered (Figure 1C). The histological observations revealed a clear decrease in villi height from the duodenum to the ileum (Figure 1A, B) in both the CON and LPM fowls, as well as the absence of histological lesions.
Histology measurements of the intestine were performed to monitor the effects of the LPM on villus height or crypt depth. The results of the morphometric analysis of the small intestine are displayed in Table 4.
Leaf powder mixture supplementation induced a significant increase in the VH in the duodenum (1046.57±199.55 µm vs 1121.73±211.15 µm, P<0.05) and ileum (482.44±102.28 µm vs 593.15±148.24 µm, P<0.001). Moreover, LPM supplementation significantly (P<0.001) increased the CD of the duodenum (303.44±11.84 µm vs 245.78±59.35 µm) and the ileum (201.05±77.25 µm vs 148.24±30.58 µm). The treatment induced a decrease of VH:CD in the duodenum (3.69±1.24 vs 4.25±1.39, P<0.05) and in the ileum (2.95±0.61 vs 3.25±1.09, P=0.145).
The histomorphometric analysis revealed that a powder mixture of olive, rosemary, and laurel leaves induces a significant increase in the VH of the duodenum and the ileum of laying hens. The increase of intestinal VH is related to the enhancement of the absorptive surface area, due to a high expression of brush border enzymes capable of greater digestion and absorption of available nutrients [87]. Thus, the findings suggest that the administered LPM exerts some beneficial effects on the absorptive capacity of the intestinal mucosa. The LPM supplementation produced a significant increase in the CD of the duodenum and the ileum. This result justifies the VH increase because the intestinal crypts consist of proliferating cell types that differentiate in villus epithelial cells (see [88] for references).
It has been shown that an increase in the depth of crypts, which serve as a cellular source for intestinal epithelium, is an indicator of cell renewal rate. However, some researchers believe that deep crypts also reflect long villi [89,90], while others argue that increased CD shows faster cell turnover in response to inflammation or loss of epithelial cells [91]. Villus height and crypt depth represent an indirect indication of the maturity and functional capacity of enterocytes; the longer the villi and crypts are, the greater number of enterocytes are present [92]. It is well known that crypt lengthening is mediated by a spectrum of local, immune, and neuro–humoral factors, as well as by diet composition [88,93,94]. It has been reported that CD is directly representative of the intestinal environment and may be used as an indicator of intestinal health [95].
Compared to the control group, the VH:CD ratio in LMP-supplemented laying hens decreased in the duodenum and ileum. There are no studies on the histological effects of adding rosemary and laurel to the diet on the avian intestine, whereas a few studies concerning the effect of adding olive on the small intestine have been published. As in this study, an increase in CD and a lower VH:CD ratio have been found in the duodenum and jejunum of broilers supplemented with olive pomace [96]. This result was interpreted as a presumed increase in epithelial turnover. However, our findings do not completely match the results obtained by Pirman et al. (2021) [16] in broilers fed with olive leaf extract, because in that investigation no difference was detected in the VH:CD ratio of the duodenum, whereas a reduction of the VH:CD ratio was observed in the ileum. It has been suggested that the variation in the results in different trials may depend on the composition of the phytogenic feed additives, level of application or methods for extracting bioactive compounds from herbs [97].
3.3. Concentration of short chain fatty acids (SCFAs) in caecum contents
To describe whether the leaf powder mixture affected gut function, the concentrations of SCFAs were determined (Table 5).
Most of the SCFA analyzed in the caecum chymus varied according to the diet of the laying hens.
The acetate concentration in the LPM group, accounting for the largest proportion of total SCFAs, was significantly higher (P<0.05) than that in the CON group. Likewise, a higher concentration of propionic acid (P<0.05), butyric acid (P<0.05) and isobutyric acid (P<0.01) were found in the LPM group compared to the CON group. No significant differences in the levels of isocaproic and heptanoic acids were observed in the caecal contents between the LPM group and the CON group.
In this study, the acetate production was the highest, followed by propionate and butyrate which showed the lowest cumulative production, in agreement with other researchers [98,99]. The butyrate concentration was lower compared to propionate, in contrast to other studies [95,100]. Isobutyric acid was significantly higher (P<0.01) in the LPM group than in the CON group. It belongs to branched short-chain fatty acids that are mainly produced during the fermentation of branched-chain amino acids (valine, leucine, and isoleucine) by the intestinal microbiota, mainly by genera Bacteroides and Clostridium [101]. It has been observed that Bacteroides fragilis in an environment with slowly fermentable carbon sources can shift its metabolism towards protein fermentation [102].
In the biosynthesis of SCFAs from dietary fiber and carbohydrate fermentation, different mechanisms are involved: acetate via the Wood–Ljungdahl pathway or acetyl-CoA; two molecules of acetyl-CoA for butyrate production; from the acrylate, succinate, and propanediol pathways for propionate [103]. The production of propionate [104] and acetate seems to be associated with the increased presence of Bacteroides spp. while butyrate is produced mainly by the Firmicutes phylum [105]. Bifidobacterium and Lactobacillus produce acetic and lactic acid in large quantities. Firmicutes species decompose polysaccharides and produce butyrate, and Bacteroidetes species degrade complex carbohydrates and mainly synthesize propionate [106,107]. Furthermore, it should be noted that cross-metabolism between microorganisms also greatly affects the final balance of intestinal SCFAs [102]. The formation of butyrate from acetate or lactate is considerably lower than the conversion of butyrate into propionate and is very scarce between propionate and acetate [108].
SCFAs influence intestine functionality. SCFAs are an important source of energy for enterocytes: up to 50% of their daily requirement [109,110]. SCFAs represent the main luminal anions; their increase reduces the pH to about 4.8. Acetic acid suppresses intestinal apoptosis and promotes mucin production [111]. The three main SCFAs affect intestinal epithelial cell function, promoting intracellular permeability by modifying the expression or distribution of the tight junction and zonulin [112]. Acetate activates the nucleotide-binding oligomerization domain 3 (NLRP3) inflammasome [113] and promotes intestinal barrier integrity [114]. Butyrate improves the balance of the gut microbiota [115], enhances intestinal barrier function [116], and increases mucin activity via MUC2 gene expression [117]. The protective effects of propionate are also evident in the intestinal barrier [118]. The short-chain fatty acids derived from the intestine are assimilated in the metabolism of host carbohydrates and lipids [108]. SCFAs that are not metabolized in the gut enter the portal circulation. When they arrive in the liver, SCFAs become substrates for the energy production of hepatocytes via oxidation [119]; propionate can become a substrate for gluconeogenesis [120]. Acetate is also used for the synthesis of fatty acids and cholesterol [121].
SCFAs are among the most important products of microbial fermentation in the gut [122,123] and their concentrations in the caecum may vary according to the dominant microbiota [98]. The caecum is the main site for microbial fermentation, where undigested carbohydrates are transformed into short-chain fatty acids [123].
The caecum is considered the fundamental part of the distal intestine, due to the highest concentration of intestinal microorganisms that affect health and performance [124]. The microbial community plays its role in digesting complex nutrients and protecting poultry from pathogenic colonization via a process of competitive exclusion [125]. In this study, the LPM may have had beneficial effects at the microbiota level.
The microbial composition can be modulated by polyphenol content in leaves, plants, and their extracts. The control mechanisms of the microbial community via plant extracts and dietary supplements (polyphenols, herbs, spices) are not well defined. Their action is supposed to take place through antimicrobial activity against pathogens [126] or by improving the metabolic function of microbes, such as protein and amino acid metabolism and lipid biosynthesis [127]. Rosemary extract is rich in carnosic acid, is able to modulate the microbiota of the caecum and the fecal excretion of SCFAs [128], and reduces the Firmicutes/Bacteroides ratio [129]. The major constituent of laurel leaf, 1,8 cineol, is responsible for its antibacterial activity [130]. It has been reported that olive leaf extract significantly stimulates the abundance of beneficial Lactobacillus and Bifidobacterium spp. as well as reducing the abundance of Escherichia coli in the caecum [131,132]. E. coli is commonly referred to as colibacillosis. It has been hypothesized that the inhibition of the growth of E. coli is attributable to the ability of polyphenols to bind to cell membranes and to disturb their function by altering the permeability and the driving force of the proton through the loss of H+ -ATPasis and functions related to the membrane [133,134].
The positive effect of phenols on bacterial growth, such as the beneficial Lactobacillus and Bifidobacterium spp., is due to their allowing these phenols to be used as substrates. Lactobacilli can metabolize polyphenols in the growth process, and polyphenols in turn provide energy to cells. Moreover, polyphenols can intensify the consumption of nutrients, such as carbohydrates [135].
The bacterial population includes the beneficial bacteria Lactobacillus spp. and Bifidobacteria spp.; Salmonella spp. and Campylobacter spp., which are pathogenic to humans; and Escherichia coli and Clostridium [124,136], which are pathogenic to poultry.
The mechanism by which the intestinal microflora may reduce Enterobacteriaceae (including Salmonella) is the bacteriostatic effect of SCFAs in the caecum [137]. Beneficial Lactobacillus spp. is known for the secretion of lactic acid, which lowers intestinal pH and inhibits pathogenic growth [138].
Butyrate and propionate have inhibitory effects on Salmonella, which frequently infects poultry farms [139,140]. Sun and O’Riordan (2013) [141] reported that SCFAs at high concentrations disrupt metabolic reactions in bacteria when small, non-ionized acids cross bacterial membranes and dissociate into protons and anions in the cytoplasm. The protons lead to the acidification of intracellular compartments, disturbing metabolic reactions, and anions disrupt osmotic balance. In addition, small peptides or proteins secreted by beneficial bacteria kill bacteria by forming pores in the cell membrane and disrupting enzymatic function.
3.4. Biochemical parameters
Blood represents the main index for evaluating the physiological, nutritional, and general pathological status. Biochemical serum constituents showed significant differences between dietary treatments (Table 6). The results indicated that dietary inclusion of 6g/kg feed of the LPM resulted in a significant increase of serum total protein (P<0.05) and tended to increase the triglyceride levels (P<0.10) compared to the control. Moreover, supplementation with the LPM significantly decreased blood total cholesterol (P<0.05) and LDL cholesterol (P<0.01), while HDL cholesterol was significantly higher (P<0.01) compared to the CON. Briefly, these results indicate that adding the LPM to the diet can reduce the blood levels of cholesterol, triglycerides and LDL. There was no significant difference (P>0.05) between the groups in the serum content of NEFA.
The higher values of blood proteins affected by the administration of the LPM may reflect the improvement of digestibility and metabolism, as previously reported [142].
As regards the effects of dietary phytochemical natural additives, olive leaf supplementation caused a decrease in blood cholesterol levels in Japanese quails [143] and in broiler chicks [81]. According to Talhaoui et al. (2015) [144], the triacetylated hydroxytyrosol and hydroxytyrosol content of olive leaves has a hypolipidemic effect and lowers total serum cholesterol, triglycerides, and low-density lipoprotein cholesterol (LDL-C), and their antioxidant activities prevent the lipid peroxidation process. The oleuropein and hydroxytyrosol found in olive leaves are known to prevent LDL oxidation and to inhibit 3-hydroxy-3-methyglutaryl coenzyme A, which plays an important role in cholesterol synthesis in the liver [30]. Ahmed et al. (2017) [77] indicated that the hypocholesterolemic action of phenolic compounds of olive leaves may be due to the inhibition of dietary cholesterol absorption in the intestine, or its production by the liver or stimulation of the biliary secretion of cholesterol and cholesterol excretion in the feces. Elazab et al. (2022) [145] reported that a possible explanation for the reduction of cholesterol and triglycerides is the stimulation of the conversion of cholesterol into bile acids that are excreted from the body through enterohepatic circulation. Torki et al. (2018) [146] reported that adding essential oils of rosemary to the diet of laying hens decreased the serum concentration of triglyceride and serum cholesterol. Radwan et al. (2008) [147] showed that adding rosemary to the diet decreased total lipid, total cholesterol and LDL cholesterol in hens. Bolukbasi et al. (2008) [148] reported that adding essential oils of rosemary reduced serum cholesterol and triglyceride in laying hens. Chalabi et al. (2020) [149] reported a serum reduction of TC, TG, LDL, and increased HDL in rats treated with ethanolic extract from laurel leaves. Ali and Al-Shuhaib (2021) [82] observed that the addition of crushed laurel leaves (2-3 g/kg of feed) to the diet led to an increase in the concentration of HDL and a decrease in TC, TG, LDL in broilers.
Torki et al. (2018) [146] suggest that the mechanism by which phytochemical additives can reduce cholesterol is by reducing lipid absorption in the intestine by binding bile acids, which results in increasing lipid excretion and reducing blood lipid concentration. The reduction of serum cholesterol can occur through a reduction in the denovo synthesis of cholesterol or by improving the excretion of bile acids. The conversion of cholesterol to bile acids through the stimulation of the hepatic activity of cholesterol-7-hydroxylase, is a key pathway for the removal of cholesterol from the body [150]. Altogether, these effects would lead to a reduction in serum cholesterol concentrations. Accordingly, the current findings suggest that the foliar mixture (olive, rosemary, laurel) can improve the lipid profile of laying hens.
The current study revealed that the addition of the LPM to the hens’ diet had no adverse effect on egg yolk cholesterol content (9.63 vs 9.59 mg/g yolk; data not shown). These results indicate a non-consecutive response to blood cholesterol levels and the cholesterol content of egg yolk and agree with the findings of other authors [151,152]. Cholesterol deposition in the egg yolk can be affected by nutrition [153,154], while different studies indicate negligible effects [7,155]. Furthermore, other studies showed that yolk cholesterol content appears constant; it cannot vary and is independent of dietary factors [156]. The yolk cholesterol level cannot be changed because it is required to ensure embryo development [157]. The liver is the main site of cholesterol synthesis and accumulates cholesterol from the blood when it synthesizes lipoproteins. Plasma LDL is the main carrier transporting endogenous cholesterol. According to Griffin (1992) [158], yolk precursors are synthesized in the liver of the laying hen and transported in the plasma to the ovary. Therefore, the cholesterol content of the yolk is primarily dependent on the cholesterol content of triglyceride-rich lipoproteins. The inhibition of cholesterol synthesis can reduce the rate of synthesis and secretion of lipoproteins by the liver but has little effect on the composition of the lipoproteins that are secreted. Fennema (1993) [159] stated that variations in total yolk lipid content are more influenced by bird genetic strain than a diet.
3.5. Oxidative status
The effect of the dietary LPM supplement on the oxidative status of hens is reported in Table 7.
Serum levels of total oxidative stress (TOS) and ROMs were significantly lower (P<0.001) in the hens treated with the LPM compared with the control. The LPM group also showed a higher total antioxidant status (TAS) than the control (469.71 vs 393.78 umol TRX/L), but the differences were not statistically significant (P>0.05).
The independent evaluation of TOS or TAS may not accurately reflect the oxidative stress, due to a dynamic balance between TOS and TAS (Wu et al., 2017). TOS represents the oxidizing state of animal/patient and is usually estimated through direct measurement of relatively stable ROS family members, such as hydrogen peroxide (H2O2), peroxyl radicals, and DNA damage markers. The measurement of only one of the oxidant or antioxidant parameters does not provide proper information about the oxidative status. TOS and TAS contribute to the overall health status [160] and are indicative of the physiological status of animals [161]. For this reason, in our study, TOS, TAS, and ROMs were measured in serum samples for an overall evaluation of the physiological status of the animals.
One of the objectives of this study was to evaluate the effects of the LPM blend of olive, rosemary and laurel on the oxidative state of the hens. The polyphenol content was highest in olive leaves, followed by rosemary and laurel (Table 1), in accordance with the results of other research [36,162,163]. It has been shown that most phenolic compounds possess antioxidant activity in vitro and in vivo. Wenk (2002) [164] reported that the increase in performance associated with the supplementation of poultry diets with plant-origin materials is mainly due to their polyphenolic contents.
In this study, the antioxidant activity of the LPM could be attributed to their polyphenol constituents and mainly to oleuropein and its derivative hydroxytyrosol, for olive leaves [24,25], to 1.8 cineole, for laurel leaves [32,165], and to the antioxidant properties of carnosol and carnosic acid, for rosemary [51,166]. Most phenolic compounds in the olive leaves or olive leaf extract showed antioxidant activity in vitro [167] and in vivo [23]. This activity in the olive leaf extract manifests through its standard one-electron reduction potential by either donating hydrogen or electrons or by breaking the free radical chain reaction and by preventing the chelation of metal ions [168]. Turkez et al. (2012) [169] reported that olive leaf extract showed beneficial influences against the genetic and oxidative damage of permethrin by increasing TAS and reducing TOS in rats. Oleuropein supplementation increased TAS and reduced TOS in quails subjected to heat stress [170].
In this study, LPM also led to numerically higher TAS serum levels in hens. Moreover, the lower ROM serum levels observed in treated hens (Table 7) express the capability of the LPM to reduce free radical production and oxidative stress and are associated with lower TOS levels. Falade et al. (2022) [163] found that the ability of aqueous extracts of rosemary and laurel leaves to inhibit TBARs can protect the brain from the damaging effect of free radicals [17]. The content of total antioxidant capability was significantly increased with rosemary or laurel leaf supplementation in lambs [172] and rabbits [145].
On the other hand, mixed proportions of herbs and plants would influence the antioxidant properties [173] because the total antioxidant capacity is attributed to their additive and/or synergistic effect [57], probably in line with the results obtained in this study. However, the underlying mechanism of the action caused by the olive, rosemary, and laurel leaves is needed for future research.
3.6. Immunomodulatory effects
In our study, LPM supplementation decreased IL-6 and IL-1 (P<0.01) and TNF-α (P<0.05) concentrations compared to the CON (Table 8).
The results suggested that the leaf powder blend (olive, rosemary, laurel) supplementation suppressed systemic inflammation in hens.
Inflammation represents a common final pathway related to oxidative processes. The molecular mechanisms of the interaction between oxidative stress and inflammation and their interaction with cell death are unclear [174]. It has been suggested that oxidative stress increases apoptosis through several signaling pathways activated by permeabilization of the mitochondrial membrane, which induces activation of caspase-3 [175]. Antioxidant properties and the immunological effect of phytochemical dietary additives merit a lot of attention because strong oxidation affects the health of laying hens.
Adipokines are involved in the regulation of several physiological processes [176], including control of oxidative stress [177]. Miliaraki et al. (2022) [178], in an oxidant/antioxidant pathway, reported an expression of the cytokines TNF-α and IL-6 strongly correlated with an increase in the TOS/TAC ratio. In agreement with our results, it seems that the leaf powder blend supplementation in the laying hens’ diet was effective in enhancing the antioxidant ability and immune system of birds.
Our results agree with those relating to the separate use of the individual plants in terms of the effects on the immune system. Vezza et al. (2017) [179] showed that olive leaf extract decreased the levels of inflammatory markers such as IL-1, TNF-α and iNOS in mice models of colitis and reduced the production of proinflammatory mediators (IL-1, IL-6, and TNF-α) in intestinal mucosal samples from Crohn’s disease patients. De Cicco et al. (2020) [180] revealed that olive leaf extract suppressed the induction of pro-inflammatory mediators: tumor necrosis factor (TNF-α) and interleukin IL-6 and IL-1β, promoting the shift of M1 macrophage toward less inflammatory M2-cells via the modulation of the associated NF-κB and proliferator-activated receptor gamma (PPARγ) signaling pathways. Kaneko et al. (2019) [181] reported that olive leaf extract has an inhibitory effect on NLRP3 inflammasome activation and pro-IL-1β protein expression, a multiprotein complex that plays a pivotal role in regulating the innate immune system and inflammatory signaling. Lee et al. (2019) [182] noted that 1,8-cineole of laurel leaves reduced the expression of proinflammatory cytokines by suppressing NLRP3 inflammasome activation. Farouk et al. (2022) [183] reported a significant increase in immunoglobulin G with a significant decline in IL-6 level in broilers treated with rosemary leaf powder (5 g/kg ration). Tsai et al. (2013) [184] reported that ethanolic rosemary extract treatment, in the progression of inflammation, significantly suppressed the secretion and mRNA expression of proinflammatory cytokines, including IL-1β and TNF-α, due to diminished nuclear factor kappa-B (NF-κB) activation and Toll-like receptor (TLR2) mediated signaling pathways.
The blend of phytochemicals improves the immune system. Aroche et al. (2018) [185] indicate that dietary inclusion of 0.50% of a mixed powder of plant leaves of Anacardium occidentale (60%), Psidium guajava (20%), and Morinda citrifolia (20%) enhances the immune status of broiler chickens.
3.7. Liver enzymes
The effects of experimental treatments on liver enzymes are presented in Table 9.
In hens supplemented with the LPM, significant differences in the serum activity of ALT, AST (P<0.01), and ALP (P<0.05) were observed compared to the control group. There were no significant differences between groups in bilirubin levels.
There is little information in the literature on the dietary effects of plant leaves and spices on liver enzymes in animals. Measurement of the serum activities of AST, ALT, and ALP, indicative of liver injury, is a valuable tool in determining a safe inclusion rate of non-conventional feed or feed additives in birds [186]. The high activity of AST, ALP, and ALT in the blood is a bioindicator of liver damage, and this is evidence of the existence of liver injury due to toxicity [187].
The results of this study indicate that a blend of leaf powder (olive, rosemary, and laurel) can have a hepatoprotective effect in hens. This finding is compatible with many studies that explain the significant decrease of AST, ALT, and ALP due to the free radical scavenging activity of antioxidant phytochemicals in plant leaves or extracts. Khalil (2004) [188] reported on the hepatoprotective activity of an aqueous extract of olive leaves, with a significant decrease in the serum content of ALT, ALP, and AST, attributed to their antioxidant properties, which can protect the liver from damage. Abdel-Wahhab et al. (2011) [189] examined the chemoprotective effect of rosemary extract in Wistar albino rats, improving all biochemical parameters and the liver’s histological picture. Al-Attar and Shawush (2015) [190] showed that the extracts of olive and rosemary leaves possess hepatoprotective properties against liver cirrhosis induced by thioacetamide by inhibiting the physiological and histopathological alterations attributed to their antioxidant activities. Treatment with laurel leaf extract resulted in a decrease in ALT, AST, and ALP in diabetic rats compared to the control [17,191]. Abdel-Azeem et al. (2018) [192] reported that laurel leaf supplementation in rabbits’ diets showed a significant decrease in AST and ALT blood plasma.
Thus, the hepatoprotective activity of the foliar powder mixture recorded in this study may be owing to the antioxidant properties of its natural components, present in olive and rosemary leaves, their combination [190], and in laurel leaves [17,191]. Ultimately, the phytochemical additives of the leaf powder mixture (olive, rosemary, and laurel) have beneficial effects on liver functions.
4. Conclusions
This is the first study on the dietary use of the mixture of olive, rosemary, and laurel leaves, and the effects on performance, egg quality, blood biochemical, and gut morphophysiological parameters in laying hens. In this study, neither body weight nor egg quality parameters were influenced by the LPM supplementation. The results showed that the addition of 6g/kg feed of LPM (2g/kg olive + 2g/kg laurel + 2g/kg rosemary) to the diet improved antioxidant activity by decreasing serum TOS and a tendential decrease of TAS and ROM levels.
The synergy among the LPM constituents also improved the immune system, by reducing the production of pro-inflammatory cytokines: IL-6, TNF-α and IL-1β. Likewise, adding the LPM to the diet improved the lipid profile, reducing total cholesterol and LDL cholesterol and increasing HDL cholesterol. Moreover, adding LPM to the diet showed an improvement in liver enzyme activity by reducing the ALP, AST, and ALT serum levels.
The significant increase of SCFAs (acetic, propionic, butyric, and isobutyric, acids) established in the caecum were found in the LPM-treated group.
The results showed that LPM supplementation exerted synergistic effects on intestinal morphology by promoting villi height, crypt depth and improving physiological activity in the intestinal tract by increasing SCFA production.
Taken together, the results indicate that adding the LPM to the laying hens’ diet was effective in enhancing the antioxidant ability and immune system of the birds, the serum lipid profile, and the functionality of liver and the intestinal tract. This could be the key to increasing the health and integrity of farm animals raised without antibiotics.
These findings provide new insights into the dietary use of a leaf powder combination of olive, laurel and rosemary. However, the underlying mechanism of the action caused by the olive, rosemary, and laurel leaves is needed for future research to optimize their use in the diet of laying hens. Due to the wide dissemination and low cost of these plants, this study is an attempt to promote a sustainable avian food industry.
Author Contributions
Conceptualization, Angela Gabriella D'Alessandro and Giovanni Martemucci; Formal analysis, Angela Gabriella D'Alessandro; Funding acquisition, Angela Gabriella D'Alessandro; Investigation, Angela Gabriella D'Alessandro, Salvatore Desantis, Giuseppe Fracchiolla, Riccardo Porrelli and Giovanni Martemucci; Methodology, Angela Gabriella D'Alessandro, Salvatore Desantis, Giuseppe Fracchiolla, Riccardo Porrelli and Giovanni Martemucci; Writing – original draft, Angela Gabriella D'Alessandro, Salvatore Desantis and Giovanni Martemucci; Writing – review & editing, Angela Gabriella D'Alessandro, Salvatore Desantis, Giuseppe Fracchiolla, Riccardo Porrelli and Giovanni Martemucci. All authors have read and agreed to the published version of the manuscript.
Funding
This research was co-financed by the European Union, European Regional Development Funds and by National Funds of Greece and Italy, Interreg V-A Greece–Italy 2014–2020. Project title: “Innovative use of olive, winery and cheese waste by products in animal nutrition for the production of functional foods from animals (Inno.trition)” (Mis. Code: 5003778).
Institutional Review Board Statement
This study was performed to avoid any animal suffering according with the Italian Legislative Decree 26/2014.
Data Availability Statement
All data are available in the present manuscript.
Acknowledgments
The authors wish to thank Dr Francesco Vizzarri for his assistance with lab analysis and data collection, and the Department of Pharmacy-Drug Sciences, University of Bari Aldo Moro, Bari, 70125, Italy for the use of TECAN infinite M1000Pro plate reader.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abdelli, N.; Solà-Oriol, D.; Pérez, J.F. Phytogenic feed additives in poultry: achievements, prospective and challenges. Animals 2021, 11, 3471. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Salem, H.M.; El-Tahan, A.M.; Soliman, M.M.; Youssef, G.B.A.; Taha, A.E-; Soliman, S.M-; Ahmed, A.E-; El-Kott, A.F-; Al Syaad, K.M.; Swelum, A.A. Alternatives to antibiotics for organic poultry production: types, modes of action and impacts on bird's health and production. Poult. Sci. 2022, 101, 101696. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, S.; Banday, M.T.; Shakeel, I.; Adil, S.; Mir, M.S.; Beigh, Y.A.; Amin, U. Histomorphological studies of broiler chicken fed diets supplemented with either raw or enzyme treated dandelion leaves and fenugreek seeds. Vet. World 2016, 9, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Gadde, U.; Kim, W.H; Oh, S.T.; Lillehoj, HS. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: a review. Anim. Health. Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef] [PubMed]
- Seidavi, A.; Tavakoli, M.; Asroosh, F.; - Scanes, C.G.; Abd El-Hack, M.E.; Naiel, M.A.E.; Taha, A.E.; Aleya, L.; El-Tarabily, K.A.; Swelum, A.A. Antioxidant and antimicrobial activities of phytonutrients as antibiotic substitutes in poultry feed. Environ. Sci. Pollut. Res. Int. 2022, 2, 5006–5031. [Google Scholar] [CrossRef]
- Chemat, F.; Rombaut, N.; Sicaire, A.G.; Meullemiestre, A.; Fabiano-Tixier, A.S.; Abert-Vian, M. Ultrasound assisted extraction of food and natural products. Mechanisms, techniques, combinations, protocols and applications. A review. Ultrason. Sonochem 2017, 34, 540–560. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.E.; Shehata, A.M.; Alzahrani, S.O.; Shafi, M.E.; Mesalam, N.M.; Taha, A.E.; Swelum, A.A.; Arif, M.; Fayyaz, M.; Abd El-Hack, M.E. The role of polyphenols in poultry nutrition. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1851–1866. [Google Scholar] [CrossRef]
- Luo, J.; Song, J.; Liu, L.; Xue, B.; Tian, G.; Yang, Y. Effect of epigallocatechin gallate on growth performance and serum biochemical metabolites in heat-stressed broilers. Poult. Sci. 2018, 97, 599–606. [Google Scholar] [CrossRef]
- Simitzis, P.; Spanou, D.; Glastra, N.; Goliomytis, M. Impact of dietary quercetin on laying hen performance, egg quality and yolk oxidative stability. Anim. Feed. Sci. Technol. 2018, 239, 27–32. [Google Scholar] [CrossRef]
- Surai, P. Polyphenol compounds in the chicken/animal diet: from the past to the future. J. Anim. Physiol. Anim. Nutr. 2014, 98, 19–31. [Google Scholar] [CrossRef]
- Petti, S.; Scully, C. Polyphenols, oral health and disease: A review. J. Dent. 2009, 37, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Galli, G.M.; Da Silva, A.S.; Biazus, A.H.; Reis, J.H.; Boiago, M.M.; Topazio, J.P.; Migliorini, M.J.; Guarda, N.S.; Moresco, R.N.; Ourique, A.F.; Santos, C.G.; Lopes, L.S.; Baldissera, M. D.; Stefani, L.M. Feed addition of curcumin to laying hens showed anticoccidial effect, and improved egg quality and animal health. Res. Vet. Sci. 2018, 118, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Abou-Elkhair, R.; Abdo Basha, H.; Abd El Naby, W.; Ajarem, J.S.; Maodaa, S.N.; Allam, A.A.; Naiel, M.A.E. Effect of a diet supplemented with the Moringa oleifera seed powder on the performance, egg quality, and gene expression in Japanese laying quail under heat stress. Animals 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Oviedo-Rondon, E.O. Holistic view of intestinal health in poultry. Anim. Feed. Sci. Technol. 2019, 250, 1–8. [Google Scholar] [CrossRef]
- Dosoky, WM.; Zeweil, H.S.; Ahmed, M.H.; Zahran, S.M.; Shaalan, M., M.; Abdelsalam, N.R.; Abdel-Moneim, A.E.; Taha. AE.; El-Tarabily, K.A.; Abd El-Hack, M.E. Impacts of onion and cinnamon supplementation as natural additives on the performance, egg quality and immunity in laying Japanese quail. Poult. Sci. 2021, 100. [Google Scholar] [CrossRef] [PubMed]
- Pirman, T.; Rezar, V.; Vrecl, M.; Salobir, J.; Levart, A. Effect of olive leaves or marigold petal extract on oxidative stress, gut fermentative activity, and mucosa morphology in broiler chickens fed a diet rich in n-3 polyunsaturated fats. J. Poult. Sci. 2021, 58, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, R.R.; Omer, A.K.; Yener, Z.; Uyar, A.; Ahmed, A.K. Biomedical effects of Laurus nobilis L. leaf extract on vital organs in streptozotocin-induced diabetic rats: Experimental research. Ann. Med. Surg. 2021, 61, 188–197. [Google Scholar] [CrossRef]
- Delgado Pertinez, M.; Chesson, A.; Provan, G.J.; Garrido, A.; Gomez Cabrera, A. Effect of different drying systems for the conservation of olive leaves on their nutritive value for ruminants. Ann. Zootechn. 1998, 47, 141–150. [Google Scholar] [CrossRef]
- Şahin, S.; Bilgin, M. Olive tree (Olea europaea L.) leaf as a waste by-product of table olive and olive oil industry: a review. J. Sci. Food Agric. 2018, 98, 1271–1279. [Google Scholar] [CrossRef]
- Contreras, M.D.M.; Lama-Muñoz, A.; Espínola, F.; Moya, M.; Romero, I.; Castro, E. Valorization of olive mill leaves through ultrasound-assisted extraction. Food Chem. 2020, 314. [Google Scholar] [CrossRef]
- Bulotta, S.; Corradino, R.; Celano, M.; D’Agostino, M.; Maiuolo, J.; Oliverio, M.; Procopio, A.; Iannone, M.; Rotiroti, D.; Russo, D. Antiproliferative and antioxidant effects on breast cancer cells of oleuropein and its semisynthetic peracetylated derivatives. Food Chem. 2011, 127, 1609–1614. [Google Scholar] [CrossRef]
- Cavaca, L.A.; López-Coca, I.M.; Silvero, G.; Afonso, C.A. The olive-tree leaves as a source of high-added value molecules: Oleuropein. Stud. Nat. Prod. Chem. 2020, 64, 131–180. [Google Scholar]
- Jemai, H.; Fki, I.; Bouaziz, M.; Bouallagui, Z.; El Feki, A.; Isoda, H.; Sayadi, S. Lipid-lowering and antioxidant effects of hydroxytyrosol and its triacetylated derivative recovered from olive tree leaves in cholesterol-fed rats. J. Agric. Food Chem. 2008, 56, 2630–2636. [Google Scholar] [CrossRef]
- Benavente-García, O.; Castillo, J.; Lorente, J.; Ortuño, A.D.R.J.; Del Rio, J.A. Antioxidant activity of phenolics extracted from Olea europaea L. leaves. Food Chem. 2000, 68, 457–462. [Google Scholar] [CrossRef]
- Erbay, Z.; Icier, F. The importance and potential uses of olive leaves. Food Rev. Int. 2010, 26, 319–334. [Google Scholar] [CrossRef]
- Huang, Y.L.; Oppong, M.B.; Guo, Y.; Wang, L.Z.; Fang, S.M.; Deng, Y.R.; Gao, X.M. The Oleaceae family: a source of secoiridoids with multiple biological activities. Fitoterapia 2019, 136, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Nediani, C.; Ruzzolini, J.; Romani, A.; Calorini, L. Oleuropein, a bioactive compound from Olea europaea L., as a potential preventive and therapeutic agent in non-communicable diseases. Antioxidants 2019, 8, 1–26. [Google Scholar] [CrossRef]
- Romani, A.; Mulas, S.; Heimler, D. Polyphenols and secoiridoids in raw material (Olea europaea L. leaves) and commercial food supplements. Eur. Food Res. Technol. 2017, 243, 429–435. [Google Scholar] [CrossRef]
- El-Damrawy, S.Z.; Khalifah, M.M.; Fares, W.A. Dietary olive leaf and antioxidative status in chickens “performance, some physiological traits and immunological responses of Mandarah chicks supplemented olive leaves powder in their diets". Egypt. Poult. Sci. J. 2013, 33, 279–287. [Google Scholar]
- Cayan, H.; Erener, G. Effect of olive leaf (Olea europaea) powder on laying hens performance, egg quality and egg yolk cholesterol levels. Asian-Australas J. Anim. Sci. 2015, 28, 538–43. [Google Scholar] [CrossRef]
- Bahşi, M.; Ciftci, M.; Simsek, U.G.; Azman, M.A.; Özdemir, G.; Yilmaz, Ö.; Dalkiliç, B. Effects of olive leaf extract (oleuropein) on performance, fatty acid levels of breast muscle and some blood parameters in Japanese quail (Coturnix coturnix Japonica) reared in different stocking densities. Ankara Univ. Vet. Fak. Derg 2016, 63, 61–68. [Google Scholar]
- Singletary, K. Bay leaf: Potential health benefits. Nutr. Today 2021, 56, 202–208. [Google Scholar] [CrossRef]
- Dias, M.I.; Barros, L.; Dueñas, M.; Alves, R.C.; Oliveira, M, B.; Santos-Buelga, C.; Ferreira, I.C. Nutritional and antioxidant contributions of Laurus nobilis L. leaves: would be more suitable a wild or a cultivated sample? Food Chem. 2014, 156, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Statti, G.; Uzunov, D.; Menichinia, F. Comparative chemical composition and antioxidant activities of wild and cultivated Laurus nobilis L. leaves and Foeniculum vulgare subsp. piperitum (Ucria) Coutinho seeds. Biol. Pharm. Bul. 2006, 29, 2056–2064. [Google Scholar] [CrossRef] [PubMed]
- Carocho, M.; Ferreira, I.C.F.R. A review on antioxidants, prooxidants and related controversy: natural and synthetic compounds, screening and analysis methodologies and future perspectives. Food Chem. Toxicol. 2013, 51, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Škerget, M.; Kotnik, P.; Hadolin, M.; Hraš, A.R.; Simonič, A.M.; Knez, Ž. Phenols, proanthocyanidins, flavones and flavonols in some plant materials and their antioxidant activities. Food Chem. 2005, 89, 191–198. [Google Scholar] [CrossRef]
- Lu, M.; Yuan, B.; Zeng, M.; Chen, J. Antioxidant capacity and major phenolic compounds of spices commonly consumed in China. Food Res. Intern. 2011, 44, 530–536. [Google Scholar] [CrossRef]
- Broadhurst, C.L.; Polansky, M.M.; Anderson, R.A. Insulin-like biological activity of culinary and medicinal plant aqueous extracts in vitro. J. Agric. Food Chem. 2000, 48, 849–852. [Google Scholar] [CrossRef]
- Khan, A.; Zaman, G.; Anderson, R.A. Bay leaves improve glucose and lipid profile of people with type 2 diabetes, J. Clin. Biochem. Nutr. 2009, 44, 52–56. [Google Scholar] [CrossRef]
- Kilic, A.; Hafizoglu, H.; Kollmannsberger, H.; Nitz, S. Volatile constituents and key odorants in leaves, buds, flowers and fruits of Laurus nobilis L. J. Agr. Food Chem. 2004, 52, 1601–1606. [Google Scholar] [CrossRef]
- Said, CM.; Hussein, K. Determination of the chemical and genetic differences of Laurus collected from three different geographic and climatic areas in Lebanon. Eur. Sci. J. 2014, 2, 412–419. [Google Scholar]
- Ramos, C.; Teixeira, B.; Batista, I.; Matos, O.; Serrano, C.; Neng, N.R.; Nogueira, J.M.F.; Nunes, M.L.; Marques, M. Antioxidant and antibacterial activity of essential oil and extracts of bay leave Laurus nobilis Linnaeus (Lauraceae) from Portugal. Nat. Prod. Res. 2012, 6, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Taban, A.; Saharkhiz, M.J.; Niakousari, M. Sweet bay (Laurus nobilis L.) essential oil and its chemical composition, antioxidant activity and leaf micromorphology under different extraction methods. Sustain. Chem. Pharm. 2018, 9, 12–18. [Google Scholar] [CrossRef]
- Ulbricht, C.; Abrams, TR.; Brigham, A.; Ceurvels, J.; Clubb, J.; Curtiss, W.; DeFranco Kirkwood, C.; Giese, N.; Hoehn, K.; Iovin, R.; Isaac, R.; Rusie, E.; Grimes Serrano, J.M.; Varghese, M.; Weissner, W.; Windsor, R.C. An evidence-based systematic review of rosemary (Rosmarinus officinalis) by the Natural Standard Research Collaboration. J. Diet. 2010, 7, 351–413. [Google Scholar] [CrossRef] [PubMed]
- Borrás-Linares, I.; Stojanović, Z.; Quirantes-Piné, R.; Arráez-Román, D.; Švarc-Gajić, J.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Rosmarinus officinalis leaves as a natural source of bioactive compounds. Int. J. Mol. Sci. 2014, 15, 20585–20606. [Google Scholar] [CrossRef] [PubMed]
- Rašković, A.; Milanović, I.; Pavlović, N.; Ćebović, T.; Vukmirović, S.; Mikov, M. Antioxidant activity of rosemary (Rosmarinus officinalis L.) essential oil and its hepatoprotective potential. BMC Complement Altern. Med. 2014, 14, 225. [Google Scholar] [CrossRef] [PubMed]
- Rizwana, H.; Bokahri, N.A.; Rashed, S.A.; Shehri, S.A.; Awad, M.A.; Merghani, N.; Tabasuum, H. Characterizing silver nanoparticles biosynthesized from Salvia rosmarinus and assessing their in vitro antifungal and cytotoxic activities against phytopathogens and cervical cells. J. Anim. Plant Sci. 2022, 32, 764–774. [Google Scholar]
- Aguilar, F.; Autrup, H.; Barlow, S.; Castle, L.; Crebelli, R.; Dekant, W.; Engel, Kh.; Gonard, N.; Gott, D.; Grilli, S. Use of rosemary extracts as a scientific food additive of the panel on food additives, aromas, technological and material adjuvants in contact with food. Efsa. J. 2008, 721, 1–29. [Google Scholar]
- Rice-Evans, C.A.; Miller, N.J.; Bolwell, P.G.; Bramley, P.M.; Pridham, J.B. The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free Radical Res. 1995, 22, 375–383. [Google Scholar] [CrossRef]
- Angioni, A.; Barra, A.; Cereti, E.; Barile, D.; Coïsson, J. D.; Arlorio, M.; Dessi, S.; Coroneo, V.; Cabras, P.C. Chemical composition, plant genetic differences, antimicrobial and antifungal activity investigation of the essential oil of Rosmarinus officinalis L. J. Agric. Food. Chem. 2004, 52, 3530–3535. [Google Scholar] [CrossRef]
- Aruoma, OI.; Halliwell, B.; Aeschbach, R.; Löligers, J. Antioxidant and pro-oxidant properties of active rosemary constituents: carnosol and carnosic acid. Xenobiotica 1992, 22, 257–68. [Google Scholar] [CrossRef] [PubMed]
- Wijeratne, S.S.; Cuppett, S.L. Potential of rosemary (Rosemarinus officinalis L.) diterpenes in preventing lipid hydroperoxide-mediated oxidative stress in Caco-2 cells. J. Agric. Food Chem. 2007, 55, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Shan, B.; Cai, YZ.; Sun, M.; Corke, H. Antioxidant capacity of 26 spice extracts and characterization of their phenolic constituents. J. Agric. Food Chem. 2005, 53, 7749–7759. [Google Scholar] [CrossRef]
- de Falco, B.; Grauso, L.; Fiore, A.; Bonanomi, G.; Lanzotti, V. Metabolomics and chemometrics of seven aromatic plants: carob, eucalyptus, laurel, mint, myrtle, rosemary and strawberry tree. Phytochem. Anal. 2022, 33, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Nieto, G.; Ros, G.; Castillo, J. Antioxidant and antimicrobial properties of rosemary (Rosmarinus officinalis, L.): A review. Medicines (Basel) 2018, 5, 98. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, J.R.; Camargo, S.E.A.; De Oliveira, L.D. Rosmarinus officinalis L.(rosemary) as therapeutic and prophylactic agent. J. Biomed. Sci. 2019, 26, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.C.; Zhang, H.Y.; Deng, J.S.; Wu, S.X.; Cui, Y.; Yang, M. Chemical constituents and pharmacological activities of Rosmarinus officinalis herba. Chin. J. Exper. Tradit. Medic. Form. 2019, 211–218. [Google Scholar]
- Afonso, MS.; de O Silva, AM.; Carvalho, EB.; Rivelli, DP.; Barros, SB.; Rogero, MM.; Lottenberg, AM.; Torres, RP.; Mancini-Filho, J. Phenolic compounds from Rosemary (Rosmarinus officinalis L.) attenuate oxidative stress and reduce blood cholesterol concentrations in diet-induced hypercholesterolemic rats. Nutr. Metab. (Lond) 2013, 10, 19. [Google Scholar] [CrossRef]
- El-Demerdash, F.M.; El-Sayed, R.A.; Abdel-Daim, M.M. Hepatoprotective potential of Rosmarinus officinalis essential oil against hexavalent chromium-induced hematotoxicity, biochemical, histological, and immunohistochemical changes in male rats. Environ. Sci. Pollut. Res. 2021, 28, 17445–17456. [Google Scholar] [CrossRef]
- Cimrin, T. Thyme (Thymbra spicata L.), rosemary (Rosmarinus officinalis L.) and vitamin E supplementation of laying hens. S. Afr. J. Anim. Sci. 2019, 49, 912–919. [Google Scholar] [CrossRef]
- Christaki, E.; Giannenas, I.; Bonos, E.; Florou-Paneri, P. Innovative uses of aromatic plants as natural supplements in nutrition. In Feed Additives; Florou-Paneri, P., Christaki, E., Giannenas, I., Eds.; Academic Press: London, UK, 2020; pp. 19–34. [Google Scholar]
- Alagawany, M.; Abd El-Hack, M.A. The effect of rosemary herb as a dietary supplement on performance, egg quality, serum biochemical parameters, and oxidative status in laying hens. J. Anim Feed Sci. 2015, 24, 341–347. [Google Scholar] [CrossRef]
- Celi, P.; Cowieson, A.J.; Fru-Nji, F.; Steinert, R.E.; Kluenter, A.-M.; Verlha, V. Gastrointestinal functionality in animal nutrition and health: new opportunities for sustainable animal production. Anim. Feed Sci. Technol. 2017, 234, 88–100. [Google Scholar] [CrossRef]
- Mahfuz, S.; Shang, Q.; Piao, X. Phenolic compounds as natural feed additives in poultry and swine diets: a review. J. Anim. Sci. Biotechnol. 2021, 12, 48. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, C. 2015. Some strategies for the stabilization of long chain n-3 PUFA-enriched foods: a review. Eur. J. Lipid Sci. Tecnol. 2015, 117, 1853–1866. [Google Scholar] [CrossRef]
- Haak, L.; Raes, K.; Van Dyck, S.; De Smet, S. Effect of dietary rosemary and alpha-tocopheryl acetate on the oxidative stability of raw and cooked pork following oxidized linseed oil administration. Meat Sci. 2008, 78, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Hashemzadeh-Cigari, F.; Khorvash, M.; Ghorbani, GR.; Kadivar, M.; Riasi, A.; Zebeli, Q. Effects of supplementation with a phytobiotics-rich herbal mixture on performance, udder health, and metabolic status of Holstein cows with various levels of milk somatic cell counts. J. Dairy Sci. 2014, 97, 7487–97. [Google Scholar] [CrossRef]
- Guo, F.C.; Kwakkel, R.P.; Soede, J.; Williams, B.A.; Verstegen, M.W.A. Effect of a Chinese herb medicine formulation, as an alternative for antibiotics, on performance of broilers. Br. Poult. Sci. 2004, 45, 793–797. [Google Scholar] [CrossRef]
- AOAC international, 2005. Official Methods of Analysis. 18th ed. Association of Analytical Chemists, AOAC International, Arlington Virginia, USA, 2005.
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of the folin-ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar]
- NRC. Nutrient requirements of poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Erel, O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef]
- Lashkari, S.; Taghizadeh, A.; Seifdavati, J.; Salem, A.Z.M. Qualitative characteristics, microbial populations and nutritive values of orange pulp ensiled with nitrogen supplementation. Slovak J. Anim. Sci. 2014, 47, 90–99. [Google Scholar]
- Botsoglou, E.; Govaris, A.; Fletouris, D.; Iliadis, S. Olive leaves (Olea europea L.) and α-tocopheryl acetate as feed antioxidants for improving the oxidative stability of α-linolenic acid-enriched eggs. J. Anim. Physiol. Anim. Nutr. (Berl). 2013, 97, 740–53. [Google Scholar] [CrossRef] [PubMed]
- Christaki, E.V.; Bonos, E.M.; Florou-Paneri, P.C. Comparative evaluation of dietary oregano, anise and olive leaves in laying Japanese quails. Brazil. J. Poult. Sci. 2011, 13, 97–101. [Google Scholar] [CrossRef]
- Ahmed, M.M.; El-Saadany, A.S.; Shreif, E.Y. Effect of dietary olive leaves extract (oleuropein) supplementation on productive, physiological and immunological parameters in bandarah chickens 2- during production period. Egypt. Poult. Sci. J. 2017, 37, 277–292. [Google Scholar]
- Karaalp, M.; Elmastas, M.; Genc, N.; Sezer, M.; Yavuz, M.; Ozkan, M. Bay laurel (Laurus nobilis L.) in Japanese quails feeding 1. Performance and egg quality parameters. J. Anim. Vet. Adv. 2011, 10, 1883–1889. [Google Scholar] [CrossRef]
- Hajiazizi, F.; Torki, M.; Habibian, M. Effects of rosemary essential oil and zinc on performance, egg quality traits, and some serum metabolites in laying hens. J. Livest Sci. Technol. 2016, 4, 1–6. [Google Scholar]
- Botsoglou, N.; Florou-Paneri, P.; Botsoglou, E.; Dotas, V.; Giannenas, I.; Koidis, A.; Mitrakos, P. The effect of feeding rosemary, oregano, saffron and a-tocopheryl acetate on hen performance and oxidative stability of eggs. S. Afr. J. Anim. Sci. 2005, 35, 143–15. [Google Scholar] [CrossRef]
- Erener, G.; Ocak, N.; Ozturk, E.; Cankaya, S.; Ozkanca, R.; Altop, A. Evaluation of olive leaf extract as a growth promoter on the performance, blood biochemical parameters, and caecal microflora of broiler chickens. Rev. Bras. Zootec. 2020, 49, e20180300. [Google Scholar] [CrossRef]
- Ali, N.A.-L.; Al-Shuhaib, M.B.S. Highly effective dietary inclusion of laurel (Laurus nobilis) leaves on productive traits of broiler chickens. Acta Sci. – Anim. Sci. 2021, 43, e52198. [Google Scholar] [CrossRef]
- Abo Ghanima, M.M.; Elsadek, M.F.; Taha, A.E.; Abd El-Hack, M.E.; Alagawany, M.; Ahmed, B.M.; Elshafie, M.M.; El-Sabrout, K. Effect of housing system and rosemary and cinnamon essential eils on layers performance, egg quality, haematological traits, blood chemistry, immunity, and antioxidant. Animals (Basel) 2020, 10, 245. [Google Scholar] [CrossRef]
- Ferdous, M.F.; Arefin, M.S.; Rahman, M.M.; Ripon, M.M.R.; Rashid, M.H.; Sultana, M.R.; Hossain, M.T.; Ahammd, M.U.; Rafiq, K. Beneficial effects of probiotic and phytobiotic as growth promoter alternative to antibiotic for safe broiler production. J. Adv. Vet. Anim. Res. 2019, 6, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Lipiński, K.; Mazur, M.; Antoszkiewicz, Z.; Purwin, C. Polyphenols in monogastric nutrition – A review. Ann. Anim. Sci. 2017, 17, 41–58. [Google Scholar] [CrossRef]
- Martínez, Y.; Yero, O.M.; Liu, G.; Ren, W.; Bertot, R.R.; Yimenez, Y.F.; Gonzalez, C.O.; Del Toro, M.I.; Arocho, R.; Navarro, M.V.; Nyachoti, C.M. Effect of dietary supplementation with Anacardium occidentale on growth performance and immune and visceral organ weights in replacement laying pullets. J. Food. Agric. Environ. 2013, 13, 1352–1357. [Google Scholar]
- Rezaei, M.; KarimiTorshizi, M.A.; Wall, H.; Ivarsson, E. Body growth, intestinal morphology and microflora of quail on diets supplemented with micronised wheat fibre. Brit. Poult. Sci. 2018, 59, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Galosi, L.; Desantis, S.; Roncarati, A.; Robino, P.; Bellato, A.; Ferrocino, I.; Santamaria, N.; Biagini, L.; Filoni, L.; Attili, A.R.; Rossi, G. Positive influence of a probiotic mixture on the intestinal morphology and microbiota of farmed guinea fowls (Numida meleagris). Front. Vet. Sci. 2021, 8, 743899. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhang, H.J.; Yu, S.H.; Wu, S.G.; Yoon, I.; Quigley, J.; Gao, Y.P.; Qi, G.H. Effects of yeast culture in broiler diets on performance and immunomodulatory functions. Poult Sci. 2008, 87, 1377–84. [Google Scholar] [CrossRef] [PubMed]
- Hamedi, S.; Rezaian, M.; Shomali, T. Histological changes of small intestinal mucosa of cocks due to sunflower meal single feeding. Am. J. Anim. Vet. Sci. 2011, 6, 171–175. [Google Scholar]
- Gao, Y.Y.; Zhang, X.L.; Xu, L.H.; Peng, H.; Wang, C.K.; Bi, Y.Z. Encapsulated blends of essential oils and organic acids improved performance, intestinal morphology, cecal microflora, and jejunal enzyme activity of broilers. Czech. J. Anim. Sci. 2019, 64, 189–1898. [Google Scholar] [CrossRef]
- Hampson, D.J. Alterations in piglet small intestinal structure at weaning. Res. Vet. Sci. 1986, 40, 32–40. [Google Scholar] [CrossRef]
- Pearson, J.P.; Brownlee, I.A. The interaction of large bowel microflora with the colonic mucus barrier. Int. J. Inflam. 2010, 2010, 321426. [Google Scholar] [CrossRef]
- Desantis, S.; Galosi, L.; Santamaria, N.; Roncarati, A.; Biagini, L.; Rossi, G. Modulation of morphology and glycan composition of mucins in Guinea fowl (Numida meleagris) intestine by the multistrain probiotic Slab51. Animals (Basel) 2021, 11, 495. [Google Scholar] [CrossRef] [PubMed]
- Meimandipour, A.; Soleimanifarjam, A.; Azhar, K.; Hair-Bejo, M.; Shuhaimi, M.; Nateghi, L.; Yazid, A.M. Age effects on short-chain fatty acids concentrations and pH values in the gastrointestinal tract of broiler chickens. Arch. Fur Geflugelkunde 2011, 75, 164–168. [Google Scholar]
- Herrero-Encinas, J.; Menoyo, D.; Blanch, M.; Pastor, J.J.; Rochell, S.J. Response of broiler chickens fed diets supplemented with a bioactive olive pomace extract from Olea europaea to an experimental coccidial vaccine challenge. Poul. Sci. 2021, 100, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Cross, D.E.; McDevitt, R.M.; Hillman, K.; Acamovic, T. The effect of herbs and their associated essential oils on performance, dietary digestibility and gut microflora in chickens from 7 to 28 days of age. Br. Poult. Sci. 2007, 48, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Yang, H.; Wang, X.; Xia, W.; Lv, W.; Xiao, Y.; Zou, X. Early intervention with cecal fermentation broth regulates the colonization and development of gut microbiota in broiler chickens. Front. Microbiol. 2019, 10, 1422. [Google Scholar] [CrossRef]
- Oost, MJ.; Velkers, F.C.; Kraneveld, A.D.; Venema, K. Development of the in vitro cecal chicken alimentary tract model-2 to study microbiota composition and function. Front. Microbiol. 2021, 12, 726447. [Google Scholar] [CrossRef] [PubMed]
- González-Ortiz, G.; Olukosi, O.A.; Jurgens, G.; Apajalahti, J.; Bedford, M.R. Short-chain fatty acids and ceca microbiota profiles in broilers and turkeys in response to diets supplemented with phytase at varying concentrations, with or without xylanase. Poult. Sci. 2020, 99, 2068–2077. [Google Scholar] [CrossRef]
- Aguirre, M.; Eck, A.; Koenen, M.E.; Savelkoul, P.H.M.; Budding, A.E.; Venema, K. Diet drives quick changes in the metabolic activity and composition of human gut microbiota in a validated in vitro gut model. Res. Microbiol. 2016, 167, 114–125. [Google Scholar] [CrossRef]
- Rios-Covian, D.; Salazar, N.; Gueimonde, M.; de los Reyes-Gavilan, C.G. Shaping the metabolism of intestinal Bacteroides population through diet to improve human health. Front. Microbiol. 2017, 8, 376. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From dietary fiber to host physiology: short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Salonen, A.; Lahti, L.; Salojarvi, J.; Holtrop, G.; Korpela, K.; Duncan, S.H.; Date, P.; Farquharson, F.; Johnstone, A.M.; Lobley, G.E.; Louis, P.; Flin, H.J.; de Vos, W.M. Impact of diet and individual variation on intestinal microbiota composition and fermentation products in obese men. ISME J. 2014, 8, 2218–2230. [Google Scholar] [CrossRef] [PubMed]
- Baothman, O.A.; Zamzami, M.A.; Taher, I.; Abubaker, J.; Abu-Farha, M. The role of gut microbiota in the development of obesity and diabetes. Lipids Health Dis. 2016, 15, 108. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Pandit, R.J.; Hinsu, A.T.; Patel, N.V.; Koringa, P.; Jakhesara, S.J.; Thakkar, J.R.; Shah, T.M.; Limon, G.; Psifidi, A.; Guitian, J.; Hume, D.A.; Tomley, F.M.; Rank, D.N.; Raman, M.; Tirumurugaan, K.G.; Blake, D.P.; Joshi, C.G. Microbial diversity and community composition of caecal microbiota in commercial and indigenous Indian chickens determined using 16s rDNA amplicon sequencing. Microbiome 2018, 6, 115. [Google Scholar] [CrossRef] [PubMed]
- Besten, G.D.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Awad, W.A.; Dublecz, F.; Hess, C.; Khayal, B.; Aschenbach, J.R.; Hess, M. Campylobacter jejuni colonization promotes the translocation of Escherichia coli to extra-intestinal organs and disturbs the short-chain fatty acids profiles in the chicken gut. Poult. Sci. 2016, 95, 2259–2265. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Tyagi, A.; Kumar, A.; Panwar, S.; Grover, S.; Saklani, A.C.; Hemalatha, R.; Batish, V.K. Adhesion of Lactobacilli and their anti-infectivity potential. Crit. Rev. Food Sci. Nutr. 2017, 57, 2042–2056. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, J.; Shi, Y.; Su, W.; Chen, J.; Zhang, Z.; Wang, G.; Wang, F. Short chain fatty acid acetate protects against ethanol-induced acute gastric mucosal lesion in mice. Biol. Pharm. Bull. 2017, 40, 1439–1446. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, Y.; Wang, P.; Huang, Y.; Wang, F. Short-chain fatty acids manifest stimulative and protective effects on intestinal barrier function through the inhibition of NLRP3 inflammasome and autophagy. Cell. Physiol. Biochem. 2018, 49, 190–205. [Google Scholar] [CrossRef]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; McKenzie, C.I.; Hijikata, A.; Wong, C.; Binge, L.; Thorburn, A.N.; Chevalier, N.; Ang, C.; Marino, E.; Robert, R.; Offermanns, S.; Teixeira, M.M.; Moore, R.J.; Flavell, R.A.; Fagarasan, S.; Mackay, C.R. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef]
- Nowarski, R.; Jackson, R.; Gagliani, N.; de Zoete, M.R.; Palm, N.W.; Bailis, W.; Low, J.S.; Harman, C.C.D.; Graham, M.; Elinav, E.; Flavell, R.A. Epithelial IL-18 equilibrium controls barrier function in colitis. Cell 2015, 163, 1444–1456. [Google Scholar] [CrossRef] [PubMed]
- Borda-Molina, D.; Mátis, G.; Mackei, M.; Neogrády, Z.; Huber, K.; Seifert, J.; Camarinha-Silva, A. Caeca microbial variation in broiler chickens as a result of dietary combinations using two cereal types, supplementation of crude protein and sodium butyrate. Front. Microbiol. 2021, 11, 617800. [Google Scholar] [CrossRef] [PubMed]
- Mathewson, N.D.; Jenq, R.; Mathew, A.V.; Koenigsknecht, M.; Hanash, A.; Toubai, T.; Oravecz-Wilson, K.; Wu, S.R.; Sun, Y.; Rossi, C. Gut microbiome-derived metabolites modulate intestinal epithelial cell damage and mitigate graft-versus-host disease. Nat. Immunol. 2016, 17, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Gaudier, E.; Jarry, A.; Blottiere, H.M.; de Coppet, P.; Buisine, M.P.; Aubert, J.P.; Laboisse, C.; Cherbut, C.; Hoebler, C. Butyrate specifically modulates MUC gene expression in intestinal epithelial goblet cells deprived of glucose. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G1168–G1174. [Google Scholar] [CrossRef]
- Tong, L.-C.; Wang, Y.; Wang, Z.-B.; Liu, W.-Y.; Sun, S.; Li, L.; Su, D.-F.; Zhang, L.-C. Propionate ameliorates dextran sodium sulfate-induced colitis by improving intestinal barrier function and reducing inflammation and oxidative stress. Front. Pharmacol. 2016, 7, 253. [Google Scholar] [CrossRef]
- Schoenfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef]
- Brüssow, H.; Parkinson, S.J. Parkinson, S.J. You are what you eat. Nat. Biotechnol. 2014, 32, 243–245. [Google Scholar] [CrossRef]
- Boets, E.; Gomand, S.V.; Deroover, L.; Preston, T.; Vermeulen, K.; De Preter, V.; Hamer, H.M.; Van den Mooter, G.; De Vuyst, L.; Courtin, C.M.; Annaert, P.; Delcour, J.A.; Verbeke, K.A. Systemic availability and metabolism of colonic-derived short-chain fatty acids in healthy subjects: a stable isotope study. J. Physiol. 2017, 595, 541–555. [Google Scholar] [CrossRef]
- Clench, M.; Mathias, J. The avian cecum: a review. Wilson Bull. 1995, 10, 93–121. [Google Scholar]
- Jamroz, D.; Jakobsen, K.; Bach, K.K.; Wiliczkiewicz, A.; Orda, J. Digestibility and energy value of non-starch polysaccharides in young chickens, ducks and geese, fed diets containing high amounts of barley. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2002, 131, 657–668. [Google Scholar] [CrossRef]
- Stanley, D.; Hughes, R.J.; Moore, R.J. Microbiota of the chicken gastrointestinal tract: influence on health, productivity and disease. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef] [PubMed]
- Waite, D.W.; Taylor, M.W. Characterizing the avian gut microbiota: membership, driving influences, and potential function. Front. Microbiol. 2014, 5, 223. [Google Scholar] [CrossRef] [PubMed]
- Diaz Carrasco, J.M.; Casanova, N.A.; Fernández Miyakawa, M.E. Microbiota, gut health and chicken productivity: what is the connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Wang, J.; Yu, L.; Zhang, Q.; Chen, K.; Liu, B. Modulation of growth performance and intestinal microbiota in chickens fed plant extracts or virginiamycin. Front. Microbiol. 2019, 10, 1333. [Google Scholar] [CrossRef]
- Romo-Vaquero, M.; Selma, M.-V.; Larrosa, M.; Obiol, M.; García-Villalba, R.; González-Barrio, R.; Issaly, N.; Flanagan, J.; Roller, M.; Tomás-Barberán, F.A.; García-Conesa, M.-T. A rosemary extract rich in carnosic acid selectively modulates caecum microbiota and inhibits -glucosidase activity, altering fiber and short-chain fatty acids fecal excretion in lean and obese female rats. PLoS ONE 2014, 9, e94687. [Google Scholar] [CrossRef]
- He, X.; Zhang, M.; Li, S.T.; Li, X.; Huang, Q.; Zhang, K.; Zheng, X.; Xu, XT.; Zhao, DG.; Ma, Y.Y. Alteration of gut microbiota in high-fat diet-induced obese mice using carnosic acid from rosemary. Food Sci. Nutr. 2022, 10, 2325–2332. [Google Scholar] [CrossRef]
- Derwich, E.; Benziane, Z.; Boukir, A. Chemical composition and antibacterial activity of leaves essential oil of Laurus nobilis from Morocco. Aust. J. Basic Appl. Sci. 2009, 3, 3818–3824. [Google Scholar]
- Jabri, J.; Kacem, H.; Yaich, H.; Abid, K.; Kamoun, M.; Rekhis, J.; Malek, A. Effect of olive leaves extract supplementation in drinking water on zootechnical performances and cecal microbiota balance of broiler chickens. J. New Sci. Sustain Livest. Manag. 2017, 4, 69–75. [Google Scholar]
- Xie, P.; Deng, Y.; Huang, L.; Zhang, C. Effect of olive leaf (Olea europaea L.) extract addition to broiler diets on the growth performance, breast meat quality, antioxidant capacity and caecal bacterial populations. Ital. J. Anim. Sci. 2022, 21, 1246–1258. [Google Scholar] [CrossRef]
- Lin, Y.T.; Vattem, D.A.; Labbe, R.G.; Shetty, K. Enhancement of antioxidant activity and inhibition of Helicobacter pylori by phenolic phytochemical enriched alcoholic beverages. Process Biochem. 2005, 40, 2059–2065. [Google Scholar] [CrossRef]
- Cardona, F.; Andres-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–22. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, A.; Bartolome, B.; Martínez-Rodríguez, AJ.; Pueyo, E.; Martín-Alvarez, PJ.; Moreno-Arribas, M.V. Potential of phenolic compounds for controlling lactic acid bacteria growth in wine. Food Control. 2008, 19, 835–841. [Google Scholar] [CrossRef]
- Amit-Romach, E.; Sklan, D.; Uni, Z. Microflora ecology of the chicken intestine using 16S ribosomal DNA primers. Poult. Sci. 2004, 83, 1093–1098. [Google Scholar] [CrossRef] [PubMed]
- Corrier, D.E.; Nisbet, D.J.; Scanlan, C.M.; Hollister, A.G.; Deloach, J.R. Control of Salmonella typhimurium colonization in broiler chicks with a continuous-flow characterized mixed culture of cecal bacteria. Poult. Sci. 1995, 74, 916–24. [Google Scholar] [CrossRef] [PubMed]
- Travers, M.A.; Florent, I.; Kohl, L.; Grellier, P. Probiotics for the control of parasites: an overview. J. Parasitol. Res. 2011, 2011, 610769. [Google Scholar] [CrossRef] [PubMed]
- Van Immerseel, F.; Fievez, V.; De Buck, J.; Pasmans, F.; Martel, A.; Haesebrouck, F.; Ducatelle, R. Microencapsulated short-chain fatty acids in feed modify colonization and invasion early after infection with Salmonella enteritidis in young chickens. Poult. Sci. 2004, 83, 69–74. [Google Scholar] [CrossRef]
- Vermeulen, K.; Verspreet, J.; Courtin, C.M.; Haesebrouck, F.; Ducatelle, R.; Immerseel, F.V. Reduced particle size wheat bran is butyrogenic and lowers Salmonella colonization, when added to poultry feed. Vet. Microbiol. 2017, 198, 64–71. [Google Scholar] [CrossRef]
- Sun, Y.; O’Riordan, M.X.D. Regulation of bacterial pathogenesis by intestinal short-chain fatty acids. Adv. Appl. Microbiol. 2013, 85, 93–118. [Google Scholar]
- Attia, Y.A.; Bovera, F.; El-Tahawy, W.S.; El-Hanoun, A.M.; Al-Harthi, M.A.; Habiba, H.I. Productive and reproductive performance of rabbits does as affected by bee pollen and/or propolis, inulin and/or mannan-oligosaccharides. World Rabbit Sci. 2015, 2, 273–282. [Google Scholar] [CrossRef]
- Sarica, S.; Toptas, S. Effects of dietary oleuropein supplementation on growth performance, serum lipid concentrations and lipid oxidation of Japanese quails. J. Anim. Physiol. Anim. Nutr. 2014, 98, 1176–1186. [Google Scholar] [CrossRef]
- Talhaoui, N.; Taamalli, A.; Gómez-Caravaca, A.M.; Fernández-Gutiérrez, A.; SeguraCarretero, A. Phenolic compounds in olive leaves: analytical determination, biotic and abiotic influence, and health benefits. Food Res. Int. 2015, 77, 92–108. [Google Scholar] [CrossRef]
- Elazab, M.A.; Khalifah, A.M.; Elokil, A.A.; Elkomy, A.E.; Rabie, M.M.; Mansour, A.T.; Morshedy, S.A. Effect of dietary rosemary and ginger essential oils on the growth performance, feed utilization, meat nutritive value, blood biochemicals, and redox status of growing NZW rabbits. Animals 2022, 12, 375. [Google Scholar] [CrossRef] [PubMed]
- Torki, M.; Sedgh-Gooya, S.; Mohammadi, H. Effects of adding essential oils of rosemary, dill and chicory extract to diets on performance, egg quality and some blood parameters of laying hens subjected to heat stress. J. App. Anim. Res. 2018, 46, 1118–1126. [Google Scholar] [CrossRef]
- Radwan, N.L.; Hassan, R.A.; Qota, E.M.; Fayek, H.M. Effect of natural antioxidant on oxidative stability of eggs and productive and reproductive performance of laying hens. Int. J. Poult. Sci. 2008, 7, 134–150. [Google Scholar]
- Bolukbasi, S.C.; Erhan, M.K.; Kaynar, O. The effect of feeding thyme, sage and rosemary oil on laying hen performance, cholesterol and some proteins ratio of egg yolk and E. coli count in feces. Arch. Geflügelk. 2008, 72, 231–237. [Google Scholar]
- Chalabi, M.; Majeed, D.; Jasim, A.; Al-Azzawi, S. Beneficial effects of ethanolic extract of bay leaves (Laurus nobilis) on blood sugar levels in adult diabetic rats induced by alloxan monohydrate. Ann. Trop. Med. Publ. Health. 2020, 23, e175–e184. [Google Scholar] [CrossRef]
- Chiang, J.Y.L.; Ferrell, J.M. Up to date on cholesterol 7 alpha-hydroxylase (CYP7A1) in bile acid synthesis. Liver Res. 2020, 4, 47–63. [Google Scholar] [CrossRef]
- Wasburn, K.W.; Nix, DF. Genetic basis of yolk cholesterol content. Poult. Sci. 1974, 53, 109–15. [Google Scholar] [CrossRef]
- Rahimi, G. Dietary forage legume (Onobrychis altissima grossh.) supplementation on serum/yolk cholesterol, triglycerides and eggshell characteristics in laying hens. Int. J. Poult. Sci. 2005, 4, 772–776. [Google Scholar]
- Hargis, P.S.; Van Elswyk, M.E.; Hargis, B.M. Dietary modification of yolk lipid with savelha oil. Poult. Sci. 1991, 70, 874–83. [Google Scholar] [CrossRef]
- Darmawan, A.; Hermana, W.; Suci, D.M.; Mutia, R.; Sumiati; Jayanegara, A.; Ozturk, E. Dietary phytogenic extracts favorably influence productivity, egg quality, blood constituents, antioxidant and immunological parameters of laying hens: a meta-analysis. Animals (Basel) 2022, 12, 2278.
- Sharma, M.K.; Dinh, T.; Adhikari, P.A. Production performance, egg quality, and small intestine histomorphology of the laying hens supplemented with phytogenic feed additive. J. Appl. Poult. Res. 2020, 29, 362–371. [Google Scholar] [CrossRef]
- Bertechini, A.G. Mitos e verdades sobre o ovo de consumo. In 21th Conferência de Ciência e Tecnologia Avícola; Santos: São Paulo. Brasil, 2003; pp. 19–26. [Google Scholar]
- Shafey, T.M.; Cham, B.E. Altering fatty acid and cholesterol contents of eggs for human consumption. In Egg uses and processing technologies: new developments; Sim, J.S., Nakai, S., Eds.; CAB International: Washington, DC, USA, 1994; pp. 374–385. [Google Scholar]
- Griffin, H.D. Manipulation of egg yolk cholesterol: a physiologist's view. World Poult. Sci. J. 1992, 48, 101–112. [Google Scholar] [CrossRef]
- Fennema, O.R. Química de los alimentos, Zaragoza: Acribia; 1993.
- Wu, R.; Feng, J.; Yang, Y.; Dai, C.; Lu, A.; Li, J.; Liao, Y.; Xiang, M.; Huang, Q.; Wang, D.; Du, X.-B. Significance of serum total oxidant/antioxidant status in patients with colorectal cancer. PLoS ONE 2017, 12, e0170003. [Google Scholar] [CrossRef] [PubMed]
- Bulbul, T.; Ulutas, E. The effects of dietary supplementation of false flax (Camelina sativa L.) meal on performance, egg quality traits, and lipid peroxidation in laying quails. Eurasian J. Vet. Sci. 2015, 31, 8–15. [Google Scholar] [CrossRef]
- Vallverdú-Queralt, A.; Regueiro, J.; Martínez-Huélamo, M.; Alvarenga, J.F.; Leal, L.N.; Lamuela-Raventos, R. MA comprehensive study on the phenolic profile of widely used culinary herbs and spices: rosemary, thyme, oregano, cinnamon, cumin and bay. Food Chem. 2014, 154, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Falade, A.O.; Adewole K., E.; Adekola, A.O.; Ikokoh, H.A.; Okaiyeto, K.; Oguntibeju, O.O. Aqueous extract of bay leaf (Laurus nobilis) ameliorates testicular toxicity induced by aluminum chloride in rats. Vet. World 2022, 15, 2525–2534. [Google Scholar] [CrossRef]
- Wenk, C. Herbs, botanicals and other related substances. WPSA-Bremen. Germany, 2002.
- Chahal, K.; Kaur, M.; Bhardwaj, U.; Singla, N.; Kaur, A. A review on chemistry and biological activities of Laurus nobilis L. essential oil. J. Pharm. Phytochem. 2017, 6, 1153–1161. [Google Scholar]
- Loussouarn, M.; Krieger-Liszkay, A.; Svilar, L.; Bily, A.; Birtić, S.; Havaux, M. Carnosic acid and carnosol, two major antioxidants of rosemary, act through different mechanisms. Plant Physiol. 2017, 75, 1381–1394. [Google Scholar] [CrossRef]
- Silva, S.; Gomes, L.; Leitão, F.; Coelho, AV.; Boas, LV. Phenolic compounds and antioxidant activity of Olea europaea L. fruits and leaves. Food Sci. Technol. Int. 2006, 12, 385–395. [Google Scholar] [CrossRef]
- Hayes, J.E.; Allen, P.; Brunton, N.; O’Grady, M. .N.; Kerry, JP. Phenolic composition and in vitro antioxidant capacity of four commercial phytochemical products: olive leaf extract (Olea Europaea L.), lutein, sesamol and ellagic acid. Food Chem. 2011, 126, 948–955. [Google Scholar] [CrossRef]
- Turkez, H.; Togar, B.; Polat, E. Olive leaf extract modulates permethrin induced genetic and oxidative damage in rats. Cytotechnology 2012, 64, 459–64. [Google Scholar] [CrossRef] [PubMed]
- Sarıca, S.; Aydın, H.; Ciftci, G. Effects of dietary supplementation of some antioxidants on liver antioxidant status and plasma biochemistry parameters of heat-stressed quail. Turk. J. Food Agric. Sci. 2017, 5, 773–779. [Google Scholar] [CrossRef]
- Adedayo, B.C.; Oyeleye, S.I.; Okeke, B.M.; Oboh, G. Anti-cholinesterase and antioxidant properties of alkaloid and phenolic-rich extracts from pawpaw (Carica papaya) leaf: A comparative study. Flavour Fragr. J. 2020, 00, 1–8. [Google Scholar] [CrossRef]
- Khayyal, A.; El-Badawy, M.; Ashmawy, T. Effect of rosemary or laurel leaves as feed additives on performance of growing lambs. Egypt. J. Nutr. Feeds 2021, 24, 343–356. [Google Scholar] [CrossRef]
- Liu, Z.; Luo, Z.; Jia, C.; Wang, D.; Li, D. Synergistic effects of Potentilla fruticosa L. leaves combined with green tea polyphenols in a variety of oxidation systems. J. Food Sci. 2016, 81, C1091–C1101. [Google Scholar] [CrossRef]
- Wu, L.; Xiong, X.; Wu, X.; Ye, Y.; Jian, Z.; Zhi, Z.; Gu, L. Targeting oxidative stress and inflammation to prevent ischemia- Reperfusion injury. Front. Mol. Neurosci. 2020, 13, 28. [Google Scholar] [CrossRef]
- Yuan, X.; Zhu, J.; Kang, Q.; He, X.; Guo, D. Protective effect of hesperidin against sepsis-induced lung injury by inducing the heat-stable protein 70 (Hsp70)/Toll-Like Receptor 4 (TLR4)/ Myeloid differentiation primary response 88 (MyD88) pathway. Med. Sci. Monit. 2019, 25, 107–114. [Google Scholar] [CrossRef]
- Estienne, A.; Bongrani, A.; Reverchon, M.; Ramé, C.; Ducluzeau, P.H.; Froment, P.; Dupont, J. Involvement of novel adipokines, chemerin, visfatin, resistin and apelin in reproductive functions in normal and pathological conditions in humans and animal models. Int. J. Mol. Sci. 2019, 20, 4431. [Google Scholar] [CrossRef]
- Barbe, A.; Mellouk, N.; Ramé, C.; Grandhaye, J.; Staub, C.; Venturi, E.; Cirot, M.; Petit, A.; Anger, K.; Chahnamian, M.; Ganier, P.; Callut, O.; Cailleau-Audouin, E.; Metayer-Coustard, S.; Riva, A.; Froment, P.; Dupont, J. A grape seed extract maternal dietary supplementation in reproductive hens reduces oxidative stress associated to modulation of plasma and tissue adipokines expression and improves viability of offsprings. PLoS One 2020, 15, e0231131. [Google Scholar] [CrossRef]
- Miliaraki, M.; Briassoulis, P.; Ilia, S.; Michalakakou, K.; Karakonstantakis, T.; Polonifi, A.; Bastaki, K.; Briassouli, E.; Vardas, K.; Pistiki, A.; Theodorakopoulou, M.; Tavladaki, T.; Spanaki, AM.; Kondili, E.; Dimitriou, H.; Venihaki, M.; Tsiodras, S.; Georgopoulos, D.; Mantzourani, M.; Nanas, S.; Armaganidis, A.; Daikos, GL.; Papassotiriou, I.; Briassoulis, G. Oxidant/antioxidant status is impaired in sepsis and is related to anti-apoptotic, inflammatory, and innate immunity alterations. Antioxidants 2022, 11, 231. [Google Scholar] [CrossRef] [PubMed]
- Vezza, T.; Algieri, F.; Rodríguez-Nogales, A.; Garrido-Mesa, J.; Utrilla, M.P.; Talhaoui, N.; Gómez-Caravaca, A.M.; Segura-Carretero, A.; Rodríguez-Cabezas, M.E.; Monteleone, G.; Gálvez, J. Immunomodulatory properties of Olea europaea leaf extract in intestinal inflammation. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- De Cicco, P.; Maisto, M.; Tenore, G.C.; Ianaro, A. Olive leaf extract, from Olea europaea L., reduces palmitate-induced inflammation via regulation of murine macrophages polarization. Nutrients 2020, 12, 3663. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Sano, M.; Seno, K.; Oogaki, Y.; Takahashi, H.; Ohkuchi, A.; Yokozawa, M.; Yamauchi, K.; Iwata, H.; Kuwayama, T.; Shirasuna, K. Olive Leaf Extract (OleaVita) suppresses inflammatory cytokine production and NLRP3 inflammasomes in human placenta. Nutrients 2019, 11, 970. [Google Scholar] [CrossRef]
- Lee, E.H.; Shin, J.H.; Kim, S.S.; Lee, H.; Yang, S.R.; Seo, S.R. Laurus nobilis leaf extract controls inflammation by suppressing NLRP3 inflammasome activation. J. Cell Physiol. 2019, 234, 6854–6864. [Google Scholar] [CrossRef] [PubMed]
- Farouk, S.M.; Abdel-Rahman, H.G.; Abdallah, O.A.; El-Behidy, N.G. Comparative immunomodulatory efficacy of rosemary and fenugreek against Escherichia coli infection via suppression of inflammation and oxidative stress in broilers. Environ. Sci. Pollut. Res. 2022, 29, 40053–40067. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.H.; Chuang, L.T.; Lien, T.J.; Liing, Y..R.; Chen, W.Y.; Tsai, PJ. Rosmarinus officinalis extract suppresses Propionibacterium acnes-induced inflammatory responses. J. Med. Food. 2013, 16, 324–33. [CrossRef]
- Aroche, R.; Martínez, Y.; Ruan, Z.; Guan, G.; Waititu, S.; Nyachoti, C.M.; Más, D.; Lan, S. Dietary inclusion of a mixed powder of medicinal plant leaves to enhance the feed efficiency and immune function in broiler chickens. J. Chem. 2018, 394, 1–6. [Google Scholar] [CrossRef]
- Diaz, G.J.; Roldan, L.P.; Cortes, A. Intoxication of Crotalaria pallida seeds to growing broiler chicks. Vet. Hum. Toxicol. 2003, 45, 187–189. [Google Scholar]
- Valchev, I.; Kanakov, D.; Hristov, TS.; Lazarov, L.; Binev, R.; Grozeva, N.; Nikolov, Y. Investigations on the liver function of broiler chickens with experimental aflatoxicosis. Bulg. J. Vet. Med. 2014, 17, 302–313. [Google Scholar]
- Khalil, E.A.M. Evaluation of the hepatoprotective activity of an aqueous extract of olive leaves in male albino rats. Egypt. J. Hosp. Med. 2004, 15, 118–123. [Google Scholar] [CrossRef]
- Abdel-Wahhab, K.G.E.; El-Shamy, K.A.; El-Beih, N.A.E.; Morcy, F.A.; Mannaa, F.A.E. Protective effect of a natural herb (Rosmarinus officinalis) against hepatotoxicity in male albino rats. Com. Sci. 2011, 2, 9–17. [Google Scholar]
- Al-Attar, A.M.; Shawush, N.A. Influence of olive and rosemary leaves extracts on chemically induced liver cirrhosis in male rats. Saudi J. Biol. Sci. 2015, 22, 157–63. [Google Scholar] [CrossRef] [PubMed]
- Al Chalabi, S.; Majeed, D.; Jasim, A.; Al-Azzawi, K. Benefit effect of ethanolic extract of Bay leaves (Laura nobilis) on blood sugar level in adult diabetic rats induced by alloxan monohydrate. Ann. Trop. Med. Publ. Health 2020, 23, 16. [Google Scholar] [CrossRef]
- Abdel-Azeem, A.; Fayed, A.; Azoz, A. Physiological response, semen quality and blood biochemical parameters of rabbit bucks supplemented with phytogenic components during summer season of Egypt. Egypt. J. Nutr. Feeds 2018, 21, 711–724. [Google Scholar] [CrossRef]
Figure 1.
Representative histological view of the duodenum (A) and ileum (B) of the laying hens. C, Detail of villus epithelium showing the goblet cells. cr, crypts; ct, connective tissue; m, muscularis; v, villum; asterisk, goblet cell. Hematoxylin-Eosin staining. Scale bars: A = 500 µm; B, = 250 µm; C = 20 µm.
Figure 1.
Representative histological view of the duodenum (A) and ileum (B) of the laying hens. C, Detail of villus epithelium showing the goblet cells. cr, crypts; ct, connective tissue; m, muscularis; v, villum; asterisk, goblet cell. Hematoxylin-Eosin staining. Scale bars: A = 500 µm; B, = 250 µm; C = 20 µm.
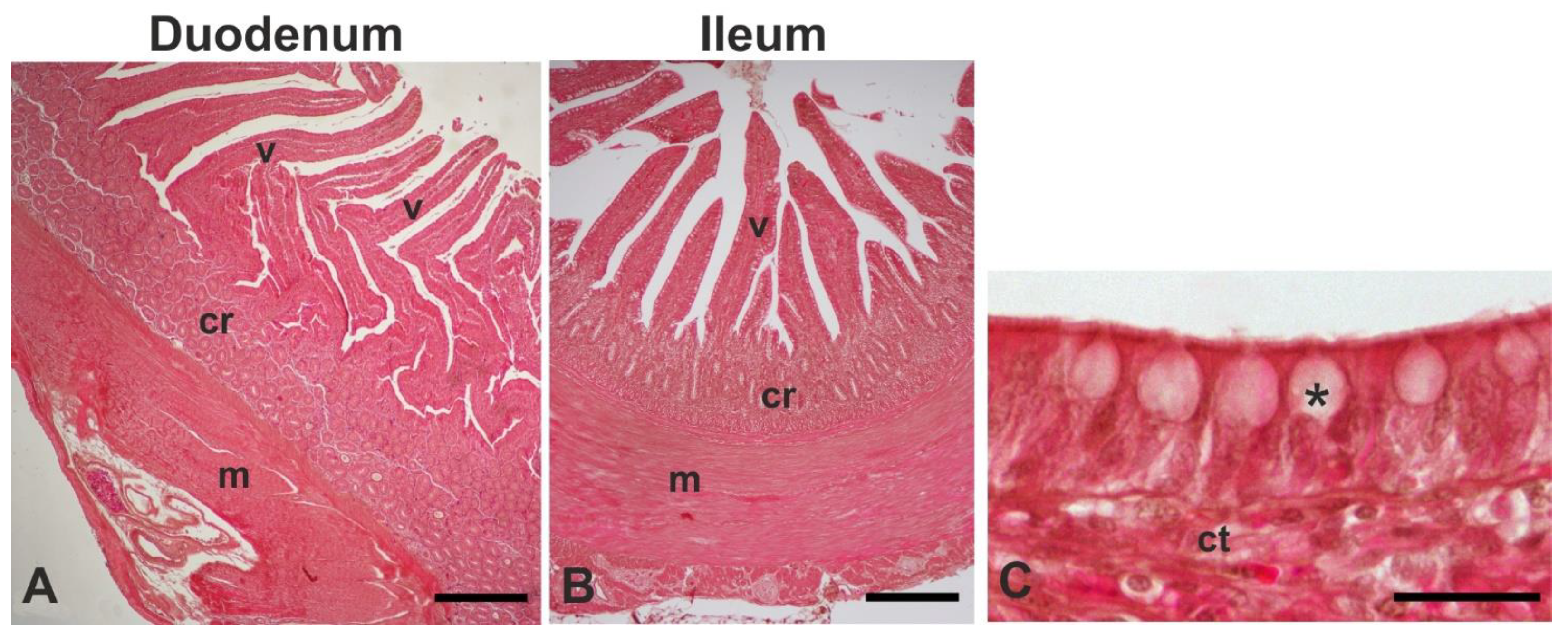

Table 1.
Chemical composition and phenol content of olive, laurel, and rosemary leaves (% DM; X ± SD).
Table 1.
Chemical composition and phenol content of olive, laurel, and rosemary leaves (% DM; X ± SD).
| Olive | Laurel | Rosemary | |
|---|---|---|---|
| Dry matter (%) | 60.57 ± 0.95 | 30.76 ± 0.83 | 31.91 ± 0.80 |
| Crude protein | 9.66 ± 0.61 | 5.13 ± 0.31 | 6.40 ± 0.02 |
| Ether extract | 2.34 ± 0.51 | 1.42 ± 0.71 | 12.85 ± 0.15 |
| Ash | 7.15 ± 0.36 | 3.75 ± 0.34 | 7.24 ± 0.93 |
| Crude fiber | 24.09 ± 1.25 | 12.67 ± 0.60 | 26.20 ± 1.80 |
| Neutral detergent fiber | 46.12 ± 1.58 | 23.85 ± 3.93 | 45.34 ± 1.01 |
| Acid detergent fiber | 28.52 ±0.87 | 14.69 ± 0.69 | 33.17 ± 0.69 |
| Acid detergent lignin | 19.46 ± 0.81 | 9.63 ± 1.94 | 22.23 ± 0.53 |
| Acid insoluble ash | 0.20 ± 0.02 | 0.11 ± 0.09 | 0.35 ± 0.05 |
| Total phenols (mg GAE/g DM) | 6.10 ± 0.80 | 3.90 ± 1.12 | 5.10 ± 1.11 |
Total phenol contents were determined as milligrams of gallic acid equivalents (GAE) per gram of dry weight.
Table 2.
Composition of the control diet (%).
| Ingredients | |
|---|---|
| Corn meal | 57.60 |
| Soybean meal (46% CP) | 22.00 |
| Sunflower flour (36% CP) | 6.00 |
| Limestone granular | 6.00 |
| Limestone | 3.30 |
| Soyabean oil | 2.50 |
| Bicalcium phosphate | 1.50 |
| Vitamin and mineral premix1 | 0.50 |
| Sodium chloride | 0.20 |
| Sodium bicarbonate | 0.15 |
| Methionine (MHA)2 | 0.14 |
| Lisine | 0.09 |
| Magnesium oxide | 0.02 |
| Metabolizable energy (kcal/kg) | 2,700 |
| Crude protein (%) | 17.80 |
| Calcium (%) | 4.10 |
| Phosphorus (available, %) | 0.45 |
| Methionine (%) | 0.31 |
| Lisine | 0.74 |
| Arginine | 0.70 |
| Threonine | 0.37 |
| Leucine | 0.74 |
| Isoleucine | 0.43 |
| Valine | 0.46 |
| Histidine | 0.25 |
| Phenylalanine | 0.48 |
| Tryptophan | 0.13 |
1Supplied per kilogram of diet: vitamin A, 10,000 IU; vitamin D3, 3,000 IU; biotin, 0.08 mg; choline chloride, 350.00 mg; folic acid, 0.80 mg; niacin, 29.50 mg; vitamin B1, 1.96 mg; vitamin B12, 0.02 mg; vitamin B2, 4.00 mg; vitamin B6, 3.96 mg; vitamin E, 25.00 mg; vitamin K3, 1.79 mg; enzymes: endo-1,4-betaglucanasi; endo-1,4-betaxilanasi; 3-fitasi; iron, 48.00 mg; iodine, 1.02 mg; manganese, 80.60 mg; calcium D-pantothenate, 8.91 mg; selenium, 0.15 mg; zinc, 79.2 mg; copper, 12.50 mg. 2methionine hydroxy analogue.
Table 3.
Effect of dietary leaf powder mixture (LPM) supplementation on egg characteristics (n = 30) in laying hens at the end of the experiment.
Table 3.
Effect of dietary leaf powder mixture (LPM) supplementation on egg characteristics (n = 30) in laying hens at the end of the experiment.
| Dietary treatment | SEM | P-value | ||
| CON | LPM | |||
| Egg weight, g | 60.64 | 61.03 | 0.53 | 0.722 |
| Shell weight, g | 7.73 | 7.79 | 0.09 | 0.783 |
| Yolk weight, g | 15.82 | 15.68 | 0.22 | 0.747 |
| Albumen weight, g | 35.60 | 35.56 | 0.41 | 0.960 |
| Yolk ratio, % | 26.15 | 25.67 | 0.30 | 0.440 |
| Albumen ratio, % | 58.67 | 58.26 | 0.38 | 0.591 |
| Shell ratio, % | 12.76 | 12.78 | 0.14 | 0.933 |
| Yolk albumen ratio, % | 44.75 | 44.21 | 0.66 | 0.690 |
| Egg shell thickness, mm | 0.43 | 0.40 | 0.025 | 0.602 |
| Yolk color fan (1-15 scale)1 | 7.69 | 7.86 | 0.16 | 0.598 |
CON, control basal diet; LPM, diet supplemented with leaf powder mixture, 6 g/kg feed containing 2 g olive leaves + 2 g laurel leaves + 2 g rosemary leaves. SEM: standard error mean. 11: pale yellow; 15: deep orange.
Table 4.
Effect of the dietary leaf power mixture (LPM) supplementation on the intestinal morphology of laying hens (n = 3) (x ± SD).
Table 4.
Effect of the dietary leaf power mixture (LPM) supplementation on the intestinal morphology of laying hens (n = 3) (x ± SD).
| Item | CON | LPM | P-value |
|---|---|---|---|
| Duodenum | |||
| Villus height (µm) | 1046.57±199.55 | 1121.73±211.15 | 0.004 |
| Crypt depth (µm) | 245.78±59.35 | 303.44±11.84 | <0.001 |
| Villus height: crypt depth | 4.25±1.39 | 3.69±1.24 | 0.046 |
| Ileum | |||
| Villus height (µm) | 482.44±102.28 | 593.15±148.24 | <0.001 |
| Crypt depth (µm) | 148.24±30.58 | 201.05±77.25 | <0.001 |
| Villus height: crypt depth | 3.25±1.09 | 2.95±0.61 | 0.145 |
CON, control basal diet; LPM, diet supplemented with leaf powder mixture, 6 g/kg feed containing 2 g olive leaves + 2 g laurel leaves + 2 g rosemary leaves.
Table 5.
Effect of dietary leaf powder mixture (LPM) supplementation on cecal concentration of short-chain fatty acids (mg/g DM) in laying hens at the end of the experiment.
Table 5.
Effect of dietary leaf powder mixture (LPM) supplementation on cecal concentration of short-chain fatty acids (mg/g DM) in laying hens at the end of the experiment.
| Dietary treatment | SEM | P value | ||
| CON | LPM | |||
| Animals, n. | 8 | 8 | ||
| Acetic acid | 20.71 | 28.11 | 1.34 | 0.022 |
| Propionic acid | 4.79 | 10.85 | 0.62 | 0.046 |
| Isobutyric acid | 0.83 | 2.40 | 0.15 | <0.001 |
| Butyric acid | 0.54 | 1.40 | 0.08 | 0.048 |
| Isocaproic acid | 1.29 | 1.66 | 0.23 | 0.304 |
| Eptanoic acid | 2.26 | 2.35 | 0.31 | 0.867 |
CON, control basal diet; LPM, diet supplemented with leaf powder mixture, 6 g/kg feed containing 2 g olive leaves + 2 g laurel leaves + 2 g rosemary leaves. SEM: standard error mean.
Table 6.
Effect of dietary leaf powder mixture (LPM) supplementation on biochemical parameters in laying hens at the end of the experiment.
Table 6.
Effect of dietary leaf powder mixture (LPM) supplementation on biochemical parameters in laying hens at the end of the experiment.
| Dietary treatment | SEM | P value | ||
| CON | LPM | |||
| Animals, n. | 30 | 30 | ||
| Total protein, g/L | 61.52 | 62.61 | 0.23 | 0.015 |
| Tryglicerides, mg/dL | 796.42 | 669.93 | 35.70 | 0.076 |
| Total cholesterol, mg/dL | 151.48 | 125.73 | 6.19 | 0.037 |
| HDL cholesterol, mmol/L | 20.61 | 24.36 | 0.63 | 0.002 |
| LDL cholesterol, mg/dL | 143.69 | 135.12 | 1.33 | 0.001 |
| NEFA, mmol/L | 1.39 | 1.31 | 0.04 | 0.331 |
CON, control basal diet; LPM, diet supplemented with leaf powder mixture, 6 g/kg feed containing 2 g olive leaves + 2 g laurel leaves + 2 g rosemary leaves. HDL = high density lipoprotein; LDL = low density lipoprotein, NEFA = not esterified fatty acids. SEM: standard error mean.
Table 7.
Effect of dietary leaf powder mixture (LPM) supplementation on oxidative status of laying hens at the end of the experiment.
Table 7.
Effect of dietary leaf powder mixture (LPM) supplementation on oxidative status of laying hens at the end of the experiment.
| Dietary treatment | SEM | P value | ||
| CON | LPM | |||
| Animals, n. | 30 | 30 | ||
| TOS, mmol H2O2 Eq/L | 23.06 | 17.97 | 0.67 | 0.002 |
| TAS, Trolox Eq/L | 393.78 | 469.71 | 24.74 | 0.11 |
| ROMs, UCarr | 39.29 | 28.07 | 1.74 | 0.001 |
CON, control basal diet; LPM, diet supplemented with leaf powder mixture, 6 g/kg feed containing 2 g olive leaves + 2 g laurel leaves + 2 g rosemary leaves. ROMs = reactive oxigen metabolites; SEM: standard error mean; TAS = total antioxidant status; TOS = total oxidative stress.
Table 8.
Effect of dietary leaf powder mixture (LPM) supplementation on serum levels of TNF-a, IL-1b and IL-6 in laying hens at the end of the experiment.
Table 8.
Effect of dietary leaf powder mixture (LPM) supplementation on serum levels of TNF-a, IL-1b and IL-6 in laying hens at the end of the experiment.
| Dietary treatment | SEM | P value | ||
| CON | LPM | |||
| Animals, n. | 30 | 30 | ||
| TNF-a, pg/mL | 25.85 | 21.07 | 1.10 | 0.029 |
| IL-1b, pg/mL | 69.71 | 57.42 | 2.66 | 0.02 |
| IL-6, pg/mL | 21.61 | 15.95 | 1.16 | 0.013 |
CON, control basal diet; IL-1b = interleukine-1b; IL-6 = interleukine-6; LPM, diet supplemented with leaf powder mixture, 6 g/kg feed containing 2 g olive leaves + 2 g laurel leaves + 2 g rosemary leaves; SEM: standard error mean; TNF-a = tumor necrosis factor.
Table 9.
Effect of dietary leaf powder mixture (LPM) supplementation on serum levels of total bilirubin and ALT, AST, and ALP in laying hens at the end of the experiment.
Table 9.
Effect of dietary leaf powder mixture (LPM) supplementation on serum levels of total bilirubin and ALT, AST, and ALP in laying hens at the end of the experiment.
| Dietary treatment | SEM | P value | ||
| CON | LPM | |||
| Animals, n. | 30 | 30 | ||
| ALP, IU/L | 275.71 | 261.98 | 2.92 | 0.017 |
| AST, IU/L | 186.27 | 176.10 | 1.22 | <0.001 |
| ALT, IU/L | 17.16 | 16.18 | 0.11 | <0.001 |
| Bilirubin, mmol/L | 0.99 | 1.03 | 0.30 | 0.299 |
ALP = alkaline phosphatase; ALT = alanine aminotransferase enzyme; AST = aspartate aminotransferase; CON, control basal diet; LPM, diet supplemented with leaf powder mixture, 6 g/kg feed containing 2 g olive leaves + 2 g laurel leaves + 2 g rosemary leaves. SEM: standard error mean.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



