
Submitted:
11 May 2023
Posted:
12 May 2023
You are already at the latest version
Abstract
Ferroptosis is a unique variety of non-apoptotic cell death, driven by massive lipid oxidation in an iron-dependent manner. Since Ferroptosis was introduced as a concept in 2012, it was shown it's essential role in the pathogenesis in neurodegenerative diseases and an important role in therapy-resistant cancer cells. Thus, a detailed molecular understanding of both canonical and alternative ferroptosis pathways are required. There is a set of widely used chemical agents to modulate ferroptosis using different pathway targets: Erastin blocks cystine-glutamate antiporter, system xc-; ML210 directly inactivate GPX4; L-buthionine sulfoximine (BSO) inhibits γ-glutamylcysteine synthetase, an essential enzyme for glutathione synthesis de novo. Most studies were focused on lipidomic profiling of model systems undergoing death in a ferroptotic modality.
Keywords:
Ferroptosis
; proteomics
; LC-MS/MS
; mouse embryonic fibroblasts
; Pfa1
; cell death
; glutathione peroxidase 4
; erastin
; ML210
; L-Buthionine-sulfoximine
; BSO
1. Summary
Ferroptosis is a unique type of non-apoptotic regulated cell death whose distinguishing features are the iron-dependent manner and overwhelming lipid peroxidation downstream of metabolic dysfunctions [1]. Lipid peroxidation is stimulated by the presence of iron in the cell, due to its ability to enter into free radical reactions according to the Fenton mechanism and generate reactive oxygen species (ROS) [2]. One of the key regulators of the ferroptosis control is a glutathione peroxidase 4 (GPX4), which is able to reduce lipid peroxides, using glutathione (GSH) as an electron donor. This enzyme is a component of the crucial ferroptosis suppressing system - the cyst(e)ine/GSH/GPX4 axis [3]. Currently, a growing number of diseases are associated with redox-stress, and many of them have known hallmarks of ferroptosis, such as lipid peroxidation [4].
There are a few widely used drugs for ferroptotic induction, among them Erastin, ML210, and L-buthionine sulfoximine (BSO). All of them inhibit different components of the cyst(e)ine/GSH/GPX4 axis. Erastin can block cystine-glutamate antiporter, system xc-, which plays the main role in cystine transportation in cells [5,6]. BSO inhibits γ-glutamylcysteine synthetase, an essential enzyme for glutathione synthesis de novo. Finally, ML210 is the most specific GPX4 inhibitor that directly inactivates it without interacting with other glutathione peroxidases [7].
Using different mechanisms of ferroptosis induction extend the ability of ferroptosis modulation that could be extremely helpful in understanding the molecular mechanisms of ferroptosis-related diseases and in researching therapeutic potential of the ferroptosis [8,9,10,11]. Since the canonical ferroptosis pathway leads to phospholipid hydroperoxides accumulation, most studies with omics were focused on lipidomic profile [12,13].
In this study, we made a bottom-up shotgun proteomic for three wide spread chemical proferroptotic inducers (Erastin, ML210 and BSO) at two time points. For each drug we performed an independent series of Liquid Chromatography-Mass Spectrometry (LC-MS/MS) in DDA mode at three conditions: control (without treatment), 24 and 48 hours after the ferroptosis induction. For each condition, we made four biological replicates, each in two or three technical replicates. We provided both the raw data, and the bioinformatically processed data at the protein level via PRoteomics IDEntifications database (PRIDE).
For our knowledge, this is the first comprehensive experimental study that estimates the proteomic shift in mouse embryonic fibroblasts for these three widely used chemical ferroptosis inducers (Erastin, ML210 and BSO). As a valuable dynamic proteome reference, this data could be helpful for further investigation of ferroptosis' role in disease pathogenesis and for searching for new biomarkers in cancer.
2. Data Description
In this study, we presented the proteomic shift during Erastin, ML210 and BSO-induced ferroptosis derived from the mouse embryonic fibroblasts cell lines with tamoxifen-induced GPX4 KO (Pfa1 line). This model (Pfa1 immortalized cell line) was previously established and is widely used for ferroptosis investigation [14]. We measured the proteome landscape dynamic for three chemical ferroptosis inducers: Erastin, ML210 and L-Buthionine-sulfoximine (BSO). For each drug, we conducted an individual series of Liquid Chromatography-Mass Spectrometry (LC-MS/MS) based shotgun proteomics in three time points: control 0h, 24h and 48h after the treatment. Four biological replicates in two or three technical replicates for each condition in each time point were made in DDA mode (Table 1). In total 27 samples in ML210 experiment; 24 samples in Erastin and 24 samples in BSO experiment.
For all samples we provided both raw and processed data using publicly available protein exchange repository PRIDE with the following unique identifiers: samples from ML210 experiment are available as PXD041327; Erastin –PXD041463; BSO – PXD041415. Raw LC-MS/MS spectra are provided as original Thermo™ RAW format files. For label free quantification (LFQ) of proteins, we used IonQuant after both MSFragger spectra alignment and merging technical replicates at the post-search analysis (see methods for details). Thus, we deposited one quantity table per treatment at the protein level for each of the biological replicates in the public repository PRIDE.
Initial quality control analysis revealed high quality samples with stable protein expression distribution across all samples in each of the experiments (Figure 1) with the median value for log2-intensities equal to 29.5 (27.5–31.5) across all samples.
Initially, we detected 3409 proteins in ML210 experiment, 3942 proteins in Erastin experiment and 3088 in BSO experiment. Removing low confidence proteins (see methods) revealed 2828 proteins in ML210, 3061 proteins - in Erastin and 2550 in BSO experiments with similar distribution of non-NA proteins across samples (Figure 2ABC). The sets of proteins detected in different experiments showed a significant overlap and most proteins (2278) were detected in all three experiments (Figure 2D).
To summarize, we provide high-quality shotgun proteome data that was obtained during ferroptosis induction by three widely used chemical agents: Erastin, ML210 and BSO. All data is publicly available on PRIDE as the following IDs: PXD041463 for Erastin, PXD041327 for ML210 and PXD041415 for BSO and include both processed and raw files. The presence of the processed data at the protein level could provide new information at the protein level and be a valuable source for searching new proteins in ferroptosis related pathogenesis (e.g. neurodegenerative diseases) or searching for new biomarkers in therapy-resistant cancers. The presence of raw data with the control samples in each of the experiments allows further accurate comparison between these data and newly generated ones using similar techniques.
3. Methods
Cell culture
Pfa1 cells which are 4-OH-TAM-inducible Gpx4−/− mouse immortalized fibroblasts were provided by Marcus Conrad (Munich, Germany). Pfa1 cells were cultured in RPMI-1640 medium (PanEko) containing glucose (2 g/L), fetal bovine serum (10%), Gibco GlutaMAX Supplement (2 mM) and pen/strep (1%) at 37 °C with 5% CO2 in a humidified incubator. For ferroptosis induction Pfa1 cells (300.000) were seeded on Corning 100 mm tissue-culture treated culture dishes (#CLS430167) and incubated overnight. On the next day cells were treated with or without: i) 0.3 µM ML210 (SML0521, Sigma-Aldrich), ii) 0.5 µM Erastin (E7781, Sigma-Aldrich), iii) 50 µM L-Buthionine-sulfoximine (B2515, Sigma-Aldrich). Cells were collected for further analysis 24h and 48h after treatment.
Protein Extraction and preparation for MS Analysis
The original trifluoroethanol (TFE) based protocol [15] was modified for sample preparation. Cells (about 1 mln) were washed (x3) with PBS, then treated with 80% cold methanol, scraped out, incubated (1h, -80C), and centrifuged. The pellet was washed with cold acetone, centrifuged and dried on air. The proteins were redissolved by ultrasonication in 120 ul of 50 mM ammonium-bicarbonate (ABC) buffer pH 8.0 mixture with TFE (1:1), then: +5 mM TCEP (1h, 50C) and +15 mM iodoacetamide (1h, RT). The samples were diluted 4 times with ABC buffer, 2 ug of trypsin/LysC mix were added, incubated (overnight, 37C), and the reaction stopped by adding formic acid (1%). Peptides were dried in vacuum centrifuge (45C, 3h), redissolved and measured with BCA assay.
LC-MS/MS Analyses
LC-MS/MS was performed on a Q Exactive HF-X mass-spectrometer coupled with UltiMate 3000 nano-flow liquid chromatography system. 1 µg of peptide mixture was loaded onto an Acclaim µ-Precolumn enrichment column (0.5 mm × 3 mm, 5 µm) at 5 μl/min flow for 5 min in isocratic mode using buffer C as the mobile phase (2% Acetonitrile, 0.1% formic acid in DI water). Then the peptides were separated on an Acclaim Pepmap C18 column (75 μm × 500 mm, 3 μm) (in ERA and ML210 experiments) or Acclaim Pepmap RSLC column (75 μm × 750 mm, 1.7 μm) (in BSO experiment) in a gradient of solvents A and B, where A: 0.1% formic acid, B: 80% acetonitrile, 0.1% formic acid in DI water, at a flow rate 0.3 µl/min. In ERA and ML210 experiments the gradient consisted of the following steps: 4% B (5 min), 4 to 33% B (100 min), 33 to 45% B (30 min), 45 to 95% B (1 min), 95% B (20 min), 95 to 4% B (1 min), 4% B (30 min) while in BSO experiment the steps were: 4% B (6 min), 4 to 30% B (95 min), 30 to 50% B (25 min), 50 to 95% B (2 min), 95% B (15 min), 95 to 4% B (2 min), 4% B (20 min).
MS/MS-analysis was performed in positive ionization mode using a NanoFlex source. The parameters for the MS/MS-analysis were as follows: emitter voltage: 2.2 kV, capillary temperature: 260°C. Panoramic scan: the mass range 350 - 1500 m/z, resolution 60,000. Tandem scan: mass range 100 m/z - precursor m/z, resolution 15,000. Isolation window: ± 0.6 Da. DDA mode was set to “top20”, the intensity cutoff limit for precursor selection: 44000, NCE: 28 units. Only ions with charge states from +1 to +6 were sent to tandem scanning. The maximum accumulation time was 60 ms for precursor ions and 45 ms for fragment ions. The AGC values for precursor ions: 3 × 106, for fragment ions: 2 × 105. All the measured precursor ions were dynamically excluded from the tandem MS/MS analysis for 30 s.
Four biological replicates were prepared, and two technical replicates were acquired for each sample: all these LC-MS2 spectra are uploaded.
Data processing and protein identification
Raw files were converted into *.mzML format using msConvert console application [16]. Next, we used MSFragger 3.5 to align spectra to the protein reference with the following parameters: true precursor tolerance of 12 ppm and fragment mass tolerance of 50 mDa [17]. As a reference we used standard mouse SwissProt proteome (UP000000589) augmented with standard contaminants of the Philosopher package [18] with FBS contaminants measured previously in our lab on FBS samples with the same instrument. To exclude false-positive results, we made target-decoy analysis on the samples with grouped technical replicates by peptideProphet [19] and used proteinProphet [20] for further statistical clarification. Label free quantification was conducted by IonQuant 1.8 [21] with match-between-run option turned on. Only proteins identified by two or more unique/razor peptides were considered for further analysis.
Author Contributions
Conceptualization, A.S., A.N. and O.K..; validation, O.K. and A.N.; formal analysis, O.K. and A.N.; investigation, D.K. and V.S.; resources, A.S.; data curation, O.K. and A.N.; writing—original draft preparation, O.K., V.S. and V.T.; writing—review and editing, A.S., O.K. and A.N.; visualization, O.K.; supervision, V.B.; project administration, A.S..; funding acquisition, A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation (RSF), grant number 21-74-00140.
Institutional Review Board Statement
Not applicable, all experiments were performed on immortalized cell line.
Informed Consent Statement
Not applicable.
Data Availability Statement
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE [22] partner repository with the following dataset identifiers: Erastin experiment - PXD041463; ML210 experiment - PXD041327; BSO - PXD041415.
Acknowledgments
We thank Dr. Marcus Conrad for providing Pfa1 cell line.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jiang, X.; Stockwell, B.R.; Conrad, M. Ferroptosis: Mechanisms, Biology and Role in Disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 266–282. [Google Scholar] [CrossRef]
- Gao, M.; Monian, P.; Pan, Q.; Zhang, W.; Xiang, J.; Jiang, X. Ferroptosis Is an Autophagic Cell Death Process. Cell Res. 2016, 26, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Friedmann Angeli, J.P.; Schneider, M.; Proneth, B.; Tyurina, Y.Y.; Tyurin, V.A.; Hammond, V.J.; Herbach, N.; Aichler, M.; Walch, A.; Eggenhofer, E.; et al. Inactivation of the Ferroptosis Regulator Gpx4 Triggers Acute Renal Failure in Mice. Nat. Cell Biol. 2014, 16, 1180–1191. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Conrad, M. The Metabolic Underpinnings of Ferroptosis. Cell Metab. 2020, 32, 920–937. [Google Scholar] [CrossRef] [PubMed]
- Dolma, S.; Lessnick, S.L.; Hahn, W.C.; Stockwell, B.R. Identification of Genotype-Selective Antitumor Agents Using Synthetic Lethal Chemical Screening in Engineered Human Tumor Cells. Cancer Cell 2003, 3, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An Iron-Dependent Form of Nonapoptotic Cell Death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Eaton, J.K.; Furst, L.; Ruberto, R.A.; Moosmayer, D.; Hilpmann, A.; Ryan, M.J.; Zimmermann, K.; Cai, L.L.; Niehues, M.; Badock, V.; et al. Selective Covalent Targeting of GPX4 Using Masked Nitrile-Oxide Electrophiles. Nat. Chem. Biol. 2020, 16, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Reichert, C.O.; de Freitas, F.A.; Sampaio-Silva, J.; Rokita-Rosa, L.; Barros, P. de L.; Levy, D.; Bydlowski, S.P. Ferroptosis Mechanisms Involved in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 8765. [Google Scholar] [CrossRef]
- Ren, J.-X.; Sun, X.; Yan, X.-L.; Guo, Z.-N.; Yang, Y. Ferroptosis in Neurological Diseases. Front. Cell. Neurosci. 2020, 14, 218. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, X.; Jin, S.; Chen, Y.; Guo, R. Ferroptosis in Cancer Therapy: A Novel Approach to Reversing Drug Resistance. Mol. Cancer 2022, 21, 47. [Google Scholar] [CrossRef]
- Nie, Q.; Hu, Y.; Yu, X.; Li, X.; Fang, X. Induction and Application of Ferroptosis in Cancer Therapy. Cancer Cell Int. 2022, 22, 12. [Google Scholar] [CrossRef] [PubMed]
- Mishima, E.; Ito, J.; Wu, Z.; Nakamura, T.; Wahida, A.; Doll, S.; Tonnus, W.; Nepachalovich, P.; Eggenhofer, E.; Aldrovandi, M.; et al. A Non-Canonical Vitamin K Cycle Is a Potent Ferroptosis Suppressor. Nature 2022, 608, 778–783. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Henry, W.S.; Ricq, E.L.; Graham, E.T.; Phadnis, V.V.; Maretich, P.; Paradkar, S.; Boehnke, N.; Deik, A.A.; Reinhardt, F.; et al. Plasticity of Ether Lipids Promotes Ferroptosis Susceptibility and Evasion. Nature 2020, 585, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Seiler, A.; Schneider, M.; Förster, H.; Roth, S.; Wirth, E.K.; Culmsee, C.; Plesnila, N.; Kremmer, E.; Rådmark, O.; Wurst, W.; et al. Glutathione Peroxidase 4 Senses and Translates Oxidative Stress into 12/15-Lipoxygenase Dependent- and AIF-Mediated Cell Death. Cell Metab. 2008, 8, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Qian, W.-J.; Mottaz, H.M.; Clauss, T.R.W.; Anderson, D.J.; Moore, R.J.; Camp, D.G.; Khan, A.H.; Sforza, D.M.; Pallavicini, M.; et al. Development and Evaluation of a Micro- and Nanoscale Proteomic Sample Preparation Method. J. Proteome Res. 2005, 4, 2397–2403. [Google Scholar] [CrossRef] [PubMed]
- Hulstaert, N.; Shofstahl, J.; Sachsenberg, T.; Walzer, M.; Barsnes, H.; Martens, L.; Perez-Riverol, Y. ThermoRawFileParser: Modular, Scalable, and Cross-Platform RAW File Conversion. J. Proteome Res. 2020, 19, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Kong, A.T.; Leprevost, F.V.; Avtonomov, D.M.; Mellacheruvu, D.; Nesvizhskii, A.I. MSFragger: Ultrafast and Comprehensive Peptide Identification in Mass Spectrometry–Based Proteomics. Nat. Methods 2017, 14, 513–520. [Google Scholar] [CrossRef]
- da Veiga Leprevost, F.; Haynes, S.E.; Avtonomov, D.M.; Chang, H.-Y.; Shanmugam, A.K.; Mellacheruvu, D.; Kong, A.T.; Nesvizhskii, A.I. Philosopher: A Versatile Toolkit for Shotgun Proteomics Data Analysis. Nat. Methods 2020, 17, 869–870. [Google Scholar] [CrossRef]
- Keller, A.; Nesvizhskii, A.I.; Kolker, E.; Aebersold, R. Empirical Statistical Model To Estimate the Accuracy of Peptide Identifications Made by MS/MS and Database Search. Anal. Chem. 2002, 74, 5383–5392. [Google Scholar] [CrossRef]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A Statistical Model for Identifying Proteins by Tandem Mass Spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef]
- Yu, F.; Haynes, S.E.; Nesvizhskii, A.I. IonQuant Enables Accurate and Sensitive Label-Free Quantification With FDR-Controlled Match-Between-Runs. Mol. Cell. Proteomics 2021, 20, 100077. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Distribution of the log2(Intensity) across all detected proteins in each of the samples (x axis). Number corresponds to the biological replicates. Ctrl - control; ml - ML210; ERA - Erastin.
Figure 1.
Distribution of the log2(Intensity) across all detected proteins in each of the samples (x axis). Number corresponds to the biological replicates. Ctrl - control; ml - ML210; ERA - Erastin.
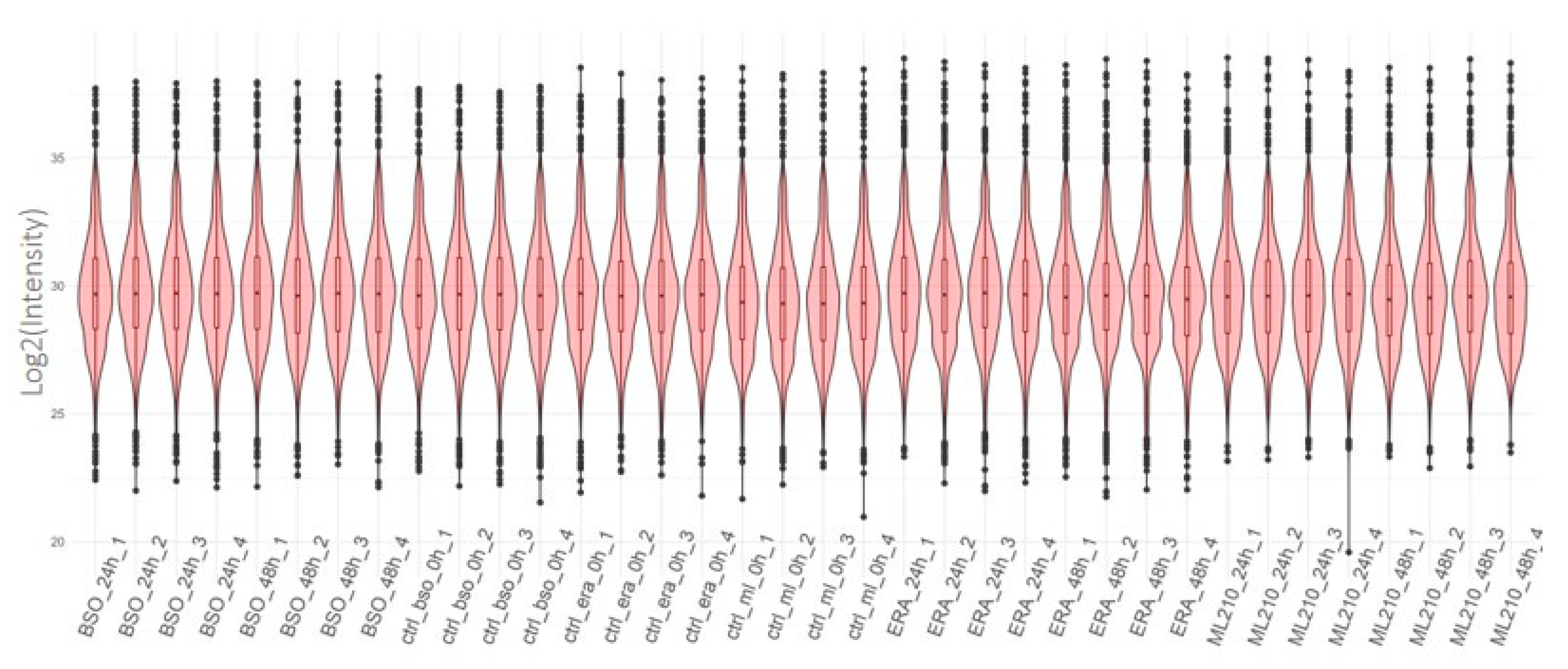
Figure 2.
Proteome data overview. (a) Number of detected proteins across biological replicates in Erastin experiment. (b) Number of detected proteins across biological replicates in ML210 experiment. (c) Number of detected proteins across biological replicates in BSO experiment. (d) Overlapping of the detected proteins across experiments. Ctrl - control; ml - ML210; ERA - Erastin. Number corresponds to the biological replicates.
Figure 2.
Proteome data overview. (a) Number of detected proteins across biological replicates in Erastin experiment. (b) Number of detected proteins across biological replicates in ML210 experiment. (c) Number of detected proteins across biological replicates in BSO experiment. (d) Overlapping of the detected proteins across experiments. Ctrl - control; ml - ML210; ERA - Erastin. Number corresponds to the biological replicates.
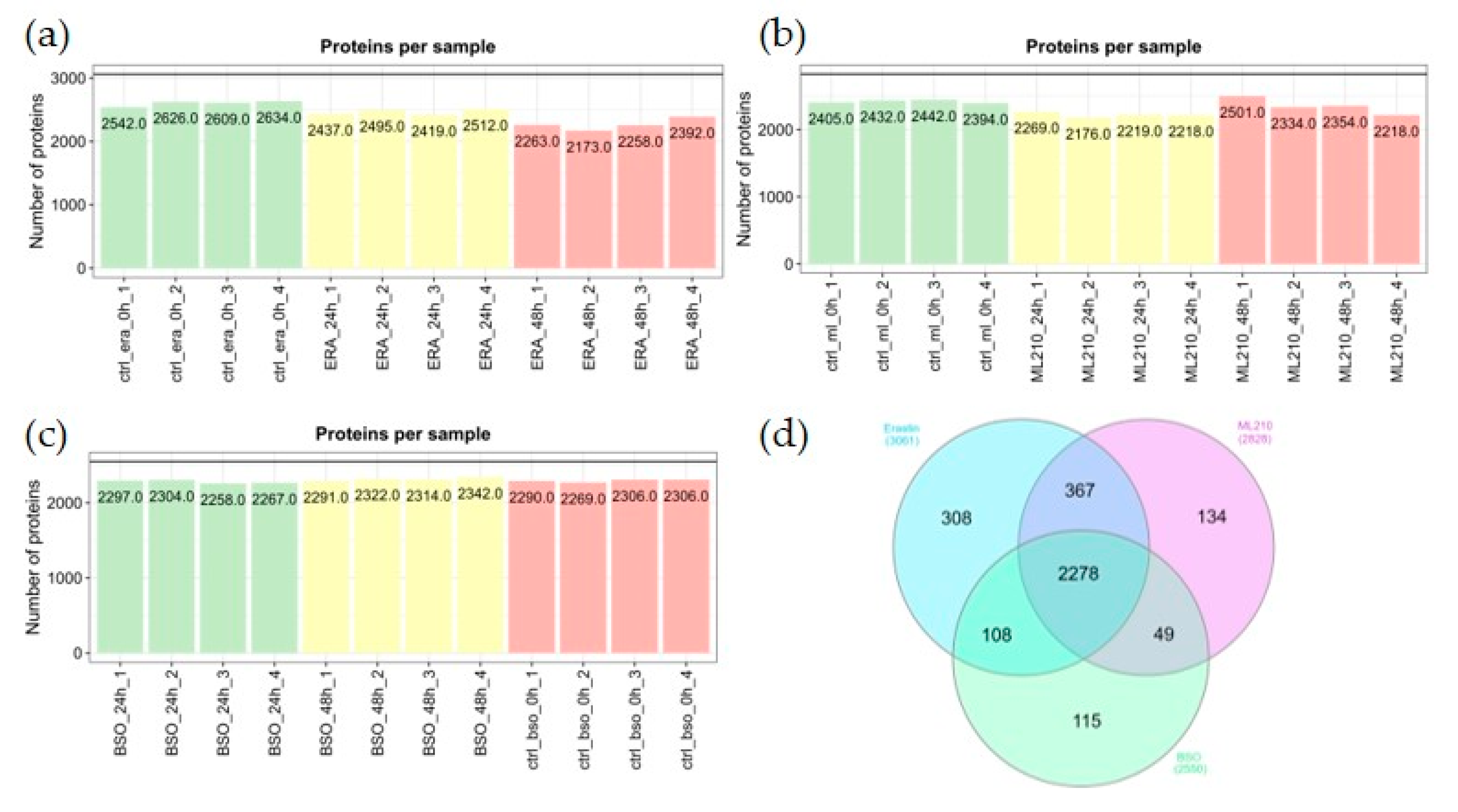

Table 1.
List of the LC-MS/MS proteome sequenced samples. Table represents the number of biological samples multiplied (x) by technical replicates in each of the conditions (ML210, Erastin, BSO) across time points that were sequenced.
Table 1.
List of the LC-MS/MS proteome sequenced samples. Table represents the number of biological samples multiplied (x) by technical replicates in each of the conditions (ML210, Erastin, BSO) across time points that were sequenced.
| Time points / Condition |
Control (0 hours) | After 24 hours | After 48 hours |
|---|---|---|---|
| Erastin | 4 х 2 | 4 х 2 | 4 х 2 |
| ML210 | 4 х 2 | 1 x 3, 3 х 2 | 2 x 3, 2 х 2 |
| BSO | 4 х 2 | 4 х 2 | 4 х 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



