
Submitted:
12 May 2023
Posted:
15 May 2023
You are already at the latest version
Abstract
In the new antibiotic era, the exponential increase of multiresistant bacterial strains become the main global health problem. Many researchers focused their efforts to explore novel or combined strategies for combating bacterial resistance. The good knowledge of molecular mechanisms of resistance and bacterial virulence factors as key targets gives us a good scenario to resolve the problem. One particularly attractive and promising way is to attack the main regulatory “network” of bacterial virulence determinants known as Quorum sensing (QS). The inhibition of QS signals will be a novel way for screening more effective Quorum sensing inhibitors (QSIs) and will put a key role in next-generation antimicrobials in the resistance battle. This determined the aim of the present review: comprehensive clarification of the regulatory mechanisms of quorum-sensing signaling pathways in Chromobacterium violaceum and discovery of potential plant quorum sensing inhibitors.
Keywords:
quorum sensing
; quorum sensing inhibitors
; Chromobacterium violaceum
; plant extracts
1. Introduction
One of the most significant events in human health history starts with the advent of antibiotics and giving people the opportunity of treating bacterial infections. Unfortunately, parallel with this key step is observed increased risk of antibiotic resistance. Moreover, the alarming tendency from the appearance of clinical resistance isolates is requiring the discovery of novel alternative ways to resolve the problem. Today is well known that bacterial cells have developed a regulatory system called QS for intracellular communication. The quorum sensing process is a cell density-dependent biochemical communication between bacteria, that allows them to receive information and respond to different environments (1). Thus, bacteria regulate their gene expression, virulence potential, pathogenicity, antibiotic resistance, etc.
Nowadays is known that the QS system is a promising target for inhibiting and controlling these bacterial activities. The evolution of different natural or synthetic molecules used as QS antagonists can be the next generation of “therapeutic substances” against antibiotic resistance (2,3). The compounds which suppress in one way or another bacterial QS cascade are called Quorum sensing inhibitors (QSIs). This concept is based on the interruption of signaling pathways, controlling virulence factors and microbial survival, which is the aim of a given antimicrobial strategy. QSIs are molecules with the potential to inhibit QS-regulated processes such as bioluminescence, fluorescence, biofilm formation and dispersal, pigment production, enzyme activity, or different reporters, thereby stopping the bacteria communications which in turn leads to control of pathogenicity. The first natural marine QSI was isolated from the Australian alga Delisea pulchra. The authors reveal that exogenous furanones produced by this marine alga reduced QS signals and swarming motility in Serratia liquefaciens MG1 (4).
During the last few years, one of the most popular microorganisms used for QS investigations is C. violaceum. Its indicator ability, related to QS-regulated characteristic - violacein biosynthesis, makes it a suitable microorganism for determining cell-to-cell signaling pathways. This advantage can be helpful for the validation of various qualitative and quantitative tests for describing and characterizing bacterial communications and regulation. The knowledge of these pathways helps identify different mechanisms for interfering with bacterial virulence. For this reason, it is essential to focus scientific efforts on discovering new ways for interrupting QS. It is well known that numerous natural and synthetic compounds have the ability to disrupt QS by interacting with the signal molecules or receptors (5–8). Many studies reveal in more detail their therapeutic and antibacterial functionality (9–13), but as anti-virulence targets are poorly recognizable. It has to be pointed out that good understanding of the QSIs' mode of action, we will answer an important question “Can we interfere or inhibit bacterial cell-to-cell signaling” network”? This provoked us to summarize the last data, about QS mechanisms in Gram-negative bacteria especially in bioreporter strains Chrormobacterium violaceum, and the application of the novel natural QSIs and their main role in this “bacterial network”.
2. Quorum sensing: bacterial communication network
In the 1970s, Nealson et. al. discovered and described QS in two luminous marine bacterial species, Vibrio fischeri and Vibrio harveyi (14–16). Since then, this bacterial feature has been found in many Gram-negative and Gram-positive species (17,18).
In essence, QS is a complex of communication mechanisms between bacteria, based on gene expression, in response to changes in cell-population density (17). This provides control over specific processes such as virulence factor expression (proteases, toxins, adhesins), biofilm formation, sporulation, symbiosis, conjugation, production of secondary metabolites, stress adaptation, horizontal DNA transfer, pigment and antibiotic synthesis, bioluminescence and synthesis of protective molecules such as biosurfactants (8,19–24). This type of bacterial communication occurs due to the synthesis and secretion of chemical signaling molecules called autoinducers (AIs) by bacteria (17,25,26). The concentration of AIs depends on the bacterial population density. In fresh cell cultures, the concentration of AIs is low, but with the increase of cell population, their concentration increase until the so-called threshold concentration is reached (4). That allows the signaling molecule to bind a receptor and activate a signaling cascade leading to a coordinated change in gene expression in the population (8,18). In Gram-negative bacteria, that belong genus Chromobacterium, the main receptors are cytoplasmic transcription factors or transmembrane two-component histidine sensor kinases (8,22). These QS-controlled processes are extremely ineffective and energy-consuming when undertaken by a single cell but effective when managed by a large bacterial group (16). One of the well-studied signals is Autoinducer 2 (AI-2), responsible for interspecies communication and regulates motility, production of virulence factors, and biofilm formation (27–29).
2.1. LuxR receptors
The group of LuxR receptors is found in Gram-negative bacteria and is subdivided into two groups known as typical LuxR-type receptors and LuxR solo receptors (8).
The typical LuxR-type receptor binds the autoinducer аcyl-homoserine lactone (AHLs), synthesized by LuxI synthase. The resulting complex activates the transcription of luciferase operon (luxICDABE) in V. fischeri (8). AHLs are small diffusible molecules with a core lactone ring and an acyl side chain. They are responsible for facilitating signaling in Gram-negative bacteria. In this group of receptors, binding is precise because they bind only specific ligands, ensuring proper communication in the environment. The specificity is achieved by modifications in the R-groups in AHLs and the number of carbon atoms. As the bacteria grow on a medium, they excrete AHLs, and when the threshold concentration is reached, they are brought back into the cells and bind to LuxR. The resulting LuxR-AHL complex binds to the Lux gene promoter, responsible for initiating bioluminescence and other QS-regulated functions (22).
LuxR solo receptors can modulate bacteria to adapt better to the environment or host organism by binding to AHLs or non-AHLs molecules (8). The best-studied solo receptors are QscR in P. aeruginosa, CviR in C. violaceum, and SdiA in E. coli.
QscR receptor in P. aeruginosa is a protein with a conserved amino-terminal AHL-binding domain and a conserved carboxy-terminal DNA-binding domain. Several studies have shown the effect of the protein on the modulation of Las and Rhl regulons, acting in the growth phase (19). It has been discovered that QscR can auto-activate its own expression (29). In addition, in mixed bacterial populations, it may be activated by other non-P. aeruginosa signals molecules, such as products from B. vietnamiensis and Roseobacter gallaeciencis (22,30). Another feature of QscR is its dose-dependent dimerization. QscR is a monomer at low concentrations, but at high concentrations, it dimerizes which is the active form of the receptor (19).
CviR in C. violaceum is thought to bind to more than 20 promoters in the bacterial genome. These promoters are responsible for a few functions, such as gene regulation, motility, coenzyme synthesis, nutrient utilization, and virulence (31,32). It has been observed that CviR affects chitinase production, suggesting that C. violaceum blocks fungal growth in water or soil and gives the bacterium a competitive advantage in the environment (30). A ligand of CviR is a C6-homoserine lactone, synthased by CviI - synthase. The CviR-CviI system is homologous to LuxI- LuxR system found first in Vibrio fischeri (32). The CviR-CviI complex regulates the synthesis of violacein, a purple pigment synthesized by C. violaceum (31,33). The formation of this complex leads to an increase in CviI expression, generating positive feedback (31,32).
The SdiA receptor found in E. coli and Salmonella, like QscR from P. aeruginosa, can recognize AHL molecules synthesized by other bacterial species. Crystallographic studies have revealed that the receptor is a symmetric dimer having an N-terminal ligand-binding domain and a C-terminal DNA binding domain (8). Another feature, established by crystallography and molecular docking technique, is the selectivity of SdiA for short-chain ligands (33). The main functions of SdiA are related to the control of bacterial virulence, cell division, and biofilm formation (8).
2.2. Bicomponent Quorum sensing receptors
Membrane-bound receptors are the best studied in Vibrio harveyi and Vibrio cholerae. In their regulatory system are used two different QS signals. One of the signals is responsible for intraspecies and the other for interspecies communication. In V. harveyi are found three bicomponent receptors- LuxN, LuxPQ, and CqsS connecting HAI-1, AI-2, and CAI-1, respectively. Four receptors have been identified in V. cholerae - LuxPQ, CqsS, CqsR, and VpsS (8). In V. harveyi, these receptors, after binding their ligands, undergo phosphorylation and transfer phosphate to the LuxU protein in the cell, which transfers it to LuxO. Phosphorylated LuxO is involved in activating the expression of five small regulatory RNAs (sRNAs). They promote the translation of AphA and inhibit the translation of LuxR (34,35). Several years ago, scientists proved that the amount of LuxN is higher than the concentration of LuxQ and CqsS and further increased in the late exponential growth phase (34). As a result of this biochemical cascade, bioluminescence, metalloproteinases, iron carrier, exopolysaccharide production, and negative type III secretion are regulated (35).
In V. cholerae, the four receptors mentioned above are histidine kinases, which, by reversible phosphorylation, regulate QS in the bacterial population. At low cell densities, the four kinases trigger a cascade identical to that in V. harveyi. At high cell densities, each receptor kinase binds to its AI, inhibits phosphorylation throughout the chain, and activates the translation of HapR, responsible for the virulence of the species. Yet, it is unclear why four kinases are needed to maintain V. cholerae colonization in hosts (36).
In Gram-negative bacteria, this bacterial communication network, in which bacteria produce and respond to specific signals and induce changes in gene expression, is the main strategy to occupy a particular niche. It is mostly used when nutrient and energy sources are limited. Most pathogenic bacteria use this “clever system” to promote infectious diseases.
3. Quorum-Sensing system in Chromobacterium violaceum
C. violaceum is a free-living Gram-negative, facultative anaerobe, non-sporulating β-proteobacterium first described in the 19th century. It dominates in a variety of ecosystems in subtropical and tropical regions and is mainly found in water and soil, also on the shores of river Negro, a big part of the Brazilian Amazon (37,38). Due to broad distribution, it is a cosmopolitan microorganism (33). It is a typical saprophyte that becomes an aggressive opportunistic pathogen causing severe, most of the time, fatal animal, and human infections with a high mortality rate (38). C. violaceum can cause respiratory and gastrointestinal infections, liver abscesses, endocarditis, meningitis, hemophagocytic syndrome, and fulminant sepsis (32) in humans by entering the bloodstream through an open wound (39). It is oxidase and catalase positive microorganism. The optimal growth temperature is at 30-35°C. C. violaceum is a rod with a rounded end and its size is 0.6–0.9 × 1.5–3.0 μm. Also, it has a single polar flagellum (32). C. violaceum is resistant to a wide range of antibiotics, mainly beta-lactams like penicillin, ampicillin, and cephalosporins (33).
These bacteria form smooth violet colonies on common laboratory media. The color comes from the violacein pigment, encoded by the vio operon, whose expression is QS-regulated. This trait is easily observed and quantified, therefore this bacteria has been widely used as a model organism for QS research in laboratories (32). Moreover, the bacteria is used to study the inhibition of AHLs-mediated QS by different compounds and for assaying the production of short-chain AHLs because AHL-QS controls the synthesis of the pigment violacein (40). Data have been reported for non-pigmented isolates, however, the pigmented cultures were found to survive longer and produce more exopolysaccharides than the non-pigmented isolates (41,42).
The ability to live in different environmental conditions is because of the energy-generating metabolism that can use a wide range of substrates by using oxidases and reductases. Thus, aerobic and anaerobic respiration are permitted. When oxygen is in total absence, fumarate and nitrate are used as final electron acceptors. In addition, the chemotactic capacity of C. violaceum is essential to survive in a diversity of environmental conditions. The genome of C. violaceum consists of a single circular chromosome of 4.75108 Mbp with a G+C content of 64.83%. The complete genome sequence reveals some key characteristics: (i) the presence of vast alternative pathways for energy metabolism, (ii) open reading frames (ORFs) for transport proteins (iii) complex systems for stress adaptation and motility, and (iv) the usage of QS to control different inducible systems, which promote flexibility and adaptability (37). In the genome are found 4431 ORFs responsible for energy generation, transport, signal transduction, motility, secretion, and secondary metabolism, which is important for proteins causing mammalian pathogenicity (38).
3.1. Quorum sensing mechanisms in Chromobacterium violaceum
C. violaceum communicates through QS via C6-homoserine lactone signal (C6-HSL) (40). This bacterium uses a LuxIR-type QS system consisting of four main components: a CviI synthase (N-hexanoyl-L-homoserine lactone synthase), AHLs diffusible molecule called AI; a CviR-cytoplasmic receptor (DNA-binding transcription factor), and target genes (43). The protein CviI synthase, a product of the cviI gene, synthesizes the AI C6-homoserine lactone (C6-HSL), CviR binds to it, and thus, gene expression is activated. Recently, it was determined the consensus DNA sequence for promoter recognition by CviR and it was found potential 53 binding sites. Further experiments confirmed that CviR binds to six different promoters and modulates the transcription of vioA (part from violacein synthesis cluster), CV_4240 (chitinase), and cviI (HSL synthase), therefore, taking part in a classical QS positive-feedback loop (40).
QS control lytic activity by exoproteases, chitinases, and virulence factors like the type VI secretion system (20,38). Furthermore, QS regulates type II (TIISS) and type III (TIIISS) secretion systems, swarming motility, lipases, flagellar proteins, collagenase, elastase, and cyanide production (32,33). In addition, QS also regulates resistance to a few antimicrobials, including bactobolin, where QS-controlled resistance is carried out by an efflux pump (43). Another important activity discovered in C. violaceum is biofilm formation, which is responsible for virulence through resistance to antibiotics, phagocytosis, and disinfectants. It has been established that in biofilms bacteria communicate through diffusible AIs (38). The secretion of the mentioned virulence factors, together with the formation of biofilm, are important for initiating infection in the host cells, and therefore, developing antimicrobial resistance (33,44).
3.2. Pigment production
Violacein is а bisindole derivative, biosynthesized by condensation of two molecules of L-tryptophan by the products of the vioABCDE operon in response to QS (31,45). It is а bioactive secondary metabolite with a putative function as a respiratory pigment that is not essential for bacterial survival and growth. Also, its role in the regulation of tryptophan synthesis has been demonstrated (32). The pigment violacein has biocidal activity against different kingdoms (bacteria, fungi, viruses, nematodes, etc.) during the microbial stationary phase of growth when cell density is high, and nutrients are limited. Hence, its production could be considered as a part of the competitive strategy to extend the duration of life of the microbial colony (20). In addition, it shows a synergistic antimicrobial effect with different antibiotics against pathogenic bacteria. Furthermore, violacein can be used as a bio-dye because of its good color tone and long-lasting stability (45). Synthesis of this visible and quantifiable pigment provides a simple way for searching for potential QSIs and gives the prospect of developing new biosensor strains. A similar application finds the biosensor strain CV026 which is mini-T5-mutant defective in AHL synthase because it lacks cviI and thus requires the addition of exogenous AHL signal molecules for violacein production. Such mutants find application in the detection of bacterial AHLs molecules in any environment (32). Furthermore, the fact that violacein production is QS-dependent, makes it a suitable marker for detecting and estimating the potential of new QSIs extracted from plants (40).
The violacein operon is negatively regulated by a new repressor protein VioS and positively by the CviI/R system. VioS does not regulate the CviI/R system. Shortly, at high cell density, the CviR protein binds AHLs and activates the expression of the vioA promoter and at the same time, the vioA promoter is suppressed by the expression of VioS, so violacein is not produced. The colonies of wild-type C. violaceum ATCC 31532 are pale. A vioS mutant that lacks this repression at the vioA promoter forms visible violet colonies (46).
C. violaceum is one of the most used bacterial species in QS research. It is widely used for finding new ways to disrupt the QS system. Violacein production is easily detected and quantified and thus is used for screening potential QSI molecules (Figure 1). Disruption of QS can decrease the secretion of virulence factors without killing the bacterium or inhibiting its growth (47). This allows reducing the selective pressure on the pathogen leading to not developing resistance. QSIs can be used as alternatives to conventional antibiotics (32).
4. Plant Inhibitors: a new way to control bacterial communication
One of the most impressive processes in microbiology is the ability of bacteria to communicate with each other via signal molecules (48). This type of bacterial communication coordinates the accumulation and responses to small molecules called AIs (7,8,49,50). The process known as QS allows the bacterial community to coordinate the gene expression, leading to the activation of specific phenotypes in the population. The most common processes that are under QS control, used by bacteria as a survival strategy, are bioluminescence, biofilm formation and dispersal, virulence factors expression, motility, pigment synthesis, sporulation, conjugation, symbiosis, antibiotic production (5–8).
During the antibiotic century, the revolution of better human health was a good scenario. Unfortunately, this development has led to an increase in bacterial resistance. Nowadays, it is necessary the discovery of new targets for inhibiting microbial pathogenicity, without stimulating microbial resistance (7). One of the most novel anti-virulence strategies is to interrupt the cascade of the QS system (51,52). Each step of the QS signaling cascade, could be a good target and result in the inhibition of pathogenicity (53). Some of the most attractive biomolecules, which could be used for this purpose, are natural QSIs (7,51). Similar inhibitors, that can mediate bacterial QS, are described in different marine algae, fungi, corals, tunicates, cyanobacteria (54–57), bacterial (7,58,59) and mammalian cells (60). Most were isolated from plant cells (61–65).
Bearing this in mind, our major interest is focused on QSIs isolated from plants, including their medicinal and anti-QS properties found in C. violaceum.
The plant kingdom is one of the most popular with their species and families, whose metabolite products have broad biological activities. It is well known about the antimicrobial activities of different plant extracts (12,13,66–68), essential oils (22,69), fractions, and their constituent, but their efficacy against QS systems are poorly understood. During the last few years, it has been found that plant extracts can act as inhibitors of QS pathways. Such active metabolites can be extracted from different parts of plant tissues such as roots, stems, leaves, bark, fruits, flowers, seeds, and green pod (70–73). Major groups of these compounds can be identified as QSIs including cyclic compounds, phenolic derivatives, nitrogen cyclics, furanones, lactones, cinnamaldehydes, alkaloids, phenolics, saponins, tannins, and terpenoids (74,75). Their functionality is different - they can inhibit bioluminescence, fluorescence, biofilm formation, pigment production, block enzyme activity, and inhibit a variety of reporters (7,12,13). These abilities depend on their chemical structure and stability. To interfere with signal acceptance, QSIs must be competitive and non-competitive molecules that prevent binding the signal by its similar receptor. It is essential to know that for competitive molecules to bind to a receptor, they must be structurally similar to the original signal molecules. Non-competitive binding molecules will bind to a site different from the signal-binding site on the receptor. Several scenarios have been known using plants molecules or metabolites as QSIs: a) homologically masking the QS signal and disrupting the bacterial communication; b) interfering with different enzymes; c) preventing the accumulation of signals; d) blocking the main receptors (22,74).
4.1. Quorum sensing inhibitory potential by plants
In the environment, plants are constantly exposed to a wide range of stress conditions. These stress factors that affect plants are temperature changes, nutrient deficiency, drought, salinity, UV radiation, lack of oxygen, pesticides, pollutants, and anthropogenic activities. Apart from environmental stress, some species such as bacteria, fungi, viruses, nematodes, and insects can cause distress. Plants have been facing most of their attackers for more than a million years. Living with their natural enemies in reciprocal evolutionary interaction, they have been learning and developing mechanisms for resisting stress and attacks. Due to this reason, plants reveal an “immune system” comparable to that of animals by biosynthesizing active compounds and secondary metabolites as a protection against infections or induce in response to pathogen attacks. Besides improving the defense against both biotic and abiotic stresses, most of the secondary metabolites have therapeutic activities: anticancer, antioxidant, antidiabetic, immunosuppressive, antifungal, anti-inflammatory, antimalarial, anti-oomycete, antibacterial, anti-fever, anti-diabetic, insecticidal, anti-biofilm and antiviral agents (9,10,12,13,68).
Lately, one of the most interesting QSIs applications is their ability to block signaling molecules produced by bacteria and consequently obstruct the bacterial virulence factors by disrupting their QS system. For this reason, the bacterial QS system is an excellent target for novel QSIs. Scientific evidence proves that the identification of the binding conformation of QSIs into the binding sites of main proteins, by molecular docking analysis, provides new information about their antagonistic characteristics (76). QSIs have been reported in many plants including medicinal plants like Syzygium cumini, Pimenta dioica, Psidium guajava, Medicago truncatula, Lotus corniculatus, Pisum sativum, Moringa oleifera, Vernonia blumeoides, Tecoma capensis, and many others (7,75,77). Their acetone, methanol, and water extracts have been proven to possess quorum sensing inhibitory activity against C. violaceum.
Our review represents summarized information on the plant QSIs, comprehensively studied in C. violaceum. C. violaceum is Gram-negative bacteria, easily cultivated on laboratory media like Blood agar, MacConkey agar, and Nutrient agar. It produces smooth violet colonies whose color comes from a violet antioxidant pigment known as violacein. The increased interest in research communities about C. violaceum is related to its phenotypic characteristics: violacein production, elastase production, biofilm formation, and cyanide production controlled from the QS system by the signal molecules - AHLs.
Many years ago, plants have been studied for their medical values (as digestives, diuretics, expectorants, and sedatives), and also for antioxidant and antimicrobial activities, which further develop the basis of modern phytotherapy. The main interests in their biological functions and mode of action for regulating bacterial communications escalated during the last years. Studies include tests on crude or ethanol, methanol, acetone, ethyl acetate, dichloromethane, hexane and water extracts, essential oils, and phytochemicals partially purified, enriched, or pure fractions. All these plant products could suppress the production of pigment violacein, biofilm formation, motility, and microbial activity in C. violaceum (Table 1).
Koh and Tham (78) screened ten Chinese medicinal plants, Prunus armeniaca, Prunella vulgaris, Nelumbo nucifera, Panax notoginseng (root and flower), Punica granatum, Areca catechu, and Imperata cylindrical for their QS activity. Seven of the extracts inhibit QS in bioreporter strain C. violaceum CV026. Part of the tested compounds have the potential to suppress violacein synthesis and six of them formed a clear zone, indicating antimicrobial activity. These results could be compared to other aqueous extracts from Ananas comosus, Musa paradiciaca, Manilkara zapota, Ocimum sanctum, Camellia sinensis L., Nymphaea tetragona and Quercus cortex whose active components were responsible only for inhibition of synthesis of pigment violacein in C. violaceum CV026 and ATCC 12472 (50,82,103). Important observations were discovered about methanol extracts from herbal plants like Pisum sativum, Trigonella foenum graecum, Myristica cinnamomea, Kigelia africana, Melicope lunuankenda, Cuminum cyminum, Costus speciosus, that proved to be inhibitors of violacein production (50,79,81,83,93,95). Bioscreening of ethanol extracts from Egypt`s ornamental and medicinal plants and such collected from Jordan, such as Adhatoda vasica, Bauhinia purpurea L., Lantana camara L., Myoporum laetum, Piper longum L., and Taraxacum officinale, Laurus nobilis L., Populus alba L., Populus nigra L., Lavandula angustifolia, Rosmarinus officinalis L., Sonchus oleraceus L., Tecoma capensis Thunb. Lindl., Jasminum sambac Ait., reveals anti-microbial activity against C. violaceum (71,87). In contrast with this, ethanol extracts from Cinnamomum zeylanicum, Ocimum basilicum, and Scutellaria baicalensis Georgi, demonstrate violacein inhibition on C. violaceum CV12472 and QS inhibition on C. violaceum CV026 (90). Similar results with ethanol extracts, but obtained from Acer monspessulanum subsp. Monspessulanum were reported by Ceylan et. al. (89). The authors determined the violacein inhibition in C. violaceum CV 12472, CV026, and the anti-QS activity of ethanol extracts. Fatima also used the same bioreporter strains to detect the QS regulatory role of ethanol seed extracts from leguminous plants Pisum sativum and Trigonella foenum graecum (78). Eight fractions, including phenolic (gallic acid, ellagic acid, epicatechin, rutin) from green pods of Acacia nilotica have been studied for their capacity to inhibit pigment production in C. violaceum 12472, as two of them can be classified as QSIs with the potential to regulate violacein production, without influencing bacterial growth. Other phenolic plant extracts of Rubus rosaefolius also have shown a similar effect on pigmentation and biofilm formation (72,97). Polyphenolic extracts from Rosa rugosa have been the focus of Zhang et. al. (101) research, with their anti-biofilm and QS inhibitory potentials. The authors proved high pigment reduction without changes in microbial growth. Indian medicinal plants, flowers seeds, barks, and fruits from Punica granatum, Syzygium cumini, Embelia ribes, Phyllanthus emblica, Terminalia bellirica, Terminalia chebula, Punica granatum, Mangifera indica, Acacia arabica, Terminalia arjuna, Thespesia populnea, Casuarina equisetifolia, were screened for the anti-QS activity where tannin-rich extracts and punicalagin influence QS mechanisms as decreased on violacein synthesis. Shukla and Bhathena (100) qualify this phenomenon in the presence of subinhibitory concentrations of tannin extracts (97,102).
The ethyl acetate fractions and eugenol of Syzygium cumini L. and Pimenta dioica L. displayed significant anti-QS activity by inhibiting the pigment production by C. violaceum (85,88,91). Extracts from different plants like Rhizophora annamalayana (bark), Astilbe rivularis, Fragaria nubicola, Osbeckia nepalensis, Dionysia revolute, Eucalyptus camaldulensis, Green tea, Amomum tsaoko, Punica granatum, and Saraca asoca barks (stem), were found to possess QS activities, but most of them against violet pigmentation of Chromobacterium (50,82,91,94,96,104).
Essential oils (EOs) have been produced in more aromatic species and stored in various plant organs, e. g., flowers, leaves, wood, roots, rhizomes, fruit seedling, and seeds. They are secondary metabolites from plant sources, characterized by natural multicomponent systems composed mainly from terpenes (monoterpenes, sesquiterpenes, and diterpenes), oxygenated compounds, which are mainly phenols, alcohols, aldehydes, ketones, esters, oxides, and hydrocarbons. Essential oils and their constituents are important for biomedical or pharmaceutic purposes due to their bactericidal, virucidal, fungicidal, analgesic, sedative, anti-inflammatory, spasmolytic, and local anesthetic properties (105,106).
Among the plant products, essential oils are most popular for their widespread use in ethnomedicine. Some of them, isolated from three species of the genus Piper growing in Colombia, Piper bredemeyer, Piper bogotense, and Piper brachypodon interfere the pigment production and proved minor effect against bacterial growth in C. violaceum CV026 as well (84). Likewise, the four Eos prepared from Cinnamomum verum, Origanum majorana, Thymus vulgaris, and Eugenia caryophyllata were evaluated as QSIs, where disruption of pigmentation production is with a lower percentage only for marjoram oil (92). Many scientists reported different EOs manifesting inhibition of violacein production, identified in Elletaria cardamomum, Eucalyptus radiate, Origanum vulgare, Melaleuca alternifolia, and Mentha suaveolens (24,98,99). Interestingly, among some EOs, like limonene from Citrus lemon, terpinen-4-ol, and pinene from Juniperus communis, and tea tree oil from Melaleuca alternifolia, that identified as QSIs of the purple pigment in C. violaceum, only cis-cis-p-menthenolide from Mentha suaveolens altered biofilm matrix during biofilm formation (98,99,107).
5. Conclusion
In this review, we try to put the accent and summarize the information on the natural QSIs, their functionality, and their main inhibitory role in C. violaceum’s QS system. We emphasize some critical points that show the effectiveness of such small molecules on broad biological activities, especially to mediate QS processes in Gram-negative bacteria. The new era of QSIs is a sufficient motive, that helps scientists to battle bacterial resistance, discovering new strategies related to isolating and synthesizing natural products or their analogs. In conclusion, this highlight on QSIs and their importance in bacterial combat will help us identify the variety of them as a target for developing new antimicrobials. This is yet to be the subject of future investigations.
Author Contributions
Literature search, writing, and image design – Petya D. Dimitrova and Tsvetozara Damyanova; Design of this review, draft, and critical revision - Tsvetelina Paunova-Krasteva
Funding
This work was supported by the National Science Fund at the Ministry of Education and Science, Bulgaria approved by Research Grant KP-06-H41/8; 30.11.2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank Dr. Stoyanka R. Stoitsova, for critical reading of the draft review.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
QS – Quorum sensing
QSIs – Quorum sensing inhibitors
AI – Autoinducer
AHLs – Acyl-homoserine lactones
References
- Kumar, P.; Sharma, S.; Tripathi, V.N. Quorum sensing in bacteria of rice rhizospheres from Chhattisgarh, India. Bioinformation 2023, 19, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Bouyahya, A.; Dakka, N.; Et-Touys, A.; Abrini, J.; Bakri, Y. Medicinal plant products targeting quorum sensing for combating bacterial infections. Asian Pac J Trop Med 2017, 10, 729–43. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Hu, W.; Tian, Z.; Yuan, D.; Yi, G.; Zhou, Y.; et al. Developing natural products as potential anti-biofilm agents. Chin Med 2019, 14, 11. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T.B.; Manefield, M.; Andersen, J.B.; Eberl, L.; Anthoni, U.; Christophersen, C.; et al. How Delisea pulchra furanones affect quorum sensing and swarming motility in Serratia liquefaciens MG1. Microbiology 2000, 146, 3237–44. [Google Scholar] [CrossRef] [PubMed]
- Parker, C.T.; Sperandio, V. Cell-to-cell signalling during pathogenesis. Cell Microbiol 2009, 11, 363–9. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, S.T.; Bassler, B.L. Bacterial Quorum Sensing: Its Role in Virulence and Possibilities for Its Control. Cold Spring Harb Perspect Med 2012, 2, a012427–a012427. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.C. Quorum sensing inhibitors: An overview. Biotechnol Adv 2013, 31, 224–45. [Google Scholar] [CrossRef]
- Yi, L.; Dong, X.; Grenier, D.; Wang, K.; Wang, Y. Research progress of bacterial quorum sensing receptors: Classification, structure, function and characteristics. Sci Total Environ 2021, 763, 143031. [Google Scholar] [CrossRef]
- Stoitsova, S.R.; Paunova-Krasteva, T.S.; Borisova, D.B. Modulation of Biofilm Growth by Sub-Inhibitory Amounts of Antibacterial Substances. In: Dhanasekaran, D.; Thajuddin, N., Eds.;. Microbial Biofilms - Importance and Applications. InTech; 2016. ISBN978-953-51-2436-8. [CrossRef]
- Trendafilova, A.; Ivanova, V.; Rangelov, M.; Todorova, M.; Ozek, G.; Yur, S.; et al. Caffeoylquinic Acids, Cytotoxic, Antioxidant, Acetylcholinesterase and Tyrosinase Enzyme Inhibitory Activities of Six Inula Species from Bulgaria. Chem Biodivers 2020, 17. [Google Scholar] [CrossRef]
- Stoitsova, S.; Paunova-Krasteva, T.; Dimitrova, P.D.; Damyanova, T. The concept for the antivirulence therapeutics approach as alternative to antibiotics: hope or still a fiction? Biotechnol Biotechnol Equip 2022, 36, 697–705. [Google Scholar] [CrossRef]
- Hemdan, B.A.; Mostafa, A.; Elbatanony, M.M.; El-Feky, A.M.; Paunova-Krasteva, T.; Stoitsova, S.; et al. Bioactive Azadirachta indica and Melia azedarach leaves extracts with anti-SARS-CoV-2 and antibacterial activities. PLOS ONE 2023, 18, e0282729. [Google Scholar] [CrossRef] [PubMed]
- Paunova-Krasteva, T.; Hemdan, B.A.; Dimitrova, P.D.; Damyanova, T.; El-Feky, A.M.; Elbatanony, M.M.; et al. Hybrid Chitosan/CaO-Based Nanocomposites Doped with Plant Extracts from Azadirachta indica and Melia azedarach: Evaluation of Antibacterial and Antibiofilm Activities. BioNanoScience 2023, 13, 88–102. [Google Scholar] [CrossRef]
- Nealson, K.H.; Platt, T.; Hastings, J.W. Cellular Control of the Synthesis and Activity of the Bacterial Luminescent System. J Bacteriol 1970, 104, 313–22. [Google Scholar] [CrossRef] [PubMed]
- Nealson, K.H.; Hastings, J.W. Bacterial Bioluminescence: Its Control and Ecological Significance. Microbiol rev 1979, 43. [Google Scholar] [CrossRef]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal–response systems in Gram-negative bacteria. Nat Rev Microbiol 2016, 14, 576–88. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.B.; Bassler, B.L. Quorum Sensing in Bacteria. Annu Rev Microbiol 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed]
- Abisado, R.G.; Benomar, S.; Klaus, J.R.; Dandekar, A.A.; Chandler, J.R. Bacterial Quorum Sensing and Microbial Community Interactions. mBio 2018, 9, e02331–17. [Google Scholar] [CrossRef]
- Chugani, S.; Greenberg, E.P. An evolving perspective on the Pseudomonas aeruginosa orphan quorum sensing regulator QscR. Front Cell Infect Microbiol 2014, 4. [Google Scholar] [CrossRef]
- Durán, N.; Justo, G.Z.; Durán, M.; Brocchi, M.; Cordi, L.; Tasic, L.; et al. Advances in Chromobacterium violaceum and properties of violacein-Its main secondary metabolite: A review. Biotechnol Adv 2016, 34, 1030–45. [Google Scholar] [CrossRef]
- Rajamanikandan, S.; Srinivasan, P. Exploring the selectivity of auto-inducer complex with LuxR using molecular docking, mutational studies and molecular dynamics simulations. J Mol Struct 2017, 1131, 281–93. [Google Scholar] [CrossRef]
- Vadakkan, K.; Choudhury, A.A.; Gunasekaran, R.; Hemapriya, J.; Vijayanand, S. Quorum sensing intervened bacterial signaling: Pursuit of its cognizance and repression. J Genet Eng Biotechnol 2018, 16, 239–52. [Google Scholar] [CrossRef] [PubMed]
- Pena, R.T.; Blasco, L.; Ambroa, A.; González-Pedrajo, B.; Fernández-García, L.; López, M.; et al. Relationship Between Quorum Sensing and Secretion Systems. Front Microbiol 2019, 10, 1100. [Google Scholar] [CrossRef] [PubMed]
- Saeki, E.K.; Kobayashi, R.K.T.; Nakazato, G. Quorum sensing system: Target to control the spread of bacterial infections. Microb Pathog 2020, 142, 104068. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.; Ahmad, F.; Dar, S.A.; Jawed, A.; Mandal, R.K.; Wahid, M.; et al. Developments in strategies for Quorum Sensing virulence factor inhibition to combat bacterial drug resistance. Microb Pathog 2018, 121, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Majdura, J.; Jankiewicz, U.; Gałązka, A.; Orzechowski, S. The Role of Quorum Sensing Molecules in Bacterial–Plant Interactions. Metabolites 2023, 13, 114. [Google Scholar] [CrossRef] [PubMed]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-Cell Communication in Bacteria. Annu Rev Cell Dev Biol. 2005, 21, 319–46. [Google Scholar] [CrossRef] [PubMed]
- De Keersmaecker, S.C.J.; Sonck, K.; Vanderleyden, J. Let LuxS speak up in AI-2 signaling. Trends Microbiol. 2006, 14, 114–9. [Google Scholar] [CrossRef]
- Stephens, K.; Bentley, W.E. Quorum Sensing from Two Engineers’ Perspectives. Isr J Chem 2023, e202200083. [Google Scholar] [CrossRef]
- Ha, C.; Park, S.J.; Im, S.J.; Park, S.J.; Lee, J.H. Interspecies signaling through QscR, a quorum receptor of Pseudomonas aeruginosa. Mol Cells 2012, 33, 53–9. [Google Scholar] [CrossRef]
- Stauff, D.L.; Bassler, B.L. Quorum Sensing in Chromobacterium violaceum: DNA Recognition and Gene Regulation by the CviR Receptor. J Bacteriol, 3871. [Google Scholar] [CrossRef]
- Kothari, V.; Sharma, S.; Padia, D. Recent research advances on Chromobacterium violaceum. Asian Pac J Trop Med. 2017, 10, 744–52. [Google Scholar] [CrossRef]
- Venkatramanan, M.; Sankar Ganesh, P.; Senthil, R.; Akshay, J.; Veera Ravi, A.; Langeswaran, K.; et al. Inhibition of Quorum Sensing and Biofilm Formation in Chromobacterium violaceum by Fruit Extracts of Passiflora edulis. ACS Omega 2020, 5, 25605–16. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, N.; Shin, J.Y.; Jung, K. Activity, Abundance, and Localization of Quorum Sensing Receptors in Vibrio harveyi. Front Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lu, Y.; Ye, X.; Emam, M.; Zhang, H.; Wang, H. Current advances in Vibrio harveyi quorum sensing as drug discovery targets. Eur J Med Chem. 2020, 207, 112741. [Google Scholar] [CrossRef] [PubMed]
- Watve, S.; Barrasso, K.; Jung, S.A.; Davis, K.J.; Hawver, L.A.; Khataokar, A.; et al. Parallel quorum-sensing system in Vibrio cholerae prevents signal interference inside the host. PLOS Pathog, 0831. [Google Scholar] [CrossRef]
- Brazilian National Genome Project Consortium. The complete genome sequence of Chromobacterium violaceum reveals remarkable and exploitable bacterial adaptability. Proc Natl Acad Sci. 2003, 100, 11660–5. [Google Scholar] [CrossRef] [PubMed]
- Kamaeva, A.A.; Vasilchenko, A.S.; Deryabin, D.G. Atomic Force Microscopy Reveals a Morphological Differentiation of Chromobacterium violaceum Cells Associated with Biofilm Development and Directed by N-Hexanoyl-L-Homoserine Lactone. PLoS ONE 2014, 9, e103741. [Google Scholar] [CrossRef] [PubMed]
- Kaniyarakkal, V.; Orvankundil, S.; Lalitha, S.K.; Thazhethekandi, R.; Thottathil, J. Chromobacterium violaceum Septicaemia and Urinary Tract Infection: Case Reports from a Tertiary Care Hospital in South India. Case Rep Infect Dis. 2016, 2016, 1–4. [Google Scholar] [CrossRef]
- de Oca-Mejía, M.M.; Castillo-Juárez, I.; Martínez-Vázquez, M.; Soto-Hernandez, M.; García-Contreras, R. Influence of quorum sensing in multiple phenotypes of the bacterial pathogen Chromobacterium violaceum. Pathog Dis. 2015, 73, 1–4. [Google Scholar] [CrossRef]
- Durán, M. ; Faljoni-Alario, A,; Durán, N. Chromobacterium violaceum and its important metabolites — review. Folia Microbiol (Praha). [CrossRef]
- Batista, J.H.; da Silva Neto, J.F. Chromobacterium violaceum Pathogenicity: Updates and Insights from Genome Sequencing of Novel Chromobacterium Species. Front Microbiol. 2017, 10, 2213. [Google Scholar] [CrossRef]
- Evans, K.C.; Benomar, S.; Camuy-Vélez, L.A.; Nasseri, E.B.; Wang, X.; Neuenswander, B.; et al. Quorum-sensing control of antibiotic resistance stabilizes cooperation in Chromobacterium violaceum. ISME J. 2018, 12, 1263–72. [Google Scholar] [CrossRef]
- Cheng WJ, Zhou JW, Zhang PP, Luo HZ, Tang S, Li JJ, et al. Quorum sensing inhibition and tobramycin acceleration in Chromobacterium violaceum by two natural cinnamic acid derivatives. Appl Microbiol Biotechnol. 2020 Jun;104:5025–37. [CrossRef]
- Gohil N, Bhattacharjee G, Gayke M, Narode H, Alzahrani KJ, Singh V. Enhanced production of violacein by Chromobacterium violaceum using agro-industrial waste soybean meal. J Appl Microbiol. 2022 Feb;132:1121–33. [CrossRef]
- Devescovi, G.; Kojic, M.; Covaceuszach, S.; Cámara, M.; Williams, P.; Bertani, I.; et al. Negative Regulation of Violacein Biosynthesis in Chromobacterium violaceum. Front Microbiol. 2017, 8, 349. [Google Scholar] [CrossRef]
- Shi, W.P.; Zeng, H.; Wan, C.X.; Zhou, Z.B. Amicoumacins from a desert bacterium: quorum sensing inhibitor against Chromobacterium violaceum. Nat Prod Res. 2021, 35, 5508–12. [Google Scholar] [CrossRef] [PubMed]
- Bassler, B.L.; Losick, R. Bacterially Speaking. Cell. 2006, 125, 237–46. [Google Scholar] [CrossRef] [PubMed]
- Bassler, B.L. How bacteria talk to each other: regulation of gene expression by quorum sensing. Curr Opin Microbiol. 1999, 2, 582–7. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Meng, F.; Gu, W.; Li, F.; Tao, Y.; Zhang, Z.; et al. Effects of Natural Products on Bacterial Communication and Network-Quorum Sensing. BioMed Res Int. 2020, 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T.B. ; Givskov. M. Quorum sensing inhibitors: a bargain of effects. Microbiology. [CrossRef]
- Fleitas Martínez, O.; Rigueiras, P.O. ; Pires Á da, S,; Porto, W.F.; Silva, O.N.; de la Fuente-Nunez, C.; et al. Interference With Quorum-Sensing Signal Biosynthesis as a Promising Therapeutic Strategy Against Multidrug-Resistant Pathogens. Front Cell Infect Microbiol, 8. [CrossRef]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat Rev Drug Discov. 2010, 9, 117–28. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Teplitski, M.; Robinson, J.B.; Bauer, W.D. Production of Substances by Medicago truncatula that Affect Bacterial Quorum Sensing. Mol Plant-Microbe Interactions. [CrossRef]
- Hentzer, M.; Givskov, M. Pharmacological inhibition of quorum sensing for the treatment of chronic bacterial infections. J Clin Invest. 2003, 112, 1300–7. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Andersen, B.; Thrane, U. The use of secondary metabolite profiling in chemotaxonomy of filamentous fungi. Mycol Res. 2008, 112, 231–40. [Google Scholar] [CrossRef]
- Dobretsov, S.; Teplitski, M.; Bayer, M.; Gunasekera, S.; Proksch, P.; Paul, V.J. Inhibition of marine biofouling by bacterial quorum sensing inhibitors. Biofouling. 2011, 27, 893–905. [Google Scholar] [CrossRef]
- Wahjudi, M.; Papaioannou, E.; Hendrawati, O.; van Assen, A.H.G.; van Merkerk, R.; Cool, R.H.; et al. PA0305 of Pseudomonas aeruginosa is a quorum quenching acylhomoserine lactone acylase belonging to the Ntn hydrolase superfamily. Microbiology. 2011, 157, 2042–55. [Google Scholar] [CrossRef]
- Terwagne, M.; Mirabella, A.; Lemaire, J.; Deschamps, C.; De Bolle, X.; Letesson, J.J. Quorum Sensing and Self-Quorum Quenching in the Intracellular Pathogen Brucellamelitensis. PLoS ONE. 2013, 8, e82514. [Google Scholar] [CrossRef]
- Teiber, J.F.; Horke, S.; Haines, D.C.; Chowdhary, P.K.; Xiao, J.; Kramer, G.L.; et al. Dominant Role of Paraoxonases in Inactivation of the Pseudomonas aeruginosa Quorum-Sensing Signal N -(3-Oxododecanoyl)- l -Homoserine Lactone. Infect Immun. 2008, 76, 2512–9. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, D.; Grossmann, G.; Séquin, U.; Brandl, H.; Bachofen, R. Effects of natural and chemically synthesized furanones on quorum sensing in Chromobacterium violaceum. BMC Microbiol. 2004, 4, 25. [Google Scholar] [CrossRef] [PubMed]
- Amaya, S.; Pereira, J.A.; Borkosky, S.A.; Valdez, J.C.; Bardón, A.; Arena, M.E. Inhibition of quorum sensing in Pseudomonas aeruginosa by sesquiterpene lactones. Phytomedicine. 2012, 19, 1173–7. [Google Scholar] [CrossRef] [PubMed]
- Martín-Rodríguez, A.J.; Ticona, J.C.; Jiménez, I.A.; Flores, N.; Fernández, J.J.; Bazzocchi, I.L. Flavonoids from Piper delineatum modulate quorum-sensing-regulated phenotypes in Vibrio harveyi. Phytochemistry. 2015, 117, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Majik, M.S.; Gawas, U.B.; Mandrekar, V.K. Next generation quorum sensing inhibitors: Accounts on structure activity relationship studies and biological activities. Bioorg Med Chem. 2020, 28, 115728. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.O.; Zedan, H.H.; Ibrahim, Y.M. Quorum sensing inhibitory effect of bergamot oil and aspidosperma extract against Chromobacterium violaceum and Pseudomonas aeruginosa. Arch Microbiol. 2021, 203, 4663–75. [Google Scholar] [CrossRef] [PubMed]
- Kazemian, H.; Ghafourian, S.; Heidari, H.; Amiri, P.; Yamchi, J.K.; Shavalipour, A.; et al. Antibacterial, anti-swarming and anti-biofilm formation activities of Chamaemelum nobile against Pseudomonas aeruginosa. Rev Soc Bras Med Trop. 2015, 48, 432–6. [Google Scholar] [CrossRef]
- Bacha, K.; Tariku, Y.; Gebreyesus, F.; Zerihun, S.; Mohammed, A.; Weiland-Bräuer, N.; et al. Antimicrobial and anti-Quorum Sensing activities of selected medicinal plants of Ethiopia: Implication for development of potent antimicrobial agents. BMC Microbiol. 2016, 16, 139. [Google Scholar] [CrossRef]
- Nikolova, I.; Paunova-Krasteva, T.; Petrova, Z.; Grozdanov, P.; Nikolova, N.; Tsonev, G.; et al. Bulgarian Medicinal Extracts as Natural Inhibitors with Antiviral and Antibacterial Activity. Plants. 2022, 11, 1666. [Google Scholar] [CrossRef]
- Olivero-Verbel, J.; Barreto-Maya, A.; Bertel-Sevilla, A.; Stashenko, E.E. Composition, anti-quorum sensing and antimicrobial activity of essential oils from Lippia alba. Braz J Microbiol. 2014, 45, 759–67. [Google Scholar] [CrossRef]
- Choo, J.H.; Rukayadi, Y.; Hwang, J.K. Inhibition of bacterial quorum sensing by vanilla extract. Lett Appl Microbiol. 2006, 42, 637–641. [Google Scholar] [CrossRef] [PubMed]
- Al-Hussaini, R.; Mahasneh, A. Microbial Growth and Quorum Sensing Antagonist Activities of Herbal Plants Extracts. Molecules. 2009, 14, 3425–35. [Google Scholar] [CrossRef] [PubMed]
- 72. Singh ,B.N.; Singh, B.R.; Singh, R.L.; Prakash, D.; Sarma, B.K.; Singh, H.B. Antioxidant and anti-quorum sensing activities of green pod of Acacia nilotica L. Food Chem Toxicol. [CrossRef]
- Singh, B.N.; Singh, B.R.; Singh, R.L.; Prakash, D.; Dhakarey, R.; Upadhyay, G.; et al. Oxidative DNA damage protective activity, antioxidant and anti-quorum sensing potentials of Moringa oleifera. Food Chem Toxicol. 2009, 47, 1109–16. [Google Scholar] [CrossRef]
- Grandclément, C.; Tannières, M.; Moréra, S.; Dessaux, Y.; Faure, D. Quorum quenching: role in nature and applied developments. FEMS Microbiol Rev. 2016, 40, 86–116. [Google Scholar] [CrossRef] [PubMed]
- Asfour, H. Anti-quorum sensing natural compounds. J Microsc Ultrastruct. 2018, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Swem, L.R.; Swem, D.L.; Stauff, D.L.; O’Loughlin, C.T.; Jeffrey, P.D.; et al. A Strategy for Antagonizing Quorum Sensing. Mol Cell. 2011, 42, 199–209. [Google Scholar] [CrossRef]
- Aliyu, A.B.; Koorbanally, N.A.; Moodley, B.; Singh, P.; Chenia, H.Y. Quorum sensing inhibitory potential and molecular docking studies of sesquiterpene lactones from Vernonia blumeoides. Phytochemistry. 2016, 126, 23–33. [Google Scholar] [CrossRef]
- Koh, K.H.; Tham, F.Y. Screening of traditional Chinese medicinal plants for quorum-sensing inhibitors activity. J Microbiol Immunol Infect. 2011, 44, 144–8. [Google Scholar] [CrossRef]
- Fatima, Q.; Zahin, M.; Khan, M.S.A.; Ahmad, I. Modulation of quorum sensing controlled behaviour of bacteria by growing seedling, seed and seedling extracts of leguminous plants. Indian J Microbiol. 2010, 50, 238–42. [Google Scholar] [CrossRef]
- Song, C.; Ma, H.; Zhao, Q.; Song, S.; Jia, Z. Inhibition of Quorum Sensing Activity by Ethanol Extract of Scutellaria baicalensis Georgi. J Plant Pathol Microbiol. 2012, 09. [Google Scholar] [CrossRef]
- Chong, Y.M.; Yin, W.F.; Ho, C.Y.; Mustafa, M.R.; Hadi, A.H.A.; Awang, K.; et al. Malabaricone C from Myristica cinnamomea Exhibits Anti-Quorum Sensing Activity. J Nat Prod. 2011, 74, 2261–4. [Google Scholar] [CrossRef] [PubMed]
- Musthafa, K.S.; Ravi, A.V.; Annapoorani, A.; Packiavathy, I.S.V.; Pandian, S.K. Evaluation of Anti-Quorum-Sensing Activity of Edible Plants and Fruits through Inhibition of the N-Acyl-Homoserine Lactone System in Chromobacterium violaceum and Pseudomonas aeruginosa. Chemotherapy. 2010, 56, 333–9. [Google Scholar] [CrossRef] [PubMed]
- Chenia, H. Anti-Quorum Sensing Potential of Crude Kigelia africana Fruit Extracts. Sensors. 2013, 13, 2802–17. [Google Scholar] [CrossRef] [PubMed]
- Olivero, V.; Jesús, T.; Pájaro, C.; Nerlis. P.; Stashenko, E. Antiquorum sensing activity of essential oils isolated from different species of the genus Piper. Vitae. 2011, 18, 77–82. [Google Scholar] [CrossRef]
- Zhou, L.; Zheng, H.; Tang, Y.; Yu, W.; Gong, Q. Eugenol inhibits quorum sensing at sub-inhibitory concentrations. Biotechnol Lett. 2013, 35, 631–367. [Google Scholar] [CrossRef] [PubMed]
- Musthafa, K.S.; Sahu, S.K.; Ravi, A.V.; Kathiresan, K. Anti-quorum sensing potential of the mangrove Rhizophora annamalayana. World J Microbiol Biotechnol. 2013, 29, 1851–8. [Google Scholar] [CrossRef]
- Zaki, A.A. Assessment of Anti-Quorum Sensing Activity for Some Ornamental and Medicinal Plants Native to Egypt. Sci Pharm. 2013, 81, 251–8. [Google Scholar] [CrossRef]
- Vasavi, H.S.; Arun, A.B.; Rekha, P.D. Inhibition of quorum sensing in Chromobacterium violaceum by Syzygium cumini L. and Pimenta dioica L. Asian Pac J Trop Biomed. 2013, 3, 954–9. [Google Scholar] [CrossRef]
- Ceylan, O.; Sahin, M.; Akdamar, G. Antioxidant and Anti-quorum Sensing Potential of Acer monspessulanum subsp. monspessulanum Extracts. Planta Med. 2016, 82, 1335–40. [Google Scholar] [CrossRef]
- Tamfu, A.N.; Kucukaydin, S.; Ceylan, O.; Sarac, N.; Duru, M.E. Phenolic Composition, Enzyme Inhibitory and Anti-quorum Sensing Activities of Cinnamon (Cinnamomum zeylanicum Blume) and Basil (Ocimum basilicum Linn). Chem Afr. 2021, 4, 759–67. [Google Scholar] [CrossRef]
- Moradi, F.; Hadi, N.; Bazargani, A. Evaluation of quorum-sensing inhibitory effects of extracts of three traditional medicine plants with known antibacterial properties. New Microbes New Infect. 2020, 38, 100769. [Google Scholar] [CrossRef] [PubMed]
- Alibi, S.; Ben Selma, W.; Ramos-Vivas, J.; Smach, M.A.; Touati, R.; Boukadida, J.; et al. Anti-oxidant, antibacterial, anti-biofilm, and anti-quorum sensing activities of four essential oils against multidrug-resistant bacterial clinical isolates. Curr Res Transl Med. 2020, 68, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiao, P.; Wang, Y.; Hao, Y. Mechanisms and Control Measures of Mature Biofilm Resistance to Antimicrobial Agents in the Clinical Context. ACS Omega. 2020, 5, 22684–90. [Google Scholar] [CrossRef] [PubMed]
- Qais, F.A.; Khan, M.S.; Ahmad, I. Broad-spectrum quorum sensing and biofilm inhibition by green tea against gram-negative pathogenic bacteria: Deciphering the role of phytocompounds through molecular modelling. Microb Pathog. 2019, 126, 379–92. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Asfour, H.Z.; Elshali, K.Z.; Shaaban, M.I.A.; Al-Attas, A.A.M.; Mohamed, G.A.A. Antimicrobial, antiquorum sensing, and antiproliferative activities of sesquiterpenes from Costus speciosus rhizomes. Pak J Pharm Sci. 2019, 32, 109–115. [Google Scholar]
- Rahman, M.R.T.; Lou, Z.; Yu, F.; Wang, P.; Wang, H. Anti-quorum sensing and anti-biofilm activity of Amomum tsaoko (Amommum tsao-ko Crevost et Lemarie) on foodborne pathogens. Saudi J Biol Sci. 2017, 24, 324–30. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, L.; Gao, J.; Liu, X.; Feng, Y.; Wu, Q.; et al. Tannin-Rich Fraction from Pomegranate Rind Inhibits Quorum Sensing in Chromobacterium violaceum and Biofilm Formation in Escherichia coli. Foodborne Pathog Dis. 2016, 13, 28–35. [Google Scholar] [CrossRef]
- Poli, J.P.; Guinoiseau, E.; de Rocca Serra, D.; Sutour, S.; Paoli, M.; Tomi, F.; et al. Anti-Quorum Sensing Activity of 12 Essential Oils on Chromobacterium violaceum and Specific Action of cis-cis-p-Menthenolide from Corsican Mentha suaveolens ssp. Insularis. Molecules. 2018, 23, 2125. [Google Scholar] [CrossRef]
- Noumi, E.; Merghni, A.; M Alreshidi, M.; Haddad, O.; Akmadar, G.; De Martino, L.; et al. Chromobacterium violaceum and Pseudomonas aeruginosa PAO1: Models for Evaluating Anti-Quorum Sensing Activity of Melaleuca alternifolia Essential Oil and Its Main Component Terpinen-4-ol. Molecules. 2018, 23, 2672. [Google Scholar] [CrossRef]
- Shukla, V.; Bhathena, Z. Broad Spectrum Anti-Quorum Sensing Activity of Tannin-Rich Crude Extracts of Indian Medicinal Plants. Scientifica. 2016, 2016, 1–8. [Google Scholar] [CrossRef]
- Zhang, J.; Rui, X.; Wang, L.; Guan, Y.; Sun, X.; Dong, M. Polyphenolic extract from Rosa rugosa tea inhibits bacterial quorum sensing and biofilm formation. Food Control. 2014, 42, 125–31. [Google Scholar] [CrossRef]
- Li, G.; Yan, C.; Xu, Y.; Feng, Y.; Wu, Q.; Lv, X.; et al. Punicalagin Inhibits Salmonella Virulence Factors and Has Anti-Quorum-Sensing Potential. Appl Environ Microbiol. 2014, 80, 6204–11. [Google Scholar] [CrossRef] [PubMed]
- Deryabin, D.; Tolmacheva, A. Antibacterial and Anti-Quorum Sensing Molecular Composition Derived from Quercus cortex (Oak bark) Extract. Molecules. 2015, 20, 17093–108. [Google Scholar] [CrossRef] [PubMed]
- Paliya, B.S.; Mathew, J.; Singh, B.N. Evaluation of Anti-quorum Sensing Potential of Saraca asoca (Family Caesalpiniaceae) against Chromobacterium violaceum and Pseudomonas aeruginosa PA01. J Pharm Res Int. 2021, 33, 71–82. [Google Scholar] [CrossRef]
- Ma, L.; Yao, L. Antiviral Effects of Plant-Derived Essential Oils and Their Components: An Updated Review. Molecules. 2020, 25, 2627. [Google Scholar] [CrossRef]
- Reyes-Jurado, F.; Navarro-Cruz, A.R.; Ochoa-Velasco, C.E.; Palou, E.; López-Malo, A.; Ávila-Sosa, R. Essential oils in vapor phase as alternative antimicrobials: A review. Crit Rev Food Sci Nutr. 2020, 60, 1641–50. [Google Scholar] [CrossRef]
- Li, J.; Zhao, X. Effects of quorum sensing on the biofilm formation and viable but non-culturable state. Food Res Int. 2020, 137, 109742. [Google Scholar] [CrossRef]
Figure 1.
Inhibition of violacein production in Chromobacterium violaceum by QSIs. .
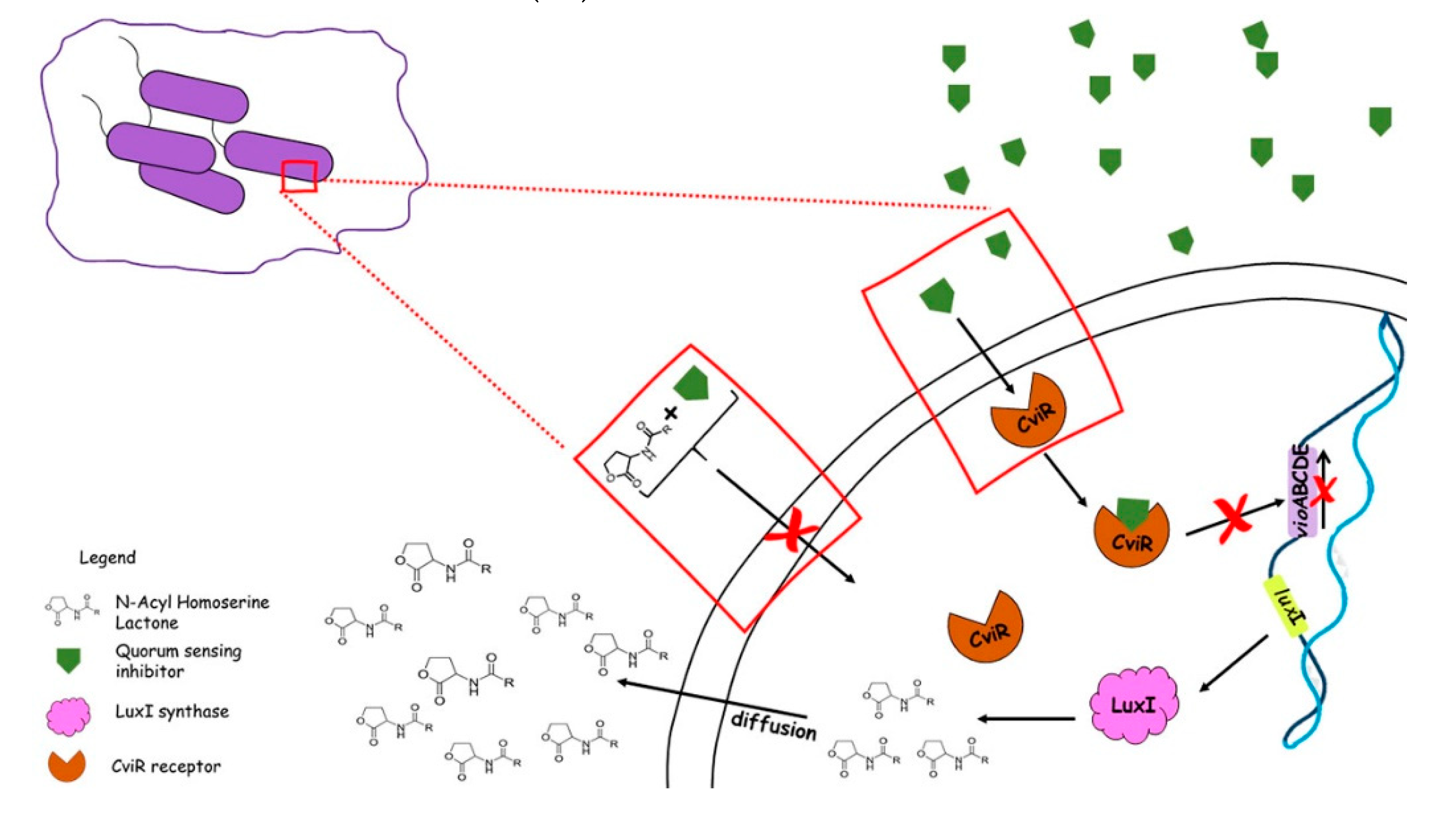

Table 1.
List of plant compounds with anti-quorum sensing activity in Chromobacterium violaceum.
| Sources of QSIs | Active component | Bacteria | Inhibition characteristics | Ref.: |
|---|---|---|---|---|
|
Prunella vulgaris (whole plant), Imperata cylindrica (underground stem), Nelumbo nucifera (leaf), Panax notoginseng (flower), Punica granatum (bark), Areca catechu (seed) |
Acetone/water extracts | C. violaceum CV026 | QS and antimicrobial activity | (78) |
|
Pisum sativum L. (seedling), Trigonella foenum graecum (seed) |
Methanol, ethanol seed extracts | C. violaceum CV026, C. violaceum ATCC 12472 | Violacein production | (79) |
| Acacia nilotica (L.) (green pod) | Phenol and polyphenol compounds |
C. violaceum ATCC 12472 |
Violacein production |
(72) |
| Scutellaria baicalensis Georgi | Ethanol extract | C. violaceum CV026 | Violacein production | (80) |
| Myristica cinnamomea King (bark) | Methanol extract, Malabaricone C | C. violaceum CV026 | Violacein |
(81) |
|
Ananas comosus, Musa paradiciaca, Manilkara zapota, Ocimum sanctum |
Fruit aqueous extracts |
C. violaceum CV026, C. violaceum ATCC 12472 |
Violacein production | (82) |
| Kigelia africana (Lam.) Benth. | Fruit ethyl acetate, dichloromethane, hexane, methanol extracts |
C. violaceum ATCC 12472, C. violaceum CV026, C. violaceum ATCC 31532 |
Antimicrobial activity, Violacein production | (83) |
|
Laurus nobilis L., Populus alba L., Populus nigra L., Lavandula angustifolia, Rosmarinus officinalis L., Sonchus oleraceus L., Tecoma capensis Thunb. Lindl., Jasminum sambac Ait. |
Ethanolic extracts |
C. violaceum | Antimicrobial activity | (71) |
|
Piper bredemeyer, Piper bogotense, Piper brachypodon (Benth.) |
Essential oils | C. violaceum CV026 | Violacein production, cell growth | (84) |
|
Syzygium aromaticum (L.) Merrill, Perry (clove) |
Extracts | C. violaceum CV026 | Violacein production | (85) |
| Rhizophora annamalayana Kathiresan (bark) | Bark extracts | C. violaceum ATCC 12472 | Violacein production | (86) |
|
Adhatoda vasica L. (leaves) Bauhinia purpurea L. (leaves) Myoporum laetum G. Forst. (leaves) Lantana camara L. (leaves) Piper longum L. (fruits) Taraxacum officinale F.H. Wigg. (aerial parts) |
Ethanol fractions |
C. violaceum ATCC 12472 |
Antimicrobial activity | (87) |
|
Syzygium cumini (L.) Skeels. Pimenta dioica (L.) Merr. |
Ethyl acetate fractions |
C. violaceum ATCC 12472, C. violaceum ATCC 31532, C. violaceum CV026 |
Violacein production | (88) |
|
Acer monspessulanum subsp. monspessulanum |
Ethanol, ethyl acetate extracts |
C. violaceum CV026, C. violaceum ATCC 12472 |
Violacein production, Antimicrobial activity | (89) |
|
Cinnamomum zeylanicum, Ocimum basilicum |
Ethanol extracts | C. violaceum CV026, C. violaceum ATCC 12472 | Anti-QS activities, Violacein production | (90) |
| Rubus rosaefolius | Phenolic extracts | C. violaceum ATCC 12472 | Cluster movement, Biofilm formation, Violacein production |
(50) |
|
Astilbe rivularis, Fragaria nubicola, Osbeckia nepalensis |
Extracts | C. violaceum MTCC 2656 | Violacein | |
|
Melicope lunuankenda (Gaertn.) T. G. Hartley |
Hexane, chloroform and methanol extracts |
C. violaceum CV026 | Violacein production | |
| Nymphaea tetragona | Water extracts | C. violaceum | Violacein production | |
| Camellia sinensis L. | Water extracts | C. violaceum ATCC 12472 | Violacein production | |
| Allium cepa Lineu | Phenolic compounds | C. violaceum | Violacein production, Swarming motility | |
| Elletaria cardamomum | Essential oils | C. violaceum | Violacein production | (24) |
| Eucalyptus radiate | ||||
| Origanum vulgare | ||||
| Rubus rosaefolius | Phenolic extracts | |||
| Syzygium aromaticum | Extracts | C. violaceum CV026 | QS inhibition assay, Violacein production | (91) |
| Dionysia revoluta Boiss. | ||||
| Eucalyptus camaldulensis Dehnh. | ||||
| Cinnamomum verum | Essential oils | C. violaceum CV026 | Violacein production | (92) |
| Origanum majorana | ||||
| Thymus vulgaris | ||||
| Eugenia caryophyllata | ||||
| Lemon | Essential oils | C. violaceum SZMC 6269 | Biofilm formation | (93) |
| Juniper | ||||
| Cuminum cyminum | Methanol extract | C. violaceum ATCC 12472 | Violacein production | |
| Green tea | Extracts | C. violaceum ATCC 12472 | Violacein production | (94) |
| Costus speciosus | Methanol extract | C. violaceum | Violacein production | (95) |
| Amomum tsaoko | Crude extract | C. violaceum ATCC 12472 | Violacein production | (96) |
| Punica granatum | Tannin-rich fraction | C. violaceum ATCC 12472 | Violacein production | (97) |
| Mentha suaveolens ssp. insularis | Essential oils | C. violaceum wild-type strain - 103350T | Violacein production, Biofilm formation | (98) |
| Melaleuca alternifolia | Essential Oils | C. violaceum ATCC 12472 | Violacein production | (99) |
|
Syzygium cumini, Embelia ribes |
Tannin-Rich extracts |
C. violaceum ATCC 12472, C. violaceum ATCC 31532, C. violaceum CV026 |
Violacein production | (100) |
|
Phyllanthus emblica, Terminalia bellirica, Terminalia chebula | ||||
| Punica granatum | Pericarp | |||
| Mangifera indica | Flowers, seed kernel | |||
|
Acacia arabica, Terminalia arjuna, Thespesia populnea, Casuarina equisetifolia |
Barks | |||
| Rosa rugosa tea | Polyphenol (RTP) extract | C. violaceum CV026 | Violacein production | (101) |
| Punica granatum L. | Punicalagin | C. violaceum ATCC 12472 | Violacein production, Growth |
(102) |
| Quercus cortex (Oak bark) | Phytochemicals | C. violaceum CV026 | Violacein production, Growth |
(103) |
| Saraca asoca barks (stem) | Extracts | C. violaceum ATCC 12472 | Violacein production, Anti-QS activities |
(104) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



