
Submitted:
12 May 2023
Posted:
15 May 2023
You are already at the latest version
Abstract
Cross-species investigations on cancer invasiveness are a new approach that identified new biomarkers potentially useful for improving tumor diagnosis and prognosis in clinical medicine and veterinary science. In this study, we combined proteomic analysis of four experimental rat malignant mesothelioma (MM) tumors with analysis of ten patient-derived cell lines to identify common features associated to mitochondrial proteome rewiring. The comparison of significant abundance changes between invasive and non-invasive rat tumors rat tumors gave a list of 433 proteins, including twenty-six proteins reported to be exclusively located in mitochondria. Next, we analyzed the differential expression of genes encoding the mitochondrial proteins of interest in five primary epithelioid and five primary sarcomatoid human MM cell lines, and the most impressive increase was observed in the expression of the long-chain acyl coenzyme A dehydrogenase (ACADL). To evaluate the role of this enzyme in the migration/invasiveness, two epithelioid and two sarcomatoid human MM cell lines derived from patients with the highest and lowest overall survival were studied. Interestingly, sarcomatoid vs epithelioid cell lines were characterized by higher migration and fatty oxidation rates, in agreement with ACADL findings. These results suggest that evaluating mitochondrial proteins in MM specimens might identify tumors with higher invasiveness.
Keywords:
malignant mesothelioma
; metabolism
; mitochondria
; long-chain specific acyl-CoA dehydrogenase
; fatty acid β-oxidation
; biomarker
1. Introduction
The role of mitochondria, at the crossroads of many studies related to cancer invasiveness, has been extensively investigated over the last fifteen years [1]. Their involvement in motility and invasion, microenvironment, plasticity, and colonization was recently reviewed [2]. Since the pioneering work of Ishikawa et al. demonstrating the role of mtDNA transfer in the acquisition of high metastatic potential [3], cancer cells were shown to acquire mitochondria from neighboring cells to acquire phenotypic characteristics, including stemness, representing gain of functions for tumors, enhancing their invasive properties [4]. The interplay between mitochondrial dynamics and extracellular matrix (ECM) remodeling was emphasized [5], and the molecular mechanisms linking dysregulated fission / fusion to tumor progression and metastasis deciphered [6]. Together with an update view of mitochondria dysfunction on tumor glycolysis [7], these important breakthroughs have a profound impact on new therapeutic strategies aiming at overcoming hypoxic and chemorefractory tumors [8]. Finally, the upregulation of mitochondrial proteins involved both in ATP production and drug resistance [9], and/or immune-resistance [10] could lead to new therapies. In parallel, all these studies could also shed light on new biomarkers for better predicting cancer chemosensitivity [11].
In this context, proteomics-based investigations provide crucial insights into the role of mitochondrial proteome rewiring [12,13,14]. The development of high-throughput proteomics techniques, combined with the use of experimental models of increasing invasiveness, has led to the identification of new proteins of interest both in rats and humans [15]. Given this methodological background, in this study we aim to focus on mitochondrial proteins, which appear to play an important role in metabolic rewiring and the invasiveness process both in experimental tumor models and tumor cells from patients.
2. Materials and Methods
2.1. Collection of rat tumor tissues for proteomic analyses
The formalin-fixed paraffin embedded (FFPE) tissue samples used in this study were collected from the same groups of Fisher F344 rats with four different experimental mesotheliomas at increasing stages of invasiveness, as previously described [16]. To generate the tumors, the experimental procedures used for in vivo manipulations at the UTE-IRS UN (Unité Thérapeutique Expérimentale de l’Institut de Recherche en Santé de l’Université de Nantes) between 2011 and 2015 followed the European Union guidelines for the care and use of laboratory animals in research protocols (approval #01257.03 from the French Ministry of Higher Education and Research (MESR)). The rats were purchased from Charles River Laboratories (L’Arbresle, 69210, France) and the experiments were approved by the ethics committee for animal experiments (CEEA) of the Pays de la Loire Region registered under the number 2011.38. The non-invasive M5-T2, mildly invasive F4-T2, moderately invasive F5-T1 and deeply invasive M5-T1 tumors were collected after intraperitoneal injection of 3 x 106 cells of the corresponding cell lines (https://technology-offers.inserm-transfert.com/offer/, recorded as RT00418, RT00419, RT00421 and RT00417, respectively) into syngeneic rats.
2.2. Proteomic analyses
For each sample analyzed, four-five 20-µm-thick sections of tumor tissue were scratched with a scalpel and collected in a 1.5-mL Eppendorf® microtube, and all the material collected was deparaffinized in three successive xylene washes, then rehydrated in 100%, 95%, 70% and 50% ethanol solutions. The pellets were vacuum-dried, and the dried tissues resuspended in 200 µL of Rapigest SF (Waters, Milford, MA, USA). Dithiothreitol (AppliChem, Darmstadt, Germany) was then added (5 mM final concentration), and the samples incubated in a thermo shaker at 95°C for one hour, and then sonicated twice (ultrasonic processor 75,185, Bioblock Scientific, Illkirch, 67400, France). Cystein residues were alkylated by adding 200 mM S-Methyl methanethiosulfonate at 37°C (10 mM final concentration). Sequencing-grade trypsin was added at a ratio ≥ 2 µg mm-3 tissue (at 37°C overnight), the reaction was stopped with formic acid (9% final concentration, incubation at 37°C for one hour), and the acid-treated samples were centrifuged at 16,000 g for 10 minutes. After removing the salts from the supernatant, the peptides were collected in a new Eppendorf® microtube using C18 STAGE tips, and their concentration finally determined using the Micro BCA™ Protein Assay Kit (Thermo Fisher Scientific, St Herblain, 44800, France). The rat spectral library, SWATH-MS analysis, peptide identification, and relative quantification were performed as previously described [16]. The statistical analysis of the SWATH data set and peak extraction output data matrix from PeakView was imported into MarkerView (v.2, AB Sciex Pte, Ltd., Framingham, MA, USA) for data normalization, and relative protein quantification. Proteins with a statistical p-value < 0.05 estimated by MarkerView were considered differentially expressed under different conditions.
2.3. Histology and immunohistochemical analyses
The FFPE blocs were cut with a Leica RM2255 microtome (Leica Biosystems, Nussloch, Germany). Areas of interest for both proteomic and histological analyses were selected based on examination of sections of all samples stained with hematoxylin phloxine saffron (HPS), scanned on a Nanozoomer 2.0 HT Hamamatsu. For immunohistochemistry, tumor sections were stained with anti-ACADL NBP2-92854 polyclonal antibody (Novus Biologicals, Centennial, CO 80112, USA).
2.4. Chemicals
FBS and culture medium were from Invitrogen Life Technologies. Plasticware for cell cultures was from Falcon (Becton Dickinson). The protein content was assessed with the BCA Kit from Sigma Chemical Co. Electrophoresis reagents were obtained from Bio-Rad Laboratories. Unless otherwise specified, all the other reagents were purchased from Sigma Chemical Co.
2.5. Cells
Ten primary human MM cell lines (5 epithelioid and 5 sarcomatoid), obtained during diagnostic thoracoscopies, were collected from the Biologic Bank of Malignant Mesothelioma, S. Antonio e Biagio e Cesare Arrigo Hospital (Alessandria, Italy), after written informed consent. The local Ethics Committees approved the study (#9/11/2011; #126/2016). Cultures were used within passage 10. Clinical and pathological characteristics of the MPM patients are reported in Table 1. Primary MPM cells were grown in HAM’s F12 and DMEM medium, supplemented with 10% v/v fetal bovine serum and 100 U/ml penicillin-100 μg/ml streptomycin.
2.6. Immunoblotting
Cells were rinsed with lysis buffer (150 mM NaCl; 1.0% Nonidet P-40; 50 mM Tris-Cl; pH 7.4), supplemented with the protease inhibitor cocktail, sonicated and centrifuged (13,000 ×g, for 10 min at 4°C). 20 μg of proteins were probed with the following antibodies: ACADL (ab152160, Abcam, Cambridge, UK); β-tubulin (sc-52-74, Santa Cruz Biotechnology Inc.), followed by peroxidase-conjugated secondary antibodies (Bio-Rad Laboratories, Hercules, CA). Blots were washed with Tris-buffered saline/Tween 0.01% v/v, developed with enhanced chemiluminescence (Bio-Rad Laboratories) and imaged using a ChemiDocTM Touch Imaging System device (Bio-Rad Laboratories).
2.7. Mitochondria isolation
To isolate mitochondria, cells were washed twice in 8 mL of PBS 1X, then 0.8 mL of mitochondria lysis buffer (50 mM TRIS, 100 mM KCl, 5 mM MgCl2, 1 mM EDTA and 1.8 mM ATP, pH 7.2) mixed with protease inhibitor cocktail set III (100 µL). PMSF (100 µL) and NaF (25 µL) were added to lyse the cells. Cells were scraped and collected in an Eppendorf® tube, then sonicated twice for 10 seconds at 40% power. Subsequently, samples were centrifuged at 2000 rpm for 1 minute at 4°C. The supernatant was collected into a new series of Eppendorf® tubes and centrifuged again at 13000 rpm for 5 minutes at 4°C. Pellets containing mitochondria were washed with 0.4 mL of mitochondria lysis buffer and centrifuged at 13000 rpm for 5 minutes at 4°C. Subsequently, the supernatant was aspirated and the pellets resuspended in 0.2 mL of mitochondria resuspension buffer (Sucrose 250 mM, K2HPO4 15 mM, MgCl2 2 mM and EDTA 0.5 mM, pH 7.2). The resuspended mitochondria were divided: one part was used to measure mitochondria protein content using a BCA kit (Sigma). The remaining part of resuspended mitochondria was divided into 50 µL aliquots and stored at -80°C until use.
2.8. ETC (Electron transport chain from complex I to complex III)
The electron transport between complexes I and III was measured in mitochondrial extracts obtained previously. In particular, 10 µL of mitochondria samples were put in a 96-well plate together with 160 µL of buffer A (5 mM KH2PO4, 5 mM of MgCl2, 5% w/v bovine serum albumin, pH 7.2), 100 µL of buffer B (50 mM KH2PO4, 5 mM MgCl2, 5% w/v serum bovine albumin, 0.05% saponin, pH 7.5) and freshly added 0.12 mM of cytochrome c-oxidized form and 0.2 mM of NaN3. After waiting 5 minutes to equilibrate the plate at room temperature, 30 µL of NADH (0.15 mM and diluted in buffer B) were added to each well. The reaction then started, and the absorbance was read at 550 nm for 6 minutes, with 1 read every 15 seconds. Considering only the linear part of the curve and calculating results in accordance with Lambert-Beer equations, the results obtained were expressed as nmoles of cytochrome C reduced/min/mg mitochondrial protein.
2.9. ATP
ATP quantities were measured following the Sigma-Aldrich protocol 213-579-1. First 50 µL of ATP assay mix (lyophilized powder containing luciferase, luciferin, MgSO4, DTT, EDTA, BSA, and tricine buffer salts, pH 7.8) were added to a vial for 3 minutes. Then, 50 µL of sample (mitochondria extract obtained as described in the previous steps) were rapidly added and the quantity of light was measured in a black 96-well plate in a microplate reader. The results were expressed as nmols of ATP/mg mitochondrial.
2.10. β-oxidation of fatty acid
Assays were performed using the fatty acid complete oxidation kit (ab222944; Abcam, Cambridge, UK) as per the manufacturer’s instructions. Cells were plated at 40,000 cells per well in a 96-well plate with 200 µl of medium per well and left overnight to equilibrate. The cells were washed twice with prewarmed FA-free measurement media, incubated with FA measurement media (150 µM FAO-Conjugate; 0.5 mM L-Carnitine) with extracellular O2 consumption reagent (ab197243; Abcam, Cambridge, UK), and then sealed with mineral oil. The fluorescence signal was read in a microplate reader (Ex/Em = 380/650 nM). The results were expressed as pmoles of O2/min.
2.11. Scratch assay
Cells were plated at 1x106 cells per well in a 6-well plate. After 24h, scratches using a 20-200µL pipette tip were made. The cell migration was calculated measuring distance (in µM) between the cells at T0 (immediately after the scratch) and T1 (24h after the scratch) and dividing it for 24h. The results were expressed as µM/h.
2.12. Real time PCR (RT-PCR)
Total RNA was extracted using VWR Life Science RiboZol™ RNA Extraction Reagent (VWR Life Science) and reverse-transcribed using the iScriptTM cDNA Synthesis Kit (Bio-Rad Laboratories). qRT-PCR was carried out using SYBR Green Supermix (Bio-Rad Laboratories). qPrimerDepot software (http://primerdepot.nci.nih.gov/) was used to obtain the following PCR primers (Supplementary Table S1). Gene Expression Quantitation software (Bio-Rad Laboratories) was used to assess relative gene expression levels.
2.13. Statistical analysis
All data in the text and figures are provided as a mean ± SEM. The results were analyzed using a one-way ANOVA and Tukey test. P < 0.05 was considered significant.
3. Results
3.1. Mitochondrial biomarkers involved in the acquisition of invasiveness in rat mesotheliomas
To identify a set of mitochondrial proteins involved in the acquisition of tumor invasiveness, we analyzed the proteomes of four experimental models of mesothelioma grown in immunocompetent F344 rats presenting increasing stages of invasiveness. For each tumor type, 1300 proteins were detected, and the comparison of abundance levels for the three invasive tumors ((1) mildly invasive F4-T2, (2) moderately invasive F5-T1, and (3) deeply invasive M5-T1)) versus (4) the noninvasive tumor M5-T2 (Figure 1) produced a list of 433 proteins satisfying the condition p < 0.05. The full list of genes encoding these proteins together with their full names are given in Supplementary Table S2.
In a second step, the subcellular extracellular locations of these proteins were recorded on https://www.proteinatlas.org (accessed on September 29, 2022), and 36 proteins exclusively or mainly located in mitochondria were identified. The list of mitochondrial proteins exhibiting significant abundance changes (increase or decrease in [1+2+3] vs 4) is shown in Table 2.
Twenty-six proteins were reported to be exclusively located in mitochondria, including 17 concerned with the most dramatic changes (13 increased and 4 decreased, with p < 0.01), and involved in 11 main biological functions. Among the proteins increasing in abundance with invasiveness, two were involved in fatty acid β-oxidation (FAO), encoded by Acadl [17] and Hsd17b10 (Table 1 and Figure 2A) [18], and one in adenine nucleotide metabolism (encoded by Ak2) [19]. This list also included two subunits of ATP synthase, one of the F0 membrane-spanning components (proton channel) (encoded by Atp5h) [20], the second being part of the connector linking the F1 catalytic core to F0 (encoded by Atp5o) [21]. Other increased proteins corresponded to two subunits of the cytochrome c oxidase (encoded by Mtco2, Cox5b, Cox6c2) [22], a chaperone regulating cellular stress responses (encoded by Trap1), two subunits of the isocitrate dehydrogenase (encoded by Idh3a and Idh3b) [23] and the malate dehydrogenase (encoded by Mdh2) [24], and two mitochondrial scaffolding/chaperone proteins (encoded by Phb and Phb2) [25]. The last two increased proteins participate in protein translation in mitochondria and contribute to mitochondrial genome stability and biogenesis (encoded by Tufm and Ssbp1, respectively) [26,27]. Of the four main proteins decreasing in abundance with invasiveness, two represented enzymes of sulfur metabolism (encoded by Suox and Mpst) [28], one was a peroxide reductase playing a role in protection against oxidative stress (encoded by Prdx3) [29], and the other modulated ion channels and receptors (encoded by S100a10) [30].
Of the enzymes involved in mitochondrial FAO and detected in proteomic analyses, the long-chain acyl coenzyme A dehydrogenase (encoded by Acadl) exhibited the most dramatic changes, a significant increase being observed for each individual invasive tumor (1 vs 4, 2 vs 4, 3 vs 4) (Figure 2A). Another enzyme in this metabolic pathway, also involved in branched-amino acid catabolism and encoded by Hsd17b10, exhibited a similar pattern of increase with invasiveness (Figure 2A). Conversely, for 2 additional enzymes in this pathway (encoded by Acads, and Hadh), invasiveness was associated with a decrease, while no significant change was observed for each individual comparison 1 vs 4, 2 vs 4, and 3 vs 4 for ACADS (Figure 2A).
The evolution in ACADL and HCD2 levels was associated with a parallel increase in two subunits of ATP synthase (Figure 2B), and three subunits of cytochrome oxidase (Figure 2C), suggesting a link with ATP production and flux within the electron transport chain. A similar increased level of two enzymes in the tricarboxylic cycle: malate dehydrogenase 2 and isocitrate dehydrogenase, also tended to demonstrate its involvement in the invasiveness process (Figure 2D).
3.2. Immunohistochemical study of ACADL distribution in rat tumors
Examination of ACADL expression by IHC in the four tumor models revealed pronounced differences with the level of invasiveness. The non-invasive tumor (M5-T2) was characterized by the absence of staining (Figure 3A). In contrast, the mildly invasive F4-T2, (Figure 3B) and moderately invasive F5-T1 (Figure 3C) tumors exhibited a weak, homogeneous distribution of ACADL expression within the tumor tissues. The most striking feature was the strong staining observed in the deeply invasive M5-T1 tumor (Figure 3D). Moreover, ACADL expression appeared heterogeneous within the tumor, some areas showing intense staining in external parts of the tumor, as shown on high magnifications views (Figure 3E).
3.3. Fatty acid β-oxidation supports cell invasiveness in human primary mesothelioma cell lines
Next, to determine whether our findings on rat mesothelioma tumors were confirmed in human malignant mesothelioma (MM), we analyzed the differential expression of genes encoding the different mitochondrial proteins of interest listed above (described in section 3.1., Table 1) in five primary sarcomatoid and five primary epithelioid mesothelioma cell lines (Supplementary Table S3). Interestingly, the most impressive increase was observed in the expression of ACADL. Other highly expressed genes in sarcomatoid mesothelioma cell lines were two ATP synthase subunits (ATP5H, ATPO), MTCO2, COX5B, COX 6C2, IDH3A, IDH3B and TIMM9 (Figure 4A). These findings confirm proteomic data obtained in rat tumors. To evaluate the role of ACADL in the migration/invasiveness of mesothelioma cells, we chose two primary epithelioid mesothelioma cell lines (UP1, UP2) and two primary sarcomatoid mesothelioma cell lines (UP 6, UP 7) deriving from patients with the highest and lowest OS, respectively (Supplementary Table S3), indicative of higher or lower invasive properties. In agreement with this finding, primary mesothelioma cells were characterized by low and high migration rates, respectively (Figure 4B, 4C). Migration of primary mesothelioma cells, evaluated with a scratch assay, was inhibited by addition of etomoxir, a drug that blocks FAO (Figure 4 B, 4C). Primary sarcomatoid mesothelioma cell lines have higher expression of ACADL mRNA (Figure 5A) and protein (Figure 5B), accompanied by higher activity of FAO in comparison with epithelioid mesothelioma cell lines (Figure 6A). Etomoxir did not change ACADL expression (Figure 5A, B) but it functionally inhibited FAO in primary sarcomatoid mesothelioma cell lines (Figure 6). A higher FAO rate (Figure 6A) fuels the electron transport chain, which worked faster (Figure 6B) and caused higher ATP production (Figure 6C). In addition to higher FAO, primary sarcomatoid mesothelioma cell lines have more active mitochondrial respiratory complexes and produce more ATP. All these metabolic processes are inhibited by etomoxir (Figure 6). Altogether, this data confirms that FAO supports ATP production through electron transport chain activity, providing energy for cell migration/invasiveness in sarcomatoid mesothelioma tumors.
4. Discussion
Cross-species investigations have provided new insights into universal mechanisms in biology, improving for example our understanding of oncogenic signatures in breast cancer development in humans and dogs [31]. Applied to proteomic analyses in cancer, common biomarkers of invasiveness have been identified in rat and human mesotheliomas [15]. Genomic analyses also pointed to reliable markers useful for the diagnosis and prognosis of hepatocellular carcinomas in both species [32]. To date, cross-species comparisons of important findings relevant to mitochondria have been very limited, focusing for example on detecting heteroplasmy [33]. In this study, we identified several biomarkers of interest that appear to play an important role in metabolic rewiring and invasiveness in both human and rat mesotheliomas.
As biosynthetic hubs, mitochondria consume a variety of different fuels to generate energy in the form of ATP for cancer cells, where fatty acid oxidation plays an important role [34]. Although most cancer researchers initially focused on glycolysis, glutaminolysis and fatty acid synthesis, the relevance of fatty acid oxidation in the metabolic reprogramming of cancer cells was extensively reviewed 10 years ago, emphasizing its role in NADPH production [35]. Linked to this statement, our results revealed two consistent findings regarding the FAO enzyme ACADL, observed both in humans and rats, and associated with the acquisition of invasive properties. The higher expression of ACADL was initially found to be positively correlated to prostate cancer progression [36].
Our data also agreed with the work of Yu et al. showing that ACADL was overexpressed both in cell lines and clinical specimens, being related to esophageal squamous cell carcinoma progression and poor diagnosis [37]. Another close FAO enzyme, which is encoded by HSD17B10, is also involved in branched amino acid catabolism and steroid metabolism. Our data are in line with previous published reports emphasizing its upregulation in invasive tumors. For example, Salas et al. have shown its predictive value in the response to chemotherapy in osteosarcomas [38]. Its overexpression also accelerated cell growth, enhanced cell respiration, and increased cellular resistance to cell death in pheochromocytoma [39]. Finally, and even more interestingly, Condon et al. found HSD17B10 was one of the six genes impacting the mTORC1 pathway [40], which is dysregulated and activated in cancer cells to drive survival, neovascularization, and invasion [41].
Interestingly, the increased β-oxidation rate, electron flux and ATP production observed in human sarcomatoid mesothelioma cell lines were all consistent with the increased expression of ATP synthase subunits, cytochrome c oxidase subunits, abundance changes in these proteins in rat tumors, and with our observations above on the long-chain acyl coenzyme A dehydrogenase. Fiorillo et al. have highlighted the fact that ATP-high cancer cells were phenotypically the most aggressive, with enhanced stem-like properties, multi-drug resistance potential and an increased capacity for cell migration, invasion, and metastasis [42]. Wang et al. also pointed out that its high expression was linked to poor prognosis in glioblastoma, clear cell renal cell carcinoma, and ovarian, prostate, and breast cancers [43]. Moreover, an additional role of ATP synthase in the formation of the permeability transition pore (PTP) was also recently reported as representing a mechanism controlling tumor cell death [44]. In this process, our findings also tend to confirm the important role of the subunit d of ATP synthase (encoded by (Atp5h / ATP5H), linked to the work by Chang et al. who reported the involvement of the overexpression of this subunit in venous invasion, distant metastasis of colon cancer, and finally poor survival [45].
Within the enzymes of mitochondrial metabolism involved in cancer progression, besides isocitrate dehydrogenase and malate dehydrogenase, subunits of the cytochrome c oxidase (complex IV of the respiratory chain), such as COX5B, were also reported [46]. Our results agreed with previous published literature on the impact of its high expression on tumor invasiveness and poor prognosis in patients with breast cancer [47]. More recently, further insights have confirmed its tremendous role as a growth-promoting gene both in hepatoma [48] and colorectal cancer [49]. Interestingly, the combined upregulation of COX5B and ATP5H was also reported by Yusenko et al. in renal oncocytomas [20]. Another subunit of the cytochrome c oxidase, COX6C, also upregulated in relation to invasiveness in our study, appeared to be differentially expressed in various cancers [50]. An important finding by Jang et al. showed it was detected in extracellular vesicles (EV) in the plasma of metastatic melanoma, ovarian and breast cancer patients, suggesting the classic EV production and mitochondrial pathways were interconnected [51]. In this latter study, an additional crucial observation was the presence of another inner mitochondrial membrane protein in these EVs [51], encoded by MTCO2. These breakthroughs are consistent with both the increased abundance of SODM and expression of this gene that we found in the most invasive rat tumors as well as in human mesothelioma cell lines. Linked to the tremendous increase in ACADL, the greater abundance and expression of the two subunits of isocitrate dehydrogenase tended to confirm previous observations regarding the central role of the TCA cycle in metabolic reprogramming and tumor invasiveness. Laurenti and Tennant have previously reviewed the impact of its dysregulation in cancers in association with hypoxia and increased intracellular levels of ROS [52]. Moreover, as shown by Zeng et al., the aberrant expression of IDH3A, which represented an upstream activator of HIF-1, promoted tumor growth and angiogenesis in various cancer types [53].
Besides the dramatic changes observed in ACADL associated with tumor invasiveness, we also identified another protein involved in the mitochondrial translation machinery, encoded by TUFM. This observation, which is consistent with higher abundance and expression of proteins involved in mtDNA maintenance, can be related to several interesting reports. For example, Cruz et al. found this protein in a list of five candidate biomarkers of drug-resistant ovarian cancer [54]. Interestingly, the mitochondrial translation pathway is required for increased electron transport chain activity [55], and its inhibition played a part in sensitizing renal cell carcinoma to chemotherapy [56]. Chatla et al. demonstrated that TUFM was required for increased mitochondrial biosynthesis [57]. Moreover, this latter work suggests the existence of a link with another elevated mitochondrial protein found in our study, encoded by ALDH7A1. ALDH7A1 is an enzyme which mechanistically appeared to provide cells with protection against various forms of stress through multiple pathways [58]. It is involved in stem cell pathways [59,60], and the link between its high expression and tumor invasiveness has been clearly established through the works of van den Hoogen et al. [61] and Giacalone et al. [62], in prostate cancer and lung cancer, respectively. Interestingly, in good agreement with our findings, Lee et al. also demonstrated its relationship with lipid catabolism as an energy source in pancreatic cancer cells [63]. ALDH7A1 was first known as antiquitin, and the study of its subcellular localization revealed its presence in cytosol in addition to mitochondria [64]. Finally, an intriguing feature of this enzyme, which resonated with this latter observation, was the recent work published by Babbi et al., revealing the central role played by this protein, also present in the nucleus, interacting with 23 other proteins in IntAct, and 62 in BioGRID, while ALDH7A1 represented one of the most frequent genes in KEGG metabolic pathways [65].
5. Conclusions
In conclusion, starting from a proteomic approach and following on with ad hoc biological validation, we picked up significant differences between non-invasive and invasive mesotheliomas developed in both rats and patient-derived cells, in terms of the expression of mitochondrial proteins. This suggests that mitochondrial activity plays an important role in cancer. In particular, ACADL and subunits of ATP synthase are highly expressed in invasive rat mesotheliomas, as well as in more aggressive human sarcomatoid mesothelioma cells, which have more active FAO, electron chain transport and ATP synthesis, supporting their growth and invasiveness. Evaluating mitochondrial proteins in MM specimens might identify tumors with higher invasiveness and new potential targets that can be explored to improve the treatment of this disease.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Supplementary Table S1; Supplementary Table S2; Supplementary Table S3
Author Contributions
Conceptualization, D.L.P., J.K. and C.R.; methodology, validation, G.O., L.R.; Preparation of samples for proteomics, D.L.P., A.B., C.H.; Preparation of samples and immunohistochemistry, S.B.; Software, validation, C.G; formal analysis, G.O., L.R.; data curation, D.L.P., J.K.; writing—original draft preparation, D.L.P., J.K.; writing—review and editing, D.L.P., J.K. and C.R.; supervision, D.L.P., J.K., C.R., C.G.; funding acquisition D.L.P., J.K. and C.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Italian Association of Cancer Research (IG21408 to CR), Fondazione Cassa di Risparmio di Torino (ID 2020.1648 to JK, ID 2021.05556 to CR). This research was conducted with the support of the “Mobilité internationale pour la recherche” (Mir program of the University of Angers) between the Universities of Angers (France) and Turin (Italy) in 2020-2021 and 2021-2022.
Institutional Review Board Statement
Experiments on F344 rats were conducted at the UTE-IRS UN facility (Nantes Université) following the European Union guidelines for the care and use of laboratory rodents in research protocols. The research was approved by the Ethics Committee for Animal Experiments (CEEA) of the Pays de la Loire Region (2011.38) and #0125703 of the French Ministry of Higher Education and Research (MESR).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Mr Costanzo Costamagna for the technical support. We are grateful to all the staff at the Animal Facility (UTE-IRS UN) at Nantes University for the excellent care of the rats during the experiments. We also acknowledge the IBISA MicroPICell facility (Biogenouest), a member of the national infrastructure France-Bioimaging supported by the French national research agency (ANR-10-INBS-04).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Faubert, B.; Solmonson, A.; DeBerardinis, R.J. Metabolic reprogramming and cancer progression. Science 2020 368, eaaw5473. https://doi-org.proxy.insermbiblio.inist.fr/10.1126/science.aaw5473. [CrossRef]
- Scheid, A.D.; Beadnell, T.C.; Welch, D.R. Roles of mitochondria in the hallmarks of metastasis. Br. J. Cancer 2021 124, 124-135. https://doi-org.proxy.insermbiblio.inist.fr/10.1038/s41416-020-01125-8. [CrossRef]
- Ishikawa, K.; Takenaga, K.; Akimoto, M.; Koshikawa, N.; Yamaguchi, A.; Imanishi, H.; Nakada, K.; Honma, Y.; Hayashi, J.-I. ROS-generating mitochondrial DNA mutations can regulate tumor cell metastasis. Science 2008 320, 661-664. https://doi-org.proxy.insermbiblio.inist.fr/10.1126/science.1156906. [CrossRef]
- Zampieri, L.X.; Silva-Almeida, C.; Rondeau, J.D.; Sonveaux, P. Mitochondrial transfer in cancer: a comprehensive review. Int. J. Mol. Sci. 2021 22, 3245. https://doi-org.proxy.insermbiblio.inist.fr/10.3390/ijms22063245. [CrossRef]
- Yanes, B.; Rainero, E. The interplay between cell-extracellular matrix interaction and mitochondria dynamics in cancer. Cancers 2022 14, 1433. https://doi-org.proxy.insermbiblio.inist.fr/10.3390/cancers14061433. [CrossRef]
- Boulton, D.P.; Caino, M.C. Mitochondrial fission and fusion in tumor progression to metastasis. Front. Cell Dev. Biol. 2022 10, 849962. https://doi-org.proxy.insermbiblio.inist.fr/10.3389/fcell.2022.849962. [CrossRef]
- Bononi, G.; Masoni, S.; Di Bussolo, V.; Tuccinardi, T.; Granchi, C.; Minutolo, F. Historical perspective of tumor glycolysis: a century with Otto Warburg. Semin. Cancer Biol. 2022 86 Pt 2, 325-333. https://doi-org.proxy.insermbiblio.inist.fr/10.1016/j.semcancer.2022.07.003. [CrossRef]
- Akman, M.; Belisario, D.C.; Salaroglio, I.C.; Kopecka, J.; Donadelli, M.; De Smaele, E.; Riganti, C. Hypoxia, endoplasmic reticulum stress and chemoresistance: dangerous liaisons. J. Exp. Clin. Cancer Res. 2021 40, 28. https://doi-org.proxy.insermbiblio.inist.fr/10.1186/s13046-020-01824-3. [CrossRef]
- Fiorillo, M.; Ózsvári, B.; Sotgia, F.; Lisanti, M.P. High ATP production fuels cancer drug resistance and metastasis: implications for mitochondrial ATP depletion therapy. Front. Oncol. 2021 11, 740720. https://doi-org.proxy.insermbiblio.inist.fr/10.3389/fonc.2021.740720. [CrossRef]
- Kopecka, J.; Gazzano, E.; Castella, B.; Salaroglio, I.C.; Mungo, E.; Massaia, M.; Riganti, C. Mitochondrial metabolism: inducer or therapeutic target in tumor immune-resistance? Semin. Cell Dev. Biol. 2020 98, 80-89. https://doi-org.proxy.insermbiblio.inist.fr/10.1016/j.semcdb.2019.05.008. [CrossRef]
- Xie, L.; Zhou, T.; Xie, Y.; Bode, A.M.; Cao, Y. Mitochondria-shaping proteins and chemotherapy. Front. Oncol. 2021 11, 769036. https://doi-org.proxy.insermbiblio.inist.fr/10.3389/fonc.2021.769036. [CrossRef]
- Concolino, A.; Olivo, E.; Tammè, L.; Fiumara, C.; De Angelis, M.T.; Quaresima, B.; Agosti, V.; Costanzo, F.S.; Cuda, G.; Scumaci, D. Proteomics analysis to assess the role of mitochondria in BRCA1-mediated breast tumorigenesis. Proteomes 2018 6, 16. https://doi-org.proxy.insermbiblio.inist.fr/10.3390/proteomes6020016. [CrossRef]
- Arif, T.; Stern, O.; Pittala, S.; Chalifa-Caspi, V.; Shoshan-Barmatz, V. Rewiring of cancer cell metabolism by mitochondrial VDAC1 depletion results in time-dependent tumor reprogramming: glioblastoma as a proof of concept. Cells 2019 8, 1330. https://doi-org.proxy.insermbiblio.inist.fr/10.3390/cells8111330. [CrossRef]
- MacVicar, T.; Ohba, Y.; Nolte, H.; Mayer, F.C.; Tatsuta, T.; Sprenger, H.-G.; Lindner, B.; Zhao, Y.; Li, J.; Bruns, C.; et al. Lipid signalling drives proteolytic rewiring of mitochondrial by YME1L. Nature 2019 575, 361-.https://doi-org.proxy.insermbiblio.inist.fr/10.3389/fonc.2021.769036. [CrossRef]
- Nader, J.S.; Boissard, A.; Henry, C.; Valo, I.; Verrièle, V.; Grégoire, M.; Coqueret, O.; Guette, C.; Pouliquen, D.L. Cross-species proteomics identifies CAPG and SBP1 as crucial invasiveness biomarkers in rat and human malignant mesothelioma. Cancers 2020 12, 2430. https://doi-org.proxy.insermbiblio.inist.fr/10.3390/cancers12092430. [CrossRef]
- Nader, J.S.; Abadie, J.; Deshayes, S.; Boissard, A.; Blandin, S.; Blanquart, C.; Boisgerault, N.; Coqueret, O.; Guette, C.; Grégoire, M.; et al. Characterization of increasing stages of invasiveness identifies stromal/cancer cell crosstalk in rat models of mesothelioma. Oncotarget 2018 9, 16311-16329. https://doi-org.proxy.insermbiblio.inist.fr/10.18632/oncotarget.24632. [CrossRef]
- Yu, D.-L.; Li, H.-W.; Wang, Y.; Li, C.-Q.; You, D.; Jiang, L.; Song, Y.-P.; Li, X.-H. Acyl-CoA dehydrogenase long chain expression is associated with esophageal squamous cell carcinoma progression and poor prognosis. Onco Targets Ther. 2018 11, 7643-7653. https://doi-org.proxy.insermbiblio.inist.fr/10.2147/ott.s171963. [CrossRef]
- Salas, S.; Jézéquel, P.; Campion, L.; Deville, J.-L., Chibon, F.; Bartoli, C.; Gentet, J.-C.; Charbonnel, C.; Gouraud, W.; Voutsinos-Porche, B.; et al. Molecular characterization of the response to chemotherapy in conventional osteosarcomas: predictive value of HSD17B10 and IFITM2. Int. J. Cancer 2009 125, 851-860. https://doi-org.proxy.insermbiblio.inist.fr/10.1002/ijc.24457. [CrossRef]
- Klepinin, A.; Zhang, S.; Klepinina, L.; Rebane-Klemm, E.; Terzic, A.; Kaambre, T.; Dzeja, P. Adenylate kinase and metabolic signaling in cancer cells. Front. Oncol. 2020 10, 660. https://doi-org.proxy.insermbiblio.inist.fr/10.3389/fonc.2020.00660. [CrossRef]
- Yusenko, M.V.; Ruppert, T.; Kovacs, G. Analysis of differentially expressed mitochondrial proteins in chromophobe renal cell carcinomas and renal oncocytomas by 2-D gel electrophoresis. Int. J. Biol. Sci. 2010 6, 213-224. https://doi-org.proxy.insermbiblio.inist.fr/10.7150/ijbs.6.213. [CrossRef]
- Wiebringhaus, R.; Pecoraro, M.; Neubauer, H.A.; Trachtova, K.; Trimmel, B.; Wieselberg, M.; Pencik, J.; Egger, G.; Krall, C.; Moriggl, R.; et al. Proteomic analysis identifies NDUFS1 and ATP5O as novel markers for survival outcome in prostate cancer. Cancers 2021 13, 6036. https://doi-org.proxy.insermbiblio.inist.fr/10.3390/cancers13236036. [CrossRef]
- Lamb, R.; Ozsvari, B.; Bonuccelli, G.; Smith, D.L.; Pestell, R.G.; Martinez-Outschoorn, U.E.; Clarke, R.B.; Sotgia, F.; Lisanti, M.P. Dissecting tumor metabolic heterogeneity: telomerase and large cell size metabolically define a sub-population of stem-like, mitochondrial-rich, cancer cells. Oncotarget 2015 6, 21892-21905. https://doi-org.proxy.insermbiblio.inist.fr/10.18632/oncotarget.5260. [CrossRef]
- Zeng, L.; Morinibu, A.; Kobayashi, M.; Zhu, Y.; Wang, X.; Goto, Y.; Yeom, C.J.; Zhao, T.; Hirota, K.; Shinomiya, K.; et al. Aberrant IDH3 expression promotes malignant tumor growth by inducing HIF-1-mediated metabolic reprogramming and angiogenesis. Oncogene 2015 34, 4758-4766. https://doi-org.proxy.insermbiblio.inist.fr/10.1038/onc.2014.411. [CrossRef]
- Liu, Q.; Harvey, C.T.; Geng, H.; Xue, C.; Chen, V.; Beer, T.M.; Qian, D.Z. Malate dehydrogenase 2 confers docetaxel resistance via regulations of JNK signaling and oxidative metabolism. Prostate 2013 73, 1028-1037. https://doi-org.proxy.insermbiblio.inist.fr/10.1002/pros.22650. [CrossRef]
- Bavelloni, A.; Piazzi, M.; Raffini, M.; Faenza, I.; Blalock, W.L. Prohibitin 2: at a communications crossroads. IUBMB Life 2015 67, 239-54. https://doi-org.proxy.insermbiblio.inist.fr/10.1002/iub.1366. [CrossRef]
- Wang, B.; Ao, J.; Yu, D.; Rao, T.; Ruan, Y.; Yao, X. Inhibition of mitochondrial translation effectively sensitizes renal cell carcinoma to chemotherapy. Biochem. Biophys. Res. Commun. 2017 490, 767-773. https://doi-org.proxy.insermbiblio.inist.fr/10.1016/j.bbrc.2017.06.115. [CrossRef]
- Xu, S.; Wu, Y.; Chen, Q.; Cao, J.; Hu, K.; Tang, J.; Sang, Y.; Lai, F.; Wang, L.; Zhang, R.; et al. hSSB1 regulates both the stability and the transcriptional activity of p53. Cell Res. 2013 23, 423-435. https://doi-org.proxy.insermbiblio.inist.fr/10.1038/cr.2012.162. [CrossRef]
- Murphy, B.; Bhattacharya, R.; Mukherjee, P. Hydrogen sulfide signaling in mitochondria and disease. FASEB J. 2019 33, 13098-13125. https://doi-org.proxy.insermbiblio.inist.fr/10.1096/fj.201901304R. [CrossRef]
- Ismail, T.; Kim, Y.; Lee, H.; Lee, D.-S.; Lee, H.-S. Interplay between mitochondrial peroxiredoxins and ROS in cancer development and progression. Int. J. Mol. Sci. 2019 20, 4407. https://doi-org.proxy.insermbiblio.inist.fr/10.3390/ijms20184407. [CrossRef]
- Seo, J.-S.; Svenningsson, P. Modulation of ion channels and receptors by p11 (S100A10). Trends Pharmacol. Sci. 2020 41, 487-497. https://doi-org.proxy.insermbiblio.inist.fr/10.1016/j.tips.2020.04.004. [CrossRef]
- Kim, T.-M.; Yang, S.; Seung, B.-J.; Lee, S.; Kim, D.; Ha, Y.-J.; Seo, M.-k.; Kim, K.-K.; Kim, H.S.; Cheong, J.-H.; et al. Cross-species oncogenic signatures of breast cancer in canine mammary tumors. Nat. Commun. 2020 11, 3616. https://doi-org.proxy.insermbiblio.inist.fr/10.1038/s41467-020-17458-0. [CrossRef]
- Al-Harazi, O.; Kaya, I.H.; Al-Eid, M.; Alfantoukh, L.; Al Zahrani, A.S.; Al Sebayel, M.; Kaya, N.; Colak, D. Identification of gene signature as diagnostic and prognostic blood biomarker for early hepatocellular carcinoma using integrated cross-species transcriptomic and network analyses. Front. Genet. 2021 12, 710049. https://doi-org.proxy.insermbiblio.inist.fr/10.3389/fgene.2021.710049. [CrossRef]
- Rensch, T.; Villar, D.; Horvath, J.; Odom, D.T.; Flicek, P. Mitochondrial heteroplasmy in vertebrates using ChIP-sequencing data. Genome Biol. 2016 17, 139. https://doi-org.proxy.insermbiblio.inist.fr/10.1186/s13059-016-0996-y. [CrossRef]
- Spinelli, J.B.; Haigis, M.C. The multifaceted contributions of mitochondria to cellular metabolism. Nat. Cell Biol. 2018 20, 745-754. https://doi-org.proxy.insermbiblio.inist.fr/10.1038/s41556-018-0124-1. [CrossRef]
- Carracedo, A.; Cantley, L.C.; Pandolfi, P.P. Cancer metabolism: fatty acid oxidation in the limelight. Nat. Rev. Cancer 2013 13, 227-232. https://doi-org.proxy.insermbiblio.inist.fr/10.1038/nrc3483. [CrossRef]
- Xie, B.-X.; Zhang, H. ; Wang, J.; Pang, B.; Wu, R.-Q.; Qian, X.-L.; Yu, L.; Li, S.-H.; Shi, Q.-G.; Huang, C.-F.; et al. Analysis of differentially expressed genes in LNCaP prostate cancer progression model. J. Androl. 2011 32, 170-182. https://doi-org.proxy.insermbiblio.inist.fr/10.2164/jandrol.109.008748. [CrossRef]
- Yu, D.-L.; Li, H.-W.; Wang, Y.; Li, C.-Q.; You, D.; Jiang, L.; Song, Y.-P.; Li, X.-H. Acyl-CoA dehydrogenase long chain expression is associated with esophageal squamous cell carcinoma progression and poor prognosis. Oncotargets Ther. 2018 11, 7643-7653. https://doi-org.proxy.insermbiblio.inist.fr/10.2147/OTT.S171963. [CrossRef]
- Salas, S.; Jézéquel, P.; Campion, L.; Deville, J.-L.; Chibon, F.; Bartoli, C.; Gentet, J.-C.; Charbonnel, C.; Gouraud, W.; Voutsinos-Porche, B.; et al. Molecular characterization of the response to chemotherapy in conventional osteosarcomas: predictive value of HSD17B10 and IFITM2. Int. J. Cancer 2009 125, 851-860. https://doi-org.proxy.insermbiblio.inist.fr/10.1002/ijc.24457. [CrossRef]
- Carlson, E.A.; Marquez, R.T.; Du, F.; Wang, Y.; Xu, L.; Yan, S.S. Overexpression of 17b-hydroxysteroid dehydrogenase type 10 increases pheochromocytoma cell growth and resistance to cell death. BMC Cancer 2015 15, 166. https://doi-org.proxy.insermbiblio.inist.fr/10.1186/s12885-015-1173-5. [CrossRef]
- Condon, K.J.; Orozco, J.M.; Adelman, C.H.; Spinelli, J.B.; van der Helm, P.W.; Roberts, J.M.; Kunchok, T.; Sabatini, D.M. Genome-wide CRISPR screens reveal multitiered mechanisms through which mTORC1 senses mitochondrial dysfunction. Proc. Natl. Acad. Sci. USA 2021 118, e2022120118. https://doi-org.proxy.insermbiblio.inist.fr/10.1073/pnas.2022120118. [CrossRef]
- Braun, C.; Weichhart, T. mTOR-dependent immunometabolism as Achilles’ heel of anticancer therapy. Eur. J. Immunol. 2021 51, 3161-3175. https://doi-org.proxy.insermbiblio.inist.fr/10.1002/eji.202149270. [CrossRef]
- Fiorillo, M.; Ózsvári, B.; Sotgia, F.; Lisanti, M.P. High ATP production fuels cancer drug resistance and metastasis: implications for mitochondrial ATP depletion therapy. Front. Oncol. 2021 11, 740720. https://doi-org.proxy.insermbiblio.inist.fr/10.3389/fonc.2021.740720. [CrossRef]
- Wang, T.; Ma, F.; Qian, H.-l. Defueling the cancer: ATP synthase as an emerging target in cancer therapy. Mol Ther. Oncolytics 2021 23, 82-95. https://doi-org.proxy.insermbiblio.inist.fr/10.1016/j.omto.2021.08.015. [CrossRef]
- Galber, C.; Acosta, M. J.; Minervivi, G.; Giorgio, V. The role of mitochondrial ATP synthase in cancer. Biol. Chem. 2020 401, 1199-1214. https://doi-org.proxy.insermbiblio.inist.fr/10.1515/hsz-2020-0157. [CrossRef]
- Chang, H.J.; Lee, M.R.; Hong, S.-H.; Yoo, B.C.; Shin, Y.-K.; Jeong, J.Y.; Lim, S.-B.; Choi, H.S.; Park, J.-G. Identification of mitochondrial F0F1-ATP synthase involved in liver metastasis of colorectal cancer. Cancer Sci. 2007 98, 1184-1191. https://doi-org.proxy.insermbiblio.inist.fr/10.1111/j.1349-7006.2007.00527. [CrossRef]
- Gaude, E.; Frezza, C. Defects in mitochondrial metabolism and cancer. Cancer Metab. 2014 2, 10. https://doi-org.proxy.insermbiblio.inist.fr/10.1186/2049-3002-2-10. [CrossRef]
- Gao, S.-P.; Sun, H.-F.; Fu, W.-Y.; Li, L.-D.; Zhao, Y.; Chen, M.-T.; Jin, W. High expression of COX5B is associated with poor prognosis in breast cancer. Future Oncol. 2017 13, 1711-1719. https://doi-org.proxy.insermbiblio.inist.fr/10.2217/fon-2017-0058. [CrossRef]
- Chu, Y.-D.; Lin, W.-R.; Lin, Y.-H.; Kuo, W.-H.; Tseng, C.-J.; Lim, S.-N.; Huang, Y.-L.; Huang, S.-C.; Wu, T.-J.; Lin, K.-H.; et al. COX5B-mediated bioenergetic alteration regulates tumor growth and migration by modulating AMPK-UHMK1-ERK cascade in hepatoma. Cancers 2020 12, 1646. https://doi-org.proxy.insermbiblio.inist.fr/10.3390/cancers12061646. [CrossRef]
- Chu, Y.-D.; Lim, S.-N.; Yeh, C.-T.; Lin, W.-R. COX5B-mediated bioenergetic alterations modulate cell growth and anticancer drug susceptibility by orchestrating claudin-2 expression in colorectal cancers. Biomedicines 2022 10, 60. https://doi-org.proxy.insermbiblio.inist.fr/10.3390/biomedicines10010060. [CrossRef]
- Tian, B.-X.; Sun, W.; Wang, S.-H.; Liu, P.-J.; Wang, Y.-C. Differential expression and clinical significance of COX6C in human diseases. Am. J. Transl. Res. 2021 13, 1-10. PMID: 33527004.
- Jang, S.C.; Crescitelli, R.; Cvjetkovic, A.; Belgrano, V.; Bagge, R.O.; Sundfeldt, K.; Ochiya, T.; Kalluri, R.; Lötvall, J. Mitochondrial protein enriched extracellular vesicles discovered in human melanoma tissues can be detected in patient plasma. J. Extracell. Vesicles. 2019 8, 1635420. https://doi-org.proxy.insermbiblio.inist.fr/10.1080/20013078.2019.1635420. [CrossRef]
- Laurenti, G.; Tennant, D.A. Isocitrate dehydrogenase (IDH), succinate dehydrogenase (SDH), fumarate hydratase (FH): three players for one phenotype in cancer? Biochem. Soc. Trans. 2016 44, 1111-1116. https://doi-org.proxy.insermbiblio.inist.fr/10.1042/BST20160099. [CrossRef]
- Zeng, L.; Morinibu, A.; Kobayashi, M.; Zhu, Y.; Wang, X.; Goto, Y.; Yeom, C.J.; Zhao, T.; Hirota, K.; Shinomiya, K.; et al. Aberrant IDH3 expression promotes malignant tumor growth by inducing HIF-1-mediated metabolic reprogramming and angiogenesis. Oncogene 2015 34, 4758-4766. https://doi-org.proxy.insermbiblio.inist.fr/10.1038/onc.2014.411. [CrossRef]
- Cruz, I.N.; Coley, H.M.; Kramer, H.B.; Madhuri, T.K.; Safuwan, N.A.M.; Angelino, A.R.; Yang, M. Proteomics analysis of ovarian cancer cell lines and tissues reveals drug resistance-associated proteins. Cancer Genom. Proteom. 2017 14, 35-52. https://doi-org.proxy.insermbiblio.inist.fr/10.21873/cgp.20017. [CrossRef]
- Norberg, E.; Lako, A.; Chen, P.-H.; Stanley, I.A.; Zhou, F.; Ficarro, S.B.; Chapuy, B.; Chen, L.; Rodig, S.; Shin, D.; et al. Differential contribution of the mitochondrial translation pathway to the survival of diffuse large B-cell lymphoma subsets. Cell Death Differ. 2017 24, 251-262. https://doi-org.proxy.insermbiblio.inist.fr/10.1038/cdd.2016.116. [CrossRef]
- Wang, B.; Ao, J.; Yu, D.; Rao, T.; Ruan, Y.; Yao, X. Inhibition of mitochondrial translation effectively sensitizes renal cell carcinoma to chemotherapy. Biochem. Biophys. Res. Commun. 2017 490, 767-773. https://doi-org.proxy.insermbiblio.inist.fr/10.1016/j.bbrc.2017.06.115. [CrossRef]
- Chatla, S.; Du, W.; Wilson, A.F.; Meetei, A.R.; Pang, Q. Fancd2-deficient hematopoietic stem and progenitor cells depend on augmented mitochondrial translation for survival and proliferation. Stem Cell Res. 2019 40, 101550. https://doi-org.proxy.insermbiblio.inist.fr/10.1016/j.scr.2019.101550. [CrossRef]
- Brocker, C.; Cantore, M.; Failli, P.; Vasiliou, V. Aldehyde dehydrogenase 7A1 (ALDH7A1) attenuates reactive aldehyde and oxidative stress induced cytotoxicity. Chem. Biol. Interact 2011 191, 269-277. https://doi-org.proxy.insermbiblio.inist.fr/10.1016/j.cbi.2011.02.016. [CrossRef]
- Prabhu, V.V.; Lulla, A.R.; Madhukar, N.S.; Ralff, M.D.; Zhao, D.; Kline, C.L.B.; Van den Heuvel, A.P.; Lev, A.; Garnett, M.J.; McDermott, U.; et al. Cancer stem cell-related gene expression as a potential biomarker of response for first-in-class imipridone ONC201 in solid tumors. PloS ONE 2017 12, e0180541. https://doi-org.proxy.insermbiblio.inist.fr/10.1371/journal.pone.0180541. [CrossRef]
- Bizzaro, V.; Belvedere, R.; Milone, M.R.; Pucci, B.; Lombardi, R.; Bruzzese, F.; Popolo, A.; Parente, L.; Budillon, A.; Petrella, A. Annexin A1 is involved in the acquisition and maintenance of a stem cell-like/aggressive phenotype in prostate cancer cells with acquired resistance to zoledronic acid. Oncotarget 2015 6, 25074-25092. https://doi-org.proxy.insermbiblio.inist.fr/. [CrossRef]
- Van den Hoogen, C.; van der Horst, G.; Cheung, H.; Buijs, J.T.; Pelger, R.C.M.; van der Pluijm, G. The aldehyde dehydrogenase enzyme 7A1 is functionally involved in prostate cancer bone metastasis. Clin. Exp. Metastasis 2011 28, 615-625. https://doi-org.proxy.insermbiblio.inist.fr/10.1007/s10585-011-9395-7. [CrossRef]
- Giacalone, N.J.; Den, R.B.; Eisenberg, R.; Chen, H.; Olson, S.J.; Massion, P.P.; Carbone, D.P.; Lu, B. ALDH7A1 expression is associated with recurrence in patients with surgically resected non-small-cell lung carcinoma. Future Oncol. 2013 9, 737-745. https://doi-org.proxy.insermbiblio.inist.fr/10.2217/fon.13.19. [CrossRef]
- Lee, J.-S.; Lee, H.; Woo, S. M.; Jang, H.; Jeon, Y.; Kim, H.Y.; Song, J.; Lee, W.J.; Hong, E.K.; Park, S.-J.; et al. Overall survival of pancreatic ductal adenocarcinoma is doubled by Aldh7a1 deletion in the KPC mouse. Theranostics 2021 11, 3472-3488. https://doi-org.proxy.insermbiblio.inist.fr/10.7150/thno.53935. [CrossRef]
- Wong, J.W.-Y.; Chan, C.-L.; Tang, W.-K.; Cheng, C.H.-K.; Fong, W.-P. Is antiquitin a mitochondrial enzyme? J. Cell. Biochem. 2010 109, 74-81. https://doi-org.proxy.insermbiblio.inist.fr/10.1002/jcb.22381. [CrossRef]
- Babbi, G.; Baldazzi, D.; Savojardo, C.; Martelli, P.L.; Casadio, R. Highlighting human enzymes active in different metabolic pathways and diseases: the case study of EC 1.2.3.1 and EC 2.3.1.9. Biomedicines 2020 8, 250. https://doi-org.proxy.insermbiblio.inist.fr/10.3390/biomedicines8080250. [CrossRef]
Figure 1.
Histological features of the four experimental rat mesothelioma tumor models. HPS staining, x400 (the scale bar represents 50 µm). Inserts (bottom right corner) represent general views (the scale bar representing 5 mm (left column) or 2.5 mm (right column)), with the open red arrows showing the location of the magnified areas.
Figure 1.
Histological features of the four experimental rat mesothelioma tumor models. HPS staining, x400 (the scale bar represents 50 µm). Inserts (bottom right corner) represent general views (the scale bar representing 5 mm (left column) or 2.5 mm (right column)), with the open red arrows showing the location of the magnified areas.
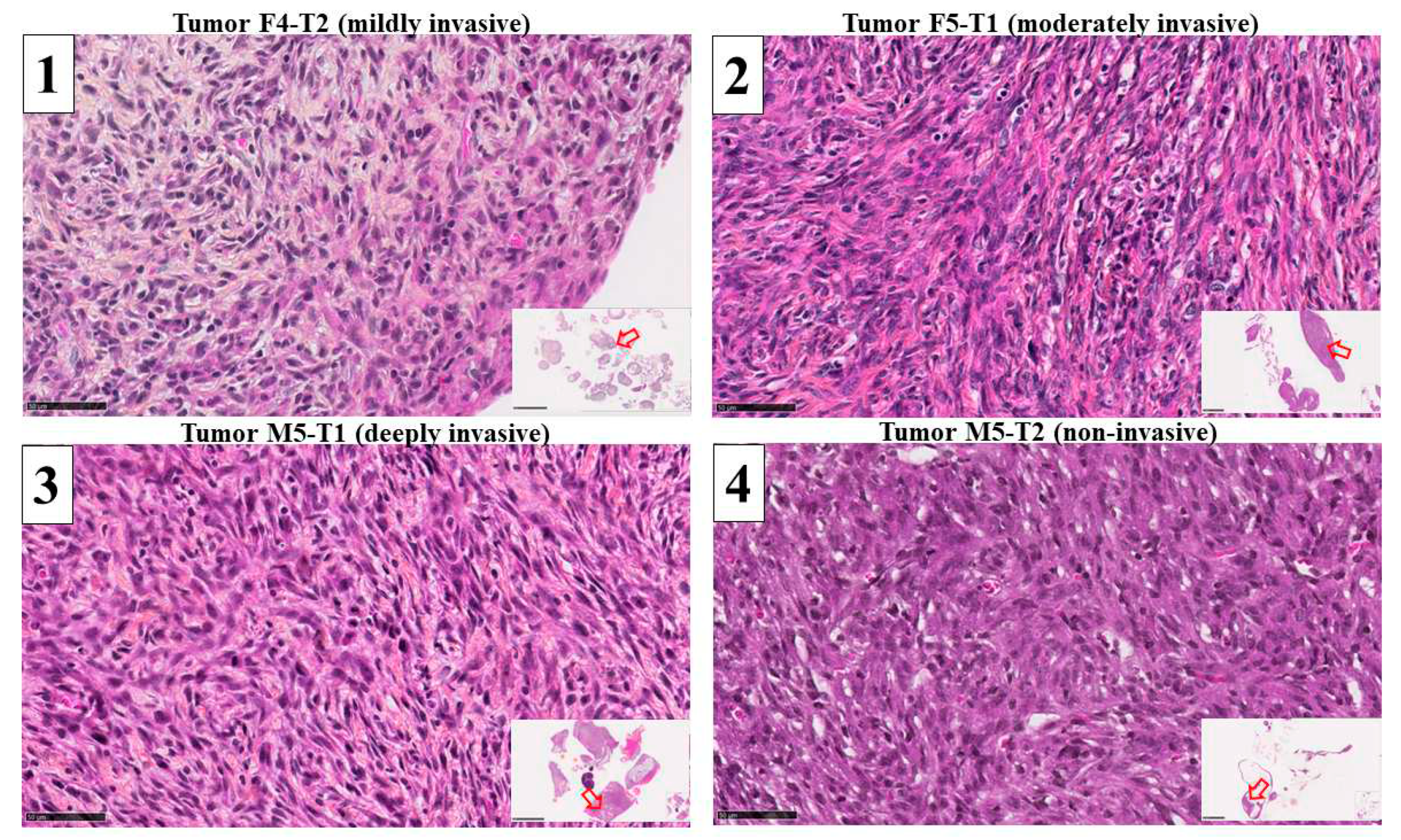
Figure 2.
Abundance changes with invasiveness, main mitochondrial proteins. A, FAO enzymes. B, ATP synthase subunits. C, Cytochrome oxidase subunits. D, TCA enzymes. Red bars represent increase and blue bars decrease, with light colors used for tendencies. Protein codes (for rattus norvegicus) are put in upper case and bold, and gene names in italics.
Figure 2.
Abundance changes with invasiveness, main mitochondrial proteins. A, FAO enzymes. B, ATP synthase subunits. C, Cytochrome oxidase subunits. D, TCA enzymes. Red bars represent increase and blue bars decrease, with light colors used for tendencies. Protein codes (for rattus norvegicus) are put in upper case and bold, and gene names in italics.
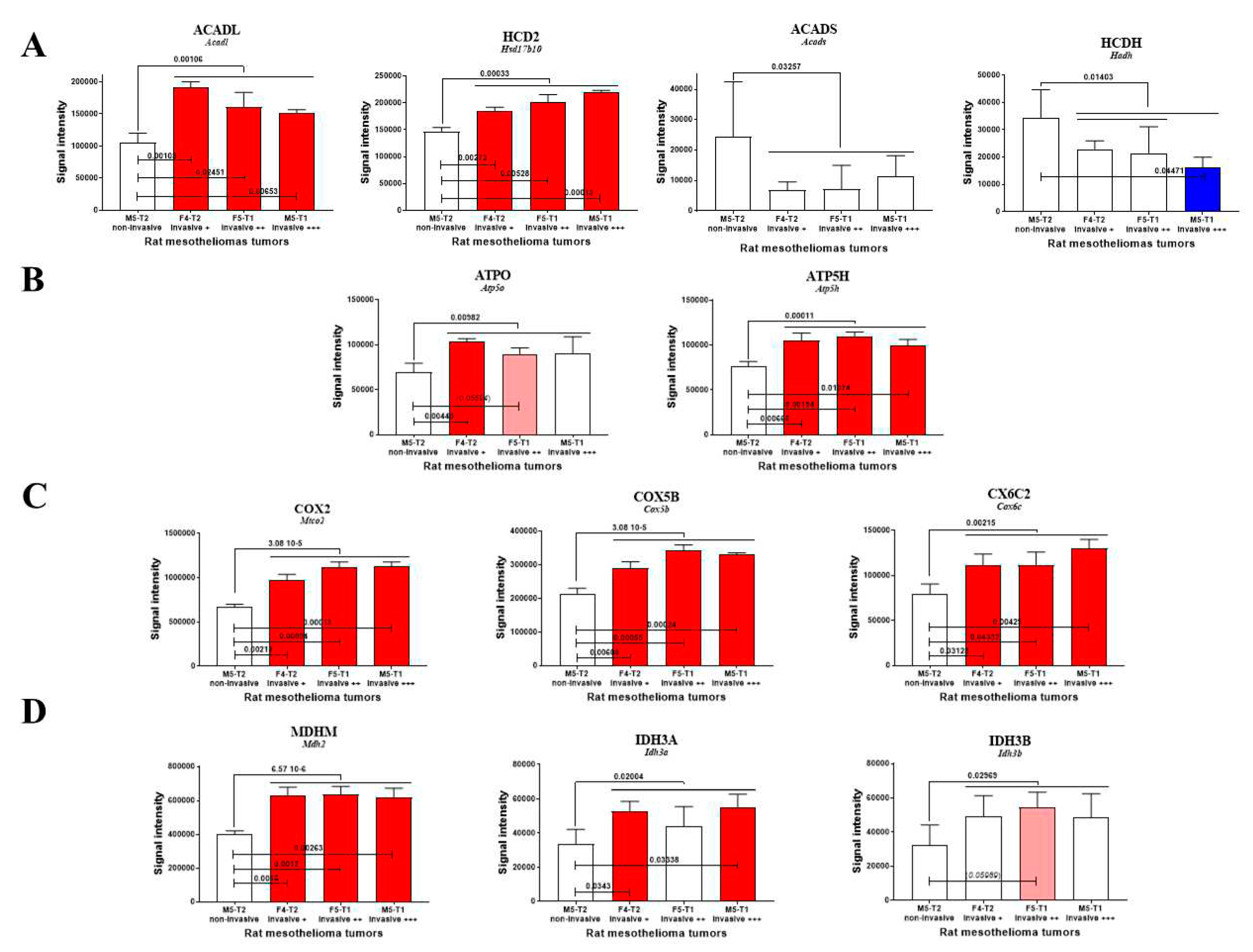
Figure 3.
Distribution of ACADL expression in rat mesothelioma tumors. A-D, Comparison of overall IHC staining with increasing invasiveness, x400 (the scale bars represent 50 µm). E, magnifications of areas of intense staining in the most aggressive, M5-T1 tumor (the scale bars represent 25 µm).
Figure 3.
Distribution of ACADL expression in rat mesothelioma tumors. A-D, Comparison of overall IHC staining with increasing invasiveness, x400 (the scale bars represent 50 µm). E, magnifications of areas of intense staining in the most aggressive, M5-T1 tumor (the scale bars represent 25 µm).
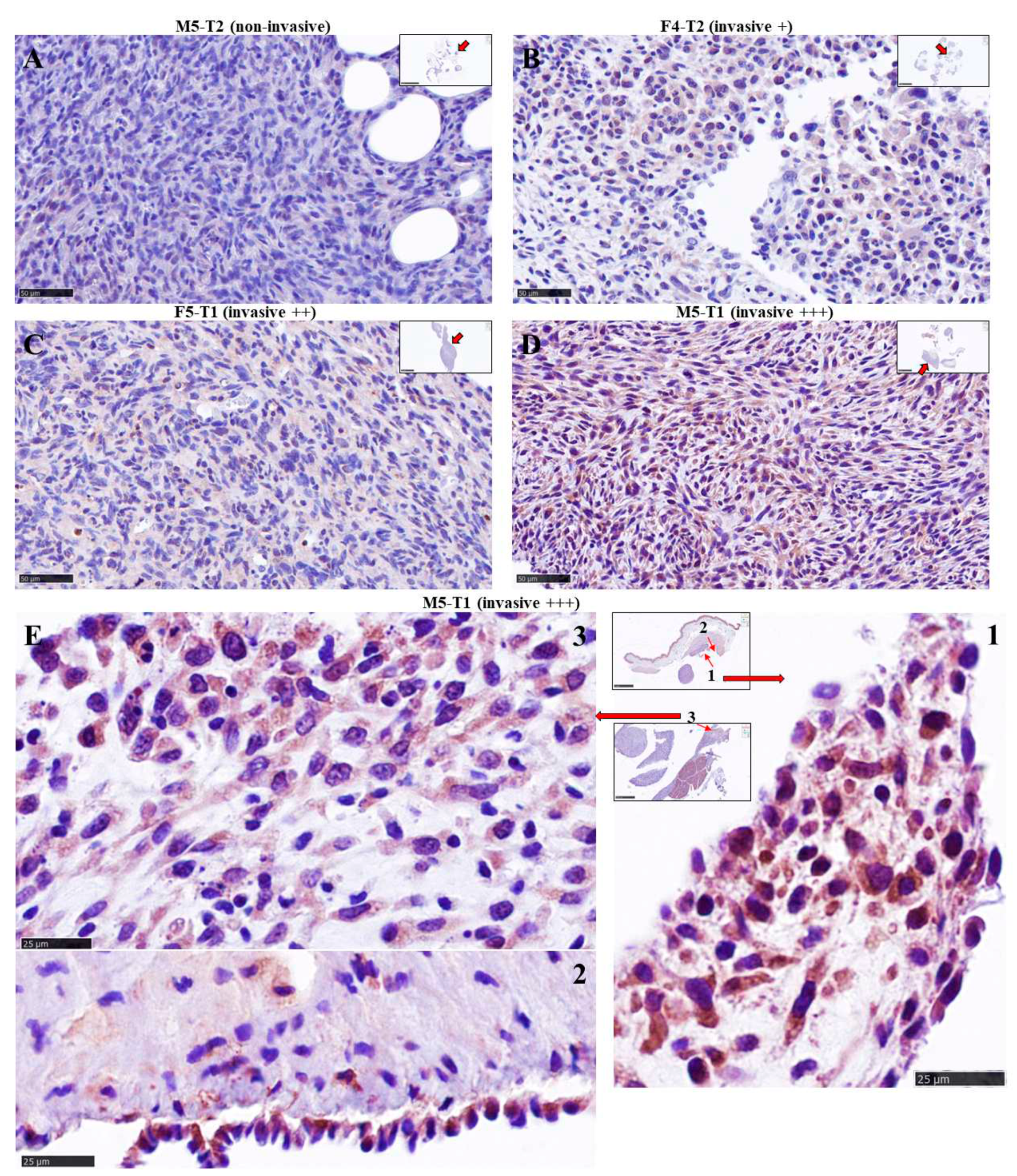
Figure 4.
Different expressions of mitochondrial genes between epithelioid and sarcomatoid MM cells. A, Mitochondrial gene expression in 10 primary MM cell lines (Supplementary Table S3) derived from 2 different histopathological subtypes, epithelioid (EPI, n = 5) and sarcomatoid (SAR, n = 5), was analyzed with real time PCR. Data are expressed as relative mean fold increase SAR vs EPI MM cells. B, C. MPM epithelioid (EPI UP1 and EPI UP2), and sarcomatoid (SAR UP6 and SAR UP7) cells were grown to confluence, then scratched and incubated for 24h in fresh medium (CTRL), or medium with 10 µM of etomoxir (ETOM). B, Representative bright-field images immediately after the scratch and after 24h. C, Cell migration. Data are presented as means ± SEM (n = 3). *p < 0.05, ***p < 0.001: ETOM treated cells vs CTRL cells; #p < 0.05, ###p < 0.001: SAR cells vs EPI cells. Scale bar is 100 µm.
Figure 4.
Different expressions of mitochondrial genes between epithelioid and sarcomatoid MM cells. A, Mitochondrial gene expression in 10 primary MM cell lines (Supplementary Table S3) derived from 2 different histopathological subtypes, epithelioid (EPI, n = 5) and sarcomatoid (SAR, n = 5), was analyzed with real time PCR. Data are expressed as relative mean fold increase SAR vs EPI MM cells. B, C. MPM epithelioid (EPI UP1 and EPI UP2), and sarcomatoid (SAR UP6 and SAR UP7) cells were grown to confluence, then scratched and incubated for 24h in fresh medium (CTRL), or medium with 10 µM of etomoxir (ETOM). B, Representative bright-field images immediately after the scratch and after 24h. C, Cell migration. Data are presented as means ± SEM (n = 3). *p < 0.05, ***p < 0.001: ETOM treated cells vs CTRL cells; #p < 0.05, ###p < 0.001: SAR cells vs EPI cells. Scale bar is 100 µm.
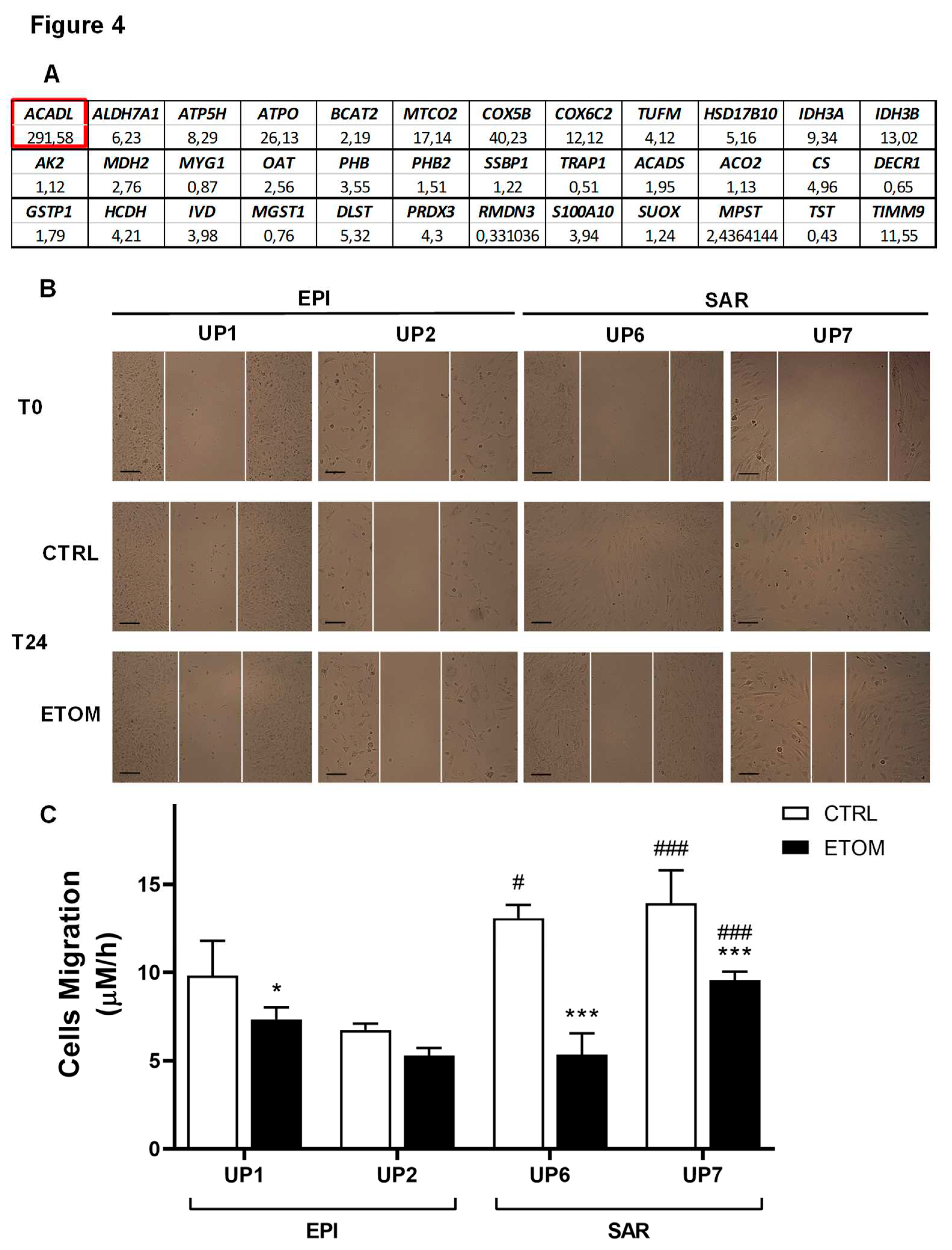
Figure 5.
Sarcomatoid MPM cells have higher expression of ACADL compared with epithelioid MM cells. Primary MM cells derived from 2 different histopathological subtypes, epithelioid (EPI UP1 and UP2), and sarcomatoid (SAR UP6 and UP7) were incubated in fresh medium (CTRL), or in medium with 10 µm of etomoxir (ETOM) for 24h then used for measurements. A, ACADL mRNA levels were measured with RT-PCR, in triplicate. Data are presented as means ± SEM (n = 3). ###p < 0.001: SAR cells vs EPI cells. B, ACADL protein was measured with immunoblotting in primary MM cell lines. GAPDH was used as a loading control. The figure is representative of 1 out of 3 experiments with similar results.
Figure 5.
Sarcomatoid MPM cells have higher expression of ACADL compared with epithelioid MM cells. Primary MM cells derived from 2 different histopathological subtypes, epithelioid (EPI UP1 and UP2), and sarcomatoid (SAR UP6 and UP7) were incubated in fresh medium (CTRL), or in medium with 10 µm of etomoxir (ETOM) for 24h then used for measurements. A, ACADL mRNA levels were measured with RT-PCR, in triplicate. Data are presented as means ± SEM (n = 3). ###p < 0.001: SAR cells vs EPI cells. B, ACADL protein was measured with immunoblotting in primary MM cell lines. GAPDH was used as a loading control. The figure is representative of 1 out of 3 experiments with similar results.
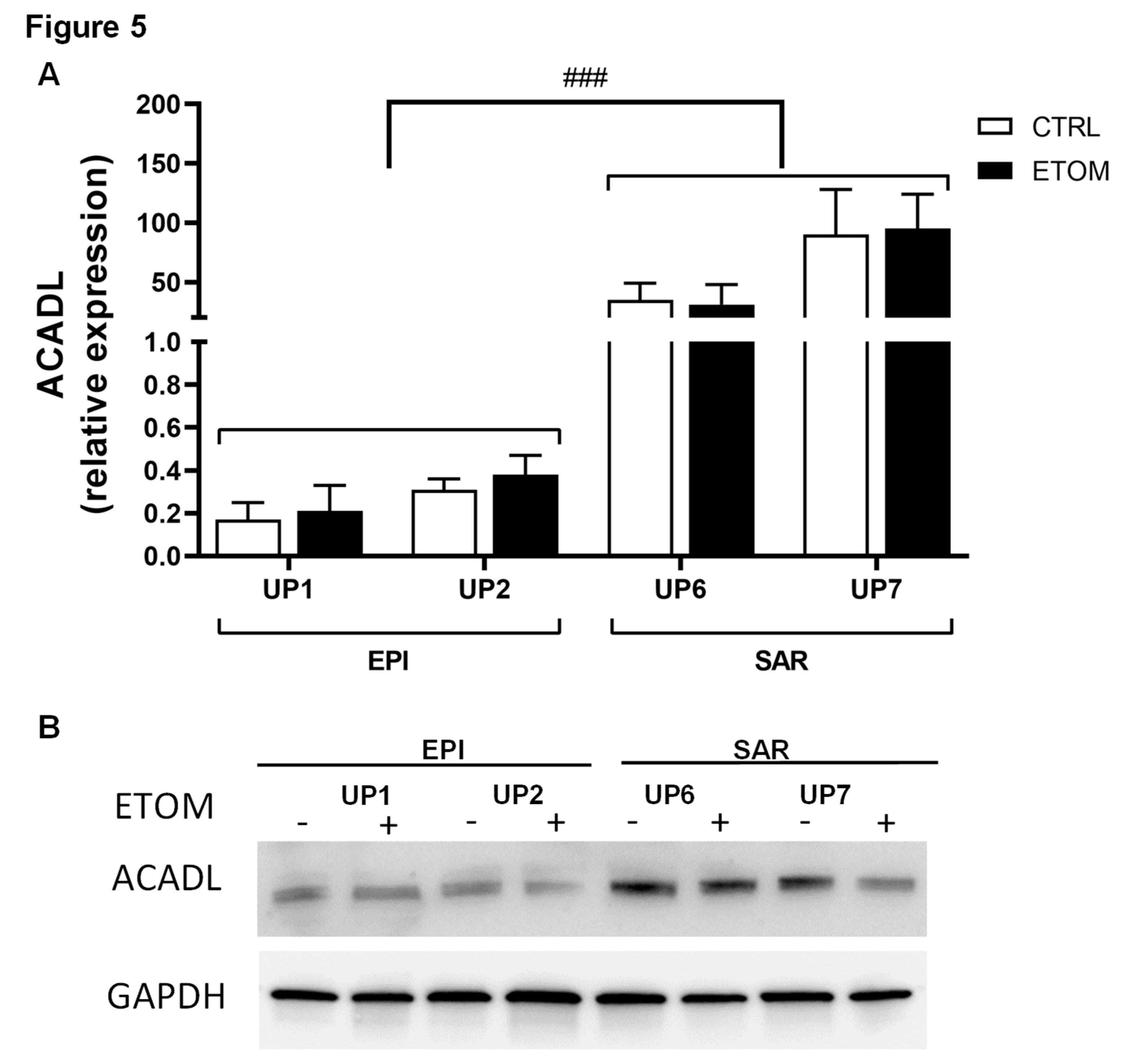
Figure 6.
Sarcomatoid MM cells have more active mitochondrial metabolism compared with epithelioid MM cells. Primary MM cells derived from 2 different histopathological subtypes, epithelioid (EPI UP1 and UP2), and sarcomatoid (SAR UP6 and SAR UP7), were grown in fresh medium (CTRL) or in medium with 10 µM of etomoxir for 24h then used for the following analysis. A, Fatty acid -oxidation was measured with fluorimetric assay in triplicate. Data are presented as means ± SEM (n = 3). ***p < 0.001: ETOM treated cells vs CTRL cells; #p < 0.05, ###p < 0.001: SAR cells vs EPI cells. B, The electron flux between Complex I and III was measured spectrophotometrically in triplicate. Data are expressed as means ± SEM (n = 3). *p < 0.05, ***p < 0.001: ETOM treated cells vs CTRL cells, ###p < 0.001: SAR cells vs EPI cells. C, ATP release was measured with a chemiluminescence-based assay in duplicate. Data are expressed as means ± SEM (n = 3). *p < 0.05, ***p < 0.001: ETOM treated cells vs CTRL cells; #p < 0.05, ###p < 0.001: SAR cells vs EPI cells.
Figure 6.
Sarcomatoid MM cells have more active mitochondrial metabolism compared with epithelioid MM cells. Primary MM cells derived from 2 different histopathological subtypes, epithelioid (EPI UP1 and UP2), and sarcomatoid (SAR UP6 and SAR UP7), were grown in fresh medium (CTRL) or in medium with 10 µM of etomoxir for 24h then used for the following analysis. A, Fatty acid -oxidation was measured with fluorimetric assay in triplicate. Data are presented as means ± SEM (n = 3). ***p < 0.001: ETOM treated cells vs CTRL cells; #p < 0.05, ###p < 0.001: SAR cells vs EPI cells. B, The electron flux between Complex I and III was measured spectrophotometrically in triplicate. Data are expressed as means ± SEM (n = 3). *p < 0.05, ***p < 0.001: ETOM treated cells vs CTRL cells, ###p < 0.001: SAR cells vs EPI cells. C, ATP release was measured with a chemiluminescence-based assay in duplicate. Data are expressed as means ± SEM (n = 3). *p < 0.05, ***p < 0.001: ETOM treated cells vs CTRL cells; #p < 0.05, ###p < 0.001: SAR cells vs EPI cells.
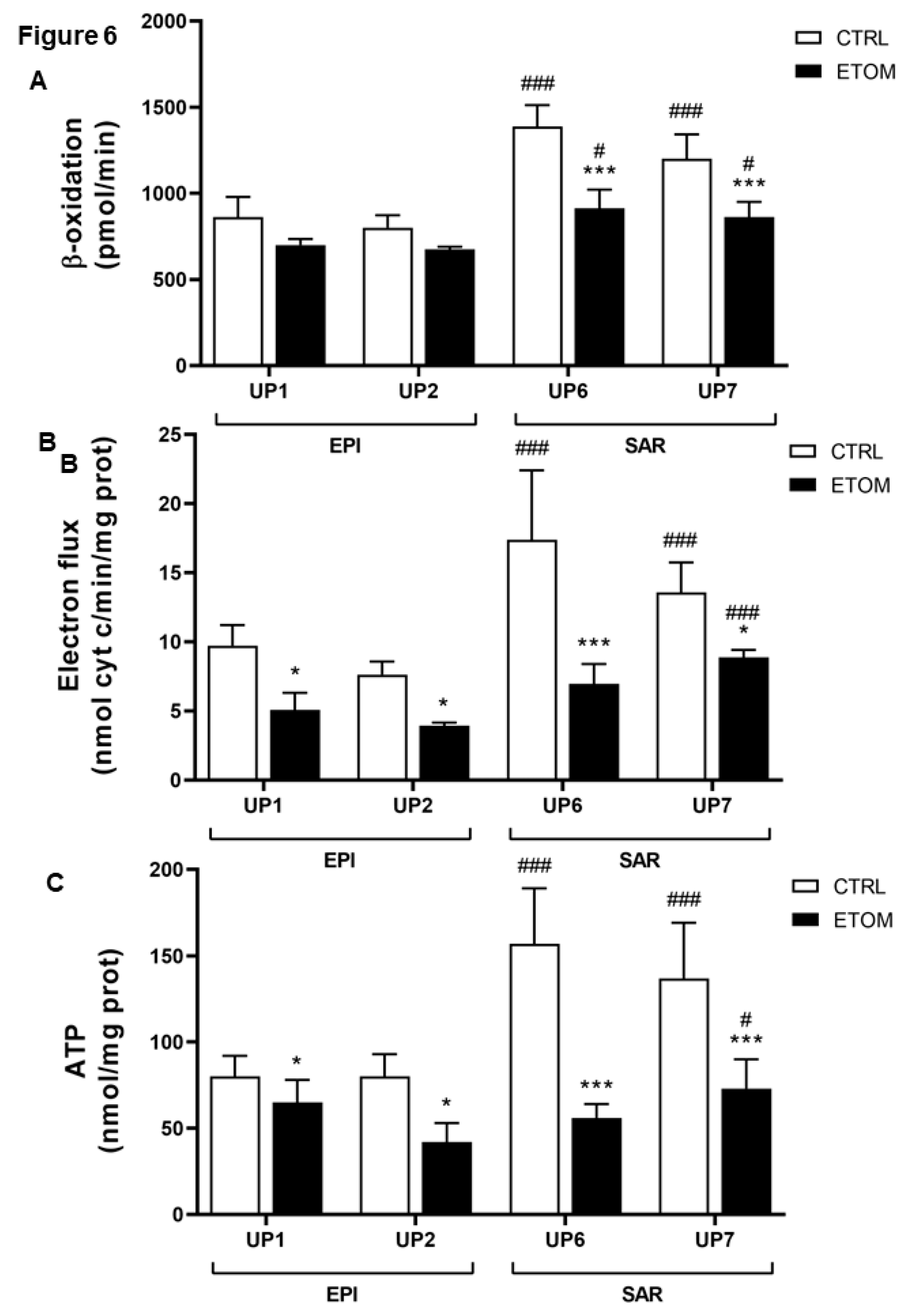

Table 1.
Origin and characteristics of human mesothelioma cell lines.
| UNP | Histotype | Gender | Age | Asbestos exposure | First line | Second line treatment | TTP | OS |
|---|---|---|---|---|---|---|---|---|
| 1 | Epithelioid | M | 74 | Unknown | Carbo+Pem | No | 7 | 11 |
| 2 | Epithelioid | F | 58 | Yes | Carbo+Pem | Pem | 6 | 13 |
| 3 | Epithelioid | M | 76 | Unknown | CisPt+Pem | No | 3 | 8 |
| 4 | Epithelioid | M | 68 | Yes | Carbo+Pem | Pem | 4 | 9 |
| 5 | Epithelioid | F | 84 | Yes | CisPt+Pem | No | 7 | 8 |
| 6 | Sarcomatoid | M | 80 | Yes | Carbo+Pem | Trabectedin | 3 | 5 |
| 7 | Sarcomatoid | F | 78 | Unknown | Pem | No | 4 | 6 |
| 8 | Sarcomatoid | M | 69 | Yes | Carbo+Pem | Trabectedin | 7 | 10 |
| 9 | Sarcomatoid | F | 74 | Unknown | Carbo+Pem | No | 5 | 7 |
| 10 | Sarcomatoid | M | 78 | Yes | Carbo+Pem | Trabectedin | 4 | 9 |
UNP: unknown patient; M: male; F: female; Carbo: carboplatin; Pem: pemetrexed; CisPt: cisplatin; TTP: time to progression; OS: overall survival.
Table 2.
Mitochondrial proteins exhibiting significant abundance changes (p < 0.05) in the three invasive rat malignant mesothelioma tumors relative to the non-invasive tumor.
Table 2.
Mitochondrial proteins exhibiting significant abundance changes (p < 0.05) in the three invasive rat malignant mesothelioma tumors relative to the non-invasive tumor.
| Code # | Gene # | Full name # | [1 + 2 + 3] vs 4 |
|---|---|---|---|
| ACADL | Acadl | Long-chain specific acyl-CoA dehydrogenase, mitochondrial | ↑ |
| AL7A1* | Aldh7a1 | Alpha-aminoadipic semialdehyde dehydrogenase | ↑ |
| ATP5H | Atp5h | ATP synthase subunit d, mitochondrial | ↑ |
| ATPO | Atp5o | ATP synthase subunit O, mitochondrial | ↑ |
| BCAT2* | Bcat2 | Branched-chain-amino-acid aminotransferase, mitochondrial | ↑ |
| COX2 | Mtco2 | Cytochrome c oxidase subunit 2 | ↑ |
| COX5B | Cox5b | Cytochrome c oxidase subunit 5B, mitochondrial | ↑ |
| CX6C2 | Cox6c2 | Cytochrome c oxidase subunit 6C-2 | ↑ |
| EFTU | Tufm | Elongation factor Tu, mitochondrial | ↑ |
| HCD2 | Hsd17b10 | 3-hydroxyacyl-CoA dehydrogenase type-2 | ↑ |
| IDH3A | Idh3a | Isocitrate dehydrogenase [NAD] subunit alpha, mitochondrial | ↑ |
| IDH3B | Idh3b | Isocitrate dehydrogenase [NAD] subunit beta, mitochondrial | ↑ |
| KAD2 | Ak2 | Adenylate kinase 2, mitochondrial | ↑ |
| MDHM | Mdh2 | Malate dehydrogenase, mitochondrial | ↑ |
| MYG1* | Myg1 | UPF0160 protein MYG1, mitochondrial | ↑ |
| OAT* | Oat | Ornithine aminotransferase, mitochondrial | ↑ |
| PHB | Phb | Prohibitin | ↑ |
| PHB2 | Phb2 | Prohibitin-2 | ↑ |
| SSBP | Ssbp1 | Single-stranded DNA-binding protein, mitochondrial | ↑ |
| TRAP1 | Trap1 | Heat shock protein 75 kDa, mitochondrial | ↑ |
| ACADS | Acads | Short-chain specific acyl-CoA dehydrogenase, mitochondrial | ↓ |
| ACON | Aco2 | Aconitate hydratase, mitochondrial | ↓ |
| CISY* | Cs | Citrate synthase, mitochondrial | ↓ |
| DECR* | Decr1 | 2, 4 dienoyl-CoA reductase, mitochondrial | ↓ |
| GSTP1* | Gstp1 | Glutathione S-transferase P | ↓ |
| HCDH | Hadh | Hydroxyacyl-CoA dehydrogenase, mitochondrial | ↓ |
| IVD* | Ivd | Isovaleryl-CoA dehydrogenase, mitochondrial | ↓ |
| MGST1* | Mgst1 | Microsomal glutathione S-transferase 1 | ↓ |
| ODO2 | Dlst | Dihydrolipoyllysine-residue succinyltransferase component of 2-oxoglutarate dehydrogenase complex, mitochondrial | ↓ |
| PRDX3 | Prdx3 | Thioredoxin-dependent peroxide reductase, mitochondrial | ↓ |
| RMD3* | Rmdn3 | Regulator of microtubule dynamics protein 3 | ↓ |
| S10AA | S100a10 | Protein S100-A10 | ↓ |
| SUOX | Suox | Sulfite oxidase, mitochondrial | ↓ |
| THTM | Mpst | 3-mercaptopyruvate sulfurtransferase | ↓ |
| THTR | Tst | Thiosulfate sulfurtransferase | ↓ |
| TIM9 | Timm9 | Mitochondrial import inner membrane translocase subunit Tim9 | ↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



