
Submitted:
13 June 2023
Posted:
14 June 2023
You are already at the latest version
Abstract
Mycobacterium tuberculosis (Mtb) the causative agent of human tuberculosis (TB) is one of the most successfully adapted-human pathogens. Human-to-human transmission occurs at high rates through aerosols containing bacteria, but the pathogen evolved prior to the establishment of crowded populations. Mtb has developed a particular strategy to ensure persistence in the host until an opportunity for transmission arise. It has refined its lifestyle to obviate the need for virulence factors such as capsules, flagella, pilli, or toxins to circumvent mucosal barriers. Instead, the pathogen uses host macrophages, where it establishes intracellular niches, for its migration into the lung parenchyma and other tissues and for the induction of long-lived latency in granulomas. Finally, at the end of the infection cycle Mtb induces necrotic cell death of macrophages to escape to extracellular milieu and instructs a strong inflammatory response required for progression from latency to disease and transmission. Common to all these events is ESAT-6, one of the major virulence factors secreted by the pathogen. This review highlights the recent advances in understanding the role of ESAT-6 in hijacking macrophage function to establish a successful infection and transmission and its use as a target for the development of diagnostic tools, and vaccines.
Keywords:
Tuberculosis
; ESAT-6
; ESX-1
; virulence factors
; T7SS
; PhoPR signal transduction
; host-pathogen interactions
; TB vaccines
; TB diagnosis
1. Introduction
Human tuberculosis (TB) is one of the world’s deadliest infectious disease [1,2,3]. In 2022 the WHO reported approximately 1.6 million deaths, with 10 million new infections and an estimated one-quarter of the human population latently infected [3]. Infection control is hampered by the limited efficacy of a 100-year-old Bacille Calmette-Guérin (BCG) vaccine and by the emergence of multidrug-resistant strains to 70 year-old antibiotics, particularly those used as first line therapy [3]. There is also a need to develop better diagnostic tools, more sensitive, using simple instruments easy to handle in remote geographical areas, and cost-effective for rapid detection of Mtb. Moreover, although new methods are being implemented [4], currently available methods do not allow differentiation between different stages of infection [4,5,6,7,8].
Mycobacterium tuberculosis (Mtb) is considered the mainly causative agent of human TB [9]. Mtb infection primarily affects the lungs, a condition that contributes to the high transmissibility by the respiratory route[9]. Although not transmitted from person to person, extrapulmonary TB is a second form of disease manifestation that particularly affects less immunocompetent individuals, namely children or people co-infected with HIV [10,11,12]. In addition to Mtb, other pathogens can also infect humans and cause TB with similar clinical symptoms [13,14,15,16,17,18,19,20]. These, include M. africanum, which is restricted to humans in West Africa [21,22,23], where it causes nearly half of all pulmonary TB [18,19,20] and the animal-adapted species M. bovis, which is estimated to cause TB in about 2% of the world’s population [24,25]. M.caprae is responsible for a smaller burden of zoonotic TB [26]. They all belong to the Mycobacterium tuberculosis species complex (MTBC) which also includes the more distant M. canettii group and additional species that cause TB in animals such as M. microti [27], M. pinnipedii, M. orygis and M. mungi [20,23,28,29,30]. Although each of these animal MTBC variants causes TB in its host species, they may trigger slight or no disease outside of their adapted host, especially in immunocompetent hosts [13,30,31]. However, conditions such as geographic prevalence of infected animals, close humans contacts, route of transmission (milk, infected meet or airborne droplets containing pathogens), and conditions such as HIV infection or other immunosuppressive conditions should be taken in account to assess the true threat to public health [32].
The animal-adapted species are more recent pathogens that emerged as a crowd-associated diseases during the Neolithic demographic expansion along with the development of animal domestication [33]. Mtb ascended long before the establishment of crowdy populations, 70 thousand years ago and accompanied the exodus of Homo sapiens from Africa during the Neolithic expansion [34]. The initial virulence of Mtb is therefore strongly adapted to the occasional availability of the population to be infected and to an evolutionary ability to survive long in the host until the opportunity for transmission arises. Consequently, Mtb infection have evolved at low human population densities exhibiting a pattern of chronic development, accompanied by decades of latency before progressing to active disease [10,11,12]. It is a well-adapted human pathogen, requiring the induction of a strong inflammation and destruction of the lung tissue for transmission and evolutionary survival [9,34,35]. This feature is unusual in most pathogens where virulence is not associated with their spread to other hosts [36].
The pathogen does not seem to require virulence factors such as the usual virulence factors pili, toxins or capsules that are essential for the invasion of epithelial barriers [9]. Nevertheless, their potential involvement should not be ignored, as a capsule and pili have recently been identified in Mtb, although their roles during infection in vivo is still unclear [37,38,39]. Mtb virulence relies on its ability to manipulate host macrophages, where it establishes intracellular niches, to cross mucosal barriers and to avoid pathogen destruction. First, Mtb subverts the endocytic pathway preventing phagolysosome fusion and proteolytic digestion [40,41,42,43,44,45]. Second, it activates innate immune responses to induce their transmigration to the lung parenchyma [46,47]. There, infected macrophages attract more permissive cells, expanding intracellular niches [9,48,49,50]. Mtb induces the adaptive responses that stimulate their containment and long-live inside granulomas [51,52,53,54]. Finally, the pathogen induces necrotic cell death of macrophages, granuloma destruction and lung cavitation for transmission [47,49,55,56]. Common to all these events is the major virulence factor the “early secreted antigenic target of 6 kDa” (ESAT-6, also called EsxA). This review highlights the role of ESAT-6 in different phases of Mtb infection and its contribution to virulence. It also points ESAT-6 as a target for the development of better diagnostic tools and future vaccines for human tuberculosis.
2. Virulence evolution among MTBC
Comparative genomic analysis among MTBC species reveals more than 99,95% sequence homology [57], but they differ mainly by large sequence polymorphisms [18] relative to Mtb reflected by the so-called regions of difference (RD) and translated into deletions [57,58,59,60,61] or punctual insertional sequences [62]. These observations reinforce the ancestral origin of Mtb reflecting a loss of genes during transmission to animals that allowed fitness gain in the new host and loss of robustness in humans. In the case of M. africanum lineage 5 (L5) is more associated with Mtb-like lineages while lineages L6 and 9 displays RD more associated with M. bovis-like animal adapted species [19,20].
Important cumulative findings from these studies have identified more pathogenic MTBC disease-causing species from less fit ones with deletions/insertions/mutations in RD regions affecting the PhoPR two-component virulence system, or the genes under the control of this signal transduction regulator [63,64]. This includes the regions called region of difference 1 (RD1) and the region of difference 8 (RD8), which are responsible for the production and secretion of the virulence factor ESAT-6 [29,62].
Particularly the RD1 region is deleted in the animal-adapted species M. microtii and was first described in BCG, where it is absent and associated with attenuation of this live vaccine during its propagation in vitro [23,65,66]. The RD8 deletion is associated with animal-adapted species and affects the regulatory PhoPR-dependent region of the espACD operon involved in ESAT-6 secretion (Figure 1). Genetic transfer of mutations affecting the PhoP binding region in M. bovis and in the closely related M. africanum L6 into Mtb sensu stricto human species, resulted in reduced ESAT-6 secretion, and lower virulence [29]. Remarkably, the deleterious effects of these mutations were partially compensated by RD8 deletions, in both species, allowing ESAT-6 secretion to some extent by creating alternative regulatory sequences [62]. The observed attenuated ESAT-6 responses contribute for the observed slower clinical progression from infection to disease when compared to Mtb [18,67]. Conversely, the insertion of the IS6110 element upstream of the PhoP binding locus resulted in the upregulation of the operon in one multidrug-resistant M. bovis strain, responsible for an unusually high human transmissibility, partially reverting the phoPR-bovis-associated fitness loss [68,69].
3. ESAT-6 is required for virulence of Mtb
ESAT-6 was first identified in 1995 by the discovery of a potent T-cell antigen in the short-term culture filtrate of Mtb [72,73]. The initial focus was on its potential use as a target for a TB vaccine to replace BCG [74]. Moreover, since patients and animals infected with MTBC respond strongly to ESAT-6 antigens, parallel emphasis was given to the development of a better diagnostic tool for TB [75].
The export of ESAT-6 from the bacilli, requires a secretory apparatus consisting of proteins that are assembled on the inner surface of the cell membrane and are strongly recognized by T cells [70,71,76]. The encoded genes are all part of the esx-1 operon (Figure 1), and there is increasing evidence that they are under selective pressure imposed by the host immune system [76]. Concomitant with these studies, comparative functional genomics among virulent, attenuated, or saprophytic mycobacterial species have contributed to the inclusion of ESAT-6 in Mtb virulence [77]. Curiously, the saprophyte species M. smegmatis (Ms) also encodes for an ESX-1 apparatus [78], however it does not appear to confer Ms virulence capabilities as demonstrated by its inability to survive in human macrophages [79] or in amoeba in the environment [9]. The emergence of the ESX-1-associated virulence in Mtb stems from the original role of ESX-1 in Ms and other GC-rich soil species in inducing horizontal gene transfer between bacteria [80,81,82]. Predatory amoeba may have contributed to the evolutionary pressure that selected mycobacterial pathogens for intracellular survival. ESAT-6 requires an extended esx-1 locus to be secreted, a putative genomic island containing the espACD locus which is adjacent to the region of difference 8 (RD8) (Figure 1). It has been hypothesized that this phenotype may be associated with independent horizontal gene transfer from pre-pathogenic mycobacteria in the contact with soil bacteria and positively selected while in amoeba phagosomes [83,84].
ESAT-6 secretion from the bacilli requires both the expression of the esx-1 locus for the type VII secretion apparatus and for the transcription of both the ESAT-6 gene (esxA) and the culture filtrate protein 10 (CFP-10) gene (esx-B) contained in the RD1 region. In addition, it requires the protein EspA which is not encoded in the esx-1 locus but in the extended espACD operon adjacent to RD8 [85] (Figure 1), which is located 260 kb upstream of esx-1 [71].
All species and strains deleted in the esx-1 locus or in the internal RD1 region, or in the the esx-1 extended locus espACD exhibits an attenuated phenotype [86]. Mutants with deletion on ESX-1 of Mtb are attenuated in virulence translated into reduced survival of mycobacteria in cultured macrophages or in experimental animal models of TB [87,88,89,90,91,92], which is consistent with the attenuation of BCG vaccine, or the species M. microti due to the deletion of the region of difference RD1 [65,66]. Likewise, the introduction of RD1 into BCG restores the virulence [90,93]. The reference laboratory strain Mtb H37Ra has a mutation in the phoP regulatory region responsible for the attenuated phenotype compared to its virulent counterpart Mtb H37Rv resulting in impaired ESAT-6 secretion [63,94]. Clinical Mtb isolates were shown to secrete higher levels of ESAT-6 than the reference laboratory strain Mtb H37Rv. Comparative analysis of genetic polymorphisms between clinical and laboratory strains revealed whiB6 (rv3862c), a gene upstream of the ESX-1 genetic locus. It encodes for a regulatory protein that activates promoters under the esx-1 and extended espACD locus responsible for ESAT-6 production and secretion, respectively [95] (Figure 1).
Clearly, the loss or gain of mycobacterial virulence is closely linked to the ability of mycobacteria to produce and secrete ESAT-6 and the extension of virulence is correlated with the amount of protein secreted.
3. ESAT-6 in Mtb pathogenesis
3.1. During the early phases of infection: the innate phase
Upon inhalation of aerosols containing the pathogen, bacilli that manage to reach the alveoli are phagocytosed by alveolar macrophages [10]. These professional phagocytic cells are permissive to Mtb and provide a critical niche for the survival of the intracellular pathogen [9,44,96,97]. In the endocytic pathway the pathogen inhibits phagosome fusion with lysosomes, prevents vesicle acidification and its proteolytic destruction by lysosomal enzymes allowing the pathogen to replicate in early phagosomes [40,41,42,43,44,45]. Recent studies showed that ESAT-6 inhibits IL-18-mediated phagolysosome fusion by regulating microRNA-30a in mycobacteria-infected macrophages [98,99]. ESAT-6 was indeed associated with the blockade of phagosomal maturation because mutants in ESAT-6 of M. marinum, a pathogen that causes a TB-like disease in fish, were found to be mainly located in lysosomes in contrast to the wild-type strain, which is located in early phagosomes [100].
However, luminal studies have challenged the dogma of the exclusive intracellular localization on phagosomes of both Mtb and M. marinum [101,102,103,104,105]. Studies using ultrastructural observations generated by electron microscopy and, more recently, the development of a Fluorescent Resonance Energy Transfer (FRET) method, demonstrate the translocation and escape of both pathogens from phagolysosomal compartments into the cytosol [101,102].
It has been shown that ESAT-6 induces phagosomal membrane rupture allowing pathogens to gain access to the cytosol, in contrast to the corresponding mutants, which instead accumulate in phagosomes. Moreover, following phagolysosomal escape, a necrotic form of cell death of infected macrophages was observed 3 to 4 days post-infection in ex-vivo studies [101].
It is unlikely that in all contexts of infection in vivo, ESAT-6 will induce complete rupture of the phagosome membrane and total escape of the bacteria into the cytosol, ending with necrotic cell death. Necrotic cell death occurring in vivo will induce a strong inflammatory response and tissue destruction. There is a possibility that punctual membrane perturbations may create local conditions for the transfer of Mtb proteins with less induced stress in the cytosol. ESAT- 6 is a pathogen-associated molecular pattern (PAMP) sensed by innate cytosolic receptors and is the major Mtb PAMP that activates the NLRP3 inflammasome [47]. This platform is required for caspase-1-mediated processing of the cytokines IL-1β and IL-18. The combination of purified ESAT-6 protein with other PAMPS such as Ag85 adds a significant impact on NLRP3 activation and IL-1β secretion demonstrating that ESAT-6 helps other Mtb PAMPS to reach the cytosol [47]. IL-1β drives neutrophil recruitment by several mechanisms and, if not controlled, neutrophils are major instructors for tissue destruction [55,106]. The mechanism is controlled by nitric oxide (NO) released by IFN𝛾-activated macrophages [55,106]. NO in turn controls the inhibition of the NLRP3 inflammasome, which regulates the amounts of cytokines secreted.
Independent studies have shown that NLRP3 activation does not always results in necrotic cell death of infected cells and tissue destruction. Transmigration of infected alveolar macrophages (AM) into the lung parenchyma is dependent on Mtb ESX-1 inducing IL-1β via the NLRP3 inflammasome. As a result, IL-1R signalling on alveolar pneumocytes affects alveolar permeability and lung tissue access without tissue destruction [46,47].
Once inside the lung, infected macrophages, activate pneumocytes surrounding the nascent granuloma to secrete matrix metalloproteinase 9 (MMP9) in an ESAT-6-dependent manner [107,108,109]. Consequently, an influx of more permissive macrophages following the MMP9 signals reach the nascent granuloma and efficiently find and perform efferocytosis of dying apoptotic infected cells [110,111]. Continuous cycles of this process allow the expansion of Mtb intracellular niches [107,108,112]. The intracellular replication and bacterial load are controlled by distinct mechanisms, some of which depend on perturbations of the ESAT-6 phagosomal membrane with concomitant cytosolic PAMPS that activate different antimicrobial mechanisms including apoptosis [48,110] and autophagy [113]. The ability to induce apoptosis is a feature of virulent strains of M. tuberculosis, in a process that involves ESAT-6 [50,114]. Moreover has also been shown that inhibition of apoptosis by non-virulent mutants of M. marinum impairs the spread of infection and bacterial expansion on nascent granulomas [52]. The apoptotic form of controlling intracellular bacterial loads counteracts the necrotic forms of death usually observed during high load infection of macrophages in vitro [115], and contributes to cell-to-cell spread of Mtb.
Autophagy is a relevant innate response to control intracellular pathogens including Mtb [116]. ESAT-6 contributes to this pathway [113,117,118]. Its functions in perturbing the phagosomal membrane will expose PAMPS including mycobacterial DNA, a signature that can be sensed by host innate cytosolic receptors. These include at least 3 cytosolic sensors, two involving the inflammasomes NOD-, LRR-, and pyrin domain-containing 3 (NLRP3) and absent in melanoma 2 (AIM2), and the third the cyclic GMP-AMP synthase (cGAS) [117]. The latter leads to the synthesis of the second messenger cyclic GMP-AMP (cGAMP), which activates the endoplasmic reticulum-associated stimulator of interferon genes (STING) and the downstream serine/ threonine-protein kinase (TBK1)-interferon regulatory factor 3 (IRF3)–IFN-I signaling pathway (Figure 2) [113,119]. TBK1 provides a bridge for Mtb destruction by targeting intracellular bacteria to the ubiquitin-mediated autophagic pathway in macrophages [113].
3.2. During latency
The latency period can last for decades after the initial infection. It is defined as a state of immune activation in the absence of disease symptoms [11]. During the innate granuloma expansion, the adaptive responses are activated, and the latency phase begins with the arrival of effector T-lymphocytes to the granuloma [10]. The adaptive granuloma is a structure that contains Mtb in the lung, but it is also the result of a strategic manipulation by the pathogen to ensure a long life in the host. A fine balance of proinflammatory microbicidal mechanisms with immunosuppressive events drives the process. Bacterial loads are kept at bay in macrophages by effector CD4+ T-cells, such as Th1 secreting the cytokines IFN𝛾 and TNFα, Th17 secreting IL-17 and, in much smaller numbers, Th2 and regulatory T-cells that counteract the inflammatory effects of the former [10]. The adaptive T-cells are constantly arriving at the granuloma, forming a surface layer, and a few infiltrating cells will license infected macrophages to exert their activity. In the case of Th1, this licensing induces macrophages to secrete IL-12 allowing efficient IFN𝛾 and TNFα release from effector T-cells. Mtb ESAT6 has been shown to induce immunosuppressive regulatory T cell populations that delay effector T cells migration into the granuloma, a process that is reversed by IL-12 [120]. In turn, IFN𝛾 and TNFα activate infected macrophages to become more microbicidal. However, Mtb resists these effects by reducing macrophage responsiveness to signaling by IFN𝛾 [121]. IFN𝛾 also contributes to NO release from activated macrophages, a mechanism required to control IL-1β and tissue destruction, helping to preserve the structure of the granuloma [55].
In the case of CD8+ T-lymphocytes, ESAT-6 in infected macrophages interacts with beta-2-microglobulin (β2M) in the endoplasmic reticulum, affecting antigen presentation to effector cytotoxic T-cells and impairing their microbicidal activity [122,123].
While macrophages become more microbicidal, the pathogen activates the two-component PhoPR system, which allows the pathogen to adapt to a stressful environment such as hypoxia and low pH, while activating the dormancy regulon that puts the bacilli in a low metabolic state [124].
Activated macrophages secrete TNFα and IL-1β which permeabilize the endothelia to feed the granuloma with newly arrived macrophages and neutrophils to be infected. All of this cell arrival dynamic is fuelled by new blood vessel formation induced by an ESAT-6 driven release of the angiogenesis factor VEGF from infected cells in the granuloma. [125].
3.2. During progression to disease
Although mycobacteria exploit macrophages to maintain a long life in intracellular niches within a host, they promote their transmission to a new host by becoming extracellular [9]. At this stage, the promotion of necrotic cell death of highly infected cells allows the release of the pathogen by a process requiring ESAT-6 [115]. Moreover, as stated before apoptosis induced by virulent Mtb favours the intracellular expansion of the pathogen, but with a reduced load per host cell [48,110] whereas intracellular Mtb replication tends to enhance necrosis [49,126]. Moreover, it has been demonstrated that Mtb is able to survive in this necrotic environment. As mentioned above, ESAT-6-mediated access of the pathogen PAMPS to the cytosol, including DNA, activates several cytosolic sensors including the NLRP3, the AIM2 and the cGAS. The first two are involved in the secretion of IL-1β, while the latter leads to the release of type I interferon (IFN-I). The crosstalk between IFN-I, and IL-1β influences the progression of the disease by controlling cell death within the granuloma [117,127,128].
Observed high levels of IFN-I in the serum of infected patients have been associated with progression to TB [129]. Type I IFNs subvert anti-tuberculous host defenses by inhibiting iNOS, the enzyme responsible for NO production, while inducing the immunosuppressive mediators IL-10 [127]. Based on the stimulation of iNOS by IFNγ and the inhibition of iNOS by IFN-I, it appears that an imbalance of IL-1β results from NLRP3 inflammasome activation [55]. Under IFNγ control, IL-1β enhances TNFα -stimulated Mtb killing and contributes to controlled neutrophil recruitment. In the context of low levels of TNFα, IL-1β stimulates cyclooxygenase-2 (COX2) to produce prostaglandin E2 (PGE2) from arachidonic acid (AA), resulting in mitochondrial membrane protection and controlled intracellular burdens via apoptosis [130]. Unlike IL-1β, IFN-I stimulates 5-lipoxygenase (5-LO) which is a competitive enzyme for COX2, to produce lipoxin A4 and leukotriene B4 from AA which leads to loss of plasma membrane integrity, cytoplasmic organelles swelling such as mitochondria and nuclei, and making cells more susceptible to necrotic cell death.
Unexpectedly, excessive TNFα in the granuloma often observed during active TB promotes a necrotic form of death called necroptosis via the production of mitochondrial reactive oxygen species (ROS). [127,128,131].
The bacteria released into the caseous centre of the granuloma find a nutrient-rich environment for massive replication. This high lipid content is mostly the result of necrotic cell death of infected foamy macrophages in an ESAT-6 dependent manner [132].
Overall, IL-1β in synergy with TNFα are major inducers of neutrophil recruitment to the lung and to the granuloma. In active TB patients, neutrophils represent the major infected cell population [49]. A synergistic mechanism can be attributed to the fact that IL-1β is required for Th17 polarization, and the cells releasing IL-17 will activate endothelia to release chemokines for neutrophil recruitment. However, transmigration occurs only after endothelial E-selectins are exposed following stimulation by both cytokines [128,133]. Neutrophils are one of the main causes of pathological tissue sequelae due to bioactive neutrophil molecules, including proteases and metalloproteinases [53,134]. Indeed, Mtb in infected cells induces human neutrophils necrosis in an ESAT-6-dependent manner, and neutrophil-produced reactive oxygen species (ROS) drive this necrosis [49]. Impaired dead cell clearance leads to severe tissue inflammation and contributes to the granuloma disruption and lung cavitation required for subsequent transmission of infection to the next host [135]. The involvement of ESAT-6 in mediating the inflammation required for transmission is indeed supported by studies showing its increased expression in the highly transmissible Mtb Beijing lineage [136] as well as in a mutant of M. bovis strain, responsible for a human outbreak in 1992 [68,69].
4. ESAT-6 from a virulence factor to diagnostic tools and vaccines for TB
Research on ESAT-6 and its involvement in several steps of Mtb pathogenesis, together with the strong antigenic recognition in TB patients, reveals its potential for therapeutic and diagnostic applications.
ESAT-6 through its duality of virulence and antigenicity, is a target for the design of more effective vaccines than BCG. The strategic design of new live attenuated vaccines should preserve Mtb antigens while removing virulence factors to prevent host damage. MTBVAC, a vaccine in development that has just entered phase 3 clinical trials (see [137]), is the only vaccine based on an attenuated Mtb strain [138]. It is conceivable that MTBVAC, by targeting epitopes from the RD1 that are missing from the BCG vaccine, could provide better protection against TB.
The attenuated virulence phenotype is based on PhoP mutants that are unable to secrete ESAT-6. The protein is synthesized but remains inside the pathogen with its antigenic potential. PhoP mutations in the attenuated vaccine strain prevent EspA translation from the espACD locus. A second deletion affects the gene required for the biosynthesis and export of phthiocerol dimycocerosates (PDIM), the major virulence-associated cell-wall lipids of Mtb. Both EspA and PDIM act together in the phagosomal secretion of ESAT-6 [105,139].
Subunit vaccine candidates are designed to boost BCG-primed responses or to induce specific immune responses as examples of the therapeutic vaccines under development to be used in conjugation with antibiotic treatment [140,141,142,143]. ESAT- 6-based subunit vaccines, already in clinical trials, include TB/FLU-04L and use a live-attenuated influenza A virus vector. Other subunit vaccines in clinical evaluation such as H6, H56:IC31 and GamTBvac are provided in non-viral delivery systems and are based on fusion immunogenic proteins, including ESAT-6 together with adjuvants [142,144,145,146].
An intranasally administered subunit vaccine combining ESAT-6 and cyclic dimeric adenosine monophosphate (c-di-AMP) was designed to promote macrophage autophagy via the STING pathway with impact on pathogen killing and humoral and cellular immune responses [147]. Other fusion proteins, such as dodicin-ESAT-6, result in the increased expression of the costimulatory molecules CD80/CD86 and the antigen presenting machinery MHC-II in a mouse model of the infection [148]. In the improvement of these combinations with ESAT-6 in subunit vaccine candidates the chaperone-like protein HtpGMtb of Mtb has been studied and structure-based design studies points for more effective antigen properties for immunization [149,150,151].
Some challenges in the development of ESAT-6-based subunit vaccines is that while CD4 T cells are maintained in the lung parenchyma due to continuous antigenic stimulation, the protective immunity is limited by functional exhaustion [152].
Another relevant application on ESAT-6 has been the development of TB diagnostics such as the IFNγ release assays (IGRAs) [153]. It was initially designed to guide preventive treatment of infected individuals at risk of developing active TB. Additionally, the ESAT-6-based IGRA allows differentiation between BCG vaccinated and unvaccinated individuals as BCG does not possess or secrete these proteins.
However, the challenges associated with subunit ESAT-6 based subunit vaccines is that they will virtually override all modern available immunodiagnostics (IGRAs and skin tests) thereby limiting the ability to distinguish immunized from infected people. To overcome this ESAT-6 free IGRA tests are under development aimed at differentiating ESAT-6 subunit responses in vaccinated individuals. These assays are based on the combination of a cocktail of proteins that are part of ESX-1 operon including the CFP-10 chaperon but excluding ESAT-6. Preliminary results show promising effects on the induction IFNγ and the chemokine biomarker IP-10 allowing distinguish subunit-vaccinated from non-vaccinated individuals [75].
An ESAT-6/CFP-10-based skin test, C-Tb, has revealed similar sensitivity for active TB compared with tuberculin skin test (TST) and QuantiFERON-TB-Gold-In-Tube (QFT), but with limited sensitivity in children and HIV-infected individuals [154]. The diagnosis of TB in children as in HIV-infected individuals is often difficult due to several factors, including the frequent extrapulmonary TB, a recurrent associated sputum smear-negative and the fact that that they have a low humoral response to mycobacterial antigens using conventional enzyme-linked immunosorbent assays (ELISA) for TB immunodiagnosis [51,155]. To overcome this, immunodiagnostic methods using antigen combinations are being developed, using “cocktails” of PhoP, ESAT-6, CFP-10 and the latency-associated antigen Acr-1. This combination of antigens includes proteins that are associated with the different stages of disease progression [156] and has been shown to significantly improve the sensitivity. Recently, another cocktail using biosynthetically derived peptides of ESAT-6 and Ag-85 allowed the detection of irrespective specific-IgG in patients’ sera, providing a reliable diagnosis of active TB in children [157].
Other situations were extrapulmonary TB is frequent is in bovine TB and the corresponding zoonotic form in humans. The available methods to distinguish M.bovis from Mtb are based in polymerase chain reaction and genomic sequencing. This imposes limitations requiring bacilli isolation difficult to obtain in this context and the lack of technical capacity in high burden countries [32].
Future directions on diagnostic tools should allow to determine differences between latent TB infection, BCG/sub-unit vaccination, and active TB infection, extrapulmonary TB with enough sensitivity to detect all cases of TB including children and HIV-infected people and MTBC species. This could revolutionize TB diagnostics and treatment strategies, allowing the development of differentiating biomarkers so relevant to evaluate the status of immune activation and/or the stage of the infection.
5. Conclusions
The studies on ESAT-6 allowed to define relevant steps in Mtb pathogenesis and to distinguish virulence from attenuated phenotypes. The lessons from this knowledge will allow us to foster our understanding of this proteinaceous army that makes Mtb such a successful human pathogen. The likelihood that this could enable us to target ESAT-6 and the host hijacking pathways involved to halt the spread of disease is a possibility for the near future. Perhaps this will open new avenues leading to the development of novel immunotherapeutic strategies to stop of TB in the 21st century.
Author Contributions
Conceptualization, E.A.; writing—original draft preparation, E.A.; writing—review and editing, E.A., D.P.; M.M and J.M.A.P.; visualization, E.A.; supervision, E.A.; All authors have read and agreed to the published version of the manuscript.
Funding
The research linked to this work was funded by Fundação para a Ciência e a Tecnologia (FCT) (grant numbers PTDC/SAU-INF/28182/2017 to E.A.; EXPL/SAU-INF/0742/2021 to D.P.; UIDB/04138/2020 to IMed-ULisboa; UIDB/04279/2020 to CIRH; and CEECINST/00070/2021 to Universidade Católica Portuguesa). M.M. is supported by a PhD fellowship from FCT with the reference 2021.07978.BD.
Acknowledgments
The authors thank Associação para o Ensino e Investigação em Microbiologia (ADEIM).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Perrin, P. Human and tuberculosis co-evolution: An integrative view. Tuberculosis 2015, 95, S112–S116. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.G. Commentary: Medicine, population, and tuberculosis. Int. J. Epidemiol. 2005, 34, 521–524. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Tuberculosis Report 2022 Factsheet. Available online: https://www.who.int/publications/m/item/global-tuberculosis-report-2022-factsheet (accessed on 20 April 2023).
- Chen, X.; Hu, T.Y. Strategies for advanced personalized tuberculosis diagnosis: Current technologies and clinical approaches. Precision Clinical Medicine 2021, 4, 35–44. [Google Scholar] [CrossRef]
- Behr, M.A.; Kaufmann, E.; Duffin, J.; Edelstein, P.H.; Ramakrishnan, L. Latent Tuberculosis: Two Centuries of Confusion. American Journal of Respiratory and Critical Care Medicine 2021, 204, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Carranza, C.; Pedraza-Sanchez, S.; de Oyarzabal-Mendez, E.; Torres, M. Diagnosis for Latent Tuberculosis Infection: New Alternatives. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Dheda, K.; Barry, C.E., 3rd; Maartens, G. Tuberculosis. The Lancet 2016, 387, 1211–1226. [Google Scholar] [CrossRef]
- Dheda, K.; Gumbo, T.; Maartens, G.; Dooley, K.E.; McNerney, R.; Murray, M.; Furin, J.; Nardell, E.A.; London, L.; Lessem, E.; et al. The epidemiology, pathogenesis, transmission, diagnosis, and management of multidrug-resistant, extensively drug-resistant, and incurable tuberculosis. The Lancet Respiratory Medicine 2017, 5, 291–360. [Google Scholar] [CrossRef]
- Cambier, C.J.; Falkow, S.; Ramakrishnan, L. Host evasion and exploitation schemes of Mycobacterium tuberculosis. Cell 2014, 159, 1497–1509. [Google Scholar] [CrossRef]
- Azevedo-Pereira, J.M.; Pires, D.; Calado, M.; Mandal, M.; Santos-Costa, Q.; Anes, E. HIV/Mtb Co-Infection: From the Amplification of Disease Pathogenesis to an “Emerging Syndemic”. Microorganisms 2023, 11. [Google Scholar] [CrossRef]
- Barry, C.E.; Boshoff, H.I.; Dartois, V.; Dick, T.; Ehrt, S.; Flynn, J.; Schnappinger, D.; Wilkinson, R.J.; Young, D. The spectrum of latent tuberculosis: rethinking the biology and intervention strategies. Nature Reviews Microbiology 2009, 7, 845–855. [Google Scholar] [CrossRef]
- Blaser, M.J.; Kirschner, D. The equilibria that allow bacterial persistence in human hosts. Nature 2007, 449, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.T.; Smith, E.G.; Banerjee, A.; Smith, R.M.M.; Dale, J.; Innes, J.A.; Hunt, D.; Tweddell, A.; Wood, A.; Anderson, C.; et al. Cluster of human tuberculosis caused by Mycobacterium bovis: evidence for person-to-person transmission in the UK. The Lancet 2007, 369, 1270–1276. [Google Scholar] [CrossRef]
- Grange, J.M. Mycobacterium bovis infection in human beings. Tuberculosis 2001, 81, 71–77. [Google Scholar] [CrossRef]
- Thoen, C.O.; LoBue, P.A. Mycobacterium bovis tuberculosis: forgotten, but not gone. The Lancet 2007, 369, 1236–1238. [Google Scholar] [CrossRef] [PubMed]
- Torres-Gonzalez, P.; Cervera-Hernandez, M.E.; Martinez-Gamboa, A.; Garcia-Garcia, L.; Cruz-Hervert, L.P.; Bobadilla-del Valle, M.; Ponce-de Leon, A.; Sifuentes-Osornio, J. Human tuberculosis caused by Mycobacterium bovis: a retrospective comparison with Mycobacterium tuberculosis in a Mexican tertiary care centre, 2000–2015. BMC Infect. Dis. 2016, 16, 657. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, E.G.; Griffiths, R.J.; Roberts, C. Pulmonary tuberculosis due to Mycobacterium bovis. Thorax 1986, 41, 685. [Google Scholar] [CrossRef] [PubMed]
- de Jong, B.C.; Hill, P.C.; Aiken, A.; Awine, T.; Martin, A.; Adetifa, I.M.; Jackson-Sillah, D.J.; Fox, A.; Kathryn, D.; Gagneux, S.; et al. Progression to Active Tuberculosis, but Not Transmission, Varies by Mycobacterium tuberculosis Lineage in The Gambia. The Journal of Infectious Diseases 2008, 198, 1037–1043. [Google Scholar] [CrossRef]
- Mostowy, S.; Onipede, A.; Gagneux, S.; Niemann, S.; Kremer, K.; Desmond Edward, P.; Kato-Maeda, M.; Behr, M. Genomic Analysis Distinguishes Mycobacterium africanum. J. Clin. Microbiol. 2004, 42, 3594–3599. [Google Scholar] [CrossRef]
- Silva, M.L.; Cá, B.; Osório, N.S.; Rodrigues, P.N.S.; Maceiras, A.R.; Saraiva, M. Tuberculosis caused by Mycobacterium africanum: Knowns and unknowns. PLOS Pathogens 2022, 18, e1010490. [Google Scholar] [CrossRef]
- de Jong, B.C.; Antonio, M.; Gagneux, S. Mycobacterium africanum—Review of an Important Cause of Human Tuberculosis in West Africa. PLoS Negl. Trop. Dis. 2010, 4, e744. [Google Scholar] [CrossRef]
- Coscolla, M.; Gagneux, S.; Menardo, F.; Loiseau, C.; Ruiz-Rodriguez, P.; Borrell, S.; Otchere, I.D.; Asante-Poku, A.; Asare, P.; Sánchez-Busó, L.; et al. Phylogenomics of Mycobacterium africanum reveals a new lineage and a complex evolutionary history. Microbial Genomics 2021, 7. [Google Scholar] [CrossRef]
- Gagneux, S. Ecology and evolution of Mycobacterium tuberculosis. Nature Reviews Microbiology 2018, 16, 202–213. [Google Scholar] [CrossRef]
- Müller, B.; Dürr, S.; Alonso, S.; Hattendorf, J.; Laisse, C.J.M.; Parsons, S.D.C.; van Helden, P.; Zinsstag, J. Zoonotic Mycobacterium bovis–induced Tuberculosis in Humans. Emerging Infectious Disease journal 2013, 19, 899. [Google Scholar] [CrossRef]
- Borham, M.; Oreiby, A.; El-Gedawy, A.; Hegazy, Y.; Khalifa, H.O.; Al-Gaabary, M.; Matsumoto, T. Review on Bovine Tuberculosis: An Emerging Disease Associated with Multidrug-Resistant Mycobacterium Species. Pathogens 2022, 11. [Google Scholar] [CrossRef]
- Prodinger, W.M.; Indra, A.; Koksalan, O.K.; Kilicaslan, Z.; Richter, E. Mycobacterium caprae infection in humans. Expert Rev. Anti Infect. Ther. 2014, 12, 1501–1513. [Google Scholar] [CrossRef]
- Tagliapietra, V.; Boniotti, M.B.; Mangeli, A.; Karaman, I.; Alborali, G.; Chiari, M.; D’Incau, M.; Zanoni, M.; Rizzoli, A.; Pacciarini, M.L. Mycobacterium microti at the Environment and Wildlife Interface. Microorganisms 2021, 9, 2084. [Google Scholar] [CrossRef] [PubMed]
- Fabre, M.; Hauck, Y.; Soler, C.; Koeck, J.-L.; van Ingen, J.; van Soolingen, D.; Vergnaud, G.; Pourcel, C. Molecular characteristics of “Mycobacterium canettii” the smooth Mycobacterium tuberculosis bacilli. Infection, Genetics and Evolution 2010, 10, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo-Asensio, J.; Malaga, W.; Pawlik, A.; Astarie-Dequeker, C.; Passemar, C.; Moreau, F.; Laval, F.; Daffé, M.; Martin, C.; Brosch, R.; et al. Evolutionary history of tuberculosis shaped by conserved mutations in the PhoPR virulence regulator. Proceedings of the National Academy of Sciences 2014, 111, 11491–11496. [Google Scholar] [CrossRef] [PubMed]
- Riojas, M.A.; McGough, K.J.; Rider-Riojas, C.J.; Rastogi, N.; Hazbón, M.H. Phylogenomic analysis of the species of the Mycobacterium tuberculosis complex demonstrates that Mycobacterium africanum, Mycobacterium bovis, Mycobacterium caprae, Mycobacterium microti and Mycobacterium pinnipedii are later heterotypic synonyms of Mycobacterium tuberculosis. International Journal of Systematic and Evolutionary Microbiology 2018, 68, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Lombard, J.E.; Patton, E.A.; Gibbons-Burgener, S.N.; Klos, R.F.; Tans-Kersten, J.L.; Carlson, B.W.; Keller, S.J.; Pritschet, D.J.; Rollo, S.; Dutcher, T.V.; et al. Human-to-Cattle Mycobacterium tuberculosis Complex Transmission in the United States. Frontiers in Veterinary Science 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- WHO. Roadmap for zoonotic tuberculosis. Available online: https://www.who.int/publications/i/item/9789241513043 (accessed on 29 May 2023).
- Wolfe, N.D.; Dunavan, C.P.; Diamond, J. Origins of major human infectious diseases. Nature 2007, 447, 279–283. [Google Scholar] [CrossRef]
- Comas, I.; Coscolla, M.; Luo, T.; Borrell, S.; Holt, K.E.; Kato-Maeda, M.; Parkhill, J.; Malla, B.; Berg, S.; Thwaites, G.; et al. Out-of-Africa migration and Neolithic coexpansion of Mycobacterium tuberculosis with modern humans. Nat. Genet. 2013, 45, 1176–1182. [Google Scholar] [CrossRef]
- Urbanowski, M.E.; Ordonez, A.A.; Ruiz-Bedoya, C.A.; Jain, S.K.; Bishai, W.R. Cavitary tuberculosis: the gateway of disease transmission. The Lancet Infectious Diseases 2020, 20, e117–e128. [Google Scholar] [CrossRef] [PubMed]
- Brites, D.; Gagneux, S. Old and new selective pressures on Mycobacterium tuberculosis. Infection, Genetics and Evolution 2012, 12, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Alteri, C.J.; Rios-Sarabia, N.; De la Cruz, M.A.; González-y-Merchand, J.A.; Soria-Bustos, J.; Maldonado-Bernal, C.; Cedillo, M.L.; Yáñez-Santos, J.A.; Martínez-Laguna, Y.; Torres, J.; et al. The Flp type IV pilus operon of Mycobacterium tuberculosis is expressed upon interaction with macrophages and alveolar epithelial cells. Frontiers in Cellular and Infection Microbiology 2022, 12. [Google Scholar] [CrossRef]
- Ramsugit, S.; Pillay, M. Pili of Mycobacterium tuberculosis: current knowledge and future prospects. Arch. Microbiol. 2015, 197, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Kalscheuer, R.; Palacios, A.; Anso, I.; Cifuente, J.; Anguita, J.; Jacobs, W.R., Jr.; Guerin, M.E.; Prados-Rosales, R. The Mycobacterium tuberculosis capsule: a cell structure with key implications in pathogenesis. Biochem. J. 2019, 476, 1995–2016. [Google Scholar] [CrossRef] [PubMed]
- Anes, E.; Pires, D.; Mandal, M.; Azevedo-Pereira, J.M. Spatial localization of cathepsins: Implications in immune activation and resolution during infections. Front. Immunol. 2022, 13, 955407. [Google Scholar] [CrossRef]
- Armstrong, J.A.; Hart, P.D. Phagosome-lysosome interactions in cultured macrophages infected with virulent tubercle bacilli. Reversal of the usual nonfusion pattern and observations on bacterial survival. J. Exp. Med. 1975, 142, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.; Bernard, E.M.; Pombo, J.P.; Carmo, N.; Fialho, C.; Gutierrez, M.G.; Bettencourt, P.; Anes, E. Mycobacterium tuberculosis Modulates miR-106b-5p to Control Cathepsin S Expression Resulting in Higher Pathogen Survival and Poor T-Cell Activation. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Pires, D.; Calado, M.; Velez, T.; Mandal, M.; Catalão, M.J.; Neyrolles, O.; Lugo-Villarino, G.; Vérollet, C.; Azevedo-Pereira, J.M.; Anes, E. Modulation of Cystatin C in Human Macrophages Improves Anti-Mycobacterial Immune Responses to Mycobacterium tuberculosis Infection and Coinfection With HIV. Front. Immunol. 2021, 12, 4693. [Google Scholar] [CrossRef]
- Pires, D.; Mandal, M.; Pinho, J.; Catalão, M.J.; Almeida, A.J.; Azevedo-Pereira, J.M.; Gaspar, M.M.; Anes, E. Liposomal Delivery of Saquinavir to Macrophages Overcomes Cathepsin Blockade by Mycobacterium tuberculosis and Helps Control the Phagosomal Replicative Niches. Int. J. Mol. Sci. 2023, 24. [Google Scholar] [CrossRef]
- Pires, D.; Marques, J.; Pombo, J.P.; Carmo, N.; Bettencourt, P.; Neyrolles, O.; Lugo-Villarino, G.; Anes, E. Role of Cathepsins in Mycobacterium tuberculosis Survival in Human Macrophages. Sci. Rep. 2016, 6, 32247. [Google Scholar] [CrossRef]
- Cohen, S.B.; Gern, B.H.; Delahaye, J.L.; Adams, K.N.; Plumlee, C.R.; Winkler, J.K.; Sherman, D.R.; Gerner, M.Y.; Urdahl, K.B. Alveolar Macrophages Provide an Early Mycobacterium tuberculosis Niche and Initiate Dissemination. Cell Host & Microbe 2018, 24, 439–446.e434. [Google Scholar] [CrossRef]
- Mishra, B.B.; Moura-Alves, P.; Sonawane, A.; Hacohen, N.; Griffiths, G.; Moita, L.F.; Anes, E. Mycobacterium tuberculosis protein ESAT-6 is a potent activator of the NLRP3/ASC inflammasome. Cell. Microbiol. 2010, 12, 1046–1063. [Google Scholar] [CrossRef] [PubMed]
- Aguilo, J.I.; Alonso, H.; Uranga, S.; Marinova, D.; Arbués, A.; de Martino, A.; Anel, A.; Monzon, M.; Badiola, J.; Pardo, J.; et al. ESX-1-induced apoptosis is involved in cell-to-cell spread of Mycobacterium tuberculosis. Cell. Microbiol. 2013, 15, 1994–2005. [Google Scholar] [CrossRef] [PubMed]
- Dallenga, T.; Repnik, U.; Corleis, B.; Eich, J.; Reimer, R.; Griffiths, G.W.; Schaible, U.E. M. tuberculosis-Induced Necrosis of Infected Neutrophils Promotes Bacterial Growth Following Phagocytosis by Macrophages. Cell Host & Microbe 2017, 22, 519–530.e513. [Google Scholar] [CrossRef]
- Derrick, S.C.; Morris, S.L. The ESAT6 protein of Mycobacterium tuberculosis induces apoptosis of macrophages by activating caspase expression. Cell. Microbiol. 2007, 9, 1547–1555. [Google Scholar] [CrossRef] [PubMed]
- Anes, E.; Azevedo-Pereira, J.M.; Pires, D. Cathepsins and Their Endogenous Inhibitors in Host Defense During Mycobacterium tuberculosis and HIV Infection. Front. Immunol. 2021, 12, 726984. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.M.; Ramakrishnan, L. The Role of the Granuloma in Expansion and Dissemination of Early Tuberculous Infection. Cell 2009, 136, 37–49. [Google Scholar] [CrossRef]
- Pagán, A.J.; Ramakrishnan, L. Immunity and Immunopathology in the Tuberculous Granuloma. Cold Spring Harb. Perspect. Med. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.J.; Linas, B.; Trevejo-Nuñez, G.J.; Kincaid, E.; Tamura, T.; Takatsu, K.; Ernst, J.D. Mycobacterium tuberculosis Infects Dendritic Cells with High Frequency and Impairs Their Function In Vivo1. The Journal of Immunology 2007, 179, 2509–2519. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.B.; Rathinam, V.A.K.; Martens, G.W.; Martinot, A.J.; Kornfeld, H.; Fitzgerald, K.A.; Sassetti, C.M. Nitric oxide controls the immunopathology of tuberculosis by inhibiting NLRP3 inflammasome–dependent processing of IL-1β. Nat. Immunol. 2013, 14, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.-W.; Jacobs Jr, W.R. Critical role for NLRP3 in necrotic death triggered by Mycobacterium tuberculosis. Cell. Microbiol. 2011, 13, 1371–1384. [Google Scholar] [CrossRef] [PubMed]
- Brosch, R.; Gordon, S.V.; Marmiesse, M.; Brodin, P.; Buchrieser, C.; Eiglmeier, K.; Garnier, T.; Gutierrez, C.; Hewinson, G.; Kremer, K.; et al. A new evolutionary scenario for the Mycobacterium tuberculosis complex. Proceedings of the National Academy of Sciences 2002, 99, 3684–3689. [Google Scholar] [CrossRef] [PubMed]
- Brosch, R.; Gordon, S.V.; Garnier, T.; Eiglmeier, K.; Frigui, W.; Valenti, P.; Dos Santos, S.; Duthoy, S.; Lacroix, C.; Garcia-Pelayo, C.; et al. Genome plasticity of BCG and impact on vaccine efficacy. Proceedings of the National Academy of Sciences 2007, 104, 5596–5601. [Google Scholar] [CrossRef]
- Cole, S.T.; Brosch, R.; Parkhill, J.; Garnier, T.; Churcher, C.; Harris, D.; Gordon, S.V.; Eiglmeier, K.; Gas, S.; Barry, C.E.; et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 1998, 393, 537–544. [Google Scholar] [CrossRef]
- Marinova, D.; Gonzalo-Asensio, J.; Aguilo, N.; Martin, C. MTBVAC from discovery to clinical trials in tuberculosis-endemic countries. Expert Review of Vaccines 2017, 16, 565–576. [Google Scholar] [CrossRef]
- Smith, N.H.; Hewinson, R.G.; Kremer, K.; Brosch, R.; Gordon, S.V. Myths and misconceptions: the origin and evolution of Mycobacterium tuberculosis. Nature Reviews Microbiology 2009, 7, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Broset, E.; Martín, C.; Gonzalo-Asensio, J. Evolutionary Landscape of the Mycobacterium tuberculosis Complex from the Viewpoint of PhoPR: Implications for Virulence Regulation and Application to Vaccine Development. mBio 2015, 6, e01289–e01215. [Google Scholar] [CrossRef] [PubMed]
- Frigui, W.; Bottai, D.; Majlessi, L.; Monot, M.; Josselin, E.; Brodin, P.; Garnier, T.; Gicquel, B.; Martin, C.; Leclerc, C.; et al. Control of M. tuberculosis ESAT-6 Secretion and Specific T Cell Recognition by PhoP. PLOS Pathogens 2008, 4, e33. [Google Scholar] [CrossRef]
- Pérez, E.; Samper, S.; Bordas, Y.; Guilhot, C.; Gicquel, B.; Martín, C. An essential role for phoP in Mycobacterium tuberculosis virulence. Mol. Microbiol. 2001, 41, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Mahairas, G.G.; Sabo, P.J.; Hickey, M.J.; Singh, D.C.; Stover, C.K. Molecular analysis of genetic differences between Mycobacterium bovis BCG and virulent M. bovis. J. Bacteriol. 1996, 178, 1274–1282. [Google Scholar] [CrossRef] [PubMed]
- Pym, A.S.; Brodin, P.; Brosch, R.; Huerre, M.; Cole, S.T. Loss of RD1 contributed to the attenuation of the live tuberculosis vaccines Mycobacterium bovis BCG and Mycobacterium microti. Mol. Microbiol. 2002, 46, 709–717. [Google Scholar] [CrossRef]
- de Jong, B.C.; Hill, P.C.; Brookes, R.H.; Gagneux, S.; Jeffries, D.J.; Otu, J.K.; Donkor, S.A.; Fox, A.; McAdam, K.P.W.J.; Small, P.M.; et al. Mycobacterium africanum Elicits an Attenuated T Cell Response to Early Secreted Antigenic Target, 6 kDa, in Patients with Tuberculosis and Their Household Contacts. The Journal of Infectious Diseases 2006, 193, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo-Asensio, J.; Pérez, I.; Aguiló, N.; Uranga, S.; Picó, A.; Lampreave, C.; Cebollada, A.; Otal, I.; Samper, S.; Martín, C. New insights into the transposition mechanisms of IS6110 and its dynamic distribution between Mycobacterium tuberculosis Complex lineages. PLoS Genet. 2018, 14, e1007282. [Google Scholar] [CrossRef] [PubMed]
- Soto, C.Y.; Menendez, M.C.; Perez, E.; Samper, S.; Gomez, A.B.; Garcia, M.J.; Martin, C. IS6110 Mediates Increased Transcription of the phoP Virulence Gene in a Multidrug-Resistant Clinical Isolate Responsible for Tuberculosis Outbreaks. J. Clin. Microbiol. 2004, 42, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, A.M.; Gey van Pittius, N.C.; DiGiuseppe Champion, P.A.; Cox, J.; Luirink, J.; Vandenbroucke-Grauls, C.M.J.E.; Appelmelk, B.J.; Bitter, W. Type VII secretion — mycobacteria show the way. Nature Reviews Microbiology 2007, 5, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Gröschel, M.I.; Sayes, F.; Simeone, R.; Majlessi, L.; Brosch, R. ESX secretion systems: mycobacterial evolution to counter host immunity. Nature Reviews Microbiology 2016, 14, 677–691. [Google Scholar] [CrossRef]
- Andersen, P.; Andersen, A.B.; Sørensen, A.L.; Nagai, S. Recall of long-lived immunity to Mycobacterium tuberculosis infection in mice. The Journal of Immunology 1995, 154, 3359–3372. [Google Scholar] [CrossRef]
- Sørensen, A.L.; Nagai, S.; Houen, G.; Andersen, P.; Andersen, A.B. Purification and characterization of a low-molecular-mass T-cell antigen secreted by Mycobacterium tuberculosis. Infection and Immunity 1995, 63, 1710–1717. [Google Scholar] [CrossRef]
- Brandt, L.; Elhay, M.; Rosenkrands, I.; Lindblad Erik, B.; Andersen, P. ESAT-6 Subunit Vaccination against Mycobacterium tuberculosis. Infection and Immunity 2000, 68, 791–795. [Google Scholar] [CrossRef]
- Ruhwald, M.; de Thurah, L.; Kuchaka, D.; Zaher, M.R.; Salman, A.M.; Abdel-Ghaffar, A.-R.; Shoukry, F.A.; Michelsen, S.W.; Soborg, B.; Blauenfeldt, T.; et al. Introducing the ESAT-6 free IGRA, a companion diagnostic for TB vaccines based on ESAT-6. Sci. Rep. 2017, 7, 45969. [Google Scholar] [CrossRef] [PubMed]
- Brodin, P.; Rosenkrands, I.; Andersen, P.; Cole, S.T.; Brosch, R. ESAT-6 proteins: protective antigens and virulence factors? Trends Microbiol. 2004, 12, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; McIntosh, F.; Radomski, N.; Dewar, K.; Simeone, R.; Enninga, J.; Brosch, R.; Rocha, E.P.; Veyrier, F.J.; Behr, M.A. Insights on the Emergence of Mycobacterium tuberculosis from the Analysis of Mycobacterium kansasii. Genome Biol. Evol. 2015, 7, 856–870. [Google Scholar] [CrossRef] [PubMed]
- Wirth, S.E.; Krywy, J.A.; Aldridge, B.B.; Fortune, S.M.; Fernandez-Suarez, M.; Gray, T.A.; Derbyshire, K.M. Polar assembly and scaffolding proteins of the virulence-associated ESX-1 secretory apparatus in mycobacteria. Mol. Microbiol. 2012, 83, 654–664. [Google Scholar] [CrossRef] [PubMed]
- Anes, E.; Peyron, P.; Staali, L.; Jordao, L.; Gutierrez, M.G.; Kress, H.; Hagedorn, M.; Maridonneau-Parini, I.; Skinner, M.A.; Wildeman, A.G.; et al. Dynamic life and death interactions between Mycobacterium smegmatis and J774 macrophages. Cell. Microbiol. 2006, 8, 939–960. [Google Scholar] [CrossRef] [PubMed]
- Coros, A.; Callahan, B.; Battaglioli, E.; Derbyshire, K.M. The specialized secretory apparatus ESX-1 is essential for DNA transfer in Mycobacterium smegmatis. Mol. Microbiol. 2008, 69, 794–808. [Google Scholar] [CrossRef] [PubMed]
- Flint, J.L.; Kowalski, J.C.; Karnati, P.K.; Derbyshire, K.M. The RD1 virulence locus of Mycobacterium tuberculosis regulates DNA transfer in Mycobacterium smegmatis. Proceedings of the National Academy of Sciences 2004, 101, 12598–12603. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Becq, J.; Gicquel, B.; Deschavanne, P.; Neyrolles, O. Horizontally acquired genomic islands in the tubercle bacilli. Trends Microbiol. 2008, 16, 303–308. [Google Scholar] [CrossRef]
- Ates, L.S.; Brosch, R. Discovery of the type VII ESX-1 secretion needle? Mol. Microbiol. 2017, 103, 7–12. [Google Scholar] [CrossRef]
- Boritsch, E.C.; Supply, P.; Honoré, N.; Seeman, T.; Stinear, T.P.; Brosch, R. A glimpse into the past and predictions for the future: the molecular evolution of the tuberculosis agent. Mol. Microbiol. 2014, 93, 835–852. [Google Scholar] [CrossRef] [PubMed]
- MacGurn, J.A.; Raghavan, S.; Stanley, S.A.; Cox, J.S. A non-RD1 gene cluster is required for Snm secretion in Mycobacterium tuberculosis. Mol. Microbiol. 2005, 57, 1653–1663. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, F.; Brosch, R. ESX/Type VII Secretion Systems—An Important Way Out for Mycobacterial Proteins. Microbiology Spectrum 2019, 7, 7–4. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.-Y.; Guo, S.; McLaughlin, B.; Morisaki, H.; Engel, J.N.; Brown, E.J. A mycobacterial virulence gene cluster extending RD1 is required for cytolysis, bacterial spreading and ESAT-6 secretion. Mol. Microbiol. 2004, 53, 1677–1693. [Google Scholar] [CrossRef]
- Guinn, K.M.; Hickey, M.J.; Mathur, S.K.; Zakel, K.L.; Grotzke, J.E.; Lewinsohn, D.M.; Smith, S.; Sherman, D.R. Individual RD1-region genes are required for export of ESAT-6/CFP-10 and for virulence of Mycobacterium tuberculosis. Mol. Microbiol. 2004, 51, 359–370. [Google Scholar] [CrossRef]
- Hsu, T.; Hingley-Wilson, S.M.; Chen, B.; Chen, M.; Dai, A.Z.; Morin, P.M.; Marks, C.B.; Padiyar, J.; Goulding, C.; Gingery, M.; et al. The primary mechanism of attenuation of bacillus Calmette–Guérin is a loss of secreted lytic function required for invasion of lung interstitial tissue. Proceedings of the National Academy of Sciences 2003, 100, 12420–12425. [Google Scholar] [CrossRef]
- Lewis, K.N.; Liao, R.; Guinn, K.M.; Hickey, M.J.; Smith, S.; Behr, M.A.; Sherman, D.R. Deletion of RD1 from Mycobacterium tuberculosis Mimics Bacille Calmette-Guérin Attenuation. The Journal of Infectious Diseases 2003, 187, 117–123. [Google Scholar] [CrossRef]
- Sassetti, C.M.; Rubin, E.J. Genetic requirements for mycobacterial survival during infection. Proceedings of the National Academy of Sciences 2003, 100, 12989–12994. [Google Scholar] [CrossRef]
- Stanley, S.A.; Raghavan, S.; Hwang, W.W.; Cox, J.S. Acute infection and macrophage subversion by Mycobacterium tuberculosis require a specialized secretion system. Proceedings of the National Academy of Sciences 2003, 100, 13001–13006. [Google Scholar] [CrossRef]
- Simeone, R.; Bottai, D.; Frigui, W.; Majlessi, L.; Brosch, R. ESX/type VII secretion systems of mycobacteria: Insights into evolution, pathogenicity and protection. Tuberculosis 2015, 95, S150–S154. [Google Scholar] [CrossRef]
- Steenken, W., Jr.; Oatway, W.H., Jr.; Petroff, S.A. Biological Studies of the Tubercle Bacillus : Iii. Dissociation and Pathogenicity of the R and S Variants of the Human Tubercle Bacillus (H(37)). J. Exp. Med. 1934, 60, 515–540. [Google Scholar] [CrossRef]
- Solans, L.; Aguiló, N.; Samper, S.; Pawlik, A.; Frigui, W.; Martín, C.; Brosch, R.; Gonzalo-Asensio, J. A Specific Polymorphism in Mycobacterium tuberculosis H37Rv Causes Differential ESAT-6 Expression and Identifies WhiB6 as a Novel ESX-1 Component. Infection and Immunity 2014, 82, 3446–3456. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.; Mandal, M.; Matos, A.I.; Peres, C.; Catalão, M.J.; Azevedo-Pereira, J.M.; Satchi-Fainaro, R.; Florindo, H.F.; Anes, E. Development of Chitosan Particles Loaded with siRNA for Cystatin C to Control Intracellular Drug-Resistant Mycobacterium tuberculosis. Antibiotics 2023, 12. [Google Scholar] [CrossRef]
- Pires, D.; Valente, S.; Calado, M.; Mandal, M.; Azevedo-Pereira, J.M.; Anes, E. Repurposing Saquinavir for Host-Directed Therapy to Control Mycobacterium tuberculosis Infection. Front. Immunol. 2021, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Behura, A.; Das, M.; Kumar, A.; Naik, L.; Mishra, A.; Manna, D.; Patel, S.; Mishra, A.; Singh, R.; Dhiman, R. ESAT-6 impedes IL-18 mediated phagosome lysosome fusion via microRNA-30a upon Calcimycin treatment in mycobacteria infected macrophages. Int. Immunopharmacol. 2021, 101, 108319. [Google Scholar] [CrossRef]
- Bettencourt, P.; Pires, D.; Anes, E. Immunomodulating microRNAs of mycobacterial infections. Tuberculosis (Edinburgh, Scotland) 2016, 97, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.; Lee, W.L.; Alexander, D.C.; Grinstein, S.; Liu, J. The ESAT-6/CFP-10 secretion system of Mycobacterium marinum modulates phagosome maturation. Cell. Microbiol. 2006, 8, 1417–1429. [Google Scholar] [CrossRef]
- Simeone, R.; Bobard, A.; Lippmann, J.; Bitter, W.; Majlessi, L.; Brosch, R.; Enninga, J. Phagosomal Rupture by Mycobacterium tuberculosis Results in Toxicity and Host Cell Death. PLOS Pathogens 2012, 8, e1002507. [Google Scholar] [CrossRef]
- van der Wel, N.; Hava, D.; Houben, D.; Fluitsma, D.; van Zon, M.; Pierson, J.; Brenner, M.; Peters, P.J. M. tuberculosis and M. leprae Translocate from the Phagolysosome to the Cytosol in Myeloid Cells. Cell 2007, 129, 1287–1298. [Google Scholar] [CrossRef]
- Osman, M.M.; Pagán, A.J.; Shanahan, J.K.; Ramakrishnan, L. Mycobacterium marinum phthiocerol dimycocerosates enhance macrophage phagosomal permeabilization and membrane damage. PLoS One 2020, 15, e0233252. [Google Scholar] [CrossRef]
- Osman, M.M.; Shanahan, J.K.; Chu, F.; Takaki, K.K.; Pinckert, M.L.; Pagán, A.J.; Brosch, R.; Conrad, W.H.; Ramakrishnan, L. The C terminus of the mycobacterium ESX-1 secretion system substrate ESAT-6 is required for phagosomal membrane damage and virulence. Proceedings of the National Academy of Sciences 2022, 119, e2122161119. [Google Scholar] [CrossRef]
- Quigley, J.; Hughitt, V.K.; Velikovsky Carlos, A.; Mariuzza Roy, A.; El-Sayed Najib, M.; Briken, V. The Cell Wall Lipid PDIM Contributes to Phagosomal Escape and Host Cell Exit of Mycobacterium tuberculosis. mBio 2017, 8, e00148–00117. [Google Scholar] [CrossRef]
- Mishra, B.B.; Lovewell, R.R.; Olive, A.J.; Zhang, G.; Wang, W.; Eugenin, E.; Smith, C.M.; Phuah, J.Y.; Long, J.E.; Dubuke, M.L.; et al. Nitric oxide prevents a pathogen-permissive granulocytic inflammation during tuberculosis. Nature Microbiology 2017, 2, 17072. [Google Scholar] [CrossRef]
- Elkington, P.T.; Green, J.A.; Emerson, J.E.; Lopez-Pascua, L.D.; Boyle, J.J.; O'Kane, C.M.; Friedland, J.S. Synergistic up-regulation of epithelial cell matrix metalloproteinase-9 secretion in tuberculosis. Am. J. Respir. Cell Mol. Biol. 2007, 37, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Volkman, H.E.; Pozos, T.C.; Zheng, J.; Davis, J.M.; Rawls, J.F.; Ramakrishnan, L. Tuberculous Granuloma Induction via Interaction of a Bacterial Secreted Protein with Host Epithelium. Science 2010, 327, 466–469. [Google Scholar] [CrossRef]
- Ha, S.-H.; Choi, H.; Park, J.-Y.; Abekura, F.; Lee, Y.-C.; Kim, J.-R.; Kim, C.-H. Mycobacterium tuberculosis–Secreted Protein, ESAT-6, Inhibits Lipopolysaccharide-Induced MMP-9 Expression and Inflammation Through NF-κB and MAPK Signaling in RAW 264.7 Macrophage Cells. Inflammation 2020, 43, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Augenstreich, J.; Arbues, A.; Simeone, R.; Haanappel, E.; Wegener, A.; Sayes, F.; Le Chevalier, F.; Chalut, C.; Malaga, W.; Guilhot, C.; et al. ESX-1 and phthiocerol dimycocerosates of Mycobacterium tuberculosis act in concert to cause phagosomal rupture and host cell apoptosis. Cell. Microbiol. 2017, 19, e12726. [Google Scholar] [CrossRef] [PubMed]
- Refai, A.; Gritli, S.; Barbouche, M.-R.; Essafi, M. Mycobacterium tuberculosis Virulent Factor ESAT-6 Drives Macrophage Differentiation Toward the Pro-inflammatory M1 Phenotype and Subsequently Switches It to the Anti-inflammatory M2 Phenotype. Frontiers in Cellular and Infection Microbiology 2018, 8. [Google Scholar] [CrossRef]
- Li, F.; Luo, J.; Xu, H.; Wang, Y.; Jiang, W.; Chang, K.; Deng, S.; Chen, M. Early secreted antigenic target 6-kDa from Mycobacterium tuberculosis enhanced the protective innate immunity of macrophages partially via HIF1α. Biochemical and Biophysical Research Communications 2020, 522, 26–32. [Google Scholar] [CrossRef]
- Watson, Robert O. ; Bell, Samantha L.; MacDuff, Donna A.; Kimmey, Jacqueline M.; Diner, Elie J.; Olivas, J.; Vance, Russell E.; Stallings, Christina L.; Virgin, Herbert W.; Cox, Jeffery S. The Cytosolic Sensor cGAS Detects Mycobacterium tuberculosis DNA to Induce Type I Interferons and Activate Autophagy. Cell Host & Microbe 2015, 17, 811–819. [Google Scholar] [CrossRef]
- Grover, A.; Izzo, A.A. BAT3 Regulates Mycobacterium tuberculosis Protein ESAT-6-Mediated Apoptosis of Macrophages. PLoS One 2012, 7, e40836. [Google Scholar] [CrossRef] [PubMed]
- Welin, A.; Eklund, D.; Stendahl, O.; Lerm, M. Human Macrophages Infected with a High Burden of ESAT-6-Expressing M. tuberculosis Undergo Caspase-1- and Cathepsin B-Independent Necrosis. PLoS One 2011, 6, e20302. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.G.; Master, S.S.; Singh, S.B.; Taylor, G.A.; Colombo, M.I.; Deretic, V. Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. Cell 2004, 119, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Wassermann, R.; Gulen Muhammet, F.; Sala, C.; Perin, Sonia G.; Lou, Y.; Rybniker, J.; Schmid-Burgk, Jonathan L.; Schmidt, T.; Hornung, V.; Cole, Stewart T.; et al. Mycobacterium tuberculosis Differentially Activates cGAS- and Inflammasome-Dependent Intracellular Immune Responses through ESX-1. Cell Host & Microbe 2015, 17, 799–810. [Google Scholar] [CrossRef]
- Yabaji, S.M.; Dhamija, E.; Mishra, A.K.; Srivastava, K.K. ESAT-6 regulates autophagous response through SOD-2 and as a result induces intracellular survival of Mycobacterium bovis BCG. Biochim. Biophys. Acta 2020, 1868, 140470. [Google Scholar] [CrossRef]
- Majlessi, L.; Brosch, R. Mycobacterium tuberculosis Meets the Cytosol: The Role of cGAS in Anti-mycobacterial Immunity. Cell Host & Microbe 2015, 17, 733–735. [Google Scholar] [CrossRef]
- Shafiani, S.; Dinh, C.; Ertelt James, M.; Moguche, Albanus O.; Siddiqui, I.; Smigiel, Kate S.; Sharma, P.; Campbell, Daniel J.; Way, Sing S.; Urdahl, Kevin B. Pathogen-Specific Treg Cells Expand Early during Mycobacterium tuberculosis Infection but Are Later Eliminated in Response to Interleukin-12. Immunity 2013, 38, 1261–1270. [Google Scholar] [CrossRef]
- Banaiee, N.; Kincaid, E.Z.; Buchwald, U.; Jacobs, W.R., Jr.; Ernst, J.D. Potent Inhibition of Macrophage Responses to IFN-γ by Live Virulent Mycobacterium tuberculosis Is Independent of Mature Mycobacterial Lipoproteins but Dependent on TLR21. The Journal of Immunology 2006, 176, 3019–3027. [Google Scholar] [CrossRef]
- Sreejit, G.; Ahmed, A.; Parveen, N.; Jha, V.; Valluri, V.L.; Ghosh, S.; Mukhopadhyay, S. The ESAT-6 Protein of Mycobacterium tuberculosis Interacts with Beta-2-Microglobulin (β2M) Affecting Antigen Presentation Function of Macrophage. PLOS Pathogens 2014, 10, e1004446. [Google Scholar] [CrossRef]
- Jha, V.; Rameshwaram, N.R.; Janardhan, S.; Raman, R.; Sastry, G.N.; Sharma, V.; Subba Rao, J.; Kumar, D.; Mukhopadhyay, S. Uncovering Structural and Molecular Dynamics of ESAT-6:β2M Interaction: Asp53 of Human β2-Microglobulin Is Critical for the ESAT-6:β2M Complexation. The Journal of Immunology 2019, 203, 1918–1929. [Google Scholar] [CrossRef]
- Bretl, D.J.; Demetriadou, C.; Zahrt, T.C. Adaptation to Environmental Stimuli within the Host: Two-Component Signal Transduction Systems of Mycobacterium tuberculosis. Microbiology and Molecular Biology Reviews 2011, 75, 566–582. [Google Scholar] [CrossRef]
- Polena, H.; Boudou, F.; Tilleul, S.; Dubois-Colas, N.; Lecointe, C.; Rakotosamimanana, N.; Pelizzola, M.; Andriamandimby, S.F.; Raharimanga, V.; Charles, P.; et al. Mycobacterium tuberculosis exploits the formation of new blood vessels for its dissemination. Sci. Rep. 2016, 6, 33162. [Google Scholar] [CrossRef] [PubMed]
- Lerner, T.R.; Borel, S.; Greenwood, D.J.; Repnik, U.; Russell, M.R.G.; Herbst, S.; Jones, M.L.; Collinson, L.M.; Griffiths, G.; Gutierrez, M.G. Mycobacterium tuberculosis replicates within necrotic human macrophages. J. Cell Biol. 2017, 216, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Mayer-Barber, K.D.; Andrade, B.B.; Oland, S.D.; Amaral, E.P.; Barber, D.L.; Gonzales, J.; Derrick, S.C.; Shi, R.; Kumar, N.P.; Wei, W.; et al. Host-directed therapy of tuberculosis based on interleukin-1 and type I interferon crosstalk. Nature 2014, 511, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Mourik, B.C.; Lubberts, E.; de Steenwinkel, J.E.M.; Ottenhoff, T.H.M.; Leenen, P.J.M. Interactions between Type 1 Interferons and the Th17 Response in Tuberculosis: Lessons Learned from Autoimmune Diseases. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O'Garra, A. Type I interferons in infectious disease. Nature Reviews Immunology 2015, 15, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Behar, S.M.; Martin, C.J.; Booty, M.G.; Nishimura, T.; Zhao, X.; Gan, H.X.; Divangahi, M.; Remold, H.G. Apoptosis is an innate defense function of macrophages against Mycobacterium tuberculosis. Mucosal Immunol. 2011, 4, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Roca Francisco, J.; Ramakrishnan, L. TNF Dually Mediates Resistance and Susceptibility to Mycobacteria via Mitochondrial Reactive Oxygen Species. Cell 2013, 153, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.G.; Cardona, P.-J.; Kim, M.-J.; Allain, S.; Altare, F. Foamy macrophages and the progression of the human tuberculosis granuloma. Nat. Immunol. 2009, 10, 943–948. [Google Scholar] [CrossRef]
- Griffin, G.K.; Newton, G.; Tarrio, M.L.; Bu, D.-x.; Maganto-Garcia, E.; Azcutia, V.; Alcaide, P.; Grabie, N.; Luscinskas, F.W.; Croce, K.J.; et al. IL-17 and TNF-α Sustain Neutrophil Recruitment during Inflammation through Synergistic Effects on Endothelial Activation. The Journal of Immunology 2012, 188, 6287–6299. [Google Scholar] [CrossRef] [PubMed]
- Poh, X.Y.; Loh, F.K.; Friedland, J.S.; Ong, C.W.M. Neutrophil-Mediated Immunopathology and Matrix Metalloproteinases in Central Nervous System – Tuberculosis. Front. Immunol. 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Dallenga, T.; Schaible, U.E. Neutrophils in tuberculosis--first line of defence or booster of disease and targets for host-directed therapy? Pathogens and Disease 2016, 74, ftw012. [Google Scholar] [CrossRef] [PubMed]
- Mahghani, G.A.; Kargar, M.; Ghaemi, E.A.; Kafilzadeh, F.; Davoodi, H. Role of ESAT-6 in pathogenicity of Beijing and non-Beijing Mycobacterium tuberculosis isolates. Microb. Pathog. 2022, 162, 105366. [Google Scholar] [CrossRef] [PubMed]
- StopTBPartnership. MTBVAC. Available online: https://newtbvaccines.org/vaccine/mtbvac/ (accessed on 29 May 2023).
- Martín, C.; Marinova, D.; Aguiló, N.; Gonzalo-Asensio, J. MTBVAC, a live TB vaccine poised to initiate efficacy trials 100 years after BCG. Vaccine 2021, 39, 7277–7285. [Google Scholar] [CrossRef] [PubMed]
- Camacho, L.R.; Constant, P.; Raynaud, C.; Lanéelle, M.-A.; Triccas, J.A.; Gicquel, B.; Daffé, M.; Guilhot, C. Analysis of the Phthiocerol Dimycocerosate Locus of Mycobacterium tuberculosis: EVIDENCE THAT THIS LIPID IS INVOLVED IN THE CELL WALL PERMEABILITY BARRIER *. J. Biol. Chem. 2001, 276, 19845–19854. [Google Scholar] [CrossRef]
- Bouzeyen, R.; Javid, B. Therapeutic Vaccines for Tuberculosis: An Overview. Front. Immunol. 2022, 13. [Google Scholar] [CrossRef]
- Bellini, C.; Horváti, K. Recent Advances in the Development of Protein- and Peptide-Based Subunit Vaccines against Tuberculosis. Cells 2020, 9, 2673. [Google Scholar] [CrossRef]
- Sable, S.B.; Posey, J.E.; Scriba, T.J. Tuberculosis Vaccine Development: Progress in Clinical Evaluation. Clin. Microbiol. Rev. 2019, 33, e00100–00119. [Google Scholar] [CrossRef]
- Dockrell, H.M.; McShane, H. Tuberculosis vaccines in the era of Covid-19 - what is taking us so long? eBioMedicine 2022, 79. [Google Scholar] [CrossRef]
- Tkachuk, A.P.; Gushchin, V.A.; Potapov, V.D.; Demidenko, A.V.; Lunin, V.G.; Gintsburg, A.L. Multi-subunit BCG booster vaccine GamTBvac: Assessment of immunogenicity and protective efficacy in murine and guinea pig TB models. PLoS One 2017, 12, e0176784. [Google Scholar] [CrossRef]
- Tkachuk, A.P.; Bykonia, E.N.; Popova, L.I.; Kleymenov, D.A.; Semashko, M.A.; Chulanov, V.P.; Fitilev, S.B.; Maksimov, S.L.; Smolyarchuk, E.A.; Manuylov, V.A.; et al. Safety and Immunogenicity of the GamTBvac, the Recombinant Subunit Tuberculosis Vaccine Candidate: A Phase II, Multi-Center, Double-Blind, Randomized, Placebo-Controlled Study. Vaccines 2020, 8, 652. [Google Scholar] [CrossRef]
- Vasina, D.V.; Kleymenov, D.A.; Manuylov, V.A.; Mazunina, E.P.; Koptev, E.Y.; Tukhovskaya, E.A.; Murashev, A.N.; Gintsburg, A.L.; Gushchin, V.A.; Tkachuk, A.P. First-In-Human Trials of GamTBvac, a Recombinant Subunit Tuberculosis Vaccine Candidate: Safety and Immunogenicity Assessment. Vaccines 2019, 7, 166. [Google Scholar] [CrossRef]
- Ning, H.; Zhang, W.; Kang, J.; Ding, T.; Liang, X.; Lu, Y.; Guo, C.; Sun, W.; Wang, H.; Bai, Y.; et al. Subunit Vaccine ESAT-6:c-di-AMP Delivered by Intranasal Route Elicits Immune Responses and Protects Against Mycobacterium tuberculosis Infection. Frontiers in Cellular and Infection Microbiology 2021, 11. [Google Scholar] [CrossRef]
- Zhao, R.; Luo, T.; Ma, P.; Ge, L.; Chen, Z.; Wang, X.; Liao, W.; Bao, L. Improvement of the immunogenicity of ESAT-6 via fusion with the dodecameric protein dodecin of Mycobacterium tuberculosis. Microb. Pathog. 2021, 155, 104890. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.-G.; Choi, S.; Woo Back, Y.; Paik, S.; Park, H.-S.; Sik Kim, W.; Kim, H.; Bin Cha, S.; Hee Choi, C.; Jae Shin, S.; et al. Rv2299c, a novel dendritic cell-activating antigen of Mycobacterium tuberculosis, fused-ESAT-6 subunit vaccine confers improved and durable protection against the hypervirulent strain HN878 in mice. Oncotarget 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Moreira, M.; Ruggiero, A.; Esposito, L.; Choi, H.-G.; Kim, H.-J.; Berisio, R. Structural features of HtpGMtb and HtpG-ESAT6Mtb vaccine antigens against tuberculosis: Molecular determinants of antigenic synergy and cytotoxicity modulation. Int. J. Biol. Macromol. 2020, 158, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, A.; Choi, H.-G.; Barra, G.; Squeglia, F.; Back, Y.W.; Kim, H.-J.; Berisio, R. Structure based design of effective HtpG-derived vaccine antigens against M. tuberculosis. Frontiers in Molecular Biosciences 2022, 9. [Google Scholar] [CrossRef]
- Moguche, A.O.; Musvosvi, M.; Penn-Nicholson, A.; Plumlee, C.R.; Mearns, H.; Geldenhuys, H.; Smit, E.; Abrahams, D.; Rozot, V.; Dintwe, O.; et al. Antigen Availability Shapes T Cell Differentiation and Function during Tuberculosis. Cell Host & Microbe 2017, 21, 695–706. [Google Scholar] [CrossRef]
- Pai, M.; Denkinger Claudia, M.; Kik Sandra, V.; Rangaka Molebogeng, X.; Zwerling, A.; Oxlade, O.; Metcalfe John, Z.; Cattamanchi, A.; Dowdy David, W.; Dheda, K.; et al. Gamma Interferon Release Assays for Detection of Mycobacterium tuberculosis Infection. Clin. Microbiol. Rev. 2014, 27, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Aggerbeck, H.; Ruhwald, M.; Hoff, S.T.; Borregaard, B.; Hellstrom, E.; Malahleha, M.; Siebert, M.; Gani, M.; Seopela, V.; Diacon, A.; et al. C-Tb skin test to diagnose Mycobacterium tuberculosis infection in children and HIV-infected adults: A phase 3 trial. PLoS One 2018, 13, e0204554. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO operational handbook on tuberculosis: module 5: management of tuberculosis in children and adolescents; World Health Organization: Geneva, 2022. [Google Scholar]
- Vidyarthi, A.; Khan, N.; Agnihotri, T.; Siddiqui, K.F.; Nair, G.R.; Arora, A.; Janmeja, A.K.; Agrewala, J.N. Antibody response against PhoP efficiently discriminates among healthy individuals, tuberculosis patients and their contacts. PLoS One 2017, 12, e0173769. [Google Scholar] [CrossRef] [PubMed]
- Araujo, Z.; Fernández de Larrea, C.; López, D.; Isern-Kebschull, J.; de Waard, J.H.; Hagel, I.; Camargo, M.; Vanegas, M.; Patarroyo, M.A. ESAT-6 and Ag85A Synthetic Peptides as Candidates for an Immunodiagnostic Test in Children with a Clinical Suspicion of Tuberculosis. Dis. Markers 2021, 2021, 6673250. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Control of ESAT-6 secretion by PhoP-dependent regulatory networks. (A) Lineages of the MTBC with the RD deletions and PhoPR mutations are highlighted [20,23,28,29]. (B) Several genes from the ESX-1 and the extended ESX-1 (espACD and espR) regions required for ESAT-6 protein synthesis and export are displayed. PhoP (blue ellipses) is a transcriptional activator and an effector of the signal transducer PhoR, that interacts with the espR, espA, and whiB6 promoters. In its phosphorylated form, PhoP binds to DNA with higher affinity. Mtb, carrying a functional PhoR, is able to sense a stress-like stimulus and subsequently phosphorylate PhoP. EspR (pink ellipses) activates the espACD locus. WhiB6 (green circles) also interacts with the promoter regions of the espA, pe35, and espB genes. The espACD locus, is activated by all these circuits and the protein translated EspA is required for ESAT-6 secretion. The RD1, absent in BCG, and of RD8, absent in M. africanum L6 and L8 animal-adapted lineages (including M. bovis), as well as polymorphisms in the espACD and whiB6 promoters (asterisks) are indicated. M. bovis and M. africanum L6, carrying a defective PhoR G71I allele, are expected to generate a low affinity binding effector due to phosphorylation impairment of PhoP. Nevertheless, ESAT-6 secretion in these species is restored to some extent by compensatory mutations in the espACD promoter region, including RD8 deletion and species-specific polymorphisms (asterisks) [62].
Figure 1.
Control of ESAT-6 secretion by PhoP-dependent regulatory networks. (A) Lineages of the MTBC with the RD deletions and PhoPR mutations are highlighted [20,23,28,29]. (B) Several genes from the ESX-1 and the extended ESX-1 (espACD and espR) regions required for ESAT-6 protein synthesis and export are displayed. PhoP (blue ellipses) is a transcriptional activator and an effector of the signal transducer PhoR, that interacts with the espR, espA, and whiB6 promoters. In its phosphorylated form, PhoP binds to DNA with higher affinity. Mtb, carrying a functional PhoR, is able to sense a stress-like stimulus and subsequently phosphorylate PhoP. EspR (pink ellipses) activates the espACD locus. WhiB6 (green circles) also interacts with the promoter regions of the espA, pe35, and espB genes. The espACD locus, is activated by all these circuits and the protein translated EspA is required for ESAT-6 secretion. The RD1, absent in BCG, and of RD8, absent in M. africanum L6 and L8 animal-adapted lineages (including M. bovis), as well as polymorphisms in the espACD and whiB6 promoters (asterisks) are indicated. M. bovis and M. africanum L6, carrying a defective PhoR G71I allele, are expected to generate a low affinity binding effector due to phosphorylation impairment of PhoP. Nevertheless, ESAT-6 secretion in these species is restored to some extent by compensatory mutations in the espACD promoter region, including RD8 deletion and species-specific polymorphisms (asterisks) [62].
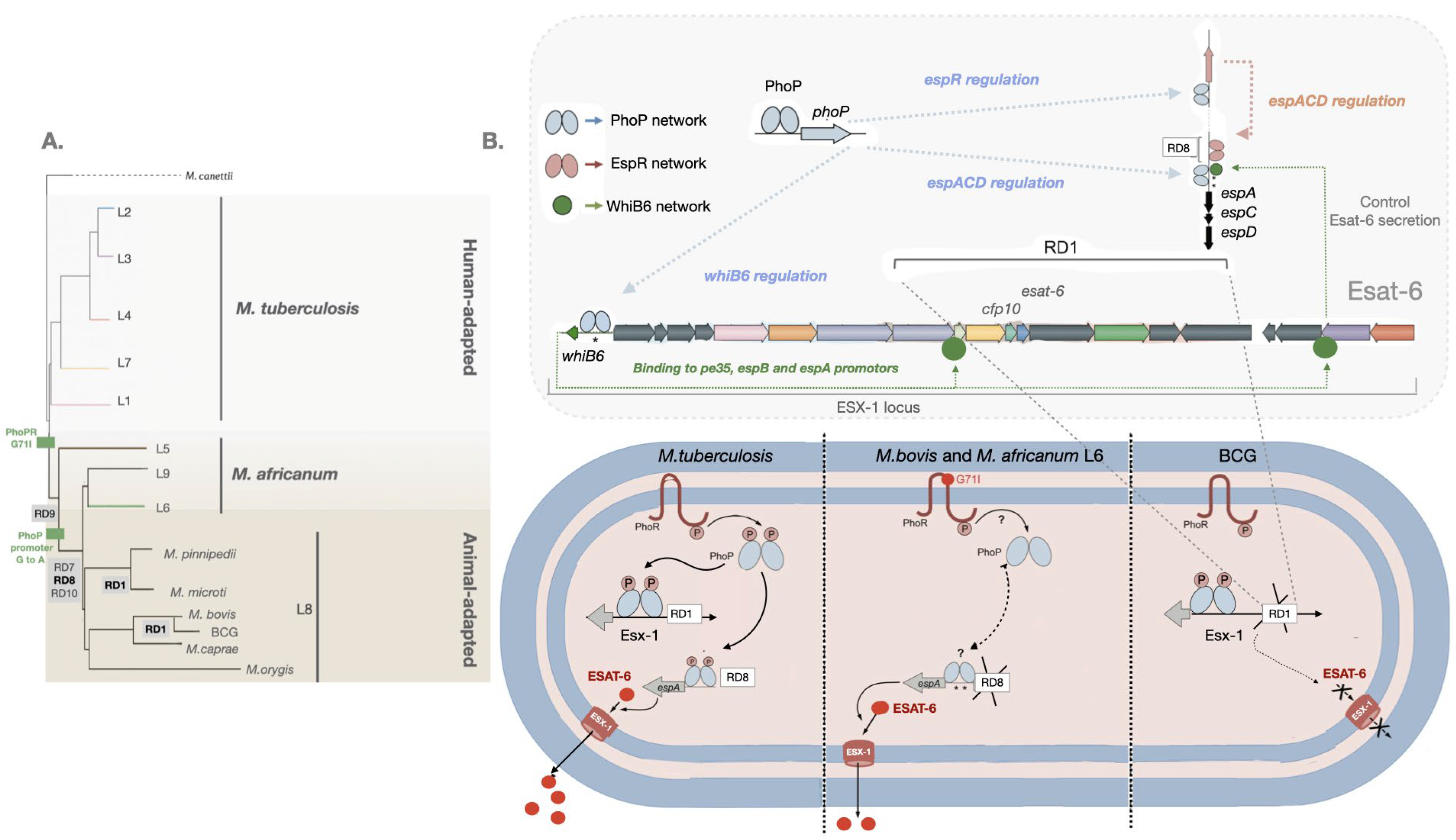
Figure 2.
Effects of ESAT-6 on the outcome of Mtb infection of host macrophages. Upon internalization into macrophages, Mtb manipulates the endocytic pathway to prevent fusion of pathogen-containing phagosomes with lysosomes. This allows the bacilli to replicate in early phagosomes (top of the figure). A few pathogens escape into the cytosol via the ESX-1 secretion apparatus, which secretes ESAT-6. ESAT-6 induces perturbations in the phagosomal membrane allowing bacilli or their PAMPS to exit the vesicle. PAMPS such as ESAT-6 or bacterial DNA activate innate sensors such as NLRP3, AIM2 to form inflammasomes required for IL-1β processing via casp-1. Under IFN𝛾 signaling regulated amounts of cytokines are secreted from of the cell, allowing the control of the intracellular bacterial loads by TNFα, which in turn induces apoptosis. In addition, IL-1β activates COX2 to produce PGE2 an eicosanoid that protects the mitochondrial inner membrane and contributes to apoptosis. Alternatively bacterial DNA activates the cGAS/cGAMP/STING pathway leading to TNK-1 activation, which will induce autophagy and therefore the control of the intracellular load of Mtb. This is what mostly happens during latent TB (black arrows). Under low IL-1β, controlled recruitment of neutrophils prevents tissue damage. During progression to active TB disease, bacterial replication induces high levels of TNFα and this, together with impairment of IFN𝛾, results in the strongest activation of inflammasomes; casp-1 stimulates gasdermin to form pores at the cell membrane inducing pyroptosis. High levels of TNFα induce necroptosis. TBK-1 phosphorylates the transcription factor IRF3 leading to the secretion of type-I IFN such as IFNβ. IFNβ is immunosuppressive in chronic infections by activating IL-10, deactivating macrophages and promoting LX4 synthesis via 5-LO. LX4 causes organelles such as mitochondria to swell and induces cell death by necrosis. Altogether this helps Mtb replicate massively in macrophages and release it into the extracellular environment (light blue lines).
Figure 2.
Effects of ESAT-6 on the outcome of Mtb infection of host macrophages. Upon internalization into macrophages, Mtb manipulates the endocytic pathway to prevent fusion of pathogen-containing phagosomes with lysosomes. This allows the bacilli to replicate in early phagosomes (top of the figure). A few pathogens escape into the cytosol via the ESX-1 secretion apparatus, which secretes ESAT-6. ESAT-6 induces perturbations in the phagosomal membrane allowing bacilli or their PAMPS to exit the vesicle. PAMPS such as ESAT-6 or bacterial DNA activate innate sensors such as NLRP3, AIM2 to form inflammasomes required for IL-1β processing via casp-1. Under IFN𝛾 signaling regulated amounts of cytokines are secreted from of the cell, allowing the control of the intracellular bacterial loads by TNFα, which in turn induces apoptosis. In addition, IL-1β activates COX2 to produce PGE2 an eicosanoid that protects the mitochondrial inner membrane and contributes to apoptosis. Alternatively bacterial DNA activates the cGAS/cGAMP/STING pathway leading to TNK-1 activation, which will induce autophagy and therefore the control of the intracellular load of Mtb. This is what mostly happens during latent TB (black arrows). Under low IL-1β, controlled recruitment of neutrophils prevents tissue damage. During progression to active TB disease, bacterial replication induces high levels of TNFα and this, together with impairment of IFN𝛾, results in the strongest activation of inflammasomes; casp-1 stimulates gasdermin to form pores at the cell membrane inducing pyroptosis. High levels of TNFα induce necroptosis. TBK-1 phosphorylates the transcription factor IRF3 leading to the secretion of type-I IFN such as IFNβ. IFNβ is immunosuppressive in chronic infections by activating IL-10, deactivating macrophages and promoting LX4 synthesis via 5-LO. LX4 causes organelles such as mitochondria to swell and induces cell death by necrosis. Altogether this helps Mtb replicate massively in macrophages and release it into the extracellular environment (light blue lines).
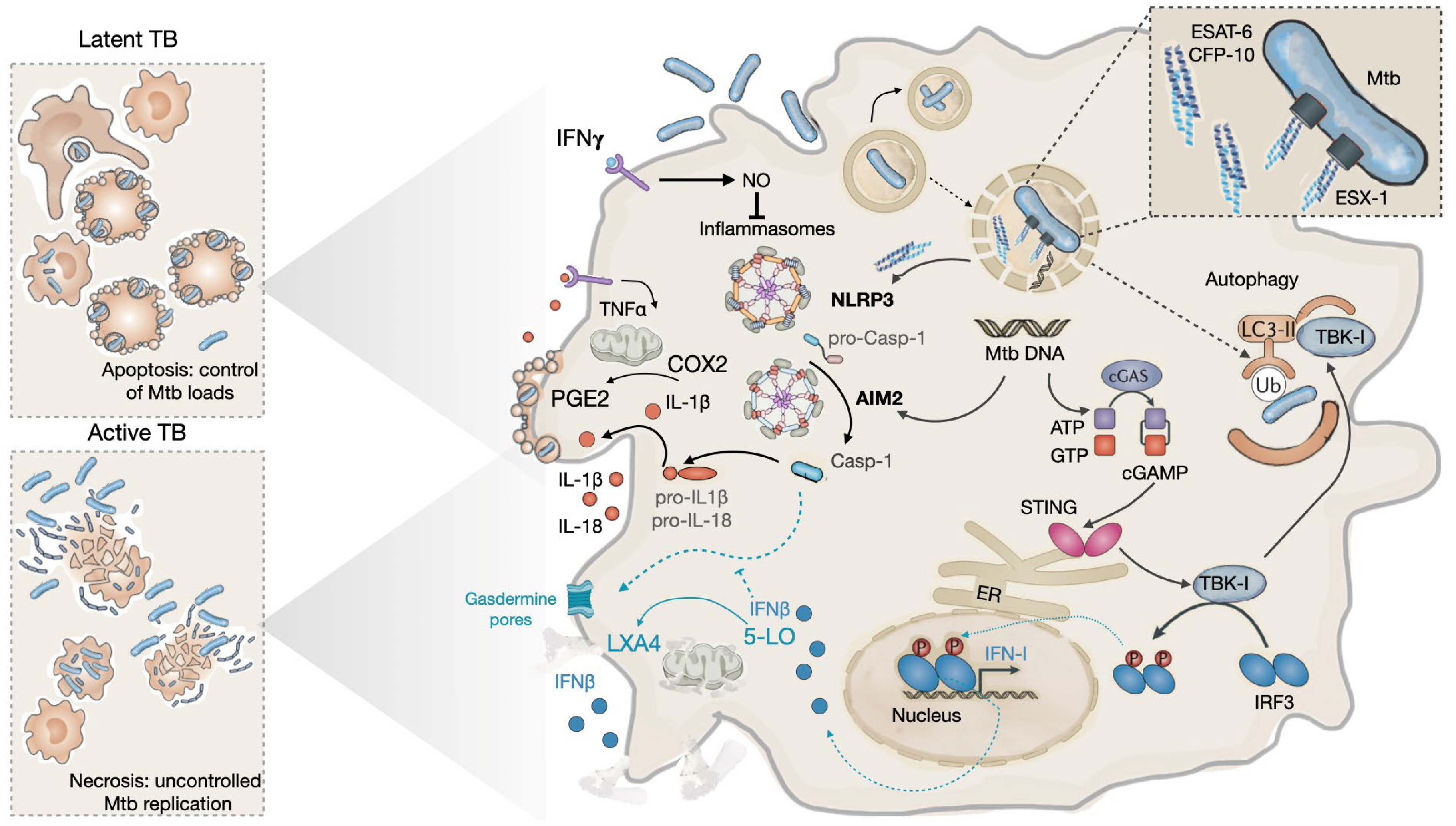
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



