
Submitted:
15 May 2023
Posted:
15 May 2023
You are already at the latest version
Abstract
This literature review was aimed to collect investigations on the in vivo evidences for bacteria associated to fermented dairy foods to behave as probiotics with beneficial effects in the prevention and treatment of various diseases. All main bacterial groups commonly present in high numbers in fermented milks or cheeses were taken into account, namely starter lactic acid bacteria (SLAB) Lactobacillus delbrueckii subsp. bulgaricus and lactis, L. helveticus, Lactococcus lactis, Strepto-coccus thermophilus, non starter LAB (NSLAB) Lacticaseibacillus spp., Lactiplantibacillus plantarum, dairy propionibacteria and other less frequently encountered species. Only studies regarding strains of proven dairy origin were considered. Studies in animal models and clinical studies showed that dairy bacteria ameliorate symptoms of inflammatory bowel disease (IBD), mucositis, metabolic syndrome, aging and oxidative stress, cancer, bone diseases, atopic dermatitis, allergies, infections and damages caused by pollutants, mild stress and depression. Immunomodulation and changes in the intestinal microbiota were the mechanisms most often involved in the observed effects. The results of the studies considered indicated that milk and dairy products are a rich source of beneficial bacteria that should be further exploited to the advantage of human and animal health.
Keywords:
dairy microorganisms
; SLAB
; NSLAB
; propionibacteria
; minor species
; in vivo studies
1. Introduction
Probiotic foods are currently defined as foods that supply “probiotics” proven to exert health-promoting effects in human trials. These effects must derive at least in part from the microorganisms present and must be distinct from those of the food matrix [1]. For probiotic foods the recommendation of providing an adequate number of microorganisms per serving, i.e. at least 109 CFU, is re-affirmed, in accordance with the recommended intake for probiotics of the Food and Agricultural Organization of the United Nations (FAO) and the World Health Organization (WHO) [2]. Indeed, the term “probiotic” can be attributed to “live microorganisms which when administered in adequate amounts confer a health benefit on the host” [FAO/WHO] later emended by the International Scientific Association for Probiotics and Prebiotics (ISAPP) as “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” [3].
Probiotic benefits include regulation of intestinal transit, normalization of perturbed intestinal microbiota, competitive exclusion of pathogens, hypolipidemic, anti-atherogenic and antioxidant effects, immunomodulation and production of bioactive compounds. These effects are strain-specific so differ for microorganisms belonging to the same species [3].
In vitro assessment of acid and bile salt tolerance prior to undertaking in vivo trials is required to indicate if a potential probiotic is able to survive in the host’s gastrointestinal tract (GIT). In vivo trials in animal models are necessary to predict beneficial effects in human beings and explain those effects by the observation of changes in tissue, cell structure and marker expression in different organs.
Bacteria naturally present in dairy products comprise strains able to behave as probiotics with efficacy proven in animal models and in clinical trials [4]. Therefore, in this review, recent studies on the beneficial effects exerted in vivo by dairy products containing potential probiotics and by bacteria isolated from dairy products were surveyed to highlight the multiple health benefits deriving from the uptake of bacteria associated to the fermentation and ripening of dairy foods.
The in vivo studies summarized in this review were retrieved by a Google Scholar (https://scholar.google.com/, accessed on 10 April 2023) and Scopus (https://www.scopus.com/search/form.uri?display=basic#basic, accessed on 15 April 2023) literature database search with the word association "name of bacterial genus”, “in vivo" and "dairy" or "milk", "cheese", "yogurt", "kefir" or "fermented milk". The origin of strains used in the trials, when not specified in the text, was derived by a search in NCBI nucleotide sequence database (https://www.ncbi.nlm.nih.gov/nuccore/) and only articles for which the strain origin could be retrieved were included in the study. The database search was restricted to the years 2019 - 2023.
All the probiotic microorganisms described in the studies under consideration were preliminarily tested for tolerance to acidic pH and at least 0.3% (w/v) bile salts, adhesion to human intestinal cell lines, presence of virulence traits and antibiotic resistance (AR).
Then specific aspects of the probiotic activity were examined, such as pathogen inhibition in vitro or co-aggregation with pathogens, exopolysaccharide (EPS) production, that is involved in immunostimulation and cholesterol assimilation and bile salts hydrolysis in vitro as from the study of Darwish et al. [5]. Genome sequences are available for many of the strains used in the trials, in accordance to the requirements for use of microorganisms as probiotics and in food products [6].
A recent review by Illikoud et al. [4] summarized the effects exerted in the amelioration of different medical conditions in animal models and clinical studies published until 2021 by bacterial strains belonging to the species used as starters in dairy products, namely Lactobacillus delbrueckii subsp. Bulgaricus and lactis, L. helveticus, Lactococcus lactis, Propionibacterium freudenreichii and Streptococcus thermophilus. Results of the in vivo trials summarized in that review article are shortly recalled here. In addition, more recent studies or relevant investigations not reported there are synthetized for the above species. Moreover, this review takes into account the non-starter dairy lactic acid bacteria (NSLAB) represented by the Lacticaseibacillus species, Lactiplantibacillus plantarum, Pediococcus acidilactici and other species less frequently isolated from dairy products. Some studies on enterococci are also reviewed, though the safety of these microorganisms is controversial. All these bacteria are naturally present in dairy products and possibly ingested in high numbers, close to 109 CFU per serving, likely to influence host health if endowed with functional traits. Preclinical studies in animal models, clinical studies and applications in the veterinary sector are reported for each bacterial group.
2. Fermented dairy products
Dairy products were shown to represent a source of bacteria able to establish, at least temporarily, in human gut. It was found that a traditional Pasta Filata fresh cheeses obtained with natural bacterial cultures contributed bacterial components to the intestinal microbiota since identical sequences of selected genetic markers, rpoD and clpP, discriminant at intra-species level, were found both in the cheese and in healty children who consumed the cheese for one week. Some of those fecal isolates were obtained after two weeks of cheese administration suspension, showing a good colonization ability [7].
An in vivo pilot study involving healthy individuals Bifidobacterium mongoliense BMONG18 detected along the whole Parmesan cheese production chain was detected in the feces of all individuals during cheese consumption. After one week of suspension B. mongoliense BMONG18 decreased revealing low persistence capacity in human gut. Despite the in vivo function of B. mongoliense BMONG18 was not elucidated, the study demonstrated that cheese consumption contributes intestinal microbiota components [8].
Illikoud et al. [4] inferred from clinical studies regarding the effects on health of fermented dairy products that their deprivation from human diet caused a lower efficiency of the innate immune response and that daily consumption of yogurt containing a conventional starter constituted by S. thermophilus and L. delbrueckii, counteracted this effect. Six reported studies showed that yogurt, cheese or sour cream consumption decreased biomarkers of inflammation in different categories of subjects, including obese women with non-alcoholic fatty liver disease and metabolic syndrome, premenopausal women, obese people, normal weight and overweight volunteers. They also cited a systematic review of the literature considering 10 trials with contrasting conclusions suggesting that the effectiveness of different fermented dairy products can vary, possibly for the different functional activities of the microorganisms present. More recent studies are summarized below.
2.1. Amelioration of intestinal health
Antimicrobial peptides of the regenerating family member 3 (REG3) family, that maintain the intestinal barrier, is reduced in the small intestines by aging, but the long-term ingestion of yogurt fermented with L. delbrueckii subsp. bulgaricus 2038 and S. thermophilus 1131 by aged mice increased its expression. The REG3 family is induced in the intestinal epithelial cells by interleukin IL-22, predominantly produced by type 3 innate lymphoid cell (ILC3) that is, in turn, induced by IL-23, produced by dendritic cells (DCs) and macrophages after stimulation of Toll-like receptors (TLRs) by bacterial cell component. In the study of Kobayashi et al. [9] oral administration of these strains to specific pathogen-free (SPF) male BALB/c mice led to Reg3g induction in the small intestine cells and production of IL-22 and IL-23 with a higher effect exerted by S. thermophilus 1131.
2.2. Amelioration of atopic dermatitis
Numerous studies have reported beneficial effects of probiotics on atopic dermatitis, e.g. for the widely used probiotic L. rhamnosus GG. Based on this evidence, a study aimed to evaluate the effect of cream cheese containing Lactococcus chungangensis CAU 28 (CAU 28) (1.4 g/kg/mouse daily) and the dry cells (1010 CFU/mouse, daily) of the same strain compared to an untreated positive control and treatment with bepotastine besilate (BB) on the amelioration of atopic dermatitis in female BALB/c mice sensitized with ovalbumin (OVA) when administered for 8 weeks [10].
Cytokines produced by regulatory T cell (Treg) cells, such as IL-10 and IL-1β and Th2 cytokines IL-4 and IL-5 were significantly lower and levels of the Th1 cytokines, IL-12, IFN-γ, and TNF-α were significantly higher in the CAU 28 and CAU 28 cream cheese groups, indicating an enhancement of the Treg-mediated suppression of Th2 immune response.
The activation of CD 86 T cell protein expression was significantly lower in the CAU 28 cream cheese group and the expression of CD 274 suppressor of adaptive immune response was significantly higher in the CAU 28 and CAU 28 cream cheese groups. Serum IgE levels and eosinophil, neutrophil, lymphocyte, and monocyte percentages significantly decreased in the latter groups. Mast cell accumulation at dorsal skin and ileal lesions was suppressed in mice treated with CAU 28 and CAU 28 cream cheese and reduced eosinophil infiltration was observed in the dorsal skin lesions of mice treated with CAU 28, CAU 28 cream cheese, and BB.
High-throughput sequencing of 16S rRNA gene from feces showed a positive correlation between CD 274 and Bacteroidales, Deferribacteraceae, Prevotellaceae, Oscillospiraceae, Rikenellaceae and Veillonellaceae, while CD 86 levels were correlated with several bacterial families, including Verrucomicrobiales and negatively correlated with Desulfovibrionaceae. Bacteroides and Akkermansia were present at significantly higher abundance in the CAU 28 cream cheese-treated group. The levels of short chain fatty acids (SCFA), associated with the maintenance of gut health and positively correlated with Lactobacillus, Bacteroides, Ruminococcus, and Akkermansia, were found to be higher in the feces of mice treated with CAU 28 cream cheese.
2.3. Allergy amelioration
Milk fermentation was found to alleviate indicators of the immune reaction in CMPA. This immune hypersensitivity condition is a complex disorder in which the main symptoms are atopic dermatitis, eczema, asthma, vomiting, and recurrent diarrhea. In CMPA, B cells induced by antigen activated CD4+ T helper (Th) cells produce IgE that bind to a high-affinity Fc receptor on the surface of mast cells or basophils that binds to allergen epitopes and triggers the release of inflammatory mediators such as histamine and mast cell protease-1 (MCPT-1). It was shown that fermentation with combined LAB could reduce the antigenicity of cow milk protein by destroying the linear epitopes in vitro. In the study of Zhao et al. [11] L. helveticus KLDS 1.8701, selected for antioxidant capacity and isolated from traditional fermented dairy products in Sinkiang, China and endowed with genes coding two cell envelope proteases (CEP), and L. plantarum KLDS 1.0386, isolated from an Inner Mongolia traditional fermented dairy product, endowed with 21 peptidase genes and transport systems, were used in single or mixed culture for the degradation of α-casein, α-lactoalbumin, and β-lactoglobulin in reconstituted skim milk. The antigenicity of the fermented milks was compared in specific-pathogen-free female Balb/c mice. On days 0, 7, 14, and 21, the mice were sensitized with a mixture of fermented milk containing L. helveticus KLDS 1.8701 and L. plantarum KLDS 1.0386, alone or in combination, and 10 µg of cholera toxin (CT) as a Th2-polarizing adjuvant. Anaphylactic shock and diarrhea were less severe and the spleen immune indices were lower in the fermented milk groups. Histamine and MCPT-1 were significantly lower in the fermented milk groups, lungs showed a normal alveolar structure without infiltrating inflammatory cells and the structure of jejunum villi did not present remarkable edema and inflammatory cell infiltration.
2.4. Amelioration of metabolic syndrome
The study of Makwana et al. [12] reported that probiotic fermented milk alone (T1) or enriched with 2% whey proteins and soy proteins (T2) and containing 108–109 CFU/ml of lactobacilli could effectively control obesity when administered for 4 weeks to adult male Wistar rats fed with high fat diets (HFD). The fermented milks were prepared with L. helveticus MTCC V3, S. thermophilus MD2, and L. rhamnosus MTCC NS6 isolated from milk and Indian style yogurts.
In the fermented milk group weight gain was 46–55% while it increased by 71% in the group receiving only the HFD. The liver weight and the weight of abdominal fat in treatments T1 and T2 were considerably reduced. Treatment with T1 and T2 produced a significant decline in triglycerides (TG) and total cholesterol but did not influence the levels of low density lipoprotein (LDL). Among alkaline phosphatase (ALP), aspartate aminotransferase (AST), and alanine aminotransferase (ALT) indicators of liver disease, associated more frequently to obese individuals than the normal population, ALT and ALP significantly decreased in T1 and T2. Release of adipocyte leptin, a regulator of food intake and energy utilization associated with hyperleptinemia, was delayed in T1 and T2. Liver sections stained with eosin and hematoxylin for T1 showed fewer and smaller lipid vacuoles. Viable counts of lactobacilli and S. thermophilus in feces during the feeding phase were of the order of 7 log CFU/ml and increased progressively during the feeding period, indicating that the probiotic lactobacilli stably colonized the intestinal tract.
2.5. Antihypertensive effects
The benefits of milk fermented by dairy LAB is exerted also by their ability to form substances with potential to ameliorate disease conditions. An example is given by angiotensin converting enzyme (ACE) inhibitors (ACE-I) that exert antihypertensive effects and the best studied ACE-I containing foods are fermented dairy products. The formation of these compounds depends on the proteolytic and peptidolytic activity of LAB that generates small peptides from milk proteins, mainly α, β and κ-caseins, during fermentation. However, the proteolytic systems in various LAB varies not only between species but also between different strains of the same species. Consequently, starter strains producing fermented milk with higher ACE-I properties must be appropriately selected.
Glazunova et al. [13] used a spontaneously hypertensive rat (SHR) model to evaluate the effect on blood pressure of reconstituted skim milk (RSM) fermented by 8 dairy starter LAB (SLAB) strains or non fermented negative control for four weeks. Rats fed with RSM fermented with Lactobacillus delbrueckii Lb100 isolated from commercial yogurt and Lactococcus lactis AM1 isolated from amasi showed a significantly decrease of systolic pressure (Psyst) down to 154 mmHg at the end of experiment with a ΔPsyst of approximately −17 mmHg. In addition, the L. delbrueckii Lb100 group showed also significantly lower levels of total cholesterol and high density lipoprotein cholesterol (HDL). In the following sections in vivo studies regarding single species or genuses of dairy bacteria are summarized.
3. Lacticaseibacillus
The Lacticaseibacillus genus comprises different species able to exert probiotic functions with a variable asset of genes that confer probiotic activities [14]. L. paracasei is the species of non-starter LAB (NSLAB) most frequently isolated from cheeses of different types [7,15,16,17].
3.1. Improvement of intestinal health
Adhesion to the intestine represents a critical parameter for probiotic action. The adhesion ability of L. casei ATCC 393 to the gastrointestinal tract of Wistar rats was examined by Saxami et al. [18]. After daily administration for 7 days and strain-specific multiplex PCR on isolates showed that the level of adhesion in cecum and colon was equal or higher than 6 Log CFU/g, reaching a minimum level required for a probiotic effect. However, the adhesion in GIT was transient, so daily consumption of the specific strain appeared necessary.
L. paracasei CIDCA 8339 with probiotic potential and safe for consumption isolates from kefir grains, when administered in fermented milk at 1010 CFU daily to 7 week-old male BALB/c mice in which acute gastric damage was induced with ethanol and HCl, protected gastric mucosa with a significant reduction in the gastric damage in 50% mice [19].
3.2. Anti-cancer effects
The remarkable anti-tumor, immunomodulatory and modulation of intestinal microbiota activity of lactobacilli administration has been linked to the increase in stimulatory cytokines that favor cytotoxicity. Consumption of probiotic bacteria has also been linked to prolonged survival of tumor-bearing mice associated with increased IL-12 production. The combination of increased IL-12 secretion with interferon gamma (IFN-γ) production and augmentation of the NK cells and CD4+ T cells populations has been verified by multiple studies.
Administration of lactobacilli has been shown to stimulate the production of cytotoxic cells, maturation of Th1 helper T cells and their cytokine production with activation signaling pathways inducing the differentiation, development and targeted intra-tumor migration of immune cell populations CD8+ T and NK that increased cancer cell apoptosis through cell interactions and secretion of anti-cancer factors.
The protective effect of L. casei ATCC 393 against CT26-induced colon carcinoma was evaluated by Aindelis et al. [20] by administering 109 CFU of the strain daily to female BALB/c mice for 13 days. At day 10, 5 × 106 CT26 cells were injected subcutaneously. In L. casei treated mice a statistically significant increase in IFN-γ was observed in Peyer’s patches three and seven days post inoculation of CT26 and IL-12 levels were found to be elevated on day 13. In spleen cells a 2.5% increase of CD45+CD8+ T cells and increased IFN-γ production, was observed. Levels of IL-12p40 were slightly but significantly increased in tumors and a two-fold increase in Granzyme B accumulation was observed. Moreover, a four fold increase in tumor-infiltrating lymphocytes (TILs) CD3+CD8+ cytotoxic T cells occurred that plays a crucial role in cancer immunotherapy. Caspase 3 was significantly active and Poly (ADP-Ribose) Polymerase 1 (PARP1), a typical apoptotic marker, increased. Accumulation of ligands for the C-C chemokine receptor and receptors that act as signals for the migration of CD8+ T cells and NK cells showed that L. casei ATCC 393 induced strong Th1 immune responses and impaired tumor growth for the production of immunostimulatory cytokines in lymphatic organs and secretion of molecules that induced the migration of cytotoxic T cells in the tumor.
This probiotic strain was used to synthesize selenium nanoparticles (SeNps). When administered by oral gavage in male BALB/c mice in which colon cancer was induced by subcutaneous injection of CT26 cells, both 108 CFUs per ml of L. casei ATCC 393 or and the derived SeNps induced a statistically significant inhibition in tumor growth with a tumor volume reduction of 77%, 54%, respectively [21].
3.3. Amelioration of metabolic syndrome
L. rhamnosus Lb102 isolated from raw milk (acc. n. KJ679486.1) was evaluated in vivo in C57BL/6 mice fed with a high-fat high-sucrose diet for 8 week. Mice treated with L. rhamnosus Lbl02 strain showed a significant decrease of body weight gain attributable to the reduction of visceral fat, indicating a decreased food efficiency. L. rhamnosus Lbl02 improved insulin sensitivity after 6 weeks of administration, proved by a rapid drop of glucose levels at 10 min after insulin injection without any change in fasting glycemia. After 8 weeks of treatment glucose tolerance improved in a oral glucose tolerance test. L. rhamnosus Lbl02 significantly reduced plasma non esterified fatty acid (NEFA) levels. In fecal samples of L. rhamnosus Lbl02-supplemented mice, the abundance of genuses Adlercreutzia and Clostridium after 4 weeks and Streptococcus and Lactobacillus after 8 weeks decreased, while Anaerovorax genus was more represented. In gut treatment with L. rhamnosus Lbl02 significantly increased gene expression of zonula occludens 1 (zo-1) and occludin, which code for important tight-junction proteins controlling epithelial integrity. Moreover, mucins 2 and 3 (muc2 and -3) gene expression increased, suggesting that the gut barrier was reinforced by a thicker mucus-layer. These results indicated that treatment with L. rhamnosus Lbl02 protected gut integrity in an obesogenic diet [22].
3.4. Immunostimulation
L. paracasei K5 dairy isolate induced a rapid and high level recruitment of leukocytes in the exudates when injected subcutaneously in air pouches in ΒALB/c male mice 3 h post injection, significantly more than for L. casei ATCC 393 and L. rhamnosus GG, with higher levels of granulocyte-colony stimulating factor (G-CSF); interleukins IL-1α, IL-1β, IL-1ra, IL- 6, and IL-16; and chemokines CCL3, CCL4, CXCL1, and CXCL2. Moreover, the administration of L. paracasei K5 decreased the expression levels of sICAM, TIMP-1, complement component 5α (C5α), macrophage colony-stimulating factor (M-CSF), and triggered receptor expressed on myeloid cells 1 (TREM-1) [23].
Kefir milk prepared from Tibetan Kefir grains containing 26 LAB and 14 yeast strains with or without addition of about 5×107 CFU L. paracasei Ž2, isolated from the same source, were administered daily to female BALB/c mice for 11 days. The L. paracasei group showed the highest level of the helper T lymphocytes (CD4+), with increased ratio of CD4+:CD8+ lymphocyte, indicative of immunostimulation. Mucin MUC-1 was upregulated in the jejunum and MUC-2 and IgA were upregulated also in cecum and ileum. The highest loads of lactobacilli were noted as biofilm on the surface of the mucin layer in the L. paracasei group. From the results authors suggested that the L. paracasei Ž2 strain alone should be used mainly for short-term stimulation of immunocompromised patients [24].
3.5. Infection mitigation
As reported by Valente et al. [25], L. rhamnosus D1 isolated from Minas cheese showed protection from L. monocytogenes infection in four-week-old conventional male BALB/c mice that received the following treatments (whole doses) by gavage each day for 2 weeks: 108 CFU of L. plantarum B7 or 108 CFU of L. rhamnosus D1, or 100 µL of 0.9% saline (control group). On day 11 of treatment, all animals were infected with 106 CFU of L. monocytogenes intravenously. After the challenge a greater weight loss was found in the mice in the L. plantarum B7 group. In contrast, the L. rhamnosus D1 group showed the greatest weight gain, indicating improvements in feed conversion that may help animals to fight infection.
Lacticaseibacillus casei LC2W, isolated from a traditional Cinese dairy product, showed inhibitory effect on E. coli O157:H7 in vivo. The two stains were labelled with different fluorescent proteins and monitored in the intestinal tracts of live mice using an in vivo imaging system. The results showed that L. casei LC2W inhibited the colonization of O157:H7 and prevention and treatment effects of this strain on the colitis severity in mice infected by O157:H7 were verified [26].
L. paracasei Zhang was isolated from koumiss, a traditional drink made from mare's milk by nomadic populations of China and Mongolia that is believed to exert curative effects on digestive diseases and other diseases, including tuberculosis, bronchitis, and anemia [27]. When 1010 CFU of this strain were administered daily for 28 days to young, middle age and elderly individuals, it modulated faecal microbiota by suppressing the potentially harmful Pseudomonas and Acinetobacter genera, elevating SCFA for a prolonged period and reducing total bile acids (TBA) [28].
L. paracasei Zhang alleviated mice mastitis induced by E. coli in lactating mice by determining an increased expression of the tight junction proteins claudin-3, occludin, and ZO-1 and decreased expression of the inflammatory cytokines TNF-α, IL-1β, and IL-6 with improvement of the histological score and of blood-milk barrier disruption. The treatments compared were intramammary infusion of 105 CFU L. paracasei Zhang alone in one group, of 103 CFU E. coli alone in another group and of 105 CFU L. paracasei Zhang and injection of 103 CFU E. coli after 24 h in a third group. In the control group only suspension buffer was infused. No redness or swelling was detected in the mammary tissues in the control, L. paracasei Zhang and L. paracasei Zhang and E. coli, groups, while redness and bleeding were observed in the E. coli group with remarkable pathological injury and presence of a large number of neutrophils in the alveolar lumen [29].
3.6. Allergy alleviation
Oral administration of L. paracasei Zhang attenuated allergy symptoms and intestinal epithelial damage induced by the allergen tropomyosin (TM) in BALB/c mice, altered the development and function of dendritic cells (DCs), T cells, and B cells, finally resulting in the change of TM-specific antibody isotypes into a tolerogenic pattern [30].
4. Lactiplanibacillus plantarum
4.1. Amelioration of intestinal health
Some studies demonstrated the beneficial effects of L. plantarum on intestinal health and microbiota composition. Zago et al. [31] found that among 90 strains of L. plantarum isolated from different cheeses, three strains with high degree of agglutination, surface hydrophobicity and good tolerance to simulated gastric juice showed no microbial translocation to liver and spleen when administered to 6-week-old BALB/c female mice for 2, 5 or 7 days in concentration 108 CFU/ml. The histological examination of the small intestine showed no lymphocyte infiltrates in the villi lamina propria, or the presence of edema or mucosal atrophy. The phagocytic activity of peritoneal macrophages and the number of IgA-producing cells were enhanced for all L. plantarum strains and feeding periods but with different profiles of immune stimulation.
L. plantarum C37 supplied at 4x109 CFU to Kunming male mice led to an increase of the viable counts of lactobacilli in feces and a decrease of the viable counts of enterococci with no significant effect on the number of bifidobacteria, Enterobacter, and Clostridium perfringens [32].
L. plantarum NWAFU-BIO-BS29 isolated from traditional Chinese fermented milk of Gansu province was evaluated for effect on intestinal health of mice. It was orally administered daily for two weeks to BALB/c white mice in 108 CFU/day or 8×109 CFU/day. The higher dose improved the intestinal length and the average value of the liver and spleen indexes. The L. plantarum group showed healthier villi and a normal structure of the intestinal mucosa with no loss of goblet cells. The SCFA profile of the administered group showed a significantly higher ability to produce butyric and valeric acids with a total increase of SCFAs concentration. The L. plantarum treatment led to the increased abundance of Firmicutes, Patescibacteria, Campylobacterota, Deferribacterota, Proteobacteria, and Cyanobacteria phyla and genuses Vampirivibrionia, Acetivibrio sp., Clostridia bacterium, rumen bacterium, Eubacterium brachy group, Halomonas, Lactobacillaceae, Gastranaerophilales, Cyanobacteria, Clostridiales bacterium CIEAF 020, Lachnoclostridium, and Streptococcus and decreased the abundance of Bacteroides and Desulfovibrio [33].
4.2. Amelioration of metabolic syndrome
Yousef et al. [34] reported that L. plantarum YS5, selected on the basis of cholesterol removal ability in vitro, when supplemented in 106–107 CFU/g of feed to male Wistar rats for eight weeks significantly decreased serum total cholesterol, TG, and LDL levels, and increased HDL to levels higher than observed in mice fed with a normal diet. The rats treated with the high fat diet and supplemented with L. plantarum YS5 showed significantly lower very low-density lipoprotein cholesterol (VLDL) compared with the normal diet group.
L. plantarum 29V isolated from raw milk was added to pasteurized honey, where in 28 days it declined from 8.49 Log CFU/ml to final numbers of 7.2 and 5.08 Log CFU/g at 4°C and 25°C respectively, and administered to adult male hypercholesterolemic Wistar rats. During the 28 day treatment, the total cholesterol, (VLDL+LDL)-cholesterol and TG levels remained lower in treated rats, though with no statistically significant differences. In addition, HDL increased and and the atherosclerosis index was significantly lower [35].
The application of L. plantarum LC38 from camel milk on wound site in a diabetic rat model significantly enhanced wound healing activity and accelerated the wound closure after 14 days of wound induction for the capacity to release organic acids lactic, succinic and citric [36].
In C57BL/6J mice fed a cholesterol-enriched diet, L. plantarum Y15, with high bile salt hydrolase (BSH) activity, effectively ameliorated weight gain and alleviated liver histopathological variations. Total cholesterol (TC), TG, and LDL levels were significantly lowered by L. plantarum administration. Metagenome analysis based on the V4 region of the 16S rRNA gene of gut microbiota showed that L. plantarum Y15 supplementation elevated the diversity of the microbial community and contrasted the increase in Proteobacteria (13.54%), increasing the relative abundances of Lactobacillus, Bifidobacterium, Bacteroides, Clostridium, Prevotella and Oscillospira. The expression levels of gene encoding FXR and SHP, that increase the catabolism of cholesterol and biosynthesis of bile acids by the α-hydroxylase CYP7A1 in the liver, were reduced by L. plantarum Y15 supplementation, while CYP7A1 gene was significantly upregulated, thus indicating that L. plantarum Y15 supplementation could lower cholesterol by upregulating cholesterol degradation genes [37].
4.3. Anti-cancer effects
It was reported that oral administration of L. plantarum in mice with colon cancer induced by challenge with murine adenocarcinoma CT26 cells led to changes in the tumor microenvironment, which favored Th1 immunological responses and the recruitment of CD3+CD8+ cytotoxic T cells and natural killer (NK) cells, leading to longer survival of the tumor-bearing mice [38].
4.4. Infection mitigation
In mice infected with Salmonella typhimurium SL1344, the intake of L. plantarum strain ACA-DC287 resulted in a decrease in the levels of pathogen cells associated with intestinal tissues and of those present in the intestinal contents [39].
In a Salmonella Typhimurium challenge, male BALB/c JUnib mice received daily 7 Log CFU of L. plantarum B7 from Brazilian artisanal cheese in fermented milk for seven days prior to infection with 5 Log CFU Salmonella Typhimurium and for seven days post-infection. Cumulative mortality evolution was delayed and weight loss reduced in mice treated with L. plantarum B7. Translocation to liver, which causes the death of mice in this Salmonella Typhimurium infection, was also less severe but not translocation to spleen. Increase of ileal levels of IFN-γ and IL-6 at days 5 and 7 after the challenge with Salmonella Typhimurium, was prevented possibly for the stimulation of TGF-β gene expression.
Less inflammatory foci surrounded by necrotic tissue were observed in liver and less augmented Payer patches and necrosis in mucosa were evident in the ileum. Intestinal villi height was higher in mice treated with L. plantarum B7. In the ileum of one animal that received L. plantarum B7, a higher diversity among the predominant ribosomal sequence variants (RSVs) and a lower abundance of the Salmonella genus RSV was observed, indicating that mechanisms other than direct pathogen reduction are associated with the ameliorated clinical and histopathological parameters [40].
L. plantarum DR7 strain was administered for 12 weeks in a double-blind RCT clinical study to young and middle aged adults, in young adults marginally reduced the duration for nasal, pharyngeal, and general flu symptoms, while in middle-aged adults it significantly reduced the duration of nasal symptoms and marginally reduced the duration of general flu symptoms and frequency of upper respiratory tract infection (URTI) in the 12 weeks study period. Plasma anti-inflammatory cytokines such as IL-4 and IL-10 increased in young adults, but not in middle-aged adults. Plasma proinflammatory cytokines IFN-γ, TNF-α and IL-1β were reduced in middle-aged adults, only IFN-γ was reduced in young adults. Therefore, in general, L. plantarum DR7 alleviated inflammatory parameters.
The administration of L. plantarum DR7 did not affect antioxidant potential in plasma, red blood cell (RBC) membrane, or hemolysate in young adults, marginally increased the antioxidant potential of RBC membranes in middle-aged adults and significantly increased this factor in all subjects. Moreover, it decreased the concentration of MDA in all subjects, upregulated the expression of plasma genes for CD44 and CD117 in young adults and downregulated the expression of plasma genes for CD4 and CD8. The administration of L. plantarum DR7 downregulated the expression of plasma genes for NKp46 and NKp30 in young adults and CD56, NKp46, and NKp30 in middle-aged adults. In general, the administration of L. plantarum DR7 led to less activation of T cells, with more nonresting and mature natural killer (NK) cells. Therefore, it was concluded that L. plantarum DR7 can protect adult populations against URTI by inducing immunomodulatory and anti-inflammatory effects while promoting mucosal barrier integrity and actions of NK cells [41].
4.5. Amelioration of aging damages
D-galactose is a well recognized aging-inducing agent that gives rise to a process similar to natural aging in animal models. Indeed, only a small amount of it is converted into glucose and metabolized, while a large amount remains unaltered causing the formation of large amounts of superoxide anions and oxidation products that damage cell macromolecules [42].
Zhang et al. [42] examined the in vivo antioxidant and gut microbiota regulation effect of its component EPS 1from L. plantarum YW11 in male mice in which oxidative stress was induced with D-galactose. The mice were subcutaneously injected with D-galactose solution and then received vitamin C (VC), to serve as anti-aging control or low-dose (LD) or high-dose (HD) EPS, or fermented milk (FM) containing 8.41 Log CFU/mL of viable L. plantarum YW11 for 12 weeks. The MDA levels of the HD group and fermented milk were similar to those of the VC, indicating an effective blocking of lipid oxidation in serum. The HD group had the highest serum activities of GSH-Px, SOD, CAT, and T-AOC and the CAT activities were higher than those in the VC group. Those enzyme activities were higher in the FM group than in the LD, indicating a dose-dependent antioxidant activity of EPS.
Zhao et al. [42] used D-galactose induced aging in Kunming mice to test the anti-aging effect of L. plantarum KSFY02 isolated from naturally fermented Xinjiang yogurt. Compared treatments were a normal group with no treatment, a positive control supplemented only with D-galactose, a group supplemented daily with 109 CFU/kg of body weight of a L. delbrueckii subsp. bulgaricus strain, a group supplemented daily with 109 CFU/kg of body weight of L. plantarum KSFY02, and a group supplemented with vitamin C as anti-aging agent. After four weeks the mice were intraperitoneally injected with D-galactose for 6 weeks, while the other treatments were continued. At the end of treatment, the indices of thymic, cerebral, cardiac, liver, spleen, and kidney indices of atrophy mice in the LAB and vitamin C treated groups were significantly higher than those of the control group, particularly in the L. plantarum LP-KSFY02 group. In this group, contents of nitric oxide (NO), malondialdehyde (MDA), GSH, and activities of the superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) in the serum were the most similar to the normal group. Mice in the anti-aging treatment groups had fewer lipid vacuoles, indicating alleviated endosmosis, and defined boundary between the epidermis and dermis, alleviated cell swelling and inflammatory infiltration, in particular, in the L. plantarum LP-KSFY02 group. Spleen lesions were found to be lowest in the L. plantarum LP-KSFY02 treatment. The mRNA expression levels for Nos1 (neuronal nitric oxide synthase), Nos3 (endothelial nitric oxide synthase), Sod1 (cuprozinc-superoxide dismutase), Sod2 (manganese superoxide dismutase), Cat (catalase), Hmox1 (heme oxygenase-1), Nfe2l2 (nuclear factor-erythroid 2 related factor 2), Gclm (γ-glutamylcysteine synthetase) and Nqo1 (NAD(P)H dehydrogenase [quinone]1) encoding proteins with anti-oxidative activities in liver and spleen increased significantly in all the anti-aging treatment groups. Expression of SOD 1 and 2, catalase (CAT), GSH 1 and 2 and β-actin (ACTB) in liver and spleen was stronger in the L. plantarum LP-KSFY02 group. Therefore, it could be concluded that the efficacy of L. plantarum LP-KSFY02 was significantly higher than the other treatments in reverting the damages of D-galactose induced aging.
4.6. Improvement of stress symptoms
Liu et al. [44] carried out the first clinical study to correlate the effects of a probiotic towards gut microbiota changes and brain neurotransmitters genes, strengthening the hypothesis of health benefits along the gut–brain axis.
L. plantarum DR7 isolated from fresh cow’s milk was evaluated in a double-blind, RCT design study randomized on men or women, aged 18–60 years old and scored moderate stress level on Cohen’s Perceived Stress Scale (PSS-10). Subjects with chronic diseases, medication assumption and, deficient in glucose-6-phosphate dehydrogenase were excluded. Daily consumption consisted of either 109 CFU/sachet of DR7 powder with maltodextrin excipient or placebo with only maltodextrin. Based on gastrointestinal questionnaire, L. plantarum DR7 decreased the frequency of defecation in all subjects, possibly by modulating the increased bowel movement triggered by the central nervous system (CNS) upon stress. No significant effects against pain and discomfort were registered in normal adults aged above 30 years old, but younger adults below 30 years reported decreased durations for both direct and indirect gastrointestinal symptoms since week-4. After L. plantarum DR7 administration, significant differences were consistently observed in the fecal microbiota, such as reduced abundance of Clostridiales, larger decrease of Selenomonadales, that exert detrimental effects on the host, increased abundance of Deltaproteobacteria and with genuses Bilophila and Desulfovibrio, producing H2S that is an important mediator for epithelial injury repair, dysbiosis prevention, and inflammation resolution, and decrease of Actinomyces. The latter genus is able to cause actinomycosis for its ability to form biofilm, induce and aggravate inflammation. Neurotransmitters β-hydroxylase (DBH), indoleamine 2,3-dioxygenase (TPH2), and tryptophan 2,3-dioxygenase (TDO) involved in the serotonin and dopamine pathways were differently regulated and changes correlated to changes in the relative abundance of specific bacterial groups in the gut microbiota. Bacteroides and Desulfovibrionales, that were higher in the L. plantarum DR7 group were negatively correlated with DBH, that catalyses the conversion of dopamine to norepinephrine, an indication of increased stress, and positively correlated with TPH2 that converts tryptophan to serotonin in the brain, where an imbalanced level of serotonin has been reported in patients with psychological disorders including mood perturbation and anxiety. DBH was positively correlated with Clostridia.
Anxiety is the first psychological reaction to stress and, if prolonged, can lead to mental illnesses including depression that affects over hundreds million people globally. Stress responses activate the autonomic nervous system and the hypothalamic-pituitary-adrenal axis with increased levels of catecholamines and glucocorticoids in blood and tissues. These hormones alter immune functions, such as antigen presentation, leukocyte trafficking and proliferation, antibody secretion and cytokine release. Long-term exposure to glucocorticoids leads to increased resistance of the glucocorticoid receptor with consequent decrease of immune cells sensitivity and activation ofinflammatory responses. L. plantarum DR7 activated the 5’-AMP-activated protein kinase (AMPK) pathway via phosphorylation. AMPK is inactivated in mice by chronic mild stress, inducing anxiety and depression-like behaviours. Therefore, Chong et al. [45] evaluated the effects of L. plantarum DR7 on stress, anxiety, depression, memory capacity and cognitive functions in stressed adults in a double-blind, randomised and placebo-controlled design study. The study participants an probiotic and placebo administration were as in the study described above.
Perceived stress score did not vary significantly in the two groups in 12 weeks. Instead, scores from the Depression, Anxiety and Stress Scale (DASS-42) questionnaire indicated that the administration of L. plantarum DR7 reduced scores in all age groups after 8 weeks. Administration of L. plantarum DR7 significantly reduced plasma cortisol, that activates the brain noradrenergicsystem triggering alertness, awareness, wakefulness and attention, and reduced pro-inflammatory cytokines levels in all subjects and increased the anti-inflammatory cytokine IL-10. The latter increased mostly in young adults after 12 weeks, while for the other subjects only a decrease of pro-inflammatory cytokines IFN-γ, TNF-α, and IL-1β was observed.
The administration of L. plantarum DR7 enhanced social emotional cognition, verbal learning, and and memory, and marginally reduced errors for associate learning in all subjects over 12 weeks benefiting mostly older adults.
The administration of L. plantarum DR7 lowered the expression of plasma neurotransmitters dopamine β-hydroxylase (DBH) and tryptophan 2,3-dioxygenase (TDO) and increased the expressions of tryptophan hydroxylase-2 (TPH2) and 5-hydroxytryptamine receptor-6 (5-HT6) over 12 weeks in all subjects. These data indicated that higher expressions of TPH and 5-HT6 potentially channelled tryptophan towards the production of serotonin.
5. Lactobacillus delbrueckii subsp. bulgaricus and lactis
The subspecies bulgaricus and lactis of Lactobacillus delbrueckii are commonly associated to dairy products. As reported by Illikoud et al. [4] L. delbrueckii subsp. bulgaricus was shown to inhibit the development of colitis-associated cancer induced in mice with azoxymethane and DSS by reducing tumor volume, attenuating the intestinal inflammation, with decreased levels of pro-inflammatory citokynes IL-6, TNF-α, IL-17, IL-23 and IL-1β. Strains of L. delbrueckii subsp. bulgaricus restored antioxidant enzyme activities in induced colitis in rats and, in combination with L. fermentum for eight weeks, favoured healing in ulcerative colitis patients by decreasing expression of IL-6, TNF-α and NF-κB p65, limiting leukocyte recruitment in the colon and the level of fecal calprotectin. In the elderly, oral supplementation of L. delbrueckii subsp. bulgaricus enhanced systemic immunity with increased NK cells and antimicrobial peptide hBD-2 and decreased IL-8 in blood, increased the concentration of saliva IgA, and natural killer (NK) cell activity and enhanced the quality of life (QOL) score. Strains of L. delbrueckii subsp. lactis were shown to mitigate gut inflammation in mice by reducing the macroscopic and microscopic symptoms of DSS-induced colitis, diminishing weight loss, improving survival and modulating the production of cytokines TGF-β, IL-6, and IL-12 in colon and TGF-β and IL-6 in the spleen with expansion of CD4+FOXP3+ regulatory T cells in the cecal lymphnodes [4].
5.1. Alleviation of hepatic injury
Liu et al. [46] evaluated the effects of the L. delbrueckii subsp. bulgaricus L7 isolated from traditional fermented Xinjiang cheese, that showed relevant antioxidant capacity based on 2,2-Diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activity, reduction ability, trolox equivalent antioxidant capacity (TEAC) and antioxidant capacity of milk that it fermented, in alcohol-induced oxidative stress and inflammatory responses in acute alcoholic liver disease (ALD) in C57BL/6 male mice.
The mice group fed with L. delbrueckii subsp. bulgaricus L7 fermented milk presented remarkably decreased oxidative stress in liver tissues caused by alcohol intake showing levels of MDA, SOD and GSH-Px activities comparable with the normal group and a group supplemented with glutathione (GSH). Serum levels of the SCFAs butyric and acetic acids, significantly reduced by alcohol feeding, were restored at normal level by the fermented milk and GSH uptake. SCFAs production is critical in preventing and decreasing inflammation and can suppress the growth of pathogenic intestinal bacteria, modulate gut-microbiota, improve lipid metabolism and the human immune function, lower intestinal pH, and promote the bioavailability of minerals such as magnesium and calcium.
Metagenomic analysis of the 16S rRNA gene hypervariable region V3-V4 showed that L. delbrueckii subsp. bulgaricus L7 administration, but not GSH, better contrasted gut dysbiosis with a decreased abundance of potential pathogens Porphyromonas sp. and Enterococcus spp., decreased Firmicutes/Bacteroidetes ratio and a less remarkable decrease of the genus Ligilactobacillus induced by alcohol. Alcohol feeding reduced the abundance of Comamonadaceae, and enriched Muribaculaceae and Clostridiales. These changes were less evident in mice fed with L. delbrueckii subsp. bulgaricus L7 and GSH. In conclusion, L. delbrueckii subsp. bulgaricus L7 exhibited a better capability than GSH to restore alcohol-induced gut dysbiosis with potential protective effects on alcohol-induced hepatic damage.
6. Lactobacillus helveticus
Based on the studies summarized by Illikoud et al. [4] dairy strains of Lactobacillus helveticus exerted anti-inflammatory properties in TNBS-induced colitis in mice with reduction of histological damage scores and weight loss, colon shortening, bleeding and diarrhea. In addition, L. helveticus strains suppressed tumorigenesis and carcinogenesis in colorectal cancer induced with azoxymethane and DSS in mice by modulating inflammation with reduction of enterocytes and IL-17-producing T cells proliferation, NF-κB activation, enhanced production of IL-10 and improving microbial homeostasis.
In healthy mice L. helveticus induced Treg, which regulate immune and inflammatory responses, suppressed the production of IL-17, IL-4 and IL-10 and induced the anti-inflammatory cytokine transforming growth factor-β1in Peyer's patch cells. Intraperitoneal and oral administration of a L. helveticus strain strongly alleviated symptoms of collagen-induced arthritis (CIA) in mice for the ability to decrease the abundance of immune cells and the subsequent production of collagen type II (CII) specific antibodies and IL-6.
In clinical studies, selected strains of L. helveticus prevented the anxiety-like behavior and negative effects on memory induced by a Western-style diet and improved behavioral, cognitive, and biochemical aberrations caused by chronic restraint stress in rats correlated with lower plasma corticosterone, adrenocorticotropic hormone (ACTH), and IL-10 levels in plasma, restoring hippocampal serotonin and norepinephrine levels. In RCT trials, L. helveticus strains modulated mucosal and humoral immunity by increasing specific IgG, and IgA, reduced the duration of respiratory infections and alleviated perennial allergic rhinitis in adults by supressing eosinophils.
6.1. Amelioration of intestinal health
Ulcerative colitis (UC) is a non-specific chronic inflammatory disease causing colon and rectum lesions, bloody diarrhea, abdominal pain, weight loss, and ulcer. It is comonly treated with 5-aminosalycilates, glucocorticoids and immunosuppressants, that only alleviate the clinical symptoms and have side effects. With the aim to develop a safer and more efficient UC treatment agent, EPS-1 produced by L. helveticus KLDS1.8701 was administered to C57BL/6 mice for 21 days and DSS colitis was induced at day 15. EPS-1 administration could effectively alleviate induced UC by reversing weight loss and colon shortening. After being treated with EPS-1, the colonic tissues showed reduced structural damage and inflammatory cell infiltration. Levels of TNF-α, IL-1β, and IL-6 of the EPS-1 group were lower and of IL-10 higher in colon in EPS-1 treated animals. The expression levels of ZO-1, Occludin, Claudin-1, and MUC2 were significantly reversed by the EPS-1 administration. EPS-1 administration significantly increased the level of acetate and butyrate SCFAs in colon [47].
Inflammatory bowel disease (IBD) comprises a range of chronic inflammations and disorders of the gastrointestinal tract, the most common forms of IBD being Crohn's disease (CD) and ulcerative colitis (UC). Anti-inflammatory agents are traditionally used to treat IBD, but alternative therapies, including probiotics, are evaluated. L. helveticus ASCC 511 (LH511) was obtained from the Dairy Innovation Australia Limited (ASCC, Werribee, Victoria, Australia). Citrulline is a non-essential amino acid that was found to ameliorate the inflammatory response by reducing the level of pro-inflammatory cytokines TNF-α, IL-6 and IFN-γ in intestinal ischemia/reperfusion model. Citrulline is produced simultaneously by nitric oxide synthase (NOS) during NO formation from arginine, and then arginine is re-generated from citrulline by argininosuccinate synthetase (ASS) and argininosccinate lyase (ASL). Some lactobacilli are able to utilize arginine by the arginine deiminase (ADI) pathway. The potential enhancement effect of combination of lactobacilli with arginine in vivo has been investigated, and a synergistic effect was shown to increase protection against liver injury induced by D-galactosamine, high-cholesterol diet and endotoxin, as well as to reduce bacterial translocation. In male C57BL/6J mice with colitis induction for 5 days with DSS, fermented milk containing L. helveticus ASCC 511 (LHFM) or L. helveticus ASCC 511 plus citrulline (Cit_LHFM) was supplied for 12 days. All milk treatment groups attenuated the weight loss. In the LHFM and Cit_LHFM the survival rate increased to 50% and 70% at day 12, respectively. Cit_LHFM improved the colonic permeability as shown by the reduced concentration of the fluorescent drug FITC-dextran administered to mice in blood. Treatment with Cit_milk, LHFM and Cit_LHFM reduced the damage in distal colon mucosa and showed the lowest score. LHFM and Cit_LHFM treatments decreased disease activity indexes, limited colon shortening and reduced the ration between colon weight and length (CW:CL). Treatment Cit_LHFM suppressed the expression of IL-4 and IL-17 induced by DSS in serum and all treatments reduced the expression of IL-6, TNF-α and IFN-γ. LHFM upregulated the expression of IL-10. The highest remediation effect, on the mRNA levels of all tight junction (TJ) proteins was observed in Cit_LHFM. Therefore, treatment by Cit_LHFM showed synergistic and superior effects on reinforcing intestinal structure and reducing inflammation [48].
6.2. Immunomodulation
Li et al. aimed to identify the most efficient bifunctional strain among 265 LAB from fermented foods based on immunomodulatory activity and fermentative ability. One selected strain, identified as L. helveticus WHH2580, was used for milk fermentation and the fermented milk was administered to specific pathogen-free male BALB/c mice that on days 28 and 29 of the administration period, mice were submitted to immunosuppression with CTX for 2 days. No significant difference was observed in the body weight between groups receiving fermented milk group or non fermented milk throughout the administration period. In the fermented milk group, a higher splenic lymphocyte proliferation and a stronger splenic NK cell cytotoxicity activity were observed [49].
6.3. Allergy alleviation
L. helveticus SBT2171 (LH2171) showed an ameliorative effect on nasal and ocular symptoms of patients with mite and house dust allergy. Therefore, a study was carried out using a previously established murine model for pollen, ovalbumin (OVA)-specific TCR-transgenic DO11.10 female mice whose T cells recognize OVA323-339. Mice inhaled OVA/saline for 1 h, twice a week for six weeks while freeze-dried L. helveticus LH2171 was given to the mice such that they could access it freely during the period of pollen allergy induction. It was observed that face scratching and sneezing were significantly reduced by L. helveticus LH2171 administration. Since the treatment did not suppress IgE production, a mechanism other than IgE inhibition was implied [50].
House dust or indoor dust is a mixture of mites, ticks, human scraps, mold, and bacteria with mites Dermatophagoides farinae and D. pteronyssinus as the most common allergen source. Allergic symptoms include runny nose, sneezing and ocular symptoms, such as itching eyes and eye swelling. Yamashita et al. [51] investigated in a randomized, double-blind, placebo-controlled, parallel group comparative study whether drinkable yoghurt (DY) containing L. helveticus LH2171 improved QOL in the subjects with daily nasal and ocular discomfort and immune markers in the blood. Active DY, containing L. helveticus LH2171, was fermented also by a strain of S. thermophilus, while only the latter was present in the placebo DY. Total LAB number was about 109 CFU/g and the L. helveticus LH2171 number was 107 CFU/g at the end of the consumption period.
Decreases in total scores of nasal and ocular discomforts at week 8 were significant in theL. helveticus LH2171 group and even more on week 12. Outdoor activity and sleep scores were significantly improved in the L. helveticus LH2171 group.
The number of sneezes also decreased significantly but not total and antigen-specific IgE levels at any experimental point. No serious adverse or side effects were noted by the responsible physician.
Allergy caused by house dust affects approximately 40% of the global population in the industrialized world and the major mechanism of the allergic response is the activation of T helper type Th2 cytokine production, which induces the production of antigen-specific immunoglobulin IgEs by B cells. Pharmacotherapy for perennial allergy, includes use of inhibitors of chemical mediators, anti-histaminic drugs, and topical steroids that can have long-term side effects. Monoclonal antibodies anti-human IgE or receptor antagonists tha regulate the signaling IL-4 and IL-13 pathways are expensive alternatives used to treat particular conditions. Noticeably, approaches that can completely cure allergies still lack [52]. Following the study of Yamashita et al. [51] on the effect of L. helveticus LH2171 in house dust allergy, another RCT was conducted to clarify the anti-allergic mechanism of and evaluate its effect on the QOL with 16 weeks test period and use of the Profile of Mood States 2 questionnaire. Patients involved were 20–65 years old males and females, who did not have chronic diseases and presented allergic rhinitis symptoms with no significant differences between groups in baseline symptoms.
In the L. helveticus LH2171 group, the severity of allergic rhinitis significantly improved in 4 - 16 weeks with significantly improved aspect of the nasal mucus, improved aqueous secretion and stuffy nose scores. The eosinophil score in the nasal fluids was significantly lower at 4 and 12 weeks. There were no significant differences in allergen-specific IgE and total IgE levels in the serum but the level of thymus and activation-regulated chemokine (TARC) was significantly lower at eight weeks in the L. helveticus LH2171. Eye itching significantly improved in the L. helveticus LH2171 group and the severity of nasal symptoms decreased in 16.0% subjects in the placebo group and in 36.8% subjects in the L. helveticus LH2171 group. It was concluded that the probiotic strain ameliorated nasal discomfort probably by suppressing eosinophils [52].
6.4. Infection mitigation
Defensins are one of the major groups of antimicrobial peptides in mammals and play critical roles in host defense. β-Defensins, produced in epithelial cells, are widely distributed in the body and strains of lactic acid bacteria have been reported to induce their expression. Since β-defensins are effective against P. gingivalis, an oral bacterium that contributes to the development of periodontal disease, increased expression of β-defensins can suppress periodontal disease. Therefore, Kobatake et al. [53] investigated the expression of β-defensins in female BALB/c mice orally administered with 109 CFU daily of L. helveticus SBT2171 for 5 weeks. Three weeks later, 108 CFU of P. gingivalis strain 381 was inoculated in oral cavity of the mice daily for 2 weeks. On day 30, alveolar bone loss was significantly suppressed in the L. helveticus group and the abundance of P. gingivalis-specific 16S rRNA isolated from gingival tissue was reduced. Furthermore, mRNA levels of Tnfα in gingival tissue were lower in the L. helveticus group on day 1. Therefore, oral administration of L. helveticus SBT2171 ameliorated gingival inflammation caused by P. gingivalis Pg381.
L. helveticus SBT2171 was administered to Sprague Dawley female rats of specific pathogen-free grade in amount of 109/CFU daily for 35 days, and 108/CFU/mL of Aggregatibacter actinomycetemcomitans HK1651 periodontitis agent were given orally from the 21st day, daily for 14 days. It was found that L. helveticus SBT2171 reduced the distance from the cement-enamel junction (CEJ) to the alveolar bone crest (ABC), indicating improved alveolar bone resorption. In addition, neat arrangement of periodontal fibers, and small blood vessels proliferation in connective tissue were observed. These results showed that L. helveticus SBT2171 attenuated damage to the hemimaxillary tissue of in periodontitis determining reduced inflammatory cell infiltration. The the number of A. actinomycetemcomitans in the L. helveticus SBT2171 group was notably decreased, as well as the expression in the gingival tissue of IL-1 β, IL-6, which significantly increases in patients with periodontal disease, and TNF- α, involved in chronic inflammation that leads to periodontal tissue damage. Moreover, the expression level of β-defensins significantly increased in the gingival tissues. Therefore, L. helveticus SBT2171 improved periodontal disease by increasing the expression of β-defensins and reducing the number of A. actinomycetemcomitans[54].
Common carp, Cyprinus carpio, is widely cultivated in the world but its farm production is challenged by infectious agents such as Aeromonas hydrophila and use of antibiotics leads to unwanted side effects such as immunosuppression, increasing of pathogen multi-drug resistance and health risks for consumers. Gum Arabic (GA) is a natural prebiotic secreted from Acacia senegal and Sengalia senegal trees and is a safe feed additive in food. It contains non-digestible glucuronic acid, rhamnose, and galactose which can be used as an energy source for gut bacteria and can be converted by the gut microbiota in SCFAs. In the study of Yousefi et al. [55] the L. helveticus type strain ATCC 15009, isolated from Emmental cheese, was administered in different combinations with GA to common carp juveniles that were fed ad libitum with different diets C (basal diet), LH1 (107 CFU/g), LH2 (109 CFU/g), GA1 (0.5% w/w), GA2 (1% w/w), LH1+GA1, and LH2+GA2, three times daily. After two weeks, fishes were transferred to challenge tanks and exposed to A. hydrophila via intraperitoneal injection. Final weight and weight gain rate were significantly higher in the fish fed diets containing prebiotics GA1 and GA2, and synbiotics GA1+LH1 and GA2+ LH2 than the control group and the highest value was obtained in the LH1+ GA1 treatment. Dietary GA and/or LH supplementation significantly increased the population of LAB in the fish intestine with no significant difference among the treatments. The treatments enhanced white blood cells, with a peak for LH1+GA1. Monocyte percentage was significantly greater in LH1 and synbiotics with also an increase of total serum Ig levels and alternative complement ACH50 activity. Alkaline phosphatase (ALP), lysozyme and Ig increased in skin mucosa. GA and/or LH diets significantly increased serum SOD and CAT activities and reduced MDA, except for LH2 treatment. The mortality rate of common carp infected with A. hydrophila at the end of the infection period showed significant decreases in all experimental groups with lowest levels for GA2. The synbiotic with lower LH and GA improved growth rate and disease resistance as efficiently as higher concentrations in carps.
6.5. Alleviation of aging damages
Strain L. helveticus KLDS1.8701 with high antioxidant capacity, was administered to mice receiving D-galactose once daily for 8 weeks to induce aging. L. helveticus KLDS1.8701 supplementation significantly decreased organic index, liver injury and endotoxin and reduced hepatic oxidative stress by modulating the Nrf-2 pathway. L. helveticus KLDS1.8701 restored the gut microbiota with increased butyrate production and decreased endotoxin production indicating that attenuation of hepatic oxidative stress was induced by the metabolites of gut microbiota [56].
The EPS purified from the culture of L. helveticus KLDS1.8701 exhibited strong scavenging properties on 2,2-diphenyl-1-picrylhydrazyl radical, superoxide radical, hydroxyl radical, and chelating activity on ferrous ion. When supplemented in vivo, the fraction EPS-1 significantly alleviated D-galactose induced liver damage by decreasing organic index, liver injury and liver oxidative stress. EPS-1 supplementation shifted the gut microbiota composition to that of the control group indicating that the mitigation of hepatic oxidative stress depended on the action on gut microbiota [57].
6.6. Improvement of stress symptoms
Ingestion of 0.2 ml of 109 CFU/ml of L. helveticus WHH1889 isolated from yogurt for five weeks in an induced chronic unpredictable mild stress (CUMS) mouse model of depression reversed the elevated level of serum corticosterone and decreased levels of hippocampal 5-hydroxytryptamine (5-HT) and its precursor 5-HTP. Moreover, the L. helveticus treatment significantly improved depressive and anxiety behaviors in mice. Furthermore, the intestinal microbiota diversity, reduced in CUMS mice, was improved by L. helveticus WHH1889, with increased colonic 5-HTP level and tryptophan hydroxylase 1 Tph1 gene expression. The findings indicated that the L. helveticus WHH1889 antidepressant-like effects were associated with the modulations of the 5-HT/5-HTP metabolism and gut microbiome. Similar results were obtained with the yogurt isolate L. lactis WHH1889, thus showing that different dairy isolates can relieve the symptoms of stress-induced depression [58].
6.7. Benefits in animal production
The intestinal health of fish is closely related to the immune function, so improving the intestinal immunity of fish is of great significance to aquaculture. Therefore, L. helveticus HML037, isolated from Inner Mongolia Koumiss, was evaluated as potential dietary supplement for M. anguillicaudatus fish by analyzing the effects on the intestinal health. The strain was administered in alginate-coated capsules containing 107, 108 and 109 CFU for 8 weeks. The groups receiving L. helveticus HML037 had significantly improved growth rate, feed conversion ratio, condition factor and survival rate. Intestinal digestive enzymes proteases, lipases, and amylases, that promote the absorption of nutrients, showed higher activities in the fish fed with L. helveticus HML037, mainly in the middle L. helveticus HML037 dose of 108 CFU/g. Moreover, L. helveticus HML037 treatments significantly increased the activity antioxidant enzymes LZM, SOD, AKP, and CAT and decreased the MDA levels. After 8 weeks TLR1, GH and IGF-1 genes were significantly up-regulated with the most significant effect the middle L. helveticus HML037 dose. The Shannon and Simpson diversity index in metagenome analysis of the intestinal microbiota indicated a higher community diversity in the L. helveticus HML037 groups with overrepresented Pseudomonas, Flavobacterium, Actinomyces, Azoarcus and Lactobacillus genuses [59].
7. Lactococcus lactis
Lactococcus lactis is one of the most common SLAB species used in the cheesemaking sector and naturally occurring in cheeses whose production implies use of process temperatures favoring mesophilic microorganisms [16]. The effects reviewed for Lactococcus lactis by Illikoud et al. [4] include prevention of DSS-induced colitis in mice, with prevented release of NO and of inflammatory factors induced by lipopolysaccharides (LPS) in RAW264.7 cells, inhibition of Salmonella invasion into intestinal epithelial cells, production of an EPS found to increase macrophage phagocytosis, spleen and thymus indices and haemolytic complement (HC(50) activity in cyclophosphamide-immunosuppressed mice. In clinical trials, milk fermented with L. lactis activated plasmacytoid dendritic cells, enhanced the ability to produce interferons, reduced symptoms such as sneezing and cumulative days of fatigue in athletes and mitigated the severity of atopic dermatitis in children [4].
A recent review by Saleena et al. [60] summarized the in vivo immunological effects demonstrated for L. lactis strains of dairy origin and these included a reduction of IL-4 production in ovalbumin-sensitized mice, amelioration of recurrent colitis in mice, increased NK cell activities, concanavalin A-induced T cell proliferation, and serum levels of tumour necrosis factor (TNF)-α, IFN-γ, IL-2, IL-4, IL-10, and IL-12 in cyclophosphamide immunosuppressed mice, antihypertensive effect of the fermented milk in spontaneously hypertensive rats, stabilization of cancer cells in the colorectal epithelium, restored T cell population in small intestinal lamina propria in aged interleukin-18 deficient mice and exerted protection against influenza virus infection. A clinical trial demonstrated that L. lactis ameliorated acute psychological stress in men.
7.1. Anti-cancer effects
Barcellos Jaskulski et al. [61] showed that a suspension of 108 CFU/mL of L. lactis R7, isolated from ricotta and able to produce nisin, exerted anti-carcinogenic potential against colorectal epithelium cancer cells induced in Wistar rats with 1,2-dimethylhydrazine (DMH) when administered for 20 days. L. lactis R7 prevented the formation of cells with malignant atypia and of large infiltrate of lymphocytes in most animals, demonstrating an improvement of the immune response. Determination of hematic glutamic-oxalacetic transaminase (GOT) and glutamic-pyruvic transaminase (GPT) levels indicated that L. lactis R7 reduced hepatic cell damage caused by the oxidative effect of DMH.
7.2. Immunomodulation
The immunostimulating effects and associated mechanisms of L. lactis GCWB1176 isolated from mozzarella cheese were investigated in cyclophosphamide-induced (CTX-induced) immunosuppressed male ICR mice. L. lactis GCWB1176 was administered intragastrically at 107 and 109 CFU daily for 16 days. Mice were treated on days 7–9 by intraperitoneal injection of CTX,. The body weight, spleen and thymus indices and the cytokine TNF-α, IFN-γ, IL-2, IL-4, IL-10 and IL-12 levels in serum of the L. lactis GCWB1176-treated group increased dose-dependently. However, all doses of L. lactis GCWB1176 enhanced NK cell activity and lymphocyte proliferation, indicating that L. lactis GCWB1176 can alleviate CTX-induced immunosuppression [62].
7.3. Improvement of stress symtoms
L. lactis WHH2078 isolated from yogurt was administered in numbers of 109 CFU/mL in mice with induced chronic unpredictable mild stress (CUMS) for 5 weeks and significantly ameliorated depressive and anxiety-like behaviors in the tail suspension test, forced swim test, sucrose preference test, and open field test. These effects were associated to a significantly reduced serum corticosterone level and restored levels of 5-hydroxytryptamine (5-HT, serotonin) and its precursor 5-hydroxytryptophan (5-HTP), and brain-derived neurotrophic factor. The gene for tryptophan hydroxylase Tph1 involved in synthesis of 5-HT, whose metabolism associated to depression, was upregulated in colon. CUMS was associated to colon dysbiosis. The high-throughput 16S rRNA gene sequencing of feces microbiota showed that L. lactis WHH2078 restored alpha diversity and the abundances of Firmicutes and Bacteroidetes, which is associated with the improvement of 5-HT metabolism [63].
7.4. Improvement of depressive behaviour
The interaction between the intestinal microbiota, the gut and the central nervous system (CNS), known as the brain–gut–microbiota axis, is increasingly studied for the possible applications in the treatment of neuropsychiatric disorders. Several preclinical studies have shown that probiotics may be used to modulate depressive-like behaviours. L. lactis subsp. cremoris LL95, a LAB isolated from cheese was investigated on this respect by supplying daily 109 CFU of it to female C57BL/6 mice for 28 days. On day 28, the L. lactis LL95 group showed an increased number of LAB CFU that was 1.4 × 109 per 100 mg of faeces vs 6.0 × 108 in the control. The weight of the hippocampus increased by 33% in female mice fed with L. lactis LL95 and a significant decrease in hippocampal ROS levels and an increase in the Ferric Reducing Antioxidant Power (FRAP) was observed in LL95-fed mice. Treatment with L. lactis LL95 resulted in a decrease in immobility time in the tail suspension test (TST), Forced Swim Test (FST) and immobile episodes and increase of latency to immobility with a reduction of depressive-Like behaviour in mice. The administration of LL95 did not affect locomotor activity in the open field test (OFT) and in the Elevated Plus Maze (EPM), suggesting anxiolytic effects [64].
8. Latilactobacillus sakei
8.1. Infection mitigation
It has been observed that Fulani tribe of Cameroon are more resistant to malaria than other West Africa ethnic groups. Fulani have a tradition of milk production, processing and consumption and this has been considered a possible explanation to resistance to malaria. Moreover, also in Sudan cow's milk fermented over a long period and added to another product (oka) to obtain Biruni, is traditionally used in the treatment of malaria.
Latilactobacillus sakei UB27 isolated from traditionally fermented milk was administered to Balb/c mice infected with the chloroquine sensitive Plasmodium berghei ANKA. After 7 or 14 days, a gradual and significant dose and duration of treatment dependent reduction of the level of malaria parasitaemia was observed in the L. sakei group. A level of 2.7×109 CFU/mL determined 100% parasitemia suppression on day 20 and prolonged survival of treated mice [65]
9. Limosilactobacillus
9.1. Alleviation of aging damages
Limosilactobacillus fermentum is one of the most common cultivable and predominant microbes in fermented dairy products. Strain L. fermentum strain MBC2 was isolated from Mozzarella di Bufala Campana (MBC) traditional Italian PDO (Protected Designation of Origin) cheese. In this cheese L. fermentum is one of the main microbiota components together with L. delbrueckii and Leuconostoc lactis. This strain was evaluated for effect on longevity using wild-type C. elegans strain N2 and CL2166 (dvIs19 [(pAF15)gst-4p::GFP::NLS] III), pept-1(lg601) and CF1038 daf-16(mu86) mutant ss living hosts. C. elegans is a small soil nematode used as an animal model for probiotic evaluation because it is easy to treat, has a short lifespan, can be safely used in the laboratory and propagates through self-fertilization. It shares two-thirds of the genes related to human diseases and is used in studies on longevity, immunity, neurodegenerative diseases, fat storage, DNA damage and apoptosis. Its simple immunity systems facilitates studies on the immune signalling pathways [Kwon et al.]. Worms were propagated on peptone-free nematode growth medium (NGM) and fed with a normal diet constituted of 10 mg of E. coli OP50 or with L. rhamnosus GG and L. fermentum separately. The wild type nematodes fed with L. fermentum MBC2 showed 50% viability at day 18, while for the controls fed with L. rhamnosus GG and E. coli OP50 50% viability was recorded at day 11 and 14, respectively, thus indicating a significantly extended lifespan for the L. fermentum MBC2 group. This effect was not observed with L. fermentum MBC2 heat-killed at 65 °C for 90 min. The intestinal microbial count of L. fermentum MBC2 group at 5 days was 2-fold and 4-fold higher than that of E. coli OP50 and L. rhamnosus GG, respectively. Moreover, nematodes fed with L. fermentum MBC2 displayed a higher motility from 6 to 9 days and a higher pharyngeal pumping frequency, an indicator of food intake ability that declines with age, at 13 days. Intracellular lipofuscin, which is a marker of aging cellular damage and is determined by autofluorescence, was lower in the group fed with L. fermentum MBC2.
Dysregulation of lipid metabolism induced by aging, determined by BODIPY staining, detected lower amounts of intestinal lipid droplets in L. fermentum MBC2 and L. rhamnosus GG-fed worms. Extended longevity was shown to correlate with enhanced resistance against oxidative damage. In transgenic C. elegans with genotype GST4::GFP, defective in GST-4, a glutathionyl S-transferase with highest activity against lipoperoxides, microscopy analysis showed that fluorescence intensity of L. fermentum MBC2-fed worms was notably lower than that of OP50- and LGG-fed animals, indicating that in L. fermentum MBC2-fed worms the production of reactive oxygen species (ROS) was effectively reduced compared to control worms at 1 and 13 days of adulthood. In the C. elegans PEPT-1 mutant, defective in one of the major regulators of fat content, L. fermentum MBC2 did not induce a pro-longevity effect, though ROS levels were reduced [66].
L. fermentum DR9, isolated from raw cow’s milk, was administered to male Sprague Dawley rats subjected with D-galactose induced aging. Rats receiving L. fermentum DR9 had higher Firmicutes/Bacteroidetes ratio, with genuses Lactobacillus, Blautia significantly higher and Prevotella, Bacteroides and. Erysiperotrichaceae were significantly lower than positive controls receiving only D-galactose. Indicating the ability to restore the phylum composition during the aging process. Aged rats receiving L. fermentum DR9 showed an higher amount of fecal 5-oxoproline, a product of GSH metabolism, that showed an active antioxidant activity [67]
9.2. Infection mitigation
L. reuteri strain 2892 isolated from camel's milk exerted protective effects against H. pylori-induced gastritis in the stomach tissue of in C57BL/6 mice. A pretreatment with this strain significantly downregulated the H. pylori virulence factor cagA gene expression, upregulated the tight junction molecules [zona occludens (ZO-1), claudin-4] and suppressed metalloproteinase (MMP)-2 and MMP-9. Moreover, L. reuteri 2892 reduced the serum concentrations of pro-inflammatory cytokines interleukin (IL)-6, IL-1β, and INF-γ and increased the anti-inflammatory cytokine, IL-10. In addition, it showed anti-oxidative stress activity by regulating the levels of SOD and MDA, suggesting that L. reuteri 2892 can attenuate H. pylori-induced gastritis [68].
10. Pediococcus acidilactici
10.1. Amelioration of metabolic syndrome
Diabetes mellitus is a life-threatening metabolic disorder that affects globally hundreds million adults and causes millions of deaths annually. Inhibition of the carbohydrates-hydrolyzing enzymes α-glucosidase and α-amylase is the most efficient intervention to reduce the postprandial glucose level in blood in diabetes treatment. Since most of the chemical compounds and protein extracts able to inhibit these enzymes are partially effective and have side effects, Al-Emran et al. [69] examined the possibility to select and evaluate the in vivo potential of probiotics of dairy origin to inhibit α-glucosidase and α-amylase. Among 77 lactic acid bacterial isolates from 20 different milk samples, four isolates inhibited both enzymes. These were identified as Pediococcus acidilactici on the basis of the pheS gene and whole genome sequence. Analysis of genome sequences by PathogenFinder indicated a nonpathogenic probability value between 75.5% and 81.2% and the presence of antibiotic resistance (AR) genes in two isolates. All four isolates were used for in vivo trial of hypoglycemic effect in Swiss Webster male mice to which about 109 CFU of the bacteria were administered daily by oral gavage for 6 weeks. Diabetes was induced after the third week with a single dose of Streptozotocin in all mice except the negative control (NC) group. A significant drop of blood glucose was observed in groups receiving three of the isolates compared with the positive control (PC) that did not receive any treatment.
In the lipid profile a significant decrease of LDL was observed in all probiotic groups, even compared to the NC group, while HDL, TG and total cholesterol slightly decreased. Body weight was significantly lower in all probiotic groups compared to all controls, also the one receiving metformin as standard diabetes treatment (PCT).
10.2. Defense from pollutants
A high heavy metal (HM)-resistant strain, P. acidilactici GR-1, isolated from yak milk, significantly alleviated chromate toxicity in mice. Therefore, it was tested in a clinical trial to investigate the potential of the strain to reduce toxic metal levels in workers of the metal mining and smelting of copper (Cu), Ni, and cobalt industry in Gansu province who had elevated levels of Cu and Ni in the urine. The participants were randomly devided in a group receiving yogurt containing 1010 CFU L. delbrueckii subsp. bulgaricus and S. thermophilus and another receiving yogurt added with P. acidilactici GR-1 in the same number for 12 weeks. The exclusion criteria were use of antibiotics within 1 month before the study, regular use of probiotic supplements, lactose intolerance, alcohol or drug abuse and presence of comorbidities. The participants were stratified according to blood Cu and Ni levels in three sub-groups with low, medium, and high levels of heavy metals. The high-level and medium-level groups of participants who consumed probiotic yogurt showed significantly decreased blood Cu levels after 4 weeks and 8 weeks. Similarly, in the high- and medium-level sets, a significant decrease in blood Ni levels was observed earlier in the probiotic yogurt group than in the conventional yogurt group. In the low-level set, no significant decrease was noted in blood Cu or Ni levels. The fecal samples showed elevated Cu and Ni levels compared with the baseline values after 12 weeks of the intervention, indicating increased HM excretion with yogurt ingestion. With the elimination of HM from the body, also oxidative stress (OS) and inflammation indicators in serum, including catalase (CAT) activity, anti-inflammatory IL-10 and IL-4 levels, and proinflammatory IL-6 and IL-1β levels, improved after 12 weeks of probiotic yogurt intake. SCFAs acetate, propionate and butyrate significantly increased in feces of the probiotic yogurt group possibly for the enrichment of SCFA-producing bacteria. After 4 weeks of probiotic yogurt consumption, P. acidilactici GR-1 was significantly enriched in the fecal samples of the participants and remained significantly higher after the completion of the trial, indicating good adherence and colonization capacity in the intestine. Metagenomic shotgun sequencing revealed that the fecal microbiota the worker group that consumed probiotic yogurt was enriched in KEGG orthologs involved in OS response, including SOD, CAT, GPX and glutathione reductase (NADPH), suggesting the induction extensive activation of HM-induced OS defense mechanisms. After the probiotic yogurt intervention ascorbate, aldarate, taurine and hypotaurine metabolism were enriched in the fecal metabolome, while GSH metabolism and pantothenate and CoA biosynthesis were enriched in the serum metabolome. Histidine, α-linolenic acid, and D-glutamine and D-glutamate metabolisms were also enriched. Each of the enriched antioxidation-related metabolites correlated positively with most of the enriched species in feces.
An animal experiment with Kunming mice administered with a broad-spectrum antibiotic that later received CuSO4 at a sublethal concentration, and the probiotic bacterial strain P. acidilactici GR-1 at a dose of 109 CFU daily it was observed that the remediating effect of P. acidilactici GR-1 was reduced thus demonstrating that the integrity of indigenous gut microbiota was essential for the protective effects [70].
11. Propionibacteria
Dairy propionibacteria (PAB), grow in cheese after lactic acid bacteria (LAB) since they can use lactic acid as main carbon source to produce propionic acid, acetic acid and carbon dioxide. PAB synthesize beneficial molecules including B9 and B12 vitamins and propionic acid that promotes gut health [4]. Among PAB Propionibacterium freudenreichii and Acidipropionibacterium acidipropionici, formerly P. acidipropionici, are included in the updated list of microorganisms with qualified presumption of safety (QPS) status of the European Food Safety Authority (EFSA) (https://zenodo.org/record/6902983#.ZDgW_fZByUk, Accessed on 13 April 2023) and the species P. freudenreichii is most frequently isolated from dairy products [71].
Illikoud et al. [4] reported that P. freudenreichii strains administered to mice in different studies protected from TNBS and DSS-induced colitis and colitis caused by the pathogen Citrobacter rodentium attenuating weight loss, oxidative stress and histopathological scores. These effects could depend on increased immunoglobulin A (IgA) secretion in the small bowel, increased expression of IL-10, restored occludin gene expression and reduced induction of TNFα, IFNγ and IL-17. Similar effects on cytokine induction were observed in mucositis, a painful inflammation and ulceration of the mucous membranes, occurring as adverse effect of chemotherapy and radiotherapy cancer treatments and induced in mice with the chemiotherapic 5-fluorouracile (5-FU). P. freudenreichii CIRM-BIA129, that is the type strain of the species ATCC 9614 of unknown but, most probably, dairy isolation source, when administered in cheese showed a high immunomodulatory effect.
P. freudenreichii KCTC 1063 isolated from Swiss type cheese protected rats from DSS-induced colitis by stimulating the expression of MUC2 or mucin 2, a glycoprotein that forms an insoluble mucous barrier on the gut epithelium, in intestinal goblet cells. This effect is explained with the production of propionate, the most effective SCFAs in MUC2 production stimulation. In a clinical pilot trial involving ulcerative colitis patients, P. freudenreichii ET-3 improved the clinical activity index and the endoscopic index. Moreover, consumption of milk fermented by P. freudenreichii JS reduced the serum level of high-sensitivity C-reactive protein (hsCRP) in a RCT in healthy adults [4].
11.1. Amelioration of metabolic syndrome
Although the alteration of the gut microbiota in obesity is not clearly defined, dysbiosis occurs in obesity and main species of the gastrointestinal microbiota and their beneficial metabolites, such as SCFAs, vitamin B12, and indole, are lost, and intestinal permeability and endotoxemia are increased, which induces inflammation and gluconeogenesis in the liver, decreases satiety in the brain and increases TG incorporation in adipose tissues. In addition, increased gut permeability maintains low-grade chronic inflammation. Among probiotics studied to improve obesity, P. freudenreichii was considered by An et al. [72] for its capacity to produce bifidogenic compounds that promote the growth of Bifidobacterium spp. in gut, vitamin B12, propionic acid, and surface proteins with immunomodulatory properties. Therefore, they evaluated the effect of live P. freudenreichii MJ2 or hkMJ2 on the prevention of lipid accumulation in adipocytes and its antiobesity and antidiabetic activity in high-fat diet (HFD)-induced obese mice. Both forms of the probiotic decreased body weight gain and liver and epididymal white adipose tissue weights in high-fat diet (HFD)-induced obese mice by 31% and 22.8%, respectively. The food efficiency ratio (FER) of the MJ2 and hkMJ2 groups significantly decreased. The expression levels of genes and proteins related to adipogenesis and lipogenesis significantly decreased, while the expression level of genes related to lipolysis (HSL and ATGL) and fatty acid β-oxidation (CPT-1α and ACOX1) increased in both P. freudenreichii MJ2 and the hkMJ2-treated groups. In addition, blood glucose and fasting insulin levels decreased in both groups with ameliorated insulin resistance, thus showing that P. freudenreichii MJ2 can alleviate obesity and metabolic syndrome.
11.2. Alleviation of aging damages
P. freudenreichii KCTC 1063 was shown to extend the lifespan of C. elegans compared with E. coli OP50 standard diet by Kwon et al. [73]. The effect was observed in daf-16, jnk-1, skn-1 or daf-7 loss-of-function mutants but not in pmk-1, sek-1, mek-1, dbl-1, daf-12 or daf-2 mutants, which suggests potential roles for these genes in P. freudenreichii-induced longevity in C. elegans. The mean life span of P. freudenreichii-fed C. elegans, compared with that of E. coli OP50-fed worms, increased by approximately 13% and the P. freudenreichii-fed worms were significantly smaller than the E. coli-fed worms. Genes related to lifespan extension and immune response, including genes involved in the p38 MAPK and the TGF-β pathways, were significantly up-regulated. Moreover, P. freudenreichii increased resistance against a human pathogen, Salmonella Typhimurium, through the activation of skn-1, which is involved in pathogen resistance in C. elegans.
11.3. Infection mitigation
Propionibacteria were tested for pathogen exclusion potential. It was reported that in poults raised until 7 weeks of age P. freudenreichii B3523 strain of dairy origin reduced the cecal colonization of Salmonella Heidelberg and this was observed also in finishing turkeys of 12 weeks of age supplemented with 1012 CFU in 18.9 l (5 gallons) water of P. freudenreichii B3523 and challenged with 108 CFU/turkey Salmonella Heidelberg at 11 weeks. In these animals cecal colonization of the pathogen was reduced by more than 2.0 log10 CFU/g and its dissemination in the spleen decreased [74].
Subsequently, Nair et al. [75] administered this strain to turkeys by adding 1010 CFU of it to 18.9 l of the drinking water until 7 or 12 weeks of age. Then challenge with Salmonella Heidelberg was carried out by administering it in numbers of 106/108 CFU to each bird at week 6 or 11 in two different experiments. Cecal microbiome analysis carried out by Illumina MiSeq showed that the relative abundance of Firmicutes on week 12 was significantly higher in the P. freudenreichii group. In the group administered with only Salmonella Heidelberg genuses Streptococcus, Gordonibacter, and Turicibacter, known to be associated with inflammatory responses in birds and mammals, showed higher relative abundance.
Genuses with increased abundance in the P. freudenreichii group two days post infection in growing turkeys were Subdoligranulum and Faecalibacterium, while the Streptococcus genus abundance decreased. Seven days post infection no differentially abundant genuses were identified.
In finishing turkeys, the abundance of Lactobacillus and Ruminococcaceae NK4A214 and UCG.010 increased two days post infection, while seven days post infection genuses with increased abundance were Lactococcus, Erysipelatoclostridium, Leuconostoc and Butyricicoccus, indicating that P. freudenreichii supplementation increased the relative abundance of beneficial microbiota that could mitigate the carriage of Salmonella Heidelberg in turkeys.
11.4. Improvement of bone health
The effects of probiotics on bone health are currently studied since an association was found between gut microbiota and osteo-immunology. Therefore Yeom et al. [76] evaluated the effect of P. freudenreichii MJ2, a probiotic with anti-inflammatory properties isolated from raw milk, on osteoblast differentiation, mineralization, and related signaling pathway in an osteoporosis mouse model. Osteoporosis is a condition in which bone mass decreases and bone structure deteriorates. This condition occurs with higher frequency in postmenopausal women because of the decrease in estrogen levels. Estrogen and progesterone therapy has as side effect an increased risk of breast cancer. Therefore, less harmful treatments to delay osteoporosis progression are highly desired. The evaluation of the effect of P. freudenreichii administration on this pathological condition was prompted from the observation that this bacterium increased the mRNA levels of genes homologous to vertebrate BMP-5 and BMP-8, which function in the development of cartilage and bone, respectively, in C. elegans.
The animal models used were ovariectomized (OVX) rats to which live P. freudenreichii MJ2 and heat killed cells (hkMJ2) were administered. After 16 weeks of treatment live/dead MJ2 administration did not show hepatotoxicity. Bone Mineral Density (BMD) of the femur in OVX rats treated with hkMJ2 increased significantly compared to the control, showing prevention of bone loss. This effect was attributed to heat stable surface proteins that were found to increase the levels of mRNAs for osteoblast differentiation-regulated genes and osteoblast mineralization in the human fetal osteoblast cell line hFOB 1.19.
P. freudenreichii MJ2 strain was also evaluated for a possible effect on the improvement of rheumatoid arthritis (RA) symptoms [77]. In this disease osteoclast differentiation is crucial for bone absorption, and osteoclasts are involved in bone destruction. P. freudenreichii MJ2 was found to decrease expression of genes and proteins related to the receptor activator of nuclear factor-κB ligand (RANKL) induced osteoclast differentiation from macrophages in the murine macrophage cell line, RAW 264.7An high dose (108 CFU) or a low dose (107 CFU) CFU of live or heat killed P. freudenreichii MJ2 were administered daily for 3 weeks to male DBA/1J mice of eight weeks. Then collagen-induced arthritis (CIA) was induced by injecting bovine type II collagen (CII) in the tail and the administration of P. freudenreichii MJ2 was continued until the end of the experiment. Live P. freudenreichii MJ2 treatment did not significantly affect the incidence of arthritis but hkMJ2 significantly decreased the arthritis score with reduced joint destruction. Bone surface/bone volume (BS/TV), BMD, trabecular thickness (Tb.Th), trabecular number (Tb.N) and bone volume/tissue volume (BV/TV) significantly increased in treated mice, while trabecular separation (Tb.sp) significantly decreased. The groups receiving P. freudenreichii MJ2 showed a lower degree of bone erosion, reduced leukocyte infiltration, and cartilage damage. The anti-inflammatory cytokine, IL-10 was significantly increased in the high dose P. freudenreichii MJ2 group, while the expression level of IL-17, which leads to the worsening of RA, was significantly decreased in all P. freudenreichii MJ2 groups. Inhibited osteoclast differentiation was attributed to a decreased expression of osteoclast differentiation-related genes and increased osteoprotegerin (OPG)/RANKL ratio, thus to the inhibition of the NF-κB signaling pathway. It was concluded that both live and dead P. freudenreichii MJ2 can prevent or improve RA in an animal model.
12. Streptococcus thermophilus
Streptococcus thermophilus is a SLAB species well adapted to fermented dairy products and is used as commercial or natural starter in yogurt and different cheese types, among which Grana type cheeses and Pasta Filata cheeses [15,16]. Illikoud et al. [4] summarized the effects of S. thermophilus strain treatment in mice with DSS or trinitrobenzene sulfonate (TNBS) induced colitis reporting that S. thermophilus strains prevented and decreased symptom intensity of DSS and TNBS induced colitis, including bodyweight loss, gastrointestinal bleeding and bacterial translocation into the colonic tissue and to the liver, macroscopic and histologic damage, immune stimulation and associated inflammation, levels of inducible nitric oxide synthase positive (iNOs+) cells in the large intestines. This effect was attributed to antioxidative activity, as inferred from the decreased content of lipid peroxides in the colon mucosa, decreased DSS induced pro-inflammatory IL-17 for a decrease in the percentage of Th17 lymphocytes in the lamina propria. For a yogurt isolate these effects were exerted also by the isolated exopolysaccharides that restored the expression of tight junction proteins claudin-1, occludin, and E-canderin.
S. thermophilus strains mitigated 5-FU induced mucositis in mice by reducing diarrhea, weight and appetite loss and restoring the intestinal architecture with maintenance of the epithelium structure in the small intestine and colon and reduced intestinal inflammation. It was suggested that the protective mechanisms exerted by S. thermophilus in intestinal inflammation may involve immunomodulation mediated by the production of folate, riboflavin and exopolysaccharides.
In clinical trials S. thermophilus oral intake improved serum collagen type II C-telopeptide and serum CRP in knee osteoarthritis and symptoms of atopic dermatitis [4].
12.1. Amelioration of metabolic syndrome
Heat inactivated S. thermophilus MN-ZLW-002, a strain isolated from a Chinese dairy product, significantly reduced weight gain after three weeks administration to male C57BL/6 HFD fed mice. Fasting blood glucose levels were significantly reduced after four and eight weeks and total TGs were significantly reduced after 12 weeks. Serum IL-1b and chemokine ligand 3 (CCL3) markers of pro-inflammatory responses decreased. Suppression of pre-adipocytes differentiation through immunity was one of the suggested mechanisms of action of heat inactivated S. thermophilus MN-ZLW-002. A possible underlying mechanisms was the regulation of the host immune system via cytokine release [78].
According to the WHO, cardiovascular diseases (CVDs) have been considered the main cause of death globally with approximately tens million affected people every year [79]. Hypercholesterolemia is a risk factor for such diseases based on a three times higher risk of heart attack in hypercholesterolemic individuals. Nonpharmacological cholesterol reduction methods are available in addition to available medications and they do not pose adverse effects. One such strategy is the use of probiotics to improve lipid metabolism [80]. It was even proposed that hypercholesterolemic people may consume probiotics and/or prebiotics as food supplements to an appropriate diet instead of cholesterol-lowering drugs [81]. S. thermophilus MH422542 isolated from an Egyptian dairy product, was selected among dairy isolates on the basis of auto-aggregation and co-aggregation, adhesion ability to human intestinal cells and to Caco-2 cells, cholesterol assimilation, β-galactosidase activity and bile salts hydrolysis. When administered to Albino rats in milk fermented by the strain for five weeks and in number of 108 CFU/mL, determined lower TG and total lipids concentrations in blood. As for other LAB tested in the same study, the group receiving S. thermophilus at 108 CFU/mL initial level exhibited a reduction of fecal Staphylococcus sp. and coliforms. The histological examination of liver did not reveal any tissue or cell shape alteration [5].
12.2. Anti-cancer effects
It was observed that S. thermophilus can reduce the risk of colorectal cancer (CRC) in mice via the β-galactosidase activity which releases galactose that perturbs cancer development signaling pathways. Therefore, Li et al. [82] examined the immunomodulatory effects of S. thermophilus ATCC 19258, isolated from pasteurized milk, on the intestinal mucosa azoxymethane-induced colon cancer in male C57BL/6J-ApcMin/J mice, which harbor a mutation in the tumor suppressor gene Apc. Mice received 2 × 107 CFU of S. thermophilus ATCC 19258 or E coli strain MG1655 (negative control) or a nonsteroidal anti-inflammatory drug that protects against CRC (positive control) by oral gavage, and they were raised until 20 weeks. The S. thermophilus-gavaged mice had a reduction in tumor number and size. The tumor-suppressive effect of S thermophilus was further validated in C57BL/6 mice injected with azoxymethane, followed by oral administration of S. thermophilus ATCC 19258. Notably, HTS analysis showed that S thermophilus administration caused the enrichment of the probiotic Bifidobacterium and Lactobacillus genuses, in particular the species B choerinum, B pseudolongum, B coryneforme, L. reuteri, L animalis, and L acidophilus. The observed activities were dependent on β-galactosidase activity, demonstrated by the fact that a β-galactosidase knockout S. thermophilus ATCC 19258 strain was ineffective, while administration of galactose decreased small-intestine tumor number in Apcmin/+ mice.
12.3. Infection mitigation
The yogurt isolate S. thermophilus KLDS 3.1003 was able to exert antimicrobial activities for the production of bacteriocin-coding peptides. It is also endowed with intact genes for acid tolerance, salt-resistance and antioxidant activities that favor its effectiveness against foodborne pathogens.
When administered orally to BALB/c mice for 14 days challenged with E.coli and S. aureus [83], it restored pathogen-disrupted blood parameters HDL, LDL, TP, TG, AST, ALT and some minerals.
Sepsis is caused by severe infections and can lead to life-threatening organ disfunction. A balanced gut microbiota has a protective role during systemic inflammation, therefore it was hypothesized that probiotic supplementation could be an aid in the treatment of sepsis. Seven-14-week-old male BALB/c (H-2Dd) mice were administered different doses of LPS, a well-known mediator of sepsis even causing septic shock in uncontrolled infection, by intraperitoneal injection and then received about 3 x 108 CFU of the yogurt isolate S. thermophilus 19 selected for its capacity to suppress the inflammatory response in vitro. All LPS-treated mice exhibited a reduction in food and water intake, but mice receiving S. thermophilus 19 showed no changes in body weight, possibly for a better recovery favored by alleviation of inflammation. The expression of TNF-α, IL-1β and IL-6 was lower in all organs in the S. thermophilus 19 group and all histopathologic scores were decreased based on structure conservation and degree of inflammatory cell infiltration for liver, lungs, small intestine and kidney. High throughput sequencing of the 16S rRNA gene of the cecum fecal microbiota revealed no effect on species richness of S. thermophilus 19 but different abundances of the genuses Fusobacterium and Flavonifractor, that could be involved in immunomodulation [84].
12.4. Alleviation of aging damage
Two strains of S. thermophilus isolated from fermented milk, T-1 and 510, were fed to four-day-old young adult C. elegans worms in alternative to E. coli OP50 normal diet. At day 11, S. thermophilus fed worms showed significantly increased survival rate and at day 14 lipofuscin lipid peroxidation product autofluorescence significantly decreased in the S. thermophilus groups. Moreover, locomotive capability indicator of muscle function was vigorous in the latter groups at day 18. As determined by qRT-PCR, feeding with S. thermophilus upregulated the expression of daf-16 encoding an orthologue of a transcription factor that regulates longevity under the conditions of oxidative stress and caloric restriction. In fact, also the expression of several antioxidant genes increased. The results of qRT-PCR indicated activation of the DAF-7/TGF-β pathway as mechanism underlying the enhanced expression of daf-16, suggesting possible analogous effects of these universal pathways in other hosts [85].
12.5. Defense from pollutants
A study investigated the effect of administration of S. thermophilus LM1012, isolated from a dairy product, on liver damage deriving from exposure to to diesel exhaust particulate matter (DEPM). S. thermophilus LM1012 was administered in 106 CFU/g to male 5-week old ICR mice daily for 14 days. Mice were then exposed to DEPM. Based on AST, ALT, and LDH levels in the blood, S. thermophilus LM1012 decreased liver injury induced by DEPM in mice [86].
13. Mixed cultures
13.1. Amelioration of intestinal health
A mixed culture (YC) of S. thermophilus MK-10 and L. bulgaricus 151 starters with synergistic effects during yogurt fermentation was administered for 8 days in numbers of 5 x 109 CFU to female specific-pathogen-free BALB/c mice after induction of colitis with DSS. The YC group showed attenuated severity of colitis, not decreased colon length and ameliorated histological signs. Myeloperoxidase (MPO) activity, an indicator of neutrophil infiltration into inflamed tissue and marker of acute inflammation, significantly decreased. YC lowered valeric acid and isovaleric acid and total putrefactive SCFA but did not show differences in the amounts of acetic, propionic, and lactic acids. Cytotoxic CD3+CD8+ T cells and CD3+CD4+CD25+FoxP3+ Tregs were downregulated in Peyer's patches and CD3+CD4+CD25+ T cells increased in spleen and CD3+CD4+CD25+FoxP3+ cells in peripheral blood mononuclear cells. IgA antibody-forming cells were downregulated in mesenteric lymph nodes and enhanced in spleen. The autors concluded that the strains possess the ability to modulate the intestinal mucosal and systemic immune system toward both IgA production and induction of Tregs cells by shifting Th1/Th2 balance [87].
13.2. Infection alleviation
L. delbrueckii subsp. bulgaricus KLDS 1.0207 and S. thermophilus KLDS 3.1003 from Mongolian indigenous dairy product and traditional yoghurt significantly suppressed the pathogenicity of E. coli ATCC 25922 and S. aureus ATCC 25923 female BALB/c mice. Standard hematological parameters of immunity showed that single cultures and co-cultures of the two strains could potentially improve immunity levels in humans [88]. When tested alone, L. delbrueckii subsp. bulgaricus KLDS 1.0207, at a 108 CFU daily dose, determined a lower weight loss for the groups receiving both pathogens, restored the serum AST, total protein and albumin levels in the E. coli group and brought AST, ALT and albumin to normal levels in the S. aureus group [89].
Pretreatment with commercial dairy protective cultures reduced mortality caused by L. monocytogenes Scott A in C. elegans. L. monocytogenes in L4/young adult nematodes infected for 24 h reduced the 50% (TD50) survival from about 10 days to about 7 days. Pretreatment by plate feeding of C. elegans with L. lactis DVS BS-10 resulted in higher times of 50% survival. TD50 values for L. plantarum Holdbac Listeria and L. plantarum Lyofast were similar to that of the negative control fed with E. coli OP50. Pretreatment of C. elegans with different protective cultures did not affect the colonization levels of L. monocytogenes, suggesting that the reduced mortality should be attributed to the enhancement of the immune response [90].
14. Enterococci as controversial probiotics
Enterococci are very commonly found as natural component of the cheese microbiota [stracciata] and a few Enterococcus species were considered for use as probiotics, namely, E. faecalis, E. faecium, E. lactis and more recently E. hirae and E. durans. Some Enterococcus strains are currently marketed to alleviate the symptoms of irritable bowel syndrome and recurrent chronic sinusitis or bronchitis. In particular, E. faecalis Symbioflor 1, isolated from the stool of a healthy human adult, has been used as a probiotic for more than 50 years without any reported adverse effect. It has also been shown to be non toxic in C. elegans. Based on several toxicological studies and on the genome sequence, this strain can be administred to humans. However, some enterococci can behave as opportunistic pathogens and are a prevalent cause of nosocomial infections such as bacteraemia, endocarditis, urinary tract infections and other infections [91]. Virulence factors that can be found in E. faecalis aggregation substance (asa1), gelatinase (gelE), collagen adhesin (ace), enterococcal surface protein (esp) and cytolysin (cylA) are sporadically detected in dairy isolates. For the possible presence of these virulence genes, Enterococcus species are not included in the QPS list [EFSA], so that in potential probiotic strains the absence of transferable AR genes and virulence determinants must be ascertained before use in pharmaceutical or food industry. On the other hand, horizontal transfer of AR genes among different strains was demonstrated in vivo in volunteers, so that AR genes represents a risk for the spread of resistance genes to indigenous enterococci, which can be responsible for infections in immunocompromised patients [92].
Among isolates from Tunisian dairy products one of the most promising strains in in vitro preliminary testing, E. faecalis OB14, led to high mortality of Galleria Mellonella larvae in the virulence test. Another strain, E. faecalis OB15, devoid of cytolysin genes, was not toxic for the larvae. MALDI TOF-MS and WGS analyses showed that E. faecalis OB15 was closely related to the E. faecalis Symbioflor 1[91]. This indicates the opportunity to test the probiotic potentialities of individual strains in in vivo models.
In a study aimed to distinguish friend from foe enterococci it was found that of 30 isolates from different dairy products carried on average four AR genes and all carried at least one AR gene. All isolates were classified as potential human pathogens by PathogenFinder analysis. However, E. faecium isolates, that were the most prevalent, showed the absence of adhesins sgrA, ecbA and scm virulence factors present in human pathogenic strains [93].
14.1. Immunomodulation
Darwish et al. [5] found that in Albino rats fed with fermented milk prepared with E. faecium MH422543, preliminarily evaluated for absence of virulence factors, 108 CFU/mL of the strain administered for five weeks determined increased G3 lymphocytes in blood, indicating an immune response. The E. faecium group had higher concentrations of TG and total lipids than other treatments compared but a remarkable suppression of LDL and 88% reduction of atherogenic indices.
14.2. Infection mitigation
Despite carrying some virulence factors and antibiotic resistance genes, E. faecalis UGRA10 isolated from a Spanish sheep's cheese was administered with a final concentration in water of 109 CFU/ml for 30 days to rainbow trouts before challenging by intraperitoneal injection of 5 × 108 CFU/fish of the fish pathogen L. garvieae CECT 5274. The cumulative survival rate of treated trouts was 50% compared with 0% for control fishes. E. faecalis UGRA10 produces enterocin AS-48, a cationic circular bacteriocin with broad bactericidal activity against most Gram-positive and some Gram-negative bacteria. Trouts inoculated with the pathogen and treated by regular dipping in AS-48 baths had a survival rate of 60% after 20 days compared with that of untreated fish (0%). Therefore, the isolated bacteriocin, which was shown not to be toxic in the trouts, can substitute use of E. faecalis UGRA10 as alternative to antibiotics to control L. garvieae infections in aquaculture [94].
The number of reports summarized in this review separated by topic and microrganisms involved are shown in Figure 1.
15. Conclusions
This review on the in vivo health promoting effects of microorganisms of dairy origin highlights that multiple advantages could derive from the evaluation of the characteristics and selection of strains with enhanced capacity to ameliorate different disease conditions. Though fermented dairy products supply high numbers of bacterial species comprising strains with probiotic properties, more studies on the in vivo functions of the microorganisms present could make available probiotics well adapted to dairy matrices that could be enriched in dairy foods to increase their functional value.
Author Contributions
G.A. and F.R. Conceptualization, methodology, data curation, writing—original draft preparation; I.D.M., P.T., L.M. and S.S. investigation, writing—review and editing.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Marco, M.L.; Sanders, M.E.; Gänzle, M.; Arrieta, M.C.; Cotter, P.D.; De Vuyst, L.; Hill, C.; Holzapfel, W.; Lebeer, S.; Merenstein, D.; Reid, G.; Wolfe, B.E.; Hutkins, R. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on fermented foods. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Food and Agricultural Organization. Probiotics in food. Health and nutritional properties and guidelines for evaluation. Available online: https://www.fao.org/3/a0512e/a0512e.pdf (accessed on 24 April 2023).
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli. L.; Canani, R.B.; Flint, H.J.; Salminen, S.; Calder, P.C.; Sanders, M.E. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol. 2014, 8, 506–514.
- Illikoud, N.; Mantel, M.; Rolli-Derkinderen, M.; Gagnaire, V.; Jan, G. Dairy starters and fermented dairy products modulate gut mucosal immunity. Immunol. Lett. 2022, 251–252, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Darwish, A.M.G.; Allam, M.G.; Shokery, E.S.; Ayad, E.H.E. Functional products fortified with probiotic LAB isolated from Egyptian dairy sources showed hypolipidemic effects in Albino rats. PLoS One 2022, 17, e0263241. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. EFSA statement on the requirements for whole genome sequence analysis of microorganisms intentionally used in the food chain. EFSA J. 2021, 19, 6506. [Google Scholar]
- Amadoro, C.; Rossi, F.; Pallotta, M.L.; Gasperi, M.; Colavita, G. Traditional dairy products can supply beneficial microorganisms able to survive in the gastrointestinal tract. LWT 2018, 93, 376–383. [Google Scholar] [CrossRef]
- Milani, C.; Duranti, S.; Napoli, S.; Alessandri, G.; Mancabelli, L.; Anzalone, R.; Longhi, G.; Viappiani, A.; Mangifesta, M.; Lugli, G.A.; Bernasconi, S.; Ossiprandi, M.C.; van Sinderen, D.; Ventura, M.; Turroni, F. Colonization of the human gut by bovine bacteria present in Parmesan cheese. Nat Commun. 2019, 10, 1286. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Honme, Y.; Sashihara, T. Lactobacillus delbrueckii subsp. bulgaricus 2038 and Streptococcus thermophilus 1131 Induce the Expression of the REG3 Family in the Small Intestine of Mice via the Stimulation of Dendritic Cells and Type 3 Innate Lymphoid Cells. Nutrients 2019, 11, 2998. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, K.; Kim, W. Cream Cheese-Derived Lactococcus chungangensis CAU 28 Modulates the Gut Microbiota and Alleviates Atopic Dermatitis in BALB/c Mice. Sci Rep. 2019, 9, 446, Erratum in: Sci Rep. 2019, 9, 7001. [Google Scholar] [CrossRef]
- Zhao, L.; Shi, F.; Xie, Q.; Zhang, Y.; Evivie, S.E.; Li, X.; Liang, S.; Chen, Q.; Xin, B.; Li, B.; Huo, G. Co-fermented cow milk protein by Lactobacillus helveticus KLDS 1.8701 and Lactobacillus plantarum KLDS 1.0386 attenuates its allergic immune response in Balb/c mice. J Dairy Sci 2022, 105, 7190–7202. [Google Scholar] [CrossRef]
- Makwana, S.; Prajapati, J.B.; Pipaliya, R.; Hati, S. Effects of probiotic fermented milk on management of obesity studied in high-fat-diet induced obese rat model. Food Prod Process and Nutr 2023, 5, 3. [Google Scholar] [CrossRef]
- Glazunova, O.A.; Moiseenko, K.V.; Savinova, O.S.; Fedorova, T.V. In Vitro and In Vivo Antihypertensive Effect of Milk Fermented with Different Strains of Common Starter Lactic Acid Bacteria. Nutrients 2022, 14, 5357. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Amadoro, C.; Pallotta, M.L.; Colavita, G. Variability of Genetic Characters Associated with Probiotic Functions in Lacticaseibacillus Species. Microorganisms 2022, 10, 1023. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Gatto, V.; Sabattini, G.; Torriani, S. An assessment of factors characterising the microbiology of Grana Trentino cheese, a Grana-type cheese. Int. J. Dairy Techn. 2012, 65, 401–409. [Google Scholar] [CrossRef]
- Del Matto, I.; Rossi, F.; Iannitto, G.; Petrone, D.; Mastrodomenico, M.T.; Alessiani, A.; Sacchini, L.; Amadoro, C.; Tucci, P.; Marino, L. Variability of the microbiota in traditional Caciocavallo, Scamorza and Caciotta cheeses manufactured with raw milk and natural cultures. Int J Dairy Technol. 2021, 74, 564–574. [Google Scholar] [CrossRef]
- Aprea, G.; Alessiani, A.; Rossi, F.; Sacchini, L.; Boni, A.; D’Angelantonio, D.; Scattolini, S.; Sperandii, A.F.; Centorotola, G.; Neri, D.; Pomilio, F.; Di Giannatale, E.; Del Matto, I.; Tucci, P.; Migliorati, G. Characterization of Lactic Acid Bacteria in Pecorino di Farindola Cheese and Manufacturing with a Lacticaseibacillus paracasei Autochthonous Culture. Appl. Sci. 2021, 11, 7897. [Google Scholar] [CrossRef]
- Saxami, G.; Ypsilantis, P.; Sidira, M.; Simopoulos, C.; Kourkoutas, Y.; Galanis, A. Distinct adhesion of probiotic strain Lactobacillus casei ATCC 393 to rat intestinal mucosa. Anaerobe 2012, 18, 417–420. [Google Scholar] [CrossRef]
- Bengoa, A.A.; Errea, A.J.; Rumbo, M.; Abraham, A.G.; Garrote, G.L. Modulatory properties of Lactobacillus paracasei fermented milks on gastric inflammatory conditions. Int. Dairy J. 2020, 111, 104839. [Google Scholar] [CrossRef]
- Aindelis, G.; Tiptiri-Kourpeti, A.; Lampri, E.; Spyridopoulou, K.; Lamprianidou, E.; Kotsianidis, I.; Ypsilantis, P.; Pappa, A.; Chlichlia, K. Immune Responses Raised in an Experimental Colon Carcinoma Model Following Oral Administration of Lactobacillus casei. Cancers (Basel) 2020, 12, 368. [Google Scholar] [CrossRef]
- Spyridopoulou, K.; Tryfonopoulou, E.; Aindelis, G.; Ypsilantis, P.; Sarafidis, C.; Kalogirou, O.; Chlichlia, K. Biogenic selenium nanoparticles produced by Lactobacillus casei ATCC 393 inhibit colon cancer cell growth in vitro and in vivo. Nanoscale Adv. 2021, 3, 2516–2528. [Google Scholar] [CrossRef]
- Le Barz, M.; Daniel, N.; Varin, TV.; Naimi, S.; Demers-Mathieu, V.; Pilon, G.; Audy, J.; Laurin, É.; Roy, D.; Urdaci, M.C.; St-Gelais, D.; Fliss, I.; Marette, A. In vivo screening of multiple bacterial strains identifies Lactobacillus rhamnosus Lb102 and Bifidobacterium animalis ssp. lactis Bf141 as probiotics that improve metabolic disorders in a mouse model of obesity. FASEB J. 2019, 33, 4921–4935. [Google Scholar]
- Chondrou, P.; Karapetsas, A.; Kiousi, D.E.; Vasileiadis, S.; Ypsilantis, P.; Botaitis, S.; Alexopoulos, A.; Plessas, S.; Bezirtzoglou, E.; Galanis, A. Assessment of the Immunomodulatory Properties of the Probiotic Strain Lactobacillus paracasei K5 In Vitro and In Vivo. Microorganisms 2020, 8, 709. [Google Scholar] [CrossRef] [PubMed]
- Karaffová, V.; Mudroňová, D.; Mad’Ar, M.; Hrčková, G.; Faixová, D.; Gancarčíková, S.; Ševčíková, Z.; Nemcová, R. Differences in Immune Response and Biochemical Parameters of Mice Fed by Kefir Milk and Lacticaseibacillus paracasei Isolated from the Kefir Grains. Microorganisms 2021, 9, 831. [Google Scholar] [CrossRef] [PubMed]
- Valente, G.L.C, Acurcio, L.B, Freitas, L.P.V, Nicoli, J.R, Silva, A.M, Souza, M.R, Penna, C.F.A.M. Short communication: In vitro and in vivo probiotic potential of Lactobacillus plantarum B7 and Lactobacillus rhamnosus D1 isolated from Minas artisanal cheese. J Dairy Sci. 2019, 102, 5957–5961.
- Wang, G.; Zhang, Y.; Song, X.; Xia, Y.; Lai, P.F.; Ai, L. Lactobacillus casei LC2W can inhibit the colonization of Escherichia coli O157:H7 in vivo and reduce the severity of colitis. Food Funct. 2019, 10, 5843–5852. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, D.; Sun, Z.; Wu, R.; WChenu, X.; Chen, W.; Meng, H.; Hu, S.; Zhang, H. Complete genome sequence of Lactobacillus casei Zhang, a new probiotic strain isolated from traditional homemade koumiss in Inner Mongolia, China. J Bacteriol. 2010, 192, 5268–5269. [Google Scholar] [CrossRef] [PubMed]
- Kwok, L.Y.; Wang, L.; Zhang, J.; Guo, Z.; Zhang, H. A pilot study on the effect of Lactobacillus casei Zhang on intestinal microbiota parameters in Chinese subjects of different age. Benef. Microbes. 2014, 5, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Liu, G.; Wang, W.; Wang, Y.; Cao, Z.; Yang, H.; Li, S. Lactobacillus casei Zhang Counteracts Blood-Milk Barrier Disruption and Moderates the Inflammatory Response in Escherichia coli-Induced Mastitis. Front Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Fu, L.; Xie, M.; Wang, C.; Qian, Y.; Huang, J.; Sun, Z.; Zhang, H.; Wang, Y. Lactobacillus Casei Zhang Alleviates Shrimp Tropomyosin-Induced Food Allergy by Switching Antibody Isotypes through the NF-κB-Dependent Immune Tolerance. Mol Nutr Food Res 2020, 64, e1900496. [Google Scholar] [CrossRef]
- Zago, M.; Fornasari, M.E.; Carminati, D.; Burns, P.; Suàrez, V.; Vinderola, G.; Reinheimer, J.; Giraffa, G. Characterization and probiotic potential of Lactobacillus plantarum strains isolated from cheeses. Food Microbiol. 2011, 28, 1033–1040. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Zhang, L.; Zhao, Y.; Niu, C.; Yang, Z.; Li, S. Potential probiotic characterization of Lactobacillus plantarum strains isolated from Inner Mongolia "Hurood" cheese. J Microbiol Biotechnol. 2014, 24, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Ismael, M.; Gu, Y.; Cui, Y.; Wang, T.; Yue, F.; Qin, Y.; Lü, X. Probiotic of Lactiplantibacillus plantarum NWAFU-BIO-BS29 Isolated from Chinese Traditional Fermented Milk and Its Potential Therapeutic Applications Based on Gut Microbiota Regulation. Foods 2022, 11, 3766. [Google Scholar] [CrossRef] [PubMed]
- Nami, Y.; Bakhshayesh, R.V.; Manafi, M.; Hejazi, M.A. Hypocholesterolaemic activity of a novel autochthonous potential probiotic Lactobacillus plantarum YS5 isolated from yogurt. LWT, 2019, 111, 876–882. [Google Scholar] [CrossRef]
- Zambou, N.F.; Barry, R.B.; Bindzi, J.M.; Kenfack, C.H.M.; Bemmo, U.L.K. Viability and in vivo Hypocholesterolemic Effect of Lactobacillus plantarum 29V in Local Honey. J Adv Biol Biotechnol. 2021, 24, 24–33. [Google Scholar]
- Chouikhi, A.; Ktari, N.; Bardaa, S.; Hzami, A.; Ben Slima, S.; Trabelsi, I.; Asehraou, A.; Ben Salah, R. A novel probiotic strain, Lactiplantibacillus plantarum LC38, isolated from Tunisian camel milk promoting wound healing in Wistar diabetic rats. Arch Microbiol. 2021, 204, 24. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, S.; Cui, J.; Guo, T.; Zhang, J. Effect of bile salt hydrolase-active Lactobacillus plantarum Y15 on high cholesterol diet induced hypercholesterolemic mice. CyTA - Journal of Food 2021, 19, 408–417. [Google Scholar] [CrossRef]
- Hu, J.; Wang, C.; Ye, L.; Yang, W.; Huang, H.; Meng, F.; Shi, S.; Ding, Z. Anti-tumour immune effect of oral administration of Lactobacillus plantarum to CT26 tumour-bearing mice. J. Biosci. 2015, 40, 269–279. [Google Scholar] [CrossRef]
- Fayol-Messaoudi, D.; Coconnier-Polter, M.H.; Moal, V.L. .; Atassi, F..; Berger, C.N.; Servin, A.L. The Lactobacillus plantarum strain ACA-DC287 isolated from a Greek cheese demonstrates antagonistic activity in vitro and in vivo against Salmonella enterica serovar Typhimurium. J Appl Microbiol. 2007, 103, 657–665. [Google Scholar] [CrossRef]
- Acurcio, L.B.; Bastos, R.W.; de Cicco Sandes, S.H.; de Carvalho Guimarães, A.C..; Alves, C.G.; dos Reis, D.C.; Wuyts, S.; Cantini Nunes, Á.; Cassali, G.D.; Lebeer, S.; Resende de Souza, M.; Nicoli, J.R. Protective effects of milk fermented by Lactobacillus plantarum B7 from Brazilian artisanal cheese on a Salmonella enterica serovar Typhimurium infection in BALB/c mice. J. Funct. Foods 2017, 33, 436–445.
- Chong, H.X.; Yusoff, N.A.A.; Hor, Y.Y.; Lew, L.C.; Jaafar, M.H.; Choi, S.B.; Yusoff, M.S.B.; Wahid, N.; Abdullah, M.F.I.L.; Zakaria, N.; Ong, K.L.; Park, Y.H.; Liong, M.T. Lactobacillus plantarum DR7 improved upper respiratory tract infections via enhancing immune and inflammatory parameters: A randomized, double-blind, placebo-controlled study. J Dairy Sci. 2019, 102, 4783–4797. [Google Scholar] [CrossRef]
- Zhao, X.; Yi, R.; Zhou, X.; Mu, J.; Long, X.; Pan, Y.; Song, J.L.; Park, K.Y. Preventive effect of Lactobacillus plantarum KSFY02 isolated from naturally fermented yogurt from Xinjiang, China, on D-galactose-induced oxidative aging in mice. J Dairy Sci. 2019, 102, 5899–5912. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, X.; Jiang, Y.;, Zhao, W.; Guo, T.; Cao ,Y.; Teng, J.; Hao, X.; Zhao, J.; Yang, Z. Antioxidant status and gut microbiota change in an aging mouse model as influenced by exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibetan kefir. J Dairy Sci. 2017, 100, 6025–6041. [CrossRef] [PubMed]
- Liu, G.; Chong, H.X.; Chung, F.Y.; Li, Y.; Liong, M.T. Lactobacillus plantarum DR7 Modulated Bowel Movement and Gut Microbiota Associated with Dopamine and Serotonin Pathways in Stressed Adults. Int J Mol Sci. 2020, 21, 4608. [Google Scholar] [CrossRef] [PubMed]
- Chong, H.X.; Yusoff, N.A.A.; Hor, Y.Y.; Lew, L.C.; Jaafar, M.H.; Choi, S.B.; Yusoff, M.S.B.; Wahid, N.; Abdullah, M.F.I.L.; Zakaria, N.; Ong, K.L.; Park, Y.H.; Liong, M.T. Lactobacillus plantarum DR7 alleviates stress and anxiety in adults: a randomised, double-blind, placebo-controlled study. Benef Microbes 2019, 10, 355–373. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Liu, M.; Yang, S.; Shen, C.; Wang, X.; Liu, W.; Guo, Y. Fermented milk of cheese-derived Lactobacillus delbrueckii subsp. bulgaricus displays potentials in alleviating alcohol-induced hepatic injury and gut dysbiosis in mice. Food Res Int. 2022, 157. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, S.; Cui, J.; Guo, T.; Zhang, J.; Li, B. Alleviative Effects of Exopolysaccharide Produced by Lactobacillus helveticus KLDS1.8701 on Dextran Sulfate Sodium-Induced Colitis in Mice. Microorganisms 2021, 9, 2086. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.W.; El-Nezami, H.; Corke, H.; Ho, C.S.; Shah, N.P. L-citrulline enriched fermented milk with Lactobacillus helveticus attenuates dextran sulfate sodium (DSS) induced colitis in mice. J Nutr Biochem. 2022, 99. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, S.; Chen, L.; Chen, C.; Ren, X.; Zheng, Z.; Weng, L.; Ge, H.; Wang, J.; Liu, G.; Ye, X. Immunomodulatory effects of L. helveticus WHH2580 fermented milk on an immunosuppressed murine model. J Funct Foods, 2022, 99, 105353. [Google Scholar] [CrossRef]
- Makino, T.; Yamashita, M.; Takeuchi, N.; Kabuki, T.; Hattori, M.; Yoshida, T. Lactobacillus helveticus SBT2171 alleviates allergic symptoms in a murine model for pollen allergy. Biosci Biotechnol Biochem. 2019, 83, 2298–2306. [Google Scholar] [CrossRef]
- Yamashita, M.; Kobatake, E.; Obuchi, S.; Iwai, M.; Ichikawa, K.; Kabuki, T.; Enomoto, T. Intake safety of Lactobacillus helveticus SBT2171 and its effects on nasal and ocular symptoms associated with mites and house dust: An open-label study. Functional Foods in Health and Disease 2019, 9, 52–78. [Google Scholar] [CrossRef]
- Yamashita, M.; Miyoshi, M.; Iwai, M.; Takeda, R.; Ono, T.; Kabuki, T. Lactobacillus helveticus SBT2171 Alleviates Perennial Allergic Rhinitis in Japanese Adults by Suppressing Eosinophils: A Randomized, Double-Blind, Placebo-Controlled Study. Nutrients 2020, 12, 3620. [Google Scholar] [CrossRef] [PubMed]
- Kobatake, E.; Kobayashi, R.; Kabuki, T.; Kurita-Ochiai, T. Lactobacillus helveticus SBT2171 upregulates the expression of β-defensin and ameliorates periodontal disease caused by Porphyromonas gingivalis. Microbiol Immunol. 2019, 63, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Shi, R.; Guan, D.; Wu, Y.; Qian, W. Lactobacillus helveticus Prevents Periodontitis Induced by Aggregatibacter actinomycetemcomitans in Rats by Regulating β-Defensins. Comput Math Methods Med 2022, 2022, 4968016. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, M.; Naderi Farsani, M.; Ghafarifarsani, H.; Raeeszadeh, M. Dietary Lactobacillus helveticus and Gum Arabic improves growth indices, digestive enzyme activities, intestinal microbiota, innate immunological parameters, antioxidant capacity, and disease resistance in common carp. Fish Shellfish Immunol. 2023, 135, 108652. [Google Scholar] [CrossRef]
- Li, B.; Evivie, S.E.; Lu, J.; Jiao, Y.; Wang, C.; Li, Z , Liu, F.; Huo, G. Lactobacillus helveticus KLDS1.8701 alleviates d-galactose-induced aging by regulating Nrf-2 and gut microbiota in mice. Food Funct. 2018, 9, 6586–6598. [CrossRef]
- Li, B.; Du, P.; Smith, E.E.; Wang, S.; Jiao, Y.; Guo, L.; Huo, G.; Liu, F. In vitro and in vivo evaluation of an exopolysaccharide produced by Lactobacillus helveticus KLDS1.8701 for the alleviative effect on oxidative stress. Food Funct. 2019, 10, 1707–1717. [Google Scholar] [CrossRef]
- Gao K, Chen CL, Ke XQ, Yu YX, Chen S, Liu GC, Wang HF, Li YJ. Ingestion of Lactobacillus helveticus WHH1889 improves depressive and anxiety symptoms induced by chronic unpredictable mild stress in mice. Benef Microbes. 2022, 13, 473–488. [CrossRef]
- Yang, G.; Cui, X.; Liu, S.; Lu, J.; Hou, X.; Meng, W.; Wu, B.; Su, Y.; Zhang, H.; Zheng, W.; Fang, Y. Effects of dietary Lactobacillus helveticus on the growth rate, disease resistance and intestinal health of pond loach (Misgurnus anguillicaudatus). Aquaculture 2021, 544, 737038. [Google Scholar] [CrossRef]
- Saleena, L.A.K.; Teo, M.Y.M.; How, Y.H.; In, L.L.A.; Pui, L.P. Immunomodulatory action of Lactococcus lactis. Journal of Bioscience and Bioengineering, 2022, 135, 1–9. [Google Scholar] [CrossRef]
- Barcellos Jaskulski, I.; Uecker, J.; Bordini, F.; Moura, F.; Gonçalves, T.; Garcia Chaves, N.; Camargo, F.; Borelli Grecco, F.; Fiorentini, Â.M.; Padilha da Silva, W.; Andreazza, R.; Pieniz, S. In vivo action of Lactococcus lactis subsp. lactis isolate (R7) with probiotic potential in the stabilization of cancer cells in the colorectal epithelium. Process Biochemistry, 2020, 91, 165–171. [Google Scholar] [CrossRef]
- Jin, S.W.; Lee, G.H.; Jang, M.J.; Hong, G.E.; Kim, J.Y.; Park, G.D.; Jin, H.; Kim, H.S.; Choi, J.H.; Choi, C.Y.; Lee, S.G.; Jeong, H.G.; Hwang, Y.P. Immunomodulatory Activity of Lactococcus lactis GCWB1176 in Cyclophosphamide-Induced Immunosuppression Model. Microorganisms 2020, 8, 1175. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Farzi, A.; Ke, X.; Yu, Y.; Chen, C.; Chen, S.; Yu, T.; Wang, H.; Li, Y. Oral administration of Lactococcus lactis WHH2078 alleviates depressive and anxiety symptoms in mice with induced chronic stress. Food Funct. 2022, 13, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Ramalho, J.B.; Soares, M.B.; Spiazzi, C.C.; Bicca, D.F.; Soares, V.M.; Pereira, J.G.; da Silva, W.P.; Sehn, C.P.; Cibin, F.W.S. In Vitro Probiotic and Antioxidant Potential of Lactococcus lactis subsp. cremoris LL95 and Its Effect in Mice Behaviour. Nutrients 2019, 11, 901. [Google Scholar] [CrossRef] [PubMed]
- Toukam, L.L.; Tatsinkou Fossi, B.; Taiwe, G.S.; Bila, R.B.; Feugaing Sofeu.; D.D, Ivo.; E.P, Achidi.; E.A. In vivo antimalarial activity of a probiotic bacterium Lactobacillus sakei isolated from traditionally fermented milk in BALB/c mice infected with Plasmodium berghei ANKA. J Ethnopharmacol 2021, 280, 114448.
- Schifano, E.; Zinno, P.; Guantario, B.; Roselli, M.; Marcoccia, S.; Devirgiliis, C.; Uccelletti, D. The Foodborne Strain Lactobacillus fermentum MBC2 Triggers pept-1-Dependent Pro-Longevity Effects in Caenorhabditis elegans. Microorganisms 2019, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Hor, Y.Y.; Lew, L.C.; Jaafar, M.H.; Lau, A.S.; Ong, J.S.; Kato, T.; Nakanishi, Y.; Azzam, G.; Azlan, A.; Ohno, H.; Liong, M.T. Lactobacillus sp. improved microbiota and metabolite profiles of aging rats. Pharmacol Res. 2019, 146. [Google Scholar] [CrossRef]
- Forooghi Nia, F.; Rahmati, A.; Ariamanesh, M.; Saeidi, J.; Ghasemi, A.; Mohtashami, M. The Anti-Helicobacter pylori effects of Limosilactobacillus reuteri strain 2892 isolated from Camel milk in C57BL/6 mice. World J Microbiol Biotechnol. 2023, 39, 119. [Google Scholar] [CrossRef] [PubMed]
- Al-Emran, H.M.; Moon, J.F.; Miah, M.L.; Meghla, N.S.; Reuben, R.C.; Uddin, M.J.; Ibnat, H.; Sarkar, S.L.; Roy, P.C.; Rahman, M.S.; Alam, A.S.M.R.U.; Islam, O.K.; Jahid, I.K. Genomic analysis and in vivo efficacy of Pediococcus acidilactici as a potential probiotic to prevent hyperglycemia, hypercholesterolemia and gastrointestinal infections. Sci Rep. 2022, 12, 20429. [Google Scholar] [CrossRef]
- Feng, P.; Yang, J.; Zhao, S.; Ling, Z.; Han,.;Wu, Y.; Salama, E-S.; Kakade, A.; Khan, A.; Jin, W.; Zhang, W.; Jeon, B-H.; Fan, J.; Liu, M.; Mamtimin, T.; Liu, P Li, X. Human supplementation with Pediococcus acidilactici GR-1 decreases heavy metals levels through modifying the gut microbiota and metabolome. npj Biofilms and Microbiomes 2022, 8, 1.
- Rossi, F.; Torriani, S.; Dellaglio, F. Identification and clustering of dairy propionibacteria by RAPD-PCR and CGE-REA methods. J Appl Microbiol. 1998, 85, 956–964. [Google Scholar] [CrossRef]
- An, M.; Park, Y.H.; Lim, Y.H. Antiobesity and antidiabetic effects of the dairy bacterium P. freudenreichii MJ2 in high-fat diet-induced obese mice by modulating lipid metabolism. Sci Rep. 2021, 11, 2481. [Google Scholar] [CrossRef] [PubMed]
- Kwon, G.; Lee, J.; Lim, Y.H. Dairy Propionibacterium extends the mean lifespan of Caenorhabditis elegans via activation of the innate immune system. Sci Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Nair, D.V.T.; Vazhakkattu Thomas, J.; Dewi, G.; Noll, S.; Brannon, J.; Kollanoor Johny, A. Reduction of Multidrug-Resistant Salmonella enterica Serovar Heidelberg Using a Dairy-Originated Probiotic Bacterium, Propionibacterium freudenreichii freudenreichii B3523, in Growing Turkeys. Journal of Applied Poultry Research, 2019, 28, 356–363. [Google Scholar] [CrossRef]
- Nair, D.V.T.; Johnson, T.J.; Noll, S.L.; Kollanoor Johny, A. Effect of supplementation of a dairy-originated probiotic bacterium, Propionibacterium freudenreichii subsp. freudenreichii, on the cecal microbiome of turkeys challenged with multidrug-resistant Salmonella Heidelberg. Poult Sci. 2021, 100, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Yeom, J.; Ma, S.; Lim, Y.-H. Probiotic P. freudenreichii MJ2 Enhances Osteoblast Differentiation and Mineralization by Increasing the OPG/RANKL Ratio. Microorganisms 2021, 9, 673. [Google Scholar] [CrossRef] [PubMed]
- Yeom, J.; Yim, D.J.; Ma, S.; Lim, Y.-H. Propionibacterium freudenreichii Inhibits RANKL-Induced Osteoclast Differentiation and Ameliorates Rheumatoid Arthritis in Collagen-Induced Arthritis Mice. Microorganisms 2022, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Liang, H.; Luo, Y.; Li, Z.; He, F.; Han, X.; Zhang, L. Anti-adipogenesis and metabolism-regulating effects of heat-inactivated Streptococcus thermophilus MN-ZLW-002. Lett Appl Microbiol, 2021, 72, 677–687. [Google Scholar] [CrossRef]
- WHO. Cardiovascular Diseases. 2020. Available online: https://www.who.int/health-topics/cardiovascular-diseases#tab=tab_1 (accessed on 30 April 2023).
- Ishimwe, N.; Daliri, E.B.; Lee, B.H.; Fang, F.; Du, G. The perspective on cholesterol-lowering mechanisms of probiotics. Mol. Nutr. Food Res. 2015, 59, 94–105. [Google Scholar] [CrossRef]
- WHO. Diet, nutrition and the prevention of chronic diseases. World Health Organ. Tech. Rep. Ser. 2003, 916, 1–149. [Google Scholar]
- Li, Q.; Hu, W.; Liu, W.X.; Zhao, L.Y.; Huang, D.; Liu, X.D.; Chan, H.; Zhang, Y.; Zeng, J.D.; Coker, O.O.; Kang, W.; Ng, S.S.M.; Zhang. L.; Wong, S.H.; Gin, T.; Chan, M.T.V.; Wu, J.L.; Yu, J.; Wu, W.K.K. Streptococcus thermophilus Inhibits Colorectal Tumorigenesis Through Secreting β-Galactosidase. Gastroenterology. 2021, 160, 1179–1193.e14.
- Evivie, S.E.; Ogwu, M.C.; Abdelazez, A.; Bian, X.; Liu, F.; Li, B.; Huo, G. Suppressive effects of Streptococcus thermophilus KLDS 3.1003 on some foodborne pathogens revealed through in vitro, in vivo and genomic insights. Food Funct. 2021, 11, 6573–6587. [Google Scholar] [CrossRef]
- Han, F.; Wu, G.; Zhang, Y.; Zheng, H.; Han, S.; Li, X.; Cai, W.; Liu, J.; Zhang, W.; Zhang, X.; Hu, D. Streptococcus thermophilus Attenuates Inflammation in Septic Mice Mediated by Gut Microbiota. Front Microbiol 2020, 11, 598010. [Google Scholar] [CrossRef] [PubMed]
- Desaka, N.; Ota, C.; Nishikawa, H.; Yasuda, K.; Ishii, N.; Bito, T.; Kishinaga, Y.; Naito, Y.; Higashimura, Y. Streptococcus thermophilus extends lifespan through activation of DAF-16-mediated antioxidant pathway in Caenorhabditis elegans. J Clin Biochem Nutr. 2022, 70, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Jeon, M.G.; Kim, T.R.; Lee, J.Y.; Kim, H.S.; Ji, Y.; Holzapfel, W.H.; Bae, D.; Choi, C.Y.; Hwang, Y.P. Hepatoprotective Effects of Streptococcus thermophilus LM1012 in Mice Exposed to Air Pollutants. J Med Food. 2020, 23, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Wasilewska, E.; Zlotkowska, D.; Wroblewska, B. Yogurt starter cultures of Streptococcus thermophilus and Lactobacillus bulgaricus ameliorate symptoms and modulate the immune response in a mouse model of dextran sulfate sodium-induced colitis. J Dairy Sci. 2019, 102, 37–53. [Google Scholar] [CrossRef]
- Evivie, S.E.; Abdelazez, A.; Li, B.; Bian, X.; Li, W.; Du, J.; Huo, G.; Liu, F. In vitro Organic Acid Production and In Vivo Food Pathogen Suppression by Probiotic S. thermophilus and L. bulgaricus. Front Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Evivie, S.E.; Abdelazez, A.; Li, B.; Lu, S.; Liu, F.; Huo, G. Lactobacillus delbrueckii subsp. bulgaricus KLDS 1.0207 Exerts Antimicrobial and Cytotoxic Effects in vitro and Improves Blood Biochemical Parameters in vivo Against Notable Foodborne Pathogens. Front Microbiol. 2020, 11. [Google Scholar]
- Aljasir, S.F.; D'Amico, D.J. Probiotic potential of commercial dairy-associated protective cultures: In vitro and in vivo protection against Listeria monocytogenes infection. Food Res Int. 2021, 149. [Google Scholar] [CrossRef]
- Baccouri, O.; Boukerb, A.M.; Farhat, L.B.; Zébré, A.; Zimmermann, K.; Domann, E.; Cambronel, M.; Barreau, M.; Maillot, O.; Rincé, I.; Muller, C.; Marzouki, M.N.; Feuilloley, M.; Abidi, F.; Connil, N. Probiotic Potential and Safety Evaluation of Enterococcus faecalis OB14 and OB15, Isolated From Traditional Tunisian Testouri Cheese and Rigouta, Using Physiological and Genomic Analysis. Front Microbiol 2019, 10, 881. [Google Scholar] [CrossRef]
- Lester, C.H.; Frimodt-Møller, N.; Sørensen, T.L.; Monnet, D.L.; Hammerum, A.M. In vivo transfer of the vanA resistance gene from an Enterococcus faecium isolate of animal origin to an E. faecium isolate of human origin in the intestines of human volunteers. Antimicrob Agents Chemother. 2006, 50, 596–9. [Google Scholar] [CrossRef]
- Apostolakos, I.; Tsigkrimani, M.; Paramithiotis, S.; Mataragas, M. Whole-Genome Sequencing and Comparative Genomic Analysis of Enterococcus spp. Isolated from Dairy Products: Genomic Diversity, Functional Characteristics, and Pathogenic Potential. Appl. Sci. 2022, 12, 9620. [Google Scholar] [CrossRef]
- Baños, A.; Ariza, J.J.; Nuñez, C.; Gil-Martínez, L.; García-López, J.D.; Martínez-Bueno, M.; Valdivia, E. Effects of Enterococcus faecalis UGRA10 and the enterocin AS-48 against the fish pathogen Lactococcus garvieae. Studies in vitro and in vivo. Food Microbiol. 2019, 77, 69–77. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Reports summarized in this review for each topic and dairy microorganisms involved.
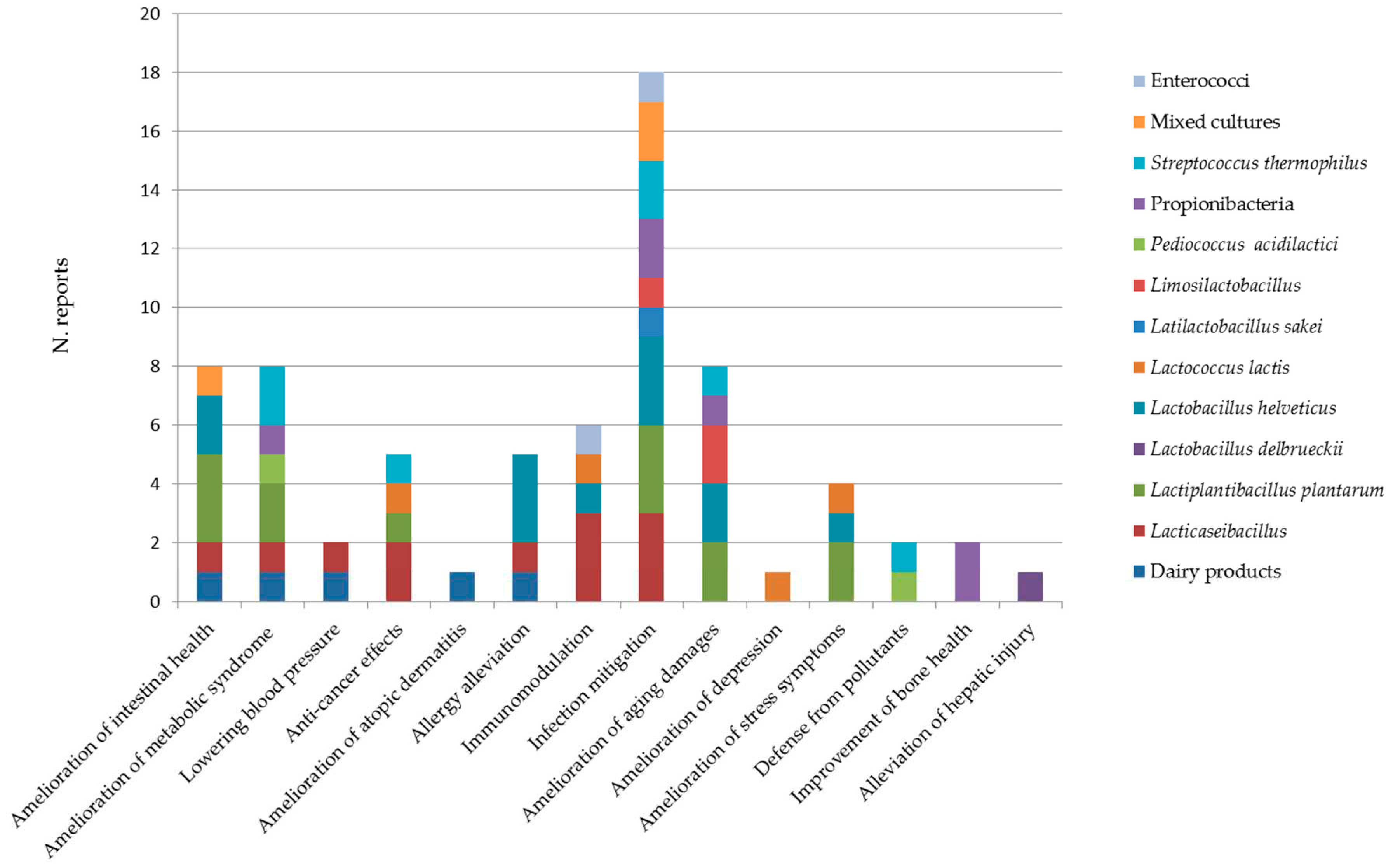
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



