Submitted:
15 May 2023
Posted:
16 May 2023
You are already at the latest version
Abstract
The antibodies targeting invasive proteins of Plasmodium falciparum (Pf) merozoites is a clever strategy against malaria parasite. The recent finding that Plasmodium merozoites surface enolase is a target for growth neutralizing antibodies has generated interest in identifying its re-ceptor (s) on erythrocytes. In the present study, several biochemical experiments were undertaken involving recombinant Pf enolase and antibody against it to search for its cognate receptor. The binding of rPfeno to erythrocytes was selectively sensitive to trypsin but resistant to neuramini-dase. Failure of yeast enolase binding to human red blood cells under similar conditions indicat-ed high specificity of parasite Pfeno. The mass spectrometric and immunological analyses of pro-teins obtained in pull downs and co-immunoprecipitation samples led to identification of band 3 as an interactor/receptor for Plasmodium enolase. Structural characterization of sequences of band 3 - rPfeno interacting regions revealed C-terminal exocytic regions of band 3 to be binding to Pfeno. Similarly, band 3 binding regions of rPfeno were also identified. Identification of receptor-ligand (band 3: Pfeno) pair paves the way for antimalarial anti-Pf enolase-based vaccine, anti-Pf enolase antibodies as drug and also for developing novel small molecule-based invasion inhibi-tory therapeutics including peptidomimetics that could disrupt the band 3: Pfeno protein-protein interactions.
Keywords:
Malaria
; Plasmodium enolase
; Band 3
; antibodies
; hRBCghosts
1. Introduction
Malaria is one of the most devastating parasitic infections, associated with a huge burden of morbidity and mortality [1]. P. falciparum, the causative agent for the deadly disease is an obligatory intracellular parasite. In its blood stage of infection, it invades erythrocytes making erythrocyte invasion an attractive target for malaria intervention. Invasion of host cells require interaction between parasite ligands and host cell receptors. In a bid to evade challenges, P. falciparum dabbles in extensive functional redundancy by deploying multiple invasion pathways that use different proteins as parasite ligands pairing with different receptors on erythrocytes. Parasite proteins which act as ligands come from two major families: Duffy binding like or erythrocyte binding like (DBL/EBL) and the reticulocyte binding like (RBL) proteins [2,3,4]. In P. falciparum, there are four and five known ligands of EBL class (EBL-175, EBL-1, EBA-140 and EBA-181) and RBL class (Rh1, Rh2a, Rh2b, Rh4 and Rh5) respectively [5]. The erythrocyte receptors that P. falciparum engages for invasion could be placed in two categories: the ones with sialic acid (SA) and the others without SA [6,7,8,9]. Accordingly, P. falciparum invasion pathways are either SA-dependent (SAD) or SA-independent (SAID). Interestingly, the malaria parasite has an uncanny ability to spontaneously switch from one invasion phenotype to another presumably to circumvent the host immune response [10]. Erythrocyte Glycophorins A-C are sialylated glycoproteins that serve as receptors for EBA-175, EBA-140 and EBL-1 respectively [7,11,12,13]. The other receptors that have been shown to be used by the parasite include complement receptor 1 (CR1) [14,15,16], basigin [17] and band 3 [18]. CR1 and basigin serve as receptors for Rh4 [4,15] and Rh5 [17] respectively. Apart from these, there are a host of other merozoite cell surface proteins that are believed to participate in the complex events of host cell invasion [5,19]. These include MSP1 that forms protein-protein complex with band 3 of erythrocyte [18], and PfMTRAP that binds to erythrocyte Semaphorin 7A [20]. However, there may be many more parasite ligands that are yet to be identified [21,22]. Thus, the parasite proteins expressed on the cell surface of invasive stages (i. e. sporozoites, merozoites and ookinetes) deserve to be examined for their functional involvement in host cell invasion. P. falciparum enolase (Pfeno), a moonlighting glycolytic enzyme has emerged as one such antigen. Surface expression of this protein has been observed in all three invasive forms of the parasite [23,24,25,26]. Exposure to antibodies against recombinant Pfeno (rPfeno) shows potent growth/invasion inhibitory effect on in vitro blood stage cultures presumably due to the blockade of merozoite surface enolase [26,27,28]. Likewise, antibody blockade of ookinete surface enolase is known to bring about inhibition of the ookinete traversal across the mosquito midgut epithelium [24]. Indeed, an enolase binding protein (EBP) has been isolated from mosquito gut epithelium that acts as host cell receptor for binding to ookinete cell surface enolase [29]. Molecular mechanisms of membrane bound enolase function in merozoite invasion of erythrocyte are yet to be explored. Recent observations about the ability of a monoclonal antibody directed against a unique pentapeptide (-104EWGWS108-) epitope of Pfeno to bring about potent invasion inhibition [28] indicate Pf enolase to serve as an invasion ligand. This would imply the presence of a receptor for merozoite enolase on host cell surface. In the present study, a series of experiments were undertaken in an attempt to identify and characterize the erythrocyte receptor for Pfeno. Multiple binding assays were performed to affinity pull down or co-immuno-precipitate the Pfeno binding human red blood cells (hRBC) membrane protein (s) and identify the pulled-out proteins by mass spectrometry and western blotting. Our results indicate that band 3 of hRBC is a Plasmodium merozoite cell surface enolase binding protein. In the present study we have also identified the peptide sequences in Pf enolase and band 3 that make the band 3 -Pf enolase interface which seems to facilitate invasion.
2. Materials and Methods
All the chemicals used were of analytical grade. The sources of different reagents were as given: Protein A-Sepharose (Pierce), Sepharose beads coupled to glutathione (GE Healthcare , Uppsala Sweden), Yeast enolase , anti-yeast enolase antibodies (α-Yeno), anti-band 3 antibodies (α-band 3), anti-spectrin antibody (α-spectrin), anti MSP119 antibody and Integrin alpha 7 antibodies (Sigma-Aldrich), Protease inhibitor cocktail (Roche Applied Science, Indianapolis, USA), HRP conjugated anti-mouse IgG secondary antibody (Molecular Probes, NJ, USA), Vectashield-mounting medium (Vector laboratories CA, USA), Biotin-labelled peptide library (15-mer peptides with five overlapping amino acids) covering the full sequence of Pfeno (Mimotopes, Australia) , synthetic band 3 peptides ( GenScript, USA Inc). Maxi-pro ELISA plates (Nunc), ready to use ABTS (, 2'-Azino-Bis-3-Ethylbenzothiazoline-6-Sulfonic Acid) (Sigma Aldrich, St. Louis, MO, USA), PVDF membrane (Millipore), Complete RPMI (cRPMI: RPMI with 0.5% Albumax II), Dibutyl phthalate, Iodoacetamide, Trypsin, Chymotrypsin, Neuraminidase and Coomassie stain (Sigma Aldrich). Sequencing grade Trypsin (Promega Madison, WI, USA).
2.1. Purification of recombinant P. falciparum enolase (rPfeno), preparation of Human RBC ghost, packed RBCs, and raising anti-rPfeno antibodies (α-rPfeno):
Recombinant P. falciparum enolase (rPfeno) [30,31], mouse anti-enolase antibodies (α-rPfeno) [31] and Human Red Blood Cell ghosts (hRBCG) were prepared as described earlier [32]. Starting with human blood taken in heparinized tube, packed RBCs were prepared by centrifuging the sample at 14000 x g for 20 min. Cellular pellet was washed thrice with incomplete RPMI medium. The washed hRBCs were stored at 4OC and used within 48 hr after collection. The α-rPfeno antibody was raised in Swiss mice in the animal house facility at TIFR and obtained as described earlier [31].
2.2. Erythrocytes binding Assays:
Packed hRBCs (100 µl) were incubated with varying concentrations of rPfeno or Yeast enolase (Yeno as control samples) in RBC binding buffer (50 mM Tris -HCl pH 8, 200 mM NaCl, 1mM EDTA, 2.5 mM MgCl2, 2 mM dithiothreitol and 1% glycerol) for 2 hours (hrs) at 37O C. Samples were centrifuged and hRBC pellet was washed twice with the binding buffer. This pellet was then layered on 100 µl of dibutyl phthalate and centrifuged (14000 x g , 5 min). The aqueous supernatant and the organic liquid layers were removed by aspiration. The RBC pellet was washed with phosphate buffer saline (PBS :10 mM Na-Phosphate, 137 mM NaCl, 2.7 mM KCl, pH 7.4). Recombinant Pfeno bound to the hRBCs was eluted using 25 µl of 2M-NaCl in PBS. The desalted eluate was run on 10% SDS-PAGE and the bound rPfeno / Yeno were visualized using western blot with α-rPfeno and α-Yeno antibodies respectively. In another experiment 100 µl of packed hRBCs in PBS were treated with trypsin, chymotrypsin (0.1 and 0.5 mg/ml) and neuraminidase (25 and 66 mU/ml) for 12 hours at 37OC. Post treatment the enzymes were inactivated by incubating with 1mM PMSF for 1 hr. Samples were centrifuged (2100 x g, 10 min) and the pellet was washed five times with PBS containing 0.1 mM PMSF. The enzyme treated hRBCs were then used for binding assay [33] as described above. Protein samples were analyzed on 10% SDS-PAGE. Western blotting was performed to detect the bound enolase using appropriate antibodies (α-Pfeno / α-Yeno). For binding with human RBC ghosts (hRBCG), ~100 µg protein equivalent ghost membranes were incubated with varying amounts of rPfeno (0, 5 and 25 µg) or Yeno (0, 5 and 100 µg) in PBS for 1 hr at 37O C. Post incubation, hRBCG membranes were collected by centrifugation and washed with PBS to remove unbound rPfeno. After each wash, absorbance at 280 nm was checked and wash step was repeated till the absorbance dropped to zero. Samples were analyzed on 10% SDS-PAGE and bound rPfeno was visualized by western blotting.
2.3. Blot Overlay assay:
The blot overlay assay or Far-western blot was done in accordance with the published protocol [34]. The blot bearing hRBCG proteins was incubated with varying concentrations of rPfeno (or Yeno as negative control) in protein binding buffer (PBB) and washed with PBST. The presence of hRBCG protein bound enolase was detected by probing the blot with α-rPfeno or α-Yeno antibodies.
2.4. Pull Down assay:
In a scouting experiment, purified hRBCG were solubilized using different detergents (CHAPS, NP-40 and Triton-X100) to identify the detergent for maximal extraction of hRBCG proteome [35]. In brief, the solubilized hRBCG proteins were incubated with purified glutathione S transferase tagged rPfeno (GST-rPfeno) for overnight at 4OC. For pulling down the GST-rPfeno bound with hRBCG protein(s), GSH- Sepharose 4B resin was added and allowed to incubate for 1 hr. In another set of experiments (for detection of non-specific binding), a control sample consisting of solubilized hRBCG were incubated with the Glutathione S-Sephadex-beads (GSH-B).The glutathione beads from the two sets of samples were separated by centrifugation (2000 rpm, 2 min), six washes of PBS were given, washed beads were suspended in SDS-PAGE sample buffer, boiled and proteins in the supernatant were resolved on 10% SDS PAGE, followed by silver staining. The bands (other than GST-rPfeno) were cut and processed for protein identification by mass spectroscopy.
2.5. Immuno-precipitation (IP) and reverse IP using Protein-A (PrA):
Protein A beads (PrA-B) (20 µl) were incubated with 2 µl α-Pfeno polyclonal antibody (1:200) for 6-8 hrs at room temperature (RT) and PrA-α-Pfeno beads were washed six times with PBS to remove unbound α-Pfeno. In a separate set, solubilized hRBCG proteins were incubated with rPfeno (in PBS, pH 7.4) overnight at 4OC. The solubilization was performed using solubilization buffer (20 mM Tris HCl pH-8, 137 mM NaCl, 10% glycerol and 1% NP-40 in presence of protease inhibitor cocktail). PrA-α-Pfeno beads prepared as above were added to the solubilized hRBCG -rPfeno sample and incubated for 2 hrs at RT. After incubation, beads were washed with PBS and collected by centrifugation (2000 rpm, 2 minutes) and then analyzed for rPfeno-bound proteins on 10% SDS-PAGE. Western blotting was done using α-band 3, α-Spectrin and α-integrin alpha-7 antibodies. In reverse IP experiments, 20 µl of PrA-B were pre-incubated with antibodies against band 3/ Spectrin / Integrin alpha-7. These pre-incubated PrA-B were then mixed with solubilized hRBCG proteins that were pre-incubated with rPfeno overnight at 4O C and for 2 hr at RT. The PBS washed beads were collected by centrifugation, solubilized with Laemmli buffer, run on 10% SDS-PAGE and transferred to a blot which was then probed using α-Pfeno to detect rPfeno bound hRBCG proteins. In an IP coupled with blot-overlay experiment, the band 3 protein from hRBCG extract was pulled out using PrA-α-band 3 as antibody-antigen complex (PrA-α- band 3-band 3). The beads were washed and mixed with Laemmli buffer, run on 10% SDS-PAGE and proteins were transferred to a blot. This was followed by overlay with rPfeno. The washed blot was then probed with α-Pfeno to confirm direct binding of immuno purified and immobilized band 3 with purified rPfeno.
2.6. Two-dimensional Gel Electrophoresis (2DE) of hRBC ghost and blot overlay:
hRBCG (120 µg protein) were solubilized in 100 µl of rehydration buffer (7M urea, 2M thiourea, 2% CHAPS, 50 mM DTT and 2% IPG buffer). The IPG strips (11 cm, 4 - 7 pH) were hydrated overnight and used for iso-electric focusing (IEF) with the following program, step 1: 250 V for 30 mins, step 2: 4000 V for 1hr and final step 3: 15000 V hrs. The strips were equilibrated with 2% DTT followed by 2.5% IAA (Iodoacetamide) (10 minutes each) before proceeding to second dimensional 10% PAGE. Three replicate gels were used for this sample. First gel was silver stained to profile the rehydration buffer solubilized proteome of hRBCG. Second gel was used for a western blot using α- band 3 to visualize band 3 and its variants. Third gel was used to blot the proteins on to a PVDF membrane, followed by blocking and overlay with rPfeno (10 µg) in protein binding buffer (10 ml) overnight at 4O C, washing and then probing with α-Pfeno. The method used for blot overlay with rPfeno followed by western blot was the same as described above.
2.7. In-gel Digestion of 2DE protein spots and Mass spectrometric analysis:
Spots corresponding to rPfeno binding proteins observed in gel overlay assay were excised from silver-stained gel and In-gel digestion was carried out using trypsin [36]. The gel pieces were treated with 25 mM ammonium bicarbonate (ABC): acetonitrile (ACN) 1:1 v/v). This was followed by reduction using 10 mM DTT at 56 ℃ for 1 hr and alkylation with 55 mM iodoacetamide at room temperature for 45 mins. Gel pieces were washed with 25 mM ABC and dried in vacuum evaporator. Finally, trypsin (400 ng) was added to gel pieces and the mixture was incubated for 16 hrs at 37O C for digesting proteins to peptides. The peptides were eluted with 70% ACN containing 5% formic acid. Eluted solutions were dried in speed-vac evaporator at 30O C. Peptides were dissolved in 0.1% formic acid (5 µl). Extracted peptides were analyzed on a Nano LC-ESI-MS/MS (Agilent 6250-Q-TOF) as described elsewhere [37]. Briefly, the extracted peptides dissolved in 0.1 % formic acid were applied to an Agilent HPLC chip (G4240-62002) which had a compact 40 nl C18 RP enrichment column and a 75 µm x 150 mm separation column that was packed with Zorbax 300 SB-C18 (5 µm) resin. Mobile phase solvents used were, solvent A: 0.1% formic acid in water and Solvent B: 0.1% formic acid in 90% ACN. After injection of the sample, column was washed with a 3 -12% gradient of solvent B for 3 min and the peptides were eluted with linear gradients of varying slopes, viz. 12-60% B for 3 to 23 min, and 60-95% B from 23 to 27 min. The mass spectrometric analysis was performed in extended dynamic mode in 50-1700 m/z range with active exclusion 0.5 min after each MS/MS spectrum. The scan rates (spectra/sec) for MS and MS/MS were 3.66 and 2.19 respectively. The acquired data were analyzed using Mass hunter software (Agilent technologies version B.07.00). For database search Mascot (Matrix sciences version 2.3.2) in-house server was used. The mascot generating files (mgf) were submitted to updated NCBI and Swiss-Prot database with parameter taxonomy set as Homo sapiens, fixed modification carbamidomethyl (cysteine), missed cleavage max 2, peptide mass tolerance 50 ppm and MS/MS mass tolerance of 0.6 Da.
2.8. P. falciparum and P. berghei parasite preparations:
P. falciparum 3D7 were maintained in culture. Briefly, human blood from healthy adults was collected in acid citrate dextrose (ACD) as the anticoagulant. After removing the leukocytes, the erythrocytes were washed and suspended in RPMI with 0.5% Albumax (cRPMI). Parasites were maintained at 5% haematocrit in cRPMI at 37O C in a humidified chamber containing 5% CO2. The ring stage synchronized P. falciparum culture was allowed to grow for 48 hrs at which time when the egressed merozoites would be maximum in the medium, the merozoites from the supernatant were harvested (at a parasitemia of ~ 5%) by centrifugation (1500xg, 10 min), and used for binding experiments. P. berghei ANKA infected RBCs (~106 cells) were injected into Swiss mice intraperitoneally. Parasitemia was monitored every day and when it reached to ~40%, infected blood (1-2 ml) was collected in equal volume of ACD solution. For the preparation of merozoites, blood samples were centrifuged and cellular pellet was washed thrice with cRPMI. Cell pellet was suspended in 60% ice cold percoll and spun at 1500xg in a swing-out rotor at 4O C for 15 min. Cellular layer at the interphase of aqueous and percoll media was collected. The infected RBCs in this layer were rich in schizonts and trophozoites. The interphase layer cells were repeatedly passed through a 25-gauge needle to release merozoites. Cellular debris from this suspension was removed by centrifugation at 600xg and the supernatant was centrifuged at 18,000xg for 15 min. to pellet down the merozoites [38]. Both the P. falciparum and the P. berghei merozoite pellets were washed with PBS and suspended in 100 μl of non-denaturing lysis buffer (20 mM Tris-HCl, pH 8 containing 137 mM NaCl, 10% glycerol, 1% NP-40 and protease inhibitor cocktail) and incubated on ice for 30 min. Suspension was centrifuged at 20,000xg (15 min, 4OC) and supernatant was used as merozoite cellular extract. These native parasite preparations and hRBC ghost preparations (as the source for band 3 protein) were used for blot overlay, IP and reverse IP experiments.
2.9. ELISA for monitoring protein-protein interaction:
A library of biotin labelled fifteen-mer peptides (overlapping by five amino acids) covering the entire sequence of Pfeno {Accession No. Q27727, purchased from Mimotopes (Australia)} was immobilized on a streptavidin coated ELISA plate. The purity of the peptides was > 80% and the amino acid composition of each peptide was confirmed by mass spectrometric analysis. The custom ELISA plates consisted of 2 wells (wells 89 and 90) coated with control peptides. The methodology followed for ELISA was according to manufacturer’s protocol. In brief the Pfeno peptide coated wells were blocked with sodium caseinate (1%w/v) for 2 hrs at 22O C, washed with PBST (phosphate buffer saline containing 0.05% Tween 20, pH 7.4) followed by incubation with solubilized hRBCG proteins at 37O C. Plates were developed with α-band 3 monoclonal antibody (1:5000). This was followed by addition of HRP conjugated mouse secondary antibody at a dilution of 1:1000 and incubation at room temperature for 45 min. The solution was discarded, and wells were washed thrice with PBS. Ready-to-use ABTS substrate (100 µl) was added. The color was developed for 10-15 min and the OD was measured at 405 nm on a Tican Plate Reader. ELISA values for control peptides were subtracted from value of each well before plotting the data.
For the converse experiment, i. e. to monitor the rPfeno - band 3 peptide interaction, the ELISA plate wells were coated with different synthetic peptides representing various regions of band 3. Binding of these peptides to rPfeno was probed by co-incubating with 1µg of purified rPfeno in PBS (100 µl) for 6 hrs at 37O C. The wells were washed with PBST followed by incubation with α-Pfeno (1:2000) for 1 hour, washing three times with PBST, adding HRP conjugated mouse secondary antibody (1:1000) into the wells and incubating it at room temperature for 45 minutes. The supernatant was discarded, wells were washed with PBS and ready to use ABTS (200 µl) added into each well. The color was developed for 10-15 min. and absorbance was measured at 405 nm on a Tican Plate Reader.
2.10. Molecular Modelling:
The UniProt database was used to obtain the amino acid sequence of Pfeno (UniProt Accession no. Q27727; 446 residues). The structure was visualized using PyMOL 2.1.0. The 3D structure of Plasmodium falciparum enolase (Pfeno) was modelled using the crystal structure of Toxoplasma gondii enolase 1 (PDB 3OTR) [39] as a template, with the help of the homology modeller SWISS MODEL (http://swissmodel.expasy.org/). Toxoplasma gondii enolase 1 was used for modelling since it shows >90% sequence identity with Pfeno.
3. Results
3.1. Binding of rPfeno to hRBCs and hRBC ghosts:
To test for the binding of rPfeno to human red cells, rPfeno was incubated with hRBCs and the washed red cells were then run on SDS-PAGE. Western blot of such a gel was probed with α-rPfeno (Figure 1A). Lanes 2-4 showed a concentration dependent binding of rPfeno to hRBCs. The ligand concentration dependent binding isotherm for rPfeno showed a typical hyperbolic saturation response with increasing amounts of rPfeno (Figure 1B) suggesting that binding is most likely to a specific receptor.
Specific nature of this binding was further supported by the failure of yeast enolase to bind to the erythrocytes under similar experimental conditions (Figure 1C). Effect of prior treatment of hRBCs with trypsin and chymotrypsin on binding of rPfeno revealed that treatment with low concentrations of trypsin eliminated the rPfeno binding (Figure 1D, lanes 3 and 4) while it required high concentrations of chymotrypsin to observe a similar effect (Figure 1D, lanes 5 and 6). A negative control showing that hRBCs did not have any cross-reacting proteins to α-rPfeno is shown in lane 1 (Figure 1D). Untreated hRBCs incubated with rPfeno showed the normal binding as expected (positive control lane 2). The Western blots (Figure 2) show the specificity of direct binding of rPfeno to hRBCG.
The concentration dependent interaction of parasite enolase with the ghost membranes was evident from the greater binding of rPfeno in 25 µg /ml sample as compared to the 5 µg/ml sample (Figure 2A). In a control sample where rPfeno was replaced with yeast enolase and the western blot was probed with α-yeast enolase antibody (α-Yeno), there was no detectable binding of Yeno to hRBC even at a concentration as high as 100 µg/ml (Figure 2B).
3.2. Pfeno interacting membrane proteins in hRBC:
Blot overlay assay (Method section - 2.3) was used to identify the likely interactors of rPfeno in hRBCG proteins. hRBCG proteins separated on a gel were allowed to interact with rPfeno and binding was detected by western blot analysis using α-rPfeno. Figure 3A (lane 2) shows the profile of proteins in hRBCG proteome. Western blot of a replicate of lane 2 did not show any cross reactivity with α-rPfeno (Figure 3A, lane 4, a negative control) while it was able to detect purified rPfeno (Figure 3A, lane 3, a positive control). Results showed the binding of rPfeno to hRBCG proteins at molecular masses ~91, 65, 48, 35 and 25 kDa (Figure 3B, lane 2; marked with arrowheads). Further, similar experiment with yeast enolase yielded negative (no binding) results (Figure 3C, lanes 2 and 3) indicating that binding of rPfeno to hRBCG proteins was specific to parasite enolase.
3.3. Molecular Identification of rPfeno binding proteins in hRBC ghosts:
GST-tagged rPfeno was immobilized on GSH-B and used to pull-down rPfeno binding protein(s) from hRBCG extracts. A silver-stained gel of the hRBCG proteins captured using different detergents is shown in Figure 4A (lanes 2 - 4). Lane 5 shows a control sample where GSH-Beads (GSH-B) were incubated with hRBCG extract to determine any non-specific adsorption of proteins by the beads. The bands marked with rectangular boxes (A-F) shown in various detergent solubilized hRBCG pull-down samples represented the potential Pfeno binding proteins. The proteins identified in various bands using Mass spectrometry are listed in Table 1 along with their Protein scores. Two major proteins identified in these bands were spectrin and band 3. Interestingly, two other proteins ankyrin and integrin-alpha 7 were also detected. This was understood in the background of the fact that for the binding of merozoite cell surface enolase with a receptor on hRBC, it would be essential that binding domain of the receptor be exposed on the cell surface. Among the above mentioned four proteins, it is interesting to note that band 3 is the only one that has an extracellular domain [18,32]. Ankyrin and spectrin are peripheral membrane proteins associated with cytoskeletal structures and are likely to be captured in pull-down along with band 3.
In a separate experiment a pull-down assay was performed on a Triton-X100 solubilized sample of hRBCG that were pre-treated with alkaline solution to eliminate peripheral proteins [33]. Lane 1 in Figure 4B shows the protein profile of alkali treated hRBCG proteome. For the pull-down, immobilized GST-rPfeno was incubated with the solubilized extract of alkali treated hRBCG. A 91 kDa protein bound to the parasite enolase was detected in this assay (Figure 4B, lane 3). Sepharose beads (without rPfeno) did not show any non-specific binding in hRBCG extract (Figure 4B, lane 2; negative control). A western analysis of the three samples using α-band 3 to probe the blot showed the presence of band 3 in hRBCG extract (Figure 4C, lane 3) as well as in the pull-down sample (lane 1). Thus, in this assay where peripheral membrane proteins were removed from hRBCG, only band 3 was identified as an interactor of Pfeno (Figure 4C). The major protein band in the pull-down sample was of ~91 kDa besides which some low molecular weight proteins were also identified by band 3 antibody.
The pull-down protein profiles in samples solubilized using different detergents (CHAPS, NP-40 and Triton X100) showed variation in the isoforms of band 3 captured by rPfeno indicating differential solubilization properties of each detergent (Figure 4A, Table 1). CHAPS and NP-40 solubilized samples yielded lower molecular mass (25 and 48 kDa) variants of band 3 while Triton-X100 solubilized sample captured band 3 at ~91 kDa. Results presented in Figure 4B and 4C provide antibody-based identification of band 3 as the most likely candidate for Pfeno binding to hRBC.
3.4. The band 3 of human erythrocytes interacts with rPfeno:
The co-immuno-precipitate obtained using Protein A Beads (PrA-B) bound to α-Pfeno (PrA-B-αPfeno) along with the two control samples was analyzed on SDS-PAGE {Figure 5A(i)}. A western blot of the gel probed with α-band 3 showed three different forms of band 3 in hRBCG detergent extract (Figure 5 A(ii) lane 1). A control sample where PrA-B were incubated with hRBCG extract (lane 2) indicated that agarose beads did not bind any band 3 protein. In the sample where hRBCG extract was incubated with rPfeno, prominent bands were observed at ~64 and 91 kDa {Figure 5A (ii) lane 3}. In the PrA-B-αPfeno system, the pull-down of band 3 with anti-rPfeno antibody implied the formation of a ‘band 3-rPfeno complex’. Similarly, α-band 3 antibody was immobilized on Protein A beads (PrA-B-α-band 3) and used to pull-down band 3 protein from a pre-incubated mixture of solubilized hRBCG and rPfeno. Further the western blot analysis of co-immuno-precipitate (probed with α-Pfeno) showed the presence of rPfeno at ~50 kDa {Figure 5B (ii) lane 3}. As expected, there were no α-Pfeno cross-reacting proteins in hRBCG (lane 1, control) and absence of any non-specific binding of rPfeno to PrA-B (lane 2, control). Also, the Western blot analysis of a pull-down sample obtained using α-rPfeno antibody bound Protein-A beads and probed with α-Spectrin antibody did not show spectrin in co-immunoprecipitate (data not shown). The pull-down sample was analyzed on SDS-PAGE. The Coomassie staining of the gel showed protein bands at molecular masses of 25, 64 and 91 kDa {Figure 5C (i) lane 2}. A blot of this immunoprecipitated sample was over laid with rPfeno and subsequently probed to detect the bound rPfeno using α-Pfeno antibody. The rPfeno showed specific binding to all three isoforms of the band 3 {Figure 5C (ii) lane 2 marked with arrowheads}. Protein bands at 91 and 64 kDa showed more pronounced binding of rPfeno as compared to the 25 kDa form of band 3 protein. Co-immunoprecipitation of rPfeno and band 3 in the reciprocal pull-down assays revealed direct interaction of rPfeno with band 3 providing unequivocal evidence for the direct interaction of parasite enolase to band 3 protein on human RBC plasma membrane.
3.5. Native enolases from P. falciparum and P. berghei merozoites bind to hRBC band 3:
Since all the earlier binding studies were carried out with recombinant Pfeno protein, we wanted to confirm whether native Plasmodium enolase also interacts with hRBC band 3 protein. Hence blot overlay and pull-down experiments were carried out using P. falciparum culture supernatant (as a source of P. falciparum merozoites) and P. falciparum infected RBCs (iRBC). Figure 6, Panel A, lane 2 shows a blot of hRBC ghost preparation, overlaid with merozoite extract, and then probed with α-Pfeno antibodies. The bands that light up are shown with arrows and some of them correspond to degradation products of band 3 protein. In lane 3 we show a pull-down experiment using Protein A beads coupled with α-band 3 antibodies, incubated with P. falciparum iRBC extract. The pulled-down proteins were run on a 10% SDS-PAGE and the Western blot was probed with α-band 3 antibodies. The band 3 protein was indeed seen to be pulled down from the P. falciparum iRBC extract. This pull down and the blot overlay showed band patterns similar to each other and was commensurate with canonical human band 3 protein fragments on SDS-PAGE. Lane 1 is a control lane to show that the α-Pfeno antibodies reacted with rPfeno protein specifically. Our P. falciparum cultures typically had a parasitemia of about 5-10%, and we realized that an iRBC preparation would contain uninfected RBCs as well. Therefore, the band 3 protein pull down in lane 3 could be due to pull down of the protein from uninfected RBCs. In order to ascertain whether band 3 protein did interact with native Pfeno present in iRBC, pull-down experiments were carried out with Protein beads coupled to α-band 3 antibodies, incubated with P. falciparum iRBCs, which were probed with α-Pfeno antibodies (Figure 6, Panel B). The α-band 3 antibodies did pull-down Pfeno from P. falciparum iRBCs as observed in lane 3. A similar result was observed when the rodent malaria parasite P. berghei infected RBCs were incubated with protein A beads coupled with α-band 3 antibodies (Panel B, lane 4). Lane 1 shows the molecular weight markers labelling the blot, while lanes 2 and 5 are controls to show that α-Pfeno antibodies are specific, since they react with rPfeno, but not with proteins from uninfected RBCs.
It is envisaged that the human RBC band 3 protein is a relevant binding partner for parasite enolase present on the merozoite surface. Since the P. falciparum culture did not provide us with adequate amounts of purified merozoites, we decided to use P. berghei merozoites purified as described in the Methods section. Protein profiles of the merozoite (M, lane 1) and hRBCG extracts (G, lane 2) are shown in a Coomassie stained gel in Figure 6C. Western blots of M and G preparations were made and probed with α-MSP-119, α-Pfeno and α-band 3 (Figure 6C lanes 3, 5 and 7). Absence of band 3 (lane 5) and the presence of MSP-119 (lane 3) and enolase (lane 7) in these blots validated the merozoite preparation. The cross-reactivity of our α-Pfeno antibodies to murine Plasmodium parasites has been demonstrated by us earlier. Immunoblots of hRBCG proteins showed no cross reactivity with α-MSP-119 (Figure 6C, lane 4) or α-Pfeno (Figure 6C, lane 8), but as expected, showed positive signals when probed with α-band 3 (Figure 6C, lane 6). In pull-down experiments, Pbeno was pulled out from Pb merozoite extract by incubation with PrA-B-α-Pfeno. The beads (PrA-B-α-Pfeno-Pbeno) were then incubated with hRBCG extract. Presence of band 3 in this pull-down was detected in a Western blot probed with α-band 3 (Figure 6 Panel D, lane 2). Conversely, PrA-B-α-band 3 was used to pull down band 3 from hRBCG extract as PrA-B-α-band 3-band 3 complex and then incubated with merozoite extract. Presence of band 3 protein in this pull-down was detected in a Western blot probed with α-Pfeno (Figure 6 Panel D, lane 4). The possibility of non-specific binding of band 3 or Pbeno proteins to PrA-B was ruled out by incubating PrA+ merozoite extract + hRBCG extract and probing with α-band 3 antibodies (lane 1); PrA-B-αPfeno + hRBCG ghost extract and probing with α-band 3 antibodies (lane 3) and PrA + merozoite extract and probing with α-Pfeno antibodies (lane 5). These controls showed an absence of nonspecific adsorption of band 3 or Pbeno on PrA beads under the experimental conditions used here. The sample where PrA-α-Pfeno-Pbeno incubated with hRBCG extract was probed with α-band 3 showed two bands at ~25 and 65 kDa (Figure 6D, lane 2) indicating the binding of band 3 to immobilized Pbeno. The blot where PrA-B-α-band 3-band 3 was incubated with merozoite extract and probed with α-Pfeno, showed an enolase band at ~50 kDa and a proteolyzed product at a lower molecular weight (Figure 6D, lane 4). Presence of band 3 in lane 2 and Pbeno in lane 4 (marked by arrows) indicated the formation of PrA-B-α-band 3-band 3-Pbeno and PrA-B-α-Pfeno-Pbeno-band 3 complexes in the two samples. Observation of low molecular weight forms of Pbeno (Figure 6D lane 4) and band 3 (Figure 6A lane 3) are due to possible proteolysis of native forms. The experiments shown in the Figure 6 confirm the binding of native Plasmodium enolases to human band 3 protein.
3.6. Post translationally modified variants of band 3 exhibit differential binding to rPfeno:
In a two-dimensional gel electrophoresis (2DE) experiment (Method section-2.6), we attempted to separate the different variants of band 3. A silver stained 2DE of hRBCG is shown in Figure 7A. Western blot of a replicate of this where band 3 was detected using α-band 3 showed two clusters of spots (Figure 7B). 25 kDa form of band 3 got resolved into at least five different spots with pIs ranging from 4.8-5.3 while the 64 kDa form showed two spots (pI~5.18). Intriguingly, the 91 kDa form was not observed in the western blot, presumably because it did not get solubilized in 2DE sample buffer. Using a replicate blot, the binding ability of band 3 variants to rPfeno was examined in an overlay assay. Interestingly, while rPfeno binding was observed with both forms at 64 kDa, the 25 kDa forms of band 3 exhibited binding to the three variants (pI~4.93, 5.03, 5.18). Band 3 variants with highest pI~5.31 and lowest pI~4.81 failed to bind Pf enolase (Figure 7C). The individual spots corresponding to band 3 (as seen in western blot Figure 7B) were excised from the silver-stained gel and subjected to Mass-spectrometry for the identification of the proteins. Two proteins that could be identified with high Prot Score were Spectrin and band 3. The protein spot at pI~5.18 and MW~64 kDa had both band 3 and spectrin while the ~25 kDa spot with pI~5.31 showed the presence of three proteins viz. diacylglycerol kinase, chromosome 20 ORF and cadherin 12 (Table 2). Although in Western blot using α-band 3, there was a positive identification of band 3 protein, mass spectrometry failed to confirm that. This can be due to missing the spot of interest on the silver-stained gel. Amounts of protein present in other spots with MW~25 kDa were too small to be identified by mass spectrometry.
As shown in Figure 8, band 3 has 911 residues (NCBI Reference Sequence Accession NP_000333.1). Table 3 lists the sequences of ten different band 3 peptides that were sequenced in mass spectrometry experiments covering ~15% (136 residues) of band 3. The residues that could be sequenced in peptides, led to identification of band 3.
3.7. Synthetic peptide towards mapping the binding sites of band 3 and Pfeno:
Band 3 is a trans membrane protein that has its N-terminal globular region on the cytosolic face while the membrane spanning and exocytic loop regions are at the C-terminal end [35,36]. Thus, the site for parasite enolase that binds to band 3 on extra cellular surface must map on the C-terminal region of band 3. For mapping the Pfeno binding region on band 3, a set of seven different peptides belonging to the exocytic domains of band 3 were custom synthesized (Table 4).
Four of these peptides (P632-655, P731–753, P744–758 and P852-879) localize in exocytic regions of band 3. Peptide P813-826 is from a trans membrane region while the peptides P220-246 and P247-283 are from the endocytic domain of the band 3 [36]. Sequence positions of all the seven synthetic peptides are underlined (red) in Figure 8A. C-terminal region of band 3 (aa 361-911) forms the trans-membrane domains and exocytic regions of the protein (highlighted in turquoise blue in Figure 8A). Using these peptides as baits, their ability to capture rPfeno was further tested in an ELISA experiment. The results of the assay (Figure 8B) indicated that while the peptide P813-826 that localizes in trans-membrane region had no interaction with rPfeno, the three exocytic peptides (P731–753, P744–758 and P852-879) showed significant binding and are likely to form the binding region for the parasite enolase. The remaining three peptides (P220-246, P247-283 and P632-655) showed intermediate level of reactivity (Figure 8B).
Interaction of rPfeno with the two N-terminal peptides (P220-246 and P247-283) is of unclear physiological relevance as these regions are located on the cytoplasmic face of the hRBC membrane. However, the possibility of post invasion interactions between enolase and the two N terminal peptides cannot be ruled out. Another peptide P632-655 that showed an intermediate level of binding is also likely to be a part of enolase binding site. Thus, the data presented here imply that rPfeno binding region of band 3 includes sequences P632-655, P731–753, P744–758 and P852-879. It is important to realize that the method used here is largely limited to identifying sequentially continuous binding sites. Protein-protein interactions that are stabilized by secondary and/or tertiary structural features formed by non-contiguous sequence would not be identified by this approach.
For mapping the Pfeno sites that bind to band 3, a ‘Peptide Scanning’ strategy was used. The method involved synthesis of entire enolase protein sequence as short biotin tagged 15 mer overlapping peptides and testing them individually for binding to band 3 [37]. Binding of band 3 with the Pfeno peptide library was monitored using an ELISA format. As shown (Figure 9A), band 3 showed binding with peptides 16, 40, 49, 50 and 57-64. The sequences of these band 3 binding Pf enolase peptides as also their two neighboring peptides that failed to bind band 3 are shown in Figure 9B. Sequence alignment among peptides 15 - 17 indicated that the peptide 16 shares ten residues with these two non-binding peptides. Thus, if any stretch of five or ten residues in peptide 16 formed a binding site, one or the other neighboring peptides could also show some binding. Absence of such recognition with flanking peptides 15 and 17 was indicative of nonspecific binding of peptide 16. Similar is the case with peptide 40 which also showed no binding for the flanking peptides. Peptides 49 and 50 have ten residues in common (Panel B, shown in green) and both showed interaction with band 3 indicating that these residues are part of the binding domain. The N-terminal five residues of peptide 49 and the C-terminal five residues of peptide 50 are not involved in the binding to band 3. This may be the reason why the flanking peptides 48 and 51 that did not have adequate number of binding domain residues failed to interact with band 3. Thus, from these data we infer that Pf Enolase region 246GKVKIAMDAV255 is involved in binding interactions with band 3. Similar analysis of the other cluster of peptides (peptides 57-64) yielded a binding sequence of 286QLVDLYIDLVKKYPIVSIEDPFDQDDWENYAKLTAAIGKD325 as the band 3 binding domain in Pfeno. To visualize the location of this sequence in 3D structure of Pfeno, a model structure of Pfeno was generated [38] using the crystal structure of T. gondii enolase (pdb: 3OTR) [39] as template (Figure 9 C) as the two proteins have >90% sequence homology. In this structure, location of these sequences is shown in blue and green. Enolase is also known to bind plasminogen that facilitates invasion of mosquito mid gut epithelium [24]. The plasminogen interacting domain is shown in red. All apicomplexan enolases differ from host enolases in having an insert of five amino acids (104EWGWS108) (Figure 9 C, pink). This epitope has been shown to be a target for potent growth inhibiting antibodies [27,28]. It is interesting to note that the domains of Pfeno that are involved in executing one or the other moonlighting function are clustered on the same face of the molecule in the 3D model structure (Figure 9 C).
4. Discussion:
Plasmodium falciparum enolase (Pfeno) is a highly conserved glycolytic enzyme that is expressed on the surface of the merozoites. Involvement of enolase in merozoite invasion of erythrocytes was evident from the observed strong inhibition of the P. falciparum growth in blood cell cultures by α-Pfeno and reduction of parasitemia in P. yoelii infected mice immunized with α-Pfeno [26,27,28,30]. The target epitope for the inhibitory α-Pfeno antibodies was identified as the pentapeptide insert 104EWGWS108 present in all apicomplexan parasite enolases but absent in host enolases [46]. As the initial invasion event requires interaction between a relevant parasite ligand with host receptor, an attempt was made to identify and characterize the Pfeno binder/receptor on hRBC surface. Several experiments presented here indicated that rPfeno bound specifically to both hRBC and hRBCG in a ligand concentration dependent manner. In the past, the effect of treatment of hRBC with proteases (trypsin and chymotrypsin) and neuraminidase on the binding of the parasite ligands have been used to define sialic acid dependent (SAD) or independent (SAID) pathways [6,7,8,9]. We found that the binding of rPfeno was sensitive to trypsin but resistant to neuraminidase suggesting that the rPfeno binding with the receptor is of the SAID kind. Results from a host of pull-down and overlay assays combined with western blotting and/or mass spectrometry (MS/MS) (Figs. 4-7) analysis led to identification of band 3 as the enolase binding protein in hRBC cell membrane. Band 3 is an anion exchanger that mediates the exchange of the cellular HCO3- with Cl- in plasma [42,47,48]. Structurally, it has 911 amino acid residues (Figure 8A) with a glycosylation site at N642. It is a trans-membrane protein that coimmunoprecipitates with RhopH3 and MSP1 [18,49]. Two extracellular domains of band 3, viz 5C (744ASTPGAAAQIGEVKE758) and 6A (807DRILLLFKPPKYHPDVPYVK826) form the binding site for MSP1. Interestingly, MSP1 mediated invasion pathway has also been shown to be of the SAID kind [18]. The enolase binding sequences identified here consisted of four exocytic peptides viz P632-655, P731–753, P744–758 and P852-879 (Figure 8A and 8B). Thus, there is quite a bit of overlap in the binding sites for MSP1 and Pfeno on band 3. Topographically, N-terminal part of band 3 is globular and located on cytosolic side while C terminal domain forms its transmembrane and exocytic regions [42,47]. As expected, the Pfeno binding site on band 3 mapped in C-terminal exocytic domain (Figure 8). Conversely, the binding site for band 3 on Pfeno was also mapped using a peptide scanning approach. The results showed that the Pfeno regions that bind band 3 are constituted by 246GKVKIAMDAV255and286QLVDLYIDLVKKYPIVSIEDPFDQDDWENYAKLTAAIGKD325 (Figure 9B). Pfeno appears to have multiple distinct sites that relate to its diverse physiological functions. In order to visualize the spatial disposition of various functional regions on the molecular surface of Pfeno, its 3D-structure was modelled using X-ray crystallographic structure of T. gondii enolase (Figure 9 C). Apart from the catalytic site (not shown), there is a plasminogen binding site constituted by 277DKSLVK282 residues [24], band 3 binding site formed by 246GKVKIAMDAV255and 286QLVDLYIDLVKKYPIVSIEDPFDQDDWENYAKLTAAIGKD325 and an invasion inhibitory antibody binding site 104EWGWS108 [27,28,30]. Presence of distinct Pfeno binding regions in band 3 and parasite growth inhibitory Pfeno antibodies indicates that band 3 enolase binding site may facilitate initial adhesion of merozoite to hRBC. The unique pentapeptide insert in Pfeno (104EWGWS108) is likely to serve as a docking port for some (yet to be identified) molecular factor of host origin whose interaction must be critical for merozoite invasion of hRBC in an enolase mediated pathway. In many pathogens as also in malignant cells, cell surface enolase is known to act as plasminogen binder which upon activation cleaves inter-cellular matrix protein facilitating the establishment of infection [25,50,51,52,53]. However, plasminogen binding function of Pfeno is unlikely to be of any relevance in blood stage merozoite invasion of hRBC as there is no barrier of matrix proteins between the host and the parasite.
Pfeno is an unusual surface antigen that expresses on cell surface of all the three invasive stages (sporozoites, merozoites and ookinetes) in Plasmodium’s life cycle [23,24]. In a series of elegant experiments, the physiological functions of ookinete cell surface enolase were demonstrated [23,24,25,29]. In mosquito mid gut, ookinete has to cross the peritrophic matrix barrier to reach to mid gut epithelium. This task is accomplished by binding of human plasminogen (derived from the blood meal of the mosquito) to ookinete cell surface enolase [24,54]. Disruption of this inter molecular interaction between Pfeno and plasminogen by anti-Pfeno antibodies or by feeding plasminogen depleted blood resulted in drastic reduction in traversal of ookinetes across the mid gut epithelium [24,29]. Most of the ookinete traversal inhibitory activity could be accounted for by the plasminogen receptor function of Pfeno. In addition, an enolase binding protein (EBP) that expresses on the luminal surface of the mosquito midgut epithelium was also identified [29]. Although physiological significance of Pfeno-EBP interaction is not well understood, it is likely to be important in mediating host cell recognition by the parasite (similar to merozoite cell surface enolase binding to band 3 of hRBC). Peritrophic matrix in mosquito mid gut is rich in proteins that contain PWWP, a protein-protein interaction motif [55]. Recently, this motif was shown to bind to 104EWGWS108 [56] raising an interesting possibility of aromatic interactions-based assembly of a ternary complex formed by Pfeno, plasminogen and PWWP containing matrix proteins, leading to high efficiency proteolysis of peritrophic matrix. Physiological functions of Pfeno on merozoite and ookinete cell surfaces are different and well adapted for host cell invasion needs of the parasite. Thus, it appears that enolase functions very much depend on when and where this versatile protein is present. This has been seen in several organisms where enolase has multiple moonlighting functions [57]. So far, the role of Pfeno on sporozoite cell surface has not been examined. It will be interesting to examine whether it has any functional role in sporozoite invasion of hepatocytes.
In conclusion, the identification of band 3 as a major erythrocyte receptor for the highly conserved glycolytic protein Pfeno should provide new insights in understanding the role of Pfeno on merozoite cell surface. More in-vivo and in-vitro studies such as RBC invasion inhibition assay using different laboratory strains and real time interaction using Surface Plasmon Resonance (SPR), are required involving both rPfeno and band 3 for better understanding of the host parasite interaction. It is hoped that the identification of erythrocyte band 3 as a receptor for Plasmodium enolase will provide a window of opportunity to develop new peptide/small molecule-based therapeutics and possible vaccine component(s) to combat the deadly disease of malaria.
Author Contributions
GKJ, conceived, planned and designed the study, planned laboratory work, participated in data analysis and interpretation, data presentation and manuscript preparation; RV, planned and executed most experiments, analyzed data and manuscript writing; CB, performed mass-spectrometry of proteins, data analysis; CB and RV performed 2-D gel electrophoresis and overlay experiments; SS, contributed to experimental design, data interpretation and manuscript editing, DS found new perspectives in the work and contributed to uplifting the manuscript.
Funding
This research was funded by grants to GKJ and SS from the Department of Atomic Energy, Government of India through Tata Institute of Fundamental Research (TIFR) Mumbai, India.
Ethical Statement
Human red blood cells (hRBC) were collected from volunteers, after obtaining their written consents, for all the RBC experiments including in-vitro cultures of Plasmodium falciparum. The procedure for such collection, details of informed consent and the frequency of samples to be collected were in accordance with a detailed proposal approved by the Institutional Human Ethics Committee (IHEC) of Tata Institute of Fundamental Research (TIFR). The IHEC of TIFR functions as per the guidelines of Indian Council of Medical Research (ICMR), Govt. of India. TIFR Animal House is registered under CPCSEA (Committee for the Purpose of Control and Supervision of Experiments on Animals), Govt. of India (registration no. 56/1999/CPCSEA) for breeding and experiments on animals. Plasmodium berghei were maintained in mice and the study was carried out under strict accordance with the guidelines of CPCSEA, with prior approval.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data is contained within the article.
Acknowledgments
We acknowledge Dr. Sanjeeva Srivastava (Department of Bioscience and Bioengineering, IIT-Mumbai) for the use of their Mass-spectrometry facility. We thank Ms. Anasuya Moitra for help in generating 3D model structure of Pfeno and Ms Sneha Dutta (Harvard University, Boston) for reading the manuscript and making helpful suggestion.
Conflicts of Interest
The authors declare there are no competing interest associated with the manuscript.
Abbreviations
ABTS, 2, 2'-Azino-Bis-3-Ethylbenzothiazoline-6-Sulfonic Acid; αPfeno, anti-rPfeno antibodies; αBand 3, anti-Band 3 antibodies; GSH, Glutathione S-Sephadex beads; GST-rPfeno, glutathione S transferase tagged rPfeno; IP, immune-precipitation; αMSP 119, anti MSP1 antibody; αSpectrin, anti spectrin antibody; αYeno, anti-yeast enolase antibody; 2DE, Two dimensional gel electrophoresis; hRBC, human red blood cells; hRBCG , human RBC ghosts; hrs, hours;), Pbeno, Plasmodium berghei enolase; PrAB, Protein A agarose beads; rPfeno, recombinant P. falciparum enolase; RT, room temperature. iRBC, infected RBC, uRBC, uinfected RBC
References
- WHO: World malaria report 2017. Available online: http://wwwwhoint/malaria/publications/world-malaria-report-2017/en/ (accessed on 10 April 2018).
- Iyer J, Gruner AC, Renia L, Snounou G, Preiser PR: Invasion of host cells by malaria parasites: a tale of two protein families. Mol Microbiol 2007, 65, 231–249. [CrossRef] [PubMed]
- Paul AS, Egan ES, Duraisingh MT: Host-parasite interactions that guide red blood cell invasion by malaria parasites. Curr Opin Hematol 2015, 22, 220–226. [CrossRef] [PubMed]
- Tham WH, Healer J, Cowman AF: Erythrocyte and reticulocyte binding-like proteins of Plasmodium falciparum. Trends Parasitol 2012, 28, 23–30. [CrossRef] [PubMed]
- Cowman AF, Tonkin CJ, Tham WH, Duraisingh MT: The Molecular Basis of Erythrocyte Invasion by Malaria Parasites. Cell Host Microbe 2017, 22, 232–245. [CrossRef] [PubMed]
- Deas JE, Lee LT: Competitive inhibition by soluble erythrocyte glycoproteins of penetration by Plasmodium falciparum. Am J Trop Med Hyg 1981, 30, 1164–1167. [CrossRef] [PubMed]
- Lobo CA, Rodriguez M, Reid M, Lustigman S: Glycophorin C is the receptor for the Plasmodium falciparum erythrocyte binding ligand PfEBP-2 (baebl). Blood 2003, 101, 4628–4631. [CrossRef] [PubMed]
- Perkins M: Inhibitory effects of erythrocyte membrane proteins on the in vitro invasion of the human malarial parasite (Plasmodium falciparum) into its host cell. J Cell Biol 1981, 90, 563–567. [CrossRef]
- Ord RL, Rodriguez M, Yamasaki T, Takeo S, Tsuboi T, Lobo CA: Targeting sialic acid dependent and independent pathways of invasion in Plasmodium falciparum. PLoS One 2012, 7, e30251.
- Awandare GA, Nyarko PB, Aniweh Y, Ayivor-Djanie R, Stoute JA: Plasmodium falciparum strains spontaneously switch invasion phenotype in suspension culture. Sci Rep 2018, 8, 5782. [CrossRef]
- Camus D, Hadley TJ: A Plasmodium falciparum antigen that binds to host erythrocytes and merozoites. Science 1985, 230, 553–556. [CrossRef]
- Li X, Marinkovic M, Russo C, McKnight CJ, Coetzer TL, Chishti AH: Identification of a specific region of Plasmodium falciparum EBL-1 that binds to host receptor glycophorin B and inhibits merozoite invasion in human red blood cells. Mol Biochem Parasitol 2012, 183, 23–31. [CrossRef] [PubMed]
- Mayer DC, Cofie J, Jiang L, Hartl DL, Tracy E, Kabat J, Mendoza LH, Miller LH: Glycophorin B is the erythrocyte receptor of Plasmodium falciparum erythrocyte-binding ligand, EBL-1. Proc Natl Acad Sci U S A 2009, 106, 5348–5352. [CrossRef] [PubMed]
- Awandare GA, Spadafora C, Moch JK, Dutta S, Haynes JD, Stoute JA: Plasmodium falciparum field isolates use complement receptor 1 (CR1) as a receptor for invasion of erythrocytes. Mol Biochem Parasitol 2011, 177, 57–60. [CrossRef] [PubMed]
- Spadafora C, Awandare GA, Kopydlowski KM, Czege J, Moch JK, Finberg RW, Tsokos GC, Stoute JA: Complement receptor 1 is a sialic acid-independent erythrocyte receptor of Plasmodium falciparum. PLoS Pathog 2010, 6, e1000968.
- Tham WH, Wilson DW, Lopaticki S, Schmidt CQ, Tetteh-Quarcoo PB, Barlow PN, Richard D, Corbin JE, Beeson JG, Cowman AF: Complement receptor 1 is the host erythrocyte receptor for Plasmodium falciparum PfRh4 invasion ligand. Proc Natl Acad Sci U S A 2010, 107, 17327–17332. [CrossRef] [PubMed]
- Crosnier C, Bustamante LY, Bartholdson SJ, Bei AK, Theron M, Uchikawa M, Mboup S, Ndir O, Kwiatkowski DP, Duraisingh MT, et al: Basigin is a receptor essential for erythrocyte invasion by Plasmodium falciparum. Nature 2011, 480, 534–537. [CrossRef] [PubMed]
- Goel VK, Li X, Chen H, Liu SC, Chishti AH, Oh SS: Band 3 is a host receptor binding merozoite surface protein 1 during the Plasmodium falciparum invasion of erythrocytes. Proc Natl Acad Sci U S A 2003, 100, 5164–5169. [CrossRef] [PubMed]
- Cowman AF, Berry D, Baum J: The cellular and molecular basis for malaria parasite invasion of the human red blood cell. J Cell Biol 2012, 198, 961–971. [CrossRef]
- Bartholdson SJ, Bustamante LY, Crosnier C, Johnson S, Lea S, Rayner JC, Wright GJ: Semaphorin-7A is an erythrocyte receptor for P. falciparum merozoite-specific TRAP homolog, MTRAP. PLoS Pathog 2012, 8, e1003031. [Google Scholar]
- Hu G, Cabrera A, Kono M, Mok S, Chaal BK, Haase S, Engelberg K, Cheemadan S, Spielmann T, Preiser PR, et al: Transcriptional profiling of growth perturbations of the human malaria parasite Plasmodium falciparum. Nat Biotechnol 2010, 28, 91–98. [CrossRef]
- Le Roch KG, Zhou Y, Blair PL, Grainger M, Moch JK, Haynes JD, De La Vega P, Holder AA, Batalov S, Carucci DJ, Winzeler EA: Discovery of gene function by expression profiling of the malaria parasite life cycle. Science 2003, 301, 1503–1508. [CrossRef] [PubMed]
- Bhowmick IP, Kumar N, Sharma S, Coppens I, Jarori GK: Plasmodium falciparum enolase: stage-specific expression and sub-cellular localization. Malar J 2009, 8, 179. [CrossRef] [PubMed]
- Ghosh AK, Coppens I, Gardsvoll H, Ploug M, Jacobs-Lorena M: Plasmodium ookinetes coopt mammalian plasminogen to invade the mosquito midgut. Proc Natl Acad Sci U S A 2011, 108, 17153–17158. [CrossRef] [PubMed]
- Ghosh AK, Jacobs-Lorena M: Surface-expressed enolases of Plasmodium and other pathogens. Mem Inst Oswaldo Cruz 2011, 106 (Suppl. 1), 85–90. [CrossRef] [PubMed]
- Pal-Bhowmick I, Mehta M, Coppens I, Sharma S, Jarori GK: Protective properties and surface localization of Plasmodium falciparum enolase. Infect Immun 2007, 75, 5500–5508. [CrossRef] [PubMed]
- Dutta S, DasSarma P, DasSarma S, Jarori GK: Immunogenicity and protective potential of a Plasmodium spp. enolase peptide displayed on archaeal gas vesicle nanoparticles. Malar J 2015, 14, 406. [Google Scholar] [CrossRef] [PubMed]
- Dutta S, Tewari A, Balaji C, Verma R, Moitra A, Yadav M, Agrawal P, Sahal D, Jarori GK: Strain-transcending neutralization of malaria parasite by antibodies against Plasmodium falciprum enolase. Malar J 2018, 17, 17.
- Vega-Rodriguez J, Ghosh AK, Kanzok SM, Dinglasan RR, Wang S, Bongio NJ, Kalume DE, Miura K, Long CA, Pandey A, Jacobs-Lorena M: Multiple pathways for Plasmodium ookinete invasion of the mosquito midgut. Proc Natl Acad Sci U S A 2014, 111, E492–E500.
- Vora HK, Shaik FR, Pal-Bhowmick I, Mout R, Jarori GK: Effect of deletion of a plant like pentapeptide insert on kinetic, structural and immunological properties of enolase from Plasmodium falciparum. Arch Biochem Biophys 2009, 485, 128–138. [CrossRef]
- Pal-Bhowmick I, Sadagopan K, Vora HK, Sehgal A, Sharma S, Jarori GK: Cloning, over-expression, purification and characterization of Plasmodium falciparum enolase. Eur J Biochem 2004, 271, 4845–4854. [CrossRef]
- Marchesi VT, Palade GE: The localization of Mg-Na-K-activated adenosine triphosphatase on red cell ghost membranes. J Cell Biol 1967, 35, 385–404. [CrossRef] [PubMed]
- Tyagi RK, Sharma YD: Erythrocyte Binding Activity Displayed by a Selective Group of Plasmodium vivax Tryptophan Rich Antigens Is Inhibited by Patients' Antibodies. PLoS One 2012, 7, e50754.
- Wu Y, Li Q, Chen XZ: Detecting protein-protein interactions by Far western blotting. Nat Protoc 2007, 2, 3278–3284. [CrossRef] [PubMed]
- Luche S, Santoni V, Rabilloud T: Evaluation of nonionic and zwitterionic detergents as membrane protein solubilizers in two-dimensional electrophoresis. Proteomics 2003, 3, 249–253. [CrossRef] [PubMed]
- Shevchenko A, Jensen ON, Podtelejnikov AV, Sagliocco F, Wilm M, Vorm O, Mortensen P, Shevchenko A, Boucherie H, Mann M: Linking genome and proteome by mass spectrometry: large-scale identification of yeast proteins from two dimensional gels. Proc Natl Acad Sci U S A 1996, 93, 14440–14445. [CrossRef] [PubMed]
- Jindal N, Balaji C, P.B. S, Dutta S, Jarori GK: Identification of Post-translational Modificatio ns of Plsmodium yoelii Glyceraldehyde-3-phosphate dehydrogenase by Mass Spectrometry. Adv Proteomics Bioinform: APBI - 103 2017.
- Moll K, Ljungstrom I, Perlman H, Scherf A, Wahlgren. M: Methods in Malaria Research (American Type Culture Collection, Manassas, VA).2008.
- Ruan J, Mouveaux T, Light SH, Minasov G, Anderson WF, Tomavo S, Ngo HM: The structure of bradyzoite-specific enolase from Toxoplasma gondii reveals insights into its dual cytoplasmic and nuclear functions. Acta Crystallogr D Biol Crystallogr 2015, 71, 417–426. [CrossRef]
- Neubig RR, Krodel EK, Boyd ND, Cohen JB: Acetylcholine and local anesthetic binding to Torpedo nicotinic postsynaptic membranes after removal of nonreceptor peptides. Proc Natl Acad Sci U S A 1979, 76, 690–694. [CrossRef]
- Shevade S, Jindal N, Dutta S, Jarori GK: Food vacuole associated enolase in plasmodium undergoes multiple post-translational modifications: evidence for atypical ubiquitination. PLoS One 2013, 8, e72687.
- Arakawa T, Kobayashi-Yurugi T, Alguel Y, Iwanari H, Hatae H, Iwata M, Abe Y, Hino T, Ikeda-Suno C, Kuma H, et al: Crystal structure of the anion exchanger domain of human erythrocyte band 3. Science 2015, 350, 680–684. [CrossRef]
- Fujinaga J, Tang XB, Casey JR: Topology of the membrane domain of human erythrocyte anion exchange protein, AE1. J Biol Chem 1999, 274, 6626–6633. [CrossRef]
- Frank R: The SPOT-synthesis technique. Synthetic peptide arrays on membrane supports--principles and applications. J Immunol Methods 2002, 267, 13–26. [Google Scholar]
- Dutta S, Mukherjee D, Jarori GK: Replacement of Ser108 in Plasmodium falciparum enolase results in weak Mg(II) binding: role of a parasite-specific pentapeptide insert in stabilizing the active conformation of the enzyme. FEBS J 2015, 282, 2296–2308. [CrossRef] [PubMed]
- Read M, Hicks KE, Sims PF, Hyde JE: Molecular characterisation of the enolase gene from the human malaria parasite Plasmodium falciparum. Evidence for ancestry within a photosynthetic lineage. Eur J Biochem 1994, 220, 513–520. [Google Scholar]
- Hamasaki N, Okubo K: Band 3 protein: physiology, function and structure. Cell Mol Biol (Noisy-le-grand) 1996, 42, 1025–1039.
- Yu J, Steck TL: Isolation and characterization of band 3, the predominant polypeptide of the human erythrocyte membrane. J Biol Chem 1975, 250, 9170–9175. [CrossRef]
- Baldwin M, Yamodo I, Ranjan R, Li X, Mines G, Marinkovic M, Hanada T, Oh SS, Chishti AH: Human erythrocyte band 3 functions as a receptor for the sialic acid-independent invasion of Plasmodium falciparum. Role of the RhopH3-MSP1 complex. Biochim Biophys Acta 2014, 1843, 2855–2870. [Google Scholar]
- Bhattacharya S, Ploplis VA, Castellino FJ: Bacterial plasminogen receptors utilize host plasminogen system for effective invasion and dissemination. J Biomed Biotechnol 2012, 2012, 482096.
- Jong AY, Chen SH, Stins MF, Kim KS, Tuan TL, Huang SH: Binding of Candida albicans enolase to plasmin(ogen) results in enhanced invasion of human brain microvascular endothelial cells. J Med Microbiol 2003, 52, 615–622. [CrossRef]
- Liu K-J, Shih N-Y: The Role of Enolae in Tissue Invasion and Metastasis of Pathogens and Tumor Cells. J Cancer Mole 2007, 3, 45–48.
- Lahteenmaki K, Edelman S, Korhonen TK: Bacterial metastasis: the host plasminogen system in bacterial invasion. Trends Microbiol 2005, 13, 79–85. [CrossRef]
- Vinetz JM: Plasmodium ookinete invasion of the mosquito midgut. Curr Top Microbiol Immunol 2005, 295, 357–382.
- Hernandez-Romano J, Rodriguez MH, Pando V, Torres-Monzon JA, Alvarado-Delgado A, Lecona Valera AN, Ramos RA, Martinez-Barnetche J, Rodriguez MC: Conserved peptide sequences bind to actin and enolase on the surface of plasmodium berghei ookinetes. Parasitology 2011, 138, 1341–1353. [CrossRef] [PubMed]
- Mukherjee D, Mishra P, Joshi M, Thakur PK, Hosur RV, Jarori GK: EWGWS insert in Plasmodium falciparum ookinete surface enolase is involved in binding of PWWP containing peptides: Implications to mosquito midgut invasion by the parasite. Insect Biochem Mol Biol 2016, 68, 13–22. [CrossRef] [PubMed]
- Didiasova M, Schaefer L, Wygrecka M: When Place Matters: Shuttling of Enolase-1 Across Cellular Compartments. Front Ceell Dev Biol 2019, 7, 61.
Figure 1.
Binding of Pf enolase (Pfeno) with intact hRBCs: (A) 100 µl packed hRBCs were incubated with various concentrations of recombinant Pfeno (rPfeno). Following washing with PBS, bound rPfeno was eluted with NaCl (2M) and analyzed on a 10% SDS-PAGE and western blots were performed using α-rPfeno antibodies: lane 1: Purified rPfeno alone; lanes 2-4: eluted samples where 5, 10 and 20 µg respectively of rPfeno was incubated with hRBCs. (B) Variation in bound rPfeno as detected by western blot with increasing concentration of rPfeno. (C) Western blots of Yeno with hRBCs., lane 1: pure Yeno (1 µg); lanes 2 and 3: eluted samples where Yeno (5 and 50 µg) was incubated with hRBCs and then probed against α-Yeno. (D) Effect of neuraminidase and protease treatment of hRBC on binding of rPfeno. Western blots of rPfeno eluted samples. Lane 1: hRBC eluate where no rPfeno was added; In remaining samples (lanes 2-8), eluted samples from variously treated 100 µl hRBCs were incubated with 15 µg rPfeno. Lane 2: untreated; lanes 3 and 4: pre-treated with trypsin; lanes 5 and 6: pre-treated with chymotrypsin, lanes 7 and 8: pre-treated with neuraminidase.
Figure 1.
Binding of Pf enolase (Pfeno) with intact hRBCs: (A) 100 µl packed hRBCs were incubated with various concentrations of recombinant Pfeno (rPfeno). Following washing with PBS, bound rPfeno was eluted with NaCl (2M) and analyzed on a 10% SDS-PAGE and western blots were performed using α-rPfeno antibodies: lane 1: Purified rPfeno alone; lanes 2-4: eluted samples where 5, 10 and 20 µg respectively of rPfeno was incubated with hRBCs. (B) Variation in bound rPfeno as detected by western blot with increasing concentration of rPfeno. (C) Western blots of Yeno with hRBCs., lane 1: pure Yeno (1 µg); lanes 2 and 3: eluted samples where Yeno (5 and 50 µg) was incubated with hRBCs and then probed against α-Yeno. (D) Effect of neuraminidase and protease treatment of hRBC on binding of rPfeno. Western blots of rPfeno eluted samples. Lane 1: hRBC eluate where no rPfeno was added; In remaining samples (lanes 2-8), eluted samples from variously treated 100 µl hRBCs were incubated with 15 µg rPfeno. Lane 2: untreated; lanes 3 and 4: pre-treated with trypsin; lanes 5 and 6: pre-treated with chymotrypsin, lanes 7 and 8: pre-treated with neuraminidase.
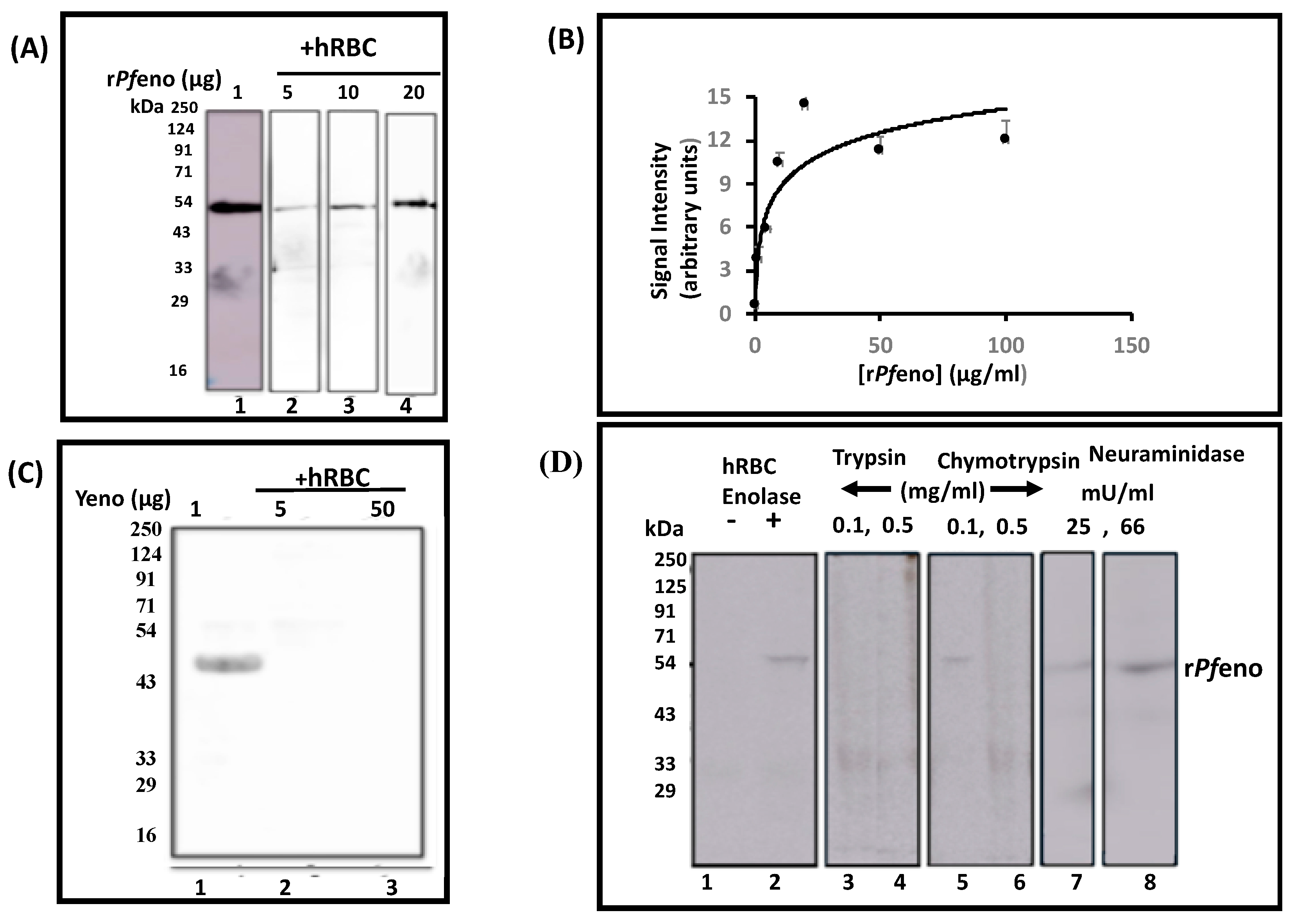
Figure 2.
Specificity of direct binding of rPfeno to hRBC ghosts (hRBCG) proteins. hRBCG (100 μg protein) were used in each sample. hRBCG were incubated with rPfeno (or Yeno), pelleted by centrifugation, washed and analyzed on a 10% SDS-PAGE. (A) Western blot of a gel visualized using α-rPfeno. Lane 1: 2 μg rPfeno (control); lanes 2 and 3: hRBCG incubated with 5 and 25 μg rPfeno respectively. (B) Western blot of Yeno samples visualized using α-Yeno. Lanes 1 and 3: 1 μg and 10 ng Yeno (controls). Lanes 2 and 4: samples where hRBCG were incubated with 5 and 100 μg of Yeno respectively. Absence of Yeno in lanes 2 and 4 indicates failure of Yeno to bind to hRBCG. Arrows mark the position of rPfeno and Yeno protein bands.
Figure 2.
Specificity of direct binding of rPfeno to hRBC ghosts (hRBCG) proteins. hRBCG (100 μg protein) were used in each sample. hRBCG were incubated with rPfeno (or Yeno), pelleted by centrifugation, washed and analyzed on a 10% SDS-PAGE. (A) Western blot of a gel visualized using α-rPfeno. Lane 1: 2 μg rPfeno (control); lanes 2 and 3: hRBCG incubated with 5 and 25 μg rPfeno respectively. (B) Western blot of Yeno samples visualized using α-Yeno. Lanes 1 and 3: 1 μg and 10 ng Yeno (controls). Lanes 2 and 4: samples where hRBCG were incubated with 5 and 100 μg of Yeno respectively. Absence of Yeno in lanes 2 and 4 indicates failure of Yeno to bind to hRBCG. Arrows mark the position of rPfeno and Yeno protein bands.
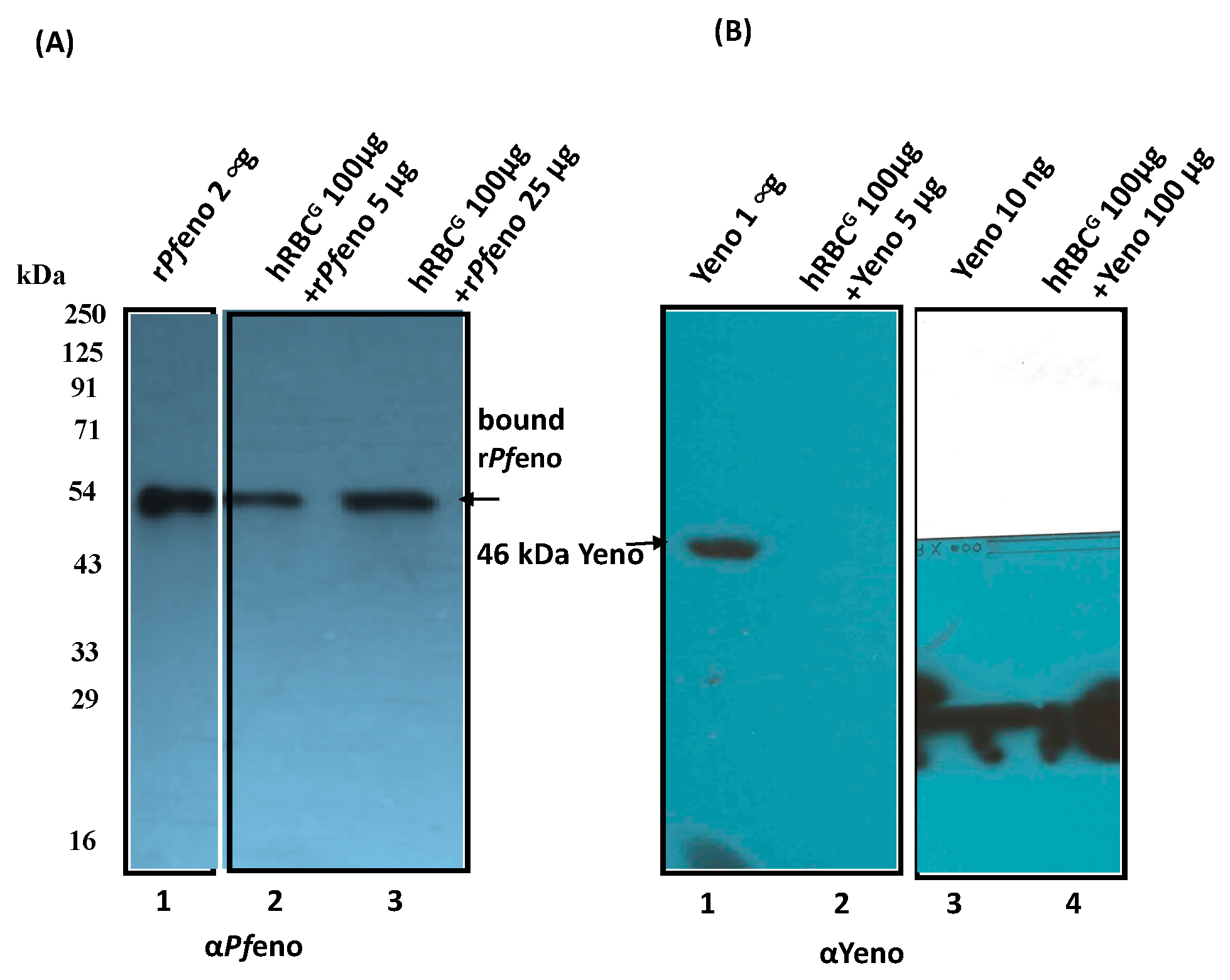
Figure 3.
Blot-overlay assays to identify potential interactor(s) of Pfeno among hRBC membrane proteins. 16 µg hRBCG membrane proteins were separated on 10% SDS-Page. (A) Coomassie stained gel. Lane 1, rPfeno; lane 2, hRBCG membrane proteins. Lanes 3 and 4: Western blot of replicates of lane 1 and 2 probed with α-rPfeno. (B) Blot of hRBCG proteins was incubated with 5 µg/ml (lane 2) rPfeno. After washing, α-rPfeno antibody was used to detect bound rPfeno. (C) Lane 1, 10 ng of Yeno, lane 2, blot of hRBCG proteins was incubated with 50 µg/ml of Yeno and anti-Yeno (α-Yeno) was used for the detection of bound Yeno.
Figure 3.
Blot-overlay assays to identify potential interactor(s) of Pfeno among hRBC membrane proteins. 16 µg hRBCG membrane proteins were separated on 10% SDS-Page. (A) Coomassie stained gel. Lane 1, rPfeno; lane 2, hRBCG membrane proteins. Lanes 3 and 4: Western blot of replicates of lane 1 and 2 probed with α-rPfeno. (B) Blot of hRBCG proteins was incubated with 5 µg/ml (lane 2) rPfeno. After washing, α-rPfeno antibody was used to detect bound rPfeno. (C) Lane 1, 10 ng of Yeno, lane 2, blot of hRBCG proteins was incubated with 50 µg/ml of Yeno and anti-Yeno (α-Yeno) was used for the detection of bound Yeno.

Figure 4.
Pull down assay for capturing rPfeno binding protein(s) in solubilized hRBC ghost. (A) Detergent solubilized hRBCG proteins were incubated with GST-tagged rPfeno immobilized on GSH beads (GSH-B). Post washing, the supernatants of Laemmli buffer treated bead bound protein(s) were analyzed on 10% SDS-PAGE (silver stained). Lane 1: purified GST-tagged rPfeno-GSH-B (GST-rPfeno). Profile of rPfeno bound hRBCG proteins solubilized with CHAPS (lane 2); NP-40 (lane 3) and Triton-X100 (lane 4). Lane 5: GSH-B incubated with hRBCG protein extracts to identify any non-specific interaction with GSH-B. Bands marked with rectangular boxes (A-F) were cut out and proteins identified using mass spectrometry. Note the absence of any bands in lane 5 indicating absence of non-specific interactions with GSH beads. Results of proteins identified are presented in Table 1. (B) Silver stain of hRBCG treated with 0.1 M NaOH for removal of peripheral membrane proteins. The ghosts were then solubilized in Triton-X100, and a pull-down assay was performed. Lane 1: alkali treated hRBCG; lane 2: GSH-B incubated with alkali treated hRBCG (control); lane 3: GSH-B incubated with alkali treated hRBCG extract + GST-tagged rPfeno. (C)Western blot using monoclonal α-band 3 antibody of three lanes from (B). Note the presence of band 3 in hRBCG extract (lane 3) and pull-down of band 3 with rPfeno (lane 1). Lack of band 3 in lane 2 indicates absence of non-specific binding of hRBCG extract proteins with beads. The arrow indicates the position of pull-down band 3. Various protein bands were sliced out from the gel lanes shown in Figure 4A. These were subjected to trypsin digestion and the extracted peptides were analyzed using LC-MS/MS. Major proteins identified are listed in Table 1.
Figure 4.
Pull down assay for capturing rPfeno binding protein(s) in solubilized hRBC ghost. (A) Detergent solubilized hRBCG proteins were incubated with GST-tagged rPfeno immobilized on GSH beads (GSH-B). Post washing, the supernatants of Laemmli buffer treated bead bound protein(s) were analyzed on 10% SDS-PAGE (silver stained). Lane 1: purified GST-tagged rPfeno-GSH-B (GST-rPfeno). Profile of rPfeno bound hRBCG proteins solubilized with CHAPS (lane 2); NP-40 (lane 3) and Triton-X100 (lane 4). Lane 5: GSH-B incubated with hRBCG protein extracts to identify any non-specific interaction with GSH-B. Bands marked with rectangular boxes (A-F) were cut out and proteins identified using mass spectrometry. Note the absence of any bands in lane 5 indicating absence of non-specific interactions with GSH beads. Results of proteins identified are presented in Table 1. (B) Silver stain of hRBCG treated with 0.1 M NaOH for removal of peripheral membrane proteins. The ghosts were then solubilized in Triton-X100, and a pull-down assay was performed. Lane 1: alkali treated hRBCG; lane 2: GSH-B incubated with alkali treated hRBCG (control); lane 3: GSH-B incubated with alkali treated hRBCG extract + GST-tagged rPfeno. (C)Western blot using monoclonal α-band 3 antibody of three lanes from (B). Note the presence of band 3 in hRBCG extract (lane 3) and pull-down of band 3 with rPfeno (lane 1). Lack of band 3 in lane 2 indicates absence of non-specific binding of hRBCG extract proteins with beads. The arrow indicates the position of pull-down band 3. Various protein bands were sliced out from the gel lanes shown in Figure 4A. These were subjected to trypsin digestion and the extracted peptides were analyzed using LC-MS/MS. Major proteins identified are listed in Table 1.
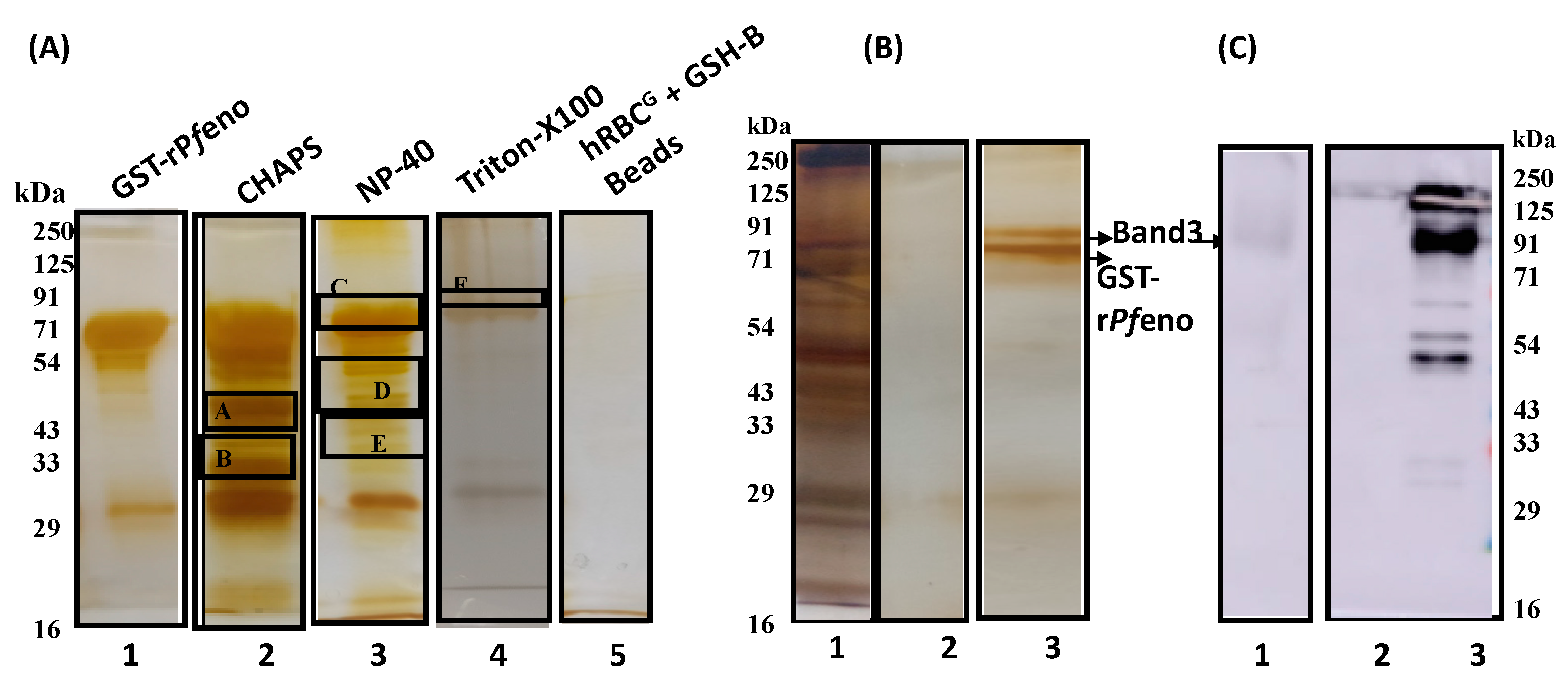
Figure 5.
Co-immuno-precipitation of rPfeno and band 3 from solubilized extract of hRBC ghost (hRBCG) incubated with rPfeno. (A) hRBCG were solubilized in detergent and the extracted proteins were incubated with rPfeno allowing the formation of rPfeno-interactor complex. For co-precipitation, α-rPfeno or α-band 3 was first immobilized on PrA forming PrA-B- α-rPfeno or PrA-B-α-band 3. Two sets of beads were incubated in two separate samples of hRBCG + rPfeno. (i): Coomassie stained gel; lane 1, detergent extracted hRBCG proteins; lane 2, PrA beads incubated with hRBCG extract (control); lane 3, PrA-B-α-rPfeno incubated with hRBCG and rPfeno sample. Proteins bound to the beads were directly eluted in gel loading buffer. (ii) Western blot of gel in (i) probed with α-band 3. Lanes 1 and 3 are positive for band 3. (B) Pull-down of band 3 protein from solubilized extract of hRBCG and rPfeno sample. PrA-α-band 3 was used to pull down band 3 along with the associated proteins. The beads were separated from hRBCG and rPfeno sample, washed and bound proteins were analysed. (i) Coomassie stained gel and (ii) Western blot was probed with α-Pfeno. Lanes 1and 2 did not show any reactivity to α-Pfeno while lane 3 was positive (rPfeno band marked with *) indicating association with hRBCG band 3. (C) (i) Coomassie stained rPfeno (lane 1) and PrA-B-α-band 3 pull down band 3 from hRBCG extract. (lane 2) (ii) Lanes 1 and 3 are a blot of lanes 1 and 2 from Figure 5C (i); lane 3 is a pull down of band 3 from the second independent sample of hRBCG. All three lanes were overlaid with rPfeno and probed with α-Pfeno. Lanes 2 and 3 reflect the observed variability in band 3 pattern in different hRBCG samples.
Figure 5.
Co-immuno-precipitation of rPfeno and band 3 from solubilized extract of hRBC ghost (hRBCG) incubated with rPfeno. (A) hRBCG were solubilized in detergent and the extracted proteins were incubated with rPfeno allowing the formation of rPfeno-interactor complex. For co-precipitation, α-rPfeno or α-band 3 was first immobilized on PrA forming PrA-B- α-rPfeno or PrA-B-α-band 3. Two sets of beads were incubated in two separate samples of hRBCG + rPfeno. (i): Coomassie stained gel; lane 1, detergent extracted hRBCG proteins; lane 2, PrA beads incubated with hRBCG extract (control); lane 3, PrA-B-α-rPfeno incubated with hRBCG and rPfeno sample. Proteins bound to the beads were directly eluted in gel loading buffer. (ii) Western blot of gel in (i) probed with α-band 3. Lanes 1 and 3 are positive for band 3. (B) Pull-down of band 3 protein from solubilized extract of hRBCG and rPfeno sample. PrA-α-band 3 was used to pull down band 3 along with the associated proteins. The beads were separated from hRBCG and rPfeno sample, washed and bound proteins were analysed. (i) Coomassie stained gel and (ii) Western blot was probed with α-Pfeno. Lanes 1and 2 did not show any reactivity to α-Pfeno while lane 3 was positive (rPfeno band marked with *) indicating association with hRBCG band 3. (C) (i) Coomassie stained rPfeno (lane 1) and PrA-B-α-band 3 pull down band 3 from hRBCG extract. (lane 2) (ii) Lanes 1 and 3 are a blot of lanes 1 and 2 from Figure 5C (i); lane 3 is a pull down of band 3 from the second independent sample of hRBCG. All three lanes were overlaid with rPfeno and probed with α-Pfeno. Lanes 2 and 3 reflect the observed variability in band 3 pattern in different hRBCG samples.
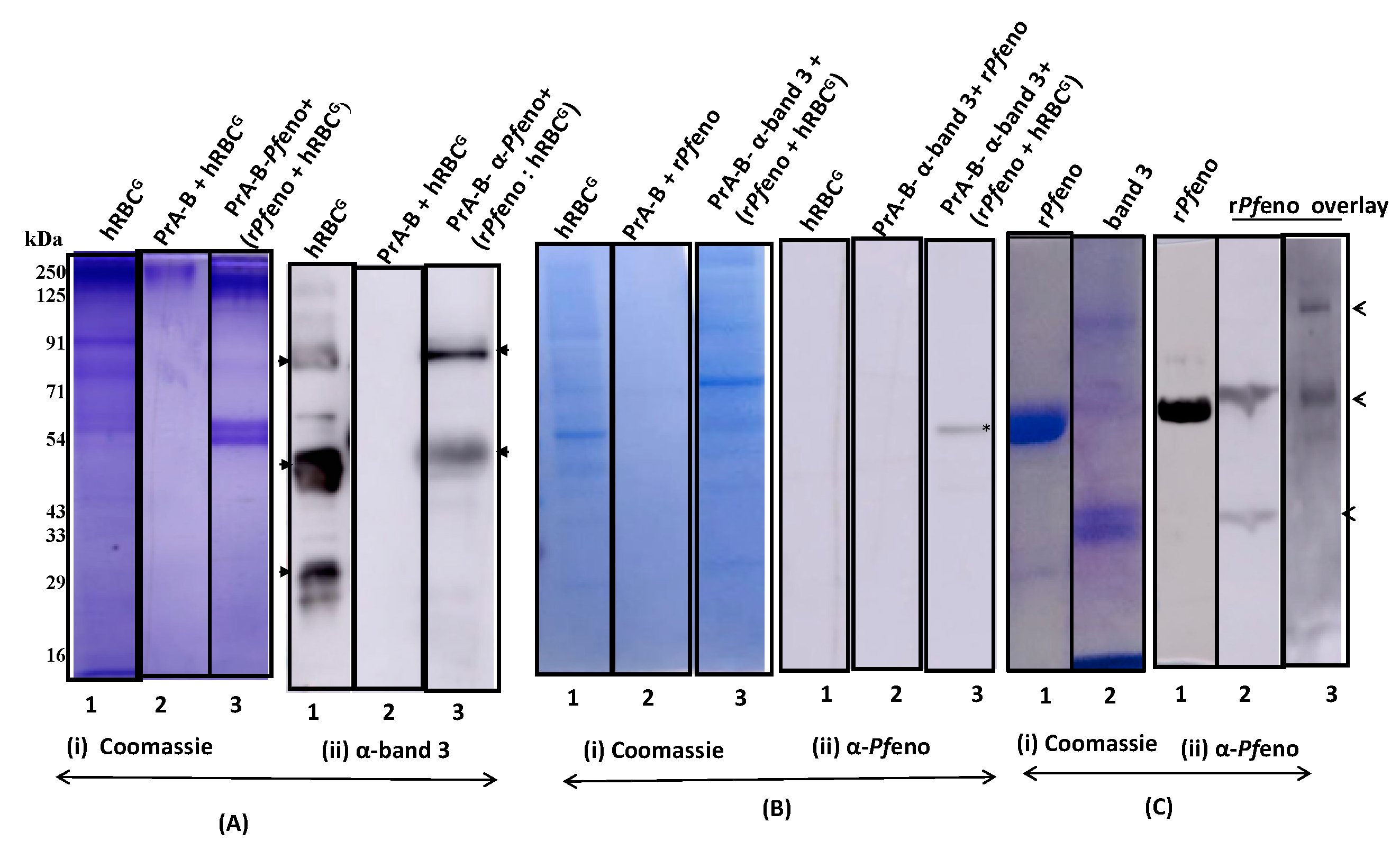
Figure 6.
Binding of native Plasmodium enolase to band 3 protein. (A) Blot-overlay assay: Lanes 1 and 2 contain 1 µg rPfeno and 16 ug hRBC ghost preparation (hRBCG) run on a 10% SDS-PAGE, the corresponding blot overlaid with supernatant from P. falciparum culture, and then probed with α-Pfeno antibodies. Arrows mark the position of hRBCG proteins which interact with native P. falciparum enolase. Lane 3: Western blot of a pull-down assay using Protein A beads coupled with α-band 3 antibodies incubated with P. falciparum infected RBC (iRBC), probed with α-band 3 antibodies. (B) Pull down experiment using P. falciparum and P. berghei iRBC. Protein A beads were bound to α-band 3 antibodies and then incubated with P. falciparum (Lane 3) and P. berghei (Lane 4) iRBC, washed and run on 10% SDS-PAGE and then probed with α-rPfeno antibodies. Lane 5 shows the control of the pull-down experiments using uninfected RBC (uRBC) extract. Lanes 1 and 2 contain molecular weight markers and 1 µg rPfeno protein, respectively. (C) Lanes 1 and 2 show protein profiles of 20 g P. berghei merozoite extract (M) and 16 µg of solubilized hRBCG (G), run on 10% SDS-PAGE and stained with Coomassie Blue. Blots of the same samples were probed with α-MSP-119 (Lanes 3 and 4), α-band 3 (Lanes 5 and 6) and α-Pfeno (Lanes 7 and 8) antibodies, respectively. Arrows mark the position of each protein (and variants arising due to proteolysis). α-MSP-119 and α-Pfeno antibodies were specific and exhibited no cross reactivity with hRBCG proteins (Lanes 4 and 8). Similarly, α-band 3 antibodies showed no cross reactivity with the P. berghei merozoite extract (Lane 5). (D) Pbeno protein was pulled down from P. berghei merozoite extract by incubation with Protein A beads (PrA-B) bound to α-Pfeno antibodies. The beads (PrA-B-α-Pfeno-Pbeno) were then incubated with solubilized hRBCG extract. Presence of band 3 protein in this pull down was detected in a Western blot probed with α-band 3 antibodies (Lane 2). Reciprocally, PrA-B-α-band 3 was used for pulling down band 3 protein from hRBCG extract, and the resultant PrA-B-α-band 3-band 3 complex was then incubated with P. berghei merozoite extract. The Western blot analysis of PrA-B-α-band3-band 3 interactor proteins, using α-Pfeno antibodies, showed the presence of enolase (Lane 4). The possibility of non-specific binding of band 3 or Pbeno proteins to PrA-B was ruled out by incubating PrA+ merozoite extract + hRBCG extract and probing with α-band 3 antibodies (Lane 1); PrA-B-αPfeno + hRBCG ghost extract and probing with α-band 3 antibodies (Lane 3) and PrA + merozoite extract and probing with α-Pfeno antibodies (Lane 5). These controls showed an absence of nonspecific adsorption of band 3 or Pbeno on PrA beads under the experimental conditions used here.
Figure 6.
Binding of native Plasmodium enolase to band 3 protein. (A) Blot-overlay assay: Lanes 1 and 2 contain 1 µg rPfeno and 16 ug hRBC ghost preparation (hRBCG) run on a 10% SDS-PAGE, the corresponding blot overlaid with supernatant from P. falciparum culture, and then probed with α-Pfeno antibodies. Arrows mark the position of hRBCG proteins which interact with native P. falciparum enolase. Lane 3: Western blot of a pull-down assay using Protein A beads coupled with α-band 3 antibodies incubated with P. falciparum infected RBC (iRBC), probed with α-band 3 antibodies. (B) Pull down experiment using P. falciparum and P. berghei iRBC. Protein A beads were bound to α-band 3 antibodies and then incubated with P. falciparum (Lane 3) and P. berghei (Lane 4) iRBC, washed and run on 10% SDS-PAGE and then probed with α-rPfeno antibodies. Lane 5 shows the control of the pull-down experiments using uninfected RBC (uRBC) extract. Lanes 1 and 2 contain molecular weight markers and 1 µg rPfeno protein, respectively. (C) Lanes 1 and 2 show protein profiles of 20 g P. berghei merozoite extract (M) and 16 µg of solubilized hRBCG (G), run on 10% SDS-PAGE and stained with Coomassie Blue. Blots of the same samples were probed with α-MSP-119 (Lanes 3 and 4), α-band 3 (Lanes 5 and 6) and α-Pfeno (Lanes 7 and 8) antibodies, respectively. Arrows mark the position of each protein (and variants arising due to proteolysis). α-MSP-119 and α-Pfeno antibodies were specific and exhibited no cross reactivity with hRBCG proteins (Lanes 4 and 8). Similarly, α-band 3 antibodies showed no cross reactivity with the P. berghei merozoite extract (Lane 5). (D) Pbeno protein was pulled down from P. berghei merozoite extract by incubation with Protein A beads (PrA-B) bound to α-Pfeno antibodies. The beads (PrA-B-α-Pfeno-Pbeno) were then incubated with solubilized hRBCG extract. Presence of band 3 protein in this pull down was detected in a Western blot probed with α-band 3 antibodies (Lane 2). Reciprocally, PrA-B-α-band 3 was used for pulling down band 3 protein from hRBCG extract, and the resultant PrA-B-α-band 3-band 3 complex was then incubated with P. berghei merozoite extract. The Western blot analysis of PrA-B-α-band3-band 3 interactor proteins, using α-Pfeno antibodies, showed the presence of enolase (Lane 4). The possibility of non-specific binding of band 3 or Pbeno proteins to PrA-B was ruled out by incubating PrA+ merozoite extract + hRBCG extract and probing with α-band 3 antibodies (Lane 1); PrA-B-αPfeno + hRBCG ghost extract and probing with α-band 3 antibodies (Lane 3) and PrA + merozoite extract and probing with α-Pfeno antibodies (Lane 5). These controls showed an absence of nonspecific adsorption of band 3 or Pbeno on PrA beads under the experimental conditions used here.
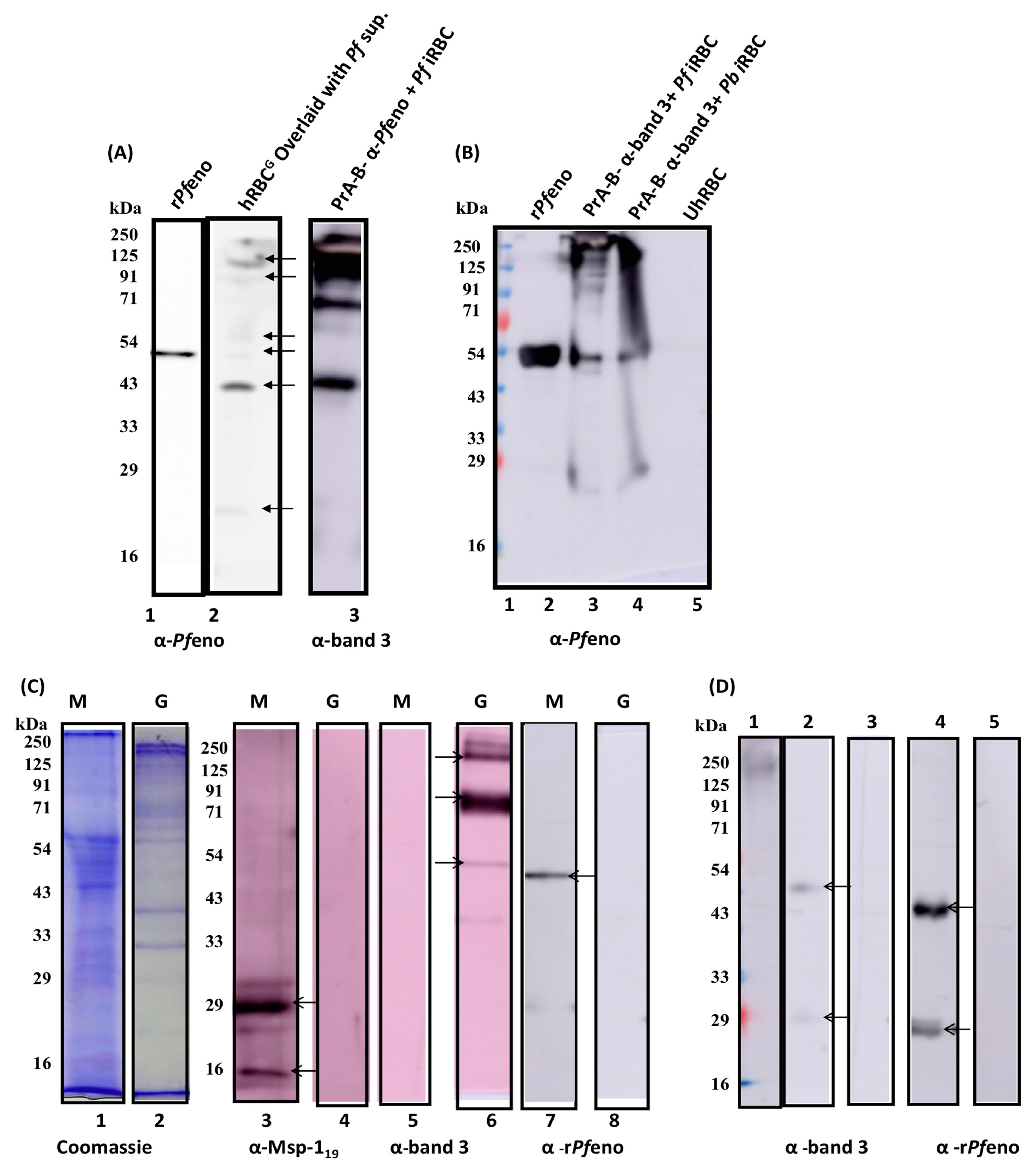
Figure 7.
Two-dimensional gel analysis of hRBC ghost proteome and identification of Pfeno binding protein(s). (A) Silver-stained gel of hRBCG. (B) Western blot of 2DE probed with α-band 3 antibodies. Proteins in boxes at near around 25 kDa and 64 kDa represent band 3 and its variants. (C) A 2DE blot of hRBCG proteome was overlaid with rPfeno protein. After removal of unbound rPfeno, blot was probed with α-rPfeno antibody. Binding of rPfeno to band 3 (rectangles) is quite evident and was marked in silver stain gel of hRBC (Panel A) to excise the bands for Mass spectrometry.
Figure 7.
Two-dimensional gel analysis of hRBC ghost proteome and identification of Pfeno binding protein(s). (A) Silver-stained gel of hRBCG. (B) Western blot of 2DE probed with α-band 3 antibodies. Proteins in boxes at near around 25 kDa and 64 kDa represent band 3 and its variants. (C) A 2DE blot of hRBCG proteome was overlaid with rPfeno protein. After removal of unbound rPfeno, blot was probed with α-rPfeno antibody. Binding of rPfeno to band 3 (rectangles) is quite evident and was marked in silver stain gel of hRBC (Panel A) to excise the bands for Mass spectrometry.

Figure 8.
(A) Amino acid sequence of band 3 Erythrocyte anion exchanger protein (NCBI Reference Sequence: NP_000333.1). Peptide sequences obtained from mass spectrometric analysis of band 3 in various pull-down assays (Figure 4, Table 3 and Table 4) are shown in red color. Regions of the protein that are exocytic in hRBCG band 3 as defined by Fuginaga et al. [36] are in blue italics. (B) ELISA for the binding of certain band 3 synthetic peptides (Peptide sequences obtained from mass spectrometric analysis of the band 3 in various pull-down assays as shown in Figure 4 and Table 3 and Table 4) to rPfeno. Plates were coated with different band 3 peptides in triplicates and rPfeno was added to each well. After washing, the bound rPfeno was estimated using αPfeno based assay. The mean ± SD value of absorbance from three experiments is shown. The sequences and locations of various peptides (also shown in Table 4) that bind to Pfeno have been underlined in panel A.
Figure 8.
(A) Amino acid sequence of band 3 Erythrocyte anion exchanger protein (NCBI Reference Sequence: NP_000333.1). Peptide sequences obtained from mass spectrometric analysis of band 3 in various pull-down assays (Figure 4, Table 3 and Table 4) are shown in red color. Regions of the protein that are exocytic in hRBCG band 3 as defined by Fuginaga et al. [36] are in blue italics. (B) ELISA for the binding of certain band 3 synthetic peptides (Peptide sequences obtained from mass spectrometric analysis of the band 3 in various pull-down assays as shown in Figure 4 and Table 3 and Table 4) to rPfeno. Plates were coated with different band 3 peptides in triplicates and rPfeno was added to each well. After washing, the bound rPfeno was estimated using αPfeno based assay. The mean ± SD value of absorbance from three experiments is shown. The sequences and locations of various peptides (also shown in Table 4) that bind to Pfeno have been underlined in panel A.

Figure 9.
Mapping of the binding domain in P. falciparum enolase for hRBCghost band 3 protein. (A) A library of biotin labelled fifteen-mer peptides covering full sequence of Pfeno was immobilized on a streptavidin coated ELISA plate. This was incubated with solubilized hRBCG proteins. Binding of band 3 to Pfeno peptides was determined using a-band 3 in standard ELISA. Peptide numbers 16, 40, 49, 50 and 57-64 (Highlighted yellow) showed affinity for band 3. The data shown are mean + SD for triplicate experiments. Single peptide showing binding in a given region (peptide no. 16 and 40) were considered nonspecific, as the two neighboring peptides having overlap of 10 residues failed to bind band 3. (B) Sequences of the Pfeno peptides involved in binding to band 3 are shown in black. Sequences of neighboring peptides that did not show any interaction with band 3 are shown in red. Comparison of the overlapping regions of binding and non-binding peptides suggested that a sequence of ten amino acid residues (shown in green) was essential for binding to band 3. (C) Modelled 3D structure showing the locations of the unique EWGWS insert (pink), the two band 3 binding regions (green & blue) and the Plasminogen binding domain (red) in Pfenolase.
Figure 9.
Mapping of the binding domain in P. falciparum enolase for hRBCghost band 3 protein. (A) A library of biotin labelled fifteen-mer peptides covering full sequence of Pfeno was immobilized on a streptavidin coated ELISA plate. This was incubated with solubilized hRBCG proteins. Binding of band 3 to Pfeno peptides was determined using a-band 3 in standard ELISA. Peptide numbers 16, 40, 49, 50 and 57-64 (Highlighted yellow) showed affinity for band 3. The data shown are mean + SD for triplicate experiments. Single peptide showing binding in a given region (peptide no. 16 and 40) were considered nonspecific, as the two neighboring peptides having overlap of 10 residues failed to bind band 3. (B) Sequences of the Pfeno peptides involved in binding to band 3 are shown in black. Sequences of neighboring peptides that did not show any interaction with band 3 are shown in red. Comparison of the overlapping regions of binding and non-binding peptides suggested that a sequence of ten amino acid residues (shown in green) was essential for binding to band 3. (C) Modelled 3D structure showing the locations of the unique EWGWS insert (pink), the two band 3 binding regions (green & blue) and the Plasminogen binding domain (red) in Pfenolase.
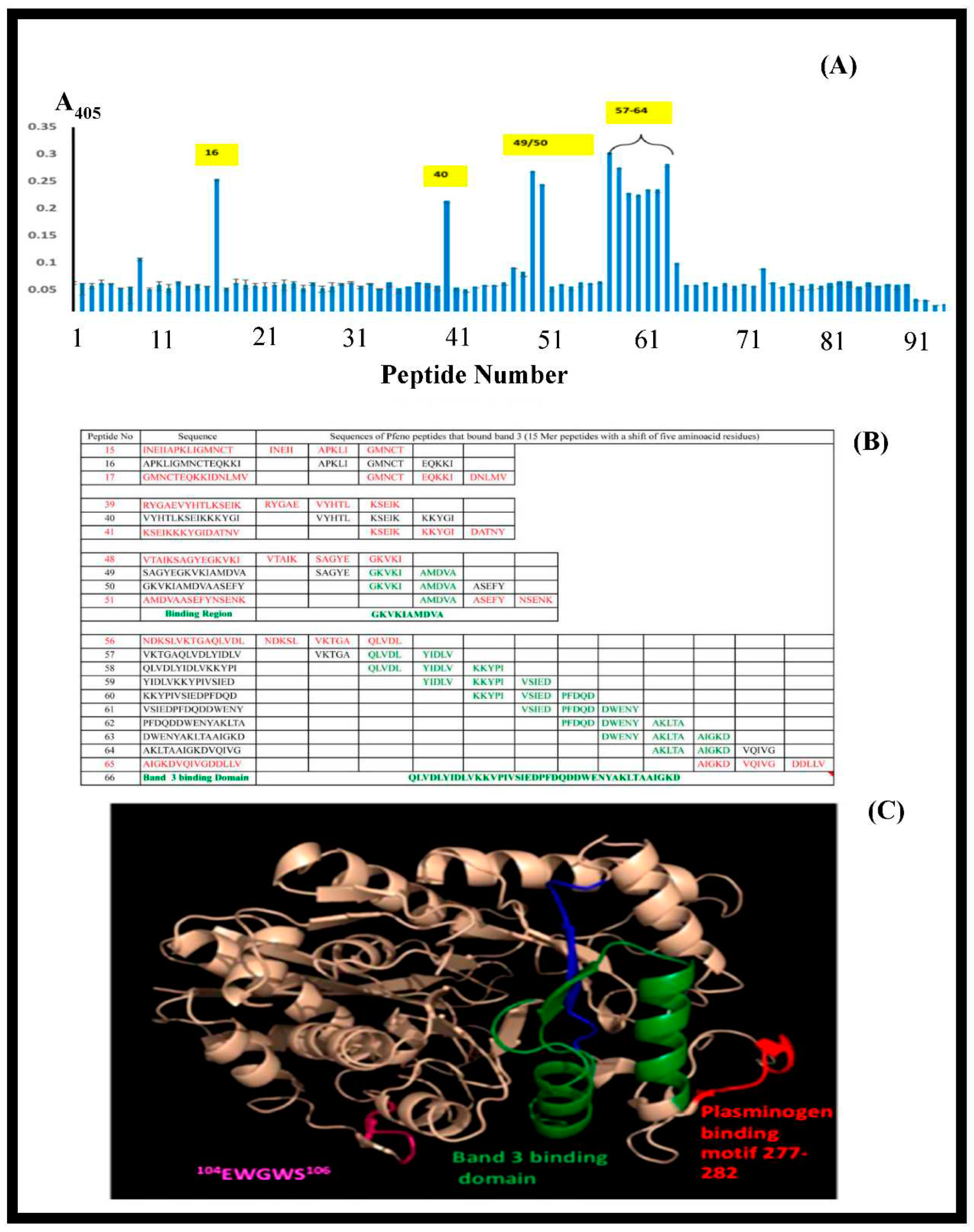

Table 1.
Mass spectrometric analysis of rPfeno interactor proteins in different detergent solubilized hRBCG. Various protein bands were sliced out from the gel lanes shown in Figure 4A. These were subjected to trypsin digestion and the extracted peptides were analysed using LC-MS/MS. Major proteins identified are listed below.
Table 1.
Mass spectrometric analysis of rPfeno interactor proteins in different detergent solubilized hRBCG. Various protein bands were sliced out from the gel lanes shown in Figure 4A. These were subjected to trypsin digestion and the extracted peptides were analysed using LC-MS/MS. Major proteins identified are listed below.
| S.No. | Detergents | Protein Bands | Proteins Identified | Protein Score |
|---|---|---|---|---|
| 1 | CHAPS | A | Spectrin | 49 |
| B | Band 3 | 81 | ||
| 2 | NP-40 | C | Spectrin | 51 |
| D | Band 3 | 71 | ||
| E | Band 3 | 83 | ||
| 3 | Triton X100 |
F | Band 3 | 89 |
Table 2.
Identification of proteins in the rectangles in panel A by mass spectrometry. Two spots were cut out from the silver-stained gel, trypsin digested and analyzed by mass spectrometry. Proteins identified are listed.
Table 2.
Identification of proteins in the rectangles in panel A by mass spectrometry. Two spots were cut out from the silver-stained gel, trypsin digested and analyzed by mass spectrometry. Proteins identified are listed.
| Protein Spot | Proteins Identified | Protein Score | Mol.Wt (kDa) |
|---|---|---|---|
| A (pI~ 5.31 MW~25kDa) |
Diacylglycerol Kinase | 42 | 102 |
| Chromosome 20 ORF | 30 | 125 | |
| Cadherin 12 | 30 | 110 | |
| B (pI~ 5.18, MW~64 kDa) |
Spectrin α chain | 50 | 281 |
| Spectrin β chain | 31 | 247 | |
| Band 3 chain P | 33 | 102 |
Table 3.
Amino acid sequences of band 3 peptides obtained in Mass spectrometric analysis of tryptic digests of protein spots. These sequences . cover ~15% of band 3.
Table 3.
Amino acid sequences of band 3 peptides obtained in Mass spectrometric analysis of tryptic digests of protein spots. These sequences . cover ~15% of band 3.
| S. No. | Peptide positions and Sequences* |
|---|---|
| 1 | G117TVLLDLQETSLAGVANQLLDR138 |
| 2 | F139IFEDQIRPQDR150 |
| 3 | I220PPDSEATVLVGR233 |
| 4 | A234DFLEQPVLGFVR246 |
| 5 | L247QEAAELEAVELPVPIR263 |
| 6 | F264LFVLLGPEAPHIDYTQLGR283 |
| 7 | A284AATLMTER292 |
| 8 | S731VTHANALTVMGKASTPGAAAQ752 |
| 9 | S851TPALSLALPFVLILTVPLR870 |
| 10 | V872LLPLIFR879 |
Table 4.
Amino acid sequence of custom synthesized band 3 peptides. Some of these peptides cover the sequences detected in Mass-spectrometry analysis of tryptic digests obtained from rPfeno binding protein samples.
Table 4.
Amino acid sequence of custom synthesized band 3 peptides. Some of these peptides cover the sequences detected in Mass-spectrometry analysis of tryptic digests obtained from rPfeno binding protein samples.
| S. No. | Amino acid sequence of band 3 peptides |
|---|---|
| 1 | I220PPDSEATLVLVGRADFLEQPVLGFVR246 |
| 2 | L247QEAAELEAVELPVPIRFLFVLLGPEAPHIDYTQLGR283 |
| 3 | L632SVPDGPKVSNSSARGWVIHPLGL655 |
| 4 | S731VTHANALTVMGKASTPGAAAQ753 |
| 5 | A744STPGAAAQIQEVKE758 |
| 6 | F813KPPKYHPDVPYVK826 |
| 7 | S852TPASLALPFVLILTVPLRRVLLPLIFRN879 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



