
Submitted:
16 May 2023
Posted:
17 May 2023
You are already at the latest version
Abstract
We survey the diversity of antimicrobial resistance profiles of Escherichia coli (E. coli) isolates and molecular genotypes by means of phylogenetic groups circulating in complex aquatic habitats, such as sewage and receiving waters bodies as well as in clinical environment in Boeotia regional district of Greece. The predominant resistant profile among all environmental and clinical isolates was to penicillins - ampicillin (AMP) and piperacillin (PIP). ESBLs resistance related profiles were observed both in environmental and clinical isolates. Phylogenetic group B2 was the predominant in clinical isolates, whilst group A was dominant in environmental isolates. We report that reclaimed river water and wastewater are reservoirs of R and MDR E. coli to commonly used antibiotics in clinical practice posing putative threats for human and animal health.
Keywords:
E. coli
; environment
; antibiotic resistance
; antibiotic resistance genes
1. Introduction
The emergence of antimicrobial resistance (AR) to nearly all clinically relevant antibiotics is a pressing health risk that could reverse a century of medical progress. AR reduces the effectiveness of antimicrobials, leading to higher morbidity and mortality rates [1,2]. Antibiotic resistant bacteria (ARB) are not confined to clinical settings, but are also spread through various routes in the ecosystem [3,4,5,6,7]. This is primarily triggered by the selective pressure of antimicrobial use in human and veterinary medicine, agriculture, and aquaculture [8,9,10]. Significant amounts of antimicrobial residues are released into the environment through various routes, including discharges from wastewater treatment plants (WWTP), disturbing the balance between sensitive and resistant bacteria populations [11,12,13,14].
WWTPs receive vast quantities of municipal and industrial waste daily, including hospital wastewater (HWW) that contains ARB and antibiotic resistance genes (ARGs) [1,14,15,16]. ARB and ARGs can evade treatment, and WWTP effluents a) provide optimal conditions for ARB proliferation and horizontal transfer of ARGs, b) are often discharged into water bodies such as rivers, seas, and lakes, and c) are reclaimed for industrial or irrigation purposes, in many countries, thus contributing to the further spread of AR in the ecosystem [3,17,18]. Systematic monitoring of wastewater provides valuable information regarding the presence and release of ARB into the environment, which is crucial for the safe reuse of treated wastewater [2,7]. Moreover, wastewater surveillance reveals the ARB and ARG that are spreading in the community.
While resistance of E. coli to last-line antibiotics that are widely used in clinical practice, livestock farming, and aquaculture is well-studied in the hospital environment, information about community and environmental settings remains limited. E. coli can cause severe infections both in humans and animals, but also is a member of the autochthonous microbiota. E. coli also represents a major reservoir of resistance genes, which can lead to treatment failures in human medicine. A number of resistance genes has been identified in E. coli, and many of these genes can be horizontally transferred. Furthermore, E. coli can act as a donor and as a recipient of resistance genes. The transmission of virulent and resistant E. coli strains between aquatic environments and humans is a major concern, and can occur through various pathways such as direct contact or via the food chain. Thus, circulation, phylogenies, and dispersal of antibiotic-resistant E. coli isolates in environmental habitats are becoming increasingly important to be studied and monitored [3,4,6,19].
To our knowledge the prevalence and diversity of ARGs in environmental E. coli isolates in Greece is still limited. Therefore, we aimed to assess AR patterns and detect ARGs related to resistant phenotypes, identify molecular genotypes, and compare resistance patterns and genotypes between clinical and environmental E. coli isolates from Livadeia area (Greece). The study also aimed to detect common phenotypes and clones among the studied settings, clinical and environmental, in Greece.
2. Materials and Methods
2.1. Sampling Locations and Collected Samples
Sampling locations and collection procedures were carefully planned to capture the spread of environmental AR in the Livadeia city (the capital of the regional unit) of Boeotia regional district, Greece (Figure 1a). This region was chosen due to its intensive agricultural and farming activities, and it is crossed by two rivers: the Erkyna river in the northern side of the city and the Boeotian river in the southeast side of the city. The area also hosts a WWTP and a general prefectural hospital that performs a semi-treatment on the HWW. The hospital provides a wide range of services to approximately 60,000 people annually, including emergency and outpatient care, and has clinics for nephrology, pathology, cardiology, surgery, orthopedics, and obstetrics- gynecology. After preliminary sedimentation, the hospital sewage is discharged into the regional WWTP.
The WWTP receives urban and HWW, with an average daily volume of 5500 m3/day at the entrance and an average hourly flow of 400m3/h. It performs primary treatment, including screening, grit collector, grease trap, oxidation ditch, and primary sedimentation, as well as biological treatment that includes nitrogen and phosphorus removal, secondary sedimentation, chlorination, sludge thickening, and dewatering [15]. The secondary treated effluents are disposed of in the Erkyna river, and are used for restricted irrigation of cropland during the irrigation season. The Erkyna river is directly influenced by the WWTP discharges and indirectly by the hospital sewage. The Erkyna river flows into the Boeotian Kifissos river at a point approximately 6 km away from the WWTP. Both rivers are used for irrigation purposes, with one irrigation project covering 16,000 acres of the studied area [20].
During the period of Summer 2019 to Spring 2021, six sequencial sampling events were conducted in Livadeia city, Boeotia regional district. A total of four samples per sampling period were collected, including a) semi-treated HWW from a septic tank outside the hospital, b) wastewater at the outlet of the regional WWTP, c) river water samples from the Erkyna river adjacent to the WWTP (RWS1), and d) river water samples from the Boeotian Kifissos river at the junction with the Erkyna river (RWS2) located 6 km downstream from the WWTP (Figure 1b). A total of 12 river water samples (six from RWS1 and six from RWS2), six wastewater samples, and six HWW samples were collected and analyzed. All of the samples were collected in sterile dark bottles (500ml volume), were placed on ice and analyzed within 12 hours post-collection. In addition, clinical isolates were collected from clinical specimens such as urine, blood and tissue from the microbiological laboratory of the hospital during the whole study period.
2.2. E. coli Isolation and Identification
E. coli isolation and identification were conducted using a standard membrane filtration technique [ISO 9308.01- 1: 2017] for all river and wastewater samples. The procedure involved filtering multiple volumes (river water: 100 ml, 10 ml, 1 ml, wastewater: 10 ml, 1 ml, 0.1 ml) of each sample through a mixed cellulose ester membrane with a diameter of 47 mm and pore size of 0.45 μm (Whatman® ME 25/21 ST). The membrane filters were then placed on Chromogenic Coliform medium (CHROMagarTM CCA, EF342, Paris, France) with and without antibiotic (CCA with 100 μg/ml ampicillin, CCA/AMP). In both culture media with and without AMP, all colonies showing positive β-D-galactosidase and β-D-glucuronidase reactions (dark blue to violet) were counted as E. coli. The CCA/AMP was used for the estimation and collection of the β-lactam- resistant isolates while CCA without AMP was used for the enumeration and isolation for all E. coli isolates (e.g sensitive and resistant to all antibiotics). Final confirmation of identification of all isolates was done through the indole biochemical test and molecular identification targeting the housekeeping β-D-glucuronidase gene uidA [21].
As for the clinical strains, they were obtained from biological fluids of hospitalized or emergency room patients, such as blood, urine, and tissue, and were identified as E. coli at the microbiological laboratory of the hospital. Specifically, the clinical samples were cultivated on Blood (Blood Agar Base CE, NCM2014, Neogen® Lansing, Michigan, USA) and MacConkey agar (MacConkey Agar No. 3 CE, NCM2018, Neogen®, Lansing, Michigan, USA) at 37°C for 24h. Following this, the isolates were identified by Micro Scan automated system according to standard biochemical tests. The isolates were stored in cryovials with Brain Heart Infusion (Scharlau Microbiology 02-599) +20% glycerol solution and transported to the Molecular Microbiology and Immunology laboratory with proper packaging and transfer conditions [22].
2.3. Antibiotic Susceptibility Testing
All isolates (environmental and clinical) were tested for their antimicrobial susceptibility by disk diffusion assays (Kirby–Bauer method) in 18 antibiotics, commonly used in clinical practice, distributed to 9 different categories: penicillins [ampicillin (AMP; 10μg), piperacillin (PIP; 30μg)], penicillin/inhibitor combinations: [amoxicillin/clavulanic acid (AMC; 20μg /10μg), piperacillin/ tazobactam (TZP; 30μg/6μg)], cephalosporins [ceftriaxone (CRO; 30μg), cefuroxime (CXM; 30μg), ceftazidime (CAZ; 10μg), cefotaxime (CTX; 5 μg), cefepime (FEP; 30μg)], cephamycins: [cefoxitin (FOX; 30μg)], monobactams [aztreonam (ATM; 30μg)], carbapenems [imipenem (IMP; 10μg), meropenem (MEM; 10μg)], aminoglycosides [amikacin (AN; 30μg), gentamicin (GM; 10μg)], quinolones [ciprofloxacin (CIP; 5μg), nalidixic acid (NAL; 30μg)] and miscellaneous agents [sulfamethoxazole-trimethoprim (SXT; 23.75μg/1.25μg)]. The interpretation of the susceptibility results for environmental and clinical isolates was performed according to EUCAST ECOFFs and clinical breakpoints criteria, respectively [23]. All isolates were characterized as sensitive/wild type (S/WT: susceptible to all antibiotics), as non-wild type (N-WT: resistant to only one antibiotic factor), as resistant (R: resistant to more than one antimicrobial agent, maximum of three different categories) and as multi-drug resistant (MDR: resistant to at least one antimicrobial agent in more than three categories). [24,25]. ESBL production was detected phenotypically by clavulanic acid synergy test [double-disk synergy test, DDST; synergy between amoxicillin/ clavulanic acid (AMC) and ceftazidime (CAZ) or cefotaxime (CTX)] [26]. The phenotypic test, carbapenem inactivation method (CIM), was implemented in isolates which exhibited decreased susceptibility to carbapenems (meropenem, imipenem) in order to detect carbapenemase production such as KPC, NDM, OXA-48, VIM, IMP and OXA-23 [26,27].
2.4. Isolation of Genomic DNA
E. coli genomic was extracted using either the boil-freezing method or the PurelinkΤΜ Genomic DNA mini kit (Invitrogen), following the manufacturer's instructions, after 24 hours of bacterial growth on nutrient agar.
2.5. PCR Amplification of Resistance Genes
All DDST-positive isolates underwent PCR to detect three different types of ESBL genes: blaTEM, blaSHV, blaCTX-M [28,29]. CIM-positive isolates were tested for the presence of carbapenemase genes (blaKPC, blaVIM, blaNDM, blaIMP, blaOXA-48, blaOXA-23) [28,30]. Isolates resistant to penicillin/inhibitor combinations and cephamycins were tested for AmpC type β-lactamases genes (blaCMY and blaFOX) [26,31], while MDR isolates exhibiting resistance to SXT were screened for the dihydropteroate synthase gene (sul1) conferring resistance to sulphonamides [32]. PCR amplicons were subjected to Sanger sequence analysis (CeMIA SA, http://cemia.eu/sangersequencing.html), as previously described [25,33]. The sequences and chromatographs were interpreted using MEGA software (https://www.megasoftware.net/), and BLAST algorithm (https://blast.ncbi.nlm.nih.gov/Blast.cgi) was used to identify antimicrobial resistance genes. DNA sequences were compared with reference antibiotic resistance genes from NCBI (https://www.ncbi.nlm.nih.gov/pathogens/refgene) and phylogenetic trees were constructed by Maximum Likelihood method to investigate any possible correlations.
2.6. Molecular Typing
2.6.1. Phylogrouping
The Triplex PCR phylogrouping method utilizes the detection of chuA and yjaA genes and DNA fragment TSPE4.C2 to classify E. coli isolates into four phylogenetic groups, A, B1, B2, and D, as per Clermont’s schema [34]. This method was employed to investigate the correlation between the origin of the sample (clinical specimens, HWW, WWTP effluents, RWS1, RWS2) and phylogenetic groups and to assess the possible association between groups and specific resistance profiles.
2.6.2. Pulsed Field Gel Electrophoresis (PFGE)
Representative MDR isolates were subjected to typing by PFGE. In total 51 E. coli isolates which were characterized as MDR, derived from different environments (6 clinical isolates, 17 from HWW, 13 from WWTP effluents, 8 from RSW1 and 7 from RSW2) and belonged to different phylogenetic groups, were subjected to genomic typing. PFGE was performed according to pulse net protocol [35]. The isolates were cultured in Nutrient broth overnight at 37°C, treated with lysozyme at 37°C for 1 hour, and then with proteinase K at 56°C for overnight incubation. After four washing steps, the DNA was digested with the rare-cutting restriction endonuclease XbaI ThermoFisher Scientific (30units/reaction) at 37°C overnight. The produced fragments of the digested genomic DNA were separated on 1% agarose gels using PFGE. Genomic profiles were visualized by staining with GelRed (Biosna) and compared visually according to Tenover et al. [36].
2.7. Statistical Analysis
Pearson's chi-square test (or Fisher's exact test in case expected values in any of the cells were below 5) was performed to examine the relationship between phylogenetic groups and origin of the sample and additionally between phylogenetic groups and resistance profiles. SPSS v.29 package was used for statistical analysis.
3. Results
3.1. E. coli Collection
The total number of E. coli colonies was determined by counting the number of characteristic colonies on the membrane filter according to ISO 9308.01-1:2017. A total of 610 colonies presumptive of E. coli (identified by their blue-violet color in CCA) were initially collected. Out of the 610 colonies, 502 (171 from WWTP, 105 from semi-treated HWW, 163 from RWS1, and 63 from RWS2 samples) were finally confirmed as E. coli using the gold standard procedures [ISO 9308.01-1:2017] and molecular uidA confirmatory test [21]. In more detail, of those 502 confirmed Ε. coli isolates, 296 (92 from WWTP, 73 from HWW, 91 from RWS1, and 40 from RWS2 samples) were collected from CCA culture media without AMP and 206 (79 from WWTP, 32 from HWW, 72 from RWS1, and 23 from RWS2 samples) from CCA/AMP. Regarding the clinical collection, a total of 139 E. coli isolates were identified and confirmed, with 104 derived from urine, 30 from blood, and 5 from patients’ tissue.
3.2. Antimicrobial Susceptibility Profiles and Assessment of Resistance Mechanisms
Considering that E. coli has no intrinsic resistance mechanisms, all isolates (environmental and clinical) were classified into specific sub-categories. For environmental isolates, 40.4% (203/502) were characterized as WT, 2.8% (14/502) as N-WT, 36.5% (183/502) as R and 20.3% (102/502) as MDR. For clinical isolates 40% (56/139) were characterized as S, 46% (64/139) as R and 14% (19/139) as MDR. Data for the characterization of resistance profiles for environmental and clinical samples are summarized in Table 1.
The resistance frequencies of the 502 environmental and 139 clinical isolates in all tested antibiotics are presented in Figure 2. Resistance to penicillins (AMP and PIP) was the most frequent among all environmental and clinical isolates followed by resistance to AMC. In more detail, 55% (275/502) of environmental isolates exhibited resistance to AMP, 53% (267/502) to PIP, and 33% (164/502) to AMC. High resistance rate to quinolones (24.9%; 125/502) was also observed and the majority of the resistant isolates were derived from HWW (33.6%; 42/125) (Figure 2). For the 139 clinical isolates, 40% (55/139) presented resistance to AMP, 33% (46/139) to PIP and 25% (35/139) to AMC. The number of different antibiotic categories in which environmental and clinical MDR isolates presented resistance are shown in Supplementary Table S1.
The resistance patterns exhibited by both environmental and clinical E. coli isolates were classified into two categories: multiple resistant patterns (MRP, resistance patterns to more than three antibiotic categories) and resistant patterns (RP, resistance patterns to maximum of three different antibiotic categories).
MRPs were further separated into six subcategories: a) MRP1- related to ESBL production, exhibiting resistance to penicillin/inhibitor combinations (such as AMC, TZP), expanded spectrum cephalosporins (such as CTX, CRO, CAZ, FEP) with or without resistance to monobactams (ATM) and positive DDST test, b) MPR2- related to ESBL production, showing resistance to expanded spectrum cephalosporins (such as CTX, CRO, CAZ, FEP) with or without resistance to monobactams (ATM) and positive DDST test, c) MRP3- related to ESBL + carbapenemase production showing resistance to expanded spectrum cephalosporins, carbapenemes (IMP, ΜΕΜ) and positive DDST and CIM test, d) MRP4- related to ESBL and AmpC production showing resistance to cephamycins (FOX) and penicillin/inhibitor combinations (AMC, TZP) in addition to resistance to expanded spectrum cephalosporins, e) MRP5- related to AmpC production exhibiting resistance to cephamycins (FOX), to penicillin/inhibitor combinations (AMC, TZP) and negative DDST test and f) other MRPs (MRP6- 10) in which resistance to penicillins and to other non β-lactam antibiotics (such as aminoglycosides, SXT and quinolones) was observed (Table 2). MRP2 and MRP1 were the most frequent MRPs among MDR environmental and clinical isolates. Specifically, 32.3% (33/102) environmental MDR isolates presented MRP2 pattern, while 29.4% (30/102) environmental and 36.8% (7/19) clinical MDR isolates presented MRP1 pattern. Furthermore, 50 environmental and eight clinical isolates with ESBL related-MRPs presented concomitant resistance to quinolones (see Table 2).
Similarly, RPs were further divided into five subcategories: a) RP1- Related to ESBL production patterns with resistance to expanded spectrum cephalosporins and positive DDS test, b) RP2- Related to AmpC production with resistance to penicillins, penicillin/inhibitor combinations and cephamycin, c) RPs3 (a-d) in which resistance to penicillins and to penicillin/inhibitor combinations with or without co-resistance to non β-lactam antibiotics such as quinolones, aminoglycosides and SXT, was observed d) RPs4 (a-g) in which resistance to penicillins with or without co-resistance to non β-lactam antibiotics, was observed and e) RP5- 6 in which resistance only to non β-lactam antibiotics was observed (Supplementary Table S2). Our results show that RP3a was the most frequent RP among 183 R environmental (30.6%; 56/183) and among 64 R clinical isolates (29.6%; 19/64). Also, four R environmental (2 from RWS1 and 2 from RWS2) and one R clinical isolates were found to be potential ESBL producers.
3.3. Resistance Genes Detection
All β-lactamase producers (n=80) were screened for β-lactamase genes. Thirty two of the 68 potential β-lactamase producers from the environment were isolated from HWW, while the remaining derived from the WWTP effluents (n=14), RWS1 (n=15) and RWS2 (n=7). Regarding the 12 clinical potential β-lactamase producers, eight, three and one were isolated from urine, blood, and tissue, respectively. The characteristics of those isolates are shown in Supplementary Table S3. BlaCTX-M-group 1-type gene was detected in 52 isolates (65%; 52/80) blaCTX-M-group 9-type gene was identified in 7 isolates (9%, 7/80) BlaTEM gene was detected in 12 isolates (15%; 12/80) and blaSHV gene in 17 isolates (21%; 17/80) (see Supplementary Table S3).
One isolate with MRP3 profile was positive for CIM test indicating the presence of carbapenemase. By molecular carbapenemase screening the isolate was found positive for blaOXA-48-type gene which was identified by sequencing coding for OXA-244 enzyme (Supplementary Tables S3 and S4). In two isolates with MRP- 4, blaCMY-2 type, and blaFOX type genes were detected, coding for AmpC type enzymes, the CMY-4 and FOX-17 respectively (Supplementary Tables S3 and S4). Detailed data for the detection rate of β-lactamase genes in E. coli isolates derived from environmental and clinical samples are summarized in Table 3.The sequencing analysis confirmed the resistance genes with an identity value of 99% to 100% (Supplementary Table S4). Finally, the sul1 gene was detected in 22/29 MDR isolates exhibiting resistance to SXT (5 clinical, 7 from HWW, 6 from WWTP, 3 from RWS1 and 1 from RWS2).
3.4. Molecular Typing Analysis
There was statistically significant correlation between the phylogenetic group and the origin of the sample [X2 (12, N= 641) = 110.63, p <0.001)] (Supplementary Table S5a-b). Group A is the predominant (48%, 242/502) in all the environmental sample sources, followed by B2 (20%, 102/502), B1 (17%, 85/502), and D (15%, 73/502) (Figure 3a). Moreover, the occurrence of group B2 was higher in E. coli isolates from wastewater samples (WWTP effluents and HWW) compared to other environmental sources evaluating the adjusted ratios (Supplementary Table S5a). In contrast to environmental, in clinical isolates, group B2 was the predominant phylogenetic group (60%; 84/139) followed by A (18%, 25/139), D (17%, 24/139) and B1 (4%, 6/139) (Figure 3a). The above comparisons are in agreement with the adjusted ratios (Supplementary Table S5a).
A chi-square test of independence showed that there was association between phylogenetic group and resistance profiles [X2 (18, N = 641) = 184.09, p <0.001] (Supplementary Table S6a-b). Group A was the dominant group among all E. coli populations, including MDR, R, WT, and N-WT, in environmental samples, while group B2 was dominant in clinical isolates (among all populations, including MDR, R, and S) (Supplementary Table S6a-b, Figure 3b). PFGE analysis revealed diverse genetic fingerprints (Supplementary Figure S1) and thus did not provide additional information on the molecular classification of E. coli isolates.
A number of isolates that produce the same β-lactamase and derived from different sources, the Maximum Likelihood phylogeny revealed the following: a) BlaCTX-M-1-like genes distributed the isolates into three groups and all clinical isolates were clustered together into the same subgroup, b) isolates which possessed blaCTX-M-9 gene and derived from RSW2 grouped together, c) isolates harboring blaSHV gene were divided into three clusters and isolates from RSW1 and WWTP effluents belonged to the same subgroup (see Supplementary Figure S2).
4. Discussion
R and MDR Enterobacterales pose an important human health issue due to the scarcity of available treatment options. In recent years, One Health approach has been adopted to recognize the role of the environment in the dissemination of ARB, including ESBL-producing E. coli [2]. E. coli is a fundamental fecal indicator in monitoring the impact of effluents on the environment. Our analysis presents data that confirm river water and reclaimed wastewater are reservoirs of R and MDR E. coli to commonly used antibiotics in clinical practice such as AMP, CIP, SXT, ESCs [8,18,19,37]. We also report antibiotic resistance to penicillins (AMP, PIP) as the most frequent among both environmental and clinical isolates, although we also observed a high quinolone resistance rate in HWW [38,39,40].
ESBL-producing E. coli, specifically CTX-M-producing isolates (subtypes of the CTX-M-1 and CTX-M-9 groups) which are the predominant types in the studied habitats (environmental and clinical) are also widely found to be isolated from various aquatic environments (such as river and lakes) as well as hospitalized patients [38,39,41,42,43]. A portion of ESBL producers isolated from patients' samples and wastewaters or river waters had the same resistance profiles, belonged to the same phylogenetic group, and carried the same resistance gene (see Supplementary Table S3).
Phylogenetic group B2 has been previously reported to predominate in hospital environments [44,45,46]. Similarly, in our clinical isolates, B2 phylogroup predominated and was also found as the second most frequent group in HWW and in WWTP effluents (Figure 3a, Supplementary Table S5a). Group B2 and group D isolates possess the chuA gene [34], which is responsible for hemin utilization and has been identified in several pathogenic E. coli strains [47,48,49]. That fact implies a strong correlation between pathogenicity and phylogenetic groups B2 and D. In our study, a portion of clinical and environmental MDR and R isolates were classified into those phylogenetic groups B2 and D (Figure 3b), highlighting the high human health risks caused by exposing to possible pathogenic R and MDR E. coli isolates deriving from enviromental sources such as rivers
The reported results reveal that treated wastewater and river water are sources of resistant bacteria. The potential reuse of treated wastewater and river water exclusively for restricted crop irrigation depending on the method of watering (e.g., spraying) may expose humans to the risk of developing gastroenteritis, particularly through droplet ingestion [50,51,52]. Regarding the risk of developing urinary tract infection (UTI), E. coli is by far the most common cause for both community and hospital-acquired UTIs. For UTI treatment, the recommended antimicrobials are SXT, CIP, and AMC. In our study, E. coli strains that were found to be MDR, including those with co-resistance to SXT, CIP, and AMC (Table 1), were isolated not only from biological fluids of patients but also from all environmental habitats (see Figure 2). This fact demonstrates that human health risks can be caused by being exposed to MDR E. coli isolates present in wastes and aquatic environments.
In our study, due to strict lockdown measures imposed during the COVID-19 pandemic, we were unable to carry out some samplings, which made seasonal analysis not feasible. Additionally, the molecular typing techniques employed did not provide adequate clustering information concerning the circulation of specific E. coli types between clinical settings and the environment. Nevertheless, this study represents the first systematic collection of E. coli isolates obtained from wastewater and river water samples from Livadeia, Greece, an area that combines urban life, husbandry, and agriculture. Despite these limitations, this work provides valuable insights into E. coli resistance profiles and genotypes present in wastewaters and aquatic habitats. The presence of AR E. coli isolates with the same MRP in clinical and HWW samples sheds light on the spread of resistant bacteria in water bodies. The reported findings suggest a potential exchange of AR bacterial populations and similar AR determinants between clinical and environmental habitats. This raises concerns for public health, as aquatic environments could serve as reservoirs for the transmission of resistance genes to various bacterial species.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: The E. coli isolates from environmental habitats and clinical specimens that exhibit multidrug resistance; Table S2: Observed patterns of R isolates (RP, resistant patterns); Table S3: Characteristics of environmental and clinical isolates harboring β-lactamasegenes; Table S4: Sequencing results for the possible β-lactamase producers; Table S5a: Examination of the relationship between phylogenetic groups and origin of the sample; Sample and Group Crosstabulation, Table S5b: Examination of the relationship between phylogenetic groups and origin of the sample; Pearson's chi-square test results; Table S6a: Examination of the relationship between phylogenetic groups and resistance profile; Resistance profile and Group Crosstabulation; Table S6b: Examination of the relationship between phylogenetic groups and resistance profile; Pearson's chi-square test results. Figure S1: PFGE analysis; Diverse PFGE patterns of E. coli isolated from clinical and environmental samples Figure S2: Maximum Likelihood phylogenetic trees for A) blaCTX-M-groups, B) blaTEM and C) blaSHV nucleotide sequences.
Author Contributions
Conceptualization, A.B.; data curation, C.D., O.P., P.G., V.M. and A.B.; formal analysis, C.D., O.P., V.M. and A.B.; funding acquisition, C.D. and A.B.; investigation, C.D., O.P.; isolates collection, C.D. and A.T.; methodology, C.D., O.P., P.G., V.M. and A.B.; experimental work, C.D., E.S., S.B.; project administration, C.D., V.M. and A.B.; supervision, P.G., V.M., and A.B.; validation, P.G., V.M., and A.B.; writing—original draft, C.D., O.P.; writing—review and editing, C.D., P.G., V.M. and A.B.; All authors have read and agreed to the final published version of the manuscript.
Funding
CD was granted a PhD Fellowship by the Special Account for Research Grants (ELKE) of the Research Committee at the University of West Attica (UNIWA), under Grant/Award Number 8068/07-04-2020. The funders played no part in the design of the study, data collection and analysis, decision to publish, or manuscript preparation.
Ethics Statement
This study has received approval from the Bioethics and Research Ethics Committee of the
University of West Attica (Reference Number: 33114/13-04-2021).
Acknowledgments
Authors would like to express their gratitude to Mr. Nikolaos Kaparos and the entire staff of the Livadeia Wastewater Treatment Plant, as well as the personnel of the Microbiology Laboratory of the General Prefectural Hospital of Livadeia for their excellent cooperation. The authors would also like to thank C. Vrysis for statistical analysis, as well as, S. Bertalli, D. Polonyphi, V. Papagianni, K. Aggelou, C. Danalia, and E. Chrysochoou for their support during the laboratory work.
Conflicts of Interest
The University of West Attica is the recipient of grant and/or sponsorship income from Gilead and GSK for research projects of which AB is the principal investigator. All other authors declare that there is no conflict of interest relevant to this work.
Data availability
All data relevant to this work are available upon reasonable request.
References
- Kim S, Aga DS. Potential ecological and human health impacts of antibiotics and antibiotic-resistant bacteria from wastewater treatment plants. J Toxicol Environ Health B Crit Rev 2007, 10, 559–573. [CrossRef] [PubMed]
- World Health Organization. G: Antimicrobial Resistance: Global Report on Surveillance; WHO, 2014.
- Zanotto C, Bissa M, Illiano E, et al. Identification of antibiotic-resistant Escherichia coli isolated from a municipal wastewater treatment plant. Chemosphere, 2016, 164, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Ben W, Wang J, Cao R, et al. Distribution of antibiotic resistance in the effluents of ten municipal wastewater treatment plants in China and the effect of treatment processes. Chemosphere, 2017, 172, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Aydin S, Aydin ME, Ulvi A, Kilic H. Antibiotics in hospital effluents: occurrence, contribution to urban wastewater, removal in a wastewater treatment plant, and environmental risk assessment. Environ Sci Pollut Res Int, 2019, 26, 544–558. [Google Scholar] [CrossRef] [PubMed]
- Cahill N, O'Connor L, Mahon B, et al. Hospital effluent: A reservoir for carbapenemase-producing Enterobacterales? Sci. Total Environ, 2019, 672, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Amarasiri, M. Amarasiri M., Sano D., Suzuki S. Understanding human health risks caused by antibiotic resistant bacteria (ARB) and antibiotic resistance genes (ARG) in water environments: Current knowledge and questions to be answered, Crit Rev Environ Sci Technol, 2020, 50, 2016–2059. [Google Scholar] [CrossRef]
- Watkinson AJ, Micalizzi GB, Graham GM, et al. Antibiotic-resistant Escherichia coli in wastewaters, surface waters, and oysters from an urban riverine system. Appl Environ Microbiol, 2007, 73, 5667–5670. [Google Scholar] [CrossRef]
- Jang J, Hur HG, Sadowsky MJ, et al. Environmental Escherichia coli: ecology and public health implications-a review. J Appl Microbiol, 2017, 123, 570–581. [Google Scholar] [CrossRef]
- Haberecht HB, Nealon NJ, Gilliland JR, et al. Antimicrobial-Resistant Escherichia coli from Environmental Waters in Northern Colorado. J Environ Public Health, 2019. [CrossRef]
- Fewtrell L, Bartram J. Water quality: guidelines, standards and health: assessment of risk and risk management for water-related infectious diseases/ edited by Lorna Fewtrell and Jamie Bartram. World Health Organization, 2001. https://apps.who.int/iris/handle/10665/42442.
- Zhang CM, Xu LM, Wang XC, et al. Effects of ultraviolet disinfection on antibiotic-resistant Escherichia coli from wastewater: inactivation, antibiotic resistance profiles and antibiotic resistance genes. J Appl Microbiol 2017, 123, 295–306. [Google Scholar] [CrossRef]
- Kaper JB, Nataro JP, Mobley HL. Pathogenic Escherichia coli. Nat Rev Microbiol, 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Carducci A, Verani M. Effects of bacterial, chemical, physical and meteorological variables on virus removal by a wastewater treatment plant. Food Environ Virol, 2013, 5, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Kokkinos P, Mandilara G, Nikolaidou A, et al. Performance of three small-scale wastewater treatment plants. A challenge for possible re use. Environ Sci Pollut Res Int, 2015, 22, 17744–17752. [Google Scholar] [CrossRef] [PubMed]
- Hong P-Y, Julian TR, Pype M-L, et al. Reusing Treated Wastewater: Consideration of the Safety Aspects Associated with Antibiotic-Resistant Bacteria and Antibiotic Resistance Genes. Water, 2018, 10, 244. [Google Scholar] [CrossRef]
- Carducci A, Donzelli G, Cioni L, et al. Quantitative Microbial Risk Assessment for Workers Exposed to Bioaerosol in Wastewater Treatment Plants Aimed at the Choice and Setup of Safety Measures. Int J Environ Res Public Health, 2018, 15, 1490. [Google Scholar] [CrossRef] [PubMed]
- Hossain MS, Ali S, Hossain M, et al. ESBL Producing Escherichia coli in Faecal Sludge Treatment Plants: An Invisible Threat to Public Health in Rohingya Camps, Cox's Bazar, Bangladesh. Front Public Health, 2021, 9, 783019. [Google Scholar] [CrossRef]
- Aslan A, Cole Z, Bhattacharya A, Oyibo O. Presence of Antibiotic-Resistant Escherichia coli in Wastewater Treatment Plant Effluents Utilized as Water Reuse for Irrigation. Water, 2018, 10, 805. [Google Scholar] [CrossRef]
- Kyriakou G, Nikolaou A, Chrysafopoulou E. Interventions for the qualitative and quantitative water resources management of the Kifissos River Basin, Technical Chamber of Greece Division of Eastern Sterea Ellada. 2009.
- Bej AK, McCarty SC, Atlas RM. Detection of coliform bacteria and Escherichia coli by multiplex polymerase chain reaction: comparison with defined substrate and plating methods for water quality monitoring. Appl Environ Microbiol, 1991, 57, 2429–2432. [Google Scholar] [CrossRef]
- World Health Organization. Guidance on regulations for the transport of infectious substances 2009-2010: applicable as from . World Health Organization. https://apps.who.int/iris/handle/10665/69899. 1 January 1066.
- European Committee on Antimicrobial Susceptibility Testing. EUCAST ECOFFs and Breakpoint tables for interpretation of MICs and zone diameters. Version 12.0, 2022. http://www.eucast.org.
- Magiorakos AP, Srinivasan A, Carey RB, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect, 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Pappa O, Vantarakis A, Galanis A, et al. Antibiotic resistance profiles of Pseudomonas aeruginosa isolated from various Greek aquatic environments. FEMS microbiology ecology 2016, 92, fiw042. [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. EUCAST Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance. Version 2.0. 2017. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Resistance_mechanisms/EUCAST_detection_of_resistance_mechanisms_170711.pdf.
- van der Zwaluw K, de Haan A, Pluister GN, et al. The carbapenem inactivation method (CIM), a simple and low-cost alternative for the Carba NP test to assess phenotypic carbapenemase activity in gram-negative rods. PLoS One, 2015, 10, e0123690. [Google Scholar] [CrossRef]
- Poirel L, Héritier C, Tolün V, Nordmann P. Emergence of oxacillinase-mediated resistance to imipenem in Klebsiella pneumoniae. Antimicrob Agents Chemother, 2004, 48, 15–22. [Google Scholar] [CrossRef]
- Woodford N, Fagan EJ, Ellington MJ. Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum (beta)-lactamases. J Antimicrob Chemother, 2006, 57, 154–155. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Laboratory manual for carbapenem and colistin resistance detection and characterisation for the survey of carbapenem- and/or colistin-resistant Enterobacteriaceae – Version 2.0. Stockholm: ECDC; 2019.
- Pérez-Pérez FJ, Hanson ND. Detection of plasmid-mediated AmpC beta-lactamase genes in clinical isolates by using multiplex PCR. J Clin Microbiol, 2002, 40, 2153–2162. [Google Scholar] [CrossRef] [PubMed]
- Hannecart-Pokorni E, Depuydt F, de wit L, et al. Characterization of the 6'-N-aminoglycoside acetyltransferase gene aac(6')-Im [corrected] associated with a sulI-type integron [published correction appears in Antimicrob Agents Chemother, 42, 485]. Antimicrob Agents Chemother. 1997, 41, 314–318. [Google Scholar] [CrossRef]
- Pappa O, Beloukas A, Vantarakis A, et al. Molecular Characterization and Phylogenetic Analysis of Pseudomonas aeruginosa Isolates Recovered from Greek Aquatic Habitats Implementing the Double-Locus Sequence Typing Scheme. Microb Ecol, 2017, 74, 78–88. [Google Scholar] [CrossRef]
- Clermont O, Bonacorsi S, Bingen E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl Environ Microbiol, 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed]
- Standard operating procedure for PulseNet PFGE of Escherichia coli O157:H7, Escherichia coli non O157 (STEC), Salmonella serotypes, Shigella sonnei and Shigella flexneri. April 2013.
- Tenover FC, Arbeit RD, Goering RV, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol, 1995, 33, 2233–2239. [Google Scholar] [CrossRef]
- Suzuki Y, Hashimoto R, Xie H, et al. Growth and antibiotic resistance acquisition of Escherichia coli in a river that receives treated sewage effluent. Sci Total Environ, 2019, 690, 696–704. [Google Scholar] [CrossRef]
- Reinthaler FF, Posch J, Feierl G, et al. Antibiotic resistance of E. coli in sewage and sludge. Water Res, 2003, 37, 1685–1690. [Google Scholar] [CrossRef]
- Hassan, SA. , Jamal SA, Kamal M. Occurrence of multidrug resistant and ESBL producing E.coli causing urinary tract infections. Australian Journal of Basic and Applied Sciences, 2011, 7, 39–43. [Google Scholar]
- Hu YY, Cai JC, Zhou HW, et al. Molecular typing of CTX-M-producing escherichia coli isolates from environmental water, swine feces, specimens from healthy humans, and human patients. Appl Environ Microbiol, 2013, 79, 5988–5996. [Google Scholar] [CrossRef] [PubMed]
- Saltoglu N, Karali R, Yemisen M, et al. Comparison of community-onset healthcare-associated and hospital-acquired urinary infections caused by extended-spectrum beta-lactamase-producing Escherichia coli and antimicrobial activities. Int J Clin Pract, 2015, 69, 766–770. [Google Scholar] [CrossRef]
- Stoll C, Sidhu JP, Tiehm A, Toze S. Prevalence of clinically relevant antibiotic resistance genes in surface water samples collected from Germany and Australia. Environ Sci Technol, 2012, 46, 9716–9726. [Google Scholar] [CrossRef] [PubMed]
- Liu H, Zhou H, Li Q, et al. Molecular characteristics of extended-spectrum β-lactamase-producing Escherichia coli isolated from the rivers and lakes in Northwest China. BMC Microbiol, 2018, 18, 125. [CrossRef]
- Lee JH, Subhadra B, Son YJ, et al. Phylogenetic group distributions, virulence factors and antimicrobial resistance properties of uropathogenic Escherichia coli strains isolated from patients with urinary tract infections in South Korea. Lett Appl Microbiol, 2016, 62, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Iranpour D, Hassanpour M, Ansari H, et al. Phylogenetic groups of Escherichia coli strains from patients with urinary tract infection in Iran based on the new Clermont phylotyping method. Biomed Res Int, 2015, 2015, 846219. [Google Scholar] [CrossRef]
- Dadi BR, Abebe T, Zhang L, et al. Distribution of virulence genes and phylogenetics of uropathogenic Escherichia coli among urinary tract infection patients in Addis Ababa, Ethiopia. BMC Infect Dis, 2020, 20, 108. [Google Scholar]
- Spurbeck RR, Dinh PC Jr, Walk ST, et al. Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract. Infect Immun, 2012, 80, 4115–4122. [Google Scholar] [CrossRef]
- Stoppe NC, Silva JS, Carlos C, et al. Worldwide Phylogenetic Group Patterns of Escherichia coli from Commensal Human and Wastewater Treatment Plant Isolates. Front Microbiol, 2017, 8, 2512. [Google Scholar] [CrossRef]
- Nagy G, Dobrindt U, Kupfer M, et al. Expression of hemin receptor molecule ChuA is influenced by RfaH in uropathogenic Escherichia coli strain 536. Infect Immun, 2001, 69, 1924–1928. [Google Scholar] [CrossRef] [PubMed]
- Mbanga J, Abia ALK, Amoako DG, et al. Quantitative microbial risk assessment for waterborne pathogens in a wastewater treatment plant and its receiving surface water body. BMC Microbiol, 2020, 20, 346. [Google Scholar]
- Sano D, Haas C.N. and Rose JB, (2019). A QMRA Framework for Sanitation Treatment Decisions. In: J.B. Rose and B. Jiménez-Cisneros, (eds) Water and Sanitation for the 21st Century: Health and Microbiological Aspects of Excreta and Wastewater Management (Global Water Pathogen Project). (J.B. Rose and B. Jiménez-Cisneros) (eds) Part 1: The Health Hazards of Excreta: Theory and Control), Michigan State University, E. Lansing, MI, UNESCO.
- Singh G, Vajpayee P, Rani N, et al. Exploring the potential reservoirs of non specific TEM beta lactamase (bla(TEM)) gene in the Indo-Gangetic region: A risk assessment approach to predict health hazards. J Hazard Mater 2016, 314, 121–128. [CrossRef] [PubMed]
Figure 1.
(a) The Boeotia regional unit is located in Central Greece and is divided into six municipalities. Livadeia serves as the capital of the Boeotia regional district. (b) The map on the right depicts the sampling locations and their relationships. The abbreviations used in the map are as follows: HWW, hospital wastewater; WWTP, wastewater treatment plant; RWS1, river water site 1 (located 100m downstream from the WWTP discharge site); RWS2, river water site 2 (located 6 km downstream from the WWTP discharge site).
Figure 1.
(a) The Boeotia regional unit is located in Central Greece and is divided into six municipalities. Livadeia serves as the capital of the Boeotia regional district. (b) The map on the right depicts the sampling locations and their relationships. The abbreviations used in the map are as follows: HWW, hospital wastewater; WWTP, wastewater treatment plant; RWS1, river water site 1 (located 100m downstream from the WWTP discharge site); RWS2, river water site 2 (located 6 km downstream from the WWTP discharge site).
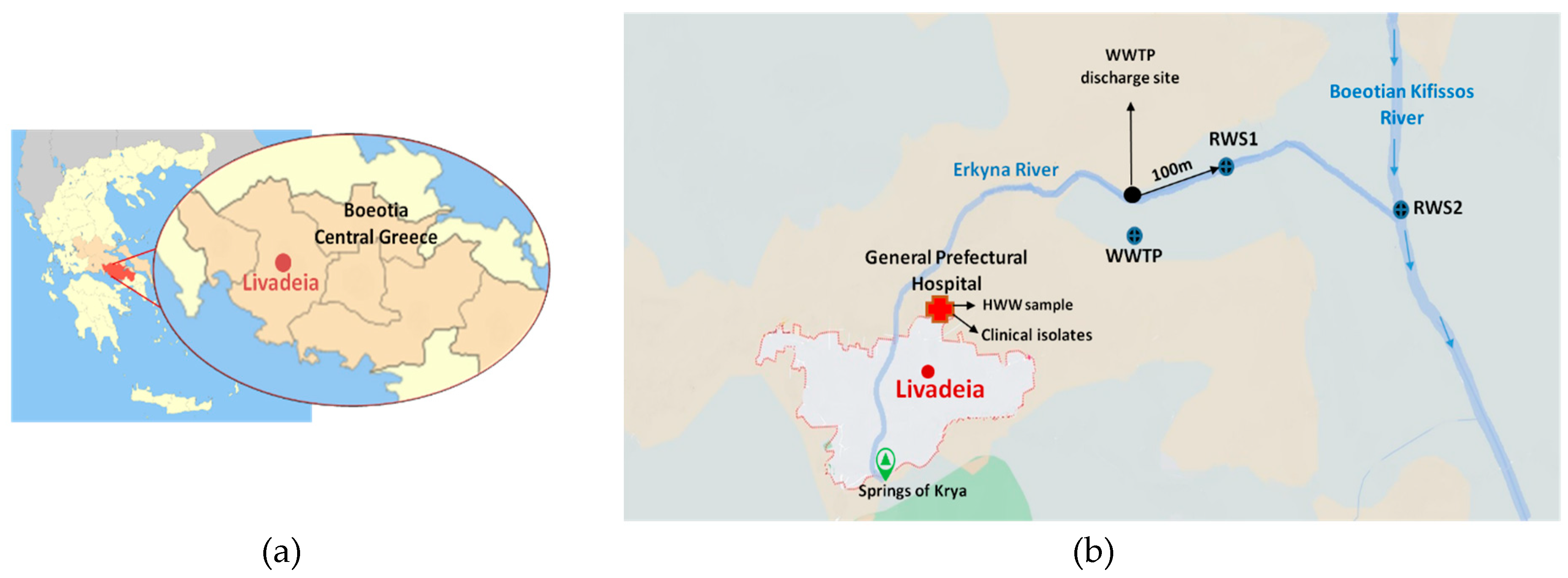
Figure 2.
The frequency rate of resistance to each antibiotic per origin of sample. Abbreviations: AMP, ampicillin; AMC, amoxicillin/clavulanic acid; TZP, piperacillin/ tazobactam; PIP, piperacillin; CXM, cefuroxime; CAZ, ceftazidime; CTX, cefotaxime; FOX, cefoxitin; FEP, cefepime; CRO, ceftriaxone; ATM, aztreonam; MEM, meropenem; IMP, imipenem; GM, gentamicin; AN, amikacin; SXT, sulfamethoxazole-trimethoprim; CIP, ciprofloxacin; NAL, nalidixic acid. HWW, hospital wastewater; WWTP, wastewater treatment plant; RWS1, river water site 1; RWS2, river water site 2.
Figure 2.
The frequency rate of resistance to each antibiotic per origin of sample. Abbreviations: AMP, ampicillin; AMC, amoxicillin/clavulanic acid; TZP, piperacillin/ tazobactam; PIP, piperacillin; CXM, cefuroxime; CAZ, ceftazidime; CTX, cefotaxime; FOX, cefoxitin; FEP, cefepime; CRO, ceftriaxone; ATM, aztreonam; MEM, meropenem; IMP, imipenem; GM, gentamicin; AN, amikacin; SXT, sulfamethoxazole-trimethoprim; CIP, ciprofloxacin; NAL, nalidixic acid. HWW, hospital wastewater; WWTP, wastewater treatment plant; RWS1, river water site 1; RWS2, river water site 2.
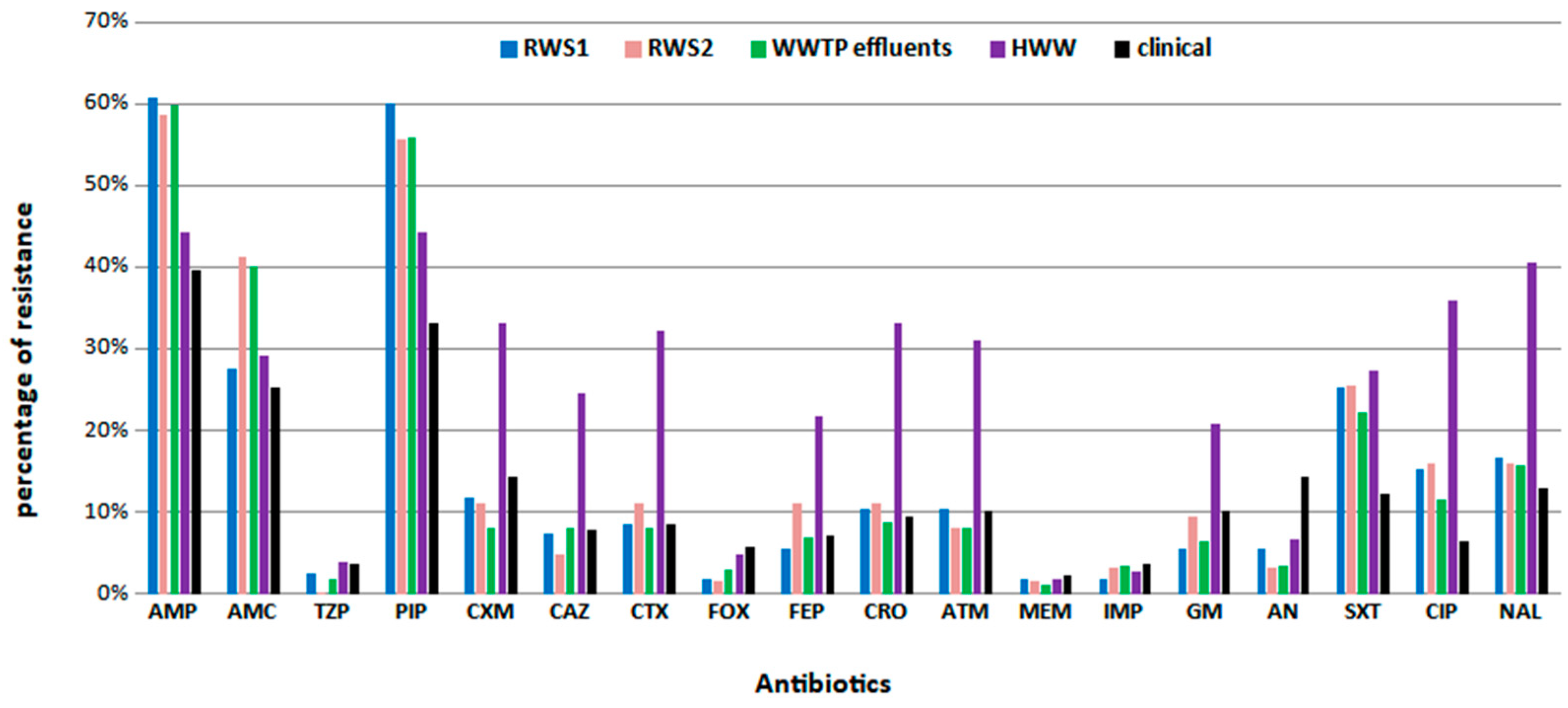
Figure 3.
(a) The distribution of each phylogenetic group among different habitats (upper figure) and (b) the relationship between phylogenetic groups and resistance profiles (lower figure). Abbreviations: RWS1, river water site 1; RWS2, river water site 2; WWTP, wastewater treatment plant; HWW, hospital wastewater; MDR, multi-drug resistant; R, resistant; N-WT, non-wild type; WT, wild type.
Figure 3.
(a) The distribution of each phylogenetic group among different habitats (upper figure) and (b) the relationship between phylogenetic groups and resistance profiles (lower figure). Abbreviations: RWS1, river water site 1; RWS2, river water site 2; WWTP, wastewater treatment plant; HWW, hospital wastewater; MDR, multi-drug resistant; R, resistant; N-WT, non-wild type; WT, wild type.

Table 1.
Resistance rate of environmental and clinical E. coli isolates.
| Environmental isolates | WT | N-WT | R | MDR |
|---|---|---|---|---|
| HWW (n=105) | 54.3% (57/105) | 3.8% (4/105) | 7.6% (8/105) | 34.3% (36/105) |
| WWTP (n=171) | 36.8% (63/171) | 3.5% (6/171) | 43.3% (74/171) | 16.4% (28/171) |
| RWS1 (n=163) | 36.8% (60/163) | 1.2% (2/163) | 45.4% (74/163) | 16.6% (27/163) |
| RWS2 (n=63) | 36.5% (23/63) | 3.1% (2/63) | 42.9% (27/63) | 17.5% (11/63) |
| Clinical isolates | S | R | MDR | |
| urine (n=104): | 41.3% (43/104) | 45.2% (47/104) | 13.5% (14/104) | |
| blood (n=30) | 43.3% (13/30) | 43.3% (13/30) | 13.4% (4/30) | |
| tissue (n=5) | - | 80% (4/5) | 20% (1/5) | |
Abbreviations: WT, wild type; N-WT, non-wild type; S, sensitive; R, resistant; MDR, multi-drug resistant; HWW, hospital wastewater; WWTP, wastewater treatment plant; RWS1, river water site 1; RWS2, river water site 2.
Table 2.
Observed patterns of MDR isolates (MRP, multiple resistant patterns).
| Environmental isolates (source) | Clinical isolates | |||
|---|---|---|---|---|
| MRP1: Related to ESBL production and resistance to penicillin/inhibitor combinations | PEN/ PEN–inhibitor/ ESCs + SXT | 1 (WWTP) | - | |
| PEN/ PEN-inhibitor/ ESCs + QNs | 1 (HWW) | - | ||
| PEN/ PEN-inhibitor/ ESCs/ ATM | 2 (1 RWS1, 1 RWS2) | - | ||
| PEN/ PEN-inhibitor/ ESCs/ ATM + QNs | 9 (3 HWW, 3 WWTP, 3 RWS1) | - | ||
| PEN/ PEN-inhibitor/ ESCs/ ATM + AMG | 2 (1 WWTP, 1 RWS2) | - | ||
| PEN/ PEN-inhibitor/ ESCs/ ATM + SXT | - | 1 | ||
| PEN/ PEN-inhibitor/ ESCs/ ATM + SXT + QNs | 2 (1 HWW + 1 RWS2) | 2 | ||
| PEN/ PEN-inhibitor/ ESCs/ ATM + AMG + QNs | 2 (HWW) | 3 | ||
| PEN/ PEN-inhibitor/ ESCs/ ATM + AMG + SXT + QNs | 11 (HWW) | 1 | ||
| Total MRP1: 37 | 30 | 7 | ||
| MRP2: Related to ESBL production | PEN/ ESCs/ ATM + QNs | 8 (4 HWW, 3 RWS1, 1 RWS2) | - | |
| PEN/ ESCs/ ATM + AMG | 3 (2 RWS1, 1 RWS2) | - | ||
| PEN/ ESCs/ ATM + SXT | 7 (6 WWTP, 1 RWS1) | - | ||
| PEN/ ESCs/ ATM + SXT + QNs | 2 (WWTP) | 1 | ||
| PEN/ ESCs/ ATM + AMG + SXT + QNs | 8 (6 HWW, 2 RWS1) | - | ||
| PEN/ ESCs/ ATM + AMG + QNs | 4 (HWW) | - | ||
| PEN/ ESCs + SXT + QNs | 1 (WWTP) | - | ||
| Total MRP2: 34 | 33 | 1 | ||
| MRP 3: Related to ESBL + carbapenemase production | PEN/ PEN-inhibitor/ ESCs/ CARB/ ATM + SXT | 1 (RWS1) | - | |
| Total MRP3: 1 | 1 | - | ||
| MRP 4: Related to ESBL + AmpC β-lactamases production | PEN/ PEN-inhibitor/ ESCs/ FOX/ ATM | 2 (1 HWW, 1 RWS1) | 1 | |
| PEN/ PEN-inhibitor/ ESCs/ FOX/ ATM + AMG + SXT | - | 1 | ||
| PEN/ PEN-inhibitor/ ESCs/ FOX/ ATM + SXT + QNs | - | 1 | ||
| PEN/ PEN-inhibitor/ ESCs/ FOX + AMG + SXT + QNs | 2 (HWW) | - | ||
| Total MRP 4: 7 | 4 | 3 | ||
|
MRP 5: Related to AmpC β-lactamases production |
PEN/ PEN-inhibitor/ FOX + AMG + QNs | 1 (WWTP) | ||
| PEN/ PEN-inhibitor/ FOX + QNs | 1 (WWTP) | |||
| PEN/ PEN-inhibitor/ NSCs/ FOX + AMG | 2 (RWS1) | 2 | ||
| Total MRP5: 6 | 4 | 2 | ||
| MPR 6 | Susceptibility to cephalosporins Penicillinase production with resistance to other non β-lactam antibiotics |
PEN/ PEN-inhibitor+ SXT+ QNs | 9 (2 WWTP, 4 RWS1, 3 RWS2) | 1 |
| Total MRP6: 10 | 9 | 1 | ||
| MRP 7 | PEN/ PEN-inhibitor+ AMG + SXT | 10 (6 WWTP, 4 RWS1) | 5 | |
| Total MRP7: 13 | 10 | 3 | ||
| MRP 8 | PEN/ PEN-inhibitor +AMG + SXT+ QNs | 7 (1 HWW, 1 WWTP, 2 RWS1, 3 RWS2) | 2 | |
| Total MRP8: 9 | 7 | 2 | ||
| MRP 9 | PEN/ PEN-inhibitor+ AMG + QNs | 1 (WWTP) | - | |
| Total MRP9: 1 | 1 | |||
| MRP10 | PEN+ AMG + SXT+ QNs | 3 (2 WWTP, 1 RWS1) | - | |
| Total MRP10: 3 | 3 | |||
total MRPs: 121. total environmental isolates with MRP 102 and total clinical isolates with MRP: 19. Abbreviations: MRP, multiple resistant patterns; ESBL, extended-spectrum-β-lactamase; PEN, penicillins; PEN–inhibitor, penicillin-inhibitor combinations; ESCs, Extended spectrum cephalosporins; SXT, sulfamethoxazole-trimethoprim; QNs, quinolones; ATM, aztreonam; AMG, aminoglycosides; CARB, carbapenems; FOX, cefoxitin; NSCs, narrow spectrum cephalosporins; HWW, hospital wastewater; WWTP, wastewater treatment plant; RWS1, river water site 1; RWS2, river water site 2.
Table 3.
Detection rate of β-lactamase genes among clinical and environmental isolates.
| β-lactamase genes | Clinical isolates | Environmental isolates | Total | ||||
|---|---|---|---|---|---|---|---|
| HWW | WWTP effluents | RWS1 | RWS2 | ||||
| ESBL genes | blaCTX-M-group 1-type | 10 | 22 | 10 | 8 | 2 | 52 |
| blaCTX-M-group 9-type | 1 | 3 | 3 | 7 | |||
| blaSHV | 1 | 12 | 2 | 3 | 17 | ||
| blaTEM | 4 | 3 | 2 | 3 | 12 | ||
| Carbapenemase genes | blaOXA-48-type | 1 | 1 | ||||
| AmpC type genes | blaCMY-2 type | 1 | 1 | ||||
| blaFOX-type | 1 | 1 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



