
Submitted:
16 May 2023
Posted:
17 May 2023
You are already at the latest version
Abstract
It was found that patients with comorbidities of obesity and diabetes have a high risk of breast cancer occurrence and face worse breast cancer outcomes. Though several reports showed the reinforced link between obesity, diabetes, and prediabetes with breast cancer, the underlying molecular mechanism is still unknown. The present study aimed to investigate the underlying molecular link between increased risks of breast cancer due to coincident diabetes or obesity using a spontaneous obese rat model with impaired glucose tolerance (WNIN/GR-Ob rat). A single dose of solubilized DMBA suspension (40 mg/kg body weight) was orally administered to the animals at the age of 60 days to induce breast tumors. The tumor incidence, latency period, tumor frequency, and tumor volume were measured. Histology, immunohistochemistry, and immunoblotting were performed to evaluate the tumor morphology and expression levels of signal molecules. It was observed that the onset of breast tumor and latency period per tumor development were early in GR-Ob rats compared to respective lean rats. It was found that 62% of obese rats were bearing tumors, and in comparison, only 21% of the lean animals developed breast tumors. Overexpression of ER, PR, Ki67, and p53 markers was observed in tumor tissues of obese rats in comparison with lean rats. The levels of the hallmarks of cell proliferation and angiogenesis involved in IGF-1/PI3K/Akt/GSK3β/β-catenin signaling pathway molecules were upregulated in obese rat breast tumors compared to lean rats. Furthermore, obesity with prediabetes is associated with changes in IGF-1 signaling and acts on PI3K/Akt/GSK3β/β-catenin signaling, which results in quick cell proliferation and development of breast tumors in obese rats than the lean rats. These results indicate that the onset of the tumor and its development was faster in a spontaneous obese rat model with impaired glucose tolerance than their lean counterparts, with a higher percentage of tumor incidence.
Keywords:
Breast cancer
; obesity
; metabolic syndrome
; rat model
; DMBA
; insulin signaling
; PI3/Akt
1. Introduction
Carcinoma of the breast is currently the leading cause of global cancer incidence with an estimated 2.3 million new cases in 2020, and the fifth leading cause of cancer mortality. Breast cancer is the most commonly diagnosed cancer among women accounting for 1 in 4 cancer cases and for 1 in 6 cancer deaths [1]. Epidemiological investigations have established a strong association of breast cancer with metabolic dysregulation that leads to diabetes and obesity [2,3,4].
Obesity, a chronic disease with increasing prevalence both in wealthy nations as well as in low- and middle-income countries, has emerged as a global epidemic. Globally, the incidence of obesity has tripled since 1975 due to sedentary lifestyle and unhealthy dietary patterns. According to the WHO Report [5], over 1 billion people worldwide are obese that includes 650 million adults, 340 million adolescents, and 39 million children. It is estimated that 1 in 5 women and 1 in 7 men will be living with obesity by 2030 [6]. Likewise, there is a significant global prevalence of diabetes mellitus among 20–79-year-olds with an estimated 10.5% (536.6 million) living with the disease in 2021 that is predicted to rise to 12.2% (783 million) by 2045 [7]. Global estimates reveal that 1 in 10 people in the world are diabetic, while 3 in 4 diabetics live in low- and middle-income countries [7]. Most importantly, over 541 million adults have Impaired Glucose Tolerance (IGT) referred to as prediabetes, which places them at high risk of type 2 diabetes (T2D). Furthermore, the probability of prediabetics developing diabetes is 3-10 folds higher than in normoglycemic individuals [8].
Accumulating evidence indicates a strong association between obesity, T2D, and several types of cancer, including breast cancer [9,10,11,12]. High body mass index (BMI), insulin resistance, increased levels of leptin and aromatase enzyme, and inflammations of breast adipose tissue are believed to contribute to obesity related post-menopausal breast cancer [12]. In a recent study, high BMI, a widely used index of obesity, was an independent factor associated with a high 21-gene recurrence score in estrogen receptor (ER)–positive, ERBB2-negative young (≤45 years) breast cancer patients [13]. Several studies have provided compelling evidence to demonstrate an increased risk of breast cancer among diabetic women as well as higher mortality and diminished quality of life [14,15,16,17]. Diabetes as well as prediabetes is associated with risk of breast cancer, especially in hormone receptor positive molecular subtypes [18].
Co-incident obesity and T2D is recognized to increase the incidence of all molecular subtypes of breast cancer and worse outcomes, besides significantly lowering the survival rate [19,20,21,22,23]. Hyperglycemia, hyperinsulinemia, and insulin resistance, cardinal features of obesity and T2D are believed to promote breast carcinogenesis [23,24]. A bi-directional relationship was observed between dysregulated glucose/insulin metabolisms with breast cancer [25]. In this clinical study, investigators found a correlation between severities of glucose/insulin metabolism with tumor and insulin resistance-related markers. The PI3K/Akt signaling pathway that plays a central role in various physiological processes and mediates the biological effects of insulin and insulin-like growth factor-1 (IGF-1) is aberrantly activated in diabetes/IGT, obesity, and breast cancer suggesting a possible link between these co-morbidities [26].
The National Institute of Nutrition has developed a unique spontaneous mutant obese rat model with impaired glucose tolerance (WNIN/GR-Ob) that can be transformed into frank diabetes by dietary manipulations [27]. WNIN/GR-Ob rat displays a set of characteristics and features associated with the other obese animal models and in addition, exhibits impaired glucose tolerance (IGT), all of which make WNIN/GR-Ob rat a suitable model of metabolic syndrome (MetS) [28]. The present study was undertaken to investigate the combined effect of obesity and IGT on the development of chemically-induced mammary tumors in the WNIN/GR-Ob rat model. Hormone receptor status as well as the expression of Ki-67, p53, and key molecules in the IGF-1/PI3K/GSK3β/β-catenin signaling pathway were assessed by immunohistochemical and immunoblotting analyses to evaluate the efficacy of 7,12-dimethylbenz[a]anthracene-(DMBA) induced mammary carcinogenesis in WNIN/GR-Ob rats as a co-incident model of IGT/obesity and breast tumor.
2. Results
2.1. Obesity accelerates the onset and development of breast cancer
The development of breast tumors was examined physically by palpation. No mammary tumors were detected in both control lean and control obese rats (Figure 1A and B). The onset of breast tumors occurred earlier in WNIN/GR-Ob rats administered with DMBA than in the lean rats with DMBA. The onset of tumor development in obese rats was observed after the 9th week of DMBA administration, whereas it was observed after the 26th week in counter-lean rats. After 32 weeks of administration of DMBA, 62% of obese rats developed mammary tumors, while only 21% of the lean animals developed breast tumors (Figure 1C). The average latency period for tumor development was 119 days in obese rats administered with DMBA compared to 211 days in the lean rats administered with DMBA (Table 1). The average tumor latency was shorter in the obese rats by 92 days when compared to lean rats. It was clear that the onset of the tumor and its development was faster in obese rats than their counter lean rats with a higher percentage of tumor incidences in obese rats than the lean rats. Rat breast tissues of all experimental animals were observed for their histomorphology by H&E staining (Figure 2). Tumor mass have lobules with pleomorphic epithelial cells with distinct variation in cell size and shape. The nuclear pleomorphism was observed with mitosis (Figure 2). Pathology of these tumors revealed that in the obese rats administered with DMBA, 20% of the rats had adenocarcinoma and 40% had fibroadenoma and all the tumors of lean rats were of adenocarcinoma.
2.2. Obesity promotes oncogenic markers expression in rat mammary gland
In the process of diagnosing breast cancer, the prognostic markers ER, PR, Ki67, and p53 proved to be the most effective. Researchers looked for several different kinds of signs of breast cancer and measured how much they were present in the breast tissue of both lean and fat rats that had been given DMBA to cause breast tumors. Positive staining areas were observed in each and every breast tumor segment, and this was true regardless of whether the rats were thin or obese. It was shown that the expression of these markers was significantly higher in the breast tissue of tumors than it was in normal breast tissue. When compared with the expression of these molecules in the tumor tissues of the lean rats, which were detected in the obese rats, there was a significant increase in the expression of these molecules in the obese rats (Figure 3).
2.3. Immunohistochemistry of insulin signaling pathway and PI3K/GSK3β/β-catenin role in breast cancer development
IGF-1 and its receptor are essential for the correct growth and development of the mammary gland. The overexpression of these molecules triggers signaling processes that are essential for the growth and survival of cancer cells. Moreover, the IGF-1-mediated activation of PI3K/GSK/-catenin was investigated in order to study potential molecular processes that may contribute to the effect of obesity and impaired glucose tolerance on breast development and progression. The concentrations of the IGF-1/PI3K/GSK3/β-catenin signaling pathway components IGF-1, IGF-1R, pIRS-1, PI3 kinase, Akt, pAkt, GSK-3, pGSK-3, β-catenin, and VEGF were evaluated. It was observed that the expression of signaling molecules IGF-1, IGF-1R, pIRS-1, PI3 kinase, pAkt, pGSK-3, β-catenin, and VEGF was elevated in the breast tumor tissues of obese rats compared to lean rats, and that the expression of these signaling molecules was elevated in tumor tissue compared to respective control breast tissues (Figure 4 and Figure 5). The levels of total GSK-3 and Akt expression were reduced (Figure 4 and Figure 5). The expression of IGF-1 and IGF-1R molecules in the breast tumor tissues of obese rats with IGT was significantly greater than that in the breast tumor tissues of lean rats. In addition, the increased production of these molecules has stimulated the increased expression of downstream molecules, such as pIRS-1, PI3 kinase, pGSK-3, β-catenin, and VEGF, leading to the proliferation, survival, and metastasis of breast tissue. Interestingly the breast tumor tissues of obese and lean rats had higher expression levels of PI3K, pAKT, pGSK-3β, and β-catenin than normal breast tissues (Figure 6).
3. Discussion
Breast cancer, the most frequent cancer in women, is a major public health issue. Several studies have provided strong evidence for a positive association between obesity, diabetes/IGT and increased risk of breast cancer development as well as recurrent metastasis [29,30,31,32,33,34,35]. The combined effect of obesity and diabetes on breast cancer outcomes has been extensively reported in humans [20,21,22,36], and undiagnosed IGT is known to affect the survival of breast cancer patients [37]. Insulin/IGF signaling is reported to be dysregulated in obesity, diabetes, and breast cancer underscoring intricate overlaps in the underlying metabolic abnormalities and disease spectrum [38]. Hence, in this study we investigated the combined effects of obesity and IGT on breast cancer risk and probable mechanisms using a genetically obese rat model with IGT for chemically induced breast cancers. The results of the present study reinforce the tenet that obesity and IGT have the propensity to progress to breast tumor genesis.
In the present investigation, spontaneous mammary tumors did not develop in the untreated obese rats until 40 weeks of age. However, DMBA administration (40 mg/kg body weight) began producing breast tumors in the 9th week and reached 62% at 40 weeks. DMBA accelerated mammary gland neoplasia in obese rats. However, the same amount of DMBA in lean littermates caused breast cancers at 26 weeks, and only 21% of animals had tumors at 40 weeks. Obese rats developed breast tumors faster after receiving the same dose of DMBA. Researchers looked for several different kinds of signs of breast cancer and measured how much they were present in the breast tissue of both lean and obese rats that had been given DMBA to cause breast tumors. DMBA also accelerated mammary cancers in mutant obese Yellow mice [39] and Zuccher rats [40,41], compared to counter treatments. However, these studies have not explained how breast cancer accelerates in these models. Previous investigations found that diet-induced or mutant-induced obesity increased tumor susceptibility. We found similar results with obese rats with decreased glucose tolerance in this investigation. This is the first preclinical rat model with obesity and IGT that shows high breast cancer susceptibility after DMBA treatment. We believe that this can be a valuable preclinical animal model to analyse underlying molecular mechanisms associated with development of comorbidities encompassing obesity, IGT and breast cancer and a valuable tool to test putative preventive/therapeutic agents.
ER and PR are important biological markers that have a key role in cellular growth, proliferation, and differentiation. Measurement of the levels of these hallmarks of breast cancer is useful as a prognostic indicator and in determining the possibility of hormonal resistance in breast cancer and treatment plan [42]. In the present study, obese animals had higher levels of all these markers in their tumor tissues than lean rats. Obesity has been suggested to increase steroid hormone receptor expression with consequent progression and proliferation of breast cancer cells [43]. An association between obesity and PR positivity was observed in ER-positive tumors [44]. The poorer survival of ER-positive breast cancer patients could depend on the tumor PR status [45]. Obese rats with IGT had a greater incidence of breast cancer than lean rats due to elevated ER and PR expression. IGT obesity may increase the risk of hormone-responsive breast cancer.
Ki67 is the most commonly used proliferative marker in breast cancer. High Ki67 expression predicts poor prognosis [46]. Ki67 distinguishes breast cancer molecular subgroups. It was classified as luminal-A or luminal-B based on the Ki67 value [47]. DMBA-induced breast cancers of obese rats had greater Ki67 expression with ER and PR positivity than lean rat breast tumors. Breast cancer has been clinically verified using the Ki67 as a proliferative marker [48]. The tumor suppressor gene TP53 encodes p53. In normal cells, ubiquitylation and proteasome activity destroy the p53 protein, which has a short half-life [49]. Mutations in the p53 gene stabilize a protein post-transcriptionally, causing cell accumulation [50], 18–25% of initial breast tumors have p53 mutations [51]. The IHC-detected p53 expression in the breast cancer was associated with an aggressive, metastatic phenotype and awful outcomes [52,53]. Obese rats develop breast cancer faster than lean rats due to DMBA's aggressive tumor induction and increased p53 expression in breast tumor tissues.
The major contributing factors of obesity and T2D/IGT that influence the risk of cancer were increased levels of growth factors such as insulin, IGF-1, steroid and peptide hormones, and inflammatory markers [54]. Aberrant activation of IGF-1 signaling has been documented in breast cancer tissues [55,56,57]. IGF-1 binds to its cognate receptor to induce phosphorylation of IRS-1 and triggers a cascade of events that eventually results in breast cancer development, progression, and metastasis [58,59]. ER is known to enhance the expression and activation of IGF-1R [60,61]. Activation of IRS-1 has been reported in ER positive breast cancer [62]. Furthermore, the crosstalk between ER and IRS-1 increases the risk of breast cancer [55]. Although some studies on obese rodent animal models showed the association of obesity with increased breast cancer [63,64], the mechanism underlying the association between obesity and breast cancer has not been delineated. Here we demonstrate that administration of DMBA induced increased expression of IGF-1, IGF-1R, pIRS, and ER in the tumor tissues of obese rats with subsequent activation of downstream molecules in the signaling pathway that could promote tumor development.
Phosphorylation of IRS-1 stimulates PI3K/Akt signaling that plays a pivotal role in the cell proliferation, cell survival, migration, and differentiation [65]. Inappropriate activation of PI3K/Akt signaling has been reported in diverse malignancies including breast cancer [66,67,68]. Our findings indicate that in obese breast tumors, PI3K stimulates PDK1, which phosphorylates Akt kinase. Akt phosphorylates GSK3 at Ser9, inhibiting its activity and stabilizing and accumulating β-catenin, which induces cell proliferation (Figure 7). The aberrant expression of β-catenin was associated with adverse outcomes of breast cancer [69,70,71]. The lower expression of these signaling molecules in lean rat tumor tissues may not stimulate this signaling cascade, resulting in delayed breast tumor induction and low tumor incidence.
In conclusion, the WNIN/GR-Ob rats provide an ideal model to study how obesity and impaired glucose tolerance affect chronic disease progression, particularly breast cancer. DMBA-treated GR-Ob obese rats developed breast tumors earlier and at a higher rate than lean rats. Obesity and IGT enhance the IGF-1 response, which promotes cancer by blocking apoptosis and increasing cell proliferation. IGF-1-mediated activation of PI3K/Akt/GSK-3 signalling with β-catenin nuclear accumulation upregulates transcription factors of cell proliferation, resulting in higher tumor development and progression in obese rats, as shown by higher VEGF and Ki67 expression. These findings underscore the importance of understanding the relationship between obesity and breast cancer to develop effective strategies for prevention and treatment.
4. Materials and Methods
4.1. Animal grouping and housing
Obese mutant rats with characteristics of abnormal response to glucose load (IGT), hyperinsulinemia, hypertriglyceridemia, hypercholesterolemia, and hyperleptinaemia were used in the study. Littermate-lean rats were used as controls. A total of 32 female rats (16 lean rats and 16 obese rats) 60 days of age were used for the investigation. The animals were housed individually in polycarbonate cages and autoclaved paddy husk was used as bedding material. Twelve hours of light-dark photoperiodicity with standard lighting conditions were maintained in the experimental rooms. The temperature, relative humidity, and air changes were kept constant at 22±2◦C and 55±10%, 14-16 respectively. Both the lean and obese rats were allowed 3 days of acclimatization and subsequently divided into four groups of eight animals each based on their body weight. Lean rats that did not receive any treatment served as lean control, whereas lean rats administered with DMBA are used as lean tumor group. Untreated obese rats as obese control, while obese rats received DMBA administration are considered as obese tumor group. All the animals received sterile standard rodent chow (AIN93M) diet and water ad libitum. The study was reviewed and approved by the Institutional Animal Ethical Committee (P10F/IAEC/NIN/5/2018/GBP/WNIN GR-Ob). The experiment was conducted in the animal facility, ICMR-National Institute of Nutrition, Hyderabad, India in compliance with the guidelines prescribed by the Committee for the Purpose of Control and Supervision on Experiments on Animals (CPCSEA).
4.2. DMBA preparation and administration
Mammary carcinogenesis was induced by administration of DMBA (Sigma Aldrich, USA). DMBA was dissolved in refined sesame oil (20 mg/mL) and stirred slowly using a magnetic stirrer until complete dissolution. A single dose of solubilized DMBA suspension (40 mg/kg body weight) was oral administered to the animals at age 60 days. The control rats received an equal amount of sesame oil. Based on a pilot study, since a single dose of DMBA at 40 mg/kg body weight was found to induce mammary tumours in both lean and obese rats without causing any mortality, this dose was administered for the experimental study.
4.3. Measurements of tumor growth parameters
To determine the incidence and latency of tumor formation, rats were palpated at the thoracic and abdominal-inguinal mammary glands once a week starting from one week after the administration of DMBA. Tumor parameters like the percentage of tumor bearing animals per group (tumor incidence), the period from carcinogen administration to the appearance of the first tumor (latency period), the average tumor number per group (tumor frequency), and tumor size were measured. Tumor size was measured by recording the length and width of each tumor using a digital caliper in each group. The volume (V) of tumors was calculated according to the formula: V = π × S1 2× S2/12, where S1 and S2 are tumor diameters, assuming S1<S2 [72].
4.4. Gross necropsy
After completion of the feeding schedule (32 weeks after administration of DMBA), rats were fasted overnight and euthanized by CO2 inhalation. A gross necropsy of the rats was carried out to examine abnormalities. Rats were shaved at the breast regions; hair was removed and mammary tumors were excised and normal breast regions were excised from control rats which served as a control for further studies. The in situ examination was carried out, after opening the viscera and the major organs like the brain, heart, lungs, liver, spleen, kidneys, and pancreas were separated from the viscera and cleaned from fat, blotted on a filter paper, and weighed (Sartorius analytical balance with a 0.1 gm sensitivity). The breast tumor tissues and normal breast tissues along with the other organs were fixed in a fixative solution containing 10% formalin in sodium phosphate buffer at pH 7.4. These tissues were processed for histopathological and immunohistochemical analyses.
4.5. Hematoxylin & Eosin staining
For analysis of tumor morphology, a smaller representative portion was taken from the freshly collected samples after animal necroscopy. For the histopathological study, H&E staining was done for both breast tumors and normal breast tissues. Paraffin tissue sections of 4μm thickness were made using a microtome (Jinhua Yidi Medical Appliance Co., Ltd, China), followed by processing with an automatic tissue processor (Thermo Scientific, USA). Hematoxylin and Eosin staining was done using an autostainer (Sakura Tissue-Tek DRS 2000 automated slide stainer). The H&E stained slides were investigated for the detection of breast cancer metastatic stages, and images were acquired at 10x and 40x magnification with a Leica microscope. (Leica Microsystems, Germany).
4.6. Immunohistochemistry
The formalin-fixed breast tissues were embedded in paraffin, and transverse sections (4 μm) mounted in gelatin-coated slides. Immunohistochemical analysis was performed using a Vectastain Elite ABC kit (Vector Laboratories, USA) that exploits the Avidin-Biotin Complex (ABC) method. Deparaffinized sections were processed in 10 mM sodium citrate buffer (pH 6.0) and heated for 5 minutes (antigen retrieval step). After blocking with 3% horse serum provided with the kit, the primary antibody (1:500 dilutions) was added to the sections and allowed to incubate overnight at 4 °C. In the present study, the following primary antibodies were utilized: estrogen receptor (ER), progesterone receptor (PR), Ki-67, p53, insulin-like growth factor 1(IGF-1), insulin-like growth factor 1receptor (IGF-1R), phospho insulin receptor substrate 1 (pIRS-1), phosphoinositide 3 kinase (PI3K), protein kinase B (Akt), p-AktSer473, glycogen synthase kinase-3 beta (GSK-3β), pGSK-3βSer9, β-catenin and vascular endothelial growth factor (VEGF). All the antibodies were procured from Cell Signaling Technology (CST), USA. After primary antibody incubation, the sections were washed three times for 5 min in 20 mM phosphate buffer saline (PBS pH 7.4). After washing with PBS, slides were incubated for 1 h at room temperature with a biotinylated secondary antibody (1:500 dilutions) solution and DAB was used as the chromogen. The negative controls were run simultaneously with the omission of the primary antibody. After staining, the sections were counterstained with hematoxylin. The sections were then dehydrated through ethanol and xylene before coverslips with Paramount. The DAB staining was visualized in the bright field using a Leica microscope (Leica microsystems, Germany) at 10x and 40 x magnifications.
4.7. Immunoblotting
The total proteins were extracted from breast tissues by homogenizing with 100 mM Tris-HCl buffer, pH 7.4 on ice with mortar and pestle, and the homogenate was centrifuged at 12,000 ×g for 30 min. The protein concentrations were estimated by the method of Lowry et al. (1951). Equal amounts of protein from tissue extracts were separated using 12% SDS-PAGE and transferred to 0.2 µm nitrocellulose blotting membrane (GE Healthcare Life science, USA) at 80 V for 1.5 h using a Bio-Rad transblot apparatus. To determine the uniformity of loading and transfer, membranes were stained with Ponceau S. The membrane was blocked for 2 h in phosphate buffer saline containing 20 mM sodium phosphate buffer, pH 7.4, and 5% nonfat dry milk powder at room temperature. Immunoblotting was performed by incubating the blot at 4oC overnight with primary antibodies of IGF-1, phospho-IRS-1, PI3K, Akt, phospho-Akt, GSK-3β, phospho-GSK-3β, and β-catenin (1:1000 dilution). After overnight primary antibody incubation, the membrane was washed 3 times with PBS and then the blot was incubated for 2 h with HRP-tagged anti-rabbit/anti-mouse respective secondary antibody with a dilution of 1:10,000. Equal loading of the protein samples was assessed by probing the membrane with a 1:1000 dilution of the β-tubulin loading control antibody. The immunoblots were developed with enhanced chemiluminescence detection reagents (Bio-Rad Laboratories, USA). The images were analyzed and quantified using Image J software [73].
4.8. Statistical analysis
All statistical analysis was performed using GraphPad Prism software 8.0 version. The data are expressed as mean ± standard mean error (SEM). P values were determined using one-way ANOVA followed by Tukey's multiple comparison tests.
Author Contributions
Conceptualization, GBR and NS.; methodology, GBR, NS, PRK; formal analysis, PRK, RK, NS and GBR.; investigation, PRK, DE, URA, UKP; resources, GBR and NS.; data curation, PRK, RK, NS and GBR; writing—original draft preparation, PRK; writing—review and editing, PRK, RK, NS and GBR; visualization, PRK, UKP; supervision, GBR; project administration, GBR; funding acquisition, NS and GBR. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Science and Engineering Research Board (EMR/2016/001984), Government of India (GBR and NS).
Institutional Review Board Statement
The study was reviewed and approved by the Institutional Animal Ethical Committee (P10F/IAEC/NIN/5/2018/GBP/WNIN GR-Ob). The experiment was conducted in the animal facility of ICMR-National Institute of Nutrition, Hyderabad, India in compliance with the guidelines prescribed by the Committee for the Purpose of Control and Supervision on Experiments on Animals (CPCSEA).
Data Availability Statement
The data presented in this study are available with corresponding author upon request.
Acknowledgments
URA is supported by National Postdoctoral Fellowship from the Science and Engineering Research Board, Government of India. The authors are grateful to Dr MV Surekha, Department of Pathology, National Institute of Nutrition for the help with histological evaluation of tissue sections.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sung, H.; Ferlay, J.; Siegel, R. L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. , Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer. J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.; Chun, M.; Oh, Y. T.; Noh, O. K.; Kim, L. , Metabolic Comorbidities and Medical Institution Utilization Among Breast Cancer Survivors: A National Population-based Study. Korean J. Intern. Med. 2020, 35, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Buono, G.; Crispo, A.; Giuliano, M.; De Angelis, C.; Schettini, F.; Forestieri, V.; Lauria, R.; De Laurentiis, M.; De Placido, P.; Rea, C. G.; Pacilio, C.; Esposito, E.; Grimaldi, M.; Nocerino, F.; Porciello, G.; Giudice, A.; Amore, A.; Minopoli, A.; Botti, G.; De Placido, S.; Trivedi, M. V.; Arpino, G. , Metabolic Syndrome and Early-Stage Breast Cancer Outcome: Results from a Prospective Observational Study. Breast Cancer Res. Treat. 2020, 182, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Kennard, K.; Buckley, M. E.; Sizer, L. M.; Larson, S.; Carter, W. B.; Frazier, T. G.; Carp, N. Z. Metabolic Syndrome: Does This Influence Breast Cancer Outcomes in the Triple-Negative Population? Breast Cancer Res. Treat. 2021, 186, 53–63. [Google Scholar] [CrossRef] [PubMed]
- https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight.
- https://www.worldobesity.org/resources/resource-library/world-obesity-atlas-2022.
- https://diabetesatlas.org/atlas/tenth-edition/.
- Wilson, M. L. , Prediabetes: Beyond the Borderline. Nurs. Clin. North. Am. 2017, 52, 665–677. [Google Scholar] [CrossRef]
- Ennis, C. S.; Llevenes, P.; Qiu, Y.; Dries, R.; Denis, G. V. The Crosstalk Within the Breast Tumor Microenvironment in Type II Diabetes: Implications for Cancer Disparities. Front. Endocrinol. 2022, 13, 1044670. [Google Scholar] [CrossRef]
- Lu, Y.; Hajjar, A.; Cryns, V. L.; Trentham-Dietz, A.; Gangnon, R. E.; Heckman-Stoddard, B. M.; Alagoz, O. Breast Cancer Risk for Women with Diabetes and the Impact of Metformin: A Meta-analysis. Cancer Med. 2022. [Google Scholar] [CrossRef]
- Luís, C.; Dias, J.; Firmino-Machado, J.; Fernandes, R.; Pereira, D.; Baylina, P.; Fernandes, R.; Soares, R. A Retrospective Study in Tumour Characteristics and Clinical Outcomes of Overweight and Obese Women with Breast Cancer. Breast Cancer Res. Treat. 2023, 198, 89–101. [Google Scholar] [CrossRef]
- Pati, S.; Irfan, W.; Jameel, A.; Ahmed, S.; Shahid, R.K. Obesity and Cancer: A Current Overview of Epidemiology, Pathogenesis, Outcomes, and Management. Cancers 2023, 15. [Google Scholar] [CrossRef]
- Lee, J.; Kim, H.; Bae, S. J.; Ji, J. H.; Lee, J. W.; Son, B. H.; Ahn, S. H.; Jeong, J.; Lee, S. B.; Ahn, S. G. Association of Body Mass Index With 21-Gene Recurrence Score Among Women with Estrogen Receptor-Positive, ERBB2-Negative Breast Cancer. JAMA Netw Open. 2022, 5, e2243935. [Google Scholar] [CrossRef]
- Calip, G. S.; Yu, O.; Boudreau, D. M.; Shao, H.; Oratz, R.; Richardson, S. B.; Gold, H. T. Diabetes and Differences in Detection of Incident Invasive Breast Cancer. Cancer Causes Control. 2019, 30, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Arce, L.; Robles-Rodríguez, N.; Fernández-Feito, A.; Llaneza-Folgueras, A.; Encinas-Muñiz, A. I.; Lana, A. Type 2 Diabetes and all-cause mortality among Spanish women with breast cancer. Cancer Causes Control. 2022, 33, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Gill, A. A.; Zahm, S. H.; Jatoi, I.; Shriver, C. D.; McGlynn, K. A.; Zhu, K. Diabetes and Overall Survival among Breast Cancer Patients in the U.S. Military Health System. Cancer Epidemiol. Biomarkers Prev. 2018, 27, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Jacob, L.; Kostev, K.; Rathmann, W.; Kalder, M. Impact of Metformin on Metastases in Patients with Breast Cancer and Type 2 Diabetes. J. Diabetes Complications. 2016, 30, 1056–1059. [Google Scholar] [CrossRef] [PubMed]
- Crispo, A.; Augustin, L. S.; Grimaldi, M.; Nocerino, F.; Giudice, A.; Cavalcanti, E.; Di Bonito, M.; Botti, G.; De Laurentiis, M.; Rinaldo, M.; Esposito, E.; Riccardi, G.; Amore, A.; Libra, M.; Ciliberto, G.; Jenkins, D. J.; Montella, M. Risk Differences Between Prediabetes and Diabetes According to Breast Cancer Molecular Subtypes. J. Cell Physiol. 2017, 232, 1144–1150. [Google Scholar] [CrossRef]
- Bernard, J. J.; Wellberg, E. A. The Tumor Promotional Role of Adipocytes in the Breast Cancer Microenvironment and Macroenvironment. Am. J. Pathol. 2021, 191, 1342–1352. [Google Scholar] [CrossRef]
- Maskarinec, G.; Jacobs, S.; Park, S. Y.; Haiman, C. A.; Setiawan, V. W.; Wilkens, L. R.; Le Marchand, L. Type II Diabetes, Obesity, and Breast Cancer Risk: The Multiethnic Cohort. Cancer Epidemiol. Biomarkers Prev. 2017, 26, 854–861. [Google Scholar] [CrossRef]
- Maskarinec, G.; Sadakane, A.; Sugiyama, H.; Brenner, A.; Tatsukawa, Y.; Grant, E. Type 2 Diabetes, Obesity, and Breast Cancer Risk Among Japanese Women of the Atomic Bomb Survivor Cohort. Cancer Epidemiol. 2019, 60, 179–184. [Google Scholar] [CrossRef]
- Maskarinec, G.; Haraldsdóttir, Á.; Einarsdóttir, K.; Aspelund, T.; Tryggvadóttir, L.; Harris, T. B.; Gudnason, V.; Torfadóttir, J. E. Type 2 Diabetes and Obesity in Midlife and Breast Cancer Risk in the Reykjavik Cohort. Cancer Causes Control. 2019, 30, 1057–1065. [Google Scholar] [CrossRef]
- McTiernan, A. Diet and Prognosis in Women with Breast Cancer. Cancer Epidemiology, Biomarkers & Prevention: A Publication of the American Association for Cancer Research, Cosponsored by the American Society of Preventive Oncology. 2021, 30, 252–254. [Google Scholar]
- Huang, Y.; Cai, X.; Qiu, M.; Chen, P.; Tang, H.; Hu, Y.; Huang, Y. Prediabetes and the risk of cancer: A meta-analysis. Diabetologia. 2014, 57, 2261–2269. [Google Scholar] [CrossRef] [PubMed]
- Luque, R. M.; López-Sánchez, L. M.; Villa-Osaba, A.; Luque, I. M.; Santos-Romero, A. L.; Yubero-Serrano, E. M.; Cara-García, M.; Álvarez-Benito, M.; López-Mirand, A. J.; Gahete, M. D.; Castaño, J. P. Breast Cancer is Associated to Impaired Glucose/Insulin Homeostasis in Premenopausal Obese/Overweight Patients. Oncotarget. 2017, 8, 81462–81474. [Google Scholar] [CrossRef] [PubMed]
- Cohen, D. H.; LeRoith, D. Obesity, Type 2 Diabetes, and Cancer: The Insulin and IGF Connection. Endocr. Relat. Cancer. 2012, 19, F27–F45. [Google Scholar] [CrossRef]
- Harishankar, N.; Vajreswari, A.; Giridharan, N. V. WNIN/GR-Ob - An Insulin-Resistant Obese Rat Model from Inbred WNIN Strain. Indian J. Med. Res. 2011, 134, 320–329. [Google Scholar] [PubMed]
- Godisela, K. K.; Reddy, S. S.; Kumar, C. U.; Saravanan, N.; Reddy, P. Y.; Jablonski, M. M.; Ayyagari, R.; Reddy, G. B. Impact of Obesity with Impaired Glucose Tolerance on Retinal Degeneration in a Rat Model of Metabolic Syndrome. Mol. Vis. 2017, 23, 263–274. [Google Scholar] [PubMed]
- Schlesinger, S.; Neuenschwander, M.; Barbaresko, J.; Lang, A.; Maalmi, H.; Rathmann, W.; Roden, M.; Herder, C. Prediabetes and Risk of Mortality, Diabetes-Related Complications and Comorbidities: Umbrella Review of Meta-Analyses of Prospective Studies. Diabetologia. 2022, 65, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, N. M.; Jaradat, S. K.; Alhusban, A.; Tahaineh, L. Glycosylated Hemoglobin A1c Is Associated with Anthropometric Measurements and Tumor Characteristics in Breast Cancer Patients. Int. J. Womens Health. 2020, 12, 139–149. [Google Scholar] [CrossRef]
- Cao, Y.; Xia, B.; Zhang, Z.; Hu, D.; Huang, X.; Yuan, J.; Li, F. Association of Body Fat Distribution and Risk of Breast Cancer in Pre- and Postmenopausal Women. Obesity Facts. 2023. [Google Scholar] [CrossRef]
- Kang, C.; LeRoith, D.; Gallagher, E. J. Diabetes, Obesity, and Breast Cancer. Endocrinology. 2018, 159, 3801–3812. [Google Scholar] [CrossRef]
- Belardi, V.; Gallagher, E. J.; Novosyadlyy, R.; LeRoith, D. Insulin and IGFs in Obesity-Related Breast Cancer. J. Mammary Gland Biol. Neoplasia. 2013, 18, 277–289. [Google Scholar] [CrossRef]
- Ecker, B. L.; Lee, J. Y.; Sterner, C. J.; Solomon, A. C.; Pant, D. K.; Shen, F.; Peraza, J.; Vaught, L.; Mahendra, S.; Belka, G. K.; Pan, T. C.; Schmitz, K. H.; Chodosh, L. A. Impact of Obesity on Breast Cancer Recurrence and Minimal Residual Disease. Breast Cancer Res. 2019, 21, 41. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S. L.; Cahyono, R.; Prabowo, D.; Avanti, W. S.; Choridah, L.; Dwianingsih, E. K.; Harahap, W. A. Aryandono, T. Metabolic Comorbidities and the Association with Risks of Recurrent Metastatic Disease in Breast Cancer Survivors. BMC Cancer. 2021, 21, 590. [Google Scholar] [CrossRef] [PubMed]
- Lopez, R.; Agullo, P.; Lakshmanaswamy, R. Links between Obesity, Diabetes and Ethnic Disparities in Breast Cancer among Hispanic Populations. Obes. Rev. 2013, 14, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Monzavi-Karbassi, B.; Gentry, R.; Kaur, V.; Siegel, E. R.; Jousheghany, F.; Medarametla, S.; Fuhrman, B. J.; Safar, A. M.; Hutchins, L. F.; Kieber-Emmons, T. Pre-Diagnosis Blood Glucose and Prognosis in Women with Breast Cancer. Cancer Metabolism. 2016, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Vella, V.; Lappano, R.; Bonavita, E.; Maggiolini, M.; Clarke, R. B.; Belfiore, A.; De Francesco, E. M. Insulin/IGF Axis and the Receptor for Advanced Glycation End Products: Role in Meta-Inflammation and Potential in Cancer Therapy. Endocr. Rev. 2023. [Google Scholar] [CrossRef]
- Wolff, G. L.; Kodell, R. L.; Cameron, A. M.; Medina, D. Accelerated Appearance of Chemically Induced Mammary Carcinomas in Obese Yellow (A vy /A) (BALB/c X VY) F1 Hybrid Mice. J. Toxicol. Environ. Health. 1982, 10, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Hakkak, R.; Holley, A. W.; Macleod, S. L.; Simpson, P. M.; Fuchs, G. J.; Jo, C. H.; Kieber-Emmons, T.; Korourian, S. Obesity Promotes 7,12-dimethylbenz(a)anthracene-induced Mammary Tumor Development in Female Zucker Rats. Breast Cancer Res. 2005, 7, R627–R633. [Google Scholar] [CrossRef]
- Hakkak, R.; MacLeod, S.; Shaaf, S.; Holley, A. W.; Simpson, P.; Fuchs, G.; Jo, C. H.; Kieber-Emmons, T.; Korourian, S. Obesity Increases the Incidence of 7,12-dimethylbenz(a)anthracene-Induced Mammary Tumors in an Ovariectomized Zucker Rat model. Int. J. Oncol. 2007, 30, 557–563. [Google Scholar] [CrossRef]
- Kabel, A. M. Tumor Markers of Breast Cancer: New prospectives. J. Oncol. Sci. 2017, 3, 5–11. [Google Scholar] [CrossRef]
- Bhardwaj, P.; Au, C. C.; Benito-Martin, A.; Ladumor, H.; Oshchepkova, S.; Moges, R.; Brown, K. A. Estrogens and Breast Cancer: Mechanisms Involved in Obesity-Related Development, Growth and Progression. J. Steroid Biochem. Mol. Biol. 2019, 189, 161–170. [Google Scholar] [CrossRef]
- Chauhan, R.; Trivedi, V.; Rani, R.; Singh, U. A Comparative Analysis of Body Mass Index with Estrogen Receptor, Progesterone Receptor and Human Epidermal Growth Factor Receptor 2 Status in Pre- and Postmenopausal Breast Cancer Patients. J. Midlife Health. 2020, 11, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Oudanonh, T.; Nabi, H.; Ennour-Idrissi, K.; Lemieux, J.; Diorio, C. Progesterone Receptor Status Modifies the Association Between Body Mass Index and Prognosis in Women Diagnosed with Estrogen Receptor Positive Breast Cancer. Int. J. Cancer. 2020, 146, 2736–2745. [Google Scholar] [CrossRef] [PubMed]
- de Azambuja, E.; Cardoso, F.; de Castro, G.; Colozza, M.; Mano, M. S.; Durbecq, V.; Sotiriou, C.; Larsimont, D.; Piccart-Gebhart, M. J.; Paesmans, M. Ki-67 as Prognostic Marker in Early Breast Cancer: A Meta-analysis of Published Studies Involving 12 155 Patients. Br. J. Cancer. 2007, 96, 1504–1513. [Google Scholar] [CrossRef] [PubMed]
- Inic, Z.; Zegarac, M.; Inic, M.; Markovic, I.; Kozomara, Z.; Djurisic, I.; Inic, I.; Pupic, G.; Jancic, S. Difference between Luminal A and Luminal B Subtypes According to Ki-67, Tumor Size, and Progesterone Receptor Negativity Providing Prognostic Information. Clinical Medicine Insights. Oncology. 2014, 8, 107–111. [Google Scholar] [CrossRef]
- Skjervold, A. H.; Pettersen, H. S.; Valla, M.; Opdahl, S.; Bofin, A. M. Visual and Digital Assessment of Ki-67 in Breast Cancer Tissue - A Comparison of Methods. Diagn. Pathol. 2022, 17, 45. [Google Scholar] [CrossRef]
- Lukashchuk, N.; Vousden, K. H. Ubiquitination and Degradation of Mutant p53. Mol. Cell. Biol. 2007, 27, 8284–8295. [Google Scholar] [CrossRef]
- Yang, P.; Du, C. W.; Kwan, M.; Liang, S. X.; Zhang, G. J. The Impact of p53 in Predicting Clinical Outcome of Breast Cancer Patients with Visceral Metastasis. Sci. Rep. 2013, 3, 2246. [Google Scholar] [CrossRef]
- Alsner, J.; Yilmaz, M.; Guldberg, P.; Hansen, L. L.; Overgaard, J. Heterogeneity in the Clinical Phenotype of TP53 Mutations in Breast Cancer Patients. Breast Cancer Res. 2000, 2, P4.04. [Google Scholar] [CrossRef]
- Jasar, D.; Smichkoska, S.; Kubelka, K.; Filipovski, V.; Petrushevska, G. Expression of p53 Protein Product in Triple Negative Breast Cancers and Relation with Clinical and Histopathological Parameters. Prilozi (Makedonska akademija na naukite i umetnostite. Oddelenie za medicinski nauki). 2015, 36, 69–79. [Google Scholar]
- Bae, S. Y.; Nam, S. J.; Jung, Y.; Lee, S. B.; Park, B.-W.; Lim, W.; Jung, S. H.; Yang, H. W.; Jung, S. P. Differences in Prognosis and Efficacy of Chemotherapy by p53 Expression in Triple-Negative Breast Cancer. Breast Cancer Res. Treat. 2018, 172, 437–444. [Google Scholar] [CrossRef]
- Zhong, W.; Wang, X.; Wang, Y.; Sun, G.; Zhang, J.; Li, Z. Obesity and Endocrine-Related Cancer: The Important Role of IGF-1. Front. Endocrinol. 2023, 14, 1093257. [Google Scholar] [CrossRef] [PubMed]
- Kaaks, R.; Johnson, T.; Tikk, K.; Sookthai, D.; Tjønneland, A.; Roswall, N.; Overvad, K.; Clavel-Chapelon, F.; Boutron-Ruault, M. C.; Dossus, L.; et al. Insulin-Like Growth Factor I and Risk of Breast Cancer by Age and Hormone Receptor Status-A Prospective Study within the EPIC Cohort. Int. J. Cancer. 2014, 134, 2683–2690. [Google Scholar] [CrossRef] [PubMed]
- Murphy, N.; Knuppel, A.; Papadimitriou, N.; Martin, R. M.; Tsilidis, K. K.; Smith-Byrne, K.; Fensom, G.; Perez-Cornago, A.; Travis, R. C.; Key, T. J.; Gunter, M. J. Insulin-Like Growth Factor-1, Insulin-Like Growth Factor-Binding Protein-3, and Breast Cancer Risk: Observational and Mendelian Randomization Analyses with ∼430 000 Women. Ann. Oncol. 2020, 31, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Farabaugh, S. M.; Boone, D. N.; Lee, A. V. Role of IGF1R in Breast Cancer Subtypes, Stemness, and Lineage Differentiation. Front. Endocrinol. 2015, 6, 59. [Google Scholar] [CrossRef] [PubMed]
- Christopoulos, P. F.; Msaouel, P.; Koutsilieris, M. The Role of the Insulin-Like Growth Factor-1 System in Breast Cancer. Mol. Cancer. 2015, 14, 43. [Google Scholar] [CrossRef] [PubMed]
- Monson, K. R.; Goldberg, M.; Wu, H.-C.; Santella, R. M.; Chung, W. K.; Terry, M. B. Circulating Growth Factor Concentrations and Breast Cancer Risk: A Nested Case-Control Study of IGF-1, IGFBP-3, and Breast Cancer in a Family-Based Cohort. Breast Cancer Res. 2020, 22, 109. [Google Scholar] [CrossRef] [PubMed]
- Kahlert, S.; Nuedling, S.; van Eickels, M.; Vetter, H.; Meyer, R.; Grohe, C. Estrogen Receptor Alpha Rapidly Activates the IGF-1 Receptor Pathway. J. Biol. Chem. 2000, 275, 18447–18453. [Google Scholar] [CrossRef] [PubMed]
- Yoshimaru, T.; Komatsu, M.; Miyoshi, Y.; Honda, J.; Sasa, M.; Katagiri, T. Therapeutic Advances in BIG3-PHB2 Inhibition Targeting the Crosstalk Between Estrogen and Growth Factors in Breast Cancer. Cancer Sci. 2015, 106, 550–558. [Google Scholar] [CrossRef]
- Fox, E. M.; Kuba, M. G.; Miller, T. W.; Davies, B. R.; Arteaga, C. L. Autocrine IGF-I/insulin Receptor Axis Compensates for Inhibition of AKT in ER-Positive Breast Cancer Cells with Resistance to Estrogen Deprivation. Breast Cancer Res. 2013, 15, R55. [Google Scholar] [CrossRef]
- Giles, E. D.; Wellberg, E. A. Preclinical Models to Study Obesity and Breast Cancer in Females: Considerations, Caveats, and Tools. J. Mammary Gland Biol. Neoplasia. 2020, 25, 237–253. [Google Scholar] [CrossRef]
- Liu, C.; Wu, P.; Zhang, A.; Mao, X. Advances in Rodent Models for Breast Cancer Formation, Progression, and Therapeutic Testing. Front. Oncol. 2021, 11, 593337. [Google Scholar] [CrossRef] [PubMed]
- Ekyalongo, R. C.; Yee, D. Revisiting the IGF-1R as a Breast Cancer Target. NPJ Precis. Oncol. 2017, 1, 14. [Google Scholar] [CrossRef]
- Lien, E. C.; Dibble, C. C.; Toker, A. PI3K Signaling in Cancer: Beyond AKT. Curr. Opin. Cell Biol. 2017, 45, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Sefidbakht, S.; Saeedipour, H.; Saffar, H.; Mirzaian, E. Determination of β-catenin Expression in Breast Cancer and Its Relationship with Clinicopathologic Parameters. Asian Pac. J. Cancer Prev. 2021, 22, 3493–3498. [Google Scholar]
- Yuan, Y.; Long, H.; Zhou, Z.; Fu, Y.; Jiang, B. PI3K-AKT-Targeting Breast Cancer Treatments: Natural Products and Synthetic Compounds. Biomolecules 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, S.; Sun, Y.; Li, L. The Expression of β-catenin in Different Subtypes of Breast Cancer and its Clinical Significance. Tumour Biol. 2014, 35, 7693–7698. [Google Scholar] [CrossRef]
- Xu, J.; Prosperi, J. R.; Choudhury, N.; Olopade, O. I.; Goss, K. H. β-Catenin is Required for the Tumorigenic Behavior of Triple-Negative Breast Cancer Cells. PLoS ONE. 2015, 10, e0117097. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, M.; Xu, F.; Jiang, S. Wnt Signaling in Breast Cancer: Biological Mechanisms, Challenges and Opportunities. Mol. Cancer. 2020, 19, 165. [Google Scholar] [CrossRef]
- Bojková, B.; Kajo, K.; Kisková, T.; Kubatka, P.; Žúbor, P.; Solár, P.; Péč, M.; Adamkov, M. Metformin and Melatonin Inhibit DMBA-Induced Mammary Tumorigenesis in Rats Fed a High-Fat Diet. Anticancer Drugs. 2018, 29, 128–135. [Google Scholar] [CrossRef]
Figure 1.
Induction and development of tumors in lean and obese rats upon DMBA oral administration (40 mg/kg body weight). Panel A: Graphical representation of tumor induction and development in lean and obese rats with DMBA administration Panel B: Actual images of lean and obese rats with and without tumors. Panel C: Quantitative representation of percentage of tumor bearing animals with duration.
Figure 1.
Induction and development of tumors in lean and obese rats upon DMBA oral administration (40 mg/kg body weight). Panel A: Graphical representation of tumor induction and development in lean and obese rats with DMBA administration Panel B: Actual images of lean and obese rats with and without tumors. Panel C: Quantitative representation of percentage of tumor bearing animals with duration.
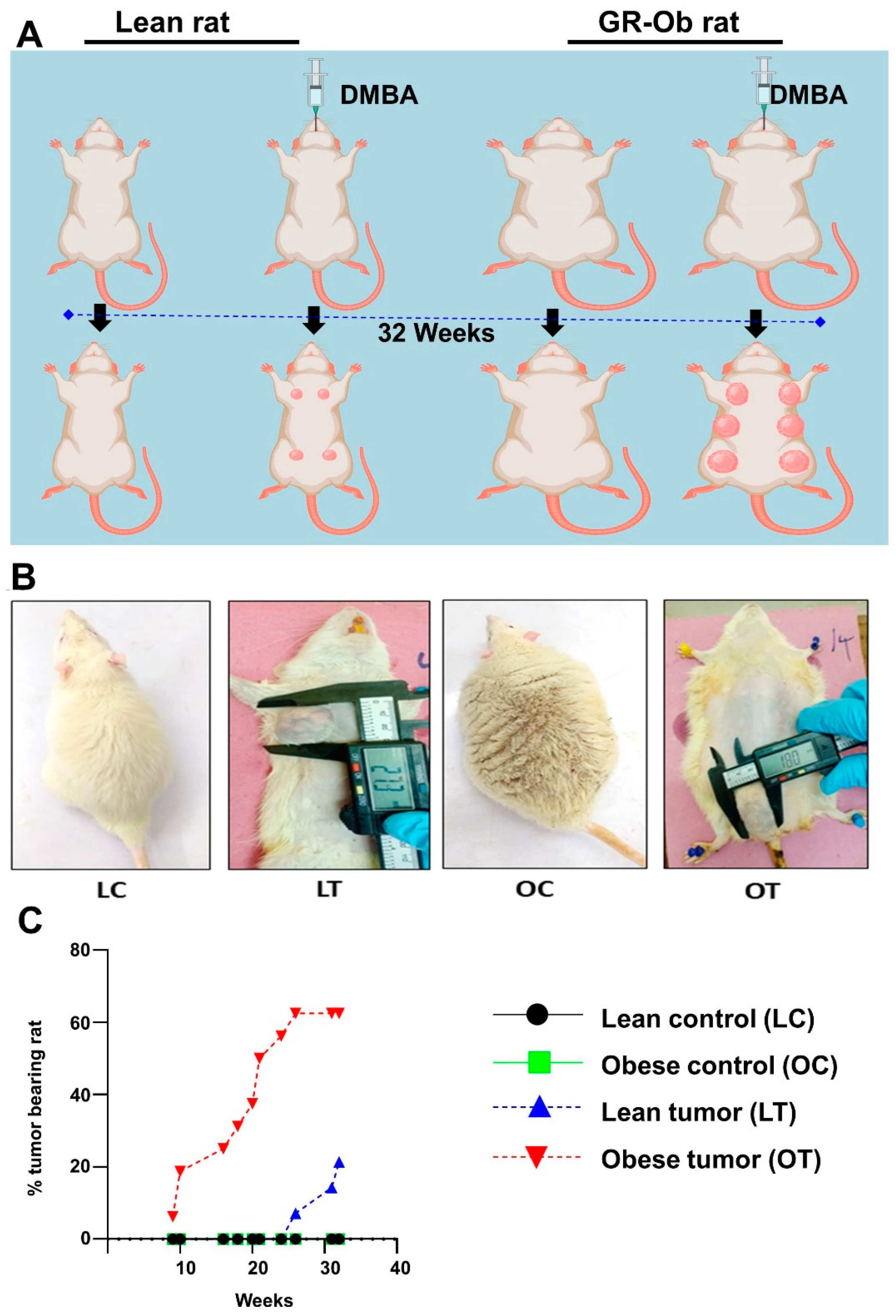
Figure 2.
Histology (H&E staining) of breast tissues of lean control (A), lean tumor (B), obese control (C) and obese tumor (D). Tumor mass both lean and obese rat breast tissues have lobules with pleomorphic epithelial cells with distinct variation in cell size and shape.
Figure 2.
Histology (H&E staining) of breast tissues of lean control (A), lean tumor (B), obese control (C) and obese tumor (D). Tumor mass both lean and obese rat breast tissues have lobules with pleomorphic epithelial cells with distinct variation in cell size and shape.
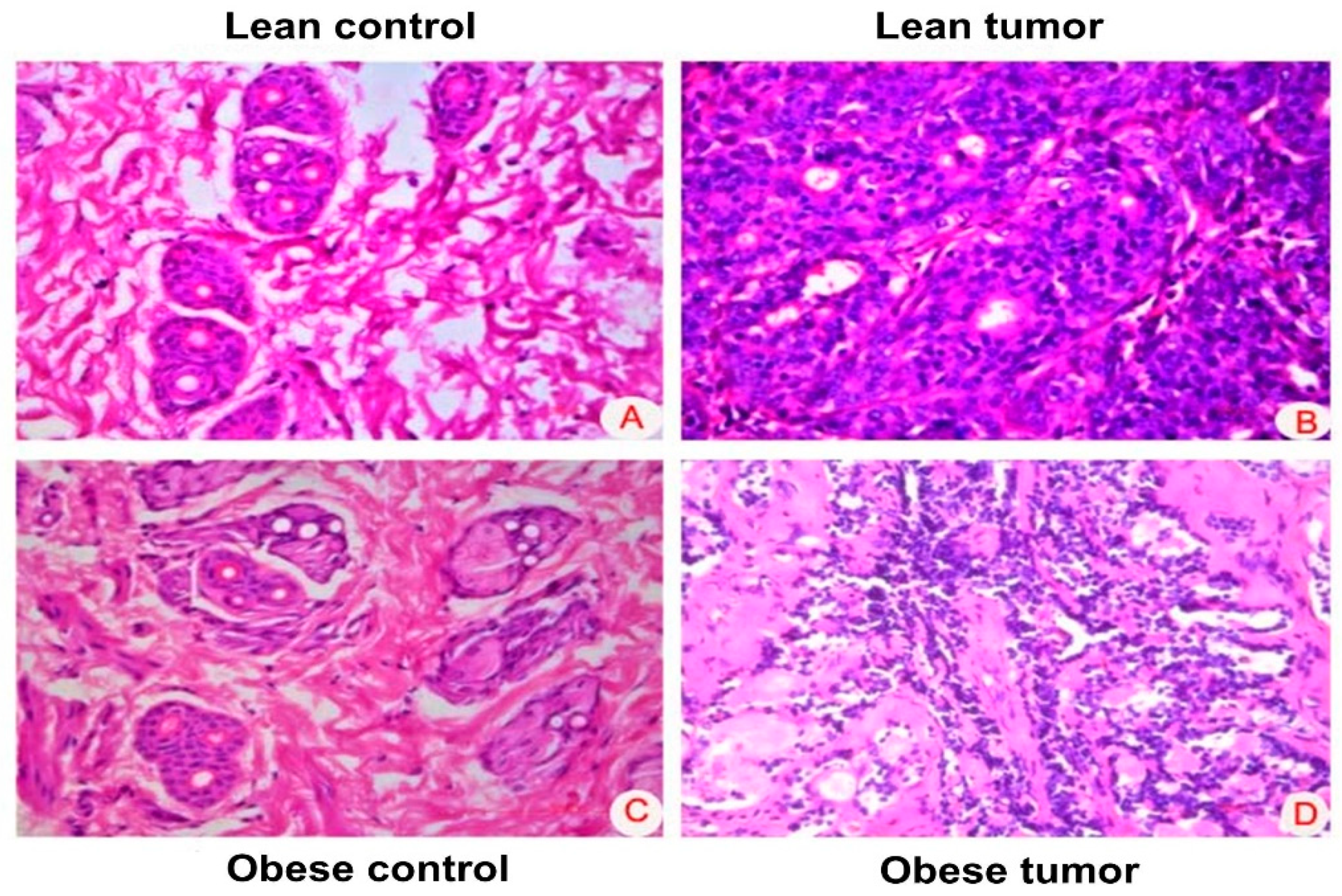
Figure 3.
Expression of oncogenic markers in rat mammary gland. Representative images of immunohistochemical staining of ER, PR, Ki67 and p53 in breast tissues of lean control, lean tumor, obese control and obese tumor.
Figure 3.
Expression of oncogenic markers in rat mammary gland. Representative images of immunohistochemical staining of ER, PR, Ki67 and p53 in breast tissues of lean control, lean tumor, obese control and obese tumor.
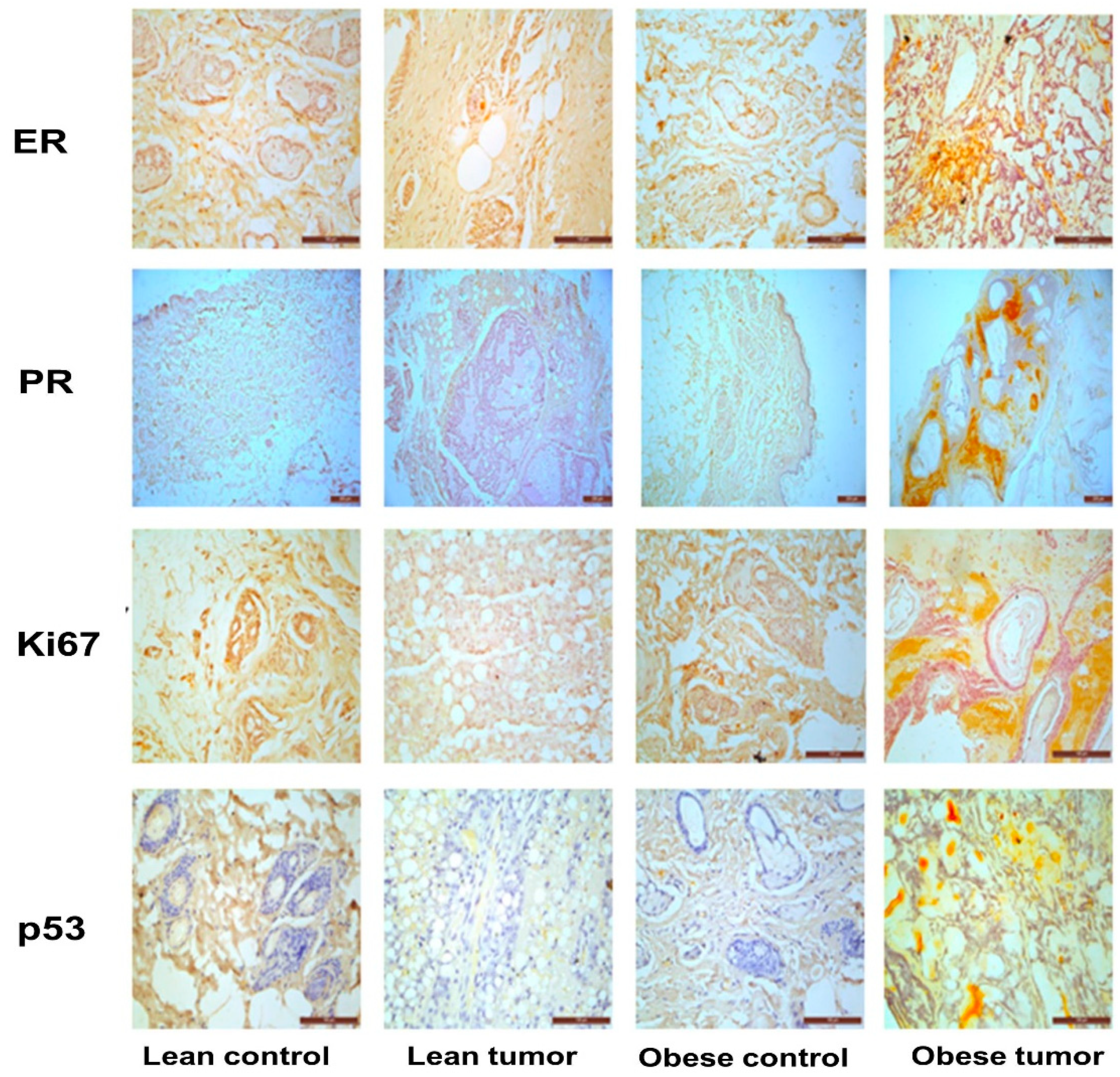
Figure 4.
Expression of insulin and PI3K/Akt signaling pathway molecules by immunostaining. Representative images of immunohistochemical staining of IGF-1, IGF-1R, pIRS, PI3K, Akt and pAkt in breast tissues of lean control, lean tumor, obese control and obese tumor.
Figure 4.
Expression of insulin and PI3K/Akt signaling pathway molecules by immunostaining. Representative images of immunohistochemical staining of IGF-1, IGF-1R, pIRS, PI3K, Akt and pAkt in breast tissues of lean control, lean tumor, obese control and obese tumor.
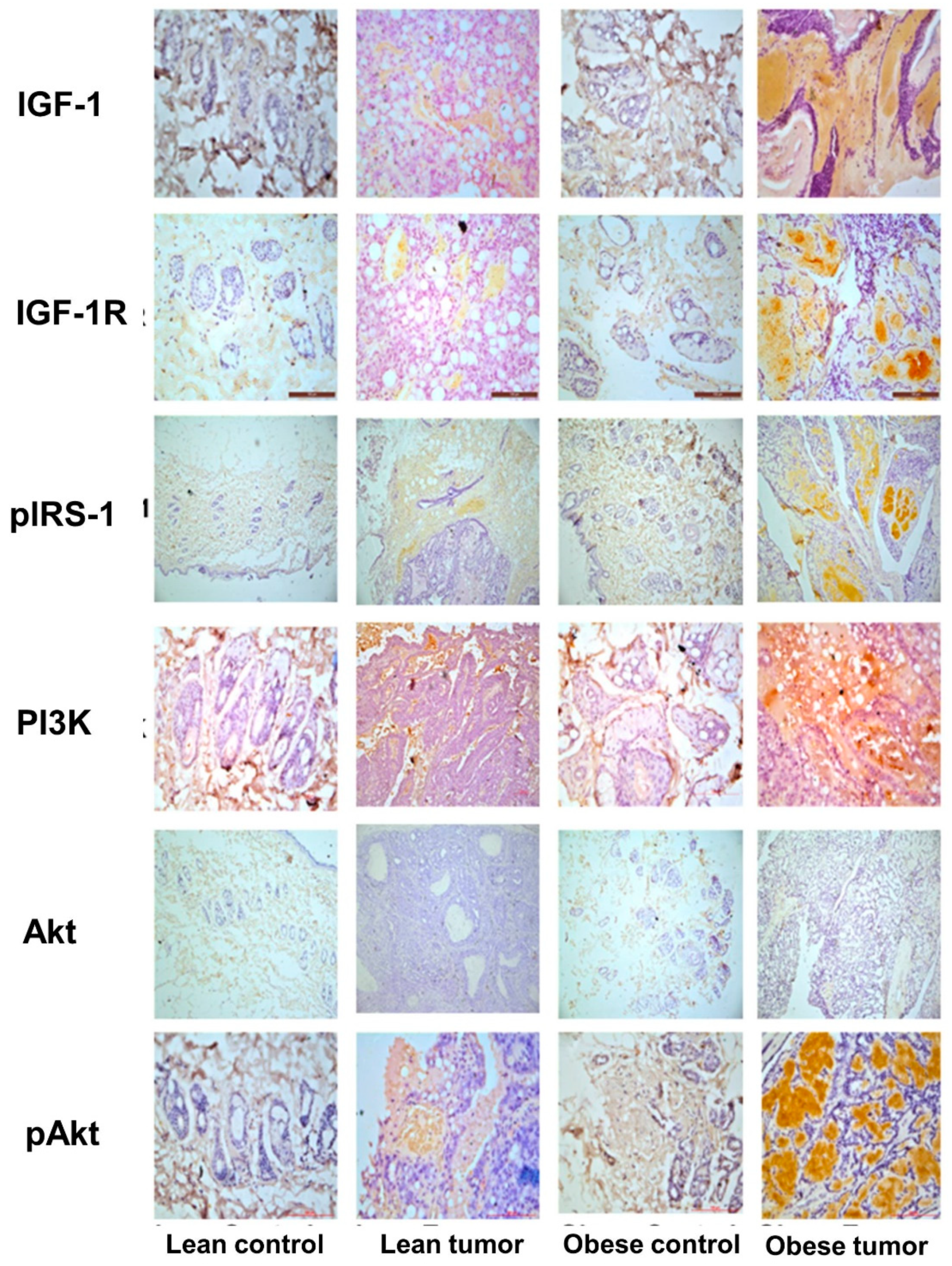
Figure 5.
Expression of GSK/catenin signaling pathway molecules by immunostatining. Representative images of immunohistochemical staining of GSK-3β, p GSK-3β, β-catenin and VEGF in breast tissues of lean control, lean tumor, obese control and obese tumor.
Figure 5.
Expression of GSK/catenin signaling pathway molecules by immunostatining. Representative images of immunohistochemical staining of GSK-3β, p GSK-3β, β-catenin and VEGF in breast tissues of lean control, lean tumor, obese control and obese tumor.
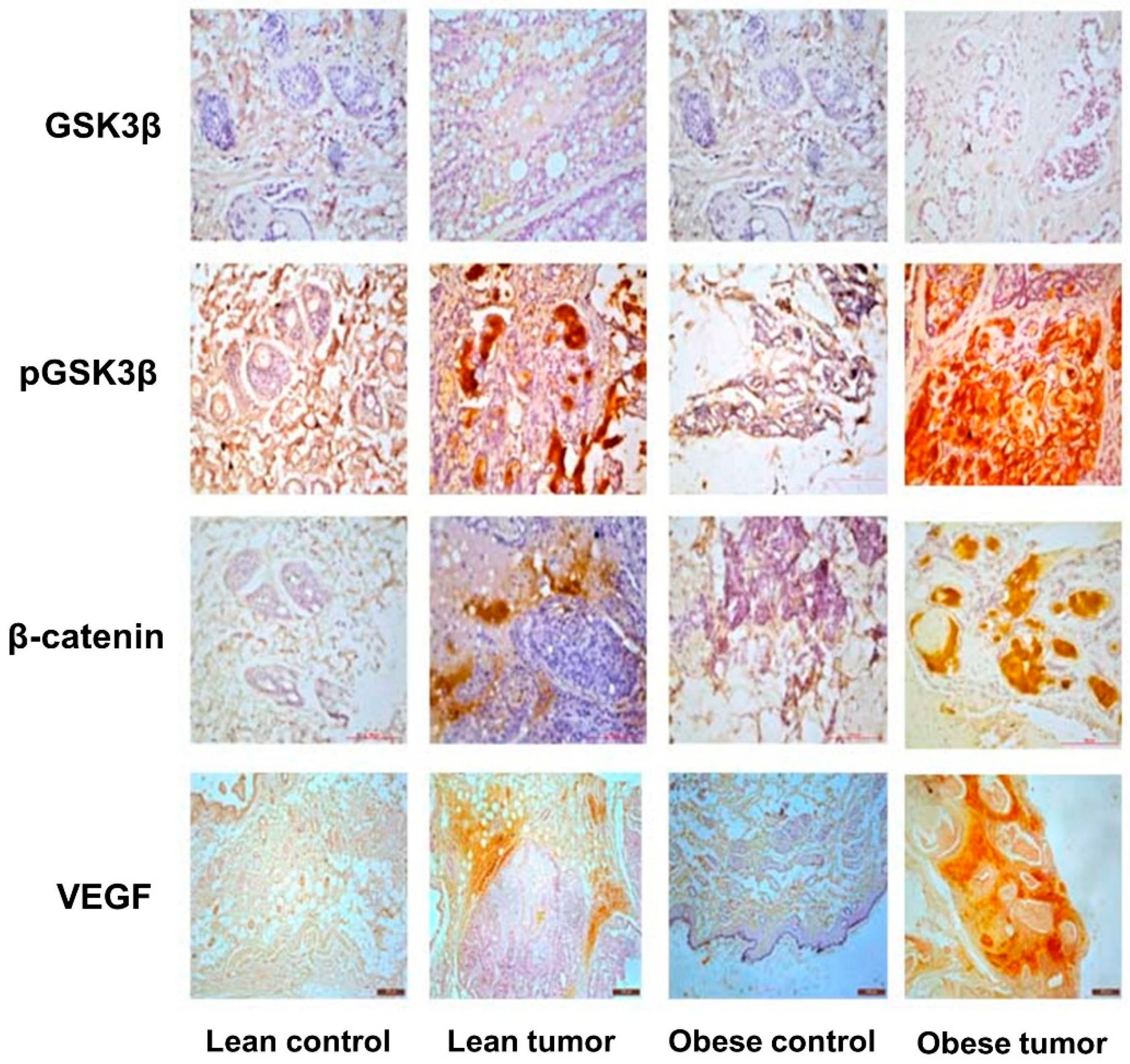
Figure 6.
Expression of PI3K/Akt/GSK-3β/ β-catenin signalling molecules by immunoblotting. Panel A: Images of immunoblots and Panel B: Quantitative data of immunoblots. The levels of pAkt, PI3K, p GSK-3β (ser9) and β-catenin were determined in the breast tissues of lean control, lean tumor, obese control and obese tumor.
Figure 6.
Expression of PI3K/Akt/GSK-3β/ β-catenin signalling molecules by immunoblotting. Panel A: Images of immunoblots and Panel B: Quantitative data of immunoblots. The levels of pAkt, PI3K, p GSK-3β (ser9) and β-catenin were determined in the breast tissues of lean control, lean tumor, obese control and obese tumor.
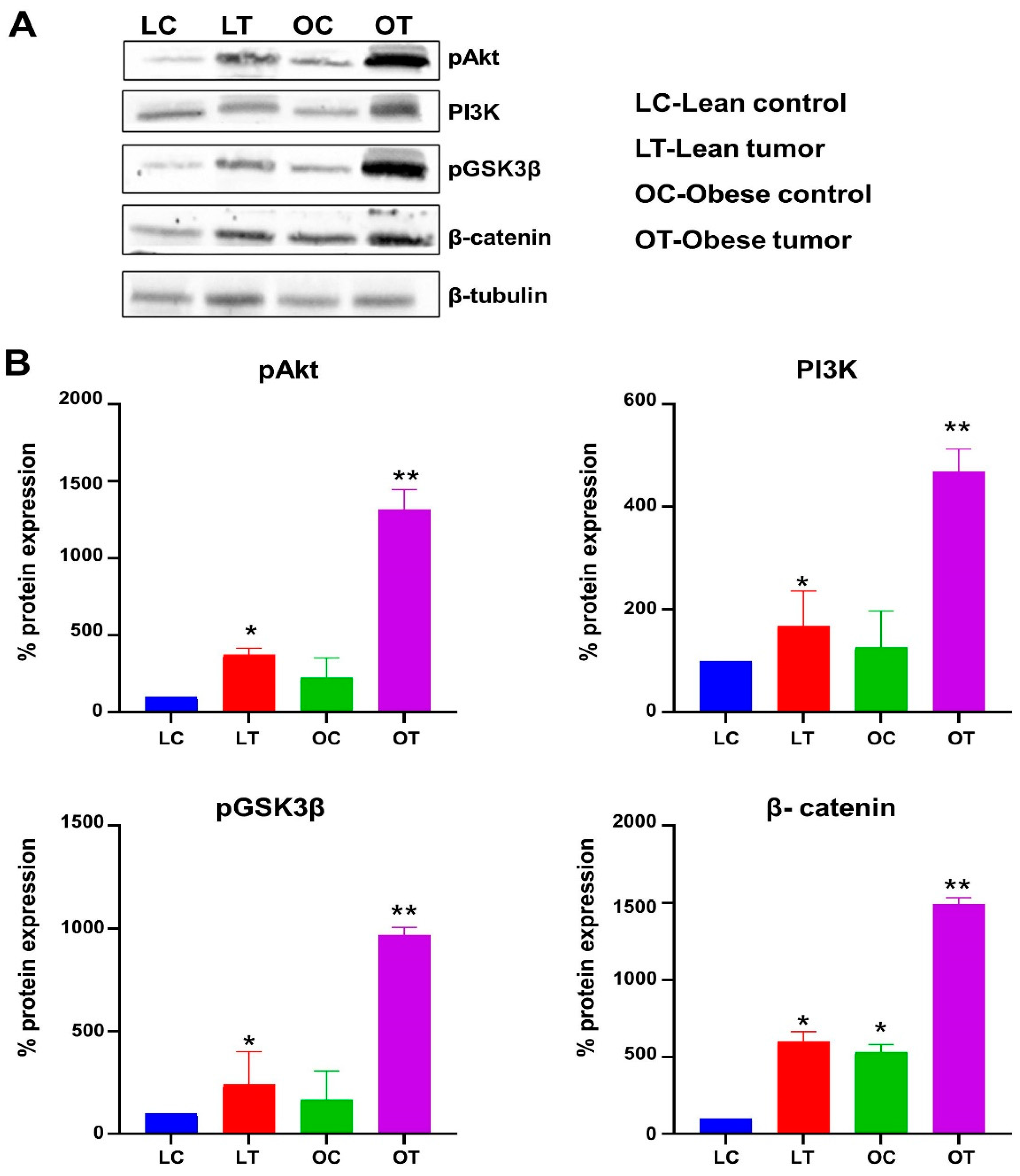
Figure 7.
Schematic representation of IGF-1/PI3K/Akt/GSK-3β/β-catenin signaling. In obese rat breast tumor tissues, the higher-level expression of IGF-1 results in binding to IGF-1R. After binding of IGF-1 ligand to its receptor IGF-1R, IRS-1 is phosphorylated to initiate the down-stream substrates PI3K. Subsequently, Akt is activated in response to PI3K signaling and get phosphorylated. The activated Akt phosphorylates GSK-3β at ser9, leading in GSK-3β inhibited and ultimately results in stabilization and accumulation β-catenin which results in higher cell proliferation (Figure 7B). In lean rats’ lower levels of IGF-1 cannot trigger the downstream cascade and reduced levels of IGF-1R, pIRS, pAkt, GSK-3β (ser9) in lean tumor tissues subsequently, activated GSK-3β stimulates degradation of GSK-3β at ser9 (Figure 7A).
Figure 7.
Schematic representation of IGF-1/PI3K/Akt/GSK-3β/β-catenin signaling. In obese rat breast tumor tissues, the higher-level expression of IGF-1 results in binding to IGF-1R. After binding of IGF-1 ligand to its receptor IGF-1R, IRS-1 is phosphorylated to initiate the down-stream substrates PI3K. Subsequently, Akt is activated in response to PI3K signaling and get phosphorylated. The activated Akt phosphorylates GSK-3β at ser9, leading in GSK-3β inhibited and ultimately results in stabilization and accumulation β-catenin which results in higher cell proliferation (Figure 7B). In lean rats’ lower levels of IGF-1 cannot trigger the downstream cascade and reduced levels of IGF-1R, pIRS, pAkt, GSK-3β (ser9) in lean tumor tissues subsequently, activated GSK-3β stimulates degradation of GSK-3β at ser9 (Figure 7A).
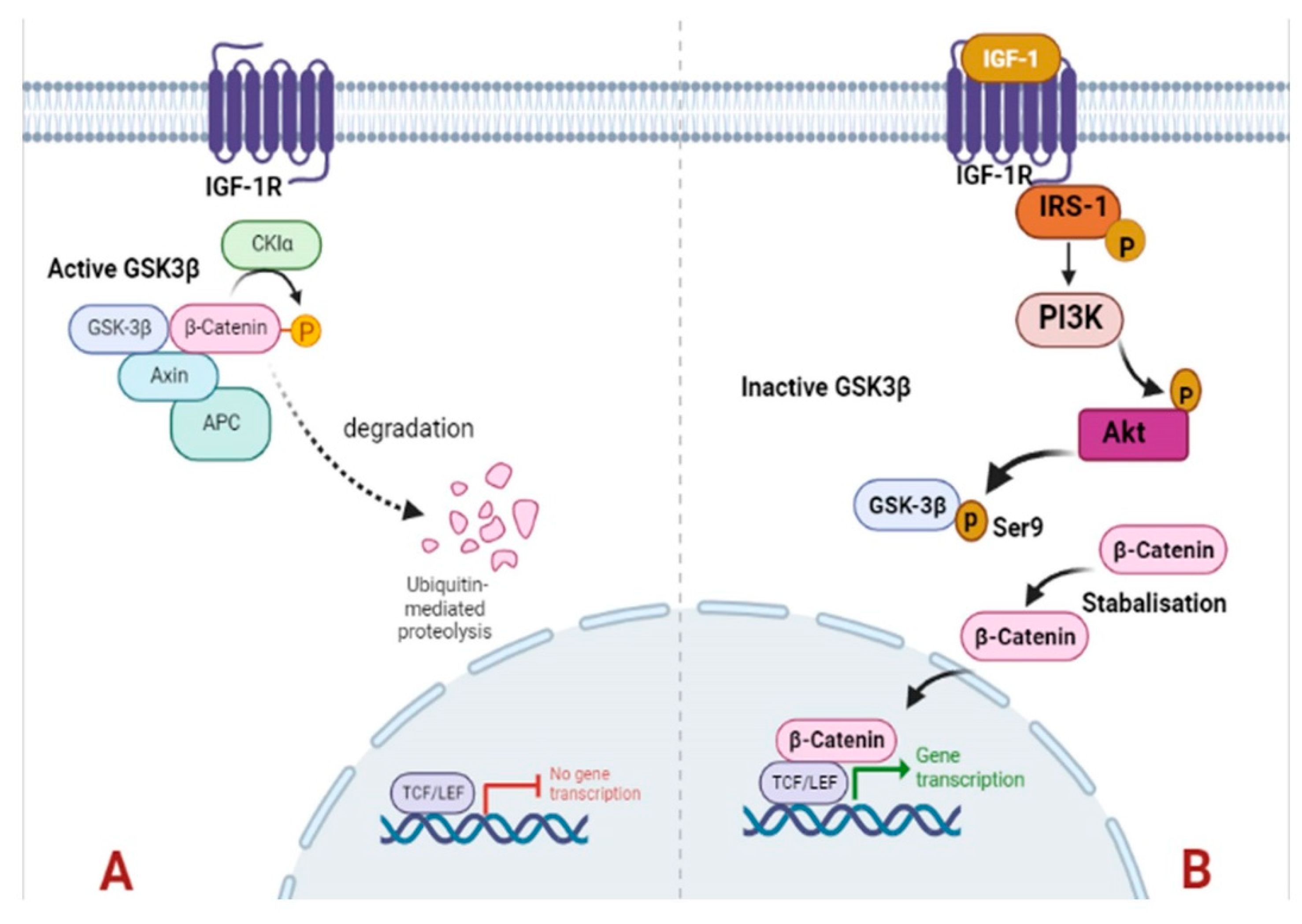

Table 1.
Tumor percentage, latency period average tumor volume and total number of tumors in obese and lean rats. Data are average of eight animals in each group,.
Table 1.
Tumor percentage, latency period average tumor volume and total number of tumors in obese and lean rats. Data are average of eight animals in each group,.
| Control | DMBA-Treated | |||
|---|---|---|---|---|
| Lean | Obese | Lean | Obese | |
| Incidence percentage | 0 | 0 | 21.42 | 62.5 |
| Latency period | 0 | 0 | 211 | 119 |
| Average tumor volume (cm2) | 0 | 0 | 7.4 | 3.06 |
| Cumulative tumor volume (cm2) | 0 | 0 | 22.22 | 24.54 |
| Total Number of tumors | 0 | 0 | 3 | 14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



