
Submitted:
16 May 2023
Posted:
17 May 2023
You are already at the latest version
Abstract
Classic Hodgkin lymphoma (cHL) is a lymphoid neoplasm composed of rare neoplastic Hodgkin and Reed-Sternberg (HRS) cells surrounded by a reactive tumor microenvironment (TME) with suppressive properties against anti-tumor immunity. TME is mainly composed of T-cells (CD4 helper, CD8 cytotoxic and regulatory), and tumor-associated macrophages (TAMs) but the impact of these cells on the natural course of the disease is not absolutely understood. TME contributes to the immune evasion of neoplastic HRS cells through production of various cytokines and/or aberrant expression of immune checkpoint molecules, in ways that have not been fully understood yet. Herein, we present a comprehensive review of findings regarding the cellular components and the molecular features of the immune TME in cHL, its correlation with treatment response and prognosis as well as the potential targeting of the TME with novel therapies. Among all cells, macrophages appear to be a most appealing target for immunomodulatory therapies, based on their functional plasticity and antitumor potency.
Keywords:
Hodgkin lymphoma
; tumor microenvironment
; tumor associated macrophages
; CD169+ macrophages
; immune evasion
; immunosuppression
1. Introduction
Hodgkin lymphoma (HL) is a B-cell lymphoid neoplasm accounting for approximately 10% of all new lymphoma diagnoses in the western world. The incidence of the disease is about 2-3/100,000 with bimodal peaks of diagnoses in the third and eighth decades of life. The mortality rate of HL is 0.4/100,000 per year, depicting the high cure rates achieved with classic chemotherapy, radiotherapy and targeted immunotherapy, although prognosis is inferior for patients at relapse [1,2].
Histologically, 95% of HL cases are termed as classic HL (cHL) with the rest of the diagnoses accounting for nodular lymphocyte predominant HL (NLPHL), while cHL is further subdivided into four subtypes: nodular sclerosis, mixed cellularity, lymphocyte-rich and lymphocyte depletion, each with different pathologic and clinical features.
Regardless of the histologic subtypes, the pathologic hallmark of cHL is the neoplastic Hodgkin and Reed-Sternberg cells (HRS). The latter ones are bi- or multi-nucleated and arise from the mono-nucleated Hodgkin cells through incomplete cytokinesis and re-fusion of daughter cells [3]]. Given their clonal Ig rearrangements and Ig genes somatic hyper-mutations, HRS cells are believed to stem from pre-apoptotic germinal center B-cells, although they have suppressed part of their B-cell gene expression program. By immunohistochemistry, HRS cells are typically positive for CD30 and variably positive for CD15 [4,5,6].
Multiple genetic mutations have been identified in HRS cells and most of them are those that affect molecular signaling pathways regulating cell survival and proliferation, primarily the NF-κΒ, JAK/STAT, PI3K/AKT and NOTCH1 pathways [7]. Another commonly detected genetic alteration in HRS cells is the amplification of the genes PD-L1 and PD-L2, leading to overexpression of the encoded immune checkpoint proteins, which regulate immune evasion [8].
Although HRS cells are considered a histologic hallmark of cHL tumors, these cells are rare, representing only 1% of the cellular composition of the tumor and are embedded within an abundant reactive cellular infiltrate. This non-neoplastic immune population is composed of T- and B-lymphocytes, neutrophils, eosinophils, macrophages, plasma cells, NK-cells, dendritic cells and mast cells, which are recruited and educated by the HRS cells. Specifically, HRS cells produce several cytokines such as CCL5, CCL17, CCL20, CCL22, CXCL9, CXCL10 along with IL-5, IL-8 and IL-9 that promote recruitment of the immune cells [4,9]. These reactive cells, in co-operation with stromal cells, fibroblasts and endothelial cells, shape a unique tumor microenvironment (TME) which is in close crosstalk with the neoplastic population. TME is believed to support the survival and promote the proliferation of HRS cells. Moreover, TME has been shown to have suppressive properties against anti-tumor immunity, thus allowing immune evasion of the neoplasm through production of various cytokines and aberrant expression of immune checkpoint molecules [10]. In this way, the TME might be involved in treatment resistance and hence, it could possibly provide targets for novel therapeutic strategies.
In this review, we summarize important research findings regarding the cellular and molecular composition of the TME in cHL, with emphasis on macrophage populations. We also shed light on a recently studied subtype of macrophages, those CD169+, and focus on how the TME and HRS cells achieve immune evasion and how the communication between neoplastic and immune cells is correlated with disease outcome and treatment response.
2. Overview of cellular components of TME in cHL
T-cells
T-cells in the TME of HL have been the object of intense investigation, among all other immune cells, due to their abundance and functional plasticity.
CD4+ T-cells
The most abundant cellular population in the TME of HL is comprised of CD4+ T-cells, with T-helper (Th) phenotype, which tend to gather around HRS cells in formations called rosettes. These cells express T-cell exhaustion markers including PD-1, TOX and TOX2 [11]. A high percentage (>75%) of tumor infiltrating CD4+ T-cells was associated with decreased freedom from treatment failure (FFTF) [12]. Initially, it was believed that the TME of cHL is dominated by Th2 CD4+ T-cells, with their increased number predicting improved disease-free survival (DFS) and event-free survival (EFS) [13]. Th2 cells are generally involved in type 2 immune response, mediated by interleukin (IL)-4, IL-5, IL-9, and IL-13, which participate in anti-helminthic immunity and tissue regeneration [14]] Nevertheless, more recent research revealed that Th cells in the TME of HL primarily polarized towards Th1 phenotype [15,16]. Th1 immunity is based on IL-2, interferon (IFN)-γ and tumor necrosis factor (TNF)-β and regulates cell mediated immune reactions that also protect against tumor cells [17,18].
FOXP3+ regulatory T-cells (Tregs) are also present in the TME of cHL. These cells restrict Th-mediated immune responses, through secretion of immunosuppressive cytokines. In this way they are involved in sustaining self-tolerance but also inhibit antitumor immunity [19]. In cHL, low numbers of FOXP3+ Tregs in TME, in combination with high numbers of cytolytic T-cells, were correlated with shortened survival [20]. Additionally, the Tregs/Th17 ratio in cHL patients was found to be positively associated with survival, thus implying that higher Th17 infiltration might characterize more aggressive disease course [21,22].
T-cells in cHL express a variety of immune checkpoint regulators, including CTLA-4, PD-1 and LAG-3, shaping a unique immunosuppressive TME that enables HRS cells to escape antitumor immunity, as discussed later.
CD8+ T-cells
CD8+ T-cells are generally known as cytolytic T-cells (CTLs) due to their capacity to directly kill infected or neoplastic cells, after recognizing antigens bound to MHC (major histocompatibility complex)-I molecules on their surface and are, therefore, considered as important mediators of anti-tumor immunity, along with other major cytolytic cells, the NK-cells. In case of cHL, CD8+ T-cell subpopulation is less abundant than the CD4+ one, with contradictory results regarding its prognostic value. Alonso-Álvarez et al found that high numbers of CD8+ T-cells predict better outcome in patients treated with ABVD as first line therapy [12]. In contrast, the presence of activated CTLs (positive for TIA-1 and granzyme B) in the TME of cHL has been correlated with decreased survival in the relapsed/refractory (R/R) setting [20,23]. Regardless of their prognostic role, CD8+ T-cells seem to have an important role in shaping the TME of cHL, since they express immune checkpoint molecules such as programmed death (PD)-1, indoleamine 2,3-Dioxygenase 1 (IDO)-1 and TIM-3 more frequently than CD4+ T-cells [24] However, these cells seem to have diverged from their cytotoxic role against neoplastic cells. A subset of CD8+ T-cells was identified in cHL TME that shares phenotypic and functional characteristics with T-follicular helper cells. Specifically, they co-express CXCR5 and ICOS, Bcl-6, PD-1 and CD200 and show deficient cytotoxicity and low IFN-γ secretion [25]. The function of CD8+ T-cells might be negatively influenced by Galectin-1 produced by HRS cells [26].
B-cells
The presence and prognostic value of non-neoplastic B-cells of TME in cHL has been studied by independent research groups, based on their negative impact in several solid malignancies. Interestingly, high proportions of CD20+ background cells in TME was correlated with increased overall survival (OS), while low B-cell counts were associated with shortened progression-free survival (PFS) and OS among patients treated with BEACOPP-based regimens, thus B-cells show both prognostic and predictive value [27,28]. A possible explanation for B-cells favorable effect might be the competition with neoplastic cells for survival and growth signals, although more research is needed to clarify whether all B-cells or specific sub-populations have a favorable predictive impact, since the presence of the PAX5+/CD38+ sub-population was shown to correlate with adverse outcomes [29,30].
Plasma cells
Data on the role of plasma cells in the TME of cHL has been scarce so far. Tumor infiltration by CD138+ plasma cells is associated with advanced disease stage, eosinophil infiltration and the presence of B-symptoms and a tendency towards inferior OS and EFS [31]. Additionally, elevated polyclonal serum free light chains in patients with cHL showed correlation with decreased survival and it has been assumed that these light chains are produced by plasma cells of the TME, since HRS are considered incapable of secreting immunoglobulins [32].
NK-cells
Although NK cells are innate lymphoid cells with known anti-tumor cytotoxic activity, in the case of HL ME these cells seem to be numerically and functionally diminished. The inhibition of NK cytotoxic activity is primarily mediated by ligands found in TME which bind to NK inactivating receptors [33]. Furthermore, in patients with HL, the ratio of CD56dimDNAM-1pos NK cells over CD56dimDNAM-1neg NK cells is reduced, indicating a shift of NK phenotype towards the less cytotoxic DNAM-1neg population. Even CD56dimDNAM-1pos NK cells were found to show impaired cytotoxic activity in HL patients compared to healthy individuals [34]. One possible explanation for the limited NK population in HL TME might be the induction of apoptosis triggered by the binding of Fas-L of HRS cells to Fas receptor of NK cells [22].
Myeloid cells
Myeloid derived suppressor cells (MDSCs) are a heterogeneous population of immature myeloid cells, expressing CD11b and CD33, which exert immunosuppressive roles when infiltrating tumors. Their immunosuppressive effect is mainly towards T-cells, since their high expression of Arginase-I (Arg-I) is believed to deprive T-cells in TME of L-arginine which is essential for their function [35,36]. High tissue levels of Arg-I positive myeloid cells was associated with inferior disease outcomes in HL [37]. Similarly, a subset of MDSCs, circulating CD34+ MDSCs, were found to negatively influence the PFS of patients with HL [38]. Interestingly, MDSCs were reduced in patients after treatment with brentuximab vedotin (BV) and baseline serum Arg-I levels emerged as a potential predictive biomarker for BV treatment response [39].
Eosinophils represent one of the most typical cellular populations found in HL biopsies and they are believed to develop a close crosstalk with HRS cells via CD30-CD30L binding, but their prognostic value remains a matter of debate. Although research indicated that eosinophilic tumor infiltration strongly correlates with FFTF, Axdorph et al did not find any association with clinical outcome, thus implying the need for further relevant investigation [40,41].
Tumor associated neutrophils (TANs), similarly to tumor associated macrophages (TAMs), appear with variable effects in TME, from suppressing anti-tumor immunity to cytotoxicity against neoplastic cells [42]. Due to this and to the immunohistochemichal overlap with MDSCs, research on the prognostic impact of neutrophils in HL has been limited to their peripheral blood counts rather than tumor infiltration. Indeed, in cHL a high absolute neutrophil count to a high absolute lymphocyte count ratio is an independent prognostic factor for patients’ reduced OS [43].
Mast cells
Evidence on the prognostic role of mast cells in HL is controversial, although their biological properties have been well-described. Mast cells are the predominant cells of the TME that express CD30L, the ligand for CD30 receptor of HRS cells, thus indicating close interaction with the neoplastic population [44]. Additionally, mast cells are believed to promote tumor growth by induction of neovascularization and fibrosis, functions that can be inhibited by bortezomib, thus providing a potential therapeutic target [45]. The hypothesized negative prognostic impact, in terms of reduced relapse-free survival, of mast cells in TME of HL was indeed demonstrated by Molin et al [46] although other researchers did not find a correlation between mast cells infiltration and prognosis [47].
Dendritic cells
Dendritic cells (DCs) have also been studied in the TME of HL with variable results, depending on the specific DC subtype. CD123+ plasmacytoid DCs are the most abundant DC type in cHL, although they do not seem to correlate with disease-specific survival and they produce reduced amounts of IFN-a compared to healthy individuals, implying an immune functional defect [48,49]. As for myeloid DCs, most of them in cHL TME are identified as a mature CD83+ subtype whose number is positively associated with improved disease specific survival of patients [49]. Finally, the presence of follicular DCs in most subtypes of HL was found to predict a favorable outcome [50]. Patients with cHL were also found to have lower counts of all subtypes of circulating DCs compared to healthy individuals [51].
Tumor Associated Macrophages
In general, macrophages are derived from mononuclear cells and have multiple roles including, but not limited to, phagocytosis, antigen presentation to other immune cells and tissue remodeling. Among all immune cells found in the TME of cHL, macrophages have attracted most research interest. This is because of the great plasticity of these cells, indicated by their ability to acquire different phenotypes that influence the tumor microenvironment towards an immunosuppressive or inflammatory state. Accordingly, macrophages variably influence disease progression, and this is probably the reason why the association of TAMs with disease outcomes, treatment response and patients’ survival has been so challenging over years.
TAMs are recruited in TME through GM-CSF, CCL2, CCL5, CCL7, and CXCL1, which are secreted by neoplastic cells. There, TAMs are programmed towards M1 or M2 phenotypes. Although initially considered as distinct subtypes, this view has been criticized as oversimplified and the current notion is that M1 and M2 phenotypes actually represent the extremities of a continuum spectrum [52,53].
M1 phenotype is triggered by GM-CSF, IFN-γ and lipopolysaccharides and is characterized by cytotoxic, pro-inflammatory and anti-neoplastic effects mediated by secretion of TNF-a, NO, Chemokine (C-X-C motif) ligand (CXCL) 9, CXCL10, CXCL11, IL-1, IL-6, IL-12, IL-23 and ROS by M1 macrophages. On the other hand, M2 polarized macrophages, driven by M-CSF, TGF-B, IL-4, IL-10 and IL-13, are believed to promote wound healing, angiogenesis and tumor growth by producing tumor growth factor (TGF)-β, IL-10, CC motif chemokine ligand (CCL) 17, CCL18, CCL22, CD206, CD204 and CD163 [54]. Apparently, the balance between the anti-neoplastic and pro-tumorigenic phenotypes is crucial for determining disease outcomes and a tempting field for therapeutic interventions (Figure 1).
In cHL HRS cells can lead TAMs to polarize towards the tumor-promoting M2 phenotype, via secretion of TGF-β and IL-13. In turn, M2 macrophages support the survival of HRS cells, partially through activation of STAT3 signaling pathway [54]. As previously discussed, TAMs actively participate in shaping the protective niche around HRS cells and express several immunosuppressive molecules such as IDO-1 and immune checkpoint proteins such as PD-L1 and CD86, through which they interact with immune cells of the TME, further promoting suppression of the anti-tumor immunity [55,56]. Another mechanism in which TAMs might promote tumor growth is supposed to be the induction of genetic instability of HRS cells, probably through release of free radicals which contribute to a mutagenic microenvironment. This was based on the observation that TP53 amplification in neoplastic cells, linked with poorer patients’ survival, was associated with increased infiltration by M2 macrophages [57].
Regarding the prognostic implications of macrophages infiltration of cHL, there have been multiple studies with rather contradictory results, probably reflecting the heterogeneity of macrophage sub-populations and the difficulties identifying each macrophage phenotype, given the lack of a definite immunohistochemistry marker. Many available studies usually utilize CD68 as a universal macrophage marker. CD163 is used by other research groups for further characterization of M2 polarized macrophages, although other researchers doubt whether CD163 is an ideal marker for detecting the M2 subgroup of CD68+ macrophages [58].
Steidl et al were the first to prove that an increased number of TAMs (CD68+) is strongly associated with shortened survival in patients with cHL, using immunohistochemistry [59]. Similarly, several groups confirmed the negative correlation of CD68+ TAMs in HL with patients’ survival and/or response to treatment [60,61,62,63,64,65,66,67], while the prognostic value of CD68+ macrophages was not reproduced by other groups [53,68,69,70,71].
When CD163 was used as a TAMs marker, with the rationale to focus on M2 polarized macrophages, it has also been found that increased infiltration correlates with poorer outcomes [65,66,67,70,72] including response to nivolumab [73], but this was doubted by others [71].
In the only meta-analysis conducted concerning the prognostic impact of TAMs in cHL, Guo et al found that high density of either CD68+ or CD163+ TAMs in the TME translates into poorer OS and PFS [74].
Interestingly, Werner et al demonstrated that both a very high and a very low number of infiltrating TAMs (CD68+ or CD163+) is associated with worst outcomes, compared with intermediate TAMs levels, while Karihtala et al indicated that the adverse prognostic effect of TAMs in cHL is immune checkpoint-dependent, because only PD-L1+ and IDO-1+ TAMs were associated with inferior outcomes, but not TAMs on the whole [58,75].
Based on the above findings, reprogramming macrophages of the TME in cHL towards the anti-tumorigenic M1 phenotype has been attempted, as in the case of PI3Kδ/γ inhibitor RP6530 [76]. Other therapeutic strategies which exploit TAMs biology with promising results include inhibition of TAMs recruitment in the TME or direct targeting of TAMs with nanomaterials [52].
CD169+ macrophages: A new regulator of antitumor immunity
CD169+ macrophages constitute a subpopulation distinct from M1 and M2 phenotypes, as they can simultaneously express markers of both M1 and M2 subtypes. Normally, they are primarily detected in the metallophilic marginal zone of the spleen and in the medulla and the subcapsular sinus of lymph nodes, but they can also be found in the intestine, liver, and bone marrow. Based on their localization, CD169+ macrophages (also known as Siglec-1 positive macrophages) basically function as “gatekeepers” of secondary lymphoid organs, since they are the first cell type that captures antigens in lymph nodes and spleen, present them to other immune cells and, thus, help the activation of T-cells and initiate adaptive immune responses. Apart from viral and bacterial inflammatory responses, CD169+ macrophages participate in immune tolerance induced by apoptotic cells clearance [77,78]. More interestingly, it has been demonstrated that CD169+ macrophages phagocytize dead tumor cells transported via lymphatic flow and present tumor associated peptides to CD8+ T-cells, whose cytotoxic activity is augmented, which is considered a crucial step for the induction of antitumor immunity [79,80].
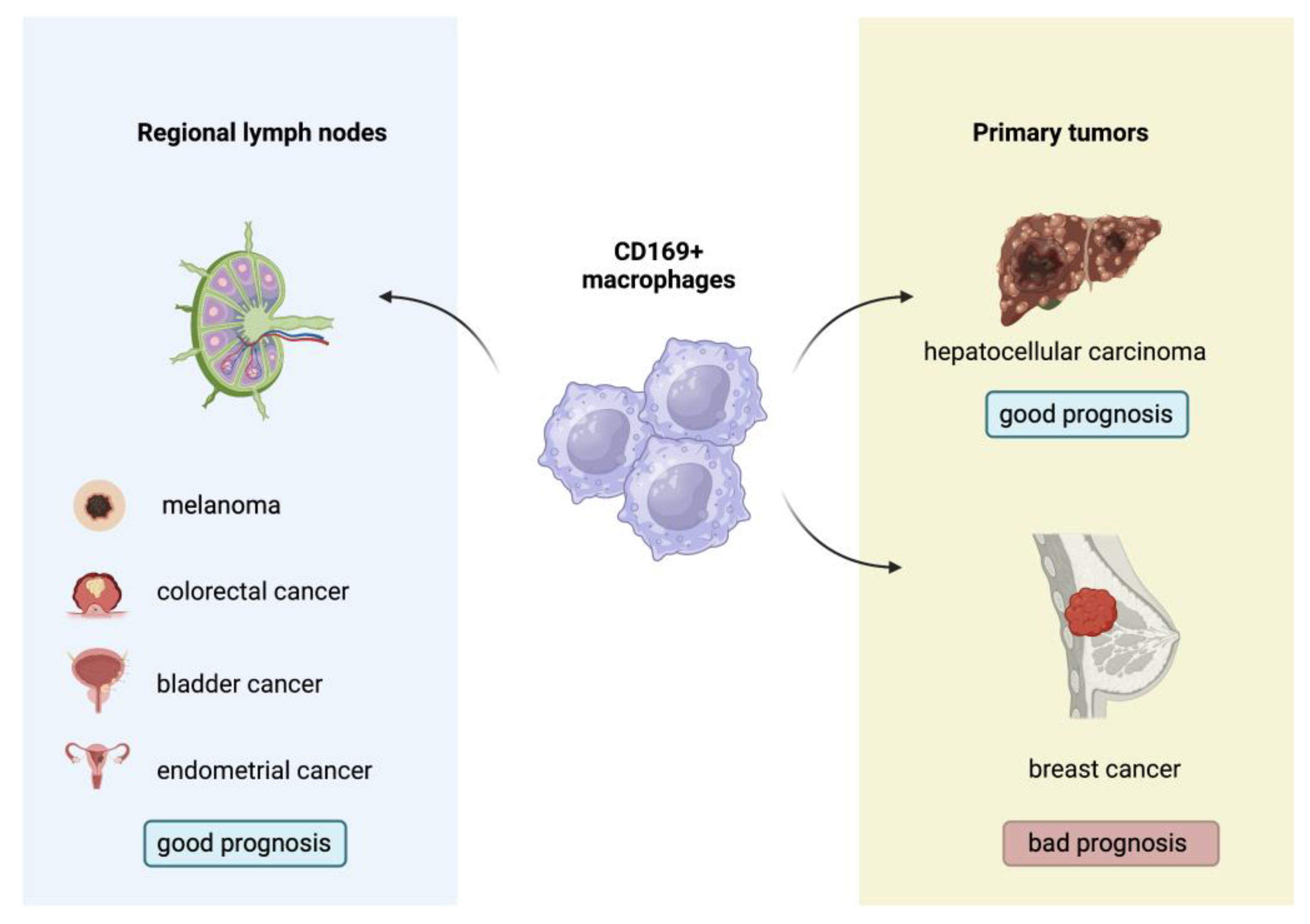
Indeed, the biological and prognostic role of CD169+ macrophages in several human malignancies has gathered research interest. A higher concentration of CD169+ macrophages in the primary tumors or regional lymph nodes has been associated with improved outcomes in patients with melanoma, hepatocellular carcinoma, colorectal, bladder and endometrial cancers, with contradictory results in breast cancer [81,82,83,84,85,86,87] (Figure 2). Moreover, Marmey et al investigated by immunohistochemistry on paraffin-embedded tissues the expression of CD169 in 51 cases of B-cell non-Hodgkin lymphomas (including diffuse large B-cell lymphomas, B-chronic lymphocytic leukemias, follicular lymphomas, mantle cell lymphomas and splenic marginal zone lymphomas). Only splenic marginal zone lymphomas (15 cases) showed remarkable increase in CD169+ cells, with preferential distribution in the splenic cords of the red pulp [88]. These CD169+ cells were also positive for CD14 (monocyte/macrophage marker) and it has been hypothesized that the CD169+/CD14+ cells observed in the splenic cords of splenic marginal zone lymphomas might have a dendritic cell differentiation potential [88]. In this latter study, however, no information about any prognostic implication of the CD169 immunostaining patterns in B-cell non-Hodgkin lymphomas was reported [88]. Interestingly, to the best of our knowledge, there is no published study regarding the potential prognostic impact of the immunohistochemical expression patterns of the CD169+ macrophages in the TME of cHL. This investigation could permit a better understanding of the interactions taking place in the TME of cHL and might indicate a new prognostic biomarker or even a therapeutic target, given the already discussed role of M1 and M2 subpopulations in cHL.
3. Immune Evasion
Multiple ways by which HRS cells manage to escape antitumor immune surveillance have been identified. Overall, the neoplastic HRS cells with their altered gene-expression profiling seem to hide from antitumor immune cells. On the other hand, as previously discussed, HRS attract, through cytokines, several immune cells, which in turn shape an immunosuppressive TME that further allows tumor cells evade the physiological antitumor immune responses.
At first place, HRS cells commonly exhibit copy number gains of 9p24.1 locus, which is associated with enhanced expression of PD-1 ligands (PD-L1 and PD-L2) in up to 97% of cases. These ligands bind to PD-1 receptor on T-cells, which inhibits their activation and proliferation, leading to T-cell exhaustion. Interestingly, in cHL, PD-L1 is also expressed by TAMs which co-localize with HRS cells, thus augmenting the immunosuppressive signals against host immunity [11,55,89]. HRS cells have been shown capable of transferring PD-L1 molecules to TAMs’ surface, via trogocytosis, a process that enables neighboring cells exchange membrane molecules, which underlines the role of HRS cells’ in actively shaping their microenvironment to their benefit [90]. The immunosuppression via PD-1 interactions has provided the biologic rationale for anti-PD1 blockade therapy with nivolumab or pembrolizumab, which, in HL, has achieved the highest response rates among human malignancies. Indeed, increased PD-L1 expression on HRS cells has been shown to predict better PFS with PD-1 blockade treatment, functioning as a predictive biomarker [91]. Additionally, the TME of cHL is highly enriched for CTLA-4+ T-cells, while HRS cells and TAMs provide the corresponding inhibitory ligand, CD86. The interacting T-cells and TAMs are gathered around HRS cells, forming a protective niche for the neoplastic cellular population [56] On the contrary, CD8+ cytolytic T-cells are located in a distance from this niche, which apparently minimizes their destructive effects on HRS cells [92]. Double therapeutic targeting of the immune checkpoints, CTLA-4 and PD-1 has been proposed as a promising alternative, especially for relapsed or refractory disease. A third important immune checkpoint molecule is LAG-3, expressed on both CD4+ T-cells, NK-cells and rarely HRS cells, which acts synergistically with CTLA-4 and PD-1 towards CD8+ T-cell suppression and promotion of Treg function. This checkpoint offers an additional target, under investigation, for immunotherapy, alone or in combination with anti-PD-1, since it is considered as one of the major ways of acquired resistance to anti-PD-1 treatment [24,93,94]. A high expression level of all these checkpoint proteins by TME immune cells, mainly CD4+ and CD8+ T-cells and TAMs, is associated with shortened OS, apparently reflecting the immunosuppressive state maintained by these proteins and this could justify therapeutic combination of immunotherapy with classic chemotherapy regimens [24]. Another immune checkpoint protein, TIM-3, is generally expressed by innate immune cells and T-cells and decreases macrophage activation while promoting MDSCs. In cHL TIM-3 is variably expressed by HRS cells and surrounding immune cells, emerging as an additional target for therapies aiming at restoring the hosts’ anti-tumor immunity by blocking the immunosuppressive checkpoint proteins [95].
HRS cells have also developed means to escape from cytotoxic immune cells. Down-regulation of B2MG and MHC-I expression probably allows neoplastic cells avoid interactions with CD8+ T-cells. Similarly, inactivating mutations of the CD58 gene have been detected, especially in cells derived from advanced-stage disease patients. Since CD58 is a receptor recognized by cytolytic CD8+ and NK cells, such mutations could further protect HRS cells from immune lysis [96,97].
Extracellular vesicles (EVs) secreted by HRS cells carry TNF-α, ADAM10 and sCD30 that are believed to participate in the education of stromal cells, especially fibroblasts, to support the survival of HRS cells or even protect them from anti-cancer treatments such as BV [98].
Apart from the direct cellular interactions, the immunosuppressive state of the TME in cHL is largely attributed to the inhibitory function of multiple cytokines and other immune-effective molecules. These include IL-10 and TGF-β produced by Th2 CD4+ T-cells, but also galectin-1 which is mainly produced by HRS cells and exerts multiple inhibitory effects on T-cell populations [26]. Histiocytes, dendritic cells and endothelial cells in the TME express IDO [99]. This enzyme catabolizes tryptophan, the depletion of which inhibits T-cell function and induces T- and NK-cell apoptosis and its expression in the TME of HL, although restricted in specific histologic subtypes, was found to be an independent prognostic factor for inferior survival [26,99,100]. Even serum kynurenine/tryptophan ratio significantly correlates to OS, further demonstrating the importance of IDO in disease pathophysiology as well as the potential of targeting IDO for a therapeutic benefit [101].
Finally, as discussed above, the lymphocytic population of the TME in cHL is highly composed of Tregs, which have innate immunosuppressive functions as they primarily induce primarily T-cell suppression [102]
5. Conclusions
There is a body of evidence that the unique TME of cHL is vital for the survival of neoplastic HRS cells as well as tumor growth, by providing multiple molecular signals that promote cellular proliferation and protection against the host’s anti-tumor immunity. Each cellular type of the TME, rather than being random by-standers, seems to contribute to forming the immunosuppressive niche for HRS cells, although the whole extent of the molecular interactions among immune cells, or between immune cells and HRS cells might not be fully elucidated yet.
As far as TAMs are concerned, they seem to play a crucial role in shaping the TME and determining the direction of immune responses towards anti-tumor activity or to immunosuppressive and tumor-promoting phenotype. The exact balance between the two macrophage phenotypes, due to the cellular plasticity of TAMs, is probably the reason under the divergent findings of original studies attempting to correlate TAMs with patients’ survival in cHL, although this might also be attributed to more technical issues, such as defining the most appropriate immunohistochemistry markers.
It is, therefore, reasonable to suggest that more extensive research of macrophage populations, such as the CD169+ ones, could shed light on unknown aspects of the TME in cHL and might also provide another prognostic biomarker. Whether this, as well as, multiple other previously discussed prognostic biomarkers will soon be utilized in clinical practice and decision making, remains to be explored, given the complexity of incorporating all these prognostic information into one unified prognostic scoring system.
As far as potential therapeutic implications are concerned, the successful use of immune checkpoint inhibitors has given hope that other sides of tumor immune evasion could be targeted to restore the host’s anti-tumor immunity against lymphoma cells. In this direction, reprogramming the macrophage pool of TME or their monocyte precursors towards the tumor-attacking phenotype, commonly referred to as M2, seems to be a promising strategy. Deeper understanding of the multilevel interactions within TME of cHL could also guide therapeutic strategies for patients who relapse after standard therapy or are resistant to treatment with immune checkpoint inhibitors. In that way, patients with HL could benefit from more personalized treatments based on their TME, to further increase the cure and survival rates of the disease.
Author Contributions
Conceptualization, P.K. and A.P-B.; writing—original draft preparation, V.G.; writing—review and editing, V.G., A.P-B., A.M., E.H., P.K.; visualization, E.H.; supervision, P.K. E.H..; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable
Conflicts of Interest
The authors declare no conflict of interest.
References
- Eichenauer, D.A.; Aleman, B.M.P.; Andre, M.; Federico, M.; Hutchings, M.; Illidge, T.; Engert, A.; Ladetto, M.; Committee, E.G. Hodgkin lymphoma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann Oncol 2018, 29, iv19–iv29. [Google Scholar] [CrossRef]
- Ansell, S.M. Hodgkin lymphoma: 2023 update on diagnosis, risk-stratification, and management. Am J Hematol 2022, 97, 1478–1488. [Google Scholar] [CrossRef]
- Rengstl, B.; Newrzela, S.; Heinrich, T.; Weiser, C.; Thalheimer, F.B.; Schmid, F.; Warner, K.; Hartmann, S.; Schroeder, T.; Kuppers, R.; et al. Incomplete cytokinesis and re-fusion of small mononucleated Hodgkin cells lead to giant multinucleated Reed-Sternberg cells. Proc Natl Acad Sci U S A 2013, 110, 20729–20734. [Google Scholar] [CrossRef]
- Weniger, M.A.; Kuppers, R. Molecular biology of Hodgkin lymphoma. Leukemia 2021, 35, 968–981. [Google Scholar] [CrossRef]
- Satou, A.; Takahara, T.; Nakamura, S. An Update on the Pathology and Molecular Features of Hodgkin Lymphoma. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Kuppers, R.; Schwering, I.; Brauninger, A.; Rajewsky, K.; Hansmann, M.L. Biology of Hodgkin's lymphoma. Ann Oncol 2002, 13 (Suppl. 1), 11–18. [Google Scholar] [CrossRef]
- Brauninger, A.; Schmitz, R.; Bechtel, D.; Renne, C.; Hansmann, M.L.; Kuppers, R. Molecular biology of Hodgkin's and Reed/Sternberg cells in Hodgkin's lymphoma. Int J Cancer 2006, 118, 1853–1861. [Google Scholar] [CrossRef]
- Roemer, M.G.; Advani, R.H.; Ligon, A.H.; Natkunam, Y.; Redd, R.A.; Homer, H.; Connelly, C.F.; Sun, H.H.; Daadi, S.E.; Freeman, G.J.; et al. PD-L1 and PD-L2 Genetic Alterations Define Classical Hodgkin Lymphoma and Predict Outcome. J Clin Oncol 2016, 34, 2690–2697. [Google Scholar] [CrossRef]
- Skinnider, B.F.; Mak, T.W. The role of cytokines in classical Hodgkin lymphoma. Blood 2002, 99, 4283–4297. [Google Scholar] [CrossRef]
- de Charette, M.; Houot, R. Hide or defend, the two strategies of lymphoma immune evasion: potential implications for immunotherapy. Haematologica 2018, 103, 1256–1268. [Google Scholar] [CrossRef]
- Takahara, T.; Satou, A.; Tsuzuki, T.; Nakamura, S. Hodgkin Lymphoma: Biology and Differential Diagnostic Problem. Diagnostics (Basel) 2022, 12. [Google Scholar] [CrossRef]
- Alonso-Alvarez, S.; Vidriales, M.B.; Caballero, M.D.; Blanco, O.; Puig, N.; Martin, A.; Penarrubia, M.J.; Zato, E.; Galende, J.; Barez, A.; et al. The number of tumor infiltrating T-cell subsets in lymph nodes from patients with Hodgkin lymphoma is associated with the outcome after first line ABVD therapy<sup/>. Leuk Lymphoma 2017, 58, 1144–1152. [Google Scholar] [CrossRef]
- Schreck, S.; Friebel, D.; Buettner, M.; Distel, L.; Grabenbauer, G.; Young, L.S.; Niedobitek, G. Prognostic impact of tumour-infiltrating Th2 and regulatory T cells in classical Hodgkin lymphoma. Hematol Oncol 2009, 27, 31–39. [Google Scholar] [CrossRef]
- Walker, J.A.; McKenzie, A.N.J. T(H)2 cell development and function. Nat Rev Immunol 2018, 18, 121–133. [Google Scholar] [CrossRef]
- Cader, F.Z.; Schackmann, R.C.J.; Hu, X.; Wienand, K.; Redd, R.; Chapuy, B.; Ouyang, J.; Paul, N.; Gjini, E.; Lipschitz, M.; et al. Mass cytometry of Hodgkin lymphoma reveals a CD4(+) regulatory T-cell-rich and exhausted T-effector microenvironment. Blood 2018, 132, 825–836. [Google Scholar] [CrossRef]
- Greaves, P.; Clear, A.; Owen, A.; Iqbal, S.; Lee, A.; Matthews, J.; Wilson, A.; Calaminici, M.; Gribben, J.G. Defining characteristics of classical Hodgkin lymphoma microenvironment T-helper cells. Blood 2013, 122, 2856–2863. [Google Scholar] [CrossRef]
- Romagnani, S. Th1/Th2 cells. Inflamm Bowel Dis 1999, 5, 285–294. [Google Scholar] [CrossRef]
- Nishimura, T.; Nakui, M.; Sato, M.; Iwakabe, K.; Kitamura, H.; Sekimoto, M.; Ohta, A.; Koda, T.; Nishimura, S. The critical role of Th1-dominant immunity in tumor immunology. Cancer Chemother Pharmacol 2000, 46 Suppl, S52–61. [Google Scholar] [CrossRef]
- Cretney, E.; Kallies, A.; Nutt, S.L. Differentiation and function of Foxp3(+) effector regulatory T cells. Trends Immunol 2013, 34, 74–80. [Google Scholar] [CrossRef]
- Alvaro, T.; Lejeune, M.; Salvado, M.T.; Bosch, R.; Garcia, J.F.; Jaen, J.; Banham, A.H.; Roncador, G.; Montalban, C.; Piris, M.A. Outcome in Hodgkin's lymphoma can be predicted from the presence of accompanying cytotoxic and regulatory T cells. Clin Cancer Res 2005, 11, 1467–1473. [Google Scholar] [CrossRef]
- Dehghani, M.; Kalani, M.; Golmoghaddam, H.; Ramzi, M.; Arandi, N. Aberrant peripheral blood CD4(+) CD25(+) FOXP3(+) regulatory T cells/T helper-17 number is associated with the outcome of patients with lymphoma. Cancer Immunol Immunother 2020, 69, 1917–1928. [Google Scholar] [CrossRef]
- Ferrarini, I.; Rigo, A.; Visco, C.; Krampera, M.; Vinante, F. The Evolving Knowledge on T and NK Cells in Classic Hodgkin Lymphoma: Insights into Novel Subsets Populating the Immune Microenvironment. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef]
- Koreishi, A.F.; Saenz, A.J.; Persky, D.O.; Cui, H.; Moskowitz, A.; Moskowitz, C.H.; Teruya-Feldstein, J. The role of cytotoxic and regulatory T cells in relapsed/refractory Hodgkin lymphoma. Appl Immunohistochem Mol Morphol 2010, 18, 206–211. [Google Scholar] [CrossRef]
- Karihtala, K.; Leivonen, S.K.; Karjalainen-Lindsberg, M.L.; Chan, F.C.; Steidl, C.; Pellinen, T.; Leppa, S. Checkpoint protein expression in the tumor microenvironment defines the outcome of classical Hodgkin lymphoma patients. Blood Adv 2022, 6, 1919–1931. [Google Scholar] [CrossRef]
- Le, K.S.; Ame-Thomas, P.; Tarte, K.; Gondois-Rey, F.; Granjeaud, S.; Orlanducci, F.; Foucher, E.D.; Broussais, F.; Bouabdallah, R.; Fest, T.; et al. CXCR5 and ICOS expression identifies a CD8 T-cell subset with T(FH) features in Hodgkin lymphomas. Blood Adv 2018, 2, 1889–1900. [Google Scholar] [CrossRef]
- Gandhi, M.K.; Moll, G.; Smith, C.; Dua, U.; Lambley, E.; Ramuz, O.; Gill, D.; Marlton, P.; Seymour, J.F.; Khanna, R. Galectin-1 mediated suppression of Epstein-Barr virus specific T-cell immunity in classic Hodgkin lymphoma. Blood 2007, 110, 1326–1329. [Google Scholar] [CrossRef]
- Jachimowicz, R.D.; Pieper, L.; Reinke, S.; Gontarewicz, A.; Plutschow, A.; Haverkamp, H.; Frauenfeld, L.; Fend, F.; Overkamp, M.; Jochims, F.; et al. Whole-slide image analysis of the tumor microenvironment identifies low B-cell content as a predictor of adverse outcome in patients with advanced-stage classical Hodgkin lymphoma treated with BEACOPP. Haematologica 2021, 106, 1684–1692. [Google Scholar] [CrossRef]
- Panico, L.; Tenneriello, V.; Ronconi, F.; Lepore, M.; Cantore, N.; Dell'Angelo, A.C.; Ferbo, L.; Ferrara, F. High CD20+ background cells predict a favorable outcome in classical Hodgkin lymphoma and antagonize CD68+ macrophages. Leuk Lymphoma 2015, 56, 1636–1642. [Google Scholar] [CrossRef]
- Calabretta, E.; d'Amore, F.; Carlo-Stella, C. Immune and Inflammatory Cells of the Tumor Microenvironment Represent Novel Therapeutic Targets in Classical Hodgkin Lymphoma. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Tudor, C.S.; Distel, L.V.; Eckhardt, J.; Hartmann, A.; Niedobitek, G.; Buettner, M. B cells in classical Hodgkin lymphoma are important actors rather than bystanders in the local immune reaction. Hum Pathol 2013, 44, 2475–2486. [Google Scholar] [CrossRef]
- Gholiha, A.R.; Hollander, P.; Hedstrom, G.; Sundstrom, C.; Molin, D.; Smedby, K.E.; Hjalgrim, H.; Glimelius, I.; Amini, R.M.; Enblad, G. High tumour plasma cell infiltration reflects an important microenvironmental component in classic Hodgkin lymphoma linked to presence of B-symptoms. Br J Haematol 2019, 184, 192–201. [Google Scholar] [CrossRef]
- Thompson, C.A.; Maurer, M.J.; Cerhan, J.R.; Katzmann, J.A.; Ansell, S.M.; Habermann, T.M.; Macon, W.R.; Weiner, G.J.; Link, B.K.; Witzig, T.E. Elevated serum free light chains are associated with inferior event free and overall survival in Hodgkin lymphoma. Am J Hematol 2011, 86, 998–1000. [Google Scholar] [CrossRef]
- Chiu, J.; Ernst, D.M.; Keating, A. Acquired Natural Killer Cell Dysfunction in the Tumor Microenvironment of Classic Hodgkin Lymphoma. Front Immunol 2018, 9, 267. [Google Scholar] [CrossRef]
- Stannard, K.A.; Lemoine, S.; Waterhouse, N.J.; Vari, F.; Chatenoud, L.; Gandhi, M.K.; Martinet, L.; Smyth, M.J.; Guillerey, C. Human peripheral blood DNAM-1(neg) NK cells are a terminally differentiated subset with limited effector functions. Blood Adv 2019, 3, 1681–1694. [Google Scholar] [CrossRef]
- Raber, P.; Ochoa, A.C.; Rodriguez, P.C. Metabolism of L-arginine by myeloid-derived suppressor cells in cancer: mechanisms of T cell suppression and therapeutic perspectives. Immunol Invest 2012, 41, 614–634. [Google Scholar] [CrossRef]
- Elliott, L.A.; Doherty, G.A.; Sheahan, K.; Ryan, E.J. Human Tumor-Infiltrating Myeloid Cells: Phenotypic and Functional Diversity. Front Immunol 2017, 8, 86. [Google Scholar] [CrossRef]
- Bertuzzi, C.; Sabattini, E.; Agostinelli, C. Immune Microenvironment Features and Dynamics in Hodgkin Lymphoma. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Romano, A.; Parrinello, N.L.; Vetro, C.; Forte, S.; Chiarenza, A.; Figuera, A.; Motta, G.; Palumbo, G.A.; Ippolito, M.; Consoli, U.; et al. Circulating myeloid-derived suppressor cells correlate with clinical outcome in Hodgkin Lymphoma patients treated up-front with a risk-adapted strategy. Br J Haematol 2015, 168, 689–700. [Google Scholar] [CrossRef]
- Romano, A.; Parrinello, N.L.; Chiarenza, A.; Motta, G.; Tibullo, D.; Giallongo, C.; La Cava, P.; Camiolo, G.; Puglisi, F.; Palumbo, G.A.; et al. Immune off-target effects of Brentuximab Vedotin in relapsed/refractory Hodgkin Lymphoma. Br J Haematol 2019, 185, 468–479. [Google Scholar] [CrossRef]
- Axdorph, U.; Porwit-MacDonald, A.; Grimfors, G.; Bjorkholm, M. Tissue eosinophilia in relation to immunopathological and clinical characteristics in Hodgkin's disease. Leuk Lymphoma 2001, 42, 1055–1065. [Google Scholar] [CrossRef]
- von Wasielewski, R.; Seth, S.; Franklin, J.; Fischer, R.; Hubner, K.; Hansmann, M.L.; Diehl, V.; Georgii, A. Tissue eosinophilia correlates strongly with poor prognosis in nodular sclerosing Hodgkin's disease, allowing for known prognostic factors. Blood 2000, 95, 1207–1213. [Google Scholar] [CrossRef]
- Masucci, M.T.; Minopoli, M.; Carriero, M.V. Tumor Associated Neutrophils. Their Role in Tumorigenesis, Metastasis, Prognosis and Therapy. Front Oncol 2019, 9, 1146. [Google Scholar] [CrossRef]
- Koh, Y.W.; Kang, H.J.; Park, C.; Yoon, D.H.; Kim, S.; Suh, C.; Kim, J.E.; Kim, C.W.; Huh, J. Prognostic significance of the ratio of absolute neutrophil count to absolute lymphocyte count in classic Hodgkin lymphoma. Am J Clin Pathol 2012, 138, 846–854. [Google Scholar] [CrossRef]
- Molin, D.; Fischer, M.; Xiang, Z.; Larsson, U.; Harvima, I.; Venge, P.; Nilsson, K.; Sundstrom, C.; Enblad, G.; Nilsson, G. Mast cells express functional CD30 ligand and are the predominant CD30L-positive cells in Hodgkin's disease. Br J Haematol 2001, 114, 616–623. [Google Scholar] [CrossRef]
- Mizuno, H.; Nakayama, T.; Miyata, Y.; Saito, S.; Nishiwaki, S.; Nakao, N.; Takeshita, K.; Naoe, T. Mast cells promote the growth of Hodgkin's lymphoma cell tumor by modifying the tumor microenvironment that can be perturbed by bortezomib. Leukemia 2012, 26, 2269–2276. [Google Scholar] [CrossRef]
- Molin, D.; Edstrom, A.; Glimelius, I.; Glimelius, B.; Nilsson, G.; Sundstrom, C.; Enblad, G. Mast cell infiltration correlates with poor prognosis in Hodgkin's lymphoma. Br J Haematol 2002, 119, 122–124. [Google Scholar] [CrossRef]
- Keresztes, K.; Szollosi, Z.; Simon, Z.; Tarkanyi, I.; Nemes, Z.; Illes, A. Retrospective analysis of the prognostic role of tissue eosinophil and mast cells in Hodgkin's lymphoma. Pathol Oncol Res 2007, 13, 237–242. [Google Scholar] [CrossRef]
- Shodell, M.; Kempin, S.; Siegal, F.P. Plasmacytoid dendritic cell and CD4 + T cell deficiencies in untreated Hodgkin disease: implications for susceptibility to opportunistic infections. Leuk Lymphoma 2014, 55, 2656–2657. [Google Scholar] [CrossRef]
- Tudor, C.S.; Bruns, H.; Daniel, C.; Distel, L.V.; Hartmann, A.; Gerbitz, A.; Buettner, M.J. Macrophages and dendritic cells as actors in the immune reaction of classical Hodgkin lymphoma. PLoS One 2014, 9, e114345. [Google Scholar] [CrossRef]
- Alavaikko, M.J.; Blanco, G.; Aine, R.; Lehtinen, T.; Fellbaum, C.; Taskinen, P.J.; Sarpola, A.; Hansmann, M.L. Follicular dendritic cells have prognostic relevance in Hodgkin's disease. Am J Clin Pathol 1994, 101, 761–767. [Google Scholar] [CrossRef]
- Galati, D.; Zanotta, S.; Corazzelli, G.; Bruzzese, D.; Capobianco, G.; Morelli, E.; Arcamone, M.; De Filippi, R.; Pinto, A. Circulating dendritic cells deficiencies as a new biomarker in classical Hodgkin lymphoma. Br J Haematol 2019, 184, 594–604. [Google Scholar] [CrossRef]
- Hourani, T.; Holden, J.A.; Li, W.; Lenzo, J.C.; Hadjigol, S.; O'Brien-Simpson, N.M. Tumor Associated Macrophages: Origin, Recruitment, Phenotypic Diversity, and Targeting. Front Oncol 2021, 11, 788365. [Google Scholar] [CrossRef]
- Cencini, E.; Fabbri, A.; Sicuranza, A.; Gozzetti, A.; Bocchia, M. The Role of Tumor-Associated Macrophages in Hematologic Malignancies. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Xie, Y.; Yang, H.; Yang, C.; He, L.; Zhang, X.; Peng, L.; Zhu, H.; Gao, L. Role and Mechanisms of Tumor-Associated Macrophages in Hematological Malignancies. Front Oncol 2022, 12, 933666. [Google Scholar] [CrossRef]
- Carey, C.D.; Gusenleitner, D.; Lipschitz, M.; Roemer, M.G.M.; Stack, E.C.; Gjini, E.; Hu, X.; Redd, R.; Freeman, G.J.; Neuberg, D.; et al. Topological analysis reveals a PD-L1-associated microenvironmental niche for Reed-Sternberg cells in Hodgkin lymphoma. Blood 2017, 130, 2420–2430. [Google Scholar] [CrossRef]
- Patel, S.S.; Weirather, J.L.; Lipschitz, M.; Lako, A.; Chen, P.H.; Griffin, G.K.; Armand, P.; Shipp, M.A.; Rodig, S.J. The microenvironmental niche in classic Hodgkin lymphoma is enriched for CTLA-4-positive T cells that are PD-1-negative. Blood 2019, 134, 2059–2069. [Google Scholar] [CrossRef]
- Hancic, S.; Grskovic, P.; Gasparov, S.; Ostojic Kolonic, S.; Dominis, M.; Korac, P. Macrophage Infiltration Correlates with Genomic Instability in Classic Hodgkin Lymphoma. Biomedicines 2022, 10. [Google Scholar] [CrossRef]
- Werner, L.; Dreyer, J.H.; Hartmann, D.; Barros, M.H.M.; Buttner-Herold, M.; Grittner, U.; Niedobitek, G. Tumor-associated macrophages in classical Hodgkin lymphoma: hormetic relationship to outcome. Sci Rep 2020, 10, 9410. [Google Scholar] [CrossRef]
- Steidl, C.; Lee, T.; Shah, S.P.; Farinha, P.; Han, G.; Nayar, T.; Delaney, A.; Jones, S.J.; Iqbal, J.; Weisenburger, D.D.; et al. Tumor-associated macrophages and survival in classic Hodgkin's lymphoma. N Engl J Med 2010, 362, 875–885. [Google Scholar] [CrossRef]
- Touati, M.; Delage-Corre, M.; Monteil, J.; Abraham, J.; Moreau, S.; Remenieras, L.; Gourin, M.P.; Dmytruk, N.; Olivrie, A.; Turlure, P.; et al. CD68-positive tumor-associated macrophages predict unfavorable treatment outcomes in classical Hodgkin lymphoma in correlation with interim fluorodeoxyglucose-positron emission tomography assessment. Leuk Lymphoma 2015, 56, 332–341. [Google Scholar] [CrossRef]
- Cuccaro, A.; Annunziata, S.; Cupelli, E.; Martini, M.; Calcagni, M.L.; Rufini, V.; Giachelia, M.; Bartolomei, F.; Galli, E.; D'Alo, F.; et al. CD68+ cell count, early evaluation with PET and plasma TARC levels predict response in Hodgkin lymphoma. Cancer Med 2016, 5, 398–406. [Google Scholar] [CrossRef]
- Greaves, P.; Clear, A.; Coutinho, R.; Wilson, A.; Matthews, J.; Owen, A.; Shanyinde, M.; Lister, T.A.; Calaminici, M.; Gribben, J.G. Expression of FOXP3, CD68, and CD20 at diagnosis in the microenvironment of classical Hodgkin lymphoma is predictive of outcome. J Clin Oncol 2013, 31, 256–262. [Google Scholar] [CrossRef]
- Jakovic, L.R.; Mihaljevic, B.S.; Perunicic Jovanovic, M.D.; Bogdanovic, A.D.; Andjelic, B.M.; Bumbasirevic, V.Z. The prognostic relevance of tumor associated macrophages in advanced stage classical Hodgkin lymphoma. Leuk Lymphoma 2011, 52, 1913–1919. [Google Scholar] [CrossRef]
- Mohamed, O.; El Bastawisy, A.; Allahlobi, N.; Abdellateif, M.S.; Zekri, A.R.N.; Shaarawy, S.; Korany, Z.; Mohanad, M.; Bahnassy, A.A. The role of CD68+ macrophage in classical Hodgkin lymphoma patients from Egypt. Diagn Pathol 2020, 15, 10. [Google Scholar] [CrossRef]
- Yoon, D.H.; Koh, Y.W.; Kang, H.J.; Kim, S.; Park, C.S.; Lee, S.W.; Suh, C.; Huh, J. CD68 and CD163 as prognostic factors for Korean patients with Hodgkin lymphoma. Eur J Haematol 2012, 88, 292–305. [Google Scholar] [CrossRef]
- Tan, K.L.; Scott, D.W.; Hong, F.; Kahl, B.S.; Fisher, R.I.; Bartlett, N.L.; Advani, R.H.; Buckstein, R.; Rimsza, L.M.; Connors, J.M.; et al. Tumor-associated macrophages predict inferior outcomes in classic Hodgkin lymphoma: a correlative study from the E2496 Intergroup trial. Blood 2012, 120, 3280–3287. [Google Scholar] [CrossRef]
- Kamper, P.; Bendix, K.; Hamilton-Dutoit, S.; Honore, B.; Nyengaard, J.R.; d'Amore, F. Tumor-infiltrating macrophages correlate with adverse prognosis and Epstein-Barr virus status in classical Hodgkin's lymphoma. Haematologica 2011, 96, 269–276. [Google Scholar] [CrossRef]
- Agur, A.; Amir, G.; Paltiel, O.; Klein, M.; Dann, E.J.; Goldschmidt, H.; Goldschmidt, N. CD68 staining correlates with the size of residual mass but not with survival in classical Hodgkin lymphoma. Leuk Lymphoma 2015, 56, 1315–1319. [Google Scholar] [CrossRef]
- Kayal, S.; Mathur, S.; Karak, A.K.; Kumar, L.; Sharma, A.; Bakhshi, S.; Raina, V. CD68 tumor-associated macrophage marker is not prognostic of clinical outcome in classical Hodgkin lymphoma. Leuk Lymphoma 2014, 55, 1031–1037. [Google Scholar] [CrossRef]
- Klein, J.L.; Nguyen, T.T.; Bien-Willner, G.A.; Chen, L.; Foyil, K.V.; Bartlett, N.L.; Duncavage, E.J.; Hassan, A.; Frater, J.L.; Kreisel, F. CD163 immunohistochemistry is superior to CD68 in predicting outcome in classical Hodgkin lymphoma. Am J Clin Pathol 2014, 141, 381–387. [Google Scholar] [CrossRef]
- Azambuja, D.; Natkunam, Y.; Biasoli, I.; Lossos, I.S.; Anderson, M.W.; Morais, J.C.; Spector, N. Lack of association of tumor-associated macrophages with clinical outcome in patients with classical Hodgkin's lymphoma. Ann Oncol 2012, 23, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Zaki, M.A.; Wada, N.; Ikeda, J.; Shibayama, H.; Hashimoto, K.; Yamagami, T.; Tatsumi, Y.; Tsukaguchi, M.; Take, H.; Tsudo, M.; et al. Prognostic implication of types of tumor-associated macrophages in Hodgkin lymphoma. Virchows Arch 2011, 459, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Gusak, A.; Fedorova, L.; Lepik, K.; Volkov, N.; Popova, M.; Moiseev, I.; Mikhailova, N.; Baykov, V.; Kulagin, A. Immunosuppressive Microenvironment and Efficacy of PD-1 Inhibitors in Relapsed/Refractory Classic Hodgkin Lymphoma: Checkpoint Molecules Landscape and Macrophage Populations. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Cen, H.; Tan, X.; Ke, Q. Meta-analysis of the prognostic and clinical value of tumor-associated macrophages in adult classical Hodgkin lymphoma. BMC Med 2016, 14, 159. [Google Scholar] [CrossRef] [PubMed]
- Karihtala, K.; Leivonen, S.K.; Bruck, O.; Karjalainen-Lindsberg, M.L.; Mustjoki, S.; Pellinen, T.; Leppa, S. Prognostic Impact of Tumor-Associated Macrophages on Survival Is Checkpoint Dependent in Classical Hodgkin Lymphoma. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef]
- Locatelli, S.L.; Careddu, G.; Serio, S.; Consonni, F.M.; Maeda, A.; Viswanadha, S.; Vakkalanka, S.; Castagna, L.; Santoro, A.; Allavena, P.; et al. Targeting Cancer Cells and Tumor Microenvironment in Preclinical and Clinical Models of Hodgkin Lymphoma Using the Dual PI3Kdelta/gamma Inhibitor RP6530. Clin Cancer Res 2019, 25, 1098–1112. [Google Scholar] [CrossRef]
- Liu, Y.; Xia, Y.; Qiu, C.H. Functions of CD169 positive macrophages in human diseases (Review). Biomed Rep 2021, 14, 26. [Google Scholar] [CrossRef]
- Grabowska, J.; Lopez-Venegas, M.A.; Affandi, A.J.; den Haan, J.M.M. CD169(+) Macrophages Capture and Dendritic Cells Instruct: The Interplay of the Gatekeeper and the General of the Immune System. Front Immunol 2018, 9, 2472. [Google Scholar] [CrossRef]
- Affandi, A.J.; Olesek, K.; Grabowska, J.; Nijen Twilhaar, M.K.; Rodriguez, E.; Saris, A.; Zwart, E.S.; Nossent, E.J.; Kalay, H.; de Kok, M.; et al. CD169 Defines Activated CD14(+) Monocytes With Enhanced CD8(+) T Cell Activation Capacity. Front Immunol 2021, 12, 697840. [Google Scholar] [CrossRef]
- Asano, K.; Nabeyama, A.; Miyake, Y.; Qiu, C.H.; Kurita, A.; Tomura, M.; Kanagawa, O.; Fujii, S.; Tanaka, M. CD169-positive macrophages dominate antitumor immunity by crosspresenting dead cell-associated antigens. Immunity 2011, 34, 85–95. [Google Scholar] [CrossRef]
- Komohara, Y.; Ohnishi, K.; Takeya, M. Possible functions of CD169-positive sinus macrophages in lymph nodes in anti-tumor immune responses. Cancer Sci 2017, 108, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Ohnishi, K.; Shiota, T.; Motoshima, T.; Sugiyama, Y.; Yatsuda, J.; Kamba, T.; Ishizaka, K.; Komohara, Y. CD169-positive sinus macrophages in the lymph nodes determine bladder cancer prognosis. Cancer Sci 2018, 109, 1723–1730. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.Q.; Jiang, Z.Z.; Li, L.; Wu, Y.; Zheng, L. CD169 identifies an anti-tumour macrophage subpopulation in human hepatocellular carcinoma. J Pathol 2016, 239, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Briem, O.; Kallberg, E.; Kimbung, S.; Veerla, S.; Stenstrom, J.; Hatschek, T.; Hagerling, C.; Hedenfalk, I.; Leandersson, K. CD169(+) Macrophages in Primary Breast Tumors Associate with Tertiary Lymphoid Structures, T(regs) and a Worse Prognosis for Patients with Advanced Breast Cancer. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, K.; Komohara, Y.; Saito, Y.; Miyamoto, Y.; Watanabe, M.; Baba, H.; Takeya, M. CD169-positive macrophages in regional lymph nodes are associated with a favorable prognosis in patients with colorectal carcinoma. Cancer Sci 2013, 104, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Ohnishi, K.; Miyashita, A.; Nakahara, S.; Fujiwara, Y.; Horlad, H.; Motoshima, T.; Fukushima, S.; Jinnin, M.; Ihn, H.; et al. Prognostic Significance of CD169+ Lymph Node Sinus Macrophages in Patients with Malignant Melanoma. Cancer Immunol Res 2015, 3, 1356–1363. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, K.; Yamaguchi, M.; Erdenebaatar, C.; Saito, F.; Tashiro, H.; Katabuchi, H.; Takeya, M.; Komohara, Y. Prognostic significance of CD169-positive lymph node sinus macrophages in patients with endometrial carcinoma. Cancer Sci 2016, 107, 846–852. [Google Scholar] [CrossRef]
- Marmey, B.; Boix, C.; Barbaroux, J.B.; Dieu-Nosjean, M.C.; Diebold, J.; Audouin, J.; Fridman, W.H.; Mueller, C.G.; Molina, T.J. CD14 and CD169 expression in human lymph nodes and spleen: specific expansion of CD14+CD169- monocyte-derived cells in diffuse large B-cell lymphomas. Hum Pathol 2006, 37, 68–77. [Google Scholar] [CrossRef]
- Liu, W.R.; Shipp, M.A. Signaling pathways and immune evasion mechanisms in classical Hodgkin lymphoma. Blood 2017, 130, 2265–2270. [Google Scholar] [CrossRef]
- Kawashima, M.; Higuchi, H.; Kotani, A. Significance of trogocytosis and exosome-mediated transport in establishing and maintaining the tumor microenvironment in lymphoid malignancies. J Clin Exp Hematop 2021, 61, 192–201. [Google Scholar] [CrossRef]
- Roemer, M.G.M.; Redd, R.A.; Cader, F.Z.; Pak, C.J.; Abdelrahman, S.; Ouyang, J.; Sasse, S.; Younes, A.; Fanale, M.; Santoro, A.; et al. Major Histocompatibility Complex Class II and Programmed Death Ligand 1 Expression Predict Outcome After Programmed Death 1 Blockade in Classic Hodgkin Lymphoma. J Clin Oncol 2018, 36, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Menendez, V.; Solorzano, J.L.; Fernandez, S.; Montalban, C.; Garcia, J.F. The Hodgkin Lymphoma Immune Microenvironment: Turning Bad News into Good. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Hatic, H.; Sampat, D.; Goyal, G. Immune checkpoint inhibitors in lymphoma: challenges and opportunities. Ann Transl Med 2021, 9, 1037. [Google Scholar] [CrossRef] [PubMed]
- Vardhana, S.; Younes, A. The immune microenvironment in Hodgkin lymphoma: T cells, B cells, and immune checkpoints. Haematologica 2016, 101, 794–802. [Google Scholar] [CrossRef] [PubMed]
- El Halabi, L.; Adam, J.; Gravelle, P.; Marty, V.; Danu, A.; Lazarovici, J.; Ribrag, V.; Bosq, J.; Camara-Clayette, V.; Laurent, C.; et al. Expression of the Immune Checkpoint Regulators LAG-3 and TIM-3 in Classical Hodgkin Lymphoma. Clin Lymphoma Myeloma Leuk 2021, 21, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Abdul Razak, F.R.; Terpstra, M.; Chan, F.C.; Saber, A.; Nijland, M.; van Imhoff, G.; Visser, L.; Gascoyne, R.; Steidl, C.; et al. The mutational landscape of Hodgkin lymphoma cell lines determined by whole-exome sequencing. Leukemia 2014, 28, 2248–2251. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Schneider, S.; Zuhlke-Jenisch, R.; Klapper, W.; Sundstrom, C.; Hartmann, S.; Hansmann, M.L.; Siebert, R.; Kuppers, R.; Giefing, M. Alterations of the CD58 gene in classical Hodgkin lymphoma. Genes Chromosomes Cancer 2015, 54, 638–645. [Google Scholar] [CrossRef]
- Casagrande, N.; Borghese, C.; Aldinucci, D. Current and Emerging Approaches to Study Microenvironmental Interactions and Drug Activity in Classical Hodgkin Lymphoma. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Choe, J.Y.; Yun, J.Y.; Jeon, Y.K.; Kim, S.H.; Park, G.; Huh, J.R.; Oh, S.; Kim, J.E. Indoleamine 2,3-dioxygenase (IDO) is frequently expressed in stromal cells of Hodgkin lymphoma and is associated with adverse clinical features: a retrospective cohort study. BMC Cancer 2014, 14, 335. [Google Scholar] [CrossRef]
- Kim, M.S.; Park, T.I.; Son, S.A.; Lee, H.W. Immunohistochemical Features of Indoleamine 2,3-Dioxygenase (IDO) in Various Types of Lymphoma: A Single Center Experience. Diagnostics (Basel) 2020, 10. [Google Scholar] [CrossRef]
- Masaki, A.; Ishida, T.; Maeda, Y.; Ito, A.; Suzuki, S.; Narita, T.; Kinoshita, S.; Takino, H.; Yoshida, T.; Ri, M.; et al. Clinical significance of tryptophan catabolism in Hodgkin lymphoma. Cancer Sci 2018, 109, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Marshall, N.A.; Christie, L.E.; Munro, L.R.; Culligan, D.J.; Johnston, P.W.; Barker, R.N.; Vickers, M.A. Immunosuppressive regulatory T cells are abundant in the reactive lymphocytes of Hodgkin lymphoma. Blood 2004, 103, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the spectrum of macrophages in the Hodgkin lymphoma microenvironment.
Figure 1.
Schematic representation of the spectrum of macrophages in the Hodgkin lymphoma microenvironment.
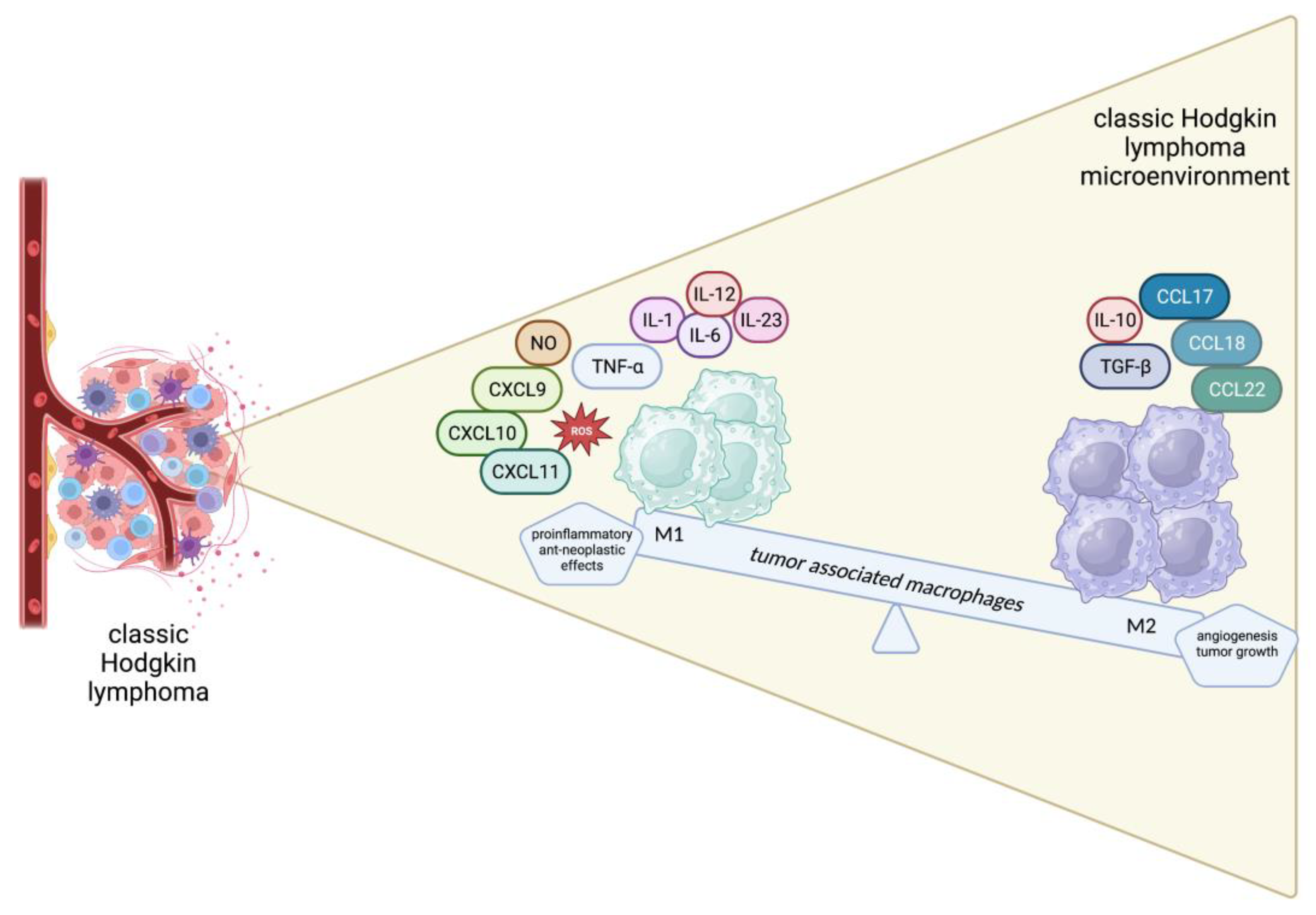
Figure 2.
Prognostic role of CD169+ macrophages in several solid malignancies.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



