
Submitted:
15 May 2023
Posted:
17 May 2023
You are already at the latest version
Abstract
Montes de María is the best-preserved tropical dry forest fragment in the Colombian Caribbean, making it an ideal location for studying the impacts of human disturbance on local ecosystems. This study analyzed the ecological structure of diurnal butterflies in both forested and disturbed areas using 16 circular plots to identify relationships between alpha and beta diversity, and the geographic distance between disturbed areas and native forests, using a range of metrics, including range-abundance and rarefaction curves, nonlinear models, and the Bray–Curtis dissimilarity index. Results revealed three distinct species assemblages associated with forest, intermediate disturbed areas (IDA), and disturbed areas (DA). Nonlinear models show that IDA are more diverse than forest and DA. However, forests have more beta diversity among plots than IDA and DA. Indicator species for each butterfly assemblage were also identified. Thus, although new butterfly species assemblages emerge from a new human land-scape, it is clear that species that only occur within dry forest fragments are lost when forest fragments disappear. Overall, these findings have important implications for conservation efforts and understanding how human disturbance affects biodiversity in tropical ecosystems.
Keywords:
structure
; diversity
; anthropic disturbance
; conservation
; Lepidoptera
1. Introduction
Neotropical seasonally dry forest (NSDF) are one of the most biodiverse ecosystems in the world [1], and in the Americas, these are distributed discontinuously from Mexico to Argentina and throughout the Caribbean [2]. In the Neotropical region, these forests are highly threatened by human intervention [2], mainly due to logging, agriculture, and livestock, and suffer the effects of the expansion of urban areas [3]. In Colombia, these forests are distributed mainly over the inter-Andean valleys, with some isolated fragments towards southern regions of the country in the Patía River valley and, to a greater extent, in the Colombian Caribbean [3]. Currently, less than 4% of the original biome cover remains, and another 5% represent relics with some degree of anthropic disturbance. In other words, only less than 10% of NSDF remains in Colombia [3]. In fact, of the 533,099 ha that historically compose the dry forests of the Caribbean, only 37.97% still remain in a relatively good state of conservation, suggesting a strong anthropogenic disturbance impact on biotic communities [4,5].
A disturbance is, by definition, a discrete event in time that alters the ecosystem, community, population structure, resources, substrate availability, or the physical environment [6]. The understanding of the patterns that comprise communities in disturbed sites is currently a central objective in ecology, primarily since it is known that a disturbance affects the temporal or spatial variability of community structure (beta diversity) or both, as well as species richness (alpha diversity) [7]. Disturbances can cause convergence in community composition (lower beta diversity) by increasing suitable habitats for disturbance-tolerant species [8]. Alternatively, disturbance can lead to divergence in community composition (high beta diversity) by increasing the environmental filter [9].
One of the challenges in studying disturbance is understanding the mechanisms by which species diversity is maximized in a heterogeneous landscape [10]. Hypothetically, the areas closest to the forests are little disturbed, and the degree of disturbance increases at a larger distance, passing through areas of intermediate disturbance before reaching highly disturbed areas. In this sense, the intermediate disturbance hypothesis (IDH) is one of the most fundamental concepts. According to the IDH, there is a unimodal relationship between disturbance and diversity. An intermediate level of disturbance leads to higher levels of alpha biodiversity [11] due to compensations between the ability of species to compete, colonize areas, and tolerate disturbance [12]. Although the IDH has influenced ecological theory, management, and conservation, its predictions are not always accurate [13].
In butterflies, e.g., diversity increases significantly as disturbance frequency increases [14,15]. Conversely, Addo-Fordjour, et al. [16] and León-Cortés, et al. [17] have found that the diversity of butterflies in disturbed habitats is considerably lower than in semi-natural preserved areas. Nevertheless, the mechanisms by which the disturbance influences the community structure are not generalizable since they depend on how the impact on the biota modifies the environmental conditions and the colonization of species [15,18]. However, how disturbances affect the community structure is unknown in tropical dry forest butterflies.
As sensitive to ecosystem changes, butterflies are a model group to understand how human actions have influenced the community structure of tropical dry forest biota [14,15,16,17,19]. Butterflies as model groups would be a strategy to generate information on the relationship of adjacent areas on the faunal diversity within dry forests without the need to explore all the biological groups, i.e., from an autoecological perspective [20].
Communities can be structured taxonomically/phylogenetically when co-occurring species are highly or lowly phylogenetically related [21]. In the former, species communities shape the so-called phylogenetic clustering, and in the latter, phylogenetic overdispersion [21]. Some studies in disturbed ecosystems have found phylogenic overdispersal in sites with intermediate disturbance [22,23]. Nonetheless, there is also evidence that the disturbance generates an environmental filter that only allows the growth of phylogenetically related species, and, therefore, a phylogenetic grouping is generated [24,25]. However, as well as richness, the behavior of the phylogenetic structure is not a pattern that can be extended to all taxonomic groups, and, therefore, the impact of disturbance on dry forest butterfly communities is little known [24].
Thus, analyzing the effect of disturbance on the structure of the Lepidoptera-Rophalocera butterfly communities of the tropical dry forest would provide a better understanding of how alpha and beta diversity are altered in areas with a disturbance gradient. In this sense, Montes de María, in the Colombian Caribbean, constitutes a unique setting to study how the species assemblages change in environments modified by man in the tropical dry forest –mainly increasing– and the knowledge that helps biodiversity preservation and restoration.
Montes de María has been classified as one of the dry forest areas in the best state of conservation, mainly due to the implementation of the Protective Forest Reserve Serranía de Coraza and Montes de María [26]. However, agricultural activities have generated a mosaic landscape comprised of patches of forest vegetation, pastures, and crops [5,27], and, therefore, human disturbance has modified the diversity and composition of butterflies. Nevertheless, how disturbance has structured the butterfly community is unknown. Herein we analyze the effect of disturbance on the community structure of five Lepidoptera-Rophalocera diurnal families (Papilionidae, Nymphalidae, Pieridae, Riodinidae, and Lycaenidae) currently distributed in seasonally dry forests of the Coraza reserve, department of Sucre, Colombia. Under this approach, the following questions want to be answered: How does species richness (alpha diversity) and turnover (beta diversity) change according to the disturbance gradient in this area? Are there assemblages of species that allow differentiating areas with different degrees of disturbance, within which a homogenization process in beta diversity and taxonomic/phylogenetic structure has been generated in the most disturbed communities? If these assemblages have been shaped to the degree of disturbance, are there indicator species for each of the assemblages?
2. Materials and Methods
2.1. Study Area
The Protective Forest Reserve Serranía de Coraza is located in the municipality of Colosó in Montes de María. Montes de María is situated in the northeastern region of the department of Sucre in the Colombian Caribbean plain [26] (Figure 1). Coraza shows temperatures between 25-28°C and precipitation of 896-1,233 mm per year, with a dry seasonal period from November to February where rainfall decreases to less than 200 mm. The average relative humidity is 83.5% [26,28].
The vegetation type belongs to a tropical dry forest with Fabaceae, Malvaceae, Meliaceae, Sapindaceae, Capparaceae, Rubiaceae, and Cactaceae as the most important plant families. The most abundant species are Ampelocera edentula Kuhlm, Aspidosperma polyneuron Mull. Arg, Brosimum alicastrum Swartz, Myrcia fallax (Rich.) DC., and Simira cordifolia (Hook. F) Steyerm [29]. However, like much of the Colombian tropical dry forest, these forest fragments are found in a mosaic of vegetation comprised of forests, crops, and grazing-livestock areas [5].
2.2. Biological Material
Three linear transects of approximately 1,200 m long were designed to identify the impact of the disturbance on the butterfly communities of the tropical dry forest in the study area. Sixteen circular plots were carried out, eight in the forest and eight in disturbed areas (pasture and crops). Each circular plot has a radius of 15 m and were spread from each other by 60 m [30]. A total of 48 plots (Figure 1) were sampled. Within each plot, butterfly individuals were collected using an entomological net, and Van Someren Rydon traps baited with an attractive mixture of banana, rum, and sugarcane juice to attract frugivorous butterflies. These include typical species of tropical and subtropical areas, mainly of the Nymphalidae family [31]. Trap sampling was carried out during five effective hours per plot, located 2 m from the ground. Once the traps were installed, they were visited twice daily to collect the specimens. The collection with an entomological net was carried out between 8:00 and 16:00 hours by three fieldwork personnel in each plot, with a total effort of 5 h of sampling per plot. Hours to collect species with crepuscular habits were not included, as the sociopolitical conditions of the area did not guarantee the safety of the personnel. Although these families show a relatively stable taxonomy compared to other groups [32], e.g., Hesperiidae exhibit complexity and taxonomy problems in many genera, leading to underestimating their diversity in ecological studies. Therefore, this family can muddy the outcome of community structure studies [33]. Hence, butterflies from all families were collected, excluding the Hesperiidae family.
The sampling was carried out during five field trips lasting eight days each between October and November 2014 and April and May 2015, where the highest rainfall peaks occurred, coinciding with the butterfly population peaks, according to Freitas, Iserhard, Santos, Junia, Danilo, Douglas, Augusto, Onildo, Gustavo and Mauricio [31]. Once the material was collected, it was stored in Milano paper envelopes [34] and transferred to the laboratory for later identification. Data regarding plot number, locality (department and municipality), date, name of the collector, altitude, and geographic coordinates were collected for each specimen.
2.3. Taxonomical Composition
The captured specimens were rehydrated in humid chambers for a minimum of 24 h for their softening and subsequent assembly in the Biological Conservation Laboratory of Universidad de Sucre. Wing extension was performed following the protocol suggested by Triplehorn and Johnson [36]. Taxonomic identification was carried out mainly by comparison with photographs of type specimens deposited on the "Butterflies of America" website [37], field guides [38,39,40], and material deposited in the Entomological Museum of Universidad Nacional de Colombia, Medellín headquarters. For those individuals for whom it was not possible to approximate to a taxonomic entity through morphology, it was necessary to perform dissections of the genital organs using a liquid 10% KOH solution in a water bath for 15 min following the methodology of Andrade-C, et al. [41], for later observation in a stereoscope at 35X (Leika K100, ) and accurate taxonomic identification.
2.4. Community Structure Analyses
The expected number of species per cover type (forest and disturbed areas) was calculated through the rarefaction and extrapolation-interpolation curves to establish whether the samplings were representative of each transect. This method, described by Chao, et al. [42], uses the sample and a completeness curve made with twice the size of the smallest reference sample to be compared, with a 95% confidence interval obtained by resampling 100 bootstrap pseudoreplicates. This analysis was implemented with the R iNEXT package [43] following the parameters established by Chao, Gotelli, C, Elizabeth, MA, Colwell and Ellison [42] and Colwell, et al. [44].
The distances of each plot within each transect were taken from Google Earth®, and the watersheds of the Coraza reserve were used as a reference base to establish if there are assemblages of species related to areas with different degrees of disturbance and evaluate the intermediate disturbance hypothesis. Watersheds are defined as lines drawn on the highest peaks or pinnacles that enclose a basin area; therefore, these are the least accessible areas for cultivation and livestock [45] and the least disturbed by such activities. For the current study, these divisions correspond to the forest areas most distant from the disturbed zones.
The above data were used to evaluate the change in richness and abundance associated with the distance from each plot to the least disturbed area of its transect (watershed), using linear and nonlinear regression models (linear, quadratic, potential, exponential, logistic, and Gaussian). Model fit was assessed using the Akaike Information Criterion (AIC) [46], where the value closest to zero AIC indicates the best fit to the data. There is a relationship between species composition (richness and abundance) and geographic distance described by a Gaussian model, with mean maximum values in plots of intermediate distances (1,001-1,250 m), both for richness (Figure 2A) and for abundance (Figure 2B). Based on these results, nine ranges were generated based on the distance from the watershed to the most disturbed areas (0-250 m, 251-500 m, 501-750 m, 751-1,000 m, 1001-1,250 m, 1,251-1,500 m, 1,501-1,750 m, 1,800-2,050 m, and >2,050).
To establish assemblages of species possibly related to areas with different types of disturbance levels using the beta diversity [47] through the Bray-Curtis index (βBRY) together with their balanced variation components (βBRY.BAL) and abundance gradients (βBRY.GR) a new species and range abundance matrix was built. Beta diversity, according to Baselga [47] metrics were calculated utilizing the R Betapart package [48]. Results of the dissimilarity matrices from Bray-Curtis, allowed the establishment of three assemblages: (i) Forest (F), (ii) intermediate disturbed (IDA), and (iii) disturbed (DA) areas. Subsequently, a new species dataset was constructed, which, together with the three types of areas (F, IDA, and DA), were used to analyze whether there are homogenization processes of beta diversity in areas with higher disturbance. In this order, the effective number of communities (βD) and the Whitaker beta diversity index (βw) were used. In the first case, the multiplicative gamma partition βD= ϒD/ αD [49] in the R Entropart package was employed [50]. In the second case, the Whitaker beta diversity index (βw) was calculated as β= ϒ/ α in the PAST version 3.2 program Hammer, et al. [51].
Analyzes to explore how butterfly community structure responds to disturbance through taxonomic/phylogenetic homogenization or heterogenization processes that allow comparing communities from the point of view of the taxonomic/phylogenetic relationships between species were carried out [52]. Warwick and Clarke [53] generated indices to approach the phylogenetic structure of communities measuring divergence and phylogenetic regularity [54]. The taxonomic distinctiveness index (Δ*) represents the taxonomic relationships between the species of an assemblage without considering richness or abundance. Therefore, it is considered the most sensitive univariate measure to analyze community structure [53]. The taxonomic distinctiveness variation index (Λ+) allows the detection of the taxonomic equitability of assemblages, i.e., it measures the degree to which specific taxa are over or underrepresented in the samples [55,56]. The taxonomic indices were calculated with the PRIMER v.6 program.
Finally, an indicator species analysis (IndVal%) was carried out to establish if there are indicator species in each assemblage. This methodology is key to identifying those taxa associated with greater weight in the different units that comprise the landscape, being of higher relevance those that identify the forest or an undisturbed zone as a reference ecosystem for the area [57]. The species considered typical (indicators) of a habitat condition were those with InVal≥50% [58]. These analyzes were performed in the PAST 4.11 program [51].
3. Results
3.1. Taxonomic Composition
Eight hundred and ninety individuals from five families were collected, belonging to 15 subfamilies, 27 tribes, 63 genera, and 80 species (Table 1). The most representative family was Nymphalidae, with 605 individuals representing 67% of the catches, followed by Pieridae (23%), Papilionidae (7%), Riodinidae (2%), and Lycaenidae (1%). Likewise, concerning richness, the following families are highlighted: Nymphalidae (62%), Pieridae (17%), Papilionidae (9%), Riodinidae (5%), and Lycaenidae (5%). The subfamilies with the highest abundance were Biblidinae (272 individuals), Nymphalinae (159 individuals), and Coliadinae (138 individuals).
The most abundant species for the forest are H. februa (56 individuals), Junonia genoveva (Cramer, 1780) (33 individuals), Myscelia leucocyana (C. Felder & R. Felder1861) (27 individuals), Nica flavilla (Godart, 1824) (18 individuals) and Zaretis ellops (Menetries, 1855) (16 individuals) that represent 17% of the total number of individuals collected in the study. On the other hand, in the disturbed areas (pasture and crops), 11 species were recorded as the most abundant. The most representative are J. evarete (111 individuals), Eurema daira (62 individuals), Mestra hersilia (Fabricius, 1776) (60 individuals), Hamadryas feronia (Linnaeus, 1758) (35 individuals), and Ascia monuste (Linnaeus, 1764) (30 individuals).
3.2. Ecological Structure of Communities
The species completeness of the study area was 89.04%, suggesting that the sampling was representative, reaching 93.7% for the forest and 97.1% for the disturbed areas (Figure 3). For each type of cover, sampling completeness estimates differ slightly by increasing the size of the reference sample in terms of individuals to double (from 302 to 604 in forest and 600 to 1,200 in disturbed areas). This means that, even if more individuals are collected in the study area, the sampling continues to represent both the forest and disturbed areas (Figure 3A). When comparing the species richness concerning the number of individuals between the types of coverage (forest and disturbed areas), it was possible to establish, with a confidence interval of 95%, that the expected richness of butterflies would continue to be higher in disturbed areas than in forests even when all extant species were recorded (Figure 3B).
The cluster analysis results based on Bray-Curtis allowed distinguishing three large groups, within which all plots located between 250-750 m correspond to the forest area, those between 751-1,500 m to IDA, and those between 1,800-2,050 to DA (Figure 4). In the case of nesting and species turnover results, βBRY.BAL = 0.937 and βBRY.GRA = 0.027. The beta diversity analyses show a general pattern in which the forest area records higher values (βD = 1.63; βw =0.94). On the contrary, IDA registers lower values than the forest area (βD =1.51; βw = 0.61), and DA has lower values than IDA (βD = 1.48; βw =0.60).
The taxonomic distinctiveness model (Δ*) and its variation (Λ+) built at the level of the communities analyzed (F, IDA, and DA) showed that they are within the 95% confidence intervals. In this sense, Δ* was 49.56 for F, 52.26 for IDA, and 52.11 for DA. On the other hand, the variation values in taxonomic distinctiveness (Λ+) were 92.41 for F, 102.51 for IDA, and 90.30 for DA.
A total of 21 specific species were found for B (undisturbed), IDA, and DA assemblages through the InVal analysis (Figure 5). In the forest, a total of nine species were found, among which Archaeoprepona demophon, Hamadryas februa februa, Itaballia demophile calydonia, Morpho helenor, Morpho helenor peleides, Myscelia leucocyana, Nica flavilla, Siderone galanthis and Zaretis ellops are typical of this cover type (InVal≥50%, p=0.001). In the case of areas with intermediate disturbance, with values of InVal≥50% and p=0.001, Biblis hyperia, Eurema daira, Hamadryas feronia, Junonia evarete, Mestra hersilia, Phoebis sennae, and Temenis laothoe are specific taxa of these areas. Finally, in disturbed areas, only Battus polydamas, Danaus eresimus, Danaus gilippus, and Dryadula phaetusa are typical species of these areas (InVal≥60% and p=0.001).
4. Discussion
4.1. Taxonomic Composition
In terms of composition, the diversity of families in Montes de María is similar to those found in other tropical dry forest fragments in Colombia [59]. Nymphalidae was the most representative family regarding the number of individuals and species recorded in most studies in Colombia. In this sense, in the Patía River basin, Millan, et al. [60], Gaviria-Ortiz and Henao-Bañol [61], and Henao-Bañol and Andrade-C [62] registered 60 and 90 species, respectively, while [63] identified 78. In the Magdalena River valley, Peña and Reinoso [64] found 64 species, while for the Caribbean, Prince-Chacon, et al. [65] reported 38. Lamas (2004) stated that Riodinidae has the highest richness after Nymphalidae. However, these groups, together with the Lycaenidae family, have recorded the lowest species richness in other studies of butterflies in the tropical dry forest [63,66], mainly due to their small size and dark colors [67]. In addition, most of the species of these families tend to fly in higher strata and rest in the upper part of the trees, making its capture difficult within the forest [68].
Biblidinae is the most abundant subfamily with the highest richness. Regarding studies on the richness of this subfamily in dry forest fragments in the Caribbean region, Montero, et al. [69] and Vargas Zapata, et al. [70] found that their species showed the highest values with 13 and 10 taxa, respectively, contrary to the study reported by Vargas-Zapata, et al. [71] in the department of Atlántico with only six species. In this context, Vargas Zapata, Martinez Hernandez, Gutierrez Moreno, Prince Chacon, Herrera Colon and Torres Periñan [70] point out that the high richness of this subfamily may be related to the ability of its species to use a wide variety of resources in different plant strata. The abundance results of Biblidinae may be related to their larval stage since these groups feed on plants belonging to the Malvaceae, Euphorbiaceae, Moraceae, and Sapindaceae families [72], typical of these dry ecosystems. Herazo-Vitola, Mendoza-Cifuentes and Mercado-Gómez [29] point out that the Malvaceae, Sapindaceae, and Euphorbiaceae are of great relevance in terms of richness and dominance in the flora of Montes de María.
4.2. Community Structure Analysis
The results obtained in the current study suggest that, in the event of the transformation of the tropical dry forest of Montes de María by the expansion of the agricultural frontier (crops and pastures), species loss will occur, and, thus, the transformation of the community structure of butterflies in the forest. The regression outcomes and the cluster analyses based on the dissimilarity matrices allowed identifying species assemblages correlated with the geographical distance (m). Preserved forest areas and areas transformed by crops and pastures were found. Three zones were highlighted in the present study: undisturbed forest area, intermediate disturbed area, and disturbed area. The regressions show that forest areas are characterized by having low diversity, increasing in the intermediate disturbed areas and decreasing in the disturbed areas.
These data support the intermediate disturbance hypothesis proposed by Connell [73], where an increase in diversity is observed in intermediate areas. This pattern in diversity has also been found in humid forests where butterfly diversity values are higher in an intermediate area than in the disturbed forest [16,17,74]. The formation of these species assemblages may be the product of the long history of anthropic transformation that the Coraza reserve has endured in Montes de María. When forest fragments are cleared for cattle grazing or crop cultivation, the habitat for butterflies disappears, leading to the dispersion of species to nearby fragments.
Increases in species richness are often due to the invasion of disturbed areas by generalist and widely distributed species [19]. According to Vanschoenwinkel, et al. [75], diversity increases because many forest species are more tolerant to disturbance than expected by chance. Thus, extinction rates mediated by stochastic events such as crop and pasture implementation are not necessarily deterministic (i.e., species have similar extinction probabilities). In other words, disturbance can promote alpha diversity under these conditions.
Disturbance was also found to affect another butterfly community structure dimension. Beta diversity decreases as disturbance increases. The forest has higher beta diversity compared to IDA and DA; however, IDA is higher than DA. The diversity homogenization process of the most disturbed communities may result from the ecological filter [76]. Environmental conditions, e.g., higher temperature and solar radiation in disturbed areas compared to forest areas can create barriers that prevent not only movement between forest species and disturbed areas but also the formation of communities with some functional traits that increase their tolerance towards more environmentally stressful areas such as pastures and crops [77,78]. However, these aspects must be evaluated in-depth, where the functional traits of butterflies and an analysis of their phylogenetic structure can be measured.
The divergence and phylogenetic regularity analysis measured through the taxonomic distinctiveness of the three types of assemblages show no apparent differences among IDA, forest, and DA. However, IDA are slightly more diverse than DA and forest. These outcomes suggest that species of IDA have a slightly higher phylogenetic separation between species or greater evolutionary distances between their taxa than the rest of the assemblages [79]. Nevertheless, although the IDA assemblage has a higher species richness, the distribution of its species in the higher taxonomic categories (phylogenetic divergence) has a value equivalent to that of the DA assemblage [79].
Conversely, the low values in the forest indicate that the species are more closely related at lower taxonomic levels (e.g., several species belonging to the same genus or family). Therefore, these species are more phylogenetically related than those in disturbed areas. This study proposes two arguments to explain the high phylogenetic relationship or a phylogenetic grouping in dry forest butterfly communities [80,81]. The first is based on ecological forces such as the ecological or environmental filter, within which the environment is a filter that only allows species with particular traits or phenotypes to establish and persist in the forest [82]. The second is more evolutive; a phylogenetic conservatism niche was formed, and, therefore, closely related species are more similar in ecological, morphological, and functional traits than distantly related species since they have inherited it from their ancestors [83].
On the other hand, outcomes from IDA and DA show a community with higher phylogenetic overdispersion, i.e., with a higher number of less taxonomically related taxa. Webb, et al. [84] suggest that overdispersion in communities is mainly the product of competitive exclusion. Mostly taxonomically related species have a higher morphological similarity in their behavior and eating habits. Similar species tend to compete for the same resources when exposed to a new niche. However, those that become the leading taxa have greater ease in adapting to obtain resources more easily, generating exclusion by competition of other phylogenetically related species [21].
In the case of the Λ+ results, a difference from Δ* is observed, as IDA and the forest area have higher values than DA. The high values of Λ+ are related to an excessive or insufficient representation of some taxonomic groups (irregularities in the taxonomic tree). In IDA and forest, species are concentrated in the subfamilies Charaxinae, Coliadinae, Papilioninae, and Nymphalinae. The low Λ+ values in the disturbed areas indicate that the variance in the distribution of their species in the higher taxonomic categories is lower (lower Λ+ value), which can be reflected in the fact that their species are grouped mainly in Coliadinae. In other words, the supraspecific distribution is more equitable than the assemblages in the areas with intermediate disturbance and the forest [79]. These results suggest that the species found in the areas with intermediate disturbance show a phylogenetic regularity similar to that of the forest. Therefore, the influence of ecological processes, such as competition, has influenced the under or overrepresentation of taxa from their communities. However, in both cases, it is necessary to analyze functional traits to obtain a higher precision on what ecological or evolutionary processes influenced the butterfly communities in the interior at the two levels of disturbance and the forest.
Another effect of the disturbance on butterflies in dry forests is not only the generation of new communities but also that the species in these assemblages have adapted to their new habitats to such an extent that their distribution is restricted to these new areas, so they no longer occur in the forest. Biblis hyperia, Eurema daira, Hamadryas feronia, Junonia evarete, Mestra hersilia, Phoebis sennae, Temenis laothoe, Battus polydamas, Danaus eresimus, Danaus gilippus, and Dryadula phaetusa are exclusive species in IDA and DA. When a forest is disturbed, species of butterflies sensitive to disturbance tend to disappear, while the more tolerant species persist [85]. This study suggests that the species of butterflies associated with IDA and DA can be considered tolerant taxa toward extreme anthropic events because they can efficiently take advantage of the resources the environment offers [86].
An indicative species of the forest area is Archaeoprepona demophon (Linnaeus, 1758), belonging to the subfamily Charaxinae. A. demophon prefers decomposing fruits, which is supported by the large number of individuals found in the baited traps. Forests in the study area have species of the Sapindaceae, Moraceae, Anacardiaceae, and Myrtaceae families that produce sweet and edible fruits, which decompose when falling to the ground, and butterflies can absorb their sugars [60]. Morpho helenor was found inside the forest and depends on little-disturbed stream edges and forest interiors [60] because it has complex habitat requirements. In addition, species such as Hamadryas februa februa, Itaballia demophile calydonia, Myscelia leucocyana, Nica flavilla, Siderone galanthis, and Zaretis ellops were also exclusive to the forest. This can be explained by the availability of resources for adults, host plants for larvae, or the environmental humidity due to nearby water sources [63] offered by forests, unlike other cover types.
Interestingly, all nine species of butterflies registered as exclusive to the forest showed less than three individuals. According to the scale proposed by Fagua [87], species with fewer than three individuals are considered rare, and their occurrence is related to discontinuous flight periods or alternate imago emergences in different butterfly species. Master [88] proposed that the occurrence of rare species within the forest may indicate areas of interest for conservation, suggesting the importance of preserving the dry ecosystems of the Protective Forest Reserve Serranía de Coraza.
5. Conclusions
Seasonally dry forests are currently considered one of the ecosystems with the highest degree of threat, mainly due to anthropic actions such as livestock, cultivation, and urban expansion. These actions have been modifying the structure of biotic communities. The diversity has been modified, with a reduction in richness and possible changes in species composition. However, hypotheses on the behavior of their communities, such as the intermediate disturbance, had not been tested in this type of ecosystem in the Colombian Caribbean. In this sense, the results of the current study corroborate that the anthropic disturbance allowed the formation of areas with intermediate disturbance, which shows a higher alpha diversity, both taxonomic and phylogenetic.
In the same way, it is clear that these effects have shaped the formation of two new assemblages of butterfly species associated with intermediate disturbed and disturbed areas, within which there is a homogeneity process in beta diversity, a product of a progressive decrease in diversity. Likewise, these new communities also show a phylogenetic homogeneity process where most of their species exhibit a higher taxonomic relationship than expected by chance. This may be the product of ecological processes such as competition or the ecological filter. However, it is necessary to address the analysis of functional traits and, thus, establish new hypotheses about the structure formation of the associated communities with disturbed areas.
In the same way, these new communities show species that are indicative or exclusive to them since they are not found in other areas. This suggests that some forest taxa are not very sensitive to cover changes, but there are also species that are more malleable to environmental changes. According to the above, the transformation of forest to agricultural cover would imply local losses of species associated with tropical dry forests. This result is highly relevant, as butterflies can quickly generate new species assemblages according to environmental changes. It is also clear that species that only occur within a forest are lost when cutting down a forest.
Author Contributions
Conceptualization C.E.G-S. and J.D.M-G.; methodology, C.E.G-S. and J.D.M-G; formal analysis, C.E.G-S. and J.D.M-G.; investigation, Y.L.M-G., C.E.G-S., and J.D.M-G.; resources, Y.L.M-G. and J.D.M-G.; data curation, Y.L.M-G. and C.E.G-S.; writing—original draft preparation, J.D.M-G.; writing—review and editing, J.D.M-G. and C.E.G-S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
On behalf of Universidad de Sucre, ANLA approved the biological collection of butterfly specimens in the study area (permit number 0391 of 2016).
Data Availability Statement
The data are available from the corresponding author upon request.
Acknowledgments
We would like to thank Pedro Álvarez, Daniel Peralta, and Luis Andrés Severiche for their assistance in collecting the butterflies and tracing the transects in the field. Likewise, to Universidad de Sucre for lending its facilities for research development.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Linares-Palomino, R.; Oliveira-Filho, A.T.; Pennington, R.T. Neotropical seasonally dry forests: diversity, endemism, and biogeography of woody plants. In Seasonally dry tropical forest. Ecology and conservation, Dirzo, R., Young, H., Mooney, H., Ceballos, G., Eds.; Island Press/Center for Resource Economics: 2011; pp. 3–21.
- Banda, K.; Delgado-Salinas, A.; Dexter, K.G.; Linares-Palomino, R.; Oliveira-Filho, A.; Prado, D.; Pullan, M.; Quintana, C.; Riina, R.; Rodríguez, G.M.; et al. Plant diversity patterns in neotropical dry forests and their conservation implications. Science 2016, 353, 1383–1387. [Google Scholar] [CrossRef]
- Pizano, C.; Garcia, H. El Bosque Seco Tropical en Colombia; Instituto de Investigacion de Recursos Biologicos Alexander von Humboldt (IAvH): Bogota, 2014. [Google Scholar]
- García, H.; Corzo, G.; Isaac, P.; Etter, A. Distribución y estado actual de los remanentes del bioma de bosque seco tropical en Colombia: Insumos para su conservación. In El bosque seco tropical en Colombia, Pizano, C., García, H., Eds.; Instituto de Investigacion de Recursos Biológicos, Alexander von Humboldt: Bogota, D.C, 2014; Volume 90, pp. 228–251. [Google Scholar]
- Olascuaga, D.; Sánchez-Montaño, R.; Mercado-Gómez, J. Análisis de la vegetación sucesional en un fragmento de bosque seco tropical en Toluviejo-Sucre (Colombia). Colombia forestal 2016, 19, 23–40. [Google Scholar] [CrossRef]
- White, P.; Pickett, S. Natural disturbance and patch dynamics: an introduction. In The Ecology of Natural Disturbance and Patch Dynamics, Pickett, S.T.A., White, P.S., Eds.; Academic Press: San Diego, 1985; pp. 3–13. [Google Scholar]
- Séguin, A.; Gravel, D.; Archambault, P. Effect of disturbance regime on alpha and beta diversity of rock pools. Diversity 2014, 6, 1–17. [Google Scholar] [CrossRef]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How should beta-diversity inform biodiversity conservation? Trends in Ecology & Evolution 2016, 31, 67–80. [Google Scholar] [CrossRef]
- Myers, J.A.; Chase, J.M.; Crandall, R.M.; Jiménez, I. Disturbance alters beta-diversity but not the relative importance of community assembly mechanisms. Journal of Ecology 2015, 103, 1291–1299. [Google Scholar] [CrossRef]
- Dornelas, M. Disturbance and change in biodiversity. Philosophical transactions of the Royal Society of London. Series B, Biological sciences 2010, 365, 3719–3727. [Google Scholar] [CrossRef] [PubMed]
- Bendix, J.; Wiley, J.J.; Commons, M.G. Intermediate disturbance and patterns of species richness. Physical Geography 2017, 38, 393–403. [Google Scholar] [CrossRef]
- Santillan, E.; Seshan, H.; Constancias, F.; Drautz-Moses, D.I.; Wuertz, S. Frequency of disturbance alters diversity, function, and underlying assembly mechanisms of complex bacterial communities. Biofilms and Microbiomes 2019, 5, 8. [Google Scholar] [CrossRef]
- Svensson, J.R.; Lindegarth, M.; Jonsson, P.R.; Pavia, H. Disturbance-diversity models: what do they really predict and how are they tested? Proceedings of the Royal Society B: Biological Sciences 2012, 279, 2163–2170. [Google Scholar] [CrossRef]
- Raguso, R.A.; Llorente-Bousquets, J. The Butterflies (Lepidoptera) of the Tuxtlas Mts., Veracruz, México, Revisited: Species-Richness and Habitat Disturbance. The Journal of Research on the Lepidoptera 1990, 29. [Google Scholar] [CrossRef]
- Spitzer, K.; Jaros̆, J.; Havelka, J.; Leps̆, J. Effect of small-scale disturbance on butterfly communities of an Indochinese montane rainforest. Biological Conservation 1997, 80, 9–15. [Google Scholar] [CrossRef]
- Addo-Fordjour, P.; Osei, B.; Kpontsu, E. Butterfly community assemblages in relation to human disturbance in a tropical upland forest in Ghana, and implications for conservation. Journal of Insect Biodiversity 2015, 3, 1–18. [Google Scholar] [CrossRef]
- León-Cortés, J.L.; Caballero, U.; Miss-Barrera, I.D.; Girón-Intzin, M. Preserving butterfly diversity in an ever expanding urban landscape? A case study in the highlands of Chiapas, MÈxico. Journal of Insect Conservation 2019, 23, 405–418. [Google Scholar] [CrossRef]
- Vellend, M.; Verheyen, K.; Flinn, K.; Jacquemyn, H.; Kolb, A.; Van calster, H.; Peterken, H.; Graae, B.; Bellemare, J.; Honnay, O.; et al. Homogenization of forest plant communities and weakening of species–environment relationships via agricultural land use. Journal of Ecology 2007, 95, 565–573. [Google Scholar] [CrossRef]
- Ghazoul, J. Impact of logging on the richness and diversity of forest butterflies in a tropical dry forest in Thailand. Biodiversity & Conservation 2002, 11, 521–541. [Google Scholar] [CrossRef]
- Mercado-Gómez, J.; Prieto-Torres, D.A.; Gonzalez, M.; Morales-Puentes, M.; Escalante, T.; Rojas, O. Climatic affinities of neotropical species of Capparaceae: an approach from ecological niche modelling and numerical ecology. Botanical Journal Of The Linnean Society 2020, 193, 1–13. [Google Scholar] [CrossRef]
- Webb, C. Exploring the phylogenetic structure of ecological communities: an example for rain forest trees. The American Naturalist 2000, 156, 145–155. [Google Scholar] [CrossRef]
- Feng, G.; Svenning, J.-C.; Mi, X.; Jia, Q.; Rao, M.; Ren, H.; Bebber, D.P.; Ma, K. Anthropogenic disturbance shapes phylogenetic and functional tree community structure in a subtropical forest. Forest Ecology and Management 2014, 313, 188–198. [Google Scholar] [CrossRef]
- Santo-Silva, E.E.; Santos, B.A.; Arroyo-Rodríguez, V.; Melo, F.P.L.; Faria, D.; Cazetta, E.; Mariano-Neto, E.; Hernández-Ruedas, M.A.; Tabarelli, M. Phylogenetic dimension of tree communities reveals high conservation value of disturbed tropical rain forests. Diversity and Distributions 2018, 24, 776–790. [Google Scholar] [CrossRef]
- Checa, M.F.; Donoso, D.; Levy, E.; Mena, S.; Rodriguez, J.; Willmott, K. Assembly mechanisms of neotropical butterfly communities along an environmental gradient. bioRxiv 2019, 632067. [Google Scholar] [CrossRef]
- Pellissier, L.; Alvarez, N.; Espíndola, A.; Pottier, J.; Dubuis, A.; Pradervand, J.-N.; Guisan, A. Phylogenetic alpha and beta diversities of butterfly communities correlate with climate in the western Swiss Alps. Ecography 2013, 36, 541–550. [Google Scholar] [CrossRef]
- Mercado-Gómez, J.; Solano, C.; Hoffman, W. Recursos florales usados por dos especies de Bombus en un fragmento de bosque subandino (Pamplonita-Colombia). Recia 2017, 9, 31–37. [Google Scholar]
- Sampedro, M.A.; Gómez, F.H.; Ballut, D.G. Estado de la vegetación en localidades abandonadas por “desplazamiento”, en los montes de María Sucre, Colombia Recia 2014, 6, 184–193. 6.
- Aguilera, M. La Mojana: riqueza natural y potencial económico; Banco de la República. Serie de documentos de trabajo sobre economía regional Nº 48. Cartagena, Colombia: 2005; p. 122.
- Herazo-Vitola, F.; Mendoza-Cifuentes, H.; Mercado-Gómez, J. Estructura y composición florística del bosque seco tropical en los Montes de María (Sucre – Colombia). Ciencia en desarrollo 2017, 8, 79–90. [Google Scholar] [CrossRef]
- Elias, M.; Gompert, Z.; Jiggins, C.; Willmott, K. Mutualistic Interactions Drive Ecological Niche Convergence in a Diverse Butterfly Community. PLoS Biol 2008, 6, 300–308. [Google Scholar] [CrossRef]
- Freitas, L.; Iserhard, C.; Santos, J.; Junia, O.; Danilo, R.; Douglas, A.; Augusto, H.; Onildo, a.-F.; Gustavo, A.; Mauricio, U.-P. Studies with butterfly bait traps: an overview. Revista Colombiana de Entomología 2014, 40, 203–212. [Google Scholar]
- Fraija, N.; Medina, G. Caracterización de la fauna del orden lepidóptera (rhopalocera) en cinco diferentes localidades de los llanos orientales colombianos. Acta Biológica Colombiana 2006, 11, 55–68. [Google Scholar]
- Villanueva-Espinoza, R.; Condo, F. Sinopsis de la familia Acanthaceae en el Perú. Revista Forestal del Perú 2019, 34, 21–40. [Google Scholar] [CrossRef]
- Borror, D.; Delong, D.; Triplehorn, C.; Johnson, N. An Introduction of the study of insects; Harcourt Brace Jovanovich College: Philadelphia, 1982. [Google Scholar]
- Borror, D.; Triplehorn, C.; Johnson, N. An Introduction to the Study of Insects; Hartcourt Brace Jovanovich College: Philadelphia, 1982. [Google Scholar]
- Triplehorn, C.; Johnson, N. Borror and DeLong's lntroduction to the study of insects, Seventh Edition ed.; Thomson Brooks Cole, USA, 2005.
- Warren, A.D.; Davis, K.J.; Stangeland, E.M.; Pelham, J., P; Willmott, K.R.; Grishin, N.V. Butterflies of America. Available online: http://www.butterfliesofamerica.com/ (accessed on Illustrated Lists of American Butterflies).
- Le Crom, J.F.; Constantino, L.M.; Salazar, J.A. Mariposas de Colombia Tomo 1 Papilionoidae; CARLEC Ltda: Bogota, 2002. [Google Scholar]
- Le Crom, J.F.; Constantino, L.M.; Salazar, J.A. Mariposas de Colombia Tomo 2: Pieridae; CARLEC Ltda: Bogota, Colombia, 2004. [Google Scholar]
- Lamas, G. Atlas of Neotropical Lepidoptera. In CheckList: Part 4A Hesperioidea-Papilionoidea, J, H., Ed.; Association for Tropical Lepidoptera Scientific, 2004; p. 479.
- Andrade-C, M.; Henao-Bañol, E.; Triviño, P. Técnicas y Procesamiento para la Recoleccion, Preservacion y Montaje de Mariposas en estudios de Biodiversidad y Conservcion (Lepidoptera:Hesperioidea-Papilionoidea) Rev. Acad. Colomb. Cienc 2013, 37, 311–325. [Google Scholar]
- Chao, A.; Gotelli, N.J.; C, H.T.; Elizabeth, S.; MA, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecological Monographs 2014, 84, 45–57. [Google Scholar] [CrossRef]
- Hsieh, T.; Ma, K.; Chao, A. iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods in Ecology and Evolution 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Colwell, R.; Chao, A.; Gotelli, N.; Shang-Yi, L.; Chang Xuan, M.; Robin L, C.; Jhon T, L. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. Journal of Plant Ecology 2012, 5, 3–21. [Google Scholar] [CrossRef]
- Gonzalez-Hidalgo, J. Las divisorias de aguas como elementos del paisaje. Geographycalia 1990, 27, 47–62. [Google Scholar] [CrossRef]
- Akaike, H. Akaike, H. A new look at the statistical model identification. In Selected Papers of Hirotugu Akaike, Parzen, E., Tanabe, K., Kitagawa, G., Eds.; Springer New York: New York, 1974; pp. 215–222. [Google Scholar]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Global Ecology and Biogeography 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L. betapart: an R package for the study of beta diversity. Methods in Ecology and Evolution 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity into independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef] [PubMed]
- Marcon, E.; Hérault, B. entropart: An R Package to Measure and Partition Diversity. 2015 2015, 67, 26. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol Electron 2001, 4, 9. [Google Scholar]
- Moreno, C.; Castillo-Campos, G.; Verdú, J. Taxonomic diversity as complementary information to assess plant species diversity in secondary vegetation and primary tropical deciduous forest. Journal of Vegetation Science 2009, 20, 935–943. [Google Scholar] [CrossRef]
- Warwick, R.; Clarke, R. New ’biodiversity’ measures reveal a decrease in taxonomic distinctness with increasing stress. Marine Ecology Progress Series 1995, 129, 301–305. [Google Scholar] [CrossRef]
- Tucker, C.M.; Cadotte, M.W.; Carvalho, S.B.; Davies, T.J.; Ferrier, S.; Fritz, S.A.; Grenyer, R.; Helmus, M.R.; Jin, L.S.; Mooers, A.O.; et al. A guide to phylogenetic metrics for conservation, community ecology and macroecology. Biological Reviews 2017, 92, 698–715. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. A further biodiversity index applicable to species lists: variation in taxonomic distinctness. Marine Ecology Progress Series 2001, 216, 265–278. [Google Scholar] [CrossRef]
- Moreno, C.; Castillo-Campos, G.; Verdú, J. Taxonomic diversity as complementary information to assess plant species diversity in secondary vegetation and primary tropical deciduous forest. Journal of vegetation Science 2009, 20, 935–943. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecological Monographs 1997, 67, 345–366. [Google Scholar] [CrossRef]
- González-Valdivia, N.; Ochoa-Gaona, S.; Pozo, C.; Gordon Ferguson, B.; Rangel-Ruiz, L.J.; Arriaga-Weiss, S.L.; Ponce-Mendoza, A.; Kampichler, C. Indicadores ecológicos de hábitat y biodiversidad en un paisaje neotropical: perspectiva multitaxonómica. Revista de Biología Tropical 2011, 59, 1433–1451. [Google Scholar] [CrossRef] [PubMed]
- Henao-Bañol, E.R.; Gantiva-Q. , C.H. Mariposas (Lepidoptera: Hesperioidea-Papilionoidae) del bosque seco tropical (BST) en Colombia. Conociendo la diversidad en un ecosistema amenazado. Boletín Científico Centro de Museos Museo de Historia Natural 2020, 24, 150–196. [Google Scholar] [CrossRef]
- Millan, C.; Chacon, P.; Giraldo, A. Estudio de la comunidad de Lepidopteros diurnos en zonas naturales y sistemas productivos del municipio de Caloto ( Cauca, Colombia). Bol.cient.mus.hist.nat. 2009, 13, 185–195. [Google Scholar]
- Gaviria-Ortiz, F.; Henao-Bañol, E. Diversidad de mariposas diurnas (Hesperioidea-Papilionoidea) del parque natural regional el vinculo (Buga-Valle del Cauca). Boletín Científico centro de Museos Museo de Historia Natural 2011, 15, 115–133. [Google Scholar]
- Henao-Bañol, E.R.; Andrade-C, M. Registro del género Megaleas (Lepidoptera: Hesperiidae: Hesperiinae) para Colombia con descripción de una nueva especie. Revista Academia Colombiana de Ciencias Exactas 2013, 37, 29–35. [Google Scholar] [CrossRef]
- Orozco, S.; Muriel, S.; Palacio, J. Diversidad de Lepidopteros diurnos en una área de Bosque Seco Tropical del Occidente antioqueño Actualidades biologicas 2009, 90, 31–41. 90.
- Peña, J.; Reinoso, G. Mariposas diurnas de tres fragmentos de bosque seco tropical del alto valle del Magdalena. Tolima-Colombia. Revista de la Asociación Colombiana de Ciencias Biológicas 2016, 1, 57–66. [Google Scholar]
- Prince-Chacon, S.; Vargas-Zapata, M.; Salazar-E, J.; Martinez-Hernandez, N. Mariposas Papilionoidea y Hesperioidea (Insecta:Lepidoptera) en dos fragmentos de Bosque Seco Tropical en Corrales de San Luis, Atlantico, Colombia. Boletin de la Sociedad Entomologica Aragonesa 2011, 48, 243–252. [Google Scholar]
- Casas-Pinilla, L.; Mahecha, J.; Dumar, R.; Rios-Malaver, I. Diversidad de mariposas en un paisaje de bosque seco tropical, en la Mesa de los Santos, Santander, Colombia (Lepidoptera: Papilionoidea). SHILAP Rev Lepid 2017, 45, 83–108. [Google Scholar]
- Vargas-Zapata, M.; Boom-Urueta, C.; Seña-Ramos, L.; Echeverry-Iglesias, A.; Martinez - Hernandez, N. Composicion vegetal, preferencias alimenticias y abundancia de Biblidinae (Lepidoptera:Nymphalidae) en un fragmento de bosque seco tropical en el departamento del Atlantico, Colombia. Acta Biol Colomb 2015, 20, 79–92. [Google Scholar] [CrossRef]
- Carlos, P.; Dahners, H.W. Eumaeini (Lepidoptera: Lycaenidae) del cerro San Antonio: Dinámica de la riqueza y comportamiento de Hilltopping Rev Colomb Entomol 2006, 32, 179–190. 32.
- Montero, F.; Moreno, M.; Gutiérrez, L. Mariposas (Lepidoptera: hesperioidea y papilionoidea) asociadas a fragmentos de bosque seco tropical en el departamento del Atlántico, Colombia. Boletín Científico Centro de Museos Museo de Historia Natural 2009, 13, 157–173. [Google Scholar]
- Vargas Zapata, M.A.; Martinez Hernandez, N.J.; Gutierrez Moreno, L.C.; Prince Chacon, S.; Herrera Colon, V.; Torres Periñan, L.F. Riqueza y Abundancia de Hesperioidea y Papilionoidea(Lepidoptera) en la Reserva Natural las Delicias, Santa Marta Magdalena, Colombia. Acta Biol. Colomb 2011, 16. [Google Scholar]
- Vargas-Zapata, M.; Boom-Urueta, C.; Seña-Ramos, L.; Echeverry-Iglesias, A.; Martinez Hernandez, N. Composicion vegetal, preferencias alimenticias y abundancia de Biblidinae (Lepidoptera:Nymphalidae) en un fragmento de Bosque Seco Tropical en el departamento del Atlantico, Colombia. Acta Biologica Colombiana 2015, 20, 79–92. [Google Scholar] [CrossRef]
- Diaz, J.M. Bosque Seco Tropical en Colombia; Banco de Occidente: Cali, 2006. [Google Scholar]
- Connell, J. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef]
- Beck, J.; Schulze, C.H.; author_in_Japanese. Diversity of fruit-feeding butterflies (Nymphalidae) along a gradient of tropical rainforest succession in Borneo with some remarks on the problem of "pseudoreplicates". Lepidoptera Science 2000, 51, 89–98. [Google Scholar] [CrossRef]
- Vanschoenwinkel, B.; Buschke, F.; Brendonck, L. Disturbance regime alters the impact of dispersal on alpha and beta diversity in a natural metacommunity. Ecology 2013, 94, 2547–2557. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Tucker, C.M. Should environmental filtering be abandoned? Trends in Ecology & Evolution 2017, 32, 429–437. [Google Scholar] [CrossRef]
- Heino, J.; Melo, A.S.; Bini, L.M. Reconceptualising the beta diversity-environmental heterogeneity relationship in running water systems. Freshwater Biology 2015, 60, 223–235. [Google Scholar] [CrossRef]
- Pavoine, S.; Baguette, M.; Stevens, V.M.; Leibold, M.A.; Turlure, C.; Bonsall, M.B. Life history traits, but not phylogeny, drive compositional patterns in a butterfly metacommunity. Ecology 2014, 95, 3304–3313. [Google Scholar] [CrossRef]
- Pérez Hernández, C. Distintividad taxonómica: Evaluación de la diversidad en la estructura taxonómica en los ensambles. In La biodiversidad en un mundo cambiante: Fundamentos teóricos y metodológicos para su estudio, Moreno, C., Ed.; Universidad Autónoma del Estado de Hidalgo/Libermex: Ciudad de México, 2019; pp. 285–306. [Google Scholar]
- Angulo, D.F.; Ruiz-Sanchez, E.; Sosa, V. Niche conservatism in the Mesoamerican seasonal tropical dry forest orchid Barkeria (Orchidaceae). Evolutionary Ecology 2012, 26 991–1010.
- Swenson, N.G.; Enquist, B. Opposing assembly mechanisms in a Neotropical dry forest: implications for phylogenetic and functional community ecology. Ecology 2009, 90, 2161–2170. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community assembly, coexistence and the environmental filtering metaphor. Functional Ecology 2015, 29, 592–599. [Google Scholar] [CrossRef]
- Wiens, J.J.; Graham, C.H. Niche conservatism: integrating evolution, ecology, and conservation biology. Annual review ecology and systematics 2005, 36, 519–539. [Google Scholar] [CrossRef]
- Webb, C.; Ackerly, D.; McPeek, M.; Donoghue, M. Phylogenies and community ecology. Annual Review of Ecology and Systematics 2002, 33, 475–505. [Google Scholar] [CrossRef]
- Robinson, W.H. Insects and mite pests in the human environment. In Urban entomology; Chapman & Hall: Gran Bretaña, 1996; p. 430. [Google Scholar]
- Ramirez -Restrepo, L.; Chacon, P.; Constantino, L. Diversidad de Mariposas Diurnas ( lepidoptera:Papilionoidea y Hesperoidea) en Santiago de Cali, Valle del Cauca, Colombia. Revista Colombiana de Entomologia 2007, 33, 54–63. [Google Scholar] [CrossRef]
- Fagua, G. Comunidad de mariposas y artropodofauna asociada con el suelo de tres tipos de vegetación de la Serranía de Taraira (Vaupés, Colombia). Una prueba del uso de mariposas como bioindicadores. Revista Colombiana de Entomología. 1996, 22, 143–151. [Google Scholar] [CrossRef]
- Master, L.L. Assesing treats and setting priorices for conservation. Conservation Biology 1991, 5, 559–563. [Google Scholar] [CrossRef]
Figure 1.
Study area. (A) Colombia, South America; (B) Department of Sucre; location of Reserva Forestal Protectora Serranía de Coraza in red; (C) Linear transects and sampling plots.
Figure 1.
Study area. (A) Colombia, South America; (B) Department of Sucre; location of Reserva Forestal Protectora Serranía de Coraza in red; (C) Linear transects and sampling plots.
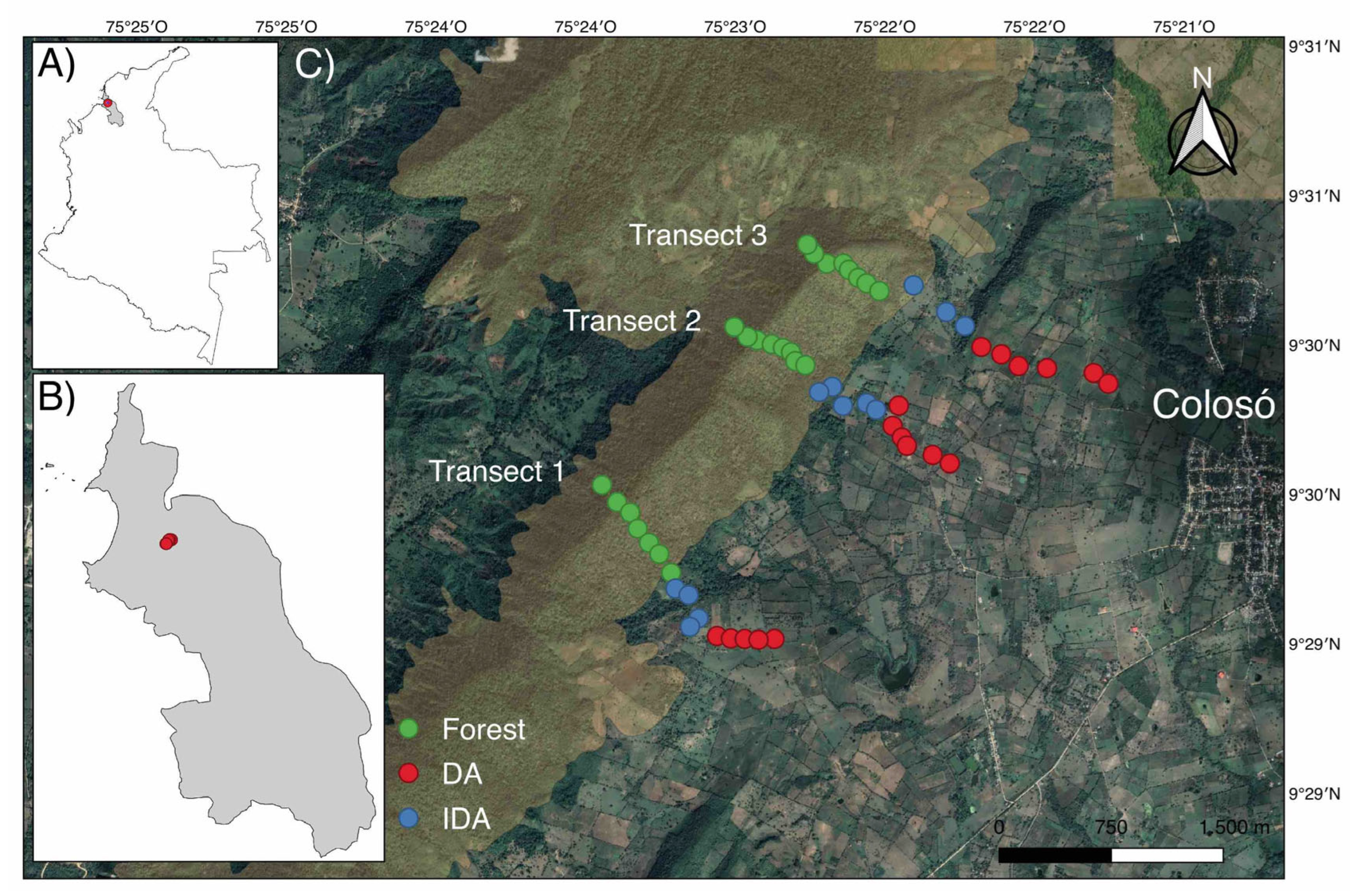
Figure 2.
Relationship between the distance to the watersheds and the diversity of diurnal butterflies in the Serranía de Coraza, described by a Gaussian non-linear regression model, in red line; 95% confidence intervals, in blue lines. (A) richness; (B) abundance.
Figure 2.
Relationship between the distance to the watersheds and the diversity of diurnal butterflies in the Serranía de Coraza, described by a Gaussian non-linear regression model, in red line; 95% confidence intervals, in blue lines. (A) richness; (B) abundance.
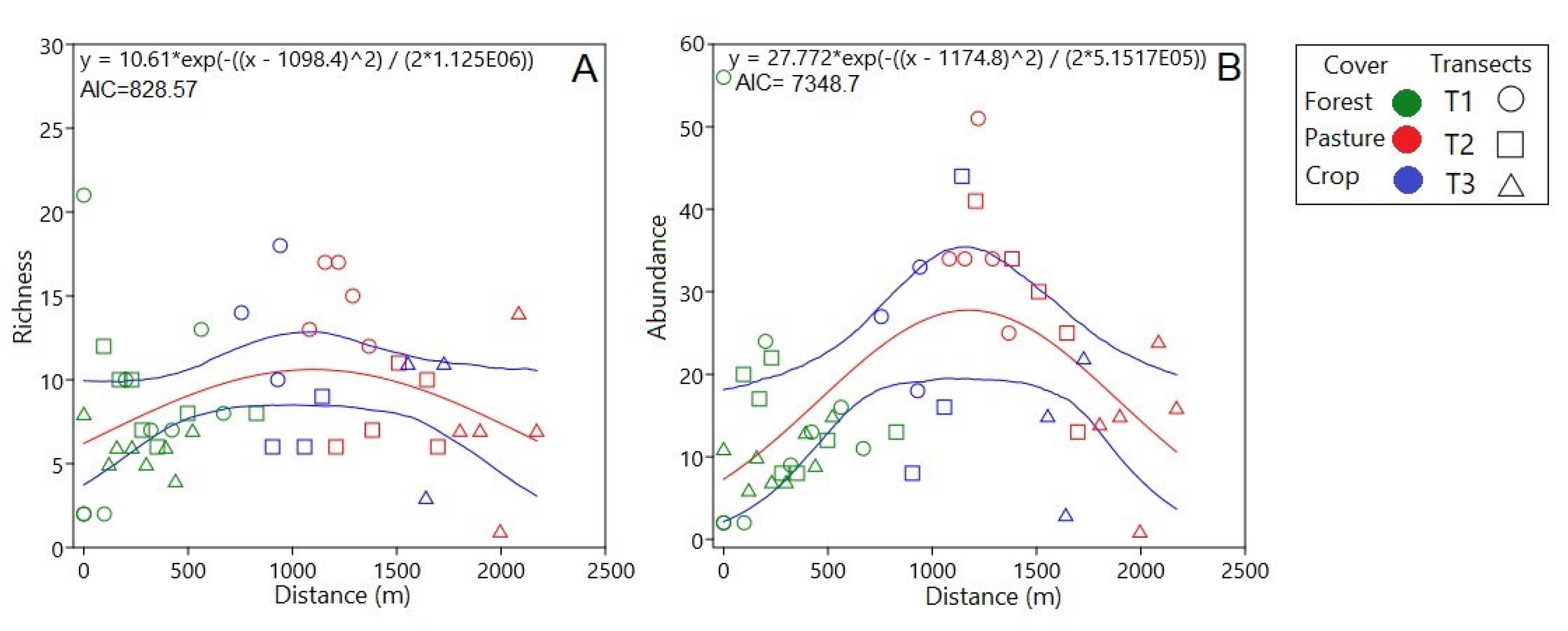
Figure 3.
Rarefaction and interpolation-extrapolation curves, based on the diurnal butterfly community by cover type (forest or disturbed); solid lines represent the estimation by interpolation, and dashed lines represent extrapolation. (A) Sampling completeness; (B) Richness estimation.
Figure 3.
Rarefaction and interpolation-extrapolation curves, based on the diurnal butterfly community by cover type (forest or disturbed); solid lines represent the estimation by interpolation, and dashed lines represent extrapolation. (A) Sampling completeness; (B) Richness estimation.
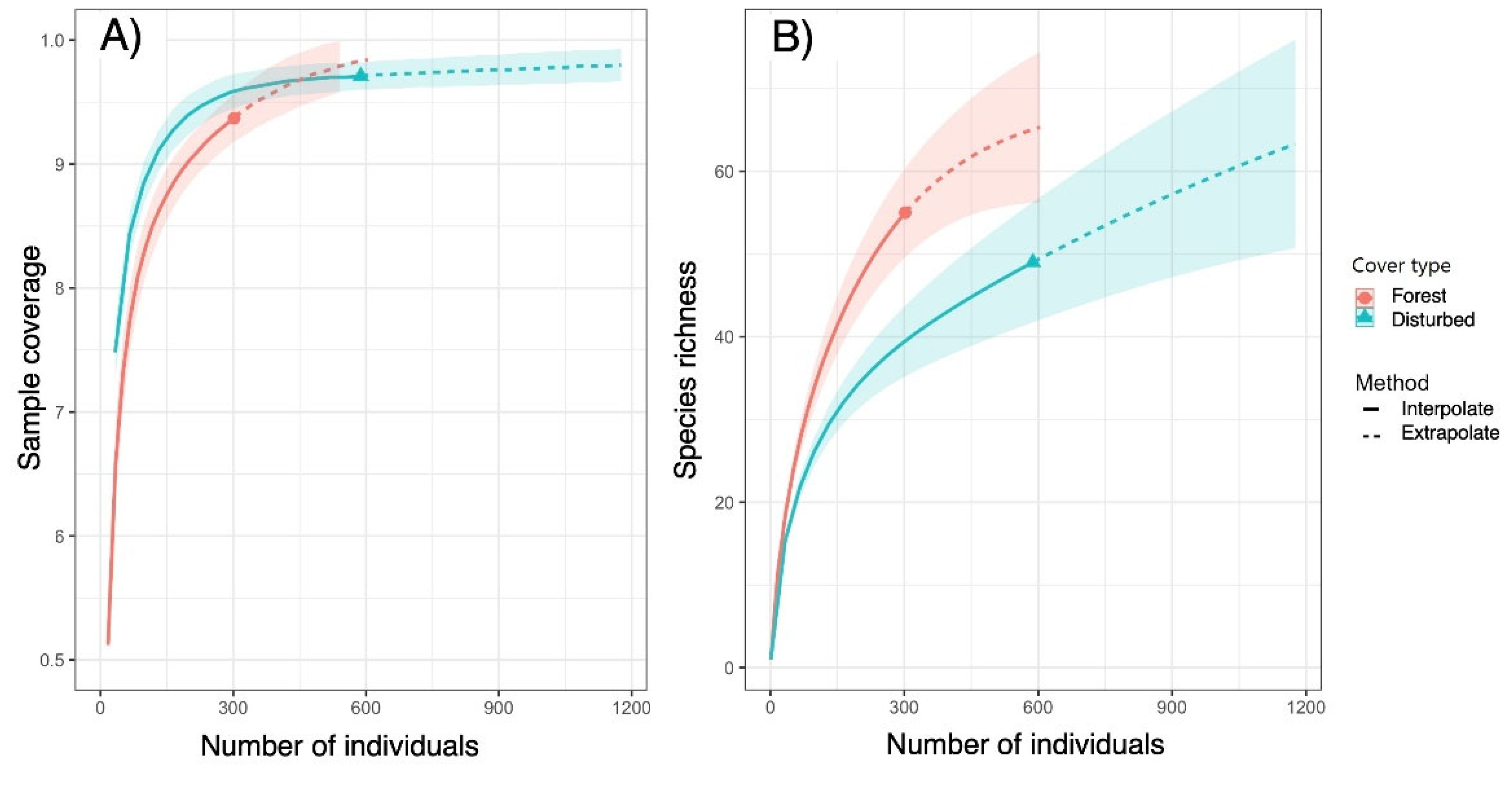
Figure 4.
Cluster analysis, based on Bray-Curtis distance, of the sampling plots grouped by the distance to the watersheds.
Figure 4.
Cluster analysis, based on Bray-Curtis distance, of the sampling plots grouped by the distance to the watersheds.
Figure 5.
Indicator species analysis (IndVal%) for undisturbed areas or forests, areas with intermediate disturbance, and disturbed areas. Species with values above 70% (reddish tones) are considered as indicators; gray boxes denote species with significances (p value< 0.05) of the indicator values.
Figure 5.
Indicator species analysis (IndVal%) for undisturbed areas or forests, areas with intermediate disturbance, and disturbed areas. Species with values above 70% (reddish tones) are considered as indicators; gray boxes denote species with significances (p value< 0.05) of the indicator values.
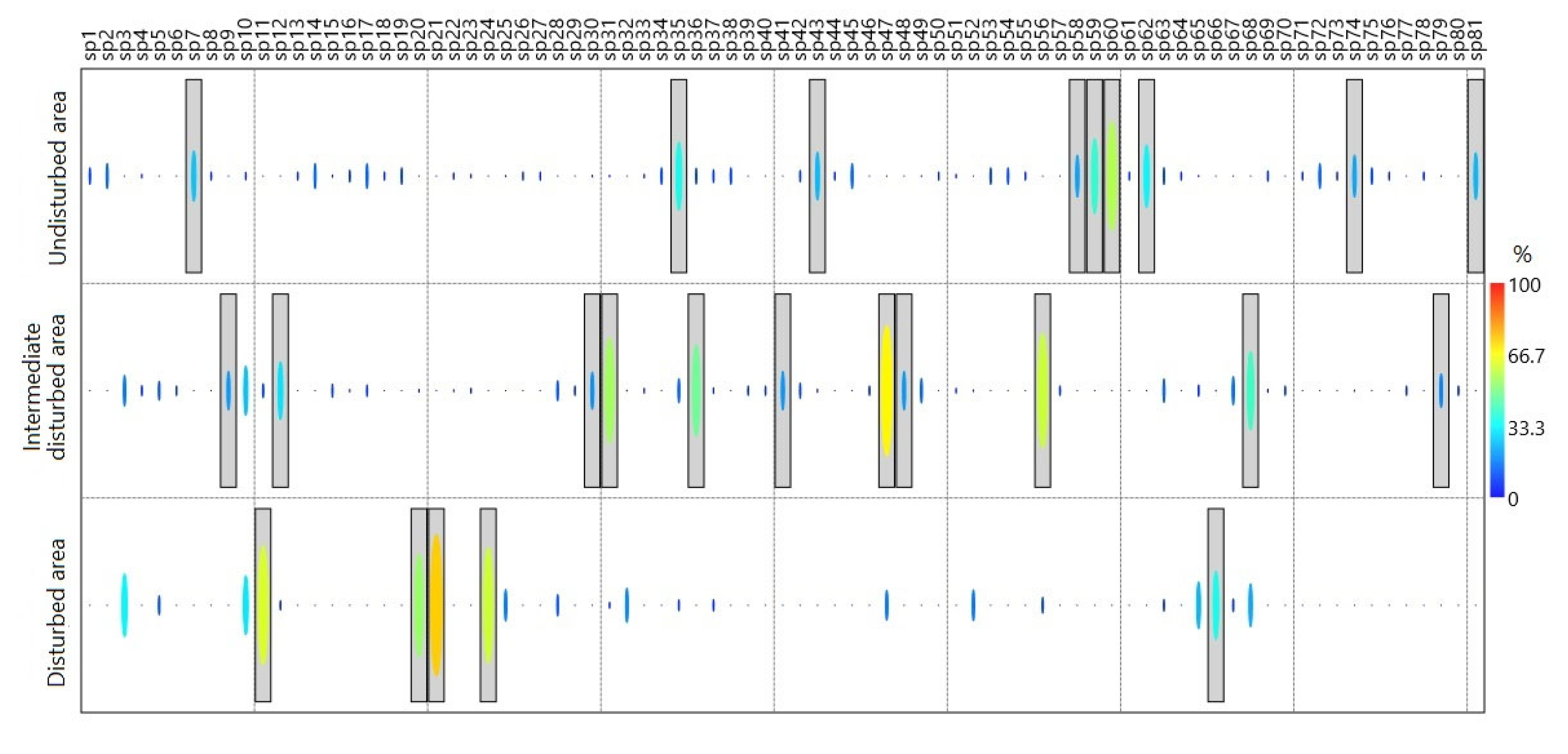

Table 1.
List of diurnal butterfly species identified in the Coraza reserve in three types of cover, forest, pasture and crops. (1) Exclusive to the forest, (2) Occurring both in the forest and in pasture-crop, and (3) Exclusive to pasture-crop
Table 1.
List of diurnal butterfly species identified in the Coraza reserve in three types of cover, forest, pasture and crops. (1) Exclusive to the forest, (2) Occurring both in the forest and in pasture-crop, and (3) Exclusive to pasture-crop
| Adelpha fessonia ernestoi 1 | Euptoieta hegesia3 | Mesosemia carissima 1 |
| Adelpha iphicleola 1 | Eurema agave3 | Mestra hersilia3 |
| Agraulis vanillae3 | Eurema arbela 2 | Microtia elva3 |
| Anartia amathea3 | Eurema daira 2 | Morpho helenor 1 |
| Anartia jatrophae3 | Eurema elathea3 | Morpho helenor peleides 1 |
| Anteos maerula 3 | Fountainea halice 2 | Myscelia leucocyana 1 |
| Archaeoprepona demophon 1 | Glutophrissa drusilla 1 | Neographium anaxilaus 1 |
| Archaeoprepona demophoon 1 | Hamadryas februa ferentina 2 | Nica flavilla 1 |
| Aricoris erostratus3 | Hamadryas feronia 2 | Parides anchises serapis 2 |
| Ascia monuste 2 | Heliconius erato 2 | Parides eurimedes mycale 1 |
| Battus polydamas 2 | Heliconius ethilla 1 | Parides iphidamas 2 |
| Biblis hyperia3 | Heraclides thoasnealces3 | Phoebis agarithe3 |
| Caligo brasiliensis morpheus 1 | Hermeuptychia Hermes 1 | Phoebis argante 2 |
| Callicore pitheas 1 | Historis odius3 | Phoebis sennae3 |
| Chlosyne lacinia 2 | Hypna clytemnestra 2 | Prepona laertes 2 |
| Chlosyne poecile 2 | Itaballia demophile calydonia 2 | Pseudolycaena marsyas3 |
| Cissia themis 2 | Itaballia pandosia 1 | Pyrisitia dina 1 |
| Colobura dirce 1 | Janatella leucodesma 1 | Pyrisitia leuce 1 |
| Consul fabius 1 | Juditha sp.3 | Pyrrhogyra neaerea 1 |
| Danaus eresimus3 | Junonia evarete3 | Siderone galanthis 2 |
| Danaus gilippus 2 | Junonia genoveva3 | Siproeta stelenes 1 |
| Detritivora hermodora 2 | Leptotes cassius3 | Smyrna blomfildia 1 |
| Doxocopa pavon theodora 2 | Libytheana carinenta 1 | Strymon sp.3 |
| Dryadula phaetusa3 | Lycorea halia 2 | Taygetis laches 1 |
| Dryas iulia 2 | Marpesia chiron3 | Temenis laothoe 2 |
| Ectima erycinoides 1 | Melanis electron 1 | Thereus cithonius3 |
| Eunica tatila 1 | Memphis arginussa 1 | Zaretis ellos 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



