
Submitted:
15 May 2023
Posted:
17 May 2023
You are already at the latest version
Abstract
Improving antigen presentation is crucial for the success of immunization strategies. Yeasts are classically used as biofactories to produce recombinant proteins and are efficient vehicles for the delivery of vaccine antigens, besides present adjuvants properties. Despite the absence of epidemic outbreaks, several vaccine approaches continue to be developed for Zika virus infection. These prophylactic strategies are fundamental given the severity of clinical manifestations, mainly due to viral neurotropism. The present study aimed to evaluate in vivo the immune response induced by P. pastoris recombinant strains displaying epitopes of the Envelope (ENV) and NS1 ZIKV proteins. Intramuscular immunization with heat-attenuated yeast enhanced the secretion of IL-6, TNF-α, and IFN-γ, besides activation of CD4+ and CD8+ T cells, in BALB/c mice. P. pastoris displaying ENV epitopes induced a more robust immune response, increasing immunoglobulin production, especially IgG isotypes. Both proposed vaccines showed the potential to induce immune responses without adverse effects, confirming the safety of administering P. pastoris as a vaccine vehicle. Here we demonstrated, for the first time, the evaluation of a vaccine against ZIKV based on a multiepitope construct, using yeast as a vehicle, reinforcing the applicability of P. pastoris as a whole yeast cell vaccine.
Keywords:
Vaccine
; Antigen-delivery
; ZIKV
; Yeast
1. Introduction
Biotechnological-relevant yeast species such as Saccharomyces cerevisiae, Pichia pastoris, Hansenula polymorpha, and Kluyveromyces lactis are conventionally employed in the synthesis of immunobiological products [1]. These species have GRAS (Generally Recognized as Safe) status, guaranteeing safety in their application as a biofactory and as vehicles for vaccine antigens [2,3,4]. The most attractive aspects of this vaccine delivery are the ability to induce specific immune responses against the recombinant antigen and the adjuvant properties of the yeasts [5]. Whole yeast vaccines can induce higher antigen-specific responses than those promoted by inactivated virus vaccines or proteins conjugated to traditional adjuvants, such as aluminum salts [6,7,8].
Yeast-based vaccines have been tested against various infectious agents, including viruses and fungi [9,10,11]. Overall, recombinant antigens are proteins from the target pathogen or epitopes derived from these proteins that are critical for inducing the appropriate immune response [12,13]. Although immunostimulation associated with the administration of recombinant yeasts can occur regardless of the cellular location of the heterologous protein, the exposure of recombinant antigens on the yeast surface can increase the efficiency of this process [7]. In the systems of yeast surface display, the target protein is covalently linked to an anchor protein with a glycosylphosphatidylinositol (GPI) motif and displayed on the cell surface. Anchor proteins belong to the mannoprotein class and include agglutinins (Agα1p, Aga1p, and Aga2p), Flo1p, Sed1p, Cwp1p, Cwp2p, Tip1p, and Tir1p/Srp1p [14,15,16]. The α-agglutinin is the anchor most commonly used and can allow the disposition of 105 - 106 target proteins per cell [17].
Due to the dissemination potential, viral neurotropism, and capacity to induce neurological disorders in fetuses and adults, preventing the Zika virus (ZIKV) infection is considered an issue for global public health [18]. The information about seroprevalence and duration of immunity against ZIKV is still limited, and the re-emergence of outbreaks continues to be considered [19]. In fact, an increase in the number of infections has been reported in Southeast Asian countries in the last four years [20]. Prophylactic vaccination is one of the main prevention measures, but there are no licensed vaccines for ZIKV infection [21]. Fully protection against flaviviruses involves a combination of adaptive humoral and cellular responses [22]. Since many immunodominant epitopes for the induction of T cell-mediated responses are present in non-structural proteins, epitopes or domains of proteins such as NS1 and NS3 have been included in the design of vaccines targeting both ZIKV and DENV [23]. In this context, vaccines based on multiepitope sequences are promising platforms concerning immunogenicity, protection, and safety [24].
In a previous study, we evaluated, in vitro, the potential of P.pastoris as a vaccine platform, surface displaying two multi-epitope synthetic antigens derived from Envelope (ENV) and NS1 ZIKV proteins [25]. The main highlights of this study were the stimulus to CD4+ and CD8+ cell expansion and the secretion of cytokines such as IL-6, IL-10, and TNF-α. Hence, here we decide to investigate the vaccine responses in vivo through yeast administration in immunocompetent BALB/c mice, evaluating the viability and immunogenicity of this yeast-based vaccine.
2. Materials and Methods
Vaccine antigens and yeast strains
In this study, two strains named P. pastoris:ENV and P. pastoris:ENVNS1 were generated in a previous study performed by our group [25]. The components of the expression cassettes used to transform the yeasts were: the MF-α peptide for signaling protein secretion, a His-tag that enable immunodetection; the vaccine antigen, and the α-agglutinin anchor. Two vaccine antigen versions were designed: one with B and T cell epitopes from the ENV protein and the other with epitopes from ENV and the NS1 proteins. The selected epitopes and a schematic representation of the expression cassette and the recombinant yeast are illustrated in Figure 1A–B.
To obtain the recombinant strains, the synthetic genes encoding the multi-epitope constructs were cloned into the expression vector pPGKΔ3_Agα (non-commercial vector; de Almeida et al. 2005)[26] that allows constitutive expression and anchorage of proteins on the yeast surface (Figure 1B). The P. pastoris strain transformed with the expression cassettes was GS115 (his4; Invitrogen). The expression and antigen anchorage were confirmed through RT-PCR and immunofluoresce microscopy, respectively [25]. In addition to these previous analyses, the immunoreactivity and anchoring of the yeasts were evaluated by Yeast-ELISA. For this purpose, 100 µL of 107 cells were applied in 96-well plates for 2 hours. Then, the subsequent steps were blocking (5% BSA in PBS), labeling with Anti-HIS primary antibody (Sigma-Aldrich) 1:1000 diluted (1 hour), and with secondary Anti-IgG antibody conjugated to peroxidase (Sigma-Aldrich) 1:5000 diluted (45 minutes). Among the incubations, the samples were triple-washed with PBS-Tween (1%). The revelation was performed with TMB (3,3',5,5'-tetramethylbenzidine; Life Technologies) in the dark, and the reaction was interrupted with HCl (1N). The signal was detected on a plate reader with a wavelength set to 450 nm.
Yeast preparation
After 72h of cultivation in YPD medium, at 28°C under agitation (150 rpm), yeasts were collected after centrifugation (4500 rpm; 10 minutes) and washed twice with 1x PBS. Cells were resuspended in sterile 1x PBS to adjust the final concentration to OD600 10 in 50 µl. The yeasts were heat inactivated by incubation at 60°C for 1 hour and stored at 4°C until the moment of use.
Mice, ethical parameters, and immunization protocol
Female immunocompetent BALB/c mice, 6 – 8 weeks old, were raised and maintained in the bioterium of the Aggeu Magalhães Institute (Oswaldo Cruz Foundation – PE, Brazil) under sterile, pathogen-free conditions. All experiments involving mice followed strictly the standards established by the institutional Ethics Committee for the Use of Animals (protocol n. 110/2017). The immunization schedule was performed in two doses, one week apart, with intramuscular injection (Figure 1C). The mice were divided into three groups (n=5) defined as: G1 - mice inoculated with non-recombinant P. pastoris (nr); G2 - mice vaccinated with P. pastoris:ENV; and G3 - vaccinated with P. pastoris:ENVNS1. Before each immunization, all mice were anesthetized with xylazine hydrochloride (10 mg.Kg-1) and ketamine (115 mg.Kg-1). Each experimental group received doses of 50 µl with yeast cells in an OD60010. Twenty-one days after the first dose, all animals were anesthetized for blood collection and subsequent euthanasia. Spleens were removed for isolation and culture of splenic lymphocytes.
In vitro culture and stimulation of isolated spleen lymphocytes
Splenocytes from vaccinated animals were isolated and the mononuclear immune cells were isolated by separation with Ficoll-Paque PLUS 1.077 g.mL-1 (GE Healthcare Life Sciences) and distributed in 48-well plates, 106 cells/well. The isolated cells were restimulated in vitro with the yeasts P. pastoris:nr (for G1), P. pastoris expressing ENV epitopes (for G2), and ENVNS1 epitopes (for G3), at a concentration of 105 cells/well. The cells were incubated in RPMI medium (Sigma-Aldrich) containing 10% FBS, at 37ºC (5% CO2), in the experimental times of 24h, 48h, and 72h.
Immunological analysis
Lymphocytes isolated from blood and spleen were characterized for the presence of surface markers CD4, CD8 and CD16, by labeling the cells with the corresponding antibodies (anti-CD4-FITC, anti-CD8-PE, and anti-CD16/32-FITC; BDTM Bioscience). Serum and culture supernatant were analyzed for cytokine dosage of TNF-α, IFN-γ, IL-2, IL-4, IL-6, IL-10, and IL-17A using the BD CBA Mouse Th1/ Th2/Th17 Kit (BDTM Bioscience), following the manufacturer's instructions. IgG, IgM, IgA, and IgE immunoglobulins were measured in the serum of vaccinated mice using the Mouse Immunoglobulin Isotyping Kit (BDTM Bioscience). All acquisitions for immunological assays were performed by flow cytometry (BD ACCURI C6).
Hematological and biochemical analyzes
Blood samples were collected through a cardiac puncture, placed into EDTA-K2 tubes, and centrifuged at 3600 rpm for 10 minutes to separate serum and plasma. The hematological evaluation included global counts of red blood cells, leukocytes, platelets, determination of hematocrit, and hemoglobin concentration. The values of red blood cells, hematocrit, and hemoglobin allowed the calculation of the mean corpuscular volume and the mean corpuscular hemoglobin concentration. The blood cell count was performed in a Neubauer chamber, with differential counting of the slides stained by the Rapid Panoptic method [27]. The number of platelets was determined using the Fonio method [28]. Hemoglobin was measured by colorimetry using a spectrophotometer. The biochemical analyses were performed with Labtest Diagnosis kits (Brazil). The levels of glucose, urea, and creatinine were measured through end-point colorimetric enzymatic assays. Alkaline phosphatase was detected by the modified Roy's method, and the liver transaminases by the Reitman - Frankel method.
Statistical analysis
Graphs and statistical analysis were generated by GraphPad Prism version 7.04. The analysis of variance (ANOVA) was the applied to assess statistical differences between groups. Results with a p-value < 0.05 were considered statistically significant.
3. Results
3.1. P. pastoris can surface display vaccine antigens
The recombinant yeasts were generated and characterized in a previous study [25]. A Yeast-ELISA was performed to confirm the accessibility of anchored antigens. P. pastoris:ENV and P. pastoris:ENVNS1 showed higher immunoreactivity compared to non-recombinant yeast (NR), confirming protein anchoring (Figure 2).
3.2. Recombinant P. pastoris strains induce increased secretion of serum and splenic cytokines
The vaccine constructs promoted different cytokine patterns. Regarding serum cytokines, P. pastoris:ENV caused an increase in the levels of IL-2, TNF-α, and IL-17A. On the other hand, P. pastoris:ENVNS1 increased all analyzed cytokines, overcoming both non-recombinant yeast and P. pastoris:ENV, mainly to IL-2, IL-4, IL-10, and IL-1A (Figure 3). The response elicited by both recombinant yeasts was equivalent only to the TNF-α release. The profile of cytokines secreted by splenic lymphocytes, restimulated in vitro, was more restricted than that observed in animal serum, with a greater tendency towards a Th1 pattern. P. pastoris:ENV caused an increase in IL-6 (48h) and IFN-γ at 24h and 48h (Figure 4A,B), while P. pastoris:ENVNS1 induced IL-6 and TNF-α in 72h cultures (Figure 4A–C). There was no significant production of the other cytokines (Table S1). Non-recombinant yeast did not cause a significant stimulus compared to P. pastoris expressing vaccine antigens.
3.3. P. pastoris:ENV and P. pastoris:ENVNS1 enhance antibody production
The levels of different types and isotypes of immunoglobulins were assessed to evaluate the humoral response. Overall, the P. pastoris:ENV vaccine stood out compared to ENVNS1 and non-recombinant yeast (Figure 5). P. pastoris:ENV induced production of the IgG isotypes, all of them, especially IgG3. Both vaccine constructs elicited a similar profile for IgA and IgM (Figure 5E and 5G), superior to P. pastoris:nr. The two vaccines led to an increase in IgE, mainly P. pastoris:ENV (Figure 5F). The ratio IgG2a/IgG1 indicates the response profile type (Th1 or Th2) and was similar for all three yeast strains. Although the mean for P. pastoris:nr (1.27) was higher than the recombinants (1.14 and 1.03), the difference was not statistically significant (Figure 5H). Values greater than 1 indicate a tendency towards a Th1 response pattern. However, the proximity between the averages of IgG1 and IgG2a for all groups suggested a balance between Th1 and Th2 responses.
3.4. Recombinant yeasts stimulate cellular responses
The evaluation of the cellular response induced by vaccination was based on the analysis of CD4+, CD8+, and CD16+ T lymphocyte populations present in the blood and spleen after immunization. Regarding circulating lymphocytes, mice inoculated with non-recombinant P. pastoris promoted a higher stimulus to CD4+ T lymphocytes. No difference in the number of CD4+ T cells between the recombinant yeasts was observed (Figure 6A). On the other hand, about the number of CD8+ T cells, P. pastoris:nr and P. pastoris:ENVNS1 showed a similar pattern, superior to that induced by P. pastoris:ENV (Figure 6B). To Natural Killer (NK) CD16+ T lymphocytes, P. pastoris:ENV and P. pastoris:ENVNS1 promoted an increase in the number of cells, compared to the non-recombinant (Figure 6C).
Lymphocytes isolated from the spleen of the immunized animals were restimulated with recombinant yeasts and incubated for periods of 24h, 48h, and 72h. The most significant stimulus was observed for the CD4+ T lymphocyte subpopulation. The immunization with the recombinant yeasts, expressing ENV and ENVNS1, induced an increase in the amount of CD4+ T cells, especially when observing the 72h culture, where both were superior to non-recombinant P.pastoris (Figure 7A). The levels of CD8+ T cells were lower than the expression of CD4+ and CD16+ and substantially reduced throughout the culture period. The proportion of CD8+ T cells stimulated by P. pastoris:ENV stood out at times of 24h and 48h. After 72 hours of stimulus, the animals inoculated with P. pastoris:ENVNS1 had a higher number of CD8+ T lymphocytes (Figure 7B). About the CD16+ T cells profile, P. pastoris:ENV induced a significant increase in the first 24h however declined in 48h and 72h. Meanwhile, P. pastoris:ENVNS1 caused a more significant expansion in 48h and 72h, even though it also diminished in 72h (Figure 7C).
3.5. The whole yeast vaccines do not cause significant side effects
Animals were weighed on the first day of the immunization regimen, on the booster dose day, and 21 days after the initial dose (Table S2). Throughout the vaccine schedule, there was no weight loss or significant behavioral changes. There were no points of inflammation, swellings, neither adverse effect in the injection site. Overall, there were no biochemical alterations that would qualify clinical disturbs (Table 1). The hematological changes were relative leukocytosis and lymphopenia, besides neutrophilia in the three experimental groups.
4. Discussion
In this study, we evaluated a multiepitope vaccine based on B and T cell epitopes from Env and NS1 ZIKV proteins in a yeast surface display strategy. After a two-dose schedule, the immune response was assessed through cytokine dosage, immunophenotyping, and immunoglobulin production. BALB/c mice immunized with recombinant P. pastoris showed a serum elevation of IL-2 and TNF-α for the two tested vaccines, in addition to the induction of IL-17A. Noteworthy, besides its conventional regulatory role, IL-17A may play pro-inflammatory functions in some viral, fungal, and cancer infections [32]. Moreover, high levels of IFN-γ, TNF-α, and IL-2 secreted by CD4+ and CD8+ T lymphocytes are also detected in immunocompetent C57BL/6 mice infected with ZIKV, exhibiting a Th1 pattern [33,34].
Despite eliciting Th1 cytokines, the P. pastoris:ENVNS1 vaccine induced an increase of IL-4 and IL-10, which may be a consequence of follicular T cells (Thf) activation. In Zika virus infection, Tfh cells enhanced production of cytokines such as IL-4 and IL-21, nevertheless also act in a Th1-like manner, producing IFN-γ. Besides, this lymphocyte subpopulation influences the development of neutralizing antibodies against ZIKV [35]. The detection of cytokines as IL-2, IL-4, and IL-17 in individuals in the acute phase of ZIKV infection suggests a polyfunctional response profile characterized by Th1, Th2, and Th17 responses [36]. Concerning the splenic immune cells, the cytokine dosage indicated a Th1 profile, where P. pastoris:ENV induced an increase in IL-6 and IFN-γ levels, while P. pastoris:ENVNS1 stimulated IL-6 and TNF-α production. These pro-inflammatory cytokines compose the anti-ZIKV immune response and were also observed in vitro assays performed with these recombinant yeasts in a previous study [25,37]. The role of IL-6 resulting of the administration of whole yeast vaccines has been linked to the generation of CD4+ T cells [38]. Furthermore, increased production of TNF-α and IFN-γ is related to the activation of CD4+ and CD8+ T cells that mediate effective responses against ZIKV-infected cells [39].
Regarding circulating lymphocytes the recombinant antigens seem to modulate the CD8+ T cells since there was a significant difference between the vaccines. Non-recombinant P. pastoris and expressing ENVNS1 induced a greater stimulus than P. pastoris:ENV. The NS1 epitopes may have influenced the activation of the CD8+ T lymphocytes, which reinforces the importance of including immunodominant epitopes present in non-structural proteins in the design of vaccines for flaviviruses [39]. In addition, both recombinant yeasts elicited the expansion on the number of CD16+ T lymphocytes, mainly P. pastoris: ENVNS1 construction.
Analysis of splenic lymphocytes reflects the induction of immune memory responses [40]. The stimulus promoted by recombinant yeasts was greater for the CD4+ T cell population than for CD8+ T cells overall. P. pastoris:ENV stimulated CD4+ (48h and 72h), CD8+ (24h and 48h), and CD16+ (24h and 72h) cells. P. pastoris:ENVNS1 elicited an increase in the number of CD4+ cells (24h and 72h), CD8+ (72h) even in low levels, and CD16+ (48h and 72h). The vaccine responses here reported are in line with other studies about the importance of T cells in the context of vaccine development, indicating that a coordinated balance among the action of antibodies, Tfh CD4+, Th1 CD4+, and CD8+ T cells are essential to confer infection control and long-term protection [39,41].
Besides the cellular immune response, the recombinant yeasts induced significant production of different classes of immunoglobulins. The highest levels were observed for IgG3, IgA, and IgM. The P. pastoris:ENV vaccine promoted a significant increase in the production of all IgG isotypes. Elevations in the generation of IgG1 and IgG2a were observed in mice challenged with ZIKV PE243 [42] as well as immunized with VLPs [43]. The IgG2 isotype is predominant among neutralizing antibodies that induce protection and is important for ZIKV infection clearance [35,42]. Besides, there were elevations in IgG3 and IgM, commonly observed in the acute phase of viral infections, involved in pro-inflammatory effector mechanisms [43,44].
Both recombinant yeasts caused an increase in IgE. Antiviral functions performed by IgE have been investigated, but there is no consistent data regarding the role of this immunoglobulin in ZIKV infection [45]. Once there were no changes in eosinophil levels or signs of anaphylaxis, the contribution of IgE was possibly related to non-allergic defense mechanisms induced by the vaccine. The two vaccine constructs also promoted an increase in IgA and IgM at equivalent levels. These immunoglobulins are frequent in recent ZIKV infections, making them helpful to diagnostic assays [46].
All tested yeasts induced neutrophilia in inoculated mice. Besides acting as critical cells of innate immunity, neutrophils also contribute to adaptive immunity in the transport and presentation of antigens and the regulation of antigen-specific responses [47]. Although neutrophil activation is involved in the pathogenesis of ZIKV infection, the role of these cells in the antiviral immune response generated by vaccination is unclear [48].
Overall, the results obtained suggest the involvement of Th1 and Th2 responses, providing a polyfunctional immune response profile also observed in the ZIKV infection [33]. Similar patterns are observed in vaccine studies for ZIKV based on different platforms, such as EDIII-based subunit vaccines [49] and VLPs [50]. This study explored the biotechnological potential of P. pastoris regarding its applicability as a biofactory and antigens carrier. We observed that although previous studies show yeast immunostimulatory properties, the expression of vaccine antigens can modulate the immune response. Recombinant P. pastoris promoted an increase in immunoglobulins production and cellular immune response activation. There were no deaths, diarrhea, or weight loss in animals vaccinated with yeasts during the vaccination schedule, in agreement with previous studies using P. pastoris and other species [51].
5. Conclusions
The present work contributes with information both in the field of vaccine development for ZIKV and in the establishment of yeasts as vaccine vectors. The data presented demonstrate the development of a vaccine strategy against ZIKV based on the use of P. pastoris as a carrier of viral antigens. Despite the several immunoinformatics-based studies for predicting ZIKV epitopes, researches at the preclinical level are still scarce. The study presented here demonstrates for the first time the evaluation of a multi-epitope construct as a vaccine antigen for ZIKV, using yeast as a vehicle, through the surface display system. Together, the results point to the generation of immune responses that should be tested to verify the induction of protection against ZIKV infection. The biotechnological potential of P. pastoris was explored in different ways: as a biofactory, as an antigen carrier, and as a vaccine adjuvant, showing its versatility. The expression vector also allows the cloning of other genes for vaccine strategies aimed at other infectious agents. P. pastoris is pointed out as a cost-effective platform for vaccine production, favoring production and distribution in Low- and Middle-Income Countries. The present study brings encouraging data regarding the use of yeast as an effective and safe vehicle for the delivery of vaccine antigens.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Secretion of splenic cytokines after in vitro re-stimulus; Table S2: Table S2: Average weight of the mice groups throughout the immunization experiment. Values correspond to mean ± standard deviation.
Author Contributions
Conceptualization: Anna J. D. Silva, André L. S. de Jesus, Cristiane M. L. de Melo, Antonio C. de Freitas; Methodology: Anna J. D. Silva, Lígia R. S. Leal, Larissa S. Macêdo, Bárbara R. da S. Barros, Georon F. de Sousa, Simone da P. L. Alves, Cristiane M. L. de Melo; Data curation: Anna J. D. Silva; Formal analysis: Anna J. D. Silva, Cristiane M. L. de Melo; Original draft preparation: Anna J. D. Silva; Writing - review & editing: Anna J. D. Silva, André L. S. de Jesus, Cristiane M. L. de Melo, Antonio C. de Freitas; Supervision: Lindomar J. Pena, Antonio C. de Freitas; Funding acquisition: Antonio C. de Freitas. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CNPq Bolsa Produtividade, grant number 308489/2019-5; CAPES – Epidemias, grant number 88887.507421/ 2020-00; CNPq/MCTI/CT-BIOTEC Nº 31/2022, grant number 440315/2022-0 and Programa de Apoio a Fixação de Jovens Doutores em Pernambuco APQ-1482-2.02/22.
Data Availability Statement
Data sharing is not applicable to this article.
Acknowledgments
The authors would like to thank NPT1 (Nucleo de Plataformas Tecnologicas) and bioterium from FIOCRUZ/Aggeu Magalhaes Institute for the use of its facilities; and Dr. Fabricio Souto (Federal University of Pernambuco, PE, Brazil) for assistance during the flow cytometry assays.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, H.; Yoo, S.J.; Kang, H.A. Yeast Synthetic Biology for the Production of Recombinant Therapeutic Proteins. FEMS Yeast Res 2014, n/a-n/a. [CrossRef]
- Stubbs, A.C.; Martin, K.S.; Coeshott, C.; Skaates, S.V.; Kuritzkes, D.R.; Bellgrau, D.; Franzusoff, A.; Duke, R.C.; Wilson, C.C. Whole Recombinant Yeast Vaccine Activates Dendritic Cells and Elicits Protective Cell-Mediated Immunity. Nat Med 2001, 7, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Vogl, T.; Hartner, F.S.; Glieder, A. New Opportunities by Synthetic Biology for Biopharmaceutical Production in Pichia Pastoris. Current Opinion in Biotechnology 2013, 24, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Kumar, P. Yeast-Based Vaccines: New Perspective in Vaccine Development and Application. FEMS Yeast Research 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Ardiani, A.; Higgins, J.P.; Hodge, J.W. Vaccines Based on Whole Recombinant Saccharomyces Cerevisiae Cells: Yeast-Based Vaccines. FEMS Yeast Research 2010, 10, 1060–1069. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.; Cheng, Y.; Wang, Z.; Hu, Y.; Zhang, X.; Wu, M.; Chen, Z.; Shi, B.; Sun, S.; Shen, Y.; et al. Whole Recombinant Hansenula Polymorpha Expressing Hepatitis B Virus Surface Antigen (Yeast-HBsAg) Induces Potent HBsAg-Specific Th1 and Th2 Immune Responses. Vaccine 2009, 28, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Bazan, S.B.; Geginat, G.; Breinig, T.; Schmitt, M.J.; Breinig, F. Uptake of Various Yeast Genera by Antigen-Presenting Cells and Influence of Subcellular Antigen Localization on the Activation of Ovalbumin-Specific CD8 T Lymphocytes. Vaccine 2011, 29, 8165–8173. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Jin, S.; Karlsson, E.; Schultz-Cherry, S.; Ye, K. Yeast Surface-Displayed H5N1 Avian Influenza Vaccines. Journal of Immunology Research 2016, 2016, 1–12. [Google Scholar] [CrossRef]
- Jacob, D.; Ruffie, C.; Dubois, M.; Combredet, C.; Amino, R.; Formaglio, P.; Gorgette, O.; Pehau-Arnaudet, G.; Guery, C.; Puijalon, O.; et al. Whole Pichia Pastoris Yeast Expressing Measles Virus Nucleoprotein as a Production and Delivery System to Multimerize Plasmodium Antigens. PLoS ONE 2014, 9, e86658. [Google Scholar] [CrossRef]
- Sun, H.; Wang, L.; Wang, T.; Zhang, J.; Liu, Q.; Chen, P.; Chen, Z.; Wang, F.; Li, H.; Xiao, Y.; et al. Display of Eimeria Tenella EtMic2 Protein on the Surface of Saccharomyces Cerevisiae as a Potential Oral Vaccine against Chicken Coccidiosis. Vaccine 2014, 32, 1869–1876. [Google Scholar] [CrossRef]
- Gebauer, M.; Hürlimann, H.C.; Behrens, M.; Wolff, T.; Behrens, S.-E. Subunit Vaccines Based on Recombinant Yeast Protect against Influenza A Virus in a One-Shot Vaccination Scheme. Vaccine 2019, 37, 5578–5587. [Google Scholar] [CrossRef]
- Kim, J.-M.; Jung, D.-I.; Eom, Y.J.; Park, S.-M.; Yoo, H.-S.; Jang, Y.-S.; Yang, M.-S.; Kim, D.-H. Surface-Displayed Expression of a Neutralizing Epitope of ApxIIA Exotoxin in Saccharomyces Cerevisiae and Oral Administration of It for Protective Immune Responses against Challenge by Actinobacillus Pleuropneumoniae. Bioscience, Biotechnology, and Biochemistry 2010, 74, 1362–1367. [Google Scholar] [CrossRef] [PubMed]
- King, T.H.; Kemmler, C.B.; Guo, Z.; Mann, D.; Lu, Y.; Coeshott, C.; Gehring, A.J.; Bertoletti, A.; Ho, Z.Z.; Delaney, W.; et al. A Whole Recombinant Yeast-Based Therapeutic Vaccine Elicits HBV X, S and Core Specific T Cells in Mice and Activates Human T Cells Recognizing Epitopes Linked to Viral Clearance. PLoS ONE 2014, 9, e101904. [Google Scholar] [CrossRef]
- Wasilenko, J.L.; Sarmento, L.; Spatz, S.; Pantin-Jackwood, M. Cell Surface Display of Highly Pathogenic Avian Influenza Virus Hemagglutinin on the Surface of Pichia Pastoris Cells Using α-Agglutinin for Production of Oral Vaccines. Biotechnol Progress 2009, NA–NA. [Google Scholar] [CrossRef] [PubMed]
- Shibasaki, S.; Maeda, H.; Ueda, M. Molecular Display Technology Using Yeast—Arming Technology—. Anal. Sci. 2009, 25, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Ananphongmanee, V.; Srisala, J.; Sritunyalucksana, K.; Boonchird, C. Yeast Surface Display of Two Proteins Previously Shown to Be Protective Against White Spot Syndrome Virus (WSSV) in Shrimp. PLoS ONE 2015, 10, e0128764. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K.; Ueda, M. Cell Surface Engineering of Yeast for Applications in White Biotechnology. Biotechnol Lett 2011, 33, 1–9. [Google Scholar] [CrossRef]
- Garcia Serpa Osorio-de-Castro, C.; Silva Miranda, E.; Machado de Freitas, C.; Rochel de Camargo, K.; Cranmer, H.H. The Zika Virus Outbreak in Brazil: Knowledge Gaps and Challenges for Risk Reduction. Am J Public Health 2017, 107, 960–965. [Google Scholar] [CrossRef]
- Morabito, K.M.; Graham, B.S. Zika Virus Vaccine Development. The Journal of Infectious Diseases 2017, 216, S957–S963. [Google Scholar] [CrossRef]
- Musso, D.; Ko, A.I.; Baud, D. Zika Virus Infection — After the Pandemic. N Engl J Med 2019, 381, 1444–1457. [Google Scholar] [CrossRef]
- Zhou, K.; Li, C.; Shi, W.; Hu, X.; Nandakumar, K.S.; Jiang, S.; Zhang, N. Current Progress in the Development of Zika Virus Vaccines. Vaccines 2021, 9, 1004. [Google Scholar] [CrossRef]
- Hassert, M.; Wolf, K.J.; Schwetye, K.E.; DiPaolo, R.J.; Brien, J.D.; Pinto, A.K. CD4+T Cells Mediate Protection against Zika Associated Severe Disease in a Mouse Model of Infection. PLoS Pathog 2018, 14, e1007237. [Google Scholar] [CrossRef] [PubMed]
- Elong Ngono, A.; Young, M.P.; Bunz, M.; Xu, Z.; Hattakam, S.; Vizcarra, E.; Regla-Nava, J.A.; Tang, W.W.; Yamabhai, M.; Wen, J.; et al. CD4+ T Cells Promote Humoral Immunity and Viral Control during Zika Virus Infection. PLoS Pathog 2019, 15, e1007474. [Google Scholar] [CrossRef]
- Antonelli, A.C.B.; Almeida, V.P.; de Castro, F.O.F.; Silva, J.M.; Pfrimer, I.A.H.; Cunha-Neto, E.; Maranhão, A.Q.; Brígido, M.M.; Resende, R.O.; Bocca, A.L.; et al. In Silico Construction of a Multiepitope Zika Virus Vaccine Using Immunoinformatics Tools. Sci Rep 2022, 12, 53. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.J.D.; Jesus, A.L.S.; Leal, L.R.S.; Silva, G.A.S.; Melo, C.M.L.; Freitas, A.C. Pichia Pastoris Displaying ZIKV Protein Epitopes from the Envelope and NS1 Induce in Vitro Immune Activation. Vaccine 2021, 39, 2545–2554. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, J.R.M.; de Moraes, L.M.P.; Torres, F.A.G. Molecular Characterization of the 3-Phosphoglycerate Kinase Gene (PGK1) from the Methylotrophic YeastPichia Pastoris. Yeast 2005, 22, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Gu, L.; Zheng, P.; Chen, Z.; Dou, X.; Qin, Q.; Cai, X. Improvement of Cell Counting Method for Neubauer Counting Chamber. J Clin Lab Anal 2020, 34. [Google Scholar] [CrossRef] [PubMed]
- Fojon Martinez Reboredo, S. [Modification of the Fonio method of counting platelets and other components of the blood]. Laboratorio 1958, 26, 511–517. [Google Scholar]
- Almeida, A.S.; Faleiros, A.C.G.; Teixeira, D.N.S.; Cota, U.A.; Chica, J.E.L. Valores de Referência de Parâmetros Bioquímicos No Sangue de Duas Linhagens de Camundongos. J. Bras. Patol. Med. Lab. 2008, 44, 429–432. [Google Scholar] [CrossRef]
- Barbosa, B.D.S.; Praxedes, É.A.; Lima, M.A.; Pimentel, M.M.L.; Santos, F.A.; Brito, P.D.; Lelis, I.C.N.G.; Macedo, M.F. de; Bezerra, M.B. Haematological and Biochemical Profile of Balb-c Mice. Acta Scientiae. Vet. 2017, 45, 5. [Google Scholar] [CrossRef]
- Silva-Santana, G.; Bax, J.C.; Fernandes, D.C.S.; Bacellar, D.T.L.; Hooper, C.; Dias, A.A.S.O.; Silva, C.B.; Souza, A.M.; Ramos, S.; Santos, R.A.; et al. Clinical Hematological and Biochemical Parameters in Swiss, BALB/c, C57BL/6 and B6D2F1 Mus Musculus. Anim. Models Exp. Med. 2020, 3, 304–315. [Google Scholar] [CrossRef]
- Reynolds, J.M.; Angkasekwinai, P.; Dong, C. IL-17 Family Member Cytokines: Regulation and Function in Innate Immunity. Cytokine & Growth Factor Reviews 2010, 21, 413–423. [Google Scholar] [CrossRef]
- Lima, N.S.; Rolland, M.; Modjarrad, K.; Trautmann, L. T Cell Immunity and Zika Virus Vaccine Development. Trends in Immunology 2017, 38, 594–605. [Google Scholar] [CrossRef] [PubMed]
- Pardy, R.D.; Rajah, M.M.; Condotta, S.A.; Taylor, N.G.; Sagan, S.M.; Richer, M.J. Analysis of the T Cell Response to Zika Virus and Identification of a Novel CD8+ T Cell Epitope in Immunocompetent Mice. PLoS Pathog 2017, 13, e1006184. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Tang, J.; Liu, Z.; Liu, Y.; Huang, Y.; Xu, Y.; Hao, P.; Yin, Z.; Zhong, J.; Ye, L.; et al. ZIKV Infection Induces Robust Th1-like Tfh Cell and Long-Term Protective Antibody Responses in Immunocompetent Mice. Nat Commun 2019, 10, 3859. [Google Scholar] [CrossRef] [PubMed]
- Tappe, D.; Pérez-Girón, J.V.; Zammarchi, L.; Rissland, J.; Ferreira, D.F.; Jaenisch, T.; Gómez-Medina, S.; Günther, S.; Bartoloni, A.; Muñoz-Fontela, C.; et al. Cytokine Kinetics of Zika Virus-Infected Patients from Acute to Reconvalescent Phase. Med Microbiol Immunol 2016, 205, 269–273. [Google Scholar] [CrossRef]
- da Silva, M.H.M.; Moises, R.N.C.; Alves, B.E.B.; Pereira, H.W.B.; de Paiva, A.A.P.; Morais, I.C.; Nascimento, Y.M.; Monteiro, J.D.; de Souto, J.T.; Nascimento, M.S.L.; et al. Innate Immune Response in Patients with Acute Zika Virus Infection. Med Microbiol Immunol 2019, 208, 703–714. [Google Scholar] [CrossRef]
- Huang, H.; Li, S.; Zhang, Y.; Han, X.; Jia, B.; Liu, H.; Liu, D.; Tan, S.; Wang, Q.; Bi, Y.; et al. CD8 + T Cell Immune Response in Immunocompetent Mice during Zika Virus Infection. J. Virol. 2017, 91, e00900–17. [Google Scholar] [CrossRef]
- Li, G.; Teleki, C.; Wang, T. Memory T Cells in Flavivirus Vaccination. Vaccines 2018, 6, 73. [Google Scholar] [CrossRef]
- Flaxman, A.; Ewer, K. Methods for Measuring T-Cell Memory to Vaccination: From Mouse to Man. Vaccines 2018, 6, 43. [Google Scholar] [CrossRef]
- Grubor-Bauk, B.; Wijesundara, D.K.; Masavuli, M.; Abbink, P.; Peterson, R.L.; Prow, N.A.; Larocca, R.A.; Mekonnen, Z.A.; Shrestha, A.; Eyre, N.S.; et al. NS1 DNA Vaccination Protects against Zika Infection through T Cell–Mediated Immunity in Immunocompetent Mice. Sci. Adv. 2019, 5, eaax2388. [Google Scholar] [CrossRef]
- Lucas, C.G.O.; Kitoko, J.Z.; Ferreira, F.M.; Suzart, V.G.; Papa, M.P.; Coelho, S.V.A.; Cavazzoni, C.B.; Paula-Neto, H.A.; Olsen, P.C.; Iwasaki, A.; et al. Critical Role of CD4+ T Cells and IFNγ Signaling in Antibody-Mediated Resistance to Zika Virus Infection. Nat Commun 2018, 9, 3136. [Google Scholar] [CrossRef] [PubMed]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG Subclasses and Allotypes: From Structure to Effector Functions. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Ximenes, R.A. de A.; Miranda-Filho, D. de B.; Brickley, E.B.; Montarroyos, U.R.; Martelli, C.M.T.; Araújo, T.V.B. de; Rodrigues, L.C.; de Albuquerque, M. de F.P.M.; de Souza, W.V.; Castanha, P.M. da S.; et al. Zika Virus Infection in Pregnancy: Establishing a Case Definition for Clinical Research on Pregnant Women with Rash in an Active Transmission Setting. PLoS Negl Trop Dis 2019, 13, e0007763. [Google Scholar] [CrossRef]
- Smith-Norowitz, T.A.; Wong, D.; Kusonruksa, M.; Norowitz, K.B.; Joks, R.; Durkin, H.G.; Bluth, M.H. Long Term Persistence of IgE Anti-Influenza Virus Antibodies in Pediatric and Adult Serum Post Vaccination with Influenza Virus Vaccine. Int. J. Med. Sci. 2011, 8, 239–244. [Google Scholar] [CrossRef]
- Warnecke, J.M.; Lattwein, E.; Saschenbrecker, S.; Stöcker, W.; Schlumberger, W.; Steinhagen, K. Added Value of IgA Antibodies against Zika Virus Non-Structural Protein 1 in the Diagnosis of Acute Zika Virus Infections. Journal of Virological Methods 2019, 267, 8–15. [Google Scholar] [CrossRef]
- Vono, M.; Lin, A.; Norrby-Teglund, A.; Koup, R.A.; Liang, F.; Loré, K. Neutrophils Acquire the Capacity for Antigen Presentation to Memory CD4+ T Cells in Vitro and Ex Vivo. Blood 2017, 129, 1991–2001. [Google Scholar] [CrossRef]
- Muralidharan, A.; Reid, S.P. Complex Roles of Neutrophils during Arboviral Infections. Cells 2021, 10, 1324. [Google Scholar] [CrossRef]
- Cibulski, S.; Varela, A.P.M.; Teixeira, T.F.; Cancela, M.P.; Sesterheim, P.; Souza, D.O.; Roehe, P.M.; Silveira, F. Zika Virus Envelope Domain III Recombinant Protein Delivered With Saponin-Based Nanoadjuvant From Quillaja Brasiliensis Enhances Anti-Zika Immune Responses, Including Neutralizing Antibodies and Splenocyte Proliferation. Front. Immunol. 2021, 12, 632714. [Google Scholar] [CrossRef]
- Dai, S.; Zhang, T.; Zhang, Y.; Wang, H.; Deng, F. Zika Virus Baculovirus-Expressed Virus-Like Particles Induce Neutralizing Antibodies in Mice. Virol. Sin. 2018, 33, 213–226. [Google Scholar] [CrossRef]
- Becerril-García, M.Á.; Flores-Maldonado, O.E.; González, G.M.; García-González, G.; Hernández-Bello, R.; Palma-Nicolás, J.P. Safety Profile of Intravenous Administration of Live Pichia Pastoris Cells in Mice. FEMS Yeast Research 2022, 22, foac023. [Google Scholar] [CrossRef]
Figure 1.
Experimental design. A) The first phase of the study included the selection of immunogenic epitopes from the envelope and NS1 proteins (Silva et al. 2021); B) Expression cassettes: the sets of Env and Env + NS1 epitopes were fused to the α-agglutinin anchor protein, allowing the display of the recombinant proteins on the P. pastoris surface. PGK1: promoter; MF-α: signal peptide for protein secretion; 6H: 6x His-tag for immunodetection. C) The yeasts were subjected to heat treatment for metabolic inactivation and used for vaccination of BALB/c mice. The mice received two doses of each yeast preparation and were monitored for weight and activity throughout the 21 days of the experiment. On the last day of the schedule were performed blood collection, euthanasia, and removal of the spleen..
Figure 1.
Experimental design. A) The first phase of the study included the selection of immunogenic epitopes from the envelope and NS1 proteins (Silva et al. 2021); B) Expression cassettes: the sets of Env and Env + NS1 epitopes were fused to the α-agglutinin anchor protein, allowing the display of the recombinant proteins on the P. pastoris surface. PGK1: promoter; MF-α: signal peptide for protein secretion; 6H: 6x His-tag for immunodetection. C) The yeasts were subjected to heat treatment for metabolic inactivation and used for vaccination of BALB/c mice. The mice received two doses of each yeast preparation and were monitored for weight and activity throughout the 21 days of the experiment. On the last day of the schedule were performed blood collection, euthanasia, and removal of the spleen..
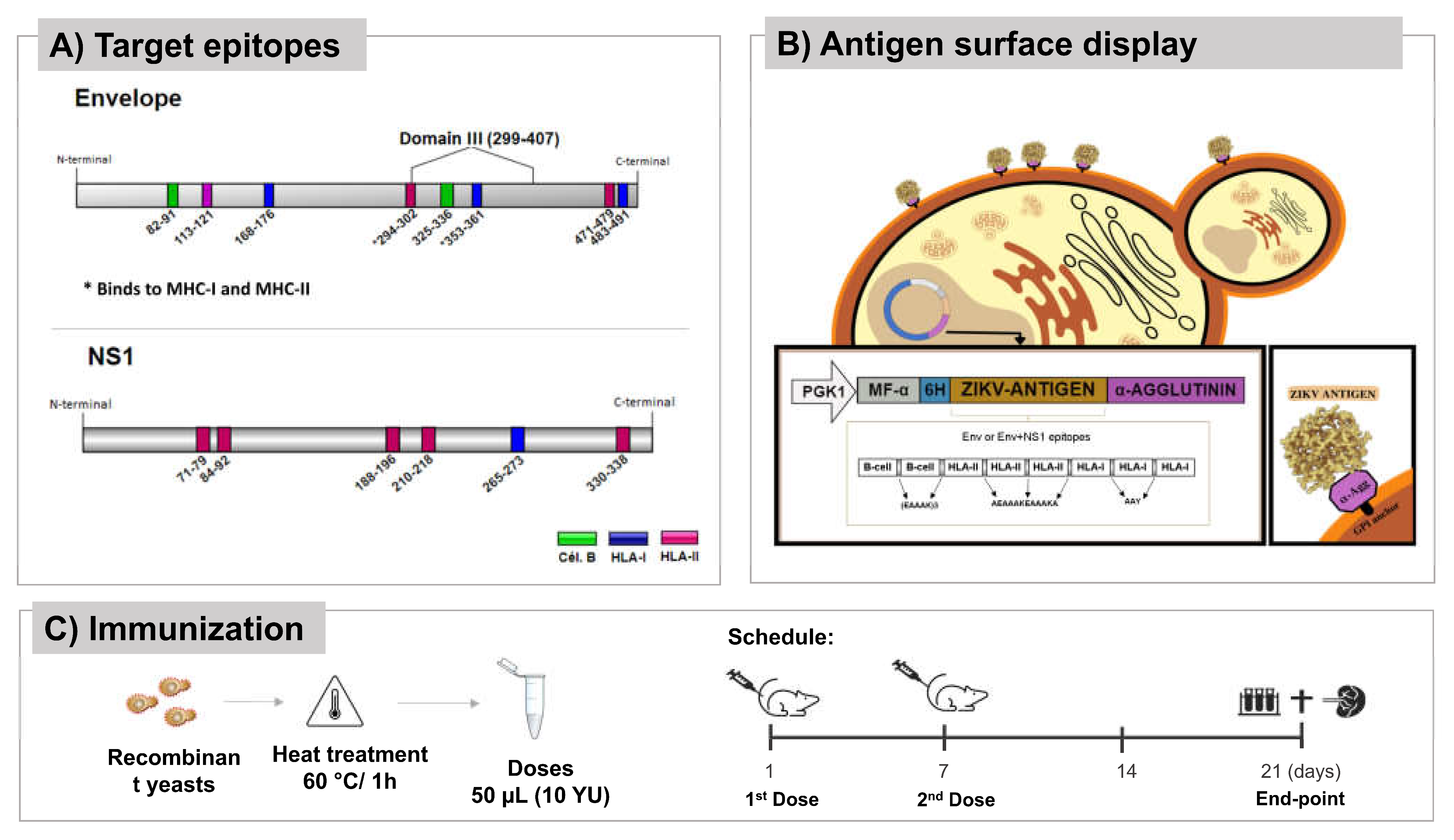
Figure 2.
Yeast-ELISA performed to evaluate the protein expression and anchorage of the yeasts utilized in the immunization assay.
Figure 2.
Yeast-ELISA performed to evaluate the protein expression and anchorage of the yeasts utilized in the immunization assay.
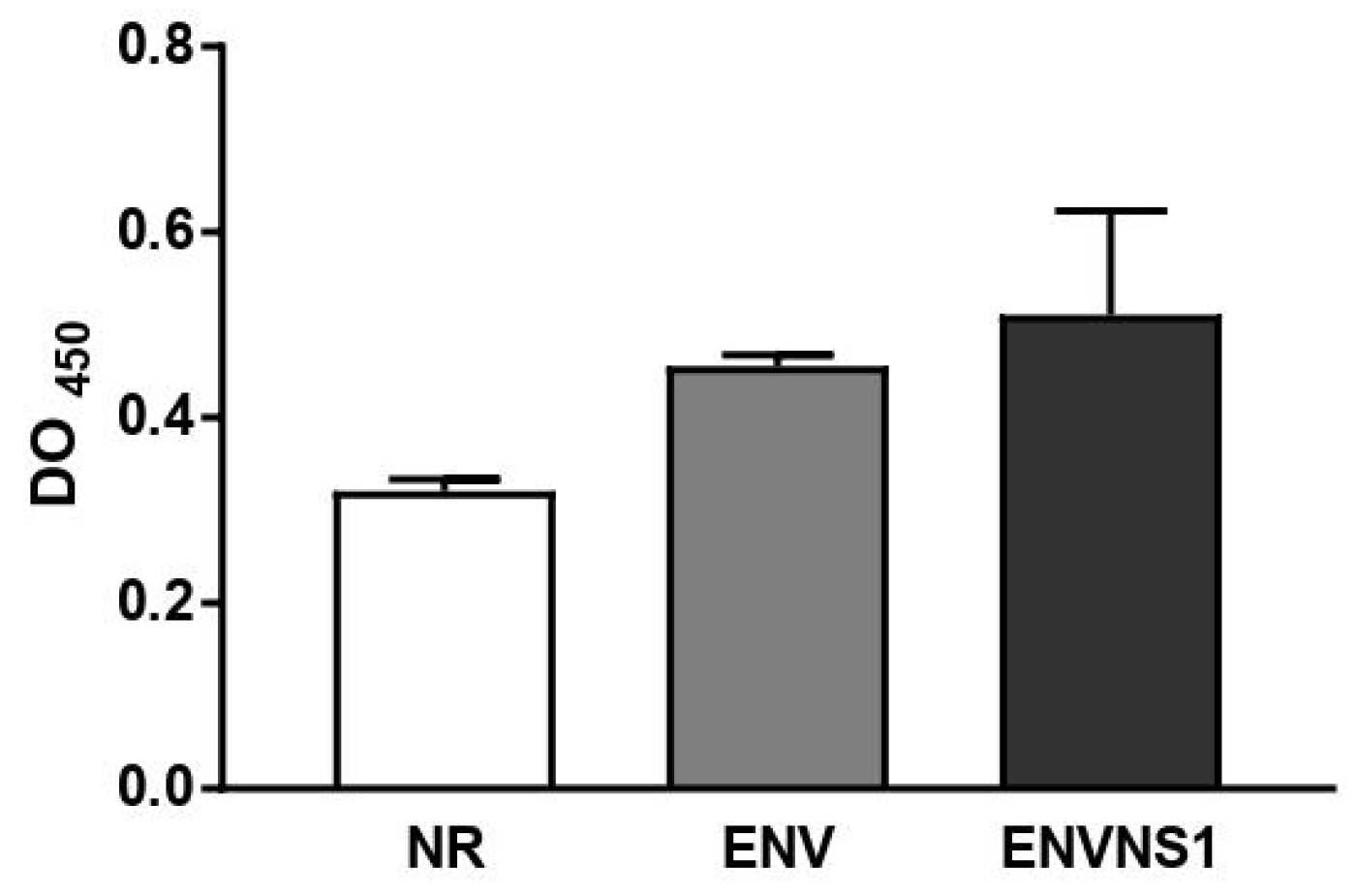
Figure 3.
Cytokines measured from the blood of mice 14 days after the booster dose. A) IL-2, B) IL-4, C) IL-17A, D) TNF-α, E) IFN-γ, F) IL-6, G) IL-10. Cytokine values were measured in pg.mL-1. Asterisks represent statistical significances (*p < 0.05, **p < 0.01, *** p<0.001). Error bars: standard deviation.
Figure 3.
Cytokines measured from the blood of mice 14 days after the booster dose. A) IL-2, B) IL-4, C) IL-17A, D) TNF-α, E) IFN-γ, F) IL-6, G) IL-10. Cytokine values were measured in pg.mL-1. Asterisks represent statistical significances (*p < 0.05, **p < 0.01, *** p<0.001). Error bars: standard deviation.
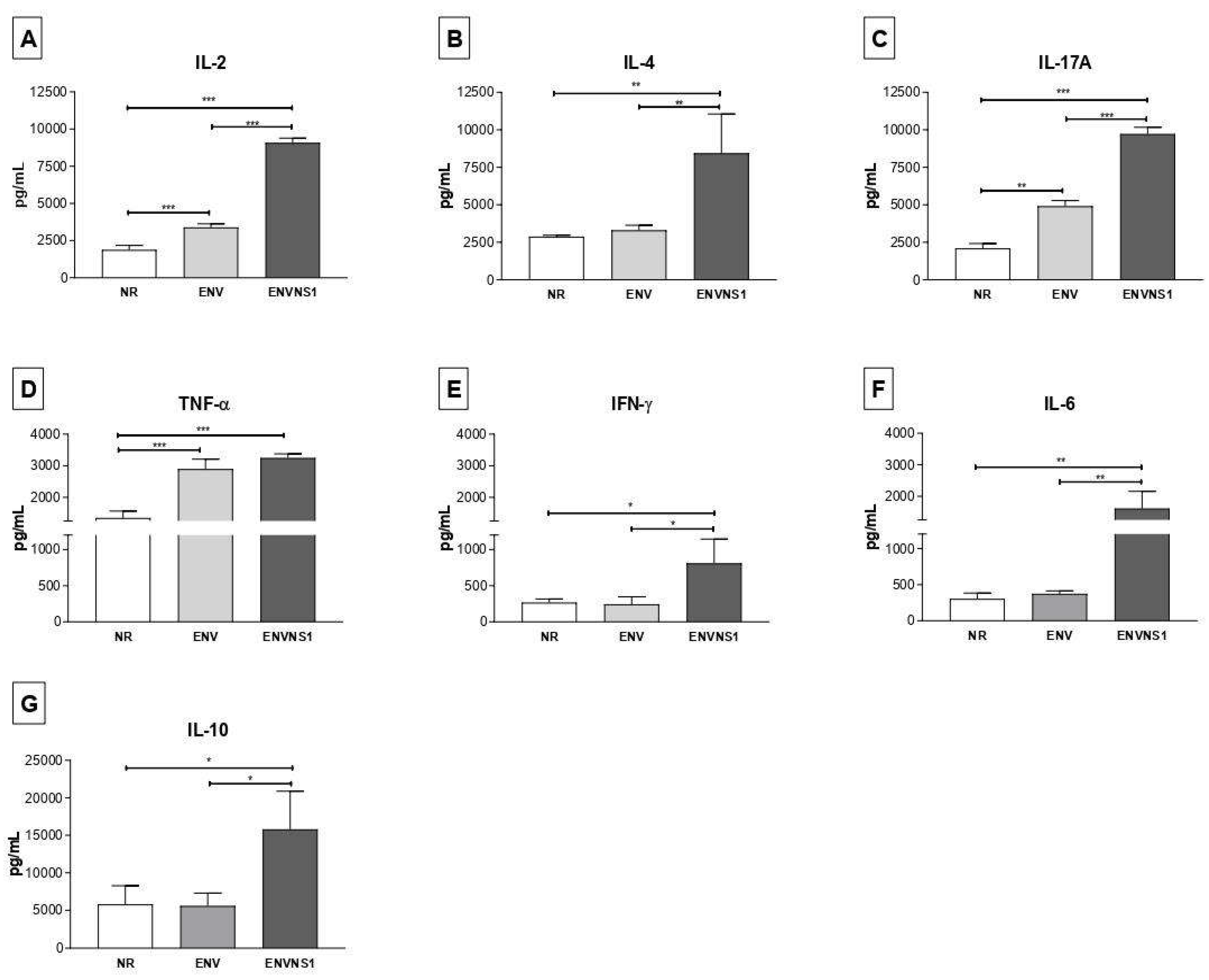
Figure 4.
Dosage cytokines in immune cells isolated from the spleens, detection obtained from the supernatant of cultures. A) IL-6, B) IFN-γ, C) TNF-α. Cytokine values were measured in pg.mL-1. Asterisks represent statistical significances (*p < 0.05, **p < 0.01, *** p<0.001). Error bars: standard deviation.
Figure 4.
Dosage cytokines in immune cells isolated from the spleens, detection obtained from the supernatant of cultures. A) IL-6, B) IFN-γ, C) TNF-α. Cytokine values were measured in pg.mL-1. Asterisks represent statistical significances (*p < 0.05, **p < 0.01, *** p<0.001). Error bars: standard deviation.
Figure 5.
Yeast-induced humoral response. The immunoglobulins IgG1, IgG2a, IgG2b, IgG3, IgA, IgM, and IgE were detected by flow cytometry. The values in MFI correspond to the mean fluorescence intensity. Asterisks represent statistical significances (*p < 0.05, **p < 0.01, *** p<0.001). Error bars: standard deviation.
Figure 5.
Yeast-induced humoral response. The immunoglobulins IgG1, IgG2a, IgG2b, IgG3, IgA, IgM, and IgE were detected by flow cytometry. The values in MFI correspond to the mean fluorescence intensity. Asterisks represent statistical significances (*p < 0.05, **p < 0.01, *** p<0.001). Error bars: standard deviation.
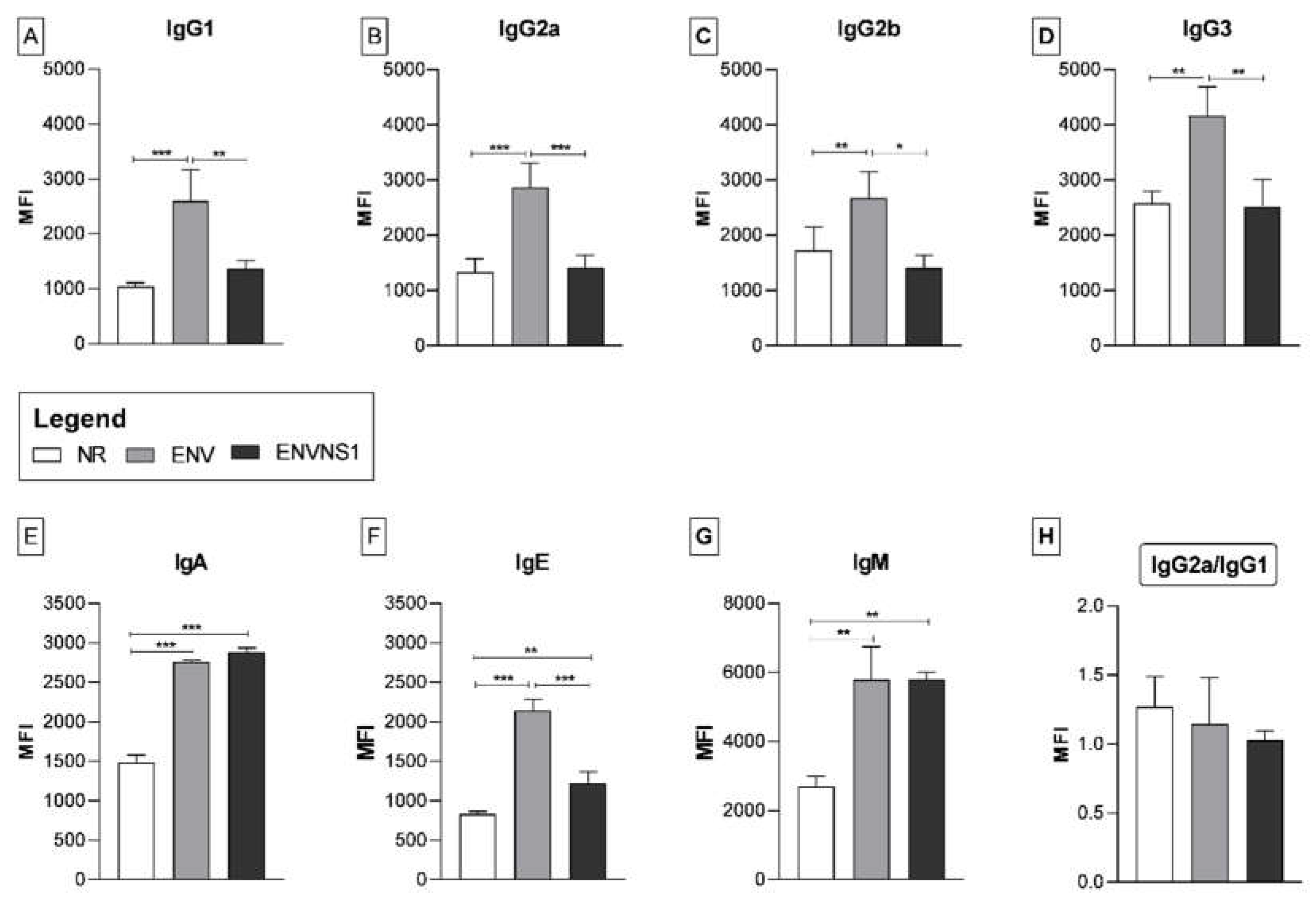
Figure 6.
Expression of surface markers in lymphocyte subpopulations present in the blood of mice after immunization. A) % of cells CD4+, B) % of cells CD8+, C) % of cells CD16+. Asterisks represent statistical significances (*p < 0.05, **p < 0.01, *** p<0.001). Error bars: standard deviation.
Figure 6.
Expression of surface markers in lymphocyte subpopulations present in the blood of mice after immunization. A) % of cells CD4+, B) % of cells CD8+, C) % of cells CD16+. Asterisks represent statistical significances (*p < 0.05, **p < 0.01, *** p<0.001). Error bars: standard deviation.
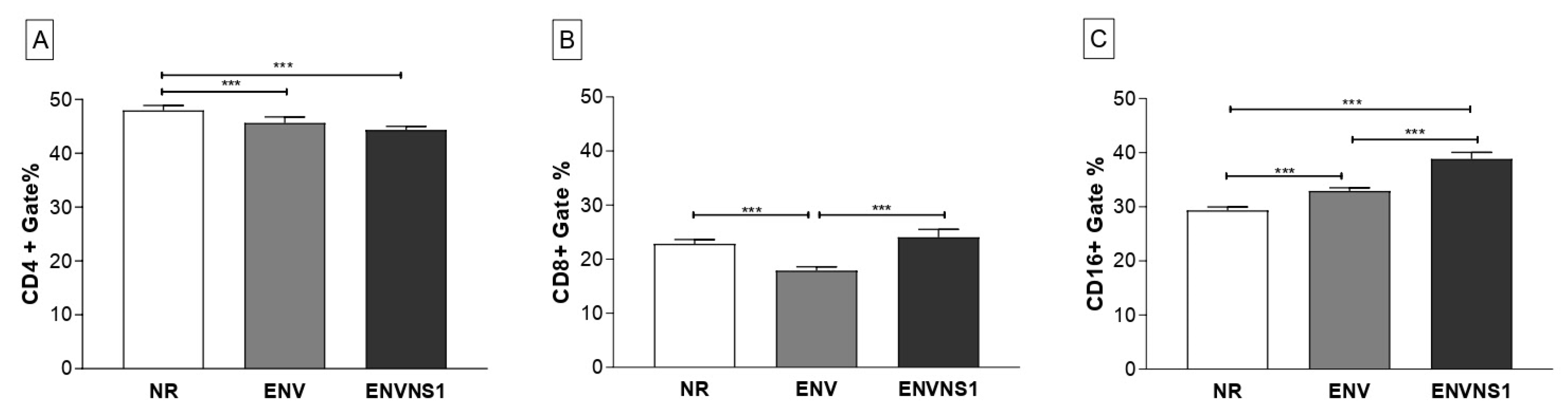
Figure 7.
Expression profile of splenic lymphocyte surface markers. A) % of cells CD4+, B) % of cells CD8+, C) % of cells CD16+. Asterisks represent statistical significances (*p < 0.05, **p < 0.01, *** p<0.001). Error bars: standard deviation.
Figure 7.
Expression profile of splenic lymphocyte surface markers. A) % of cells CD4+, B) % of cells CD8+, C) % of cells CD16+. Asterisks represent statistical significances (*p < 0.05, **p < 0.01, *** p<0.001). Error bars: standard deviation.
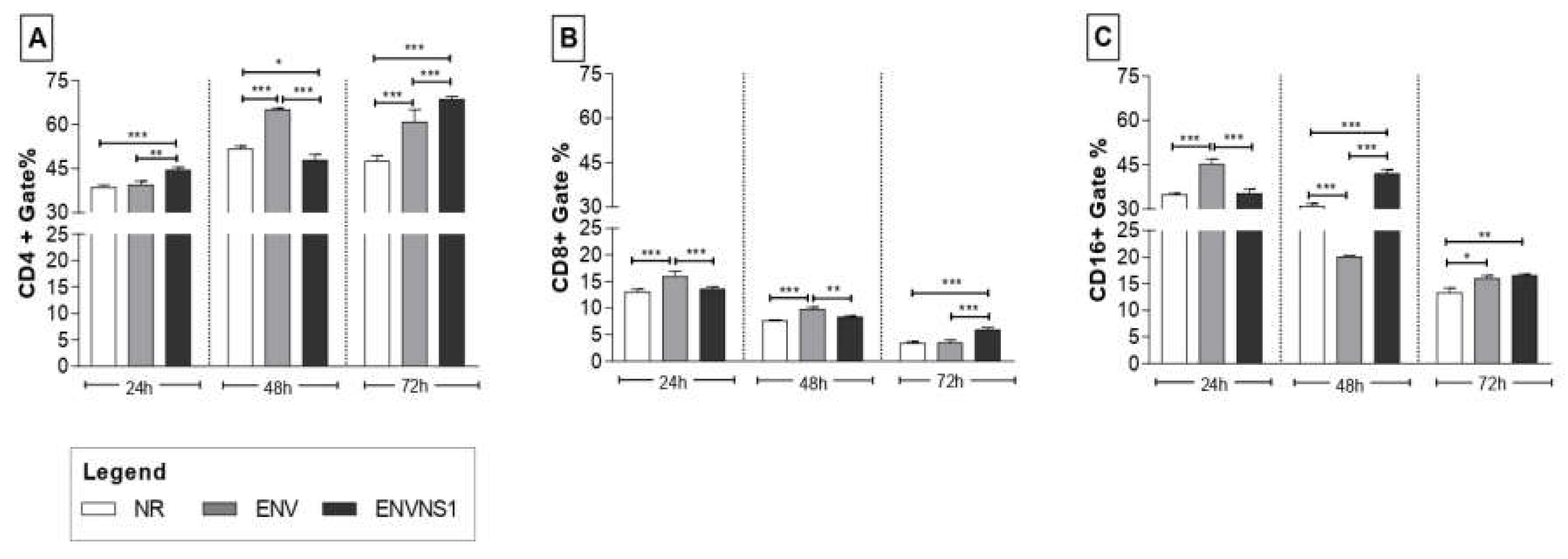

Table 1.
Hematological and biochemical parameters of vaccinated mice. Values correspond to mean ± standard deviation.
Table 1.
Hematological and biochemical parameters of vaccinated mice. Values correspond to mean ± standard deviation.
| Analysis | NR | ENV | ENVNS1 | R.V. [29,30,31] |
|---|---|---|---|---|
| Hematological | ||||
| Red blood cells (106/mm³) | 5.27 ± 0.35 | 5.32 ± 0.49 | 4.7 3± 0.41 | 7.3 ± 2.01 |
| Hemoglobin (g.dL-1) | 15.18 ± 1.10 | 16.04 ± 1.89 | 14.05 ± 1.31 | 13.82 ± 1.07 |
| Hematocrit (%) | 47.4 ± 3.36 | 48.2 ± 5.67 | 42.25 ± 3.86 | 38,44 ± 3,93 |
| MCV (fL) | 89.82 ± 1.27 | 90.35 ± 2.43 | 89.24 ± 0.62 | 60.26 ± 18.25 |
| MCHC (%) | 33.29 ± 0.06 | 33.27 ± 0.05 | 33.27 ± 0.08 | 33.00 ± 2.60 |
| Total leukocytes (10³/mm³) | 9.38 ± 1.14 | 9.16 ± 0.58 | 9.2 ± 0.42 | 6.23 ± 2.57 |
| Neutrophils (%) | 38.8 ± 5.72 | 41 ± 6.20 | 44.25 ± 2.93 | 22.96 ± 5.54 |
| Lymphocytes (%) | 56 ± 3.16 | 56.6 ± 6.07 | 53.5 ± 2.52 | 71.76 ± 5.9 |
| Eosinophils (%) | 1.6 ± 0.89 | 1.2 ± 0.45 | 1 ± 0 | 2.16 ± 1.71 |
| Monocytes (%) | 1.6 ± 0.89 | 1.2 ± 0.45 | 1.25 ± 0.5 | 2.68 ± 1 |
| Platelets (10³/mm³) | 386 ± 39.06 | 459.2 ± 11.73 | 434 ± 37.21 | 560 ± 119 |
| Biochemical tests | ||||
| Glucose (mg.dL-1) | 71.9 ± 4.34 | 85.84 ± 5.95 | 80.57 ± 13.86 | 80.75 ± 20.25 |
| AST (UI.L-1) | 133.52 ± 5.77 | 143.8 ± 0.08 | 103.02 ± 25.98 | 239.50 ± 141.20 |
| ALT (UI.L-1) | 154.4 ± 6.57 | 145.32 ± 5.11 | 145.67 ± 17.23 | 156.70 ± 57.20 |
| ALP (UI.L-1) | 215.82 ± 8.88 | 218.16 ± 4.52 | 202.6 ± 22.93 | 362.90 ± 226.60 |
RV- Reference value; MCV- Mean Corpuscular Volume; MCHC- Mean Corpuscular Hemoglobin Concentration; AST- Aspartate Aminotransferase; ALT- Alanine Aminotransferase; ALP- Alkaline phosphatase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



