
Submitted:
16 May 2023
Posted:
18 May 2023
You are already at the latest version
Abstract
Molybdenum (Mo) is vital for the activity of a small but essential group of enzymes called mo-lybdoenzymes. So far, specifically five molybdoenzymes have been discovered in eukaryotes: nitrate reductase, sulfite oxidase, xanthine dehydrogenase, aldehyde oxidase, and mARC. In order to become biologically active, Mo must be chelated to a pterin, forming the so-called Mo cofactor (Moco). Deficiency or mutation in any of the genes involved in Moco biosynthesis results in the simultaneous loss of activity of all molybdoenzymes, fully or partially preventing the normal development of the affected organism. To prevent this, the different mechanisms in-volved in Mo homeostasis must be finely regulated. Chlamydomonas reinhardtii is a unicellular, photosynthetic, eukaryotic microalga that has produced fundamental advances in key steps of Mo homeostasis over the last 30 years, which have been extrapolated to higher organisms, both plants and animals. These advances include the identification of the first two molybdate transporters in eukaryotes (MOT1 and MOT2), the characterization of key genes in Moco biosynthesis, the identification of the first enzyme that protects and transfers Moco (MCP1), the first characteriza-tion of mARC in plants, and the discovery of the crucial role of the nitrate reductase-mARC complex in plant nitric oxide production. This review aims to summarize all these advances made using C. reinhardtii as a model organism in Mo homeostasis and suggest how this microalga can continue to contribute to progress in this field.
Keywords:
Chlamydomonas
; Microalgae
; Molybdenum
; Homeostasis
; Moco
1. Chlamydomonas reinhardtii as a Model Microorganism
Microalgae are unicellular photosynthetic microorganisms that are considered one of the primary producers on our planet. Although microalgae are predominantly found in aquatic ecosystems, they can also thrive in various habitats, including soil as part of the rhizosphere [1]. Additionally, microalgae also can be found as part of animals, such as in coral reefs [2], and are a fundamental constituent of lichens [3]. In turn, microalgae have been discovered in polar regions [4]. Microalgae are responsible for a significant portion of total carbon fixation [5], making them fundamental to sustain different ecosystems [6]. However, they can also have adverse effects, such as algae blooms, which cause serious environmental, economic, and health damages [7]. The microalgae display a wide variety of cell sizes, morphologies, and architectures, and possess a diverse metabolic capacity that provides several distinctive features for scientific investigation [8]. The biotechnological use of microalgae has experienced exponential growth in recent years in various applications [9]. Notable examples of these applications include wastewater treatment, biofuel production, hydrogen production, production of high-value substances, and the use of microalgae as biofertilizers in agriculture [10]. It has been observed that the efficiency of some of these processes can be improved by the consortium of bacteria with microalgae [11].
The green microalgae C. reinhardtii (hereafter Chlamydomonas) was first isolated in a potato field in Massachusetts (USA) in 1945 [12], and since then it has become an important model organism [13] because it possesses numerous attributes that have contributed to its importance. Chlamydomonas exhibits a high growth rate, with a duplication time of approximately 8 hours. It also has easy and versatile growth in the laboratory and can prosper under mixotrophic, autotrophic, and heterotrophic conditions. This allows for the selection of mutants to study the mechanisms responsible for growth under different environmental conditions. Chlamydomonas shares basic metabolic characteristic with higher plants, and also with animals [14,15]. It has both an asexual life cycle as a unicellular haploid alga, as well as a sexual life cycle with the formation of diploid cells. This allows genetic crosses and transfer of genes/mutations to the resulting offspring. The Chlamydomonas genome has been sequenced [15], and there is a large collection of mutants, many of which have their mutation molecularly labeled [16]. In addition, a multitude of molecular tools have been developed and optimized for the study of Chlamydomonas, including techniques for transforming its three genomes (nuclear, chloroplastic, and mitochondrial). All of this has allowed for the development of a number of biochemical, metabolic, and physiological studies that have contributed to the understanding of the structure, function, and regulation of several biological processes [17]. Among others, the research topics that have used Chlamydomonas as a reference organism include the metabolism of nitrogen [18], sulfur [19], phosphorus [20], amino acid [21] and lipids [22], biosynthesis of carotenoids [23], starch [24], heme group [25], Fe-S cluster [26], chlorophyll [27] or glycerolipids [28], as well as the function of chaperones [29], proteases [30], thioredoxins [31], flagella [32], and the response to different types of stresses [33].
Chlamydomonas has also emerged as an organism with great biotechnological potential [34]. It is worth noting its use for the production of vaccines [35], antibodies [36], or in human clinical trials aimed to improve gastrointestinal health [37]. Furthermore, Chlamydomonas is also an excellent model for studying the basis of mutualistic interactions with bacteria, based on carbon-nitrogen exchange [38]. Certain mutualistic interactions have biotechnological potential applicable to bioremediation and H2 production [39,40]. The recent development of gene editing techniques such as CRISPR-Cas9 in Chlamydomonas could be a decisive step in increasing the biotechnological potential of this organism in addressing future challenges [41].
Regarding molybdenum (Mo) homeostasis, Chlamydomonas has been successfully used to uncover different steps of this process such as Mo uptake by identifying the first eukaryotic Mo transporters [42,43], Moco storage, protection and transfer by means of the identification and characterization of the Moco carrier protein (MCP1) [44,45]. Furthermore, Chlamydomonas has been used to obtain mutants affected in Mo metabolism [46], or to characterize Mo-using proteins like nitrate reductase [47] and mARC [48]. Therefore, an important part of the knowledge currently available about plant Mo homeostasis has been generated directly from Chlamydomonas, demonstrating its crucial role in this field of research over the last 30 years. In the present review we are focused on the advances in Mo homeostasis generated by using Chlamydomonas and how the knowledge acquired is resolving key aspects of this important process in other organisms.
2. The Role of Molybdenum in Living Organisms
The organisms require a sufficient cellular supply of Mo to ensure the activity of important enzymes such as nitrate reductase, sulfite oxidase, xanthine dehydrogenase, aldehyde oxidase, and mARC. Eukaryotic Mo homeostasis is a complex process that involves strategies for Mo uptake from the medium, Mo storage by either binding to other molecules in the cell or compartmentalizing into organelles, mechanisms for long-distance transport to ensure nutrient supply to the entire organism, and regulatory mechanisms to coordinate intracellular Mo content, environmental changes, and cellular responses to those changes. The different stages of Mo homeostasis, including its transport into the cell, the formation of active Mo cofactor, cellular accumulation, and the activity and regulation of different molybdoenzymes, will be discussed below, with special attention given to those aspects where the use of Chlamydomonas has produced significant advances in this field.
3. Molybdenum Uptake
With an average of 1.5 parts per million, Mo ranks as the 54th most abundant element in the Earth's crust, while in oceans, it ranks as the 25th most abundant element with an average of 10 parts per billion [49]. In the soil, the bioavailability of Mo depends on the abundance of the oxyanion molybdate (MoO42-), which is the predominant form of Mo at a pH greater than 4.2. All organisms take up Mo from the outside in the form of MoO42-. A direct relationship between Mo availability in soils and plant growth has been reported, showing a drastic decrease of growth under low Mo conditions [50]. High concentrations of molybdate also have drastic effects, for example, in Chlamydomonas the molybdate concentrations greater than 50 mM have been shown to be toxic to wild-type strains; however, Chlamydomonas mutants deficient in Mo transport are resistant to these high concentrations [51]. Therefore, a precise Mo homeostatic mechanism ensuring cellular Mo availability is required.
For quite some time it was unknown whether eukaryotes had a specific molybdate transporter or if molybdate entered non-specifically through sulfate or phosphate transporters, anions sharing structural similarity with molybdate [52]. However, experiments using different mutants of Chlamydomonas have shown that there should be at least two molybdate transporters, one mediating high affinity transport and the other one mediating low affinity transport [51] (Figure 1). The low-affinity molybdate transport activity was blocked by similar concentrations of sulfate, whereas the high-affinity transport activity was not blocked by sulfate. These transport activities were genetically linked to the loci NIT5 and NIT6 in Chlamydomonas. Although the identity of the NIT5 locus is still unknown, the NIT6 locus has been found to be the CNX1E gene [53]. As will be discussed later, CNX1E is the final enzyme involved in the biosynthesis of Moco. These results indicate that there is a close relationship between the protein involved in the last step of Moco biosynthesis (CNX1E) and the molybdate transporters, which could potentially be attributed to a physical connection between both proteins [54]. This hypothesis is reinforced by fact that CNX1E is associated with the cytoskeleton, favoring a localization close to the plasma membrane where MOT1 could be localized. However, this interesting possibility has not been proven so far and requires further experimentation in order to be confirmed. Molecularly, two families of eukaryotic molybdate transporters (MOT1 and MOT2) have been identified for the first time using Chlamydomonas. The MOT1 transporter family was mistakenly annotated in databases as belonging to the sulfate transporter SULTR family, due to the fact that they share some structural homology with them. However, this homology is low, and MOT1 transporters lack the fundamental STAS domain required for the ability to transport sulfate in the SULTR transporter family [42]. The MOT1 transporter family is characterized by having two conserved domains (FGXMPXCHG(S/A)GGLAXQ(Y/H)XFG(A/G)RXG and PXPVQPMKX(I/L)(A/G)AXA) that serve to molecularly label members belonging to this family. The Chlamydomonas MOT1 transporter shows high affinity for molybdate (Km 7 nM), is not blocked by sulfate, but is blocked by tungstate (structural analog of molybdate). Interestingly, the expression of MOT1 does not depend on the presence of molybdate in the medium but on that of nitrate, which increases its expression. This form of gene regulation may be due to the fact that the molybdoenzyme nitrate reductase is essential for growth on nitrate, and suggests that there must be a close relationship and coordination between Mo homeostasis and nitrate assimilation. This discovery in Chlamydomonas has enabled the identification of putative MOT1 member proteins in bacteria, fungi, and plants, but interestingly MOT1 seems to be not present in animal genomes including humans. In Arabidopsis thaliana, MOT1 is also a high-affinity transporter involved in transporting molybdate from the soil. Impaired plant growth occurs when MOT1 transporter is absent [55,56]. In the legume plants Lotus japonicus and Medicago truncatula, MOT1 proteins has been described to facilitate molybdate transport, whereas in L. japonicus the MOT1 reported member seems to be involved in Mo uptake by the roots [57], in M. truncatula both MOT1.2 and MOT1.3 are mediating Mo supply to the nodules during nitrogen fixation. M. truncatula mutants lacking these transporter show a deficient N-fixation activity although nitrogenase was present within the nodules [58,59]. Furthermore, it has been shown that polymorphisms in the A. thaliana MOT1.1 gene can be used as markers of leaf Mo content to detect phenotypic diversity in a plant population [60].
The MOT2 transporter family was also first molecularly identified in Chlamydomonas [43]. The members of this family do not have the typical conserved domain of MOT1 transporters and have low structural homology with MOT1. MOT2 shows lower affinity to transport molybdate (Km 550 nM) compared to MOT1 and is blocked by tungstate but not by sulfate. Interestingly, the expression of MOT2 gene does depend on the availability of Mo in the medium, being activated under conditions of low molybdate concentrations, but it does not respond to the presence of nitrate as nitrogen source, in other words, its regulation is opposite to that of MOT1. Homologs proteins of MOT2 are present in plants, animals, and humans. It has been shown that the human MOT2 is capable of transporting molybdate when expressed heterologously in yeast, suggesting a similar role in Mo homeostasis in mammals [43].
Apart from these two identified transporters, other transporters related to Mo homeostasis might be present in eukaryotes, mediating uptake, export or re-direction to organelles for storage. In this sense, the Chlamydomonas mutant db6 has been characterized as having a lower intracellular concentration of molybdate and reduced activity of the molybdoenzymes xanthine dehydrogenase and aldehyde oxidase. These effects can be restored by increasing the molybdate concentration in the medium to 10 mM. Additionally, the Chlamydomonas mutant db6 shows high resistance to elevated concentrations of tungstate in the medium [61]. These characteristics suggest that db6 is a mutant in the molybdate transport, although the molecular identity of the gene involved has not yet been resolved.
4. Moco Biosynthesis
As mentioned, in eukaryotic organisms, for Mo to be biologically active, it must complex with a pterin-derive molecule called MPT to form Moco. The different steps of this biosynthetic pathway will be shown below, focusing on those aspects where Chlamydomonas has made significant contributions. The proteins involved in the Moco biosynthetic pathway have a high degree of homology across all studied organisms. According to the different intermediates formed, this pathway can be divided into three main steps: cyclic pyranopterin monophosphate (cPMP) biosynthesis, MPT biosynthesis and Mo insertion [62]. In Chlamydomonas, 7 genes are essential for Moco biosynthesis, within these 3 steps. These genes follow the CNX (Cofactor of Nitrate reductase and Xanthine dehydrogenase) nomenclature, as occurs in plants and fungi [63].
4.1. cPMP Biosynthesis
In plants the first step of the Moco biosynthesis pathway is catalyzed in the mitochondria by the enzymes CNX2 and CNX3, and involves the conversion of a molecule of GTP into cyclic pyranopterin monophosphate (cPMP) [64] (Figure 1). In Chlamydomonas, individual mutants affected either in CNX2 or CNX3 genes have been identified. Characterization of such mutants indicates that they are unable to grow on nitrate and have elevated expression of the nitrate reductase gene due to a putative persistent nitrate positive signal [65]. Chlamydomonas CNX2 and CNX3 mutants are also resistant to high concentrations of mutagenic base analogs due to the involvement of the molybdoenzyme mARC in their detoxification [66]. Once cPMP is synthesized, it must be transported to the cytosol, which is carried out in Arabidopsis by the mitochondrial transporter ATM3 [67]. The ATM3 genes have not yet been characterized in Chlamydomonas. However, a mutant of the ATM3 gene (LMJ.RY0402.180694) is available in the molecularly tagged mutant library of Chlamydomonas [16].
4.2. MPT Biosynthesis
The second step consists of the insertion of two sulfur atoms into cPMP, which converts it to MPT. This reaction is catalyzed by MPT synthase, a tetrameric protein consisting of two large subunits (CNX6) and two small subunits (CNX7) [68] (Figure 1). To date, Chlamydomonas mutants have not been characterized in the CNX6 and CNX7 genes. However, in the library of molecularly labeled Chlamydomonas mutants [16], a mutant affected in the CNX6 gene (LMJ.RY0402.047904) is present, but not in the CNX7 gene, likely due to its small size (252 pb of predicted coding sequence). After the transfer of the sulfur atoms, MPT synthase must be resulfurated in a reaction that is mediated by MPT synthase sulfurase, which is encoded by the CNX5 gene [69]. A Chlamydomonas mutant affected in the CNX5 gene is also available [65]. Similarly to CNX2 and CNX3, Chlamydomonas CNX5 mutant is also resistant to high concentrations of mutagenic base analogs [66].
cPMP purified from Escherichia coli overexpressing strains have been used for the treatment of human Moco deficiency [70]. However, in 2021, a chemically synthesized form of cPMP, called 'Fosdenopterin', was granted FDA approval and received marketing authorization as a Nulibry product [71]. It can be assumed that the accumulation of cPMP could also occur in the Chlamydomonas CNX5 mutant. Therefore, it would be interesting to evaluate how this accumulation compares to that of E. coli or the chemically synthesized 'Fosdenopterin', to determine if the purification of cPMP from this Chlamydomonas mutant is more feasible from an economical point of view.
4.3. Mo Insertion
After MPT formation, a Mo atom coordinates to the sulfurs of MPT to form a biologically active Moco, a process that is mediated by adenylation [72,73] (Figure 1). In plants, the two-domain enzyme CNX1 catalyzes this insertion. The CNX1G domain of CNX1 catalyzes, in an Mg2+ and ATP-dependent reaction, the adenylation of MPT, forming transient MPT-AMP [74]. Then, the CNX1E domain of CNX1 catalyzes the deadenylation of MPT-AMP, a reaction that triggers the Mo insertion [75]. In Chlamydomonas, unlike most other eukaryotic organisms, the two domains CNX1G and CNX1E are encoded by two separate genes [46]. Therefore, in this aspect, Chlamydomonas resembles more to prokaryotic organisms such as E. coli where this two proteins are also separated and encoded in different genes [76]. In agreement with that, the Chlamydomonas CNX1G and CNX1E genes are able to complement mutations in the E. coli homologs mutants [46]. The mutations Nit4 and Nit6 in Chlamydomonas arise from a single alteration in CNX1E (V171A and G183D, respectively), which results in its loss of function. Additionally, it has been shown that the Chlamydomonas CNX1E gene exhibit intragenic complementation, suggesting that in Chlamydomonas CNX1E forms multimeric complexes [53]. The Moco of xanthine dehydrogenase and aldehyde oxidase family enzymes requires an additional step to become biologically active. This step involves the addition of an additional sulfur atom coordinated to Mo, a reaction catalyzed by the Moco sulfurase enzyme, encoded by the ABA3 gene [77] (Figure 1). In the other molybdoenzymes, instead of this sulfur atom, Mo is coordinated to the thiol group of a cysteine from the active center [78]. So far, Chlamydomonas mutant strains affected in ABA3 have not been reported. However, an ABA3 mutant (LMJ.RY0402.059375) seems to be also available in the labeled mutant library of Chlamydomonas [16].
5. Moco Storage
Active Moco is highly susceptible to inactivation in the presence of oxygen. Therefore, photosynthetic organisms should have some mechanism to protect this cofactor against oxidation [79]. The Chlamydomonas Moco Carrier Protein 1 (MCP1) was the first protein identified in eukaryotes to be involved in this process of Moco protection [80,81]. The main function of MCP1 seems to be the Moco protection from oxidation, but additionally this protein has also been reported to have a function in Moco storage and transference to the corresponding molybdoenzyme [45] (Figure 1). The three-dimensional structure of MCP1 has been resolved, being a tetramer with a typical Rossmann fold, and a putative Moco binding pocket has been proposed [44]. Proteins showing similar characteristics to Chlamydomonas MCP1 have also been identified and characterized in the cyanobacterium Rippkaea orientalis [82] and the microalga Volvox carteri [83]. Based on the three-dimensional structure of MCP1, orthologous proteins have also been identified in A. thaliana, which have been named MoBP (Molybdenum Binding Proteins) [84,85] however their function in Mo homeostasis still need to be clarified.
In prokaryotic organisms, proteins involved in Moco binding/storage have not been identified yet. However, in certain bacteria such as Azotobacter vinelandii, the existence of proteins that bind, accumulate, and buffer intracellular concentrations of molybdate, called molybdate binding proteins, has been described [86,87]. These Mo-binding proteins seem to be not present in eukaryotic genomes, and Mo storage may occur using different proteins or alternative strategies. In this sense it has been proposed that in plants like the legume Medicago sativa intracellular Mo excess might be chelated by organic acids such as malate or citrate [88].
6. Chlamydomonas Moco Enzymes
As previously mentioned, five molybdoenzymes have been identified in Chlamydomonas. In the next section, we will present the main advances made in the study of these enzymes, with special attention given to those in which the results obtained in Chlamydomonas has led to significant progress.
6.1. Xanthine Dehydrogenase
Plant xanthine dehydrogenase (XDH) catalyzes the oxidation of hypoxanthine to xanthine and xanthine to uric acid with NAD+ as the physiological electron acceptor [89]. Chlamydomonas is capable of using the purines xanthine and hypoxanthine as a nitrogen source [90], which indicates that it must have an active XDH enzyme to search for additional nitrogen sources when ammonia or nitrate are not present. In this regard, it has been observed that the Chlamydomonas XDH gene is strongly expressed under nitrogen-depleted conditions in the culture medium [91]. Chlamydomonas was the first microalga in which XDH was purified and characterized. Chlamydomonas XDH is a homodimeric enzyme with high molecular weight (330 kDa) that contains not only Moco but also Fe-S and FAD as cofactors [92].
6.2. Aldehyde Oxidase
Plant aldehyde oxidase (AO) catalyzes the oxidation of abscisic aldehyde to the phytohormone abscisic acid (ABA) [93]. AO and XDH share a great structural homology and have the same cofactor distribution. One reason for the high similarity in sequence and structure is that, in eukaryotes, the AO gene evolved from a duplication of the XDH gene [94]. So far, the AO protein has not been characterized in Chlamydomonas, but it is presumed to play a critical role in protecting against various types of stresses. In this regard, it has been observed that ABA protects the Chlamydomonas cells against the increase of reactive oxygen species that occur in response to salt stress [95], enhances the transport of bicarbonate and is responsible for regulating the diurnal cycle of upward and downward Chlamydomonas movement in response to gravity in the water column [96]. Chlamydomonas AO mutant (LMJ.RY0402.043929) is available in the labeled mutant library of Chlamydomonas [16], and its study could fundamentally contribute to elucidating the role of ABA in the response to different types of stresses.
6.3. Sulfite Oxidase
Plant sulfite oxidase (SO) enzyme functions as a homodimeric protein and plays a key role in sulfur catabolism by facilitating the two-electron oxidation of sulfite to sulfate using H2O as an electron acceptor and releasing hydrogen peroxide [97]. The SO seems to be protecting plant against the toxicity of sulfite in an environment with high levels of SO2 [98]. There are some differences between animal and plant SO. In vertebrates SO enzyme is localized in the mitochondrial intermembrane space and contains heme, in addition Moco, as cofactors [99]. In contrast, plant SO is localized in the peroxisome [100] and only contains Moco as a cofactor [101]. Interestingly, the Chlamydomonas enzyme resembles the SO of vertebrates more than that of plants, since it contains a heme group and has been found in the mitochondrial proteome [102]. Furthermore, Chlamydomonas SO is overexpressed in the presence of nitrate as nitrogen source. The reason behind this overexpression is not fully clear, but it could be related to the need of Chlamydomonas to regulate the balance of sulfur and nitrogen within its cellular metabolism. In the presence of nitrate, cells can use sulfur to produce other compounds necessary for their growth, such as cysteine, which may increase the need for SO to maintain the sulfur balance in the cell. Chlamydomonas SO mutant (LMJ.RY0402.221179) is available in the labeled mutant library of Chlamydomonas [16], although it has not yet been characterized.
6.4. Nitrate Reductase
The nitrate reductase (NR) enzyme is involved in two key processes: the critical step in nitrate assimilation, which is its reduction to nitrite, and the reduction of nitrite to nitric oxide (NO). The Chlamydomonas NR gene (NIA1) encodes a homodimeric protein. Each monomer has 3 prosthetic groups: FAD, heme b557, and Moco [103]. In NR, the flow of electrons goes from NAD(P)H to FAD, then to the heme b557, and finally ends in the Moco domain, where nitrate is reduced to nitrite [104]. Interestingly, the NR Moco domain is also able to use nitrite as a substrate, when it is present in high concentrations, reducing it to NO [105]. NO is an important and ubiquitous signaling molecule that is involved in many different biological processes, fundamental for cell survival under different types of stresses [106]. Two mechanisms exist for the production of NO, oxidative and reductive. The oxidative mechanism is catalyzed by the enzyme NO synthase (NOS) and uses L-arginine as a substrate [107]. However, in plants, the presence of NOS has only been determined in some microalgae such as Ostreococcus tauri [108], but not in Chlamydomonas. Therefore, in plants, the reduction of nitrite to NO through different mechanisms appears to be the most important way for NO synthesis [109]. Using Chlamydomonas it has been shown, for the first time in a photosynthetic organism, that NR can not only reduce nitrite to NO by itself but also with the assistance of another molybdoenzyme, the mARC protein [48]. This aspect will be explained bellow.
6.5. mARC
mARC has been the latest Moco enzyme discovered in eukaryotic organisms. It was firstly identified in pig liver mitochondria, and its name (mitochondrial Amidoxime Reducing Component) is derived from the first substrate characterized for this enzyme, amidoximes [110]. mARC is able to reduce the amidoxime N-hydroxylate pro-drug benzamidoxime to benzamidine, and therefore supposed to be involved in xenobiotic detoxification. However, since then, a wide variety of other mARC substrates have been identified. The first characterization of a mARC protein in a photosynthetic organism has been carried out in Chlamydomonas [66]. The mARC enzyme in Chlamydomonas has Moco showing a zinc-dependent activity. In agreement with that, proteomic studies have shown that in Chlamydomonas, zinc deficiency increases the cellular content of mARC by more than 30 times [111]. Moreover, the characterization of the Chlamydomonas mARC enzyme revealed that it belongs to the SO protein family, as it has a completely conserved cysteine within the active center that is used to chelate Moco [66]. In contrast to Human, where mARC appears to be a monomeric enzyme, Chlamydomonas mARC seems to form high molecular weight oligomeric complexes consisting of 10-12 monomers. These complexes could be involved in protecting the oxygen-labile Moco [112].
In addition to amidoximes, the human mARC is also involved in the reduction of sulfamethoxazole hydroxylamine to sulfamethoxazole [113], N4-hydroxy-L-arginine (NOHA) to arginine [114] an nitrite to NO [115]. It has been characterized that Chlamydomonas mARC can reduce the base analogue 6-hydroxylaminopurine (HAP) to adenine [66]. Therefore, it could be involved in the detoxification of mutagenic components. An interesting aspect about mARC proteins is that they can bind to different electron donors able to transfer electrons from NADH to reduce the corresponding substrates. Chlamydomonas mARC uses a cytochrome b5 and a cytochrome b5 reductase as electron donors for the HAP reduction [66].
Fascinatingly, it has been reported that Chlamydomonas mARC is also capable of reducing nitrite to the second messenger NO. However, for this action, it does not use cytochrome b5 as electron donor, but instead uses the molybdoenzyme NR [48]. In agreement with that, in Chlamydomonas, NR mutants overexpress mARC, and vice versa, and furthermore, Chlamydomonas mARC does not appear to be located in the mitochondria as in humans, but rather seems to be in the cytoplasm where its partner NR is located [48]. It was known that plant NR itself is capable of reducing nitrite to NO [116]; however, this ability is blocked in the presence of high millimolar concentrations of nitrate [117], which does not occur when NR acts as electron donor in Chlamydomonas to produce NO from nitrite reduction together with mARC [48]. Whereas C. reinhardtii has only one mARC enzyme, in the higher plant A. thaliana two mARC proteins (mARC1 and mARC2) have been identified, both of which are capable to reduce N-hydroxylated compounds, but only mARC2 seems to be involved in nitrite reduction to NO using NR as an electron donor [118]. These data suggest that the NR-mARC pair is a very efficient machinery for synthesizing NO under physiological conditions, in the presence of both nitrate and nitrite, and may have a very important role in modulating cellular NO levels. Apart from NO, Chlamydomonas complex NR-mARC has been shown to regulate the N2O production, a greenhouse gas, through the NO synthesis [119]. The mARC proteins have been proposed as moonlighting proteins [120]. Moonlighting are proteins able to perform more than one biological function within the cell, that is, they have the ability to perform different molecular tasks or participate in different cellular processes [121]. Therefore, the large number of different substrates and partners discovered for mARC support its classification as moonlighting protein.
7. Concluding Remarks and Future Outlook
In this review, we have shown how the green microalga C. reinhardtii has been used to identify various molecular components of Mo homeostasis, ranging from molybdate transport to molybdoenzyme activities. These advances have subsequently been applied to identify the corresponding molecular components in other model organisms. However, there are several issues that remain unresolved in which Chlamydomonas could also play a crucial role. For example, one of them is the mechanism mediating Mo export from the cell, being important to avoid Mo toxicity or for long-distance transport in multicellular organisms. Transporters mediating this process are still unknown. The isolation and characterization of Chlamydomonas strains resistant to high Mo concentrations could clarify this issue. Another point that remains unanswered is whether there are regulatory interconnections between the maintenance of intracellular concentration of Mo and the activity of Moco biosynthesis. In this regulation, some relationship between the molybdate transporters and the proteins that biosynthesize Moco may play a fundamental role. Therefore, it would be interesting to clarify whether there is a relationship between the molybdate transporters and the final protein involved in the biosynthesis of the Moco (CNX1E). Regarding molybdoenzymes, the role of the NR-mARC complex in the production of NO is well defined, but the role of the NO produced in the production of N2O has just begun to be studied. Surely in the future, very interesting results will be obtained studying this process in Chlamydomonas.
Author Contributions
AL conceptualization and original draft preparation; AL and M T-J wrote the paper; E L-M review and editing; All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by Gobierno de España, Ministerio de Ciencia e Innovacion (Grant PID2020-118398GB-I00), Junta de Andalucía (Grant ProyExcel_00483), the “Plan Propio” from University of Cordoba, and a grant awarded by the Torres-Gutierrez foundation.
Acknowledgments
This paper is dedicated to Emilio Fernandez Reyes for his more than 30 years of studying Mo homeostasis using Chlamydomonas reinhardtii as a reference organism. He was the driving force behind the research on Mo homeostasis in Chlamydomonas, the pillar that has allowed its advancement, and our great teacher whom we will never be able to repay for all the learnings received.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Quintas-Nunes, F.; Brandão, P.R.; Barreto Crespo, M.T.; Glick, B.R.; Nascimento, F.X. Plant Growth Promotion, Phytohormone Production and Genomics of the Rhizosphere-Associated Microalga, Micractinium Rhizosphaerae Sp. Nov. Plants 2023, 12, 651. [CrossRef]
- Maire, J.; Buerger, P.; Chan, W.Y.; Deore, P.; Dungan, A.M.; Nitschke, M.R.; van Oppen, M.J.H. Effects of Ocean Warming on the Underexplored Members of the Coral Microbiome. Integr. Comp. Biol. 2022, 62, 1700–1709. [CrossRef]
- Grimm, M.; Grube, M.; Schiefelbein, U.; Zühlke, D.; Bernhardt, J.; Riedel, K. The Lichens’ Microbiota, Still a Mystery? Front. Microbiol. 2021, 12, 623839. [CrossRef]
- Cvetkovska, M.; Vakulenko, G.; Smith, D.R.; Zhang, X.; Hüner, N.P.A. Temperature Stress in Psychrophilic Green Microalgae: Minireview. Physiol. Plant. 2022, 174, e13811.. [CrossRef]
- Falkowski, P.G. The Role of Phytoplankton Photosynthesis in Global Biogeochemical Cycles. Photosynth. Res. 1994, 39, 235–258. [CrossRef]
- Tarafdar, A.; Sowmya, G.; Yogeshwari, K.; Rattu, G.; Negi, T.; Awasthi, M.K.; Hoang, A.T.; Sirohi, R. Environmental Pollution Mitigation through Utilization of Carbon Dioxide by Microalgae. Environ. Pollut. 2023, 328, 121623. [CrossRef]
- Kazmi, S.S.U.H.; Yapa, N.; Karunarathna, S.C.; Suwannarach, N. Perceived Intensification in Harmful Algal Blooms Is a Wave of Cumulative Threat to the Aquatic Ecosystems. Biology (Basel). 2022, 11, 852. [CrossRef]
- Gerotto, C.; Norici, A.; Giordano, M. Toward Enhanced Fixation of CO2 in Aquatic Biomass: Focus on Microalgae. Front. Energy Res. 2020, 8, 213. [CrossRef]
- Parmar, P.; Kumar, R.; Neha, Y.; Srivatsan, V. Microalgae as next Generation Plant Growth Additives: Functions, Applications, Challenges and Circular Bioeconomy Based Solutions. Front. Plant Sci. 2023, 14, 1073546. [CrossRef]
- Fabris, M.; Abbriano, R.M.; Pernice, M.; Sutherland, D.L.; Commault, A.S.; Hall, C.C.; Labeeuw, L.; McCauley, J.I.; Kuzhiuparambil, U.; Ray, P.; et al. Emerging Technologies in Algal Biotechnology: Toward the Establishment of a Sustainable, Algae-Based Bioeconomy. Front. Plant Sci. 2020, 11, 279. [CrossRef]
- Zhang, B.; Li, W.; Guo, Y.; Zhang, Z.; Shi, W.; Cui, F.; Lens, P.N.L.; Tay, J.H. Microalgal-Bacterial Consortia: From Interspecies Interactions to Biotechnological Applications. Renew. Sustain. Energy Rev. 2020, 118, 109563. [CrossRef]
- Harris, E.H. Introduction into Chlamydomonas and Its Laboratory Use. Chlamydomonas Sourcebook. Oxford Acad. Press 2009.
- Salomé, P.A.; Merchant, S.S. A Series of Fortunate Events: Introducing Chlamydomonas as a Reference Organism. Plant Cell 2019, 31, 1682–1707. [CrossRef]
- Llamas, A.; Tejada-Jiménez, M.; Fernández, E.; Galván, A. Molybdenum Metabolism in the Alga Chlamydomonas Stands at the Crossroad of Those in Arabidopsis and Humans. Metallomics 2011, 3, 578–590. [CrossRef]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Maréchal-Drouard, L.; et al. The Chlamydomonas Genome Reveals the Evolution of Key Animal and Plant Functions. Science 2007, 318, 245–250. [CrossRef]
- Fauser, F.; Vilarrasa-Blasi, J.; Onishi, M.; Ramundo, S.; Patena, W.; Millican, M.; Osaki, J.; Philp, C.; Nemeth, M.; Salomamp, P.A.; et al. Systematic Characterization of Gene Function in the Photosynthetic Alga Chlamydomonas Reinhardtii. Nat. Genet. 2022, 54, 705–714. [CrossRef]
- Goodenough, U. The Chlamydomonas Sourcebook. Volume 1: Introduction to Chlamydomonas and Its Laboratory Use. Elsevier Acad. Press 2023. [CrossRef]
- Bellido-Pedraza, C.M.; Calatrava, V.; Sanz-Luque, E.; Tejada-Jiménez, M.; Llamas, A.; Plouviez, M.; Guieysse, B.; Fernández, E.; Galván, A. Chlamydomonas Reinhardtii, an Algal Model in the Nitrogen Cycle. Plants 2020, 9, 903. [CrossRef]
- Saroussi, S.; Sanz-Luque, E.; Kim, R.G.; Grossman, A.R. Nutrient Scavenging and Energy Management: Acclimation Responses in Nitrogen and Sulfur Deprived Chlamydomonas. Curr. Opin. Plant Biol. 2017, 39, 114–122. [CrossRef]
- Irihimovitch, V.; Yehudai-Resheff, S. Phosphate and Sulfur Limitation Responses in the Chloroplast of Chlamydomonas Reinhardtii. FEMS Microbiol. Lett. 2008, 283, 1–8. [CrossRef]
- Calatrava, V.; Hom, E.F.Y.; Llamas, Á.; Fernández, E.; Galván, A. Nitrogen Scavenging from Amino Acids and Peptides in the Model Alga Chlamydomonas Reinhardtii. The Role of Extracellular L-Amino Oxidase. Algal Res. 2019, 38, 101395. [CrossRef]
- Li-Beisson, Y.; Kong, F.; Wang, P.; Lee, Y.; Kang, B.-H. The Disassembly of Lipid Droplets in Chlamydomonas. New Phytol. 2021, 231, 1359–1364. [CrossRef]
- Rengel, R.; Giraldez, I.; Díaz, M.J.; García, T.; Vigara, J.; Leon, R. Simultaneous Production of Carotenoids and Chemical Building Blocks Precursors from Chlorophyta Microalgae. Bioresour. Technol. 2022, 351, 127035. [CrossRef]
- Sirohi, R.; Joun, J.; Choi, H.I.; Gaur, V.K.; Sim, S.J. Algal Glycobiotechnology: Omics Approaches for Strain Improvement. Microb. Cell Fact. 2021, 20, 163. [CrossRef]
- Calderon, R.H.; de Vitry, C.; Wollman, F.A.; Niyogi, K.K. Rubredoxin 1 Promotes the Proper Folding of D1 and Is Not Required for Heme B559 Assembly in Chlamydomonas Photosystem II. J. Biol. Chem. 2023, 299, 102968. [CrossRef]
- Przybyla-Toscano, J.; Couturier, J.; Remacle, C.; Rouhier, N. Occurrence, Evolution and Specificities of Iron-Sulfur Proteins and Maturation Factors in Chloroplasts from Algae. Int. J. Mol. Sci. 2021, 22, 3175. [CrossRef]
- Roach, T.; Baur, T.; Kranner, I. β-Cyclocitral Does Not Contribute to Singlet Oxygen-Signalling in Algae, but May Down-Regulate Chlorophyll Synthesis. Plants 2022, 11, 2155. [CrossRef]
- Yang, M.; Xie, X.; Kong, F.T.; Xie, K.P.; Yu, S.H.; Ma, J.Y.; Xue, S.; Gong, Z. Differences in Glycerolipid Response of Chlamydomonas Reinhardtii Starchless Mutant to High Light and Nitrogen Deprivation Stress Under Three Carbon Supply Regimes. Front. Plant Sci. 2022, 13, 860966. [CrossRef]
- Kreis, E.; Niemeyer, J.; Merz, M.; Scheuring, D.; Schroda, M. CLPB3 Is Required for the Removal of Chloroplast Protein Aggregates and Thermotolerance in Chlamydomonas. J. Exp. Bot. 2023, erad109. [CrossRef]
- Zou, Y.; Bozhkov, P. V. Chlamydomonas Proteases: Classification, Phylogeny, and Molecular Mechanisms. J. Exp. Bot. 2021, 72, 7680–7693. [CrossRef]
- Marchetti, G.M.; Füsser, F.; Singh, R.K.; Brummel, M.; Koch, O.; Kümmel, D.; Hippler, M. Structural Analysis Revealed a Novel Conformation of the NTRC Reductase Domain from Chlamydomonas Reinhardtii. J. Struct. Biol. 2022, 214, 107829. [CrossRef]
- Marshall, W.F. The Flagellar Length Control System: Exploring the Physical Biology of Organelle Size. Phys. Biol. 2023, 20, 021001. [CrossRef]
- Sasso, S.; Stibor, H.; Mittag, M.; Grossman, A.R. From Molecular Manipulation of Domesticated Chlamydomonas Reinhardtii to Survival in Nature. Elife 2018, 7, e39233. [CrossRef]
- Masi, A.; Leonelli, F.; Scognamiglio, V.; Gasperuzzo, G.; Antonacci, A.; Terzidis, chael A. Chlamydomonas Reinhardtii: A Factory of Nutraceutical and Food Supplements for Human Health. Molecules 2023, 28, 1185. [CrossRef]
- Dehghani, J.; Movafeghi, A.; Mathieu-Rivet, E.; Mati-Baouche, N.; Calbo, S.; Lerouge, P.; Bardor, M. Microalgae as an Efficient Vehicle for the Production and Targeted Delivery of Therapeutic Glycoproteins against SARS-CoV-2 Variants. Mar. Drugs 2022, 20, 657. [CrossRef]
- Rosales-Mendoza, S.; García-Silva, I.; González-Ortega, O.; Sandoval-Vargas, J.M.; Malla, A.; Vimolmangkang, S. The Potential of Algal Biotechnology to Produce Antiviral Compounds and Biopharmaceuticals. Molecules 2020, 25, 4049. [CrossRef]
- Fields, F.J.; Lejzerowicz, F.; Schroeder, D.; Ngoi, S.M.; Tran, M.; McDonald, D.; Jiang, L.; Chang, J.T.; Knight, R.; Mayfield, S. Effects of the Microalgae Chlamydomonas on Gastrointestinal Health. J. Funct. Foods 2020, 65, 103738. [CrossRef]
- Calatrava, V.; Tejada-Jimenez, M.; Sanz-Luque, E.; Fernandez, E.; Galvan, A.; Llamas, A. Chlamydomonas Reinhardtii, a Reference Organism to Study Algal–Microbial Interactions: Why Can’t They Be Friends? Plants 2023, 12, 788. [CrossRef]
- Fakhimi, N.; Gonzalez-Ballester, D.; Fernández, E.; Galván, A.; Dubini, A. Algae-Bacteria Consortia as a Strategy to Enhance H2 Production. Cells 2020, 9, 1353. [CrossRef]
- Torres, M.; Gonzalez-Ballester, D.; Gomez-Osuna, A.; Galván, A.; Fernandez, E.; Dubini, A. Chlamydomonas-Methylobacterium Oryzae Cooperation Leads to Increased Biomass, Nitrogen Removal, and Hydrogen Production. Bioresour. Technol. 2022, 352, 127088. [CrossRef]
- Kelterborn, S.; Boehning, F.; Sizova, I.; Baidukova, O.; Evers, H.; Hegemann, P. Gene Editing in Green Alga Chlamydomonas Reinhardtii via CRISPR-Cas9 Ribonucleoproteins. Plant Synth. Biol. Methods Mol. Biol. 2022, 2379, 45–65. [CrossRef]
- Tejada-Jiménez, M.; Llamas, A.; Sanz-Luque, E.; Galván, A.; Fernández, E. A High-Affinity Molybdate Transporter in Eukaryotes. Proc. Natl. Acad. Sci. USA 2007, 104, 20126–20130. [CrossRef]
- Tejada-Jiménez, M.; Galván, A.; Fernández, E. Algae and Humans Share a Molybdate Transporter. Proc. Natl. Acad. Sci. USA 2011, 108, 6420–6425. [CrossRef]
- Fischer, K.; Llamas, A.; Tejada-Jiménez, M.; Schrader, N.; Kuper, J.; Ataya, F.S.; Galván, A.; Mendel, R.R.; Fernández, E.; Schwarz, G. Function and Structure of the Molybdenum Cofactor Carrier Protein from Chlamydomonas Reinhardtii. J. Biol. Chem. 2006, 281, 30186–30194. [CrossRef]
- Ataya, F.S.; Witte, C.P.; Galván, A.; Igeño, M.I.; Fernández, E. Mcp1 Encodes the Molybdenum Cofactor Carrier Protein in Chlamydomonas Reinhardtii and Participates in Protection, Binding, and Storage Functions of the Cofactor. J. Biol. Chem. 2003, 278, 10885–10890. [CrossRef]
- Llamas, A.; Tejada-Jimenez, M.; González-Ballester, D.; Higuera, J.J.; Schwarz, G.; Galván, A.; Fernández, E. Chlamydomonas Reinhardtii CNX1E Reconstitutes Molybdenum Cofactor Biosynthesis in Escherichia Coli Mutants. Eukaryot. Cell 2007, 6, 1063–1067. [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galván, A.; Fernandez, E. Nitrate Reductase Regulates Plant Nitric Oxide Homeostasis. Trends Plant Sci. 2017, 22, 163–174. [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Ocaña-Calahorro, F.; Mariscal, V.; Carreras, A.; Barroso, J.B.; Galván, A.; Fernández, E. A Dual System Formed by the ARC and NR Molybdoenzymes Mediates Nitrite-Dependent NO Production in Chlamydomonas. Plant. Cell Environ. 2016, 39, 2097–2107. [CrossRef]
- Emsley, J. Nature’s Building Blocks. Oxford Univ. Press 2001.
- Lešková, A.; Javot, H.; Giehl, R.F.H. Metal Crossroads in Plants: Modulation of Nutrient Acquisition and Root Development by Essential Trace Metals. J. Exp. Bot. 2022, 73, 1751–1765. [CrossRef]
- Llamas, A.; Kalakoutskii, K.L.; Fernández, E. Molybdenum Cofactor Amounts in Chlamydomonas Reinhardtii Depend on the Nit5 Gene Function Related to Molybdate Transport. Plant, Cell Environ. 2000, 23, 1247–1255. [CrossRef]
- Mendel, R.R.; Haensh, R. Molybdoenzymes and Molybdenum Cofactor in Plants. J. Exp. Bot. 2002, 53, 1689–1698. [CrossRef]
- Chamizo-Ampudia, A.; Galvan, A.; Fernandez, E.; Llamas, A. Characterization of Chlamydomonas 102 and 104 Mutants Reveals Intermolecular Complementation in the Molybdenum Cofactor Protein CNX1E. Protist 2013, 164, 116–128. [CrossRef]
- Schwarz, G.; Schulze, J.; Bittner, F.; Eilers, T.; Kuper, J.; Bollmann, G.; Nerlich, A.; Brinkmann, H.; Mendel, R.R. The Molybdenum Cofactor Biosynthetic Protein Cnx1 Complements Molybdate-Repairable Mutants, Transfers Molybdenum to the Metal Binding Pterin, and Is Associated with the Cytoskeleton. Plant Cell 2000, 12, 2455–2471. [CrossRef]
- Tomatsu, H.; Takano, J.; Takahashi, H.; Watanabe-Takahashi, A.; Shibagaki, N.; Fujiwara, T. An Arabidopsis Thaliana High-Affinity Molybdate Transporter Required for Efficient Uptake of Molybdate from Soil. Proc. Natl. Acad. Sci. 2007, 104, 18807–18812. [CrossRef]
- Ide, Y.; Kusano, M.; Oikawa, A.; Fukushima, A.; Tomatsu, H.; Saito, K.; Hirai, M.Y.; Fujiwara, T. Effects of Molybdenum Deficiency and Defects in Molybdate Transporter MOT1 on Transcript Accumulation and Nitrogen/Sulphur Metabolism in Arabidopsis Thaliana. J. Exp. Bot. 2011, 62, 1483–1497. [CrossRef]
- Gao, J.-S.; Wu, F.-F.; Shen, Z.-L.; Meng, Y.; Cai, Y.-P.; Lin, Y. A Putative Molybdate Transporter LjMOT1 Is Required for Molybdenum Transport in Lotus Japonicus. Physiol. Plant. 2016, 158, 331–340. [CrossRef]
- Tejada-Jiménez, M.; Gil-Díez, P.; León-Mediavilla, J.; Wen, J.; Mysore, K.S.; Imperial, J.; González-Guerrero, M. Medicago Truncatula Molybdate Transporter Type 1 (MtMOT1.3) Is a Plasma Membrane Molybdenum Transporter Required for Nitrogenase Activity in Root Nodules under Molybdenum Deficiency. New Phytol. 2017, 216, 1223–1235. [CrossRef]
- Gil-Díez, P.; Tejada-Jiménez, M.; León-Mediavilla, J.; Wen, J.; Mysore, K.S.; Imperial, J.; González-Guerrero, M. MtMOT1.2 Is Responsible for Molybdate Supply to Medicago Truncatula Nodules. Plant. Cell Environ. 2019, 42, 310–320. [CrossRef]
- Forsberg, S.K.G.; Andreatta, M.E.; Huang, X.Y.; Danku, J.; Salt, D.E.; Carlborg, Ö. The Multi-Allelic Genetic Architecture of a Variance-Heterogeneity Locus for Molybdenum Concentration in Leaves Acts as a Source of Unexplained Additive Genetic Variance. PLoS Genet. 2015, 11, 1005648. [CrossRef]
- Li, W.; Fingrut, D.R.; Maxwell, D.P. Characterization of a Mutant of Chlamydomonas Reinhardtii Deficient in the Molybdenum Cofactor. Physiol. Plant. 2009, 136, 336–350. [CrossRef]
- Tejada-Jiménez, M.; Chamizo-Ampudia, A.; Calatrava, V.; Galván, A.; Fernández, E.; Llamas, A. From the Eukaryotic Molybdenum Cofactor Biosynthesis to the Moonlighting Enzyme MARC. Molecules 2018, 23, 3287. [CrossRef]
- Schwarz, G.; Mendel, R.R. Molybdenum Cofactor Biosynthesis and Molybdenum Enzymes. Annu. Rev. Plant Biol. 2006, 57, 623–647. [CrossRef]
- Kruse, I.; Maclean, A.E.; Hill, L.; Balk, J. Genetic Dissection of Cyclic Pyranopterin Monophosphate Biosynthesis in Plant Mitochondria. Biochem J 2018, 475, 495–509. [CrossRef]
- González-Ballester, D.; de Montaigu, A.; Higuera, J.J.; Galván, A.; Fernández, E. Functional Genomics of the Regulation of the Nitrate Assimilation Pathway in Chlamydomonas. Plant Physiol. 2005, 137, 522–533. [CrossRef]
- Chamizo-Ampudia, A.; Galvan, A.; Fernandez, E.; Llamas, A. The Chlamydomonas Reinhardtii Molybdenum Cofactor Enzyme CrARC Has a Zn-Dependent Activity and Protein Partners Similar to Those of Its Human Homologue. Eukaryot. Cell 2011, 10, 1270–1282. [CrossRef]
- Teschner, J.; Lachmann, N.; Schulze, J.; Geisler, M.; Selbach, K.; Santamaria-Araujo, J.; Balk, J.; Mendel, R.R.; Bittner, F. A Novel Role for Arabidopsis Mitochondrial ABC Transporter ATM3 in Molybdenum Cofactor Biosynthesis. Plant Cell 2010, 22, 468–480. [CrossRef]
- Kaufholdt, D.; Baillie, C.K.; Meinen, R.; Mendel, R.R.; Hänsch, R. The Molybdenum Cofactor Biosynthesis Network: In Vivo Protein-Protein Interactions of an Actin Associated Multi-Protein Complex. Front. Plant Sci. 2017, 8, 1946. [CrossRef]
- Nakai, Y.; Harada, A.; Hashiguchi, Y.; Nakai, M.; Hayashi, H. Arabidopsis Molybdopterin Biosynthesis Protein Cnx5 Collaborates with the Ubiquitin-like Protein Urm11 in the Thio-Modification of TRNA. J Biol Chem 2012, 287, 30874–30884. [CrossRef]
- Veldman, A.; Santamaria-Araujo, J.A.; Sollazzo, S.; Pitt, J.; Gianello, R.; Yaplito-Lee, J.; Wong, F.; Ramsden, C.A.; Reiss, J.; Cook, I.; et al. Successful Treatment of Molybdenum Cofactor Deficiency Type A with CPMP. Pediatrics 2010, 125, e1249-54. [CrossRef]
- Farrell, S.; Karp, J.; Hager, R.; Wang, Y.; Adeniyi, O.; Wang, J.; Li, L.; Ma, L.; Peretz, J.; Summan, M.; et al. Regulatory News: Nulibry (Fosdenopterin) Approved to Reduce the Risk of Mortality in Patients with Molybdenum Cofactor Deficiency Type A: FDA Approval Summary. J. Inherit. Metab. Dis. 2021, 44, 1085–1087. [CrossRef]
- Kuper, J.; Llamas, A.; Hecht, H.-J.; Mendel, R.R.; Schwarz, G. Structure of the Molybdopterin-Bound Cnx1G Domain Links Molybdenum and Copper Metabolism. Nature 2004, 430, 803–806. [CrossRef]
- Kruse, T. Function of Molybdenum Insertases. Molecules 2022, 27, 5372. [CrossRef]
- Llamas, A.; Mendel, R.R.; Schwarz, G. Synthesis of Adenylated Molybdopterin: An Essential Step for Molybdenum Insertion. J. Biol. Chem. 2004, 279, 55241–55246. [CrossRef]
- Llamas, A.; Otte, T.; Multhaup, G.; Mendel, R.R.; Schwarz, G. The Mechanism of Nucleotide-Assisted Molybdenum Insertion into Molybdopterin. J. Biol. Chem. 2006, 281, 18343–18350. [CrossRef]
- Nichols, J.D.; Rajagopalan, K. V. In Vitro Molybdenum Ligation to Molybdopterin Using Purified Components. J. Biol. Chem. 2005, 280, 7817–7822. [CrossRef]
- Selles, B.; Moseler, A.; Caubrière, D.; Sun, S.K.; Ziesel, M.; Dhalleine, T.; Hériché, M.; Wirtz, M.; Rouhier, N.; Couturier, J. The Cytosolic Arabidopsis Thaliana Cysteine Desulfurase ABA3 Delivers Sulfur to the Sulfurtransferase STR18. J. Biol. Chem. 2022, 298, 101749. [CrossRef]
- Mendel, R.R. The History of the Molybdenum Cofactor-A Personal View. Molecules 2022, 27, 4934. [CrossRef]
- Aguilar, M.; Cárdenas, J.; Fernández, E. Quantitation of Molybdopterin Oxidation Product in Wild-Type and Molybdenum Cofactor Deficient Mutants of Chlamydomonas Reinhardtii. Biochim. Biophys. Acta 1992, 1160, 269–274. [CrossRef]
- Witte, C.-P.; Igeño, M.I.; Mendel, R.; Schwarz, G.; Fernández, E. The Chlamydomonas Reinhardtii MoCo Carrier Protein Is Multimeric and Stabilizes Molybdopterin Cofactor in a Molybdate Charged Form. FEBS Lett. 1998, 431, 205–209. [CrossRef]
- Aguilar, M.; Kalakoutskii, K.; Cárdenas, J.; Fernández, E. Direct Transfer of Molybdopterin Cofactor to Aponitrate Reductase from a Carrier Protein in Chlamydomonas Reinhardtii. FEBS Lett 1992, 307, 162–163. [CrossRef]
- Krausze, J.; Hercher, T.W.; Archna, A.; Kruse, T. The Structure of the Moco Carrier Protein from Rippkaea Orientalis. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2020, 76, 453–463. [CrossRef]
- Hercher, T.W.; Krausze, J.; Yang, J.; Kirk, M.L.; Kruse, T. Identification and Characterization of the Volvox Carteri Moco Carrier Protein. Biosci. Rep. 2020, 40, BSR20202351. [CrossRef]
- Kruse, T.; Gehl, C.; Geisler, M.; Lehrke, M.; Ringel, P.; Hallier, S.; Hänsch, R.; Mendel, R.R. Identification and Biochemical Characterization of Molybdenum Cofactor-Binding Proteins from Arabidopsis Thaliana. J. Biol. Chem. 2010, 285, 6623–6635. [CrossRef]
- Kruse, T. Moco Carrier and Binding Proteins. Molecules 2022, 27, 6571. [CrossRef]
- Lawson, D.M.; Williams, C.E.; White, D.J.; Choay, A.P.; Mitchenall, L.A.; Pau, R.N. Protein Ligands for Molybdate. Specificity and Charge Stabilisation at the Anion-Binding Sites of Periplasmic and Intracellular Molybdate-Binding Proteins of Azotobacter Vinelandii. J. Chem. Soc. Dalt. Trans. 1997, 3981–3984. [CrossRef]
- Fenske, D.; Gnida, M.; Schneider, K.; Meyer-Klaucke, W.; Schemberg, J.; Henschel, V.; Meyer, A.-K.; Knöchel, A.; Müller, A. A New Type of Metalloprotein: The Mo Storage Protein from Azotobacter Vinelandii Contains a Polynuclear Molybdenum-Oxide Cluster. ChemBioChem 2005, 6, 405–413. [CrossRef]
- Steinke, D.R.; Majak, W.; Sorensen, T.S.; Parvez, M. Chelation of Molybdenum in Medicago Sativa (Alfalfa) Grown on Reclaimed Mine Tailings. J. Agric. Food Chem. 2008, 56, 5437–5442. [CrossRef]
- Yesbergenova, Z.; Yang, G.; Oron, E.; Soffer, D.; Fluhr, R.; Sagi, M. The Plant Mo-Hydroxylases Aldehyde Oxidase and Xanthine Dehydrogenase Have Distinct Reactive Oxygen Species Signatures and Are Induced by Drought and Abscisic Acid. Plant J 2005, 42, 862–876. [CrossRef]
- Pineda, M.; Cardenas, J. Transport and Assimilation of Purines in Chlamydomonas Reinhardtii. Sci. Mar. 1996, 60, 195–201.
- Schmollinger, S.; Mühlhaus, T.; Boyle, N.R.; Blaby, I.K.; Casero, D.; Mettler, T.; Moseley, J.L.; Kropat, J.; Sommer, F.; Strenkert, D.; et al. Nitrogen-Sparing Mechanisms in Chlamydomonas Affect the Transcriptome, the Proteome, and Photosynthetic Metabolism. Plant Cell 2014, 26, 1410–1435. [CrossRef]
- Perez-Vicente, R.; Pineda, M.; Cardenas, J. Isolation and Characterization of Xanthine Dehydrogenase from Chlamydomonas Reinhardtii. Physiol. Plant. 1988, 72, 101–107. [CrossRef]
- Seo, M.; Koiwai, H.; Akaba, S.; Komano, T.; Oritani, T.; Kamiya, Y.; Koshiba, T. Abscisic Aldehyde Oxidase in Leaves of Arabidopsis Thaliana. Plant J 2000, 23, 481–488. [CrossRef]
- Rodríguez-Trelles, F.; Tarrío, R.; Ayala, F.J. Convergent Neofunctionalization by Positive Darwinian Selection after Ancient Recurrent Duplications of the Xanthine Dehydrogenase Gene. Proc Natl Acad Sci USA 2003, 100, 13413–13417. [CrossRef]
- Abu-Ghosh, S.; Iluz, D.; Dubinsky, Z.; Miller, G. Exogenous Abscisic Acid Confers Salinity Tolerance in Chlamydomonas Reinhardtii During Its Life Cycle. J. Phycol 2021, 57, 1323–1334. [CrossRef]
- Al-Hijab, L.; Gregg, A.; Davies, R.; Macdonald, H.; Ladomery, M.; Wilson, I. Abscisic Acid Induced a Negative Geotropic Response in Dark-Incubated Chlamydomonas Reinhardtii. Sci. Rep. 2019, 9, 12063. [CrossRef]
- Feng, C.; Tollin, G.; Enemark, J.H. Sulfite Oxidizing Enzymes. Biochim. Biophys. Acta - Proteins Proteomics 2007, 1774, 527–539. [CrossRef]
- Randewig, D.; Hamisch, D.; Herschbach, C.; Eiblmeier, M.; Gehl, C.; Jurgeleit, J.; Skerra, J.; Mendel, R.R.; Rennenberg, H.; Hänsch, R. Sulfite Oxidase Controls Sulfur Metabolism under SO2 Exposure in Arabidopsis Thaliana. Plant. Cell Environ. 2012, 35, 100–115. [CrossRef]
- Mellis, A.T.; Roeper, J.; Misko, A.L.; Kohl, J.; Schwarz, G. Sulfite Alters the Mitochondrial Network in Molybdenum Cofactor Deficiency. Front. Genet. 2021, 11, 594828. [CrossRef]
- Nowak, K.; Luniak, N.; Witt, C.; Wüstefeld, Y.; Wachter, A.; Mendel, R.R.; Hänsch, R. Peroxisomal Localization of Sulfite Oxidase Separates It from Chloroplast-Based Sulfur Assimilation. Plant Cell Physiol 2004, 45, 1889–1894. [CrossRef]
- Eilers, T.; Schwarz, G.; Brinkmann, H.; Witt, C.; Richter, T.; Nieder, J.; Koch, B.; Hille, R.; Hänsch, R.; Mendel, R.R. Identification and Biochemical Characterization of Arabidopsis Thaliana Sulfite Oxidase: A New Player in Plant Sulfur Metabolism. J. Biol. Chem. 2001, 276, 46989–46994. [CrossRef]
- Gerin, S.; Mathy, G.; Blomme, A.; Franck, F.; Sluse, F.E. Plasticity of the Mitoproteome to Nitrogen Sources (Nitrate and Ammonium) in Chlamydomonas Reinhardtii: The Logic of Aox1 Gene Localization. Biochim. Biophys. Acta - Bioenerg. 2010, 1797, 994–1003. [CrossRef]
- Fernandez, E.; Schnell, R.; Ranum, L.P.; Hussey, S.C.; Silflow, C.D.; Lefebvre, P.A. Isolation and Characterization of the Nitrate Reductase Structural Gene of Chlamydomonas Reinhardtii. Proc. Natl. Acad. Sci. 1989, 86, 6449–6453. [CrossRef]
- Vidal, E.A.; Alvarez, J.M.; Araus, V.; Riveras, E.; Brooks, M.D.; Krouk, G.; Ruffel, S.; Lejay, L.; Crawford, N.M.; Coruzzi, G.M.; et al. Nitrate in 2020: Thirty Years from Transport to Signaling Networks. Plant Cell 2020, 32, 2094–2119. [CrossRef]
- Dean, J. V; Harper, J.E. The Conversion of Nitrite to Nitrogen Oxide(s) by the Constitutive NAD(P)H-Nitrate Reductase Enzyme from Soybean. Plant Physiol 1988, 88, 389–395. [CrossRef]
- Khan, M.; Ali, S.; Al Azzawi, T.N.I.; Yun, B.W. Nitric Oxide Acts as a Key Signaling Molecule in Plant Development under Stressful Conditions. Int. J. Mol. Sci. 2023, 24, 4782. [CrossRef]
- Gupta, K.J.; Kaladhar, V.C.; Fitzpatrick, T.B.; Fernie, A.R.; Møller, I.M.; Loake, G.J. Nitric Oxide Regulation of Plant Metabolism. Mol. Plant 2022, 15, 228–242. [CrossRef]
- Foresi, N.; Correa-Aragunde, N.; Parisi, G.; Caló, G.; Salerno, G.; Lamattina, L. Characterization of a Nitric Oxide Synthase from the Plant Kingdom: NO Generation from the Green Alga Ostreococcus Tauri Is Light Irradiance and Growth Phase Dependent. Plant Cell 2010, 22, 3816–3830. [CrossRef]
- Santolini, J.; André, F.; Jeandroz, S.; Wendehenne, D. Nitric Oxide Synthase in Plants: Where Do We Stand? Nitric Oxide 2017, 63, 30–38. [CrossRef]
- Havemeyer, A.; Bittner, F.; Wollers, S.; Mendel, R.; Kunze, T.; Clement, B. Identification of the Missing Component in the Mitochondrial Benzamidoxime Prodrug-Converting System as a Novel Molybdenum Enzyme. J Biol Chem 2006, 281, 34796–34802. [CrossRef]
- Hsieh, S.I.; Castruita, M.; Malasarn, D.; Urzica, E.; Erde, J.; Page, M.D.; Yamasaki, H.; Casero, D.; Pellegrini, M.; Merchant, S.S.; et al. The Proteome of Copper, Iron, Zinc, and Manganese Micronutrient Deficiency in Chlamydomonas Reinhardtii. Mol Cell Proteomics 2013, 12, 65–86. [CrossRef]
- Chamizo-Ampudia, A.; Galvan, A.; Fernandez, E.; Llamas, A. Study of Different Variants of Mo Enzyme CrARC and the Interaction with Its Partners CrCytb5-R and CrCytb5-1. Int. J. Mol. Sci. 2017, 18, 670. [CrossRef]
- Ott, G.; Plitzko, B.; Krischkowski, C.; Reichmann, D.; Bittner, F.; Mendel, R.R.; Kunze, T.; Clement, B.; Havemeyer, A. Reduction of Sulfamethoxazole Hydroxylamine (SMX-HA) by the Mitochondrial Amidoxime Reducing Component (MARC). Chem Res Toxicol 2014, 27, 1687–1695. [CrossRef]
- Kotthaus, J.; Wahl, B.; Havemeyer, A.; Schade, D.; Garbe-Schönberg, D.; Mendel, R.; Bittner, F.; Clement, B. Reduction of N(ω)-Hydroxy-L-Arginine by the Mitochondrial Amidoxime Reducing Component (MARC). Biochem J 2011, 433, 383–391. [CrossRef]
- Sparacino-Watkins, C.E.; Tejero, J.; Sun, B.; Gauthier, M.C.; Thomas, J.; Ragireddy, V.; Merchant, B.A.; Wang, J.; Azarov, I.; Basu, P.; et al. Nitrite Reductase and Nitric-Oxide Synthase Activity of the Mitochondrial Molybdopterin Enzymes MARC1 and MARC2. J. Biol. Chem. 2014, 289, 10345–10358. [CrossRef]
- Bender, D.; Schwarz, G. Nitrite-Dependent Nitric Oxide Synthesis by Molybdenum Enzymes. FEBS Lett 2018, 592, 2126–2139. [CrossRef]
- Rockel, P.; Strube, F.; Rockel, A.; Wildt, J.; Kaiser, W.M. Regulation of Nitric Oxide (NO) Production by Plant Nitrate Reductase in Vivo and in Vitro. J. Exp. Bot. 2002, 53, 103–110. [CrossRef]
- Maiber, L.; Koprivova, A.; Bender, D.; Kopriva, S.; Fischer-Schrader, K. Characterization of the Amidoxime Reducing Components ARC1 and ARC2 from Arabidopsis Thaliana. FEBS J. 2022, 289, 5656–5669. [CrossRef]
- Bellido-Pedraza, C.M.; Calatrava, V.; Llamas, A.; Fernandez, E.; Sanz-Luque, E.; Galvan, A. Nitrous Oxide Emissions from Nitrite Are Highly Dependent on Nitrate Reductase in the Microalga Chlamydomonas Reinhardtii. Int. J. Mol. Sci. 2022, 23, 9412. [CrossRef]
- Llamas, A.; Chamizo-Ampudia, A.; Tejada-Jimenez, M.; Galvan, A.; Fernandez, E. The Molybdenum Cofactor Enzyme MARC: Moonlighting or Promiscuous Enzyme? BioFactors 2017, 43, 486–494. [CrossRef]
- Gupta, M.N.; Uversky, V.N. Moonlighting Enzymes: When Cellular Context Defines Specificity. Cell. Mol. Life Sci. 2023, 80, 130. [CrossRef]
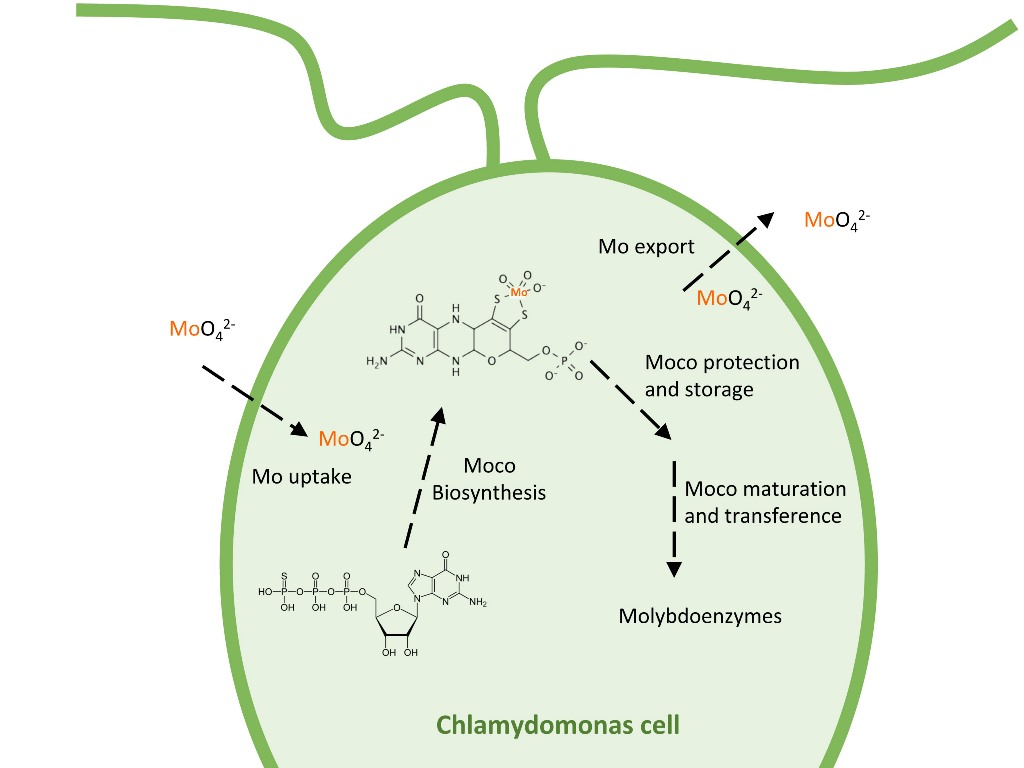
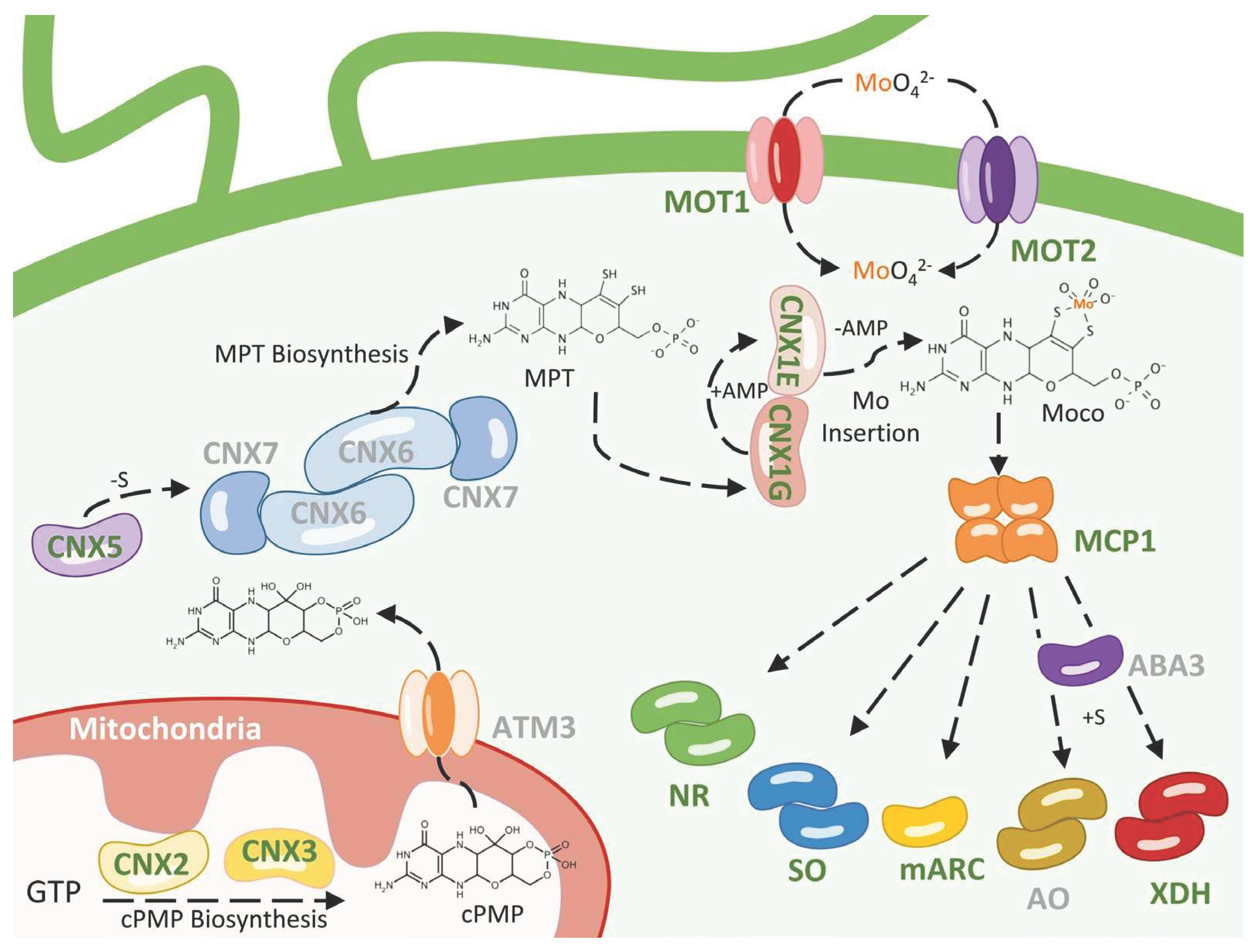
Figure 1.
Molybdenum homeostasis in Chlamydomonas. The biosynthetic machinery, maturation, and distribution of Moco in a Chlamydomonas cell are shown. The basic steps of Moco biosynthesis, from GTP to Moco, including enzymes that contain Moco, are depicted. Proteins catalyzing individual steps are shown in different colors. The names of proteins that have been notably studied in Chlamydomonas are shown in green, and the others are shown in gray. All known intermediates of the pathway are presented sequentially in the three steps in which Moco is synthesized. The MOT1 and MOT2 proteins transport molybdate-anion to the cytosol. The CNX2 and CNX3 proteins catalyze the conversion of GTP to cPMP. The mitochondrial ABC transporter ATM3 is involved in transporting cPMP from the mitochondria to the cytosol. MPT-synthase, consisting of CNX6 and CNX7, converts cPMP to MPT, which is then sulfurated by CNX5. CNX1G activates MPT by converting it to MPT-AMP, which is then transferred to CNX1E. CNX1E deadenylates MPT-AMP and incorporates Mo into MPT to produce Moco. ABA3 catalyzes the addition of a sulfur atom coordinated to Mo to the XDH and AO families of molybdoenzymes. Moco can bind to the Moco carrier protein MCP1, where it is accumulated or transferred to form the different Mo enzymes nitrate reductase (NR), sulfite oxidase (SO), xanthine dehydrogenase (XDH), aldehyde oxidase (AO), and mARC. AMP (adenosine monophosphate), for a more detailed explanation, refer to the text.
Figure 1.
Molybdenum homeostasis in Chlamydomonas. The biosynthetic machinery, maturation, and distribution of Moco in a Chlamydomonas cell are shown. The basic steps of Moco biosynthesis, from GTP to Moco, including enzymes that contain Moco, are depicted. Proteins catalyzing individual steps are shown in different colors. The names of proteins that have been notably studied in Chlamydomonas are shown in green, and the others are shown in gray. All known intermediates of the pathway are presented sequentially in the three steps in which Moco is synthesized. The MOT1 and MOT2 proteins transport molybdate-anion to the cytosol. The CNX2 and CNX3 proteins catalyze the conversion of GTP to cPMP. The mitochondrial ABC transporter ATM3 is involved in transporting cPMP from the mitochondria to the cytosol. MPT-synthase, consisting of CNX6 and CNX7, converts cPMP to MPT, which is then sulfurated by CNX5. CNX1G activates MPT by converting it to MPT-AMP, which is then transferred to CNX1E. CNX1E deadenylates MPT-AMP and incorporates Mo into MPT to produce Moco. ABA3 catalyzes the addition of a sulfur atom coordinated to Mo to the XDH and AO families of molybdoenzymes. Moco can bind to the Moco carrier protein MCP1, where it is accumulated or transferred to form the different Mo enzymes nitrate reductase (NR), sulfite oxidase (SO), xanthine dehydrogenase (XDH), aldehyde oxidase (AO), and mARC. AMP (adenosine monophosphate), for a more detailed explanation, refer to the text.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



