
Submitted:
18 May 2023
Posted:
18 May 2023
You are already at the latest version
Abstract
Considering tumors from an evolucionary perspective, successive tumor cells undergo turnover in response to stressful conditions which pressures tumor cells to adapt to a new environment. Tumor cells uses parts of themselves to communicate and share biological information, known as extracellular vesicles (EV) secretion to promote their survival and “survive to each and every metabolic cost”. Tumor microenvironment (TME) is a miscellaneous of cells, factors, extracellular matrix proteins and EVs. EVs include smalls (<160nm, exosomes) and larger (100-1,000nm, microvesicles) lipid bilayer enclosed packages of biomolecules that are shed by tumors to TME. Tumor-derived extracellular vesicles transfer adaptative stress signaling to recipient cells, reprograming these cells. Heat Shock Proteins (HSP) are more prominent stress response regulators found in exosomes. HSP-loaded exosomes actively reprogram tumor and TME cells to acquire mechanisms that contribute to tumor progression and therapy resistance. The intercellular communication mediated by HSP-loaded EVs favors the escape of tumor cells from, endoplasmic reticulum (ER) stress, hypoxia, apoptosis and anti-cancer therapies. Extracellular HSPs activate and deactivate immune response, induce cell differentiation, changes vascular homeostasis, and help to augment the pre-metastatic niche formation. Here we will explore the EV mechanism of HSP transmission among TME cells and the relevance of these intercellular communications for resistance to therapies in cancer.
Keywords:
Heat-shock-protein
; extracellular vesicles
; reprograming tumors
; tumor evolucionary pressure
1. Introduction
Cancer cell survival is challenged by internal and external stress factors, which exert selective pressure on cancer cells, driving the emergence of genetic and phenotypic diversity that allows cell survival. Harsh tumor microenvironment (TME) conditions increase intracellular misfolded and damaged proteins, thus in an evolutionary perspective, cancer adaptability involves proteotoxic stress adaptation. Proteostasis network preserve the proteome functionality by coordinating process such as protein synthesis and folding, translocation, or degradation, to promptly respond to stress conditions (Young, Barral et al. 2003, Kampinga, Mayer et al. 2019). However, severe, or persistent stress stimulus can disturb the protein-folding equilibrium and surpass the proteostasis “buffering” capacity, culminating in the induction of cell death. Heat shock proteins (HSPs) are a group of molecular chaperones that play crucial roles in cellular protection to stress and maintaining cellular homeostasis. Chaperones act as stress sensors and have a pivotal role as protectors of misfolding protein accumulation by facilitating protein folding, trafficking, assembly, and degradation (reviewed in (Amodio, Pagliara et al. 2021). The interface between environmental stress and protein homeostasis impacts the evolution and selection of more adaptive phenotypes. In a model of Drosophila cell lines, it was evidenced that the direct folding of mutated proteins provided new functions that eventually conferred selective advantages to cells to survive in hostile environments (Lindquist 2009). Here, we exploit the idea that increased chaperone expression and activity in cancer cells (Ferrarini, Heltai et al. 1992, Jäättelä 1995, Vargas-Roig, Gago et al. 1998) can be an evolutionary advantage to respond environmental stress signals that result in cancer progression and resistance to therapy.
Cancer cell survivability and proliferation require continuous communication among tumor and TME cells. The systemic release of extracellular vesicles (EVs) has been reported as a particular mechanism of cancer cells to propagate stress tolerance to other cells. EVs are small lipid bilayer-delimited particles derived from cells with sizes ranging from 100 to 1,000 nm. Exosomes (<160 nm) are the minor subtype of EVs, originating from the endosomal pathways, and the “giant” EVs microvesicles (100-1,000nm, MV) are originated from membrane blebbing. EVs can act as intercellular communication mediators, contributing to many cells biological processes, by carrying different biomolecules, such as proteins, lipids, nucleic acids, and metabolites. The release of EVs from cancer cells as an adaptative stress response to harsh conditions has been well documented. We have previously shown that acquisition of resistance to treatment by melanoma cells could be mediated by EVs derived from treated tumor cells (Andrade, Otake et al. 2019). EVs are shed by tumors as unique forms of process or reshape of cell content in a way that they can share with other cells information received by a particular population of cells. Samwise, cancer cells can receive foreign information from EVs that signalize them to survive, grow, migrate, or even die.
Considering the primary function of HSPs to maintain cellular homeostasis, it is important to explore the potential EV-loaded-HSP roles in activating the proteostasis network within the TME with prominent modulators to undergo internal proteotoxic stress (Dai, Dai et al. 2012). Although the concept of EVs preconditioning mechanism remains under construction, it is mainly attributed to EV’s internal cargo. Also, some knowledge needs to be explored: differently from external-uncovered-HSP, EVs can deliver HSPs to distinct cell types and at distant sites outside the body’s compartments in saliva or other fluids. In this review, we exploit the idea that chaperones-containing EVs are shed by cancer cells upon stress stimulus and focus our discussion on HSP-mediated stress response adaptation inside the TME that contributes to tumor progression.
2. Extracellular Vesicle (EV)-Mediated Stress Propagation during Tumor Development and Progression
Cells release EVs because of their physiology and pathophysiology. EVs can be classified into three main subtypes (Raposo and Stoorvogel 2013) according to their biogenesis and size. Apoptotic bodies are generated by membrane disintegration after injuries and cell death activation, producing diameter vesicles from 1 to 5µM. Microvesicles are particles generated by the direct outward budding of the plasma membrane of viable cells, having vesicles in a size range of around 50nm to 1,000nm in diameter. Finally, exosomes are generated by the endosomal sorting complex required for transport (ESCRT) system composed of different proteins able to interact with other proteins and promote the formation of intraluminal vesicles. Exosome formation starts after the invagination of the plasma membrane and formation of an early-sorting endosome (ESE). The ESE contains cell-surface and soluble proteins associated with the extracellular milieu in addition to content from the trans-Golgi network and endoplasmic reticulum. Late sorting endosomes (LSE) are matured ESEs that generate multivesicular bodies (MVB) containing intraluminal vesicles. MVBs form by double invagination of plasma membrane and can later fuse with lysosomes or autophagosomes to be degraded. Release of exosomes occurs after fusion of MVB to the plasma membrane being that exosomes size round 40 to 160nm in diameter (Théry, Zitvogel et al. 2002).
Studies have been conducted to investigate the role of EVs in the transfer of stress tolerance phenotype in different cancer models. Hypoxia leads to EV release and/or a higher cargo loading per vesicle and transfer a hypoxic tolerance phenotype to TME, as well as promote pro-tumoral effects, including induced proliferation, migration, angiogenesis and immunomodulation (Bister, Pistono et al. 2020). The main cellular hypoxia effect is protein-folding instability with damaged and misfolded proteins accumulation. Proteostasis instability affects the endoplasmic reticulum (ER), triggering a specific cellular state known as ER stress and the unfolded protein response (UPR) to restore the homeostasis. Mahadevan et al. demonstrated that ER stress can be transmitted from cancer cells to bone marrow-derived myeloid cells, a phenomenon known as transmissible ER stress (Mahadevan, Rodvold et al. 2011). This communication was confirmed to be organized by EVs, which was firstly attributed to cancer cell soluble factors (Jiang, Zhang et al. 2020) and compelling evidence from cancer cells submitted to ER stress inducers demonstrate that this insult increases EV secretion (Kanemoto, Nitani et al. 2016, Collett, Redman et al. 2018).
EV-mediated remote stress preconditioning have identified several EV cargos, including mRNA, microRNA and proteins that impact in HIF-1α-, UPR-, angiogenesis- and autophagy signaling into recipient cells (reviewed in (Bister, Pistono et al. 2020)). Given the critical role of HSPs in driving stress response, its activity is one of the main cellular pro-survival mechanisms and it would be logical to expect the presence of these proteins in EVs. Six major mammalian HSP families have been classified according to their molecular size: HSP60, HSP70, HSP90, HSP100, HSP40s and small heat shock proteins (sHSPs). EVs containing diverse HSP members are passively or actively released by damaged, stressed, or dead cells. The expression of HSP90 in exosomes derived from diverse normal cells was previously reported (Théry, Regnault et al. 1999, Géminard, Nault et al. 2001, van Niel, Raposo et al. 2001, Wubbolts, Leckie et al. 2003). Later, B cell-derived exosomes were reported to have increased levels of chaperones under heat stress condition (Clayton, Turkes et al. 2005). Oral squamous cell carcinoma secretes HSP90-enriched EVs and promoted expression of HSP90, TRAP1 and HSP105, that were correlated with poor prognosis in head and neck carcinoma patients (Ono, Eguchi et al. 2018). A mitochondrial chaperonin, HSP60, is secreted into exosomes as a regular process independently of cell death induction (Merendino, Bucchieri et al. 2010). A mitochondrial chaperone, GRP75/mt-HSP70, is involved in EVs secretion by breast cancer cells and its blockage decreases tumoral EVs secretion (Huang, Wu et al. 2019). The release of EV-HSP70 can also be enhanced immediately in plasma after cardio-exercising which follows in returning to the baseline quantitate amount of HSP in EV extracted from patients’ serum (Nederveen, Warnier et al. 2021).
3. Heat Shock Proteins (HSPs) Secretion by EVs Triggered by Chemo or Radiotherapy
Several reports have underlined that stress induced by anticancer therapies, as chemotherapy and radiotherapy, induce EVs secretion from TME cells, resulting in drug resistance transfer to recipient cells (reviewed in (O’Neill, Gilligan et al. 2019)). Importantly, high levels of tumoral HSPs have been reported associated with poor prognosis and resistance to therapy (reviewed in (Ciocca and Calderwood 2005)). Some authors revealed that diverse HSPs, including HSP27, HSP60, HSP70 and HSP90, have a cytoprotective activity in reducing the sensitive of tumor cells to anticancer drugs (Garrido, Schmitt et al. 2003, Gabai, Budagova et al. 2005, Dempsey, Ireland et al. 2010, Heinrich, Donakonda et al. 2016, Boudesco, Cause et al. 2018, Kumar, Siripini et al. 2020). It has become apparent that HSPs are released and are able to induce cellular responses in the extracellular milieu, including therapy resistance (Santos, Martins et al. 2017).
For many years, findings of extracellular HSPs were considered artefacts caused by cell necrosis, due to absence of a peptide secretion signal in their sequence (De Maio and Vazquez 2013). However, this concept was revised showing HSPs to be released even in the absence of cell death (Hunter-Lavin, Davies et al. 2004) and further studies described diverse unconventional pathways of HSP secretion, including inside EVs (Santos, Martins et al. 2017). Importantly, HSPs are present in exosomes released by various cancer cells (Table 1). The exact mechanism by which HSPs are incorporated within EVs is still controversial. However, the promotion of malignant features by HSP-EVs and drug resistance are extensively reported (Gastpar, Gehrmann et al. 2005, McCready, Sims et al. 2010, Peinado, Alečković et al. 2012, Tamura, Torigoe et al. 2012, Hsu, Hung et al. 2017, Ono, Eguchi et al. 2018, Taha, Ono et al. 2019).
4. Anticancer Therapies Induce HSP-EVs’ Release by Tumor Cells
Besides the advances in targeted therapies, some patients relapse even after an initial positive response to a therapy schedule and become unresponsive after a few cycles. Therapy resistance is documented to all cancer and therapy types. Up to date, the focus of drug resistance research has been on identifying genetic and epigenetic changes in cancer cells and/or cells from TME. They usually aim for prosurvival signaling, apoptotic pathways inhibition and controlled drug alteration. It was recently revealed that EVs also alter cancer cell plasticity to modify them to become chemotherapeutic-resistant (Kreger, Johansen et al. 2016, Namee and O’Driscoll 2018, Maacha, Bhat et al. 2019). EVs mediate drug resistance by reducing the concentration through exporting it from cancer cells or by dividing their cargos among TME cells (Shedden, Xie et al. 2003). These altered pathways evolve cells to diminish drug efficacy (Maacha, Bhat et al. 2019). The role of intercellular transfer of HSPs mediated by EVs in the horizontal transmission of drug resistance in multiple cancer types is explored here.
Chemo and radiotherapy, stimulates secretion of EVs by tumor cells with pro-survival and pro-metastatic capacity (Dai, Wan et al. 2005, Hurwitz, Kaur et al. 2010, Arscott, Tandle et al. 2013, Al-Mayah, Bright et al. 2015, König, Kasimir-Bauer et al. 2017, Bandari, Purushothaman et al. 2018, Emam, Ando et al. 2018, Keklikoglou, Cianciaruso et al. 2019). Lv et al. showed that paclitaxel, irinotecan, and carboplatin promote the release of HSP-bearing exosomes from HepG2 hepatocellular carcinoma cells (Lv, Wan et al. 2012). Campanella C et al. observed a decrease of HSP60 intracellular levels and an increase of nitrated HSP60 exportation via exossomes in human lung-derived carcinoma cell line (H292) after treatment with the histone deacetylase inhibitor, suberoylanilide hydroxamic acid (SAHA) (Campanella, D’Anneo et al. 2016). Similarly, evidence shows that radiotherapy also promotes the release of exosomes containing HSP72 from PC-3 and DU-145 prostate cancer cells (Hurwitz, Kaur et al. 2010). Shao et al showed that glioblastomas cells treated with temozolamide (TMZ) secreted more microvesicles with HSP90 as cargo. In a combined TMZ plus an HSP90 inhibitor (geldanalmycin) treatment, there was more GBM apoptosis and diminished vesiculation (Shao, Chung et al. 2012). Li-Hong et al showed that chemotherapeutic treatment of hepatocarcinoma cells, HepG2 and PLC/PRF/5 increase secretion of exosomes expressing HSP60, HSP70 and HSP90 on their membranes. Interestingly, HepG2 cell-derived exosomes secretion and the highest HSP levels in exosomes were observed in response to chemotherapy that HepG2 cells exhibit resistance (Lv, Wan et al. 2012).
These findings indicate that increased HSP expression on EVs may confer advantages to cancer cells to resist and survive to anticancer therapies. Notably, a comparative proteomic analysis of EVs derived from breast cancer patients that relapsed or not, showed a differential expression of HSP70 amongst both groups (Vinik, Ortega et al. 2020). Likewise, Rothammer et al observed higher HSP70 serum levels in breast cancer patients that exhibited contralateral recurrence or metastases after radiotherapy treatment (Rothammer, Sage et al. 2019). This is of particular interest, as tumor recurrence results from therapy resistance and suggests the involvement of HSPs in this phenomenon.
5. Cancer Cell Intrinsic Mechanism Modulated by HSP-EVs that Impact Therapy Response
In addition to the transfer of HSP cargo from EVs to cancer cells, HSP present in the external surface of EVs can interact with surface receptors of target cancer cells and contribute for resistance phenotype propagation. McCready J et al. described that invasive cancer cells secrete HSP90α-enrich exosomes and also identified the pro-migratory protein plasminogen as a potential client protein of these extracellular chaperones(McCready, Sims et al. 2010). Tsen F et al. revealed another mechanism by which HSP90α can modulate cancer cell migration. In this study, they have provided evidence that extracellular HSP90α binds to the subdomain II of the extracellular part of low-density lipoprotein receptor-related protein 1 (LRP-1) which signals to Akt kinases, Akt1 and Akt2, to promote cell motility(Tsen, Bhatia et al. 2013). Similarly, Ono K et al. have shown that HSP90-enriched EVs derived from metastatic oral cancer cells initiate epithelial-mesenchymal transition (EMT) in normal epithelial cells and promoted migration and invasion of tumor cells. Moreover, these EV-driven migratory events were reversed by HSP90 depletion(Ono, Sogawa et al. 2020). Tang X et al. also demonstrated that breast cancer derived-exosomes present eHSP90α on their external surface and stimulates migration of both normal stromal cells and tumor cells in a paracrine and autocrine mechanisms(Tang, Chang et al. 2019).
HSP-EVs can also mediate the communication of cancer cells with other stromal cells, such as endothelial cells and promote angiogenesis. Yukawa H et al investigated the influence of exosomes secreted from hepatocellular carcinoma cells on angiogenesis and found that HepG2-derived exosomes expressing HSP70 are incorporated by HUVECs cells and induce lumen formation(Yukawa, Suzuki et al. 2018). Noticeably, several reports have suggested a key role of HSP90 in regulating tumor angiogenesis, as multiple arms of angiogenic signaling have been described clients of this chaperone(Bohonowych, Gopal et al. 2010). Feng Q et al. reported that HSP90 is directly associated with a unique 90kDa form of VEGF (VEGF90K) on microvesicles (MVs) isolated from MDAMB231 and SKBR3 breast cancer cells. Interestingly, this association results in a sustained activation of VEGFRs and a consequent resistance to Bevacizumab. However, HSP90 inhibitors disrupt this client-protein interaction and the release of VEGF90K from the MVs restore Bevacizumab sensitivity(Feng, Zhang et al. 2017).
Taken together, all these reports clearly demonstrate that HSP-EVs can contribute for tumor heterogeneity response to anticancer therapies by inducing EMT, migration and angiogenesis. It is interesting to stress out that HSP-EVs can also propagate cancer drug resistance by interacting with and modulating crucial components of the immune response.
6. Immunological Roles of Cancer HSP-EVs That Impact Therapy Response
Many studies have been conducted to reveal the immunological consequences of tumoral HSP-EVs.All processes related to the immune system are tissue-context dependent in wich the response is occurring. In TME and in presence of EV eriched or carrying HSP’s, this idea is no different. Therefore, vesicles conatining HSP can have an anti-tumor or pro-tumor role (Figure 1). Chalmin F et al. and Diao J et al. provided mechanistic insight linking tumor-derived exosomes (TDE) and tumor-induced immunosuppression. HSP70 or HSP72 present on TDEs binds to toll-like receptor 2 (TLR2) on myeloid-derived suppressor cells (MDSC) and trigger Stat3 activation, promoting the suppressor activity of MDSC(Diao, Yang et al. 2015). Chalmin F et al. also observed that dimethyl amiloride reduced exosome secretion and Stat3 phosphorylation in MDSCs resulting in an enhanced cytotoxic effect on T cells under cyclophosphamide treatment. In line with this, Gobbo et. al evaluated the blockage of HSP70 and TLR2 association by the peptide aptamer A8, which targets the extracellular domain of HSP70 on exosomes. They observed that this peptide impaired MDSCs activity induced by cisplatin and 5-fluorouracil treatment and potentiated the antitumor effect of these chemotherapeutic drugs(Gobbo, Marcion et al. 2016). Additionally, Ono K et al. showed that HSP90-enriched-EV derived from metastatic oral cancer cells are uptaked by macrophages resulting in M2 polarization(Ono, Sogawa et al. 2020).
On the contrary, there is also evidence that HSP-EVs can modulate anti-tumor immunity. Gastpar et al. demonstrated that high-HSP70/Bag-4 surface-positive exosomes act as natural killer (NK) cells attractants and elicits a strong NK lytic capacity to HSP70 membrane-positive tumors (Gastpar, Gehrmann et al. 2005). Elsner et al. also found that HSP70-positive exosomes derived from human melanoma cells induce the activation of mouse NK cells and result in tumor growth and metastasis reduction (Elsner, Muppala et al. 2007). Additionally, the encounter of myeloma-HSP-expressing exosomes and dendritic cells efficiently stimulate their maturation to promote a T helper 1 (Th1) and cytotoxic T lymphocyte (CTL) anti-tumoral responses (Xie, Bai et al. 2010). Similarly, HSP70-enriched exosomes derived from a tumor heat-treatment promote tumor regression in murine models mediated by a Th1 immune response (Cho, Lee et al. 2009). Menay F et al showed the presence of HSP70 in the lumen and HSP90 on the surface of exosomes isolated from mice bearing a very aggressive T-cell lymphoma. The immunogenic properties of these HSP-exosomes were found to induce Th1 response in naïve-syngeneic mice, resulting in protection against secondary challenges (Menay, Herschlik et al. 2017). Sen K et al. demonstrated that the exposure of naïve murine macrophages is activated by HSP70-rich exosomes released from murine breast carcinoma cell lines post hyperthermia treatment. Moreover other anti-tumoral responses were observed, such as increased macrophage migration and release of TNF-α and RANTES, which triggered a cytotoxic response against breast cancer cells (Sen, Sheppe et al. 2020). Vega VL et al also showed that exosomes-enriched in HSP70 activate macrophages to increased TNF-α production (Vega, Rodríguez-Silva et al. 2008).
Hurwitz M D et al. showed that prostate cancer cell line PC-3 and DU-145 secretes HSP72-exosomes after irradiation treatment. These exosomes also promote increase in pro-inflammatory cytokines IL-6 and TNF-α and expression of CD8+ T and NK cells(Hurwitz, Kaur et al. 2010). HSP-EVs play a pivotal role in stimulating an antitumor immune response after anticancer therapies. LH et al have shown that hepatocarcinoma HepG2 cells secrete HSP-rich exosomes in response to paclitaxel, irinotecan and carboplatin. These secreted EVs elicit an NK cell mediated antitumor response after granzymes B production. Exosome treatment in NK cells decreased the expression of inhibitory receptor CD94 and increased the expression of activating receptors CD69, NKG2D, and NKp44(Lv, Wan, et al. 2012).
All these reports demonstrate that same HSP-EVs can sometimes act as a danger signal, increasing tumor immunogenicity and inducing an active response. On the other hand, HSP-expressing EVs can induce immunosuppression and compromise anticancer therapy efficacy. Furthermore, there is growing evidence that HSP inside or at the membrane of EVs contributes to tumor progression and resistance to therapy. However, once the timeline and order of these events are understood, physicians can use it to manage the patient’s treatment better and improve their follow-ups (Table 2).
7. Future Perspectives
Research to date strongly supports that HSPs produced upon stress promote resistance by protecting cells against apoptotic cell death (Takayama, Reed et al. 2003). For example, Cesa et al identified that inhibitors of apoptosis proteins (IAPs) are specific client substrates of HSP70 (Cesa, Shao et al. 2018). Considering that overexpression of IAPs have been associated with resistance to chemo- and radiotherapy(Hunter, LaCasse et al. 2007), it is highly likely that the delivery of HSP70 by EVs could inhibit the turnover of IAPs in target cells, favoring their accumulation and resulting in tumoral apoptosis resistance. We speculate that anticancer therapy stress increases intracellular HSPs expression that are secreted by EVs, mainly exosomes, and promote an anti-apoptotic cytoprotective phenotype in the target cancer cells conferring protection against a second therapy stressful stimulus, favoring tumor repopulation (Figure 2).
A major player in therapy resistance and tumor repopulation are cancer stem cells (CSC). One of distinguishing characteristics of CSCs are their high tolerance to oxidative stress, hypoxia and nutritional shortage (Prager, Xie et al. 2019), and it has been shown that HSP70 and HSP90 are involved in the development and maintenance of the CSC phenotype, as well as the cytoprotective machinery that allows these cells to survive stress conditions (Kabakov, Yakimova et al. 2020, Lettini, Lepore et al. 2020). These chaperones are constantly secreted by CSCs and have been widely reported as involved in cancer stemness-associated events, such as EMT, angiogenesis, treatment resistance, tumor immunosuppression and metastasis (Kabakov, Yakimova et al. 2020). Importantly, CSCs release EVs that perform a variety of biological roles in tumors, including transferring stem-like features to non-CSCs and mediating cell-cell communication in the TME (Nakano, Garnier et al. 2015, Sun, Wang et al. 2018). The ability of CSCs to release EVs that carry specific proteins and transcription factors to surrounding cells has a stronger impact on tumor heterogeneity (Al-Sowayan, Al-Shareeda et al. 2020). Consequently, the investigation of EVs transporting key molecular chaperones involved in establishing and sustaining the CSC phenotype may become very attractive. To our knowledge, the only EV-chaperones secreted by CSCs reported up to date are exosomal HSP90 and HSP70, both found in prostate cancer. Hypoxia-stressed prostate cancer cells secrete exosomes rich in HSP90 and HSP70 (Ramteke, Ting et al. 2015), which seems to play a role in the establishment of the CSC phenotype. Prostate cancer cell-organoids with CSC-like properties secret abundant amounts of HSP90 and EPCAM-containing exosomes, as well as exhibit multiple stemness markers expression (Eguchi, Sogawa et al. 2018). Furthermore, extracellular HSP90 (eHSP90) has been linked to overexpression of a cohort of stemness-associated markers and the EMT marker Snail in prostate CSCs. Additionally, eHSP90 has been implicated in boosting self-renewal, tumoroids formation and treatment resistance associated with metastatic propensity (Nolan, Kaur et al. 2017). Further research looking for HSPs in CSC-EVs and investigating the mechanisms by which they contribute to the maintenance of CSCs is needed and their modulation may represent an important weapon in the elimination of these hard-to-treat cells.
8. Concluding Remarks
The interplay between EV-mediated communication and HSP cargo has profound implications for tumor biology and therapeutic strategies. Thus, interfering in HSP-EVs has emerged as a new potential target therapy. However, interfering in HSP signaling is challenging due to the overlap between HSP family members and since they can vary widely depending on the disease context. While evidence suggests that targeting EV-HSPs may be a promising strategy for cancer therapy, it is unlikely that analyzing HSPs inside EVs alone would be a reliable method for predicting bad or good therapy response for different types of cancer. It is crucial to consider other primordial factors that can influence therapy response such as tumor stage, mutation status, history of disease, age of patient, overall health status, etc. Although HSP inhibitors could eventually lead to improved cancer treatment outcomes for some patients, to anticipate drug resistance it is crucial to better understand the crosstalk between HSP networks and other molecular factors in the TME to influence treatment response.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, MCR. and AHO.; methodology, AL.; software, AL and CMLM.; validation, CMLM., RFS. and AHO.; formal analysis, RFS.; investigation, RFS.; resources, MCR.; data curation, RFS.; writing—original draft preparation, RFS.; writing—review and editing, CMLM, MCR and AHO. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by São Paulo Research Foundation (FAPESP) for funding our work with the grants 2018/08107-2, 2019/07723-4 and 2020/15445-1.
Acknowledgments
We thank the São Paulo Research Foundation (FAPESP) for funding our work with the grants 2018/08107-2, 2019/07723-4 and 2020/15445-1.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Al-Mayah, A.; Bright, S.; Chapman, K.; Irons, S.; Luo, P.; Carter, D.; Goodwin, E.; Kadhim, M. The non-targeted effects of radiation are perpetuated by exosomes. Mutat. Res. Mol. Mech. Mutagen. 2015, 772, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Al-Sowayan, B.S.; Al-Shareeda, A.T.; Alrfaei, B.M. Cancer Stem Cell-Exosomes, Unexposed Player in Tumorigenicity. Front. Pharmacol. 2020, 11, 384. [Google Scholar] [CrossRef] [PubMed]
- Amodio, G.; Pagliara, V.; Moltedo, O.; Remondelli, P. Structural and Functional Significance of the Endoplasmic Reticulum Unfolded Protein Response Transducers and Chaperones at the Mitochondria–ER Contacts: A Cancer Perspective. Front. Cell Dev. Biol. 2021, 9, 641194. [Google Scholar] [CrossRef] [PubMed]
- Arscott, W.T.; Tandle, A.T.; Zhao, S.; Shabason, J.E.; Gordon, I.K.; Schlaff, C.D.; Zhang, G.; Tofilon, P.J.; Camphausen, K.A. Ionizing Radiation and Glioblastoma Exosomes: Implications in Tumor Biology and Cell Migration. Transl. Oncol. 2013, 6, 638–648. [Google Scholar] [CrossRef]
- Bandari, S.K.; Purushothaman, A.; Ramani, V.C.; Brinkley, G.J.; Chandrashekar, D.S.; Varambally, S.; Mobley, J.A.; Zhang, Y.; Brown, E.E.; Vlodavsky, I.; et al. Chemotherapy induces secretion of exosomes loaded with heparanase that degrades extracellular matrix and impacts tumor and host cell behavior. Matrix Biol. 2018, 65, 104–118. [Google Scholar] [CrossRef]
- Bausero, M.A.; Gastpar, R.; Multhoff, G.; Asea, A. Alternative mechanism by which IFN-gamma enhances tumor recognition: Active release of heat shock protein 72. J Immunol 2005, 175, 2900–2912. [Google Scholar]
- Bister; Pistono, C.; Huremagic, B.; Jolkkonen, J.; Giugno, R.; Malm, T. Hypoxia and extracellular vesicles: A review on methods, vesicular cargo and functions. J. Extracell. Vesicles 2020, 10, e12002. [Google Scholar] [CrossRef]
- Bohonowych, J.E.; Gopal, U.; Isaacs, J.S. Hsp90 as a Gatekeeper of Tumor Angiogenesis: Clinical Promise and Potential Pitfalls. J. Oncol. 2010, 2010, 412985. [Google Scholar] [CrossRef]
- Boudesco; Cause, S.; Jego, G.; Garrido, C. Hsp70, A Cancer Target Inside and Outside the Cell. Methods Mol. Biol. 2018, 1709, 371–396. [Google Scholar]
- Campanella, C.; Bucchieri, F.; Merendino, A.M.; Fucarino, A.; Burgio, G.; Corona, D.F.V.; Barbieri, G.; David, S.; Farina, F.; Zummo, G.; et al. The Odyssey of Hsp60 from Tumor Cells to Other Destinations Includes Plasma Membrane-Associated Stages and Golgi and Exosomal Protein-Trafficking Modalities. PLoS ONE 2012, 7, e42008. [Google Scholar] [CrossRef]
- Campanella, C.; D'Anneo, A.; Gammazza, A.M.; Bavisotto, C.C.; Barone, R.; Emanuele, S.; Cascio, F.L.; Mocciaro, E.; Fais, S.; De Macario, E.C.; et al. The histone deacetylase inhibitor SAHA induces HSP60 nitration and its extracellular release by exosomal vesicles in human lung-derived carcinoma cells. Oncotarget 2016, 7, 28849–28867. [Google Scholar] [CrossRef] [PubMed]
- Cesa, L.C.; Shao, H.; Srinivasan, S.R.; Tse, E.; Jain, C.; Zuiderweg, E.R.; Southworth, D.R.; Mapp, A.K.; Gestwicki, J.E. X-linked inhibitor of apoptosis protein (XIAP) is a client of heat shock protein 70 (Hsp70) and a biomarker of its inhibition. J. Biol. Chem. 2018, 293, 2370–2380. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.-A.; Lee, Y.-S.; Kim, S.-H.; Ko, J.-K.; Kim, C.-W. MHC independent anti-tumor immune responses induced by Hsp70-enriched exosomes generate tumor regression in murine models. Cancer Lett. 2009, 275, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Ciocca, D.R.; Calderwood, S.K. Heat shock proteins in cancer: Diagnostic, prognostic, predictive, and treatment implications. Cell Stress. Chaperones 2005, 10, 86–103. [Google Scholar] [CrossRef] [PubMed]
- Clayton, A.; Turkes, A.; Navabi, H.; Mason, M.D.; Tabi, Z. Induction of heat shock proteins in B-cell exosomes. J. Cell Sci. 2005, 118, 3631–3638. [Google Scholar] [CrossRef] [PubMed]
- Collett, G.P.; Redman, C.W.; Sargent, I.L.; Vatish, M. Endoplasmic reticulum stress stimulates the release of extracellular vesicles carrying danger-associated molecular pattern (DAMP) molecules. Oncotarget 2018, 9, 6707–6717. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Dai, S.; Cao, J. Proteotoxic stress of cancer: Implication of the heat-shock response in oncogenesis. J. Cell. Physiol. 2012, 227, 2982–2987. [Google Scholar] [CrossRef]
- Dai, S.; Wan, T.; Wang, B.; Zhou, X.; Xiu, F.; Chen, T.; Wu, Y.; Cao, X. More Efficient Induction of HLA-A*0201-Restricted and Carcinoembryonic Antigen (CEA)–Specific CTL Response by Immunization with Exosomes Prepared from Heat-Stressed CEA-Positive Tumor Cells. Clin. Cancer Res. 2005, 11, 7554–7563. [Google Scholar] [CrossRef]
- Maio, D.; Vazquez, D. Extracellular heat shock proteins: A new location, a new function. Shock. 2013, 40, 239–246. [Google Scholar] [CrossRef]
- Dempsey, N.C.; Ireland, H.E.; Smith, C.M.; Hoyle, C.F.; Williams, J.H. Heat Shock Protein translocation induced by membrane fluidization increases tumor-cell sensitivity to chemotherapeutic drugs. Cancer Lett. 2010, 296, 257–267. [Google Scholar] [CrossRef]
- Diao, J.; Yang, X.; Song, X.; Chen, S.; He, Y.; Wang, Q.; Chen, G.; Luo, C.; Wu, X.; Zhang, Y. Exosomal Hsp70 mediates immunosuppressive activity of the myeloid-derived suppressor cells via phosphorylation of Stat3. Med. Oncol. 2015, 32, 453. [Google Scholar] [CrossRef]
- Eguchi, T.; Sogawa, C.; Okusha, Y.; Uchibe, K.; Iinuma, R.; Ono, K.; Nakano, K.; Murakami, J.; Itoh, M.; Arai, K.; et al. Organoids with cancer stem cell-like properties secrete exosomes and HSP90 in a 3D nanoenvironment. PLoS ONE 2018, 13, e0191109. [Google Scholar] [CrossRef] [PubMed]
- Elsner, L.; Muppala, V.; Gehrmann, M.; Lozano, J.; Malzahn, D.; Bickeböller, H.; Brunner, E.; Zientkowska, M.; Herrmann, T.; Walter, L.; et al. The Heat Shock Protein HSP70 Promotes Mouse NK Cell Activity against Tumors That Express Inducible NKG2D Ligands. J. Immunol. 2007, 179, 5523–5533. [Google Scholar] [CrossRef] [PubMed]
- Emam, S.E.; Ando, H.; Abu Lila, A.S.; Kobayashi, S.; Shimizu, T.; Okuhira, K.; Ishima, Y.; Ishida, T. Doxorubicin Expands in vivo Secretion of Circulating Exosome in Mice. Biol. Pharm. Bull. 2018, 41, 1078–1083. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Zhang, C.; Lum, D.; Druso, J.E.; Blank, B.; Wilson, K.F.; Welm, A.; Antonyak, M.A.; Cerione, R.A. A class of extracellular vesicles from breast cancer cells activates VEGF receptors and tumour angiogenesis. Nat. Commun. 2017, 8, 14450. [Google Scholar] [CrossRef] [PubMed]
- Ferrarini, M.; Heltai, S.; Zocchi, M.R.; Rugarli, C. Unusual expression and localization of heat-shock proteins in human tumor cells. Int. J. Cancer 1992, 51, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Gabai, V.L.; Budagova, K.R.; Sherman, M.Y. Increased expression of the major heat shock protein Hsp72 in human prostate carcinoma cells is dispensable for their viability but confers resistance to a variety of anticancer agents. Oncogene 2005, 24, 3328–3338. [Google Scholar] [CrossRef] [PubMed]
- Garrido; Schmitt, E.; Candé, C.; Vahsen, N.; Parcellier, A.; Kroemer, G. HSP27 and HSP70, potentially oncogenic apoptosis inhibitors. Cell Cycle 2003, 2, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Gastpar, R.; Gehrmann, M.; Bausero, M.A.; Asea, A.; Gross, C.; Schroeder, J.A.; Multhoff, G. Heat Shock Protein 70 Surface-Positive Tumor Exosomes Stimulate Migratory and Cytolytic Activity of Natural Killer Cells. Cancer Res. 2005, 65, 5238–5247. [Google Scholar] [CrossRef]
- Gobbo; Marcion, G.; Cordonnier, M.; Dias, A.M.M.; Pernet, N.; Hammann, A.; Richaud, S.; Mjahed, H.; Isambert, N.; Clausse, V.; Rébé, C.; Bertaut, A.; Goussot, V.; Lirussi, F.; Ghiringhelli, F.; de Thonel, A.; Fumoleau, P.; Seigneuric, R.; Garrido, C. Restoring Anticancer Immune Response by Targeting Tumor-Derived Exosomes With a HSP70 Peptide Aptamer. J. Natl. Cancer Inst. 2016, 108. [Google Scholar] [CrossRef]
- Géminard, C.; Nault, F.; Johnstone, R.M.; Vidal, M. Characteristics of the Interaction between Hsc70 and the Transferrin Receptor in Exosomes Released during Reticulocyte Maturation. J. Biol. Chem. 2001, 276, 9910–9916. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, J.C.; Donakonda, S.; Haupt, V.J.; Lennig, P.; Zhang, Y.; Schroeder, M. New HSP27 inhibitors efficiently suppress drug resistance development in cancer cells. Oncotarget 2016, 7, 68156–68169. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-L.; Hung, J.-Y.; Chang, W.-A.; Lin, Y.-S.; Pan, Y.-C.; Tsai, P.-H.; Wu, C.-Y.; Kuo, P.-L. Hypoxic lung cancer-secreted exosomal miR-23a increased angiogenesis and vascular permeability by targeting prolyl hydroxylase and tight junction protein ZO-1. Oncogene 2017, 36, 4929–4942. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.-B.; Wu, J.Y.; Lillard, J.; Bond, V.C. SMR peptide antagonizes mortalin promoted release of extracellular vesicles and affects mortalin protection from complement-dependent cytotoxicity in breast cancer cells and leukemia cells. Oncotarget 2019, 10, 5419–5438. [Google Scholar] [CrossRef] [PubMed]
- Hunter, A.M.; LaCasse, E.C.; Korneluk, R.G. The inhibitors of apoptosis (IAPs) as cancer targets. Apoptosis 2007, 12, 1543–1568. [Google Scholar] [CrossRef] [PubMed]
- Hunter-Lavin, C.; Davies, E.L.; Bacelar, M.M.; Marshall, M.J.; Andrew, S.M.; Williams, J.H. Hsp70 release from peripheral blood mononuclear cells. Biochem. Biophys. Res. Commun. 2004, 324, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Hurwitz, M.D.; Kaur, P.; Nagaraja, G.M.; Bausero, M.A.; Manola, J.; Asea, A. Radiation therapy induces circulating serum Hsp72 in patients with prostate cancer. Radiother. Oncol. 2010, 95, 350–358. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, G.; Huang, L.; Yuan, Y.; Wu, C.; Li, Y. Transmissible Endoplasmic Reticulum Stress: A Novel Perspective on Tumor Immunity. Front. Cell Dev. Biol. 2020, 8, 846. [Google Scholar] [CrossRef]
- Jäättelä, M. Over-expression of hsp70 confers tumorigenicity to mouse fibrosarcoma cells. Int. J. Cancer 1995, 60, 689–693. [Google Scholar] [CrossRef]
- Kabakov, A.; Yakimova, A.; Matchuk, O. Molecular Chaperones in Cancer Stem Cells: Determinants of Stemness and Potential Targets for Antitumor Therapy. Cells 2020, 9. [Google Scholar] [CrossRef]
- Kampinga, H.H.; Mayer, M.P.; Mogk, A. Protein quality control: From mechanism to disease : EMBO Workshop, Costa de la Calma (Mallorca), Spain, April 28—May 03, 2019. Cell Stress Chaperones 2019, 24, 1013–1026. [Google Scholar] [CrossRef] [PubMed]
- Kanemoto, S.; Nitani, R.; Murakami, T.; Kaneko, M.; Asada, R.; Matsuhisa, K.; Saito, A.; Imaizumi, K. Multivesicular body formation enhancement and exosome release during endoplasmic reticulum stress. Biochem. Biophys. Res. Commun. 2016, 480, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Keklikoglou, I.; Cianciaruso, C.; Güç, E.; Squadrito, M.L.; Spring, L.M.; Tazzyman, S.; Lambein, L.; Poissonnier, A.; Ferraro, G.B.; Baer, C.; et al. Chemotherapy elicits pro-metastatic extracellular vesicles in breast cancer models. Nat. Cell Biol. 2019, 21, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Kreger, B.T.; Johansen, E.R.; Cerione, R.A.; Antonyak, M.A. The Enrichment of Survivin in Exosomes from Breast Cancer Cells Treated with Paclitaxel Promotes Cell Survival and Chemoresistance. Cancers 2016, 8. [Google Scholar] [CrossRef]
- Kumar, P.; Siripini, S.; Sreedhar, A.S. The matrix metalloproteinase 7 (MMP7) links Hsp90 chaperone with acquired drug resistance and tumor metastasis. Cancer Rep. 2020, e1261. [Google Scholar] [CrossRef]
- König, L.; Kasimir-Bauer, S.; Bittner, A.-K.; Hoffmann, O.; Wagner, B.; Manvailer, L.F.S.; Kimmig, R.; Horn, P.A.; Rebmann, V. Elevated levels of extracellular vesicles are associated with therapy failure and disease progression in breast cancer patients undergoing neoadjuvant chemotherapy. OncoImmunology 2017, 7, e1376153. [Google Scholar] [CrossRef]
- Lancaster; G.I.; Febbraio, M.A. Exosome-dependent trafficking of HSP70, a novel secretory pathway for cellular stress proteins. J. Biol. Chem. 2005, 280, 23349–23355.
- Lettini; Lepore, S.; Crispo, F.; Sisinni, L.; Esposito, F.; Landriscina, M. Heat shock proteins in cancer stem cell maintenance: A potential therapeutic target? Histol. Histopathol. 2020, 35, 25–37. [Google Scholar]
- Li, Z.; Zhuang, M.; Zhang, L.; Zheng, X.; Yang, P.; Li, Z. Acetylation modification regulates GRP78 secretion in colon cancer cells. Sci. Rep. 2016, 6, 30406. [Google Scholar] [CrossRef]
- Lindquist, S. Protein Folding Sculpting Evolutionary Change. Cold Spring Harb. Symp. Quant. Biol. 2009, 74, 103–108. [Google Scholar] [CrossRef]
- Lv, L.-H.; Wan, Y.-L.; Lin, Y.; Zhang, W.; Yang, M.; Li, G.-L.; Lin, H.-M.; Shang, C.-Z.; Chen, Y.-J.; Min, J. Anticancer Drugs Cause Release of Exosomes with Heat Shock Proteins from Human Hepatocellular Carcinoma Cells That Elicit Effective Natural Killer Cell Antitumor Responses in Vitro. J. Biol. Chem. 2012, 287, 15874–15885. [Google Scholar] [CrossRef]
- Maacha, S.; Bhat, A.A.; Jimenez, L.; Raza, A.; Haris, M.; Uddin, S.; Grivel, J.-C. Extracellular vesicles-mediated intercellular communication: roles in the tumor microenvironment and anti-cancer drug resistance. Mol. Cancer 2019, 18, 55. [Google Scholar] [CrossRef] [PubMed]
- Mahadevan, N.R.; Rodvold, J.; Sepulveda, H.; Rossi, S.; Drew, A.F.; Zanetti, M. Transmission of endoplasmic reticulum stress and pro-inflammation from tumor cells to myeloid cells. Proc. Natl. Acad. Sci. USA 2011, 108, 6561–6566. [Google Scholar] [CrossRef] [PubMed]
- McCready, J.; Sims, J.D.; Chan, D.; Jay, D.G. Secretion of extracellular hsp90α via exosomes increases cancer cell motility: a role for plasminogen activation. BMC Cancer 2010, 10, 294–294. [Google Scholar] [CrossRef] [PubMed]
- Menay, F.; Herschlik, L.; De Toro, J.; Cocozza, F.; Tsacalian, R.; Gravisaco, M.J.; Di Sciullo, M.P.; Vendrell, A.; Waldner, C.I.; Mongini, C. Exosomes Isolated from Ascites of T-Cell Lymphoma-Bearing Mice Expressing Surface CD24 and HSP-90 Induce a Tumor-Specific Immune Response. Front. Immunol. 2017, 8, 286. [Google Scholar] [CrossRef]
- Merendino, A.M.; Bucchieri, F.; Campanella, C.; Marcianò, V.; Ribbene, A.; David, S.; Zummo, G.; Burgio, G.; Corona, D.F.V.; de Macario, E.C.; et al. Hsp60 Is Actively Secreted by Human Tumor Cells. PLOS ONE 2010, 5, e9247. [Google Scholar] [CrossRef]
- Nakano; Garnier, D.; Minata, M.; Rak, J. Extracellular vesicles in the biology of brain tumour stem cells--Implications for inter-cellular communication, therapy and biomarker development. Semin. Cell Dev. Biol. 2015, 40, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Namee, N. M.; O’Driscoll, L. Extracellular vesicles and anticancer drug resistance. Biochim. Biophys. Acta Rev. Cancer 2018, 1870, 123–136. [Google Scholar] [CrossRef]
- Nolan, K.D.; Kaur, J.; Isaacs, J.S. Secreted heat shock protein 90 promotes prostate cancer stem cell heterogeneity. Oncotarget 2017, 8, 19323–19341. [Google Scholar] [CrossRef]
- O’neill, C.P.; Gilligan, K.E.; Dwyer, R.M. Role of Extracellular Vesicles (EVs) in Cell Stress Response and Resistance to Cancer Therapy. Cancers 2019, 11. [Google Scholar] [CrossRef]
- Ono, K.; Eguchi, T.; Sogawa, C.; Calderwood, S.K.; Futagawa, J.; Kasai, T.; Seno, M.; Okamoto, K.; Sasaki, A.; Kozaki, K. HSP-enriched properties of extracellular vesicles involve survival of metastatic oral cancer cells. J. Cell. Biochem. 2018, 119, 7350–7362. [Google Scholar] [CrossRef]
- Ono; Sogawa, C.; Kawai, H.; Tran, M.T.; Taha, E.A.; Lu, Y.; Oo, M.W.; Okusha, Y.; Okamura, H.; Ibaragi, S.; Takigawa, M.; Kozaki, K.I.; Nagatsuka, H.; Sasaki, A.; Okamoto, K.; Calderwood, S.K.; Eguchi, T. Triple knockdown of CDC37, HSP90-alpha and HSP90-beta diminishes extracellular vesicles-driven malignancy events and macrophage M2 polarization in oral cancer. J. Extracell. Vesicles 2020, 9, 1769373. [Google Scholar] [CrossRef]
- Peinado; Alečković, M.; Lavotshkin, S.; Matei, I.; Costa-Silva, B.; Moreno-Bueno, G.; Hergueta-Redondo, M.; Williams, C.; García-Santos, G.; Ghajar, C.; Nitadori-Hoshino, A.; Hoffman, C.; Badal, K.; Garcia, B.A.; Callahan, M.K.; Yuan, J.; Martins, V.R.; Skog, J.; Kaplan, R.N.; Brady, M.S.; Wolchok, J.D.; Chapman, P.B.; Kang, Y.; Bromberg, J.; Lyden, D. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat. Med. 2012, 18, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Prager, B.C.; Xie, Q.; Bao, S.; Rich, J.N. Cancer Stem Cells: The Architects of the Tumor Ecosystem. Cell Stem Cell 2019, 24, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Ramteke, A.; Ting, H.; Agarwal, C.; Mateen, S.; Somasagara, R.; Hussain, A.; Graner, M.; Frederick, B.; Agarwal, R.; Deep, G. Exosomes secreted under hypoxia enhance invasiveness and stemness of prostate cancer cells by targeting adherens junction molecules. Mol. Carcinog. 2015, 54, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Raposo; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef]
- Rothammer, A.; Sage, E.K.; Werner, C.; Combs, S.E.; Multhoff, G. Increased heat shock protein 70 (Hsp70) serum levels and low NK cell counts after radiotherapy—Potential markers for predicting breast cancer recurrence? Radiat Oncol 2019, 14, 78. [Google Scholar] [CrossRef]
- Santos, T.G.; Martins, V.R.; Hajj, G.N.M. Unconventional Secretion of Heat Shock Proteins in Cancer. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef]
- Sen, K.; Sheppe, A.E.F.; Singh, I.; Hui, W.W.; Edelmann, M.J.; Rinaldi, C. Exosomes released by breast cancer cells under mild hyperthermic stress possess immunogenic potential and modulate polarization. Int. J. Hyperthermia 2020, 37, 696–710. [Google Scholar] [CrossRef]
- Shao, H.; Chung, J.; Balaj, L.; Charest, A.; Bigner, D.D.; Carter, B.S.; Hochberg, F.H.; Breakefield, X.O.; Weissleder, R.; Lee, H. Protein typing of circulating microvesicles allows real-time monitoring of glioblastoma therapy. Nat. Med. 2012, 18, 1835–1840. [Google Scholar] [CrossRef]
- Shedden, K.; Xie, X.T.; Chandaroy, P.; Chang, Y.T.; Rosania, G.R. Expulsion of small molecules in vesicles shed by cancer cells: Association with gene expression and chemosensitivity profiles. Cancer Res. 2003, 63, 4331–4337. [Google Scholar] [PubMed]
- Sun, Z.; Wang, L.; Dong, L.; Wang, X. Emerging role of exosome signalling in maintaining cancer stem cell dynamic equilibrium. J. Cell. Mol. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Taha, E.A.; Ono, K.; Eguchi, T. Roles of Extracellular HSPs as Biomarkers in Immune Surveillance and Immune Evasion. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Takayama, S.; Reed, J.C.; Homma, S. Heat-shock proteins as regulators of apoptosis. Oncogene 2003, 22, 9041–9047. [Google Scholar] [CrossRef]
- Tamura, Y.; Torigoe, T.; Kutomi, G.; Hirata, K.; Sato, N. New Paradigm for Intrinsic Function of Heat Shock Proteins as Endogenous Ligands in Inflammation and Innate Immunity. Curr. Mol. Med. 2012, 12, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Chang, C.; Guo, J.; Lincoln, V.; Liang, C.; Chen, M.; Woodley, D.T.; Li, W. Tumour-Secreted Hsp90α on External Surface of Exosomes Mediates Tumour - Stromal Cell Communication via Autocrine and Paracrine Mechanisms. Sci. Rep. 2019, 9, 15108. [Google Scholar] [CrossRef]
- Théry, C.; Regnault, A.; Garin, J.; Wolfers, J.; Zitvogel, L.; Ricciardi-Castagnoli, P.; Raposo, G.; Amigorena, S. Molecular Characterization of Dendritic Cell-Derived Exosomes. J. Cell Biol. 1999, 147, 599–610. [Google Scholar] [CrossRef]
- Théry; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Tsen; Bhatia, A.; O’Brien, K.; Cheng, C.F.; Chen, M.; Hay, N.; Stiles, B.; Woodley, D.T.; Li, W. Extracellular heat shock protein 90 signals through subdomain II and the NPVY motif of LRP-1 receptor to Akt1 and Akt2, a circuit essential for promoting skin cell migration in vitro and wound healing in vivo. Mol. Cell Biol. 2013, 33, 4947–4959. [Google Scholar] [CrossRef]
- Van Niel, G.; Raposo, G.; Candalh, C.; Boussac, M.; Hershberg, R.; Cerf-Bensussan, N.; Heyman, M. Intestinal epithelial cells secrete exosome–like vesicles. Gastroenterology 2001, 121, 337–349. [Google Scholar] [CrossRef]
- Vargas-Roig, L.M.; Gago, F.E.; Tello, O.; Aznar, J.C.; Ciocca, D.R. Heat shock protein expression and drug resistance in breast cancer patients treated with induction chemotherapy. Int. J. Cancer 1998, 79, 468–475. [Google Scholar] [CrossRef]
- Vega, V. L.; Rodríguez-Silva, M.; Frey, T.; Gehrmann, M.; Diaz, J.C.; Steinem, C.; Multhoff, G.; Arispe, N.; De Maio, A. Hsp70 translocates into the plasma membrane after stress and is released into the extracellular environment in a membrane-associated form that activates macrophages. J. Immunol. 2008, 180, 4299–4307. [Google Scholar] [CrossRef]
- Vinik; Ortega, F.G.; Mills, G.B.; Lu, Y.; Jurkowicz, M.; Halperin, S.; Aharoni, M.; Gutman, M.; Lev, S. Proteomic analysis of circulating extracellular vesicles identifies potential markers of breast cancer progression, recurrence, and response. Sci. Adv. 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Wubbolts; Leckie, R.S.; Veenhuizen, P.T.; Schwarzmann, G.; Möbius, W.; Hoernschemeyer, J.; Slot, J.W.; Geuze, H.J.; Stoorvogel, W. Proteomic and biochemical analyses of human B cell-derived exosomes. Potential implications for their function and multivesicular body formation. J. Biol. Chem. 2003, 278, 10963–10972. [Google Scholar] [CrossRef]
- Wyciszkiewicz, A.; Kalinowska-Łyszczarz, A.; Nowakowski, B.; Kaźmierczak, K.; Osztynowicz, K.; Michalak, S. Expression of small heat shock proteins in exosomes from patients with gynecologic cancers. Sci. Rep. 2019, 9, 9817. [Google Scholar] [CrossRef] [PubMed]
- Xie; Bai, O.; Zhang, H.; Yuan, J.; Zong, S.; Chibbar, R.; Slattery, K.; Qureshi, M.; Wei, Y.; Deng, Y.; Xiang, J. Membrane-bound HSP70-engineered myeloma cell-derived exosomes stimulate more efficient CD8(+) CTL- and NK-mediated antitumour immunity than exosomes released from heat-shocked tumour cells expressing cytoplasmic HSP70. J Cell Mol Med 2010, 14, 2655–2666. [Google Scholar] [CrossRef] [PubMed]
- Young, J.C.; Barral, J.M.; Hartl, F.U. More than folding: localized functions of cytosolic chaperones. Trends Biochem. Sci. 2003, 28, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Yukawa, H.; Suzuki, K.; Aoki, K.; Arimoto, T.; Yasui, T.; Kaji, N.; Ishikawa, T.; Ochiya, T.; Baba, Y. Imaging of angiogenesis of human umbilical vein endothelial cells by uptake of exosomes secreted from hepatocellular carcinoma cells. Sci. Rep. 2018, 8, 6765. [Google Scholar] [CrossRef]
- Andrade, L.N.d.S.; Otake, A.H.; Cardim, S.G.B.; da Silva, F.I.; Sakamoto, M.M.I.; Furuya, T.K.; Uno, M.; Pasini, F.S.; Chammas, R. Extracellular Vesicles Shedding Promotes Melanoma Growth in Response to Chemotherapy. Sci. Rep. 2019, 9, 14482. [Google Scholar] [CrossRef]
- Nederveen, J.P.; Warnier, G.; Di Carlo, A.; Nilsson, M.I.; Tarnopolsky, M.A. Extracellular Vesicles and Exosomes: Insights from Exercise Science. Front. Physiol. 2021, 11, 604274. [Google Scholar] [CrossRef]
Figure 1.
EV-HSPs acting as a double-edged sword inside the tumor microenvironment. (HSP: Heat Shock Protein; MDSC: Myeloid-Derived Suppressor Cell; NK: Natural Killer). Created with BioRender.com.
Figure 1.
EV-HSPs acting as a double-edged sword inside the tumor microenvironment. (HSP: Heat Shock Protein; MDSC: Myeloid-Derived Suppressor Cell; NK: Natural Killer). Created with BioRender.com.
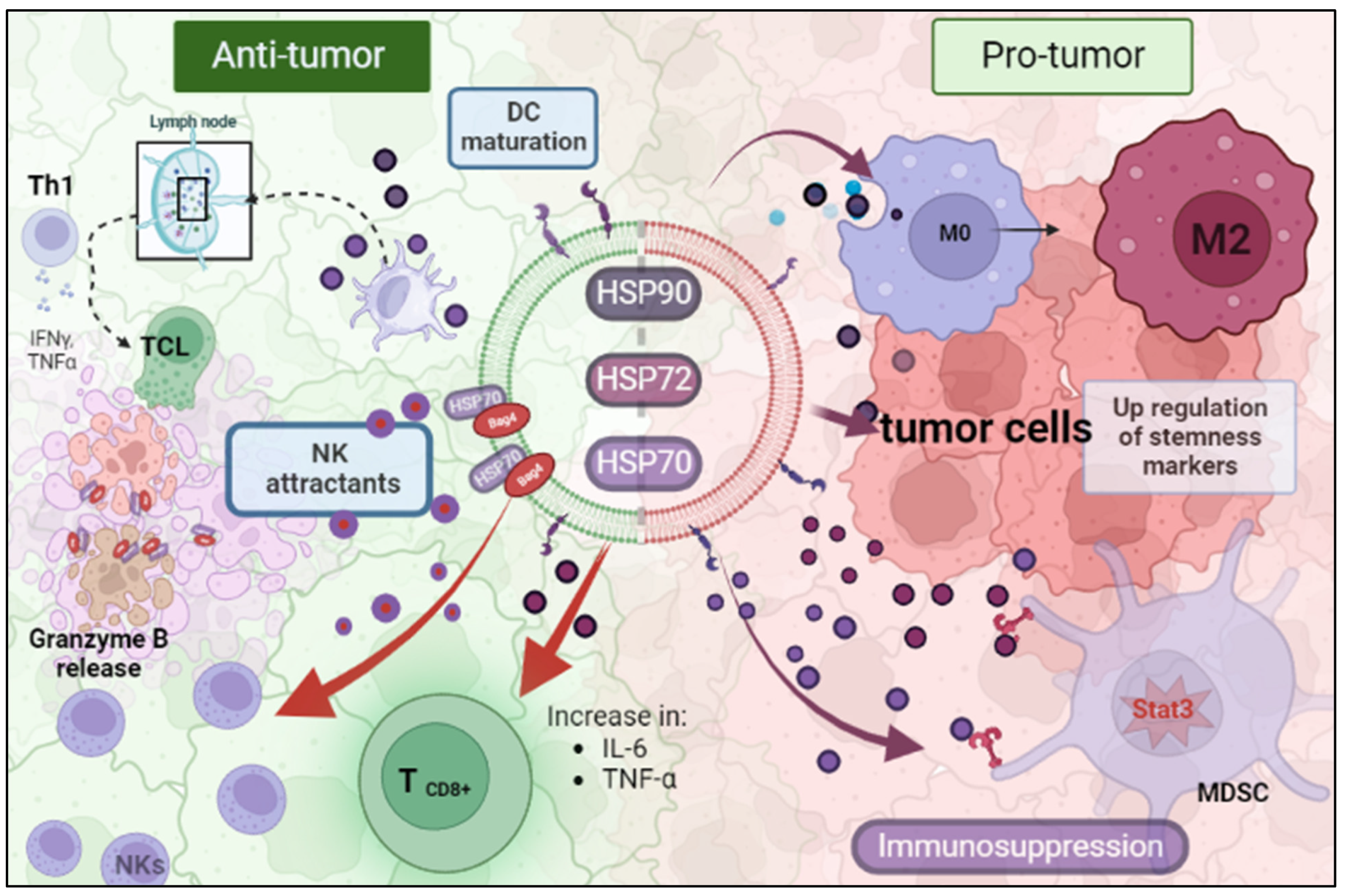
Figure 2.
Roles of Extracellular vesicles (EVs) derived from tumor microenvironment (TME) after injuries acting in tumor repopulation (ER: Endoplasmic Reticulum; HSP: Heat Shock Protein; MDSC: Myeloid-Derived Suppressor Cell; NK: Natural Killer). Created with BioRender.com.
Figure 2.
Roles of Extracellular vesicles (EVs) derived from tumor microenvironment (TME) after injuries acting in tumor repopulation (ER: Endoplasmic Reticulum; HSP: Heat Shock Protein; MDSC: Myeloid-Derived Suppressor Cell; NK: Natural Killer). Created with BioRender.com.
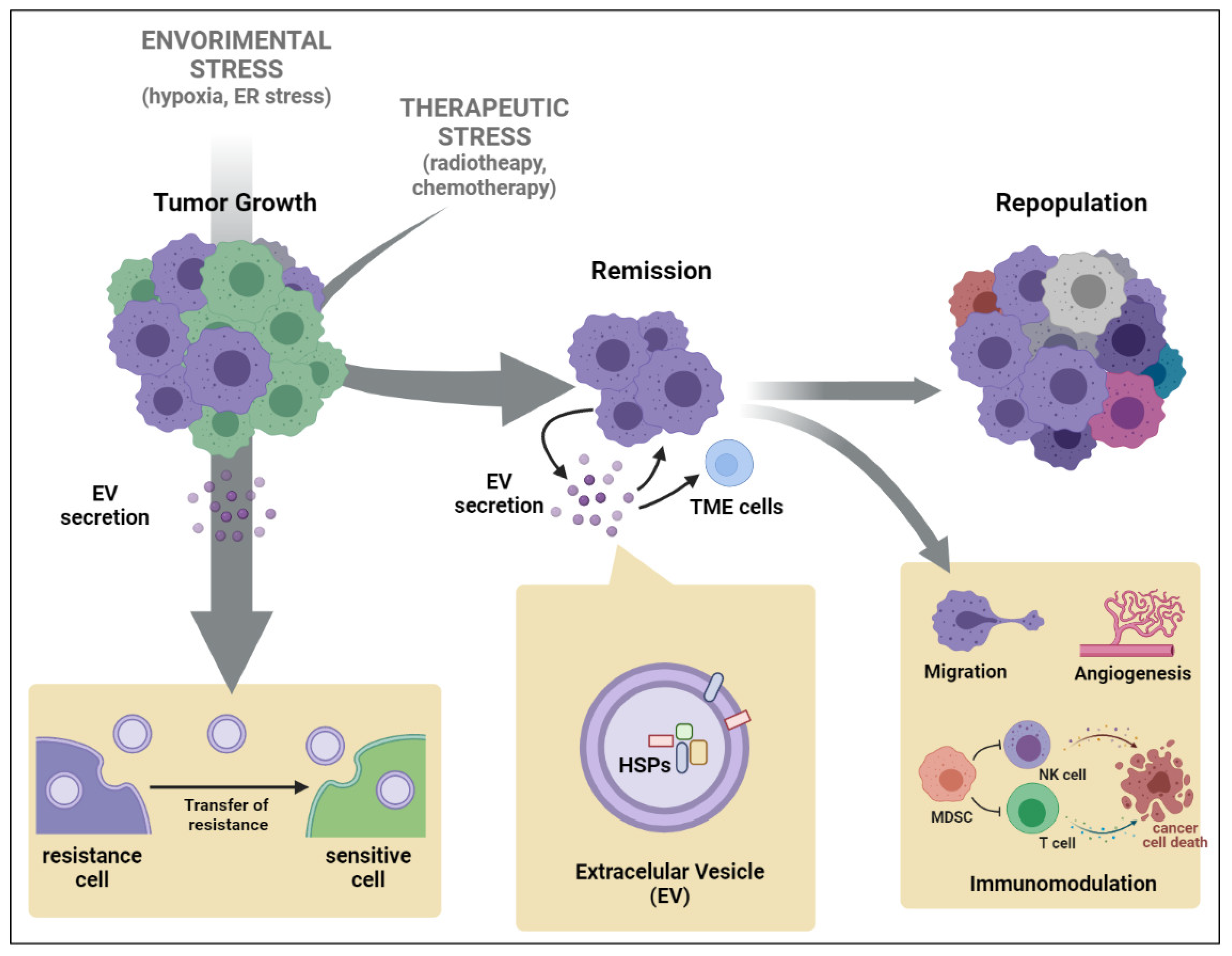

Table 1.
Chaperone-containing EVs secreted by different cellular types.
| Chaperone | Cell type | Reference |
|---|---|---|
| HSP20 | Gynecologic cancer cells | (Wyciszkiewicz, Kalinowska-Łyszczarz et al. 2019) |
| HSP27 HSC70 HSP70 HSP90 |
B cells | (Clayton, Turkes et al. 2005) |
| HSP60 | Human lung carcinoma cells | (Campanella, Bucchieri et al. 2012) |
| HSP60 HSP70 |
H292, A549 and K562 tumor cell lines | (Merendino, Bucchieri et al. 2010) |
| HSP60 HSP70 HSP90 |
Hepatocellular carcinoma cells | (Lv, Wan et al. 2012) |
| HSP70 | Human peripheral blood mononuclear cells | (Lancaster and Febbraio 2005) |
| HSP70 | Natural killer cells | (Gastpar, Gehrmann et al. 2005) |
| HSP70 | Choriocarcinoma cells | (Collett, Redman et al. 2018) |
| HSP72 | Breast adenocarcinoma cells Erythroleukemic cells |
(Bausero, Gastpar et al. 2005) |
| HSC73 | Dendritic cells | (Théry, Regnault et al. 1999) |
| HSP70 HSP90 |
Prostate cancer cell | (Ramteke, Ting et al. 2015) |
| mt-HSP70 | Breast cancer cells | (Huang, Wu et al. 2019) |
| GRP78 | Colon cancer cells | (Li, Zhuang et al. 2016) |
| HSP90 | Cancer stem cell-like | (Eguchi, Sogawa et al. 2018) |
Table 2.
EV-cargo HSPs dual role in cancer.
| Chaperone | Activity | |
|---|---|---|
| Pro-tumor | Anti-tumor | |
| HSP70 | Promote cell-survival, protect againist oxidative stress and others, promote protein folding and degradation, and promote cell migration and invasion | Induce tumor cells apoptotic death and sensibilize to chemo or radiotherapies |
| HSP72 | Promote angiogenesis, protect cancer cells from oxidative stress. Supress-apoptosis, promote cell invation and migration | Favor arresting in tumor growth, promote apoptosis, sensitize to chemotherapy |
| HSP90 | promote protein folding and stabilization of multiple proteins, promote cell survival,supress apoptosis | promote apoptosis, sensitize to chemo- and radiotherapy |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



