
Submitted:
18 May 2023
Posted:
19 May 2023
You are already at the latest version
Abstract
The channels from the superfamily of transient receptor potential (TRP) activated by reactive oxygen species (ROS) are defined as redox channels. Those with the best exposure of the cysteine residues and hence the most sensitive to oxidative stress are TRPC4, TRPC5, TRPV1, TRPV4, and TRPA1, while others, such as TRPM2 and TRPM7 are indirectly activated by ROS. Activation by ROS has different effects on the tumorigenic process: some TRP channels may, upon activation, stimulate proliferation, apoptosis, or migration of cancer cells, while others inhibit these processes, depending on the cancer type, tumoral microenvironment and finally on the methods used for evaluation. Therefore, the use of these polymodal proteins as therapeutic targets is still an unmet need, despite their draggability and modulation by simple and mostly unharmful compounds. In this review, we intend to create some cellular models of the interaction between oxidative stress, TRP channels, and inflammation. Although the crosstalk between the three actors is rather theoretical, we intended to gather the recently published data and proposed pathways of cancer inhibition using modulators of TRP proteins, hoping that the experimental data corroborated with clinical information may finally bring the results from the bench to the bedside.
Keywords:
ROS
; TRP
; inflammation
1. Introduction
Transient receptor potential (TRP) channels constitute a superfamily of cation-permeable (Na+, K+, Ca2+) ion channels that exhibit extraordinary diversity in structure, activation mechanisms, and physiological functions. Today, 30 years after the discovery of the first TRP channel, this superfamily includes more than 100 members, found in both vertebrates and invertebrates. TRP members are grouped into seven families based on structural homology: TRPC (classical or canonical), TRPV (vanilloid), TRPM (melastatin), TRPA (ankyrin), TRPML (mucolipin), TRPP (polycystin), TRPN (NOMPC - no mechanoreceptor potential C). Apart from TRPN proteins – found in invertebrates and fish, all TRP proteins have been identified in mammals (Drosophila TRP channels). A multitude of stimuli activates TRP channels as temperature, membrane potential (voltage), oxidative stress, mechanical stimuli, pH, and ligands (both endogenous and exogenous), thus illustrating their adaptability. Many TRP members are key proteins in pain perception and integration of nociceptive signals; activation of these TRP sensors leads to cellular depolarization by increasing the intracellular concentration of cations, resulting in protective responses such as pain or local inflammation [1].
As such, TRP channels modulate the functions of both excitable and non-excitable cells, mainly by calcium homeostasis [2]. The uptake of Ca2+ is specific for most TRP channels, with some members sharing a higher relative permeability to Ca2+ over Na+ (PCa/PNa). TRPV5 and TRPV6 are specifically Ca2+-selective with PCa/PNa > 100 [3], whereas TRPM4 and TRPM5 have almost insignificant calcium permeability of < 0.1 [4]. These differences come from the well-established structure of the selectivity filter and the variability of the amino acids lining the central pore. All TRP family members generally form tetramers, and each subunit shares the same structure: 6 transmembrane domains with the pore region between the fifth and the sixth domain. The contribution of TRP channels in so different cellular functions relies not only on the pore structure but also on their gating properties, which are modulated by various stimuli. Also, detailed structural models predicted the way that these complex proteins are regulated by endo- and exogenous factors.
This review envisages the role of TRP channels that are sensors for oxidative reactions in cancer cells. In the last years, the specific TRPs directly or indirectly activated by reactive oxygen species (ROS) are named redox-sensitive TRP channels [5] and we will restrain this review to these TRP family members. We will also bring into focus the work of the last ten years based on the intricate interplay between redox channels, inflammation, and cancer. Given the complexity of each of these processes addressed mainly in specific and separated studies, this review tries to fill the gaps with possible mechanisms and signaling pathways.
A key question is whether redox channels can serve as therapeutic targets or prognostic markers for cancer. To address this problem, we will compare studies proposing means to use TRP redox channels in cancer therapy and emit the addressability of these complex proteins as tumoral biomarkers.
2. The Structure of the TRP Redox Channels
The most common means by which ROS directly activate the TRP channels involves redox modification of the free thiol groups. The channels with the best exposure of the cysteine residues and in consequence, the most sensitive to oxidative stress are TRPC4, TRPC5, TRPV1, TRPV4, and TRPA1 (Figure 1).
The channels from the TRPC family bear conserved cysteines residues situated in the pore-forming region and accessible from the cytoplasm denoted C549/C554 in TRPC4 and C553/C558 in TRPC5 [6].
Channels from the TRPV family have extracellular and intracellular cysteine residues. Oxidizing compounds activate rat and chicken TRPV1 by acting on extracellular Cys residues from the cytoplasmic side. [7], whereas human TRPV1 is activated by oxidizing agents via dimerization. Using non-reducing SDS Page electrophysiology and mass spectrometry, Ogawa et al. suggest that from many Cys residues in the N terminal, Cys-127, -158, -258, -363, and -742 participate in intersubunit disulfide bond formation and are sensitive to oxidation. From these, the most sensitive residue assessed by half-maximum saturation of H2O2 was Cys 258 localized in the N-terminal domain of TRPV1 [8].
In a recent report, mutational studies revealed that cysteines from the extracellular domain of TRPV4 (C639, C645, C652, and C660) that share homology with TRPV1 cysteines are ROS sensitive [9]. Also, mutation of several intracellular cysteines (C294, C353, and C427) abolished the regulatory role of TRPV4 in the invadasome through a process directly linked to ROS sensing. Both extra and intracellular cysteine groups are linked to cell invasion related to the degradative functions of acto-adhesive and extracellular matrix [9].
TRPA1, the most ROS-sensitive channel is activated either by extracellular compounds such as hydrogen peroxide (H2O2), hydroxyl radical (HO·), nitric oxide or by endogenously generated products such as 4-hydroxinonenal (4-HNE). This sensitivity comes from both cysteine residues (Cys 621, Cys 641, Cys 665) and to a lesser extent lysine 708, localized on their N-terminal segments [10].
We also define redox channels TRPM2 and TRPM7, which are indirectly activated by ROS, but through ADP-ribose (ADPR) generated in the mitochondria by NADase hydrolyses [11]. A high-resolution structure of TRPM2 revealed that the N terminus contains four TRPM homology regions, while the C-terminus has a conserved coiled-coil tetramerization domain and a NUDT9-H domain. TRPM1/2 and NUDT9-H domains provide the binding sites for ADPR and other ADPR analogs, such as nicotinic acid adenine dinucleotide phosphate (NAADP) and deacetylation product release by sirtuins – OAADPr, which activate the channel [12].
TRPM7 is a channel negatively regulated by intracellular Mg2+. H2O2 inhibits TRPM7 currents in an Mg2+ and ATP-dependent manner [13]. Site-directed mutagenesis studies revealed that cysteine residues (C1809 and C1813) of the zinc-binding motif act as oxidative stress sensors and increase Mg2+ inhibition perturbing the interactions between the channel domain and the kinase domain [14].
Apart from cysteines, free radicals may target methionine from TRP ion channels which turns to methionine sulfoxide and alters the protein structure and in consequence channel gating and signaling pathways. For instance, mutagenesis analysis showed that the redox sensitivity of TRPV2 depends on methionine residues M528 and M607 [15].
The oxidation of the sensitive residues of redox TRP channels has various effects on the tumorigenic process and modulates the proliferation and growth of cancer cells, migration, and apoptosis, depending on the ROS sources and the interaction they have with different cellular compartments.
3. Sources of Oxidative Stress and Their Interaction with Redox TRP Channels
Oxidative stress represents the disequilibrium between the generation and accumulation of free radicals or peroxides. Free radicals are oxygens with an uneven number of electrons, which raises the capacity to interact with other molecules, whereas peroxides are compounds in which a covalent bond links two oxygen atoms. Free radicals, classified as reactive oxygen species (ROS) reactive nitrogen species (RNS), and reactive carbonyl species (RCS) are derived from exogenous and endogenous sources. Production of free radicals from exogenous sources may be consecutive to pollutant exposure, heavy metals, ingestion of some drugs, cigarette smoke, alcohol, or radiation. As defense mechanism cells respond with a rapid release of ROS defined as the oxidative burst. Instead, tumoral processes, inflammation, infection, and exercise, are endogenous sources of free radical production [16]. In animal cells, ROS are formed primarily in mitochondria and peroxisomes, but also in any other cellular compartment presenting proteins or molecules with high redox potential [17]. They are then removed or detoxified by antioxidative enzymes and antioxidants, thus helping the cells to maintain ROS at low concentrations.
The ROS with the higher cellular impact are hydrogen peroxide H2O2, O2-, hydroxyl radical: OH–, and singlet oxygen. Generally, oxidative stress and free radicals are considered harmful to human organisms by breaking down DNA and proteins, but the generation of ROS is also beneficial, as recently reviewed in [18]. For instance, the immune system uses ROS to fight against pathogens. It is documented that macrophages use ROS to destroy invasive microorganisms, consecutive to oxidative burst mediated by NADPH [19]. The concept that ROS have physiological roles stay in the data proving that mitochondrial ROS improve skin development and regeneration [20], neuronal differentiation [21], or cell apoptosis [22]. However, the deleterious effects of ROS are much better documented. Cells are damaged by the interaction with the lipids’ membrane, proteins, or DNA. In the following paragraphs, we will discuss the role of ROS on different cellular compartments and their relationship to TRP channels.
3.1. Role of TRP Channels in ROS-Induced DNA Damage
ROS induce peroxidation of fatty acids and eventually impair membrane fluidity, protein-lipid interactions, and ion transport [23]. In cancer, lipid peroxidation triggers ferroptosis -the iron-dependent form of non-apoptotic cell death- consecutive to lipid hydroperoxide accumulation [24]. Ferroptosis occurs when the accumulation of lipid peroxides surpasses the cellular defense mechanisms. Apart from its effects on neurons, cardiac cells, and immune cells, recent studies report the role of ferroptosis in cancer [25]. The enrichment of PUFA-PLs in cell membranes, the overload of iron, and the imbalance of ferroptosis defense mechanisms inhibit tumor growth and proliferation and even augment anti-tumor immunity. This statement is based on previous data reporting that CD8+ T cells suppress tumorigenesis consecutive to the activation upon immunotherapy- drive ferroptosis [26].
As reported for redox TRP channels, recent studies showed that stimulation of TRPV4 induces exocytosis and leads to human melanoma cell ferroptosis [27]. The authors showed that TRPV4 stimulation in melanoma cell lines changed mitochondria structure and crista morphology. These changes are specific to ferroptosis, experimentally assessed by changes in the expression of ferroptosis markers: iron metabolism-associated gene expression of FTH1 (ferritin heavy chain 1), ACSL4 (acyl-CoA synthetase long-chain family member 4) and PTGS2 (prostaglandin-endoperoxide synthase 2). When activated, TRPV4 triggers ferroptosis through signaling pathways linked to increased intracellular calcium levels and in the end inhibits tumorigenesis. Another study proposes the use of TRPV4 agonist GSK1016790A in combination with cisplatin (an anticancer drug that produces ferroptosis) for the treatment of oral squamous cell carcinoma [28]. However, these results are disputed by Fujii S. et. al. The authors prove that TRPV4 promotes cell proliferation [29] and reiterate the double-edged sword of both TRP activation and ROS.
The relationship between the immune system and ferroptosis is however more complicated than this, as shown by disputed results obtained from studies of interleukins and tumor necrosis factor-alpha (TNF-α). Although serum interleukin 6 (IL-6) is a mediator of inflammatory diseases and lung cancer [30] and indicates a worse prognosis [31], unexpectedly promotes ferroptosis in bronchial epithelial cells [32]. Experiments on head and neck squamous cell carcinoma and clinical data correlate ferroptosis resistance with tumor progression [33,34].
Lipid peroxidation generates short-chain aldehydes and malondialdehydes (MDAs), which in turn inactivates proteins that promote ferroptosis. A type of toxic aldehyde is 4-hydroxynonenal (4-HNE) with the ability to induce structural and functional changes by the covalent binding to different proteins. In contrast to other unstable ROS, 4-HNE is a lipid peroxidation product with a high affinity for proteins forming stable complexes. 4-HNE strongly influences the genome and has affected the differentiation, proliferation, and apoptosis of cells [35]. Specifically, 4-HNE reacts with thiol groups leading to the formation of covalent bonds and consecutively posttranslational modification of proteins.
Studies on HEK cells showed that 4-HNE inhibited TRPV1 currents and mediated Ca2+ influx. In silico analysis revealed four putative binding sites in the pore helixes of TRPV1: cysteine residues 616, 621, and 634. However, mutational studies indicated that the effect was linked to a key pore cysteine residue 621, responsible for the impaired activity of TRPV1 due to the direct binding of 4-HNE [36]. The present literature describes many physiological alterations associated with 4-HNE. We will discuss the recent reports on 4-HNE and redox TRPs in a forthcoming chapter. For now, we recapitulate the roles of redox TRPs after DNA-induced ROS damage.
3.2. Role of TRP Channels in ROS-Induced DNA Damage
ROS induce DNA damage and consecutively dysregulate cellular responses to DNA damage response (DDR). Among other inducers of DDR, anticancer agents such as the divalent platinum compound cisplatin (CDDP) and the anthracycline doxorubicin (Dox) increase ROS levels and modulate the expression of several TRP channels. The results comprise toxic effects or resistance to chemotherapies.
Cisplatin is used in the treatment of various solid cancers, but unfortunately has many different secondary effects including hearing loss. The compound directly binds to DNA, accumulates in mitochondria, and generates ROS, which mediates DDR. Several studies showed that CCDP-induced ototoxicity is related to the production of 4-HNE. The authors also described Ca2+ overload in cochlear cells and overexpression of the TRPV1 channel. The secondary effects were reversed by ursolic acid (UA), a plant-derived triterpene compound with strong antioxidant effects and a blocker of TRPV1 [37].
Doxorubicin (Dox) intercalates into DNA, inhibits topoisomerase II, changes the chromatin structure, and increases ROS production. Its therapeutic application is hampered by cardiomyopathy and symptomatic heart failure noticed in up to 5% of cancer patients [38]. In a study conducted on cardiac tissues collected from mice, the authors measured high levels of TRPA1 mRNA and protein after Dox treatment, correlated with myocardial lesions. The functional consequences were cardiac dysfunctions, attenuation of oxidative stress, an increase of proinflammatory cytokines, and endoplasmic reticulum stress [39].
TRPC1 expression predicts breast cancer vulnerability to Dox and pulsed electromagnetic field (PEMF) therapy [40]. The cumulative ROS production by combined PEMF and Dox therapy creates an oxidative environment sufficient to slow breast cancer cells growth. Overexpression of TRPC1 increased the sensitivity to this therapy in animal models. These findings prompted the authors to propose the combinatory therapeutic strategy of PEMT + Dox, specifically for cancers characterized by high TRPC1 channel expression.
Another study reported that intensive use of doxorubicin induces pathological protein interaction of TRPC3 channels with NADPH oxidase 2 (Nox2) consecutive to cardiac atrophy in mice. Ibudilast, an anti-asthmatic drug, suppressed ROS production and cytotoxicity by destabilizing the TRPC3-Nox2 complexes, without reducing TRPC3 channel activity [41].
TRPM2 had protective roles in human breast adenocarcinoma cell lines, by minimizing DNA damage. Inhibition of TRPM2 with the specific antagonist, 2-APB, decreased cancer cell proliferation by about 60% and saved non-tumoral cells. These results indicate that the use of TRPM2 as a therapeutic target in breast cancer can pave the way to a more effective treatment. Alternatively, tumors with lower TRPM2 levels may be more sensitive to chemotherapy [42].
Other studies sustain the use of agonists and antagonists of redox TRP channels for their effects on proliferation, tumor growth, or cell apoptosis. It is not always clear whether these compounds are efficient because of the interaction with channels pore or by a direct action on other cellular compartments. In the following subchapter, we review recent data on the participation of redox TRPs in tumorigenesis upon the activation by ROS.
4. Regulation of TRP Channels by Oxidative Stress in Cancer Cell Proliferation
The fine tune of ROS levels has different effects on cancer cell proliferation and uses different pathways. Most of the results related to redox TRP come from experiments showing that TRPM2 and TRPM7, indirectly activated by ROS, influence cell proliferation. Figure 2 summarizes the results presenting the signaling pathways activated in cell proliferation by TRPM2 and TRPM7.
Firstly, we discuss the compelling evidence coming from the analysis of TRPM2 in acute myeloid leukemia (AML). Using CRISPR/Cas9 technology, the results show that in TRPM2-depleted leukemia cells, ROS levels were higher, while mitochondria function was reduced. In these conditions, cell proliferation significantly decreased. The authors bring strong evidence that TRPM2 regulates mitochondria bioenergetics, cell proliferation, and autophagy in AML by synchronous modulation of several transcription factors such as HIF-1/2α, Nrf2, ATF4, and CREB [43]. Using the same technology, the study of Bao L et al. [44] strengths that TRPM2 preserves survival of neuroblastoma cell lines and xenografts after oxidative injury. Here, the authors reveal the importance of the nuclear factor erythroid 2-related factor (Nrf2).
Also, TRPM2 regulates the tumorigenic process by activating major signaling pathways: JNK, PKC, or MAPK. The effect may depend on the splice variant expressed in different tumors. For instance, the isoform, TRPM2-S, lacking the four C-terminal transmembrane domains and the putative Ca2+ pore, is the prognostic biomarker for retroperitoneal liposarcoma (RPLS), which modulated proliferation and apoptosis via ERK and AKT pathways. Under oxidative stress conditions, mimicked by high levels of H2O2, the TRPM2-S enhanced apoptosis. In normoxia conditions, this isoform promoted the proliferation of RPLS cells [45].
Xing, Y. et al. systematically explore the role of TRPM7 channels and various modulators (Clozapine, Naltriben, Proadifen, Mibefradil, FTY720, and NS8593) on several cancer cell lines, in vivo xenograft, and metastasis mouse models. The study showed that pharmacological activation of TRPM7 specifically inhibits autophagy and suppresses proliferation, as quantified by changes in microtubule associated protein 1 light chain 3 (LC3) and p62. Importantly, The TRPM7 agonists Clozapine and Naltriben elevated ROS levels, while TRPM7 silencing inhibited ROS production by impairing mitochondrial turnover [46].
Missing conspicuous data of a direct liaison between activated -ROS TRPA1/V1 and proliferation, we propose a possible link that emerged from studies that independently analyzed either the modulation by ROS or the role of TRPA1/V1 on cell proliferation.
Experiments performed on lung adenocarcinoma cells propose a bold hypothesis: the N-terminal ankyrin repeats of TRPA1 directly bind to the C-terminal proline-rich motif of FGFR2 inducing the activation of the receptor through the MAP kinase pathway [47]. TRPA1 is the major ROS sensor, and the ankyrin repeats may form protein scaffolds with unexpected functions in different tumoral cells. However, no other studies support these findings, and TRPA1 remains a proposed target especially based on the uptake of Ca2+. In response to oxidative stress, TRPA1 regulates intracellular Ca2+ levels along with the ryanodine receptor (RyR). Consecutive to the Ras/MAPK pathway activation both proteins trigger intestinal stem cell (ISC) proliferation [48].
In lung adenocarcinoma the expression of TRPA1 and TRPV1 were upregulated by the hypoxia-inducible factor 1α (HIF1α) nuclear accumulation finally leading to cell proliferation. The study proposes that the signaling pathway TRPV1-Ca2+-nNOS-NO activates proliferation and metastasis in these tumoral cells [49]. In other instances, TRPV1 hinders cell proliferation. For instance, in skin carcinoma cells, pancreatic adenocarcinoma, or human colorectal carcinoma, TRPV1 suppresses posttranslational modification of the epidermal growth factor receptor (EGFR) associated with pro-proliferative pathways [50]. The results presented so far sustain the role of TRPA1 and TRPV1 in cancer cell proliferation but are irrelevant to the modulation of channels by ROS. Data obtained from studies unrelated to ion channels prove without doubt that ROS modulate cell proliferation [51]. A possible, so far missing from experimental evidence, link between TRPA1, ROS, and proliferation is very feasible because the major endogenous activators for TRPA1 in cancer are the products of oxidative stress. A brief schematic depiction of this hypothesis is shown in Figure 2.
In cancer cells, proliferation is intimately regulated by the apoptosis of neighboring cells through a process defined as “apoptosis-induced proliferation (AiP)” [52]. Moreover, apoptotic caspases mediate the generation of ROS for promoting AiP [53]. Also, TRP channels were indicated as important regulators of apoptosis, especially by their function in Ca2+ homeostasis [54]. The crosstalk TRP-ROS-apoptosis will be covered in the following subchapter.
5. Apoptosis Induced by Modulation of Redox TRP Channels
In breast cancer cells TRPA1 suppressed apoptosis upon stimulation by H2O2, whereas TRPA1 inhibition increased cell apoptosis in response to platinum-based drugs [55]. Interestingly, while inhibition of TRPA1 was determinant for apoptosis, channel silencing had no effect. These results prove that apoptosis induced by TRPA1 is related to Ca2+ signaling and to channel conducting properties of TRPA1. In pancreatic adenocarcinoma cells, the process is different. When TRPA1 was downregulated with siRNA, cells viability decreased, and possibly, the process of apoptosis augmented, as revealed by cell cycle analysis [56]. These demonstrated that TRPA1 expression, without activation is important in proliferation and apoptosis of pancreatic ductal adenocarcinoma cells. The expression of TRPA1 is enhanced by the activation of H2O2 and Nrf2 transcription factor, a process that takes place only in serum-starved conditions.
In glioma cells (GMB), the standard of care is surgery followed by external-beam radiation with concomitant temozolomide (TMZ) therapy. TMZ produces cell apoptosis and increases ROS production, but the treatment has short-term results due to chemoresistance. A recent study showed that activating TRPA1 in glioma cells leads to mitochondrial damage and cell apoptosis and decreases TMZ resistance. This process is based on increased ROS levels and apoptosis after the pretreatment of GMB cells with a TRPA1 agonist [57].
TRPV1 expression or functional modulation by agonists and antagonists affects apoptosis in many cancer cells, as carefully revised in [50]. The relation to ROS is mentioned in a couple of studies that monitored its production. For instance, in breast cancer cells a modulator of the TRPV1 channel decreased cell viability, and increased apoptosis and ROS production [58]. These results favor the idea that an allosteric modulator may be a better putative therapeutic agent than the classical TRPV1 activator capsaicin, which induces pain when applied in high and repeated doses. The agonists and the antagonists of TRPV1 raised the interest of many groups in the therapy of cancer. A study performed on different cancer cell lines showed that capsazepin, a TRPV1 antagonist, upregulates proapoptotic proteins p53 and Bax, which in turn augments TRAIL-induced apoptosis Finally, the study suggests that induction of TRAIL receptors is linked to the ROS–JNK–CHOP pathway [59].
Overexpression of TRPV4 in lung cancer cells promotes apoptosis by stimulation of the p38 MAPK pathways [60]. In squamous cell carcinoma (SSC) apoptosis was induced by oxidative stress with 24 hours application of H2O2. Data indicated that TRPM2 was activated and promoted apoptosis via caspases-3 and -9 in cultured SCC cells [61]. In irradiated human T-cell leukemia cells TRPM2 was denoted as a "death channel" because they promoted apoptosis and G2M cell cycle arrest through autophosphorylation and activation of Ca2+/calmodulin-dependent protein kinase II (CaMKII) [62]. Because ionizing radiation confers direct and indirect oxidative stress, the study hints toward crosstalk between TRPM2, ROS, and apoptosis.
Also, clostridium botulinum neurotoxin A (BTX) inhibits proliferation, growth and induces apoptosis in neuroblastoma and glioblastoma cells. The underlying mechanism may be the upregulation of mitochondrial oxidative stress and activation of the TRPM2 channel [63]. Together, these results indicate a clear liaison of TRP redox channels with apoptosis (Figure 3) upon modulation by ROS, intuitively via the Ca2+ entry, which is a clear activator of apoptosis.
5. TRP Channels Modulation by Oxidative Stress in Cancer Cell Migration
Many studies reported that ROS modulate cell motility, and different reports suggest identical effects of several TRP channels. The evidence of a physical link or a common signaling pathway is not clear, but we review and explain these putative interactions and condense them in Figure 4, which schematically illustrates a migratory cancer cell.
Some data stand that ROS inhibits cancer cell migration. A study performed on mice with malignant melanoma and several cell lines showed that the antioxidant N-acetylcysteine (NAC) increased the number of lymph nodes, as well as the proliferation and migration of tumor cells by boosting GSH concentrations and activating RhoA [64]. These results indicate that ROS inhibit cell proliferation and migration in these tumors. One may argue that melanocytes sequester ROS [65] preventing a significant increase of transcription factors sensitive to ROS, therefore, the effect of NAC is elusive. But other experiments support the idea and qualify ROS as friends rather than foes, reducing migration of tumoral cells. For instance, studies on liver cancer cells showed that H2O2 inhibited migration via the DLC1/RhoA signaling pathway [66] (pathway 1 in Figure 4). Also, in advanced gastric cancer cells, H2O2 inhibited migration by modulation of the PI3K/Akt signaling pathway [67].
The signaling pathway of cell migration by ROS comes from a recent study that evaluated cell-cell adhesion of head and neck squamous cell carcinoma [68]. The downstream effectors of ROS activation are Src kinases and STAT3 transcription factors. Along with protein kinase C (PKC), protein kinase A (PKA), and CaM-Kinase II, Src phosphorylates many TRP channels [69]. An early study showed that Src kinases phosphorylate TRPV4 during hypotonic stress [70] and recently, TRPV4 was associated with the migration of endometrial cancer or melanoma cells [71,72]. TRPV4 induced metastasis of melanoma by Src-cofilin intracellular pathway but, unexpectedly, this study shows that TRPV4 activates the phosphorylation of Src kinase. This is a particular situation when the channel activates the phosphorylation of other proteins downstream of the migratory pathway. Even more prone to phosphorylation of other proteins are TRPM7 and TRPM6 which contain their own kinase domains [73]. Indeed, cell migration was suppressed, and so was the phosphorylation of Src, Akt, and JNK (c-Jun N-terminal kinase) kinases when TRPM7 was inhibited in bladder cancer cells [74]. Possibly, the auto-phosphorylation of these channels contributes to the migration in specific tumoral environments [75], but the hypothesis is not yet supported by data. Two hypothetical models emerge from the data presented so far. One implies that ROS activates Src which in turn phosphorylates TRPV4 and promotes cell motility. The other may state that ROS activated TRPV4/M7 which phosphorylates other kinases that will mobilize proteins for cell migration and eventually metastases. In these cases, blocking channel phosphorylation will impact cell migration, but these theories have yet to be tested.
Less speculative is the relation of TRPM2 phosphorylation with ROS. H2O2 activates and induces phosphorylation of TRPM2. The effect is blocked by the non-specific inhibitor of Src family kinases PP2 [76]. Independent of phosphorylation, TRPM2 promotes migration of gastric cancer cells, through a pathway related to Ca2+ entry and regulation of the PTEN/Akt pathway [77]. In neuroblastoma, the same pathway is activated by TRPM2, along with the transcription factors HIF-1α1, E2F1, FOXM1, and CREB. Activation of TRPM2 increased the expression of α1, αv, β1, and β5 integrins, and consecutively migration and invasion of neuroblastoma [78].
Another member of the TRPM family, TRPM8 is phosphorylated by Src, and Src inhibition downregulates TRPM8-mediated responses [79]. Most importantly, Src- regulates the inflammatory pathway NF-κB signaling pathway and it is activated by 4-HNE. In this scenario, we may picture a better image of oxidative stress and inflammation, as we can add the putative roles of TRP channels in this signaling pathway.
6. Redox TRP Channels and Inflammation
The physiological role of inflammatory molecules in cancer has been established for a long time. Lately, the tumor microenvironment has been declared the main culprit for the inflammatory response in neoplasms [80]. In many cancers, the tumor is invaded by macrophages, T cells, myeloid cells, natural killers, and many others that secrete cytokines (e.g., TNF-α and IL- cytokines) and active mediators, including ROS [81]. The relationship between ROS and inflammation is again dual. On the one hand, ROS activated the NF-κB signal pathway and JAK-STAT signal pathway, which are positive regulators of cancer cells [82]. On the other hand, NF-κB decreased tumor development in some cancers. For instance, a model of hepatocellular carcinoma (HCC) induced in mice by the tumor initiator diethylnitrosamine (DEN), prevented NF-kB-induced hepatocarcinogenesis [83]. Also, in human epidermal cells both NF-κB and oncogenic Ras trigger cell-cycle arrest [84].
In the same line, activation of redox TRP channels may have dual roles in cancers associated with the inflammatory process because they activate different signaling pathways depending on the tumor type. Moreover, redox TRPs are differently modulated by anti-inflammatory compounds (Table 1). Therefore, a better insight into the use of anti-inflammatory drugs in cancer may come from the understanding of TRP channels modulation along with the effects on the tumorigenesis process.
From all members of the TRP superfamily, TRPA1 was named “the gatekeeper for inflammation” [95]. Activation of TRPA1 in glioma cells significantly decreased antioxidant expression and increased mitochondrial ROS production. The effects were reversed by pretreatment of glioma cells with the TRPA1 antagonist, as opposed to the temozolomide (TMZ) therapy, which had lower effects on the production of mitochondrial ROS and cell apoptosis [57]. The authors propose a scenario in which GBM cells are resistant to TMZ treatment, because of ROS production and consecutive TRPA1 activation. From this study, TRPA1 emerges as a tumor suppressor protein and its activation may be useful for better therapy. In agreement with these results, TRPA1 decreased cell proliferation and migration in pancreatic adenocarcinoma cells [56]. Contradictory results were obtained from breast cancer cells, where inhibition of TRPA1 reduced tumor growth [96]. Also, melanoma cells developed oxidative stress defense mechanisms upon TRPA1 activation [97].
The latest research on TRPA1 in inflammatory diseases and the signaling pathways were exhaustively reviewed in [98]. Although the authors skipped the relation of inflammation with cancer, they present several cellular pathways. For instance, they mention that activation of TRPA1 reduces the expression of pro-inflammatory cytokines through the nuclear factor kappa-B (NF-kB) signaling pathway, described in a previous study by Lee KI et. al. [99]. Nevertheless, cancer pain is associated with TRPA1 blockade by triterpenoids or riterpenoids ([100] and channel activation by ROS, but the process takes place in nociceptors and not in tumoral cells.
Another redox channel, TRPV1 activated the inflammatory pathway Nf-kB and STAT3. Convincing evidence comes from a study on knockout mice for TRPV1, which develop colon tumors, through a mechanism tightly related to inflammation [101]. The signaling pathways were activated along with proinflammatory (IL-1 and IL-6) and invasion factors (MMP9).
The oxidative stress by-product, 4-HNE is linked to inflammatory bowel disease (IBD) and colorectal cancer (CRC). High concentrations of 4-HNE were measured in DSS-induced IBD in C57BL/6 mice, through the activation of Toll-like receptor 4 (TLR4) signaling [102]. Besides impairing the quality of life, IBD increases the risk of CRC, associated also with serious oxidative stress and high 4-HNE levels [103]. Then again, redox TRP channels may be associated with 4-HNE production and inflammation in CRC, working either as oncogenic factors or as tumor suppressors.
For instance, the expression level of TRPC5 in colon cancer patients correlates with poor prognosis and metastasis, via the HIF-1α/Twist signaling pathway [104]. Also, we know from studies on hippocampal neurons that a significant reduction in 4-HNE concentration is obtained when TRPC5 is inhibited with a specific antagonist [105]. Putting together these data, we may speculate that inhibition of 4-HNE through a specific TRPV5 antagonist could reduce metastasis of colon cancer cells.
Many studies indicated TRPV1 as a tumor suppressor of CRC, therefore its activation may be a feasible approach for this neoplasm [106]. We also know that 4-HNE inhibited TRPV1 Ca2+ current in HEK transfected cells [36], encouraging the use of the antagonist capsazepine, as a pharmacological tool for CRC.
A type of CRC, so-called colitis-associated cancer (CAC), is one of the complications of long-standing inflammatory bowel disease, characterized by the massive production of ROS and inflammatory cytokines that activate signaling pathways, such as NF-κB, STAT3, p38 MAPK, and Wnt/β-catenin, which in turn modulate the tumorigenic process [107]. A previous study investigated the pathogenesis of CAC in mice [108]. It reported that TRPV4 expression is upregulated in CAC and contributes to the progression of colonic inflammation and vascular permeability. The authors propose inhibition of TRPV4 as a therapeutic strategy for CAC, which may decrease cytokines production and tumorigenesis.
Previous studies measured high expression levels of TRPA1 and TRPM2 in mice with induced CRC and a direct relationship with expression levels of TNF-α and Casp-3 [109]. The authors propose the extract of pomegranate as a modulator of these proteins.
Oxidative stress and 4-HNE are the driving forces for esophageal squamous cell carcinoma (ESCC). In a cohort of 3 non-malignant esophageal epithelial tissues, 11 esophageal carcinoma in situ tissues, and 57 ESCC tissues from patients in the Chaoshan area, high levels of 4-HNE were linked to the degree of inflammation in ESCC [110]. In samples from patients with ESCC, TRPM7 was found as an independent prognostic factor of good postoperative survival [111].
Contrary to these results, a retrospective study of oropharyngeal squamous cell carcinoma (SCC) in 102 patients shows the presence of 4-HNE in tumor cells, but a higher HNE-immunopositivity in non-malignant cells that were localized close to the carcinoma. The study concludes that 4-HNE is a bioactive marker of OS but with a defensive, anti-cancer potential of the surrounding non-malignant cells [112]. Similarly, TRPM2 has dual roles in SCC cell lines [61]. On the one hand, high levels of plasma membrane TRPM2 expression protect against the early stages of cell growth. On the other hand, increased levels of nuclear TRPM2 during the advanced stages of cancer may stimulate tumorigenesis. Modulation of TRPM2 by agonists and antagonists would be a rather tedious mission in this case.
7. Conclusions
TRP redox channels are expressed in many cancer patients and tumoral cells, where they have dual roles as suppressors or initiators of the tumoral process. Because many cancer types show resistance to radio- or chemotherapy, the main objective of studies on TRP proteins is the finding of new targets and the understanding of signaling pathways that will meet the need for new therapeutic agents. In this context, the draggability of TRP channels and their modulation by compounds that interfere with tumorigenesis, such as ROS, is appealing to both the scientific community and pharmaceutical companies. Also, the agonists and antagonists of TRP, often natural compounds that avoid radioactive flux could be a benefit. We reviewed here studies that capture the relationship between TRP and ROS, TRP and inflammation and we try to make the link that explains the crosstalk between these actors in the tumorigenic process.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jardin, I.; Lopez, J.J.; Diez, R.; Sanchez-Collado, J.; Cantonero, C.; Albarran, L.; Woodard, G.E.; Redondo, P.C.; Salido, G.M.; Smani, T.; et al. TRPs in Pain Sensation. Frontiers in physiology 2017, 8, 392. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome biology 2011, 12, 218. [Google Scholar] [CrossRef] [PubMed]
- Vennekens, R.; Hoenderop, J.G.; Prenen, J.; Stuiver, M.; Willems, P.H.; Droogmans, G.; Nilius, B.; Bindels, R.J. Permeation and gating properties of the novel epithelial Ca(2+) channel. The Journal of biological chemistry 2000, 275, 3963–3969. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Prenen, J.; Janssens, A.; Owsianik, G.; Wang, C.; Zhu, M.X.; Voets, T. The selectivity filter of the cation channel TRPM4. The Journal of biological chemistry 2005, 280, 22899–22906. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, R.; Mori, Y. Transient receptor potential (TRP) channels: Biosensors for redox environmental stimuli and cellular status. Free radical biology & medicine 2020, 146, 36–44. [Google Scholar] [CrossRef]
- Takahashi, N.; Mori, Y. TRP Channels as Sensors and Signal Integrators of Redox Status Changes. Frontiers in pharmacology 2011, 2, 58. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Inoue, R.; Morii, T.; Takahashi, N.; Yamamoto, S.; Hara, Y.; Tominaga, M.; Shimizu, S.; Sato, Y.; Mori, Y. Nitric oxide activates TRP channels by cysteine S-nitrosylation. Nature chemical biology 2006, 2, 596–607. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, N.; Kurokawa, T.; Fujiwara, K.; Polat, O.K.; Badr, H.; Takahashi, N.; Mori, Y. Functional and Structural Divergence in Human TRPV1 Channel Subunits by Oxidative Cysteine Modification. The Journal of biological chemistry 2016, 291, 4197–4210. [Google Scholar] [CrossRef]
- Vellino, S.; Oddou, C.; Rivier, P.; Boyault, C.; Hiriart-Bryant, E.; Kraut, A.; Martin, R.; Coute, Y.; Knolker, H.J.; Valverde, M.A.; et al. Cross-talk between the calcium channel TRPV4 and reactive oxygen species interlocks adhesive and degradative functions of invadosomes. The Journal of cell biology 2021, 220. [Google Scholar] [CrossRef]
- Talavera, K.; Startek, J.B.; Alvarez-Collazo, J.; Boonen, B.; Alpizar, Y.A.; Sanchez, A.; Naert, R.; Nilius, B. Mammalian Transient Receptor Potential TRPA1 Channels: From Structure to Disease. Physiological reviews 2020, 100, 725–803. [Google Scholar] [CrossRef]
- Ding, R.; Yin, Y.L.; Jiang, L.H. Reactive Oxygen Species-Induced TRPM2-Mediated Ca(2+) Signalling in Endothelial Cells. Antioxidants (Basel) 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Malko, P.; Jiang, L.H. TRPM2 channel-mediated cell death: An important mechanism linking oxidative stress-inducing pathological factors to associated pathological conditions. Redox biology 2020, 37, 101755. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Murayama, T.; Tashiro, M.; Sakurai, T.; Konishi, M. Mg(2+)- and ATP-dependent inhibition of transient receptor potential melastatin 7 by oxidative stress. Free radical biology & medicine 2014, 72, 257–266. [Google Scholar] [CrossRef]
- Inoue, H.; Murayama, T.; Kobayashi, T.; Konishi, M.; Yokoyama, U. The zinc-binding motif of TRPM7 acts as an oxidative stress sensor to regulate its channel activity. The Journal of general physiology 2021, 153. [Google Scholar] [CrossRef] [PubMed]
- Fricke, T.C.; Echtermeyer, F.; Zielke, J.; de la Roche, J.; Filipovic, M.R.; Claverol, S.; Herzog, C.; Tominaga, M.; Pumroy, R.A.; Moiseenkova-Bell, V.Y.; et al. Oxidation of methionine residues activates the high-threshold heat-sensitive ion channel TRPV2. Proceedings of the National Academy of Sciences of the United States of America 2019, 116, 24359–24365. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative medicine and cellular longevity 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS Are Good. Trends in plant science 2017, 22, 11–19. [Google Scholar] [CrossRef]
- Sinenko, S.A.; Starkova, T.Y.; Kuzmin, A.A.; Tomilin, A.N. Physiological Signaling Functions of Reactive Oxygen Species in Stem Cells: From Flies to Man. Frontiers in cell and developmental biology 2021, 9, 714370. [Google Scholar] [CrossRef]
- Canton, M.; Sanchez-Rodriguez, R.; Spera, I.; Venegas, F.C.; Favia, M.; Viola, A.; Castegna, A. Reactive Oxygen Species in Macrophages: Sources and Targets. Frontiers in immunology 2021, 12, 734229. [Google Scholar] [CrossRef]
- Nugud, A.; Sandeep, D.; El-Serafi, A.T. Two faces of the coin: Minireview for dissecting the role of reactive oxygen species in stem cell potency and lineage commitment. Journal of advanced research 2018, 14, 73–79. [Google Scholar] [CrossRef]
- Vieira, H.L.; Alves, P.M.; Vercelli, A. Modulation of neuronal stem cell differentiation by hypoxia and reactive oxygen species. Progress in neurobiology 2011, 93, 444–455. [Google Scholar] [CrossRef] [PubMed]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free radical biology & medicine 2010, 48, 749–762. [Google Scholar] [CrossRef]
- Catala, A.; Diaz, M. Editorial: Impact of Lipid Peroxidation on the Physiology and Pathophysiology of Cell Membranes. Frontiers in physiology 2016, 7, 423. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Lei, G.; Zhuang, L.; Gan, B. Targeting ferroptosis as a vulnerability in cancer. Nature reviews. Cancer 2022, 22, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Green, M.; Choi, J.E.; Gijon, M.; Kennedy, P.D.; Johnson, J.K.; Liao, P.; Lang, X.; Kryczek, I.; Sell, A.; et al. CD8(+) T cells regulate tumour ferroptosis during cancer immunotherapy. Nature 2019, 569, 270–274. [Google Scholar] [CrossRef]
- Li, M.; Zheng, J.; Wu, T.; He, Y.; Guo, J.; Xu, J.; Gao, C.; Qu, S.; Zhang, Q.; Zhao, J.; et al. Activation of TRPV4 Induces Exocytosis and Ferroptosis in Human Melanoma Cells. International journal of molecular sciences 2022, 23. [Google Scholar] [CrossRef]
- Yahya, F.; Mohd Bakri, M.; Hossain, M.Z.; Syed Abdul Rahman, S.N.; Mohammed Alabsi, A.; Ramanathan, A. Combination Treatment of TRPV4 Agonist with Cisplatin Promotes Vessel Normalization in an Animal Model of Oral Squamous Cell Carcinoma. Medicina (Kaunas) 2022, 58. [Google Scholar] [CrossRef]
- Fujii, S.; Tajiri, Y.; Hasegawa, K.; Matsumoto, S.; Yoshimoto, R.U.; Wada, H.; Kishida, S.; Kido, M.A.; Yoshikawa, H.; Ozeki, S.; et al. The TRPV4-AKT axis promotes oral squamous cell carcinoma cell proliferation via CaMKII activation. Laboratory investigation; a journal of technical methods and pathology 2020, 100, 311–323. [Google Scholar] [CrossRef]
- Kang, D.H.; Park, C.K.; Chung, C.; Oh, I.J.; Kim, Y.C.; Park, D.; Kim, J.; Kwon, G.C.; Kwon, I.; Sun, P.; et al. Baseline Serum Interleukin-6 Levels Predict the Response of Patients with Advanced Non-small Cell Lung Cancer to PD-1/PD-L1 Inhibitors. Immune network 2020, 20, e27. [Google Scholar] [CrossRef]
- Shang, G.S.; Liu, L.; Qin, Y.W. IL-6 and TNF-alpha promote metastasis of lung cancer by inducing epithelial-mesenchymal transition. Oncology letters 2017, 13, 4657–4660. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Li, S.; Yang, Y.; Bai, Z. Interleukin-6 promotes ferroptosis in bronchial epithelial cells by inducing reactive oxygen species-dependent lipid peroxidation and disrupting iron homeostasis. Bioengineered 2021, 12, 5279–5288. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Jin, S.; Zhang, Z.; Ma, H.; Yang, X. Interleukin-6 facilitates tumor progression by inducing ferroptosis resistance in head and neck squamous cell carcinoma. Cancer letters 2022, 527, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Uz, U.; Eskiizmir, G. Association Between Interleukin-6 and Head and Neck Squamous Cell Carcinoma: A Systematic Review. Clinical and experimental otorhinolaryngology 2021, 14, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Gasparovic, A.C.; Milkovic, L.; Sunjic, S.B.; Zarkovic, N. Cancer growth regulation by 4-hydroxynonenal. Free radical biology & medicine 2017, 111, 226–234. [Google Scholar] [CrossRef]
- DelloStritto, D.J.; Sinharoy, P.; Connell, P.J.; Fahmy, J.N.; Cappelli, H.C.; Thodeti, C.K.; Geldenhuys, W.J.; Damron, D.S.; Bratz, I.N. 4-Hydroxynonenal dependent alteration of TRPV1-mediated coronary microvascular signaling. Free radical biology & medicine 2016, 101, 10–19. [Google Scholar] [CrossRef]
- Di, Y.; Xu, T.; Tian, Y.; Ma, T.; Qu, D.; Wang, Y.; Lin, Y.; Bao, D.; Yu, L.; Liu, S.; et al. Ursolic acid protects against cisplatin-induced ototoxicity by inhibiting oxidative stress and TRPV1-mediated Ca2+-signaling. International journal of molecular medicine 2020, 46, 806–816. [Google Scholar] [CrossRef]
- Rawat, P.S.; Jaiswal, A.; Khurana, A.; Bhatti, J.S.; Navik, U. Doxorubicin-induced cardiotoxicity: An update on the molecular mechanism and novel therapeutic strategies for effective management. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie 2021, 139, 111708. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, M.; Liu, J.; Ye, J.; Jiang, H.; Xu, Y.; Ye, D.; Wan, J. Inhibition of TRPA1 Attenuates Doxorubicin-Induced Acute Cardiotoxicity by Suppressing Oxidative Stress, the Inflammatory Response, and Endoplasmic Reticulum Stress. Oxidative medicine and cellular longevity 2018, 2018, 5179468. [Google Scholar] [CrossRef]
- Tai, Y.K.; Chan, K.K.W.; Fong, C.H.H.; Ramanan, S.; Yap, J.L.Y.; Yin, J.N.; Yip, Y.S.; Tan, W.R.; Koh, A.P.F.; Tan, N.S.; et al. Modulated TRPC1 Expression Predicts Sensitivity of Breast Cancer to Doxorubicin and Magnetic Field Therapy: Segue Towards a Precision Medicine Approach. Frontiers in oncology 2021, 11, 783803. [Google Scholar] [CrossRef]
- Nishiyama, K.; Numaga-Tomita, T.; Fujimoto, Y.; Tanaka, T.; Toyama, C.; Nishimura, A.; Yamashita, T.; Matsunaga, N.; Koyanagi, S.; Azuma, Y.T.; et al. Ibudilast attenuates doxorubicin-induced cytotoxicity by suppressing formation of TRPC3 channel and NADPH oxidase 2 protein complexes. British journal of pharmacology 2019, 176, 3723–3738. [Google Scholar] [CrossRef]
- Hopkins, M.M.; Feng, X.; Liu, M.; Parker, L.P.; Koh, D.W. Inhibition of the transient receptor potential melastatin-2 channel causes increased DNA damage and decreased proliferation in breast adenocarcinoma cells. International journal of oncology 2015, 46, 2267–2276. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.J.; Bao, L.; Keefer, K.; Shanmughapriya, S.; Chen, L.; Lee, J.; Wang, J.; Zhang, X.Q.; Hirschler-Laszkiewicz, I.; Merali, S.; et al. Transient receptor potential ion channel TRPM2 promotes AML proliferation and survival through modulation of mitochondrial function, ROS, and autophagy. Cell death & disease 2020, 11, 247. [Google Scholar] [CrossRef]
- Bao, L.; Festa, F.; Freet, C.S.; Lee, J.P.; Hirschler-Laszkiewicz, I.M.; Chen, S.J.; Keefer, K.A.; Wang, H.G.; Patterson, A.D.; Cheung, J.Y.; et al. The Human Transient Receptor Potential Melastatin 2 Ion Channel Modulates ROS Through Nrf2. Scientific reports 2019, 9, 14132. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Bu, F.; Ma, S.; Cananzi, F.; Zhao, Y.; Xiao, M.; Min, L.; Luo, C. The Janus-faced role of TRPM2-S in retroperitoneal liposarcoma via increasing ROS levels. Cell communication and signaling : CCS 2022, 20, 128. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Wei, X.; Wang, M.M.; Liu, Y.; Sui, Z.; Wang, X.; Zhang, Y.; Fei, Y.H.; Jiang, Y.; Lu, C.; et al. Stimulating TRPM7 suppresses cancer cell proliferation and metastasis by inhibiting autophagy. Cancer letters 2022, 525, 179–197. [Google Scholar] [CrossRef] [PubMed]
- Berrout, J.; Kyriakopoulou, E.; Moparthi, L.; Hogea, A.S.; Berrout, L.; Ivan, C.; Lorger, M.; Boyle, J.; Peers, C.; Muench, S.; et al. TRPA1-FGFR2 binding event is a regulatory oncogenic driver modulated by miRNA-142-3p. Nature communications 2017, 8, 947. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Luo, J.; He, L.; Montell, C.; Perrimon, N. Oxidative stress induces stem cell proliferation via TRPA1/RyR-mediated Ca(2+) signaling in the Drosophila midgut. eLife 2017, 6. [Google Scholar] [CrossRef]
- Nie, Y.; Feng, F.; Luo, W.; Sanders, A.J.; Zhang, Y.; Liang, J.; Chen, C.; Feng, W.; Gu, W.; Liao, W.; et al. Overexpressed transient receptor potential vanilloid 1 (TRPV1) in lung adenocarcinoma harbours a new opportunity for therapeutic targeting. Cancer gene therapy 2022, 29, 1405–1417. [Google Scholar] [CrossRef]
- Li, L.; Chen, C.; Chiang, C.; Xiao, T.; Chen, Y.; Zhao, Y.; Zheng, D. The Impact of TRPV1 on Cancer Pathogenesis and Therapy: A Systematic Review. International journal of biological sciences 2021, 17, 2034–2049. [Google Scholar] [CrossRef]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: the bright side of the moon. Experimental & molecular medicine 2020, 52, 192–203. [Google Scholar] [CrossRef]
- Moreno-Celis, U.; Garcia-Gasca, T.; Mejia, C. Apoptosis-Induced Compensatory Proliferation in Cancer. In Metastasis, Sergi, C.M., Ed.; Brisbane (AU), 2022.
- Diwanji, N.; Bergmann, A. An unexpected friend - ROS in apoptosis-induced compensatory proliferation: Implications for regeneration and cancer. Seminars in cell & developmental biology 2018, 80, 74–82. [Google Scholar] [CrossRef]
- Yang, D.; Kim, J. Emerging role of transient receptor potential (TRP) channels in cancer progression. BMB reports 2020, 53, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Chen, H.Y.; Harris, I.S.; Stover, D.G.; Selfors, L.M.; Bronson, R.T.; Deraedt, T.; Cichowski, K.; Welm, A.L.; Mori, Y.; et al. Cancer Cells Co-opt the Neuronal Redox-Sensing Channel TRPA1 to Promote Oxidative-Stress Tolerance. Cancer cell 2018, 33, 985–1003. [Google Scholar] [CrossRef] [PubMed]
- Cojocaru, F.; Selescu, T.; Domocos, D.; Marutescu, L.; Chiritoiu, G.; Chelaru, N.R.; Dima, S.; Mihailescu, D.; Babes, A.; Cucu, D. Publisher Correction: Functional expression of the transient receptor potential ankyrin type 1 channel in pancreatic adenocarcinoma cells. Scientific reports 2021, 11, 8853. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, C.; Hu, H.; Zhang, B. Activated TRPA1 plays a therapeutic role in TMZ resistance in glioblastoma by altering mitochondrial dynamics. BMC molecular and cell biology 2022, 23, 38. [Google Scholar] [CrossRef]
- Naziroglu, M.; Cig, B.; Blum, W.; Vizler, C.; Buhala, A.; Marton, A.; Katona, R.; Josvay, K.; Schwaller, B.; Olah, Z.; et al. Targeting breast cancer cells by MRS1477, a positive allosteric modulator of TRPV1 channels. PloS one 2017, 12, e0179950. [Google Scholar] [CrossRef]
- Sung, B.; Prasad, S.; Ravindran, J.; Yadav, V.R.; Aggarwal, B.B. Capsazepine, a TRPV1 antagonist, sensitizes colorectal cancer cells to apoptosis by TRAIL through ROS-JNK-CHOP-mediated upregulation of death receptors. Free radical biology & medicine 2012, 53, 1977–1987. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, J.; Liu, X. TRPV4 induces apoptosis via p38 MAPK in human lung cancer cells. Brazilian journal of medical and biological research = Revista brasileira de pesquisas medicas e biologicas 2021, 54, e10867. [Google Scholar] [CrossRef]
- Zhao, L.Y.; Xu, W.L.; Xu, Z.Q.; Qi, C.; Li, Y.; Cheng, J.; Liu, L.K.; Wu, Y.N.; Gao, J.; Ye, J.H. The overexpressed functional transient receptor potential channel TRPM2 in oral squamous cell carcinoma. Scientific reports 2016, 6, 38471. [Google Scholar] [CrossRef]
- Klumpp, D.; Misovic, M.; Szteyn, K.; Shumilina, E.; Rudner, J.; Huber, S.M. Targeting TRPM2 Channels Impairs Radiation-Induced Cell Cycle Arrest and Fosters Cell Death of T Cell Leukemia Cells in a Bcl-2-Dependent Manner. Oxidative medicine and cellular longevity 2016, 2016, 8026702. [Google Scholar] [CrossRef] [PubMed]
- Akpinar, O.; Ozsimsek, A.; Guzel, M.; Naziroglu, M. Clostridium botulinum neurotoxin A induces apoptosis and mitochondrial oxidative stress via activation of TRPM2 channel signaling pathway in neuroblastoma and glioblastoma tumor cells. Journal of receptor and signal transduction research 2020, 40, 620–632. [Google Scholar] [CrossRef] [PubMed]
- Le Gal, K.; Ibrahim, M.X.; Wiel, C.; Sayin, V.I.; Akula, M.K.; Karlsson, C.; Dalin, M.G.; Akyurek, L.M.; Lindahl, P.; Nilsson, J.; et al. Antioxidants can increase melanoma metastasis in mice. Science translational medicine 2015, 7, 308re308. [Google Scholar] [CrossRef] [PubMed]
- Meyskens, F.L., Jr.; McNulty, S.E.; Buckmeier, J.A.; Tohidian, N.B.; Spillane, T.J.; Kahlon, R.S.; Gonzalez, R.I. Aberrant redox regulation in human metastatic melanoma cells compared to normal melanocytes. Free radical biology & medicine 2001, 31, 799–808. [Google Scholar] [CrossRef]
- Ma, L.; Zhu, W.Z.; Liu, T.T.; Fu, H.L.; Liu, Z.J.; Yang, B.W.; Song, T.Y.; Li, G.R. H2O2 inhibits proliferation and mediates suppression of migration via DLC1/RhoA signaling in cancer cells. Asian Pacific journal of cancer prevention : APJCP 2015, 16, 1637–1642. [Google Scholar] [CrossRef] [PubMed]
- Savino, L.; Di Marcantonio, M.C.; Moscatello, C.; Cotellese, R.; Centurione, L.; Muraro, R.; Aceto, G.M.; Mincione, G. Effects of H(2)O(2) Treatment Combined With PI3K Inhibitor and MEK Inhibitor in AGS Cells: Oxidative Stress Outcomes in a Model of Gastric Cancer. Frontiers in oncology 2022, 12, 860760. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Hsu, J.Y.; Chu, C.T.; Chang, Y.W.; Fan, J.R.; Yang, M.H.; Chen, H.C. Loss of cell-cell adhesion triggers cell migration through Rac1-dependent ROS generation. Life science alliance 2023, 6. [Google Scholar] [CrossRef]
- Yao, X.; Kwan, H.Y.; Huang, Y. Regulation of TRP channels by phosphorylation. Neuro-Signals 2005, 14, 273–280. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, H.; Tian, W.; Yoshida, K.; Roullet, J.B.; Cohen, D.M. Regulation of a transient receptor potential (TRP) channel by tyrosine phosphorylation. SRC family kinase-dependent tyrosine phosphorylation of TRPV4 on TYR-253 mediates its response to hypotonic stress. The Journal of biological chemistry 2003, 278, 11520–11527. [Google Scholar] [CrossRef]
- Li, X.; Cheng, Y.; Wang, Z.; Zhou, J.; Jia, Y.; He, X.; Zhao, L.; Dong, Y.; Fan, Y.; Yang, X.; et al. Calcium and TRPV4 promote metastasis by regulating cytoskeleton through the RhoA/ROCK1 pathway in endometrial cancer. Cell death & disease 2020, 11, 1009. [Google Scholar] [CrossRef]
- Huang, S.; Yu, S.; Deng, R.; Liu, H.; Ding, Y.; Sun, Y.; Chen, W.; Wang, A.; Wei, Z.; Lu, Y. TRPV4 Promotes Metastasis in Melanoma by Regulating Cell Motility through Cytoskeletal Rearrangement. International journal of molecular sciences 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Cai, N.; Bai, Z.; Nanda, V.; Runnels, L.W. Mass Spectrometric Analysis of TRPM6 and TRPM7 Phosphorylation Reveals Regulatory Mechanisms of the Channel-Kinases. Scientific reports 2017, 7, 42739. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Chun, S.Y.; Kim, B.; Yoon, B.H.; Lee, J.N.; Kim, B.S.; Yoo, E.S.; Lee, S.; Song, P.H.; Kwon, T.G.; et al. Knockdown of TRPM7 prevents tumor growth, migration, and invasion through the Src, Akt, and JNK pathway in bladder cancer. BMC urology 2020, 20, 145. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.; Middelbeek, J.; Morrice, N.A.; Figdor, C.G.; Lasonder, E.; van Leeuwen, F.N. Massive autophosphorylation of the Ser/Thr-rich domain controls protein kinase activity of TRPM6 and TRPM7. PloS one 2008, 3, e1876. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Tong, Q.; Conrad, K.; Wozney, J.; Cheung, J.Y.; Miller, B.A. Regulation of TRP channel TRPM2 by the tyrosine phosphatase PTPL1. American journal of physiology. Cell physiology 2007, 292, C1746–C1758. [Google Scholar] [CrossRef] [PubMed]
- Almasi, S.; Sterea, A.M.; Fernando, W.; Clements, D.R.; Marcato, P.; Hoskin, D.W.; Gujar, S.; El Hiani, Y. TRPM2 ion channel promotes gastric cancer migration, invasion and tumor growth through the AKT signaling pathway. Scientific reports 2019, 9, 4182. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Festa, F.; Hirschler-Laszkiewicz, I.; Keefer, K.; Wang, H.G.; Cheung, J.Y.; Miller, B.A. The human ion channel TRPM2 modulates migration and invasion in neuroblastoma through regulation of integrin expression. Scientific reports 2022, 12, 20544. [Google Scholar] [CrossRef]
- Manolache, A.; Selescu, T.; Maier, G.L.; Mentel, M.; Ionescu, A.E.; Neacsu, C.; Babes, A.; Szedlacsek, S.E. Regulation of TRPM8 channel activity by Src-mediated tyrosine phosphorylation. Journal of cellular physiology 2020, 235, 5192–5203. [Google Scholar] [CrossRef]
- Arfin, S.; Jha, N.K.; Jha, S.K.; Kesari, K.K.; Ruokolainen, J.; Roychoudhury, S.; Rathi, B.; Kumar, D. Oxidative Stress in Cancer Cell Metabolism. Antioxidants (Basel) 2021, 10. [Google Scholar] [CrossRef]
- Tan, Z.; Xue, H.; Sun, Y.; Zhang, C.; Song, Y.; Qi, Y. The Role of Tumor Inflammatory Microenvironment in Lung Cancer. Frontiers in pharmacology 2021, 12, 688625. [Google Scholar] [CrossRef]
- Xia, Y.; Shen, S.; Verma, I.M. NF-kappaB, an active player in human cancers. Cancer immunology research 2014, 2, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Kamata, H.; Luo, J.L.; Leffert, H.; Karin, M. IKKbeta couples hepatocyte death to cytokine-driven compensatory proliferation that promotes chemical hepatocarcinogenesis. Cell 2005, 121, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Dajee, M.; Lazarov, M.; Zhang, J.Y.; Cai, T.; Green, C.L.; Russell, A.J.; Marinkovich, M.P.; Tao, S.; Lin, Q.; Kubo, Y.; et al. NF-kappaB blockade and oncogenic Ras trigger invasive human epidermal neoplasia. Nature 2003, 421, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Ralph, S.J.; Pritchard, R.; Rodriguez-Enriquez, S.; Moreno-Sanchez, R.; Ralph, R.K. Hitting the Bull's-Eye in Metastatic Cancers-NSAIDs Elevate ROS in Mitochondria, Inducing Malignant Cell Death. Pharmaceuticals (Basel) 2015, 8, 62–106. [Google Scholar] [CrossRef] [PubMed]
- Sevinç, S.K.; Orun, O.; Tiber, P.M.; Çıkla-Süzgün, P.; Küçükgüzel, Ş.G. Anti-Cancer Acitivity of Etodolac and Its Derivatives on Prostate and Colorectal Cancer Cell Lines. Proceedings 2018, 2, 1573. [Google Scholar]
- Tsagareli, M.G.; Nozadze, I.; Tsiklauri, N.; Gurtskaia, G. Non-steroidal anti-inflammatory drugs attenuate agonist-evoked activation of transient receptor potential channels. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie 2018, 97, 745–751. [Google Scholar] [CrossRef]
- Muller, C.; Morales, P.; Reggio, P.H. Cannabinoid Ligands Targeting TRP Channels. Frontiers in molecular neuroscience 2018, 11, 487. [Google Scholar] [CrossRef]
- Li, D.; Ilnytskyy, Y.; Ghasemi Gojani, E.; Kovalchuk, O.; Kovalchuk, I. Analysis of Anti-Cancer and Anti-Inflammatory Properties of 25 High-THC Cannabis Extracts. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Dogru, A.; Naziroglu, M.; Cig, B. Modulator role of infliximab and methotrexate through the transient receptor potential melastatin 2 (TRPM2) channel in neutrophils of patients with rheumatoid arthritis: a pilot study. Archives of medical science : AMS 2019, 15, 1415–1424. [Google Scholar] [CrossRef]
- Sharan, M.; Jha, M.; Chandel, R.; Syeda, S.; Mathur, R.; Jha, N.K.; Jha, S.K.; Goel, H.; Shrivastava, A.; Chauhan, S.; et al. Demethylation of CADM1 and SOCS1 using capsaicin in cervical cancer cell line. Naunyn-Schmiedeberg's archives of pharmacology 2023, 396, 649–657. [Google Scholar] [CrossRef]
- Pawar, J.S.; Mustafa, S.; Ghosh, I. Chrysin and Capsaicin induces premature senescence and apoptosis via mitochondrial dysfunction and p53 elevation in Cervical cancer cells. Saudi journal of biological sciences 2022, 29, 3838–3847. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Dong, J.; Tian, W.; Qiao, S.; Wang, H. Role of TRPV1 ion channel in cervical squamous cell carcinoma genesis. Frontiers in molecular biosciences 2022, 9, 980262. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.H.; Jung, S.H.; Sethi, G.; Ahn, K.S. Pleiotropic Pharmacological Actions of Capsazepine, a Synthetic Analogue of Capsaicin, against Various Cancers and Inflammatory Diseases. Molecules 2019, 24. [Google Scholar] [CrossRef] [PubMed]
- Bautista, D.M.; Pellegrino, M.; Tsunozaki, M. TRPA1: A gatekeeper for inflammation. Annual review of physiology 2013, 75, 181–200. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lin, C.; Liu, Z.; Sun, Y.; Chen, M.; Guo, Y.; Liu, W.; Zhang, C.; Chen, W.; Sun, J.; et al. Cancer cells co-opt nociceptive nerves to thrive in nutrient-poor environments and upon nutrient-starvation therapies. Cell metabolism 2022, 34, 1999–2017. [Google Scholar] [CrossRef] [PubMed]
- De Logu, F.; Souza Monteiro de Araujo, D.; Ugolini, F.; Iannone, L.F.; Vannucchi, M.; Portelli, F.; Landini, L.; Titiz, M.; De Giorgi, V.; Geppetti, P.; et al. The TRPA1 Channel Amplifies the Oxidative Stress Signal in Melanoma. Cells 2021, 10. [Google Scholar] [CrossRef]
- Yao, K.; Dou, B.; Zhang, Y.; Chen, Z.; Li, Y.; Fan, Z.; Ma, Y.; Du, S.; Wang, J.; Xu, Z.; et al. Inflammation-the role of TRPA1 channel. Frontiers in physiology 2023, 14, 1093925. [Google Scholar] [CrossRef]
- Lee, K.I.; Lee, H.T.; Lin, H.C.; Tsay, H.J.; Tsai, F.C.; Shyue, S.K.; Lee, T.S. Role of transient receptor potential ankyrin 1 channels in Alzheimer's disease. Journal of neuroinflammation 2016, 13, 92. [Google Scholar] [CrossRef]
- Maki-Opas, I.; Hamalainen, M.; Moilanen, L.J.; Haavikko, R.; Ahonen, T.J.; Alakurtti, S.; Moreira, V.M.; Muraki, K.; Yli-Kauhaluoma, J.; Moilanen, E. Pyrazine-Fused Triterpenoids Block the TRPA1 Ion Channel in Vitro and Inhibit TRPA1-Mediated Acute Inflammation in Vivo. ACS chemical neuroscience 2019, 10, 2848–2857. [Google Scholar] [CrossRef]
- Vinuesa, A.G.; Sancho, R.; Garcia-Limones, C.; Behrens, A.; ten Dijke, P.; Calzado, M.A.; Munoz, E. Vanilloid receptor-1 regulates neurogenic inflammation in colon and protects mice from colon cancer. Cancer research 2012, 72, 1705–1716. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, W.; Yang, H.; Shao, D.; Zhao, X.; Zhang, G. Intraperitoneal injection of 4-hydroxynonenal (4-HNE), a lipid peroxidation product, exacerbates colonic inflammation through activation of Toll-like receptor 4 signaling. Free radical biology & medicine 2019, 131, 237–242. [Google Scholar] [CrossRef]
- Janion, K.; Szczepanska, E.; Nowakowska-Zajdel, E.; Walkiewicz, K.; Strzelczyk, J. Lipid peroxidation and total oxidant/antioxidant status in colorectal cancer patients. Journal of biological regulators and homeostatic agents 2020, 34, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhu, Y.; Dong, Y.; Zhang, P.; Han, X.; Jin, J.; Ma, X. Overexpression of TrpC5 promotes tumor metastasis via the HIF-1alpha-Twist signaling pathway in colon cancer. Clin Sci (Lond) 2017, 131, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Park, M.K.; Choi, B.Y.; Kho, A.R.; Lee, S.H.; Hong, D.K.; Jeong, J.H.; Kang, D.H.; Kang, B.S.; Suh, S.W. Effects of Transient Receptor Potential Cation 5 (TRPC5) Inhibitor, NU6027, on Hippocampal Neuronal Death after Traumatic Brain Injury. International journal of molecular sciences 2020, 21. [Google Scholar] [CrossRef]
- Chen, Y.; Li, J.; Jin, L.; Lei, K.; Liu, H.; Yang, Y. Fibulin-5 contributes to colorectal cancer cell apoptosis via the ROS/MAPK and Akt signal pathways by downregulating transient receptor potential cation channel subfamily V member 1. Journal of cellular biochemistry 2019, 120, 17838–17846. [Google Scholar] [CrossRef]
- Wang, Z.; Li, S.; Cao, Y.; Tian, X.; Zeng, R.; Liao, D.F.; Cao, D. Oxidative Stress and Carbonyl Lesions in Ulcerative Colitis and Associated Colorectal Cancer. Oxidative medicine and cellular longevity 2016, 2016, 9875298. [Google Scholar] [CrossRef]
- Matsumoto, K.; Deguchi, A.; Motoyoshi, A.; Morita, A.; Maebashi, U.; Nakamoto, T.; Kawanishi, S.; Sueyoshi, M.; Nishimura, K.; Takata, K.; et al. Role of transient receptor potential vanilloid subtype 4 in the regulation of azoymethane/dextran sulphate sodium-induced colitis-associated cancer in mice. European journal of pharmacology 2020, 867, 172853. [Google Scholar] [CrossRef]
- Kaya, İ.; Dağ, S.; Kaya, M.M.; Tanrıverdi, E.A.; Beşeren, H.; Aşasın, G. Modulatory effect of pomegranate extract on TRPA1, TRPM2 and caspase-3 expressions in colorectal cancer induction of mice. Turkish Journal of Biochemistry 2022, 47, 612–619. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, H.; Wu, K.; Liu, Z. Expression of 4-hydroxynonenal in esophageal squamous cell carcinoma. Oncology letters 2017, 14, 35–40. [Google Scholar] [CrossRef]
- Nakashima, S.; Shiozaki, A.; Ichikawa, D.; Hikami, S.; Kosuga, T.; Konishi, H.; Komatsu, S.; Fujiwara, H.; Okamoto, K.; Kishimoto, M.; et al. Transient Receptor Potential Melastatin 7 as an Independent Prognostic Factor in Human Esophageal Squamous Cell Carcinoma. Anticancer research 2017, 37, 1161–1167. [Google Scholar] [CrossRef]
- Jakovcevic, A.; Zarkovic, K.; Jakovcevic, D.; Rakusic, Z.; Prgomet, D.; Waeg, G.; Sunjic, S.B.; Zarkovic, N. The Appearance of 4-Hydroxy-2-Nonenal (HNE) in Squamous Cell Carcinoma of the Oropharynx. Molecules 2020, 25. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The topological structure of redox TRP channels. Each subunit consists of six transmembrane domains (S1-S6); one loop between S5 and S6 containing the channel pore; N and C termini located intracellularly. Illustrated in red are the cysteine residues exposed to oxidation. TRPC4/TRPC5 have two conserved cysteine residues in the pore-forming region. TRPV1 and TRPV4 have cysteine residues intracellularly and extracellularly located. TRPA1 contains ankyrin repeats, while TRPM2 and TRPM7 have a conserved TRP box. The coiled-coil region and NUDT9-H domain from TRPM2 provide the binding site for ADPR. TRPM7 has a zinc-finger domain containing two cysteines exposed to oxidation.
Figure 1.
The topological structure of redox TRP channels. Each subunit consists of six transmembrane domains (S1-S6); one loop between S5 and S6 containing the channel pore; N and C termini located intracellularly. Illustrated in red are the cysteine residues exposed to oxidation. TRPC4/TRPC5 have two conserved cysteine residues in the pore-forming region. TRPV1 and TRPV4 have cysteine residues intracellularly and extracellularly located. TRPA1 contains ankyrin repeats, while TRPM2 and TRPM7 have a conserved TRP box. The coiled-coil region and NUDT9-H domain from TRPM2 provide the binding site for ADPR. TRPM7 has a zinc-finger domain containing two cysteines exposed to oxidation.
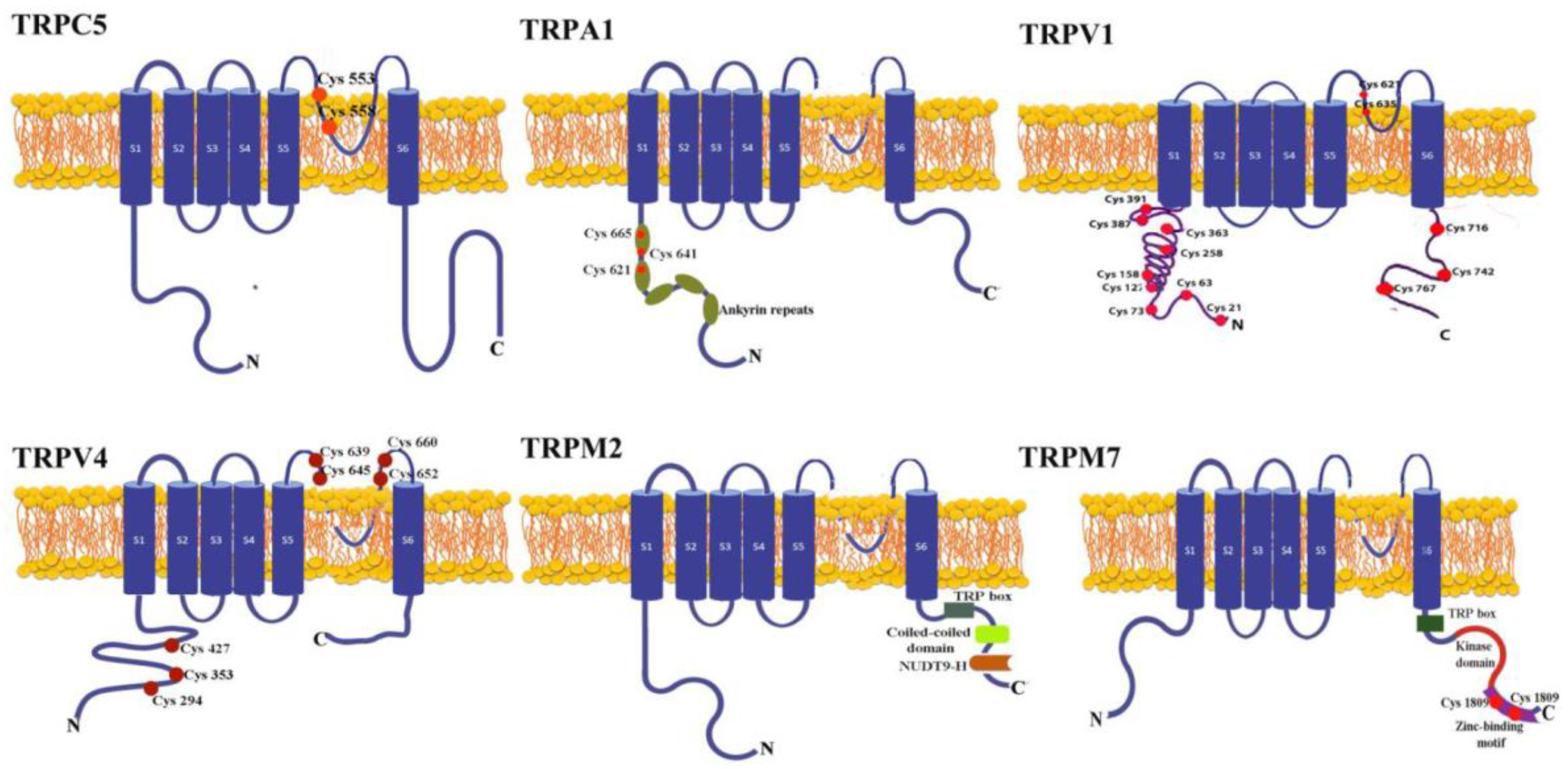
Figure 2.
Signaling pathway activated by TRP channels in cancer cell proliferation. TRPM2 activates mitochondria and the transcription factors: HIF-1/2α, Nrf2, ATF4, and CREB; induces the proliferation of acute myeloid leukemia (AML) cells. The pathway is indicated with magenta connectors.
TRPM2 activates mitochondria and the transcription factors: HIF-1/2α, Nrf2, ATF4, and CREB; induces the proliferation of acute myeloid leukemia (AML) cells. The pathway is indicated with magenta connectors.  A pathway illustrated in blue, indicates that Ca2+ uptake by TRPM2 promotes survival of neuroblastoma cells. Nrf2 activates the antioxidant response and cofactors GSH, NADPH, and NADH.
A pathway illustrated in blue, indicates that Ca2+ uptake by TRPM2 promotes survival of neuroblastoma cells. Nrf2 activates the antioxidant response and cofactors GSH, NADPH, and NADH.  TRPM2-S is activated by ROS and consecutively increases apoptosis and decreases proliferation in retroperitoneal liposarcoma (RPLS) cells, through AKT/MAPK pathways as depicted in scarlet. agonists of TRPM7 stimulate ROS production and minimize mitochondrial turnover. The connections between TRPM7, ROS production, and consecutive reduction in autophagy and proliferation are displayed in green.
TRPM2-S is activated by ROS and consecutively increases apoptosis and decreases proliferation in retroperitoneal liposarcoma (RPLS) cells, through AKT/MAPK pathways as depicted in scarlet. agonists of TRPM7 stimulate ROS production and minimize mitochondrial turnover. The connections between TRPM7, ROS production, and consecutive reduction in autophagy and proliferation are displayed in green.
TRPM2 activates mitochondria and the transcription factors: HIF-1/2α, Nrf2, ATF4, and CREB; induces the proliferation of acute myeloid leukemia (AML) cells. The pathway is indicated with magenta connectors. A pathway illustrated in blue, indicates that Ca2+ uptake by TRPM2 promotes survival of neuroblastoma cells. Nrf2 activates the antioxidant response and cofactors GSH, NADPH, and NADH. TRPM2-S is activated by ROS and consecutively increases apoptosis and decreases proliferation in retroperitoneal liposarcoma (RPLS) cells, through AKT/MAPK pathways as depicted in scarlet. agonists of TRPM7 stimulate ROS production and minimize mitochondrial turnover. The connections between TRPM7, ROS production, and consecutive reduction in autophagy and proliferation are displayed in green.
Figure 2.
Signaling pathway activated by TRP channels in cancer cell proliferation. TRPM2 activates mitochondria and the transcription factors: HIF-1/2α, Nrf2, ATF4, and CREB; induces the proliferation of acute myeloid leukemia (AML) cells. The pathway is indicated with magenta connectors. A pathway illustrated in blue, indicates that Ca2+ uptake by TRPM2 promotes survival of neuroblastoma cells. Nrf2 activates the antioxidant response and cofactors GSH, NADPH, and NADH. TRPM2-S is activated by ROS and consecutively increases apoptosis and decreases proliferation in retroperitoneal liposarcoma (RPLS) cells, through AKT/MAPK pathways as depicted in scarlet. agonists of TRPM7 stimulate ROS production and minimize mitochondrial turnover. The connections between TRPM7, ROS production, and consecutive reduction in autophagy and proliferation are displayed in green.
TRPM2 activates mitochondria and the transcription factors: HIF-1/2α, Nrf2, ATF4, and CREB; induces the proliferation of acute myeloid leukemia (AML) cells. The pathway is indicated with magenta connectors. A pathway illustrated in blue, indicates that Ca2+ uptake by TRPM2 promotes survival of neuroblastoma cells. Nrf2 activates the antioxidant response and cofactors GSH, NADPH, and NADH. TRPM2-S is activated by ROS and consecutively increases apoptosis and decreases proliferation in retroperitoneal liposarcoma (RPLS) cells, through AKT/MAPK pathways as depicted in scarlet. agonists of TRPM7 stimulate ROS production and minimize mitochondrial turnover. The connections between TRPM7, ROS production, and consecutive reduction in autophagy and proliferation are displayed in green.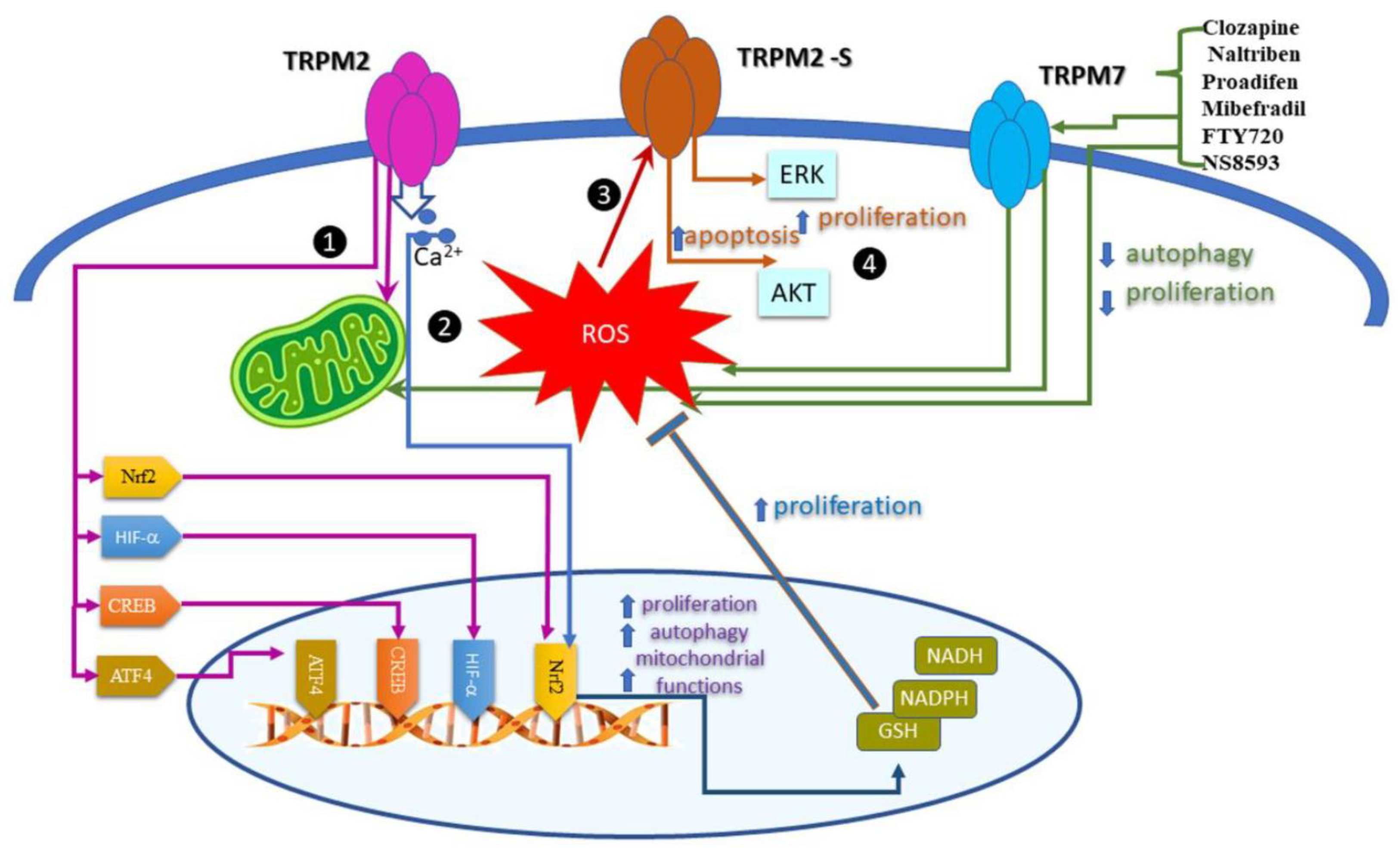
Figure 3.
Intracellular pathways activated by redox TRP channels triggering apoptosis in cancer cells Calcium entry through TRPA1 inhibited apoptosis upon ROS activation. The process involves the nuclear factor erythroid 2-related factor (Nrf2). The model is valid for breast cancer cells. In glioblastoma, temozolomide (TMZ) activates ROS, TRPA1, and apoptosis. The process is related to TMZ resistance. Concomitant application of TRPV1 agonists (MRS1477 and capsaicin) increased apoptosis and ROS production.  capsazepin, a TRPV1 antagonist, upregulates proapoptotic proteins p53 and Bax, which augment TRAIL-induced apoptosis. The activation of TRAIL receptors triggers the ROS–JNK–CHOP pathway.
capsazepin, a TRPV1 antagonist, upregulates proapoptotic proteins p53 and Bax, which augment TRAIL-induced apoptosis. The activation of TRAIL receptors triggers the ROS–JNK–CHOP pathway.
Calcium entry through TRPA1 inhibited apoptosis upon ROS activation. The process involves the nuclear factor erythroid 2-related factor (Nrf2). The model is valid for breast cancer cells. In glioblastoma, temozolomide (TMZ) activates ROS, TRPA1, and apoptosis. The process is related to TMZ resistance. Concomitant application of TRPV1 agonists (MRS1477 and capsaicin) increased apoptosis and ROS production. capsazepin, a TRPV1 antagonist, upregulates proapoptotic proteins p53 and Bax, which augment TRAIL-induced apoptosis. The activation of TRAIL receptors triggers the ROS–JNK–CHOP pathway.
Figure 3.
Intracellular pathways activated by redox TRP channels triggering apoptosis in cancer cells Calcium entry through TRPA1 inhibited apoptosis upon ROS activation. The process involves the nuclear factor erythroid 2-related factor (Nrf2). The model is valid for breast cancer cells. In glioblastoma, temozolomide (TMZ) activates ROS, TRPA1, and apoptosis. The process is related to TMZ resistance. Concomitant application of TRPV1 agonists (MRS1477 and capsaicin) increased apoptosis and ROS production. capsazepin, a TRPV1 antagonist, upregulates proapoptotic proteins p53 and Bax, which augment TRAIL-induced apoptosis. The activation of TRAIL receptors triggers the ROS–JNK–CHOP pathway.
Calcium entry through TRPA1 inhibited apoptosis upon ROS activation. The process involves the nuclear factor erythroid 2-related factor (Nrf2). The model is valid for breast cancer cells. In glioblastoma, temozolomide (TMZ) activates ROS, TRPA1, and apoptosis. The process is related to TMZ resistance. Concomitant application of TRPV1 agonists (MRS1477 and capsaicin) increased apoptosis and ROS production. capsazepin, a TRPV1 antagonist, upregulates proapoptotic proteins p53 and Bax, which augment TRAIL-induced apoptosis. The activation of TRAIL receptors triggers the ROS–JNK–CHOP pathway.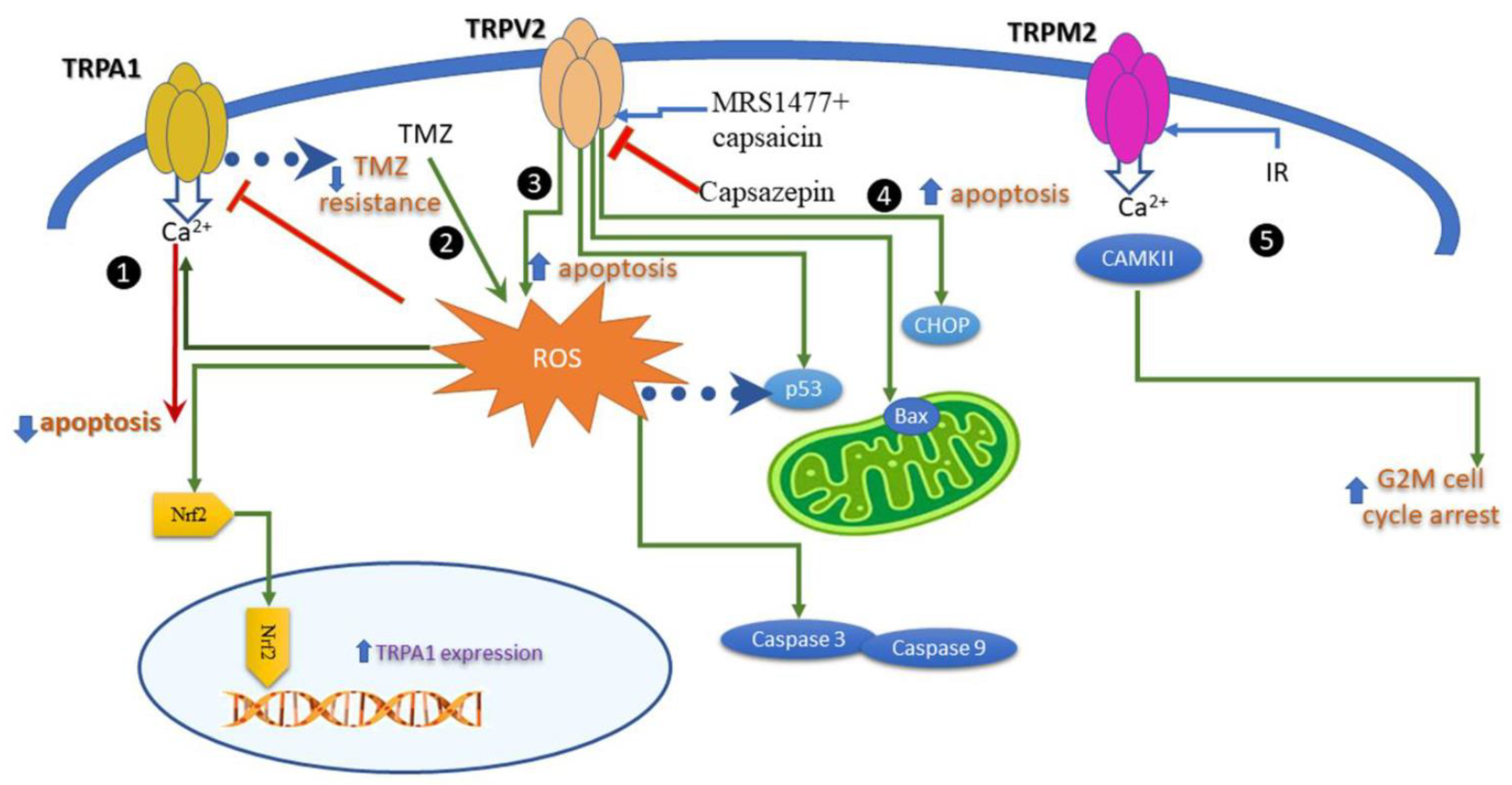
Figure 4.
TRP redox channels modulate the migration of cancer cells ROS inhibits stress fibers and consecutively migration through the downregulation of the DLC1/RhoA pathway. Migration mediated by Src kinases. ROS stimulates Src, which in turn phosphorylates TRPV4 and TRPV7. TRPV4 channel contributes to migration via Src-cofilin intracellular pathway, while the TRPV7 kinase domain modulates Src phosphorylation. Ca2+ entry via TRPM2 activates transcription factors HIF-1α1, E2F1, FOXM1, and CREB, which in turn modulate the expression of α and β integrins. The proteins α and β integrin from the focal adhesion complex engage the extracellular matrix (ECM) in cell migration.
ROS inhibits stress fibers and consecutively migration through the downregulation of the DLC1/RhoA pathway. Migration mediated by Src kinases. ROS stimulates Src, which in turn phosphorylates TRPV4 and TRPV7. TRPV4 channel contributes to migration via Src-cofilin intracellular pathway, while the TRPV7 kinase domain modulates Src phosphorylation. Ca2+ entry via TRPM2 activates transcription factors HIF-1α1, E2F1, FOXM1, and CREB, which in turn modulate the expression of α and β integrins. The proteins α and β integrin from the focal adhesion complex engage the extracellular matrix (ECM) in cell migration.
Figure 4.
TRP redox channels modulate the migration of cancer cells ROS inhibits stress fibers and consecutively migration through the downregulation of the DLC1/RhoA pathway. Migration mediated by Src kinases. ROS stimulates Src, which in turn phosphorylates TRPV4 and TRPV7. TRPV4 channel contributes to migration via Src-cofilin intracellular pathway, while the TRPV7 kinase domain modulates Src phosphorylation. Ca2+ entry via TRPM2 activates transcription factors HIF-1α1, E2F1, FOXM1, and CREB, which in turn modulate the expression of α and β integrins. The proteins α and β integrin from the focal adhesion complex engage the extracellular matrix (ECM) in cell migration.
ROS inhibits stress fibers and consecutively migration through the downregulation of the DLC1/RhoA pathway. Migration mediated by Src kinases. ROS stimulates Src, which in turn phosphorylates TRPV4 and TRPV7. TRPV4 channel contributes to migration via Src-cofilin intracellular pathway, while the TRPV7 kinase domain modulates Src phosphorylation. Ca2+ entry via TRPM2 activates transcription factors HIF-1α1, E2F1, FOXM1, and CREB, which in turn modulate the expression of α and β integrins. The proteins α and β integrin from the focal adhesion complex engage the extracellular matrix (ECM) in cell migration.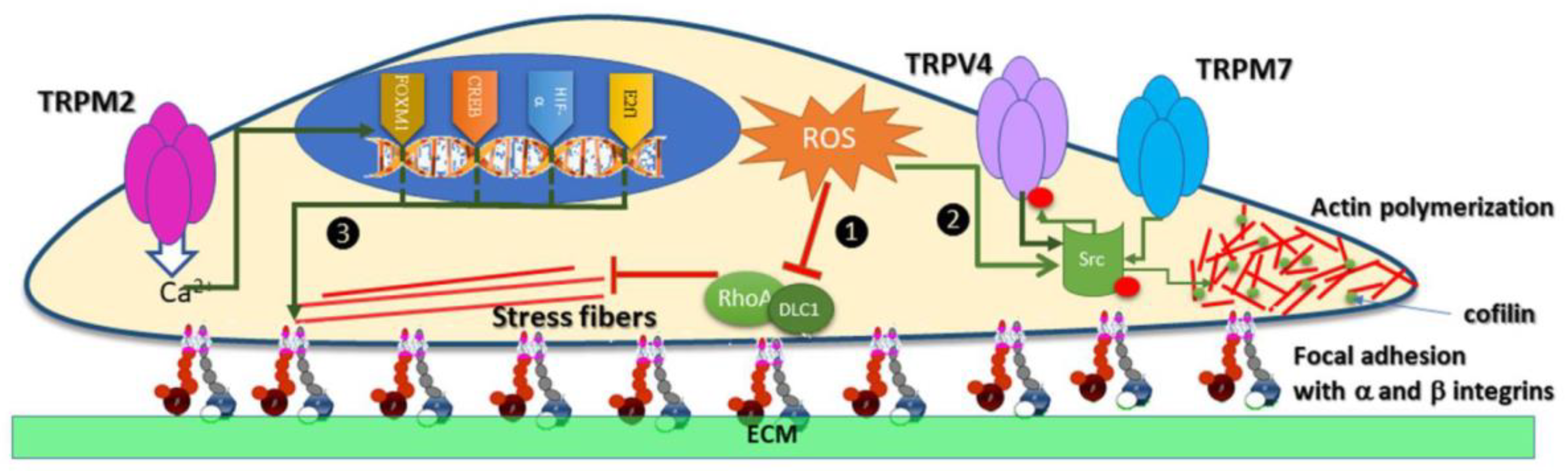

Table 1.
Anti-inflammatory drugs with effects on tumorigenesis and modulators of TRP channels.
| Anti-inflammation drug | TRP channel affected | Role in tumorigenesis | Ref. |
|---|---|---|---|
| Nonsteroidal anti-inflammatory drugs (NSAIDs), e.g., aspirin and cyclooxygenase | Inhibition of TRPC4/C5 | reduces the risk of breast cancer | [13,44] [85] |
| Etodolac (Cox-2 inhibitor) | Desensitization of TRPA1 | Inhibition of proliferation of prostate and colorectal carcinoma | [86,87] |
| Cannabinoids and Cannabis Extracts | Activation of TRPV4 and TRPA1 | Inhibits growth of breast cancer cells | [88,89] |
| Methotrexate | Inhibition of TRPM2 | Chemotherapy of breast cancer, head, and neck cancer, lymphoma | [90] |
| Capsaicin | Activation of TRPV1 | Senescence and Apoptosis in cervical cancer cells | [91,92,93] |
| Capsazepine | Inhibition of TRPV1 | Inhibits growth and survival of prostate cancer, breast cancer, colorectal cancer, oral cancer, and osteosarcoma | [94] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



