
Submitted:
18 May 2023
Posted:
19 May 2023
You are already at the latest version
Abstract
Regulatory macrophages (Mregs) are unique in that they have anti-inflammatory and immunosuppressive properties. Thus, treating inflammatory diseases using Mregs is an area of active research. Human Mregs are usually generated by culturing peripheral blood monocytes stimulated by macrophage colony-stimulating factor with interferon (IFN)-. Here, we generated Mregs with an elongated cell morphology from THP-1 cells stimulated with phorbol 12-myristate 13-acetate and cultured with arginylglycylaspartic acid and vitamin D3. These Mregs regulated macrophage function, and respectively downregulated and upregulated the expression of pro-inflammatory and immunosuppressive mediators. They also expressed Mregs-specific marker, such as dehydrogenase/reductase 9, even when exposed to inflammatory stimulants such as IFN-, lipopolysaccharide, purified xenogeneic antigen, and xenogeneic cells. The Mregs also exerted anti- inflammatory and anticoagulatory actions in response to xenogeneic cells and immunosuppressive effects on mitogen-induced Jurkat T-cell proliferation. Our method of generating functional Mregs in vitro without cytokines is simple and cost-effective.
Keywords:
regulatory macrophage
; arginylglycylaspartic acid
; vitamin D3
; anti-inflammation
; xenotrans-plantation
1. Introduction
Macrophages are phagocytic immune cells that play diverse roles in host defense against infectious agents and tissue injury [1]. Macrophages are activated and convert from one type to another according to various environmental stimuli [2]. The two representative types of macrophages are classically (M1) and alternatively (M2) activated [3]. M1 macrophages can be differentiated from resting macrophages in response to pathogens (PAMPs)- or damage (DAMPs)- associated molecular patterns derived from infectious agents or injured tissues [4,5]. They produce pro-inflammatory cytokines, nitric oxide, and reactive oxygen species (ROS) that kill pathogens. M2 macrophages are induced by colony-stimulating factor (M-CSF), interleukins (IL)-4, -10, or -13, and have anti-inflammatory and regulatory properties [6]. M2 macrophages can be further classified as M2a, M2b, M2c, and M2d subtypes according to their inducing factors and phenotypic characteristics [5]. Interleukins-4 and -13 induce M2a macrophages [7] that secrete IL-10, TGF-β, and arginase-1 [8]. Immune complexes and toll-like receptor (TLR) ligands, such as lipopolysaccharide (LPS) induce M2b cells [9] that secrete high levels of IL-10, and IL-1 receptor antagonist (IL-1Ra), and low levels of IL-12 [10,11]. Glucocorticoids, IL-10, and TGF-β induce M2c cells that secrete high levels of IL-10 and TGF-β [12]. Interleukin-6 and A2 adenosine receptor agonists induce M2d macrophages that secrete IL-10, transforming growth factor-beta (TGF-β), C-C Motif Chemokine Ligand 22 (CCL22), and vascular endothelial growth factor (VEGF) [6,13]. All these M2 macrophage subtypes are involved in the regulation of inflammation and tissue repair and are differentiated by distinct stimuli and mediators.
Regulatory macrophages (Mregs) are distinct populations of macrophages with regulatory functions that dampen inflammatory immune responses and suppress those of T cells [14,15]. Anti-inflammatory Mregs have therapeutic value for several diseases, such as inflammatory bowel disease (IBD; [16]), colitis [17], Crohn’s disease [18], leishmaniasis [19], and organ transplantation [15,20]. Human Mregs have been derived from CD14+ peripheral blood monocytes stimulated with M-CSF and then incubated with IFN-γ [21]. The induction of Mregs requires contact with a plastic surface and exposure to serum factors [20,21,22].
The most physiologically active form of vitamin D3 is 1,25-dihydroxy-vitamin D3 (vitD3) which functions in regulation of the immune system, the nervous system, and calcium balance [23] and mediates its biological effects by binding to vitamin D receptors (VDRs) [24]. This vitamin exerts anti-inflammatory effects on monocytes/macrophages [25,26] and can induce the M2 polarization of macrophages [27,28] and microglia [29].
The arginine-aspartate-glycine (RGD) motif is an important structural component of various proteins that are recognized by integrins and mediate cell-cell interaction and adhesion to the extracellular matrix [30]. Binding to the RGD motif induces a conformational change in integrin and triggers intracellular signaling, which is involved in the regulation of macrophage adhesion and activation [31]. Because of their ability to regulate macrophage functions, a cell adhesion protein containing the RGD motif [32] and integrin β3 [33] are responsible for the M2 polarization of macrophages.
Here, we established a new protocol to induce macrophage differentiation into Mregs using vitD3 and the RGD motif without cytokines, which are typically applied to generate Mregs, then characterized their phenotype and functional features.
2. Materials and Methods
2.1. Reagents and Antibodies
The following reagents were obtained from the respective suppliers: phorbol-12-myristate 13-acetate (PMA; Cayman Chemical (Ann Arbor, MI, USA), 1,25-dihydroxy vitamin D3 (Toronto Research Chemicals, Inc., Toronto, ON, Canada), Roswell Park Memorial Institute (RPMI)-1640, Dulbecco modified Eagle medium (DMEM), Dulbecco phosphate-buffered saline (DPBS), and fetal bovine serum (FBS) (Welgene, Inc., Daegu, Korea). Interferon-gamma (IFN-γ) and anti-MER-TK antibodies (R&D Systems, Minneapolis, MN, USA). Arginylglycylaspartic acid (RGD), phytohemagglutinin P (PHA), and LPS from Escherichia coli K12 (Sigma-Aldrich, St. Louis, MO, USA), carboxyfluorescein succinimidyl ester (CFSE) CellTrace™ Cell Proliferation Kit, and antibiotic-antimycotic (Invitrogen; Thermo Fisher Scientific Inc., Waltham, MA, USA), ethylenediaminetetraacetic acid (EDTA; Amresco, Inc., Solon, OH, USA), Galα1-3Galβ1-4GlcNAcβ-PAA trisaccharide (α-Gal; GlycoTech, Gaithersburg, MD, USA), anti-CD14, anti-CD274, and anti-CD80 ( eBioscience Inc. (San Diego, CA, USA), anti-CD16 (Biolegend Inc., San Diego, CA, USA), anti-DHRS9 (Abnova, Taiwan, China), anti-CD11b (BD Biosciences Inc., Franklin Lakes, NJ, USA), anti-CD209 (Serotec Bio-Rad Inc., CA, USA), FITC-conjugated and other antibodies (Santa Cruz Biotechnology Inc., Dallas, TX, USA)
2.2. Cell Culture
The human monocytic cell line, THP-1 (Korean Cell Line Bank, Seoul, Korea) was grown in RPMI-1640 medium supplemented with 5% heat-inactivated fetal bovine serum (FBS) and 1% antibiotic-antimycotic. The established porcine vascular endothelial cell line MPN-3 [34] was grown in DMEM supplemented with 10% heat-inactivated fetal bovine serum (FBS) and 1% antibiotic-antimycotic solution. Jurkat T cells (E6.1) were grown in RPMI-1640 medium supplemented with 10% heat-inactivated FBS and 1% antibiotic-antimycotic. T cells were stained with 4 μM CFSE (Invitrogen) in PBS at room temperature for 10 min, washed three times with culture medium containing 10% FBS, then stimulated with 2 μg/mL PHA. The cells were maintained at 37°C in a 5% CO2 humidified incubator.
2.3. Generation of Mregs
Figure 1 shows the protocol used to generate various human macrophage subtypes. Briefly, we incubated THP-1 cells with 10 ng/mL PMA for 48 h, then stimulated them with 100 ng/mL LPS and 100 ng/mL IFN-γ to generate M1 macrophages. The PMA-induced macrophages were incubated with 1 µg/mL RGD for 24 h, followed by 1 µM vitD3 for 24 h to generate Mregs. Cells were harvested using 5 mM EDTA on ice to avoid damage by detachment as described [35] and characterized.
2.4. Co-Culture Experiments
We assessed mitogen-induced T cell proliferation in 1 × 106 Jurkat T cells incubated with 1 mL 2 μM CFSE for 10 min at 37°C. The reaction was stopped by adding medium containing 10% FBS, then the cells were washed with the same medium three times. Labeled cells were maintained overnight before co-culture with Mregs at a ratio of 1:1. T cells and co-cultured T cells + Mregs were incubated with a final concentration of 2 μg/mL PHA for 5 days, harvested, then analyzed using flow cytometry.
We co-cultured Mregs with M1 macrophages for 3 days and then assessed their inflammatory responses to xenogeneic MPN-3 cells. Cells were harvested at 3, 6, and 12 h after co-culture, and the mRNA expression of inflammatory, anti-inflammatory, and coagulation-related mediators were analyzed using quantitative real-time polymerase chain reaction (qRT-PCR).
2.5. Preparation of RNA and qRT-qPCR
Total RNA extracted using easy-BLUETM Total RNA Extraction Kits (iNtRON Biotechnology, Inc., Seongnam-Si, South Korea) as described by the manufacturer was quantified using a MaestroNano Micro-Volume Spectrophotometer (MaestroGen Inc., Las Vegas, NV, USA). Thereafter, cDNA was synthesized using 2 μg of total RNA in Hyperscript RT master mix (GeneAll Biotechnology, Seoul, Korea) and an Invitrogen Oligo (dT) primer (Thermo Fisher Scientific Inc.). The cDNA was amplified by qRT-PCR using a LineGene 9600 Plus Fluorescent Quantitative Detection System (Hangzhou Bloer Technology Co., Ltd. [BIOER], Hangzhou, China) and EzAMPTM FAST One-Step RT-qPCR 2x Master Mix (SYBR; Elpis-Biotech Inc., Daejeon, South Korea) and the following (5′ → 3′) primer sets:
Fibrinogen-like protein 2 (fgl-2): AGCTGATGACAGCAGAGTTAGAG and AGTGATCATACAAGGCATAGAGC;
Inducible nitric oxide synthase (iNOS): ACAGCACATTCAGATCCCCA and AACACGTTCTTGGCATGCAT;
IL-1β: GGGATAACGAGGCTTATGTGC and AGGTGGAGAGCTTTCAGTTCA;
IL-6: GACCCAACCACAAATGCCAG and GAGTTGTCATGTCCTGCAGC;
IL-10: TCTCCGAGATGCCTTCAGCAGA and TCAGACAAGGCTTGGCAACCCA;
IL-12: ACGAGTGCTCCTGGCAGTAT and AGGATTCCACCCAGAGTGTG;
Indoleamine 2,3-dioxygenase (IDO): TGCAAGAACGGGACACTTTG and CCCTTCATACACCAGACCGT;
Monocyte chemoattractant protein-1 (MCP-1): CCCAAGAATCTGCAGCTAAC and GGTAGAACTGTGGTTCAAGAGG;
Tissue Factor (TF): GGGCTGACTTCAATCCATGT and GAAGGTGCCCAGAATACCAA;
Tissue necrosis factor alpha (TNF-α): TGAGCACTGAAAGCATGATCC and GGAGAAGAGGCTGAGGAACA;
Protease-activated receptor-1 (PAR-1): CATCTGTGTACACCGG and TGCCAATCACTGCC;
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH): ACAGCCTCAAGATCATCAGCAAT and AGGAAATGAGCTTGACAAAGTGG.
Results were normalized using human GAPDH as the endogenous control. The relative abundance of target mRNA in each sample was calculated from the C△t values of the target and the GAPDH housekeeping gene using the 2−△△cycle threshold (Ct) method.
2.6. Flow Cytometry
The cells were incubated with primary antibodies for 30 min at 4°C followed by PE-conjugated or FITC-conjugated secondary antibodies at 4°C for 30 min. After three washes with DPBS, the cells were resuspended in 0.4 mL PBS and cell surface protein expression was analyzed using Cytomics FC500 MLP and CXP software (Beckman Coulter Inc., Fullerton, CA, USA).
The cells were fixed in 4% formaldehyde in DPBS, permeabilized with 0.1% Triton X-100 at room temperature for 10 min, then stained with primary anti-iNOS, anti-DHRS9, and secondary antibodies to determine the expression of intracellular protein as described above for cell surface protein.
2.7. Statistical Analysis
All experiments were repeated at least three times. Significant differences among groups were assessed using one-way analysis of variance (ANOVA), followed by post hoc tests using SPSS 12.0 (SPSS Inc., Chicago, IL, USA). All data are expressed as means ± standard deviation (SD). Differences were considered statistically significant at p < 0.05.
3. Results
3.1. Morphology and Expression of Anti-Inflammatory Cytokines in THP-1 Cells Incubated with PMA, RGD, and vitD3
One of the most important changes in Mregs differentiation is cell elongation [36]. Therefore, we examined the morphology of THP-1 cells incubated with various combinations of vitD3 and RGD (Figure 1). Cells were the most elongated when incubated with 1 μg/mL RGD or 1 μM vitD3 among the assessed concentrations (Figure 2A,B). Thus, we applied this combination in subsequent experiments. The cells seemed more efficiently elongated after incubation with PMA + RGD + vitD3 than with either PMA + RGD or PMA + vitD3 (Figure 2C). We evaluated the expression of anti-inflammatory IL-10 and pro-inflammatory IL-12 using qRT-PCR to determine whether the cells generated by PMA + RGD + vitD3 had anti-inflammatory properties. Figure 3 shows enhanced and diminished mRNA expression of IL-10 and IL-12, respectively, in these cells, indicating an anti-inflammatory phenotype.
3.2. Cells Incubated with PMA, RGD, and vitD3 Express Mregs Markers
Various cell surface markers are induced by M-CSF and IFN-γ in Mregs [15]. Therefore, we examined the expression of several surface markers in macrophages incubated with PMA + RGD + vitD3 using flow cytometry to determine whether these cells express the same factors as those previously established. Generic markers of human macrophages, CD11b, CD16, and CD33, were expressed, whereas CD14 expression was low (Figure 4). Typical markers of the tissue-resident macrophages CD163, CD206, CD209, and MER-TK were significantly enhanced in cells incubated with PMA + RGD + vitD3 (Fig. 4). The expression of most surface markers, including costimulatory CD80 and CD86 and inhibitory CD274 and TREM-2 were similar between macrophages incubated with PMA + RGD + vitD3 and established Mregs [15]. Significantly less iNOS was expressed in macrophages incubated with PMA + RGD + vitD3 than in resting macrophages (Figure 4). Human Mregs selectively express the stable marker dehydrogenase/reductase 9 (DHRS9), which is involved in the initial step of synthesizing the important immune regulator retinoic acid from retinol or β-carotene ingested as a vital nutrient [22]. We found more abundant DHRS9 expression in macrophages incubated with PMA + RGD + vitD3 than in the other cells examined (Figure 5A,B). The elongated cells and surface marker expression indicated that the macrophages-induced PMA + RGD + vitD3 have the morphological and phenotypic characteristics of Mregs.
3.3. Mregs Stably Express Mregs Markers even when Exposed to Inflammatory Stimulants
The phenotypic and functional stability of Mregs is crucial factors for their clinical application, and instability under inflammatory conditions is a problem that should be addressed [12]. Since macrophage polarization can be reversed by environmental stimuli and IFN-γ stimulates polarization towards the M1 pro-inflammatory phenotype [37], we examined DHRS9 expression in Mregs incubated with IFN-γ. We found that IFN-γ did not affect the enhanced expression of DHRS9 in Mregs, indicating that the Mregs were stable (Figure 5C,D). We further investigated the phenotypic stability of several Mregs markers in a pro-inflammatory milieu using flow cytometry. We incubated the Mregs with LPS, a major component of bacterial cell walls that stimulates macrophages to generate inflammatory responses. Enhanced expression of the Mregs markers CD163, CD206, CD209, CD274, and DHRS9 (Figure 6A,B) and decreased expression of the inflammatory markers, CD80, iNOS, TLR2, and TLR4 (Figure 6C,D) were not affected in Mregs incubated with LPS. We then examined the expression of several markers in Mregs exposed to the xenogeneic antigen α-Gal, which is a sugar molecule expressed on porcine, but not human cells and thus directly activates human macrophages [38]. The expression of markers in Mregs incubated with or without α-Gal did not differ (Figure 7A). Expression of the inflammatory markers, CD80 and iNOS was significantly enhanced by α-Gal in THP-1 cells compared with control cells incubated without α-Gal, whereas that in Mregs did not differ regardless of α-Gal (Figure 7B).
3.4. Expression of Pro-Inflammatory and Immunosuppressive Mediators is Respectively Damped and Enhanced in Mregs Co-Cultured with Xenogeneic Cells
We assessed the mRNA expression of pro-inflammatory and immunosuppressive mediators in THP-1 cells, M1 macrophages derived from THP-1 cells according to the procedure described in Figure 1, and Mregs co-cultured with the porcine aortic vascular endothelial cell line MPN3 [34]. Expression of the pro-inflammatory mediators IL-1β, IL-6, IL-12, TNF-α, MCP-1, and iNOS was significantly enhanced, whereas that of anti-inflammatory IL-10 was significantly decreased in co-cultured M1 macrophages and MPN3, compared with co-cultured THP-1 and MPN3 cells. In contrast, the expression of pro-inflammatory mediators was significantly suppressed, whereas that of the immunosuppressive mediators IL-10 and IDO was obviously enhanced in co-cultured Mregs and MPN3 cells (Figure 8A). The coagulation-associated mediators fgl2, PAR-1, and TF play important roles in inflammation and coagulation during xenograft rejection [39]. We examined the mRNA expression of these factors in THP-1 cells, M1 macrophages, and Mregs co-cultured with MPN3 cells. The expression of human fgl2, PAR-1, and TF was significantly more increased in M1 and MPN3 than in THP-1 and MPN3 co-cultures, and significantly lower in co-cultured Mregs and MPN3 than in the other two groups (Figure 8B). We co-cultured Mregs with M1 and MPN3 cells at different ratios and assessed the mRNA expression of pro-inflammatory and coagulation-associated mediators to determine whether Mregs would suppress immune responses. The expression of pro-inflammatory mediators was significantly increased in M1 and MPN3, compared with THP-1 and MPN3 co-cultures (Figure 9A). However, the enhanced expression was completely inhibited at a 1:1 ratio of Mregs:M1. The expression of inflammatory mediators gradually increased as the Mregs:M1 ratio decreased and reached the levels of M1 only at a Mregs/M1 ratio of 1:4, except for MCP-1, the expression of which was completely suppressed at this ratio (Figure 9A). Similarly, the expression of coagulation-associated mediators was significantly increased in M1 and MPN3 compared with THP-1 and MPN3 co-cultures (Figure 9B). However, this was completely inhibited at a 1:1 ratio of Mregs to M1. The expression of coagulation-associated mediators gradually increased as the Mregs:M1 ratio decreased and reached levels only when the Mregs/M1 ratio was 1:4. The expression for TF, however, remained significantly suppressed at this ratio (Fig. 9B). These results indicated that our Mregs suppress the expression of inflammatory and coagulation-associated mediators of M1 macrophages in response to xenogeneic α-Gal and bacterial LPS.
3.5. Regulatory Macrophages Immunosuppress Mitogen-Induced T Cell Proliferation
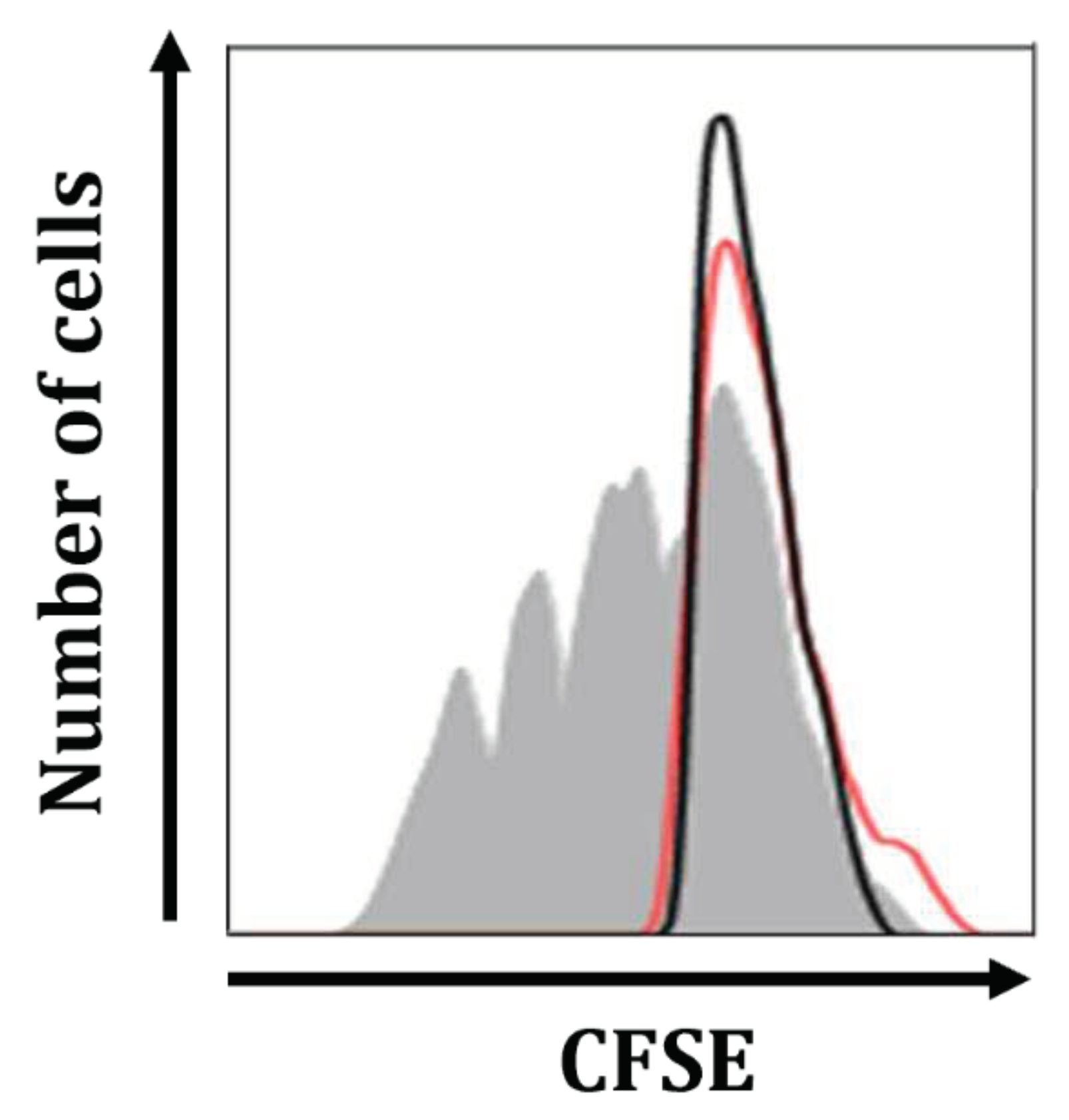
The suppression of mitogen-induced T cell proliferation is a distinct feature of Mregs [21]. Therefore, we investigated whether Mregs could suppress the mitogen-stimulated Jurkat T cell line. We co-cultured Mregs with CFSE-labeled Jurkat T cells and incubated them with the T cell mitogen, phytohemagglutinin (PHA). Figure 10 shows that T cells stimulated with PHA rapidly proliferated (gray shadow), whereas those stimulated with a 1:1 ratio of Mregs to T cells did not proliferate (red line). These results indicated that Mregs suppress mitogen-induced T cell proliferation.
4. Discussion
We induced Mregs from THP-1 monocytes incubated with PMA, RGD and vitD3. We did not use M-CSF and IFN-γ that are commonly used to generate human Mregs [11]. Our Mregs had an elongated morphology, which was in line with other findings [15,40] and were phenotypically characterized as CD11bhi, CD14lo, CD16+, CD33+, CD80lo, CD86+, iNOS-, CD163hi, CD206+, CD209+, CD274+/hi, TREM-2+, MER-TKhi, and DHRS9+/hi. Most of the markers evaluated herein are expressed in Mregs generated by others, including CD14, CD80, CD86 [15,21,41], CD163, CD206, CD274 [15,42], CD209, MER-TK [15], and DHRS9 [22]. Among these, DHRS9 is considered a specific and stable marker of Mregs because it is not expressed by other known human monocyte-derived tolerogenic or suppressive cells used for cell-based immunotherapies [22] such as immature monocyte-derived dendritic cells (DCs), tolerogenic DCs [43], rapamycin-treated DCs [44], IL-10 conditioned DCs [45] or prostaglandin E2-induced myeloid-derived suppressor cells [46]. Furthermore, DHRS9 expression is stable even when Mregs are exposed to LPS or IFN-γ [22]. Here, DHRS9 expression was stable in Mregs exposed to IFN-γ, LPS and xenogeneic α-Gal. Our Mregs had anti-inflammatory phenotypes, with upregulated IL-10 and downregulated IL-12, iNOS, CD80, TLR2, and TLR4 expression. In addition, the expression of anti-inflammatory mediators was enhanced, while that of pro-inflammatory and coagulation-related mediators was damped in response to xenogeneic antigens or cells. Furthermore, Mregs inhibited mitogen-induced T-cell proliferation, which is another cardinal feature of Mregs [21]. Taken together, the present findings showed that incubating THP-1 monocytes with PMA + RGD + vitD3 induces functional Mregs.
Regulatory macrophages can be induced by combinations of M-CSF and IFN-γ, LPS and immune complex, LPS and prostaglandin E2, anti-TNF-α monoclonal antibody and others [11]. Among them, M-CSF + IFN-γ is the most prevalent inducer, especially of human Mregs. The induction of Mregs normally requires two consecutive signals; primary signals generated by M-CSF that induce monocyte differentiation into macrophages [47], and secondary signals stimulated by IFN-γ that induce Mregs to harness anti-inflammatory properties [48]. Because PMA can induce monocyte differentiation into macrophages [49], it might have acted like M-CSF in the process of Mreg induction in the present study. Because vitD3 exerts anti-inflammatory effects on macrophages [25,26] and induces M2 polarization of macrophages through the VDR signaling mechanism [27,28], we speculate that it acts as a stimulant for secondary signals to coordinate an anti-inflammatory environment. We also used RGD to generate Mregs in addition to these two reagents. The RGD sequence is the principal integrin-binding motif present within extracellular matrix (ECM) proteins such as fibronectin [50]. Compared with other immune cells, macrophages are more dependent on continuous interaction with the ECM through integrin-fibronectin (or its RGD motif) to develop a specific phenotype and function [51]. Importantly, the ECM is extensively remodeled during inflammation and provides critical signals for macrophage migration and activation to resolve inflammation and induce wound healing [52]. A cell adhesion protein containing an RGD motif can promote M2 polarization by activating macrophage integrins with its RGD motif, suggesting that RGD-integrin-induced cellular signaling regulates M2 polarization [32]. Similarly, the RGD motif facilitates M2 polarization of macrophages in an M2-favorable environment [53,54]. We recently showed that THP-1 cells cultured on RGD- and polydopamine-coated micropatterns can induce THP-1 cell differentiation into Mregs-like cells in the absence of cytokines [40]. Integrin is a RGD counterpart that is also involved in M2 polarization. Increased expression of the integrins α4 [55] and β3 [33] is associated with M2-like characteristics. Furthermore, M2 polarization is dependent on integrin β3 through the transcription factor peroxisome proliferator-activated receptorγ (PPARγ), which interacts with VDRs [33]. Taken together, these results suggest that the RGD motif facilitates the Mregs polarization of PMA-induced macrophages in a vitD3-induced anti-inflammatory environment.
Regulatory macrophages exert immunosuppressive effects in human kidney transplantation. Hutchinson et al. found that administering donor Mregs to kidney recipients dampens the allogeneic rejection response, thus minimizing the need for conventional immunosuppressive therapy [41,42]. One mechanism of allograft acceptance by recipients is Mregs-induced Treg generation [15,21]. Macrophages mainly reside in the medullary cords of lymph nodes that are adjacent to the paracortical area where most T cells reside. Therefore, they probably interact intimately and influence each other in terms of functional activity. Macrophages that induce regulatory T cells are assumed to be Mregs [56]). In turn, interactions between Tregs and macrophages can result in polarization of the latter into regulatory phenotypes [57]. Thus, the ability of Mregs to suppress T-cell proliferation might be due to their effects on Treg expansion and immunosuppressive cytokine production [14,15].
5. Conclusions
We showed that sequential incubations with PMA, RGD and vitD3 could generate human Mregs from THP-1 monocytes. Our Mregs express Mregs markers and have both anti-inflammatory and immunosuppressive properties. Our method is simple and cost-effective because it does not require cytokines. However, our study has some limitations. We did not evaluate the capacity of Mregs to induce Treg expansion and used an established cell line instead of primary cells to generate Mregs. Thus, further studies are needed to elucidate the epigenetic and metabolic reprogramming that occurs during macrophage polarization to determine the mechanisms underlying Mregs induction. In addition, Mregs should be established using primary human monocytes to evaluate their possible applications to various inflammatory states, including endotoxemia and organ transplantation.
Author Contributions
HLP designed the study, performed experiments, statistical analysis, and drafted the manuscript. TXH designed the study, interpreted the data and reviewed the draft of the manuscript. JYK designed the study, interpreted the data, supported the research, and finalized the manuscript.
Funding
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2022R1F1A1076227). This research was supported by the Institute of Civil Military Technology Cooperation funded by the Defense Acquisition Program Administration and Ministry of Trade, Industry and Energy of Korean government under grant No. 22-CM-EC-18.
Institutional Review Board Statement
No ethics approval was required for this work as it was performed in vitro using a cell line and no primary human samples were used.
Informed Consent Statement
All authors consent to the publication of this study.
Acknowledgments
This study was partly based on the first author’s Master’s thesis submitted in 2021 to Gachon University in Korea.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M. The many faces of macrophage activation. J. Leukoc. Biol. 2003, 73, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Nau, G.J.; Richmond, J.F.L.; Schlesinger, A.; Jennings, E.G.; Lander, E.S.; Young, R.A. Human macrophage activation programs induced by bacterial pathogens. Proc. Natl. Acad. Sci. 2002, 99, 1503–1508. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, P.; Ruscitti, P.; Vadasz, Z.; Toubi, E.; Giacomelli, R. Macrophages with regulatory functions, a possible new therapeutic perspective in autoimmune diseases. Autoimmun. Rev. 2019, 18, 102369. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Martinez, F.O. ScienceDirect.Com - Immunity - Alternative Activation of Macrophages: Mechanism and Functions. Immunity 2010.
- Abdelaziz, M.H.; Abdelwahab, S.F.; Wan, J.; Cai, W.; Huixuan, W.; Jianjun, C.; Kumar, K.D.; Vasudevan, A.; Sadek, A.; Su, Z.; et al. Alternatively activated macrophages; a double-edged sword in allergic asthma. J. Transl. Med. 2020, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Wang, L.X.; Zhang, S.X.; Wu, H.J.; Rong, X.L.; Guo, J. M2b macrophage polarization and its roles in diseases. J. Leukoc. Biol. 2019, 106, 345–358. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, J.; Cao, P.; Sun, Z.; Wang, W. The characteristics of regulatory macrophages and their roles in transplantation. Int. Immunopharmacol. 2021, 91, 107322. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage Plasticity, Polarization, and Function in Health and Disease. J Cell Physiol 2018, 233. [Google Scholar] [CrossRef] [PubMed]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [PubMed]
- Fleming, B.D.; Mosser, D.M. Regulatory macrophages: Setting the Threshold for Therapy. Eur. J. Immunol. 2011, 41, 2498–2502. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, P.; Haarer, J.; Kammler, A.; Walter, L.; Tomiuk, S.; Ahrens, N.; Wege, A.K.; Goecze, I.; Zecher, D.; Banas, B.; et al. TIGIT+ iTregs elicited by human regulatory macrophages control T cell immunity. Nat. Commun. 2018, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Vos, A.C.W.; Wildenberg, M.E.; Arijs, I.; Duijvestein, M.; Verhaar, A.P.; de Hertogh, G.; Vermeire, S.; Rutgeerts, P.; van den Brink, G.R.; Hommes, D.W. Regulatory macrophages induced by infliximab are involved in healing in vivo and in vitro. Inflamm. Bowel Dis. 2012, 18, 401–408. [Google Scholar] [CrossRef] [PubMed]
- E Wildenberg, M.; Levin, A.D.; Ceroni, A.; Guo, Z.; Koelink, P.J.; Hakvoort, T.B.M.; Westera, L.; Bloemendaal, F.M.; Brandse, J.F.; Simmons, A.; et al. Benzimidazoles Promote Anti-TNF Mediated Induction of Regulatory Macrophages and Enhance Therapeutic Efficacy in a Murine Model. J. Crohn’s Colitis 2017, 11, 1480–1490. [Google Scholar] [CrossRef] [PubMed]
- Koelink, P.J.; Bloemendaal, F.M.; Li, B.; Westera, L.; Vogels, E.W.M.; van Roest, M.; Gloudemans, A.K.; Wout, A.B.v. '.; Korf, H.; Vermeire, S.; et al. Anti-TNF therapy in IBD exerts its therapeutic effect through macrophage IL-10 signalling. Gut 2019, 69, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.M.; Belew, A.T.; El-Sayed, N.M.; Tafuri, W.L.; Silveira, F.T.; Mosser, D.M. Host and parasite responses in human diffuse cutaneous leishmaniasis caused by L. amazonensis. PLOS Neglected Trop. Dis. 2018, 13, e0007152. [Google Scholar] [CrossRef]
- Conde, P.; Rodriguez, M.; van der Touw, W.; Jimenez, A.; Burns, M.; Miller, J.; Brahmachary, M.; Chen, H.-M.; Boros, P.; Rausell-Palamos, F.; et al. DC-SIGN+ Macrophages Control the Induction of Transplantation Tolerance. Immunity 2015, 42, 1143–1158. [Google Scholar] [CrossRef]
- Hutchinson, J.A.; Riquelme, P.; Sawitzki, B.; Tomiuk, S.; Miqueu, P.; Zuhayra, M.; Oberg, H.H.; Pascher, A.; Lützen, U.; Janßen, U.; et al. Cutting Edge: Immunological Consequences and Trafficking of Human Regulatory Macrophages Administered to Renal Transplant Recipients. J. Immunol. 2011, 187, 2072–2078. [Google Scholar] [CrossRef]
- Riquelme, P.; Amodio, G.; Macedo, C.; Moreau, A.; Obermajer, N.; Brochhausen, C.; Ahrens, N.; Kekarainen, T.; Fändrich, F.; Cuturi, C.; et al. DHRS9 Is a Stable Marker of Human Regulatory Macrophages. Transplantation 2017, 101, 2731–2738. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.D. Vitamin D Regulation of Immune Function; 2011; Vol. 86;
- Haussler, M.R.; Jurutka, P.W.; Mizwicki, M.; Norman, A.W. Vitamin D Receptor (VDR)-Mediated Actions of 1α,25(OH) 2 Vitamin D 3 : Genomic and Non-Genomic Mechanisms. Best Pract Res Clin Endocrinol Metab 2011, 25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Leung, D.Y.M.; Richers, B.N.; Liu, Y.; Remigio, L.K.; Riches, D.W.; Goleva, E. Vitamin D Inhibits Monocyte/Macrophage Proinflammatory Cytokine Production by Targeting MAPK Phosphatase-1. J. Immunol. 2012, 188, 2127–2135. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Jung, J.H.; Kim, J.Y. 1,25-Dihydroxyvitamin D3 up-regulates TLR10 while down-regulating TLR2, 4, and 5 in human monocyte THP-1. J. Steroid Biochem. Mol. Biol. 2014, 141, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhou, M.; Guo, Y.; Song, Z.; Liu, B. 1,25-Dihydroxyvitamin D3Promotes High Glucose-Induced M1 Macrophage Switching to M2 via the VDR-PPARγSignaling Pathway. BioMed Res. Int. 2015, 2015, 1–14. [Google Scholar] [CrossRef]
- Liang, S.; Cai, J.; Li, Y.; Yang, R. 1,25-Dihydroxy-Vitamin D3 induces macrophage polarization to M2 by upregulating T-cell Ig-mucin-3 expression. Mol. Med. Rep. 2019, 19, 3707–3713. [Google Scholar] [CrossRef]
- Verma, R.; Kim, J.Y. 1,25-Dihydroxyvitamin D3 Facilitates M2 Polarization and Upregulates TLR10 Expression on Human Microglial Cells. Neuroimmunomodulation 2016, 23, 75–80. [Google Scholar] [CrossRef]
- Barczyk, M.; Carracedo, S.; Gullberg, D. Integrins. Cell Tissue Res. 2010, 339, 269–280. [Google Scholar] [CrossRef]
- Mezu-Ndubuisi, O.J.; Maheshwari, A. The role of integrins in inflammation and angiogenesis. Pediatr. Res. 2020, 89, 1619–1626. [Google Scholar] [CrossRef]
- Soki, F.N.; Koh, A.J.; Jones, J.D.; Kim, Y.W.; Dai, J.; Keller, E.T.; Pienta, K.J.; Atabai, K.; Roca, H.; McCauley, L.K. Polarization of Prostate Cancer-associated Macrophages Is Induced by Milk Fat Globule-EGF Factor 8 (MFG-E8)-mediated Efferocytosis. J. Biol. Chem. 2014, 289, 24560–24572. [Google Scholar] [CrossRef]
- Shu, Y.; Qin, M.; Song, Y.; Tang, Q.; Huang, Y.; Shen, P.; Lu, Y. M2 polarization of tumor-associated macrophages is dependent on integrin β3 via peroxisome proliferator-activated receptor-γ up-regulation in breast cancer. Immunology 2020, 160, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kim, J.Y.; Koh, H.S.; Lee, J.P.; Kim, Y.; Kang, H.J.; Hwang, W.S.; Ahn, C. Establishment and characterization of endothelial cell lines from the aorta of miniature pig for the study of xenotransplantation. Cell Biol. Int. 2005, 29, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; So, E.C.; Strome, S.E.; Zhang, X. Impact of Detachment Methods on M2 Macrophage Phenotype and Function. J. Immunol. Methods 2015, 426, 56–61. [Google Scholar] [CrossRef] [PubMed]
- McWhorter, F.Y.; Wang, T.; Nguyen, P.; Chung, T.; Liu, W.F. Modulation of macrophage phenotype by cell shape. Proc. Natl. Acad. Sci. 2013, 110, 17253–17258. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Locati, M. Orchestration of macrophage polarization. Blood 2009, 114, 3135–3136. [Google Scholar] [CrossRef]
- Galili, U. Interaction of the natural anti-Gal antibody with α-galactosyl epitopes: a major obstacle for xenotransplantation in humans. Immunol. Today 1993, 14, 480–482. [Google Scholar] [CrossRef]
- Kim, J.Y. Macrophages in Xenotransplantation. Korean Journal of Transplantation 2019, 33. [Google Scholar] [CrossRef]
- Pham, H.L.; Yang, D.H.; Chae, W.R.; Jung, J.H.; Hoang, T.X.; Lee, N.Y.; Kim, J.Y. PDMS Micropatterns Coated with PDA and RGD Induce a Regulatory Macrophage-like Phenotype. Micromachines 2023, 14, 673. [Google Scholar] [CrossRef]
- Hutchinson, J.A.; Riquelme, P.; Brem-Exner, B.G.; Schulze, M.; Matthäi, M.; Renders, L.; Kunzendorf, U.; Geissler, E.K.; Fändrich, F. Transplant acceptance-inducing cells as an immune-conditioning therapy in renal transplantation. Transpl. Int. 2008, 21, 728–741. [Google Scholar] [CrossRef]
- Hutchinson, J.A.; Brem-Exner, B.G.; Riquelme, P.; Roelen, D.; Schulze, M.; Ivens, K.; Grabensee, B.; Witzke, O.; Philipp, T.; Renders, L.; et al. A cell-based approach to the minimization of immunosuppression in renal transplantation. Transpl. Int. 2008, 21, 742–754. [Google Scholar] [CrossRef]
- Moreau, A.; Varey, E.; Bouchet-Delbos, L.; Cuturi, M.-C. Cell therapy using tolerogenic dendritic cells in transplantation. Transplant. Res. 2012, 1, 13–13. [Google Scholar] [CrossRef] [PubMed]
- Macedo, C.; Turnquist, H.R.; Castillo-Rama, M.; Zahorchak, A.F.; Shapiro, R.; Thomson, A.W.; Metes, D. Rapamycin Augments Human DC IL-12p70 and IL-27 Secretion to Promote Allogeneic Type1 Polarization Modulated by NK Cells. Am. J. Transplant. 2013, 13, 2322–2333. [Google Scholar] [CrossRef] [PubMed]
- Gregori, S.; Tomasoni, D.; Pacciani, V.; Scirpoli, M.; Battaglia, M.; Magnani, C.F.; Hauben, E.; Roncarolo, M.-G. Differentiation of type 1 T regulatory cells (Tr1) by tolerogenic DC-10 requires the IL-10–dependent ILT4/HLA-G pathway. Blood 2010, 116, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Obermajer, N.; Muthuswamy, R.; Lesnock, J.; Edwards, R.P.; Kalinski, P. Positive feedback between PGE2 and COX2 redirects the differentiation of human dendritic cells toward stable myeloid-derived suppressor cells. Blood 2011, 118, 5498–5505. [Google Scholar] [CrossRef] [PubMed]
- Mia, S.; Warnecke, A.; Zhang, X.; Malmström, V.; Harris, R.A. An optimized Protocol for Human M2 Macrophages using M-CSF and IL-4/IL-10/TGF-β Yields a Dominant Immunosuppressive Phenotype. Scand. J. Immunol. 2014, 79, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Foucher, E.D.; Blanchard, S.; Preisser, L.; Garo, E.; Ifrah, N.; Guardiola, P.; Delneste, Y.; Jeannin, P. Erratum: IL-34 Induces the Differentiation of Human Monocytes into Immunosuppressive Macrophages. Antagonistic Effects of GM-CSF and IFN? (PLoS ONE (2014) 9:1). PLoS One 2014, 9.
- Tsuchiya, S.; Kobayashi, Y.; Goto, Y.; Okumura, H.; Nakae, S.; Konno, T.; Tada, K. Induction of maturation in cultured human monocytic leukemia cells by a phorbol diester. . 1982, 42, 1530–6. [Google Scholar] [PubMed]
- Humphries, J.D.; Byron, A.; Humphries, M.J. Integrin ligands at a glance. J. Cell Sci. 2006, 119, 3901–3903. [Google Scholar] [CrossRef]
- Jacob, S.S.; Sudhakaran, P. Molecular Mechanism Involved in Matrix Dependent Upregulation of Matrix Metalloproteinases in Monocyte/Macrophage. J. Biochem. Mol. Biol. Biophys. 2002, 6, 335–340. [Google Scholar] [CrossRef]
- Wynn, T.A.; Vannella, K.M. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef]
- Kang, H.; Jung, H.J.; Kim, S.K.; Wong, D.S.H.; Lin, S.; Li, G.; Dravid, V.P.; Bian, L. Magnetic Manipulation of Reversible Nanocaging Controls In Vivo Adhesion and Polarization of Macrophages. ACS Nano 2018, 12, 5978–5994. [Google Scholar] [CrossRef]
- Wu, L.; Kim, Y.; Seon, G.M.; Choi, S.H.; Park, H.C.; Son, G.; Kim, S.M.; Lim, B.-S.; Yang, H.-C. Effects of RGD-grafted phosphatidylserine-containing liposomes on the polarization of macrophages and bone tissue regeneration. Biomaterials 2021, 279, 121239. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, M.M.; Messer, K.S.; Ralainirina, N.; Li, H.; Leem, C.J.; Gorjestani, S.; Woo, G.; Nguyen, A.V.; Figueiredo, C.C.; Foubert, P.; et al. PI3Kγ is a molecular switch that controls immune suppression. Nature 2016, 539, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Tiemessen, M.M.; Jagger, A.L.; Evans, H.G.; van Herwijnen, M.J.C.; John, S.; Taams, L.S. CD4+CD25+Foxp3+ regulatory T cells induce alternative activation of human monocytes/macrophages. Proc. Natl. Acad. Sci. USA 2007, 104, 19446–19451. [Google Scholar] [CrossRef] [PubMed]
- Gosselin, D.; Link, V.M.; Romanoski, C.E.; Fonseca, G.J.; Eichenfield, D.Z.; Spann, N.J.; Stender, J.D.; Chun, H.B.; Garner, H.; Geissmann, F.; et al. Environment Drives Selection and Function of Enhancers Controlling Tissue-Specific Macrophage Identities. Cell 2014, 159, 1327–1340. [Google Scholar] [CrossRef]
Figure 1.
Schema of procedures to generate different types of macrophages using human THP-1 monocytes.
Figure 1.
Schema of procedures to generate different types of macrophages using human THP-1 monocytes.
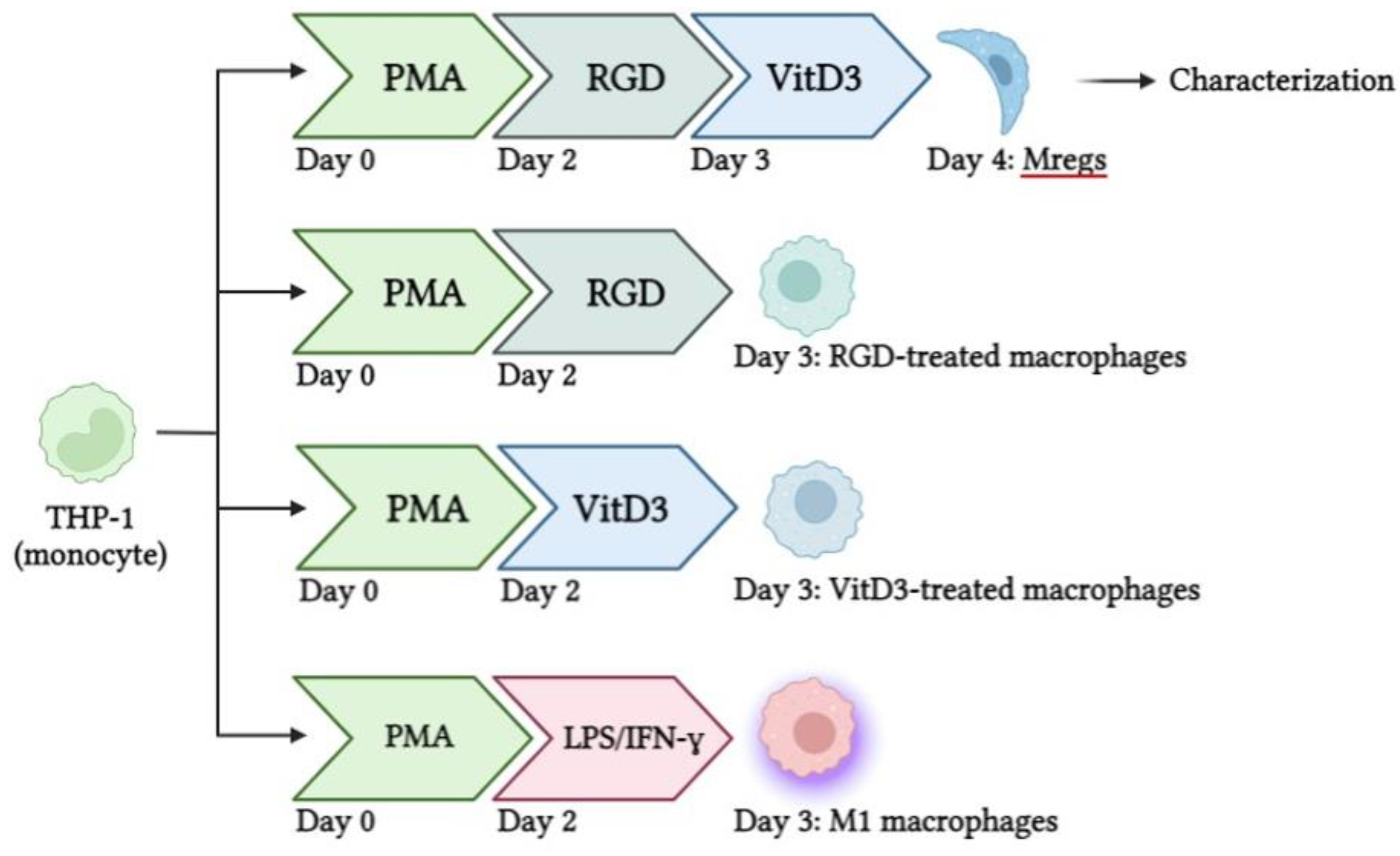
Figure 2.
Incubation with PMA, RGD, and vitD3 induces THP-1 cell elongation. We incubated THP-1 cells with PMA (10 ng/mL) for 2 days, followed by various concentrations of RGD (A) or vitD3 (B) for 1 day. Cells were incubated with PMA (10 ng/mL) for 2 days, followed by RGD (1 μg/mL), then 1 μM vitD3 for 1 day (PMA + RGD + VitD3 group; (C). Cell morphology was visualized using inverted microscope at 100 ✕ magnification.
Figure 2.
Incubation with PMA, RGD, and vitD3 induces THP-1 cell elongation. We incubated THP-1 cells with PMA (10 ng/mL) for 2 days, followed by various concentrations of RGD (A) or vitD3 (B) for 1 day. Cells were incubated with PMA (10 ng/mL) for 2 days, followed by RGD (1 μg/mL), then 1 μM vitD3 for 1 day (PMA + RGD + VitD3 group; (C). Cell morphology was visualized using inverted microscope at 100 ✕ magnification.
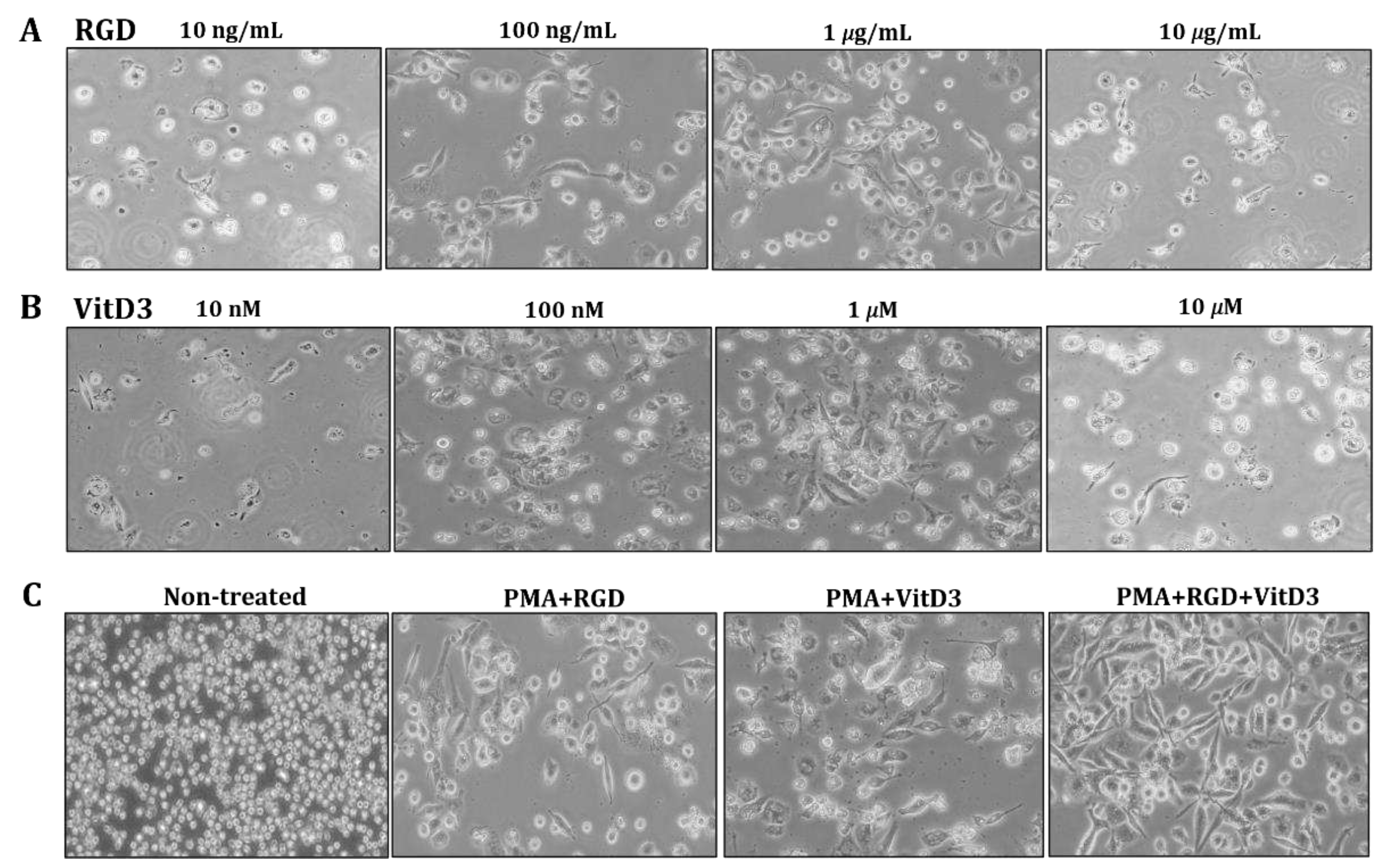
Figure 3.
Incubation with PMA, RGD, and vitD3 upregulated IL-10 and downregulated IL-12 in THP-1 cells. We incubated THP-1 cells with 10 ng/mL PMA for 2 days, washed them with DPBS, then maintained them in RPMI-1640 media for 2 days (PMA group). Cells were incubated with PMA (10 ng/mL) for 2 days, followed by RGD (1 μg/mL; PMA + RGD) or vitD3 (1 μM; PMA + VitD3) for 1 day. Cells were washed with DPBS, then maintained in RPMI-1640 media for 1 day. We incubated cells with PMA (10 ng/mL) for 2 days, followed sequentially by RGD (1 μg/mL) for 1 day and vitD3 (1 μM) for 1 day (PMA + RGD + VitD3). Cells were harvested and mRNA expression of anti-inflammatory IL-10 (A) and pro-inflammatory IL-12 (B) was quantified by qRT-PCR. *p < 0.05, **p < 0.005 vs. controls incubated with DMSO; #p < 0.05, ##p < 0.005.
Figure 3.
Incubation with PMA, RGD, and vitD3 upregulated IL-10 and downregulated IL-12 in THP-1 cells. We incubated THP-1 cells with 10 ng/mL PMA for 2 days, washed them with DPBS, then maintained them in RPMI-1640 media for 2 days (PMA group). Cells were incubated with PMA (10 ng/mL) for 2 days, followed by RGD (1 μg/mL; PMA + RGD) or vitD3 (1 μM; PMA + VitD3) for 1 day. Cells were washed with DPBS, then maintained in RPMI-1640 media for 1 day. We incubated cells with PMA (10 ng/mL) for 2 days, followed sequentially by RGD (1 μg/mL) for 1 day and vitD3 (1 μM) for 1 day (PMA + RGD + VitD3). Cells were harvested and mRNA expression of anti-inflammatory IL-10 (A) and pro-inflammatory IL-12 (B) was quantified by qRT-PCR. *p < 0.05, **p < 0.005 vs. controls incubated with DMSO; #p < 0.05, ##p < 0.005.
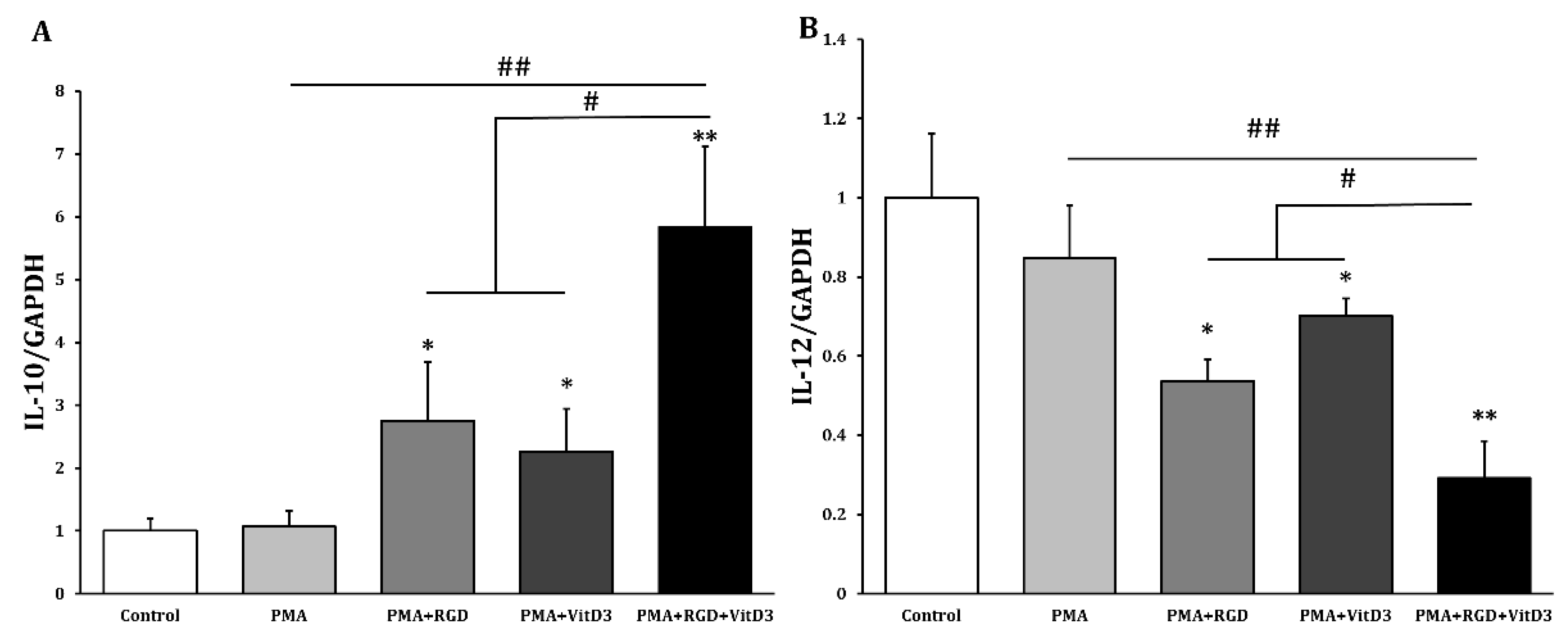
Figure 4.
Phenotypic characterization of PMA/RGD/vitD3-induced Mregs-like cells. We generated resting macrophages by incubating THP-1 cells with PMA (10 ng/mL) for 2 days, washing them with DPBS, then maintaining them in RPMI-1640 media for 2 days (resting macrophages group). Cells were incubated with PMA (10 ng/mL) for 2 days, followed by RGD (1 μg/mL) for 1 day, then with vitD3 (1 μM) for 1 day to generate Mregs. Cells were harvested and expression of cell surface or intracellular markers was assessed using flow cytometry. (A) Black lines, red lines, and gray shadows on representative flow cytometric histograms respectively indicate resting macrophages, Mregs, and controls. (B) Bar graphs show means of relative protein expression ± SD. *p < 0.05, **p < 0.005 vs. DMSO-incubated controls. #p < 0.05, ##p < 0.005.
Figure 4.
Phenotypic characterization of PMA/RGD/vitD3-induced Mregs-like cells. We generated resting macrophages by incubating THP-1 cells with PMA (10 ng/mL) for 2 days, washing them with DPBS, then maintaining them in RPMI-1640 media for 2 days (resting macrophages group). Cells were incubated with PMA (10 ng/mL) for 2 days, followed by RGD (1 μg/mL) for 1 day, then with vitD3 (1 μM) for 1 day to generate Mregs. Cells were harvested and expression of cell surface or intracellular markers was assessed using flow cytometry. (A) Black lines, red lines, and gray shadows on representative flow cytometric histograms respectively indicate resting macrophages, Mregs, and controls. (B) Bar graphs show means of relative protein expression ± SD. *p < 0.05, **p < 0.005 vs. DMSO-incubated controls. #p < 0.05, ##p < 0.005.
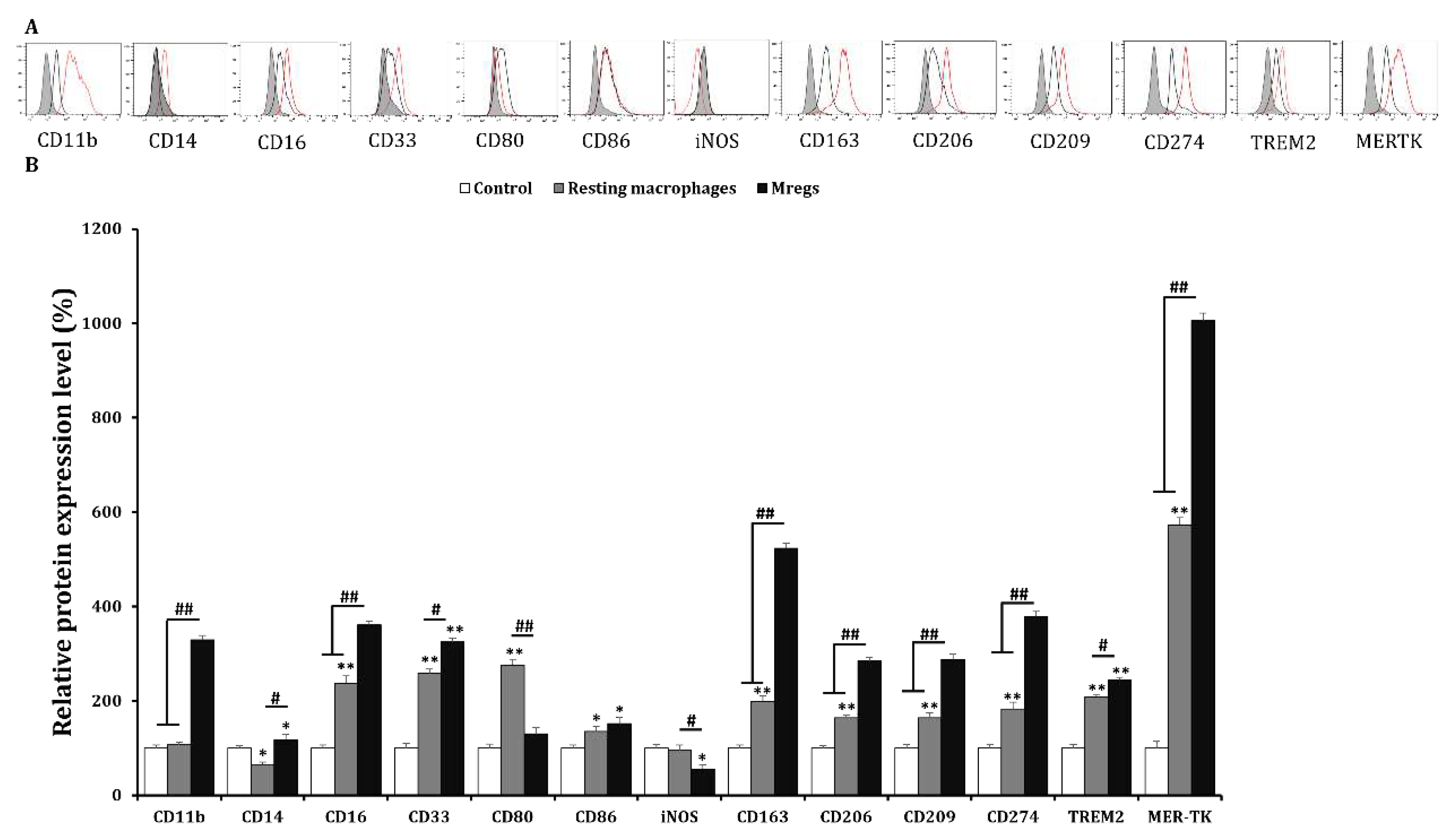
Figure 5.
DHRS9 expression in Mregs. (A) and (B) cells were incubated with PMA (10 ng/mL) for 2 days, washed with DPBS, then maintained in RPMI-1640 media for 2 days (PMA group). Cells were incubated with PMA (10 ng/mL) for 2 days, followed by (RGD 1 μg/mL; PMA+ RGD group) or vitD3 (1 μM; PMA + VitD3 group) for 1 day, washed with DPBS, then maintained in RPMI-1640 media for 1 day. Cells were incubated with PMA (10 ng/mL) for 2 days, followed by RGD (1 μg/mL) for 1 day, then 1 μM vitD3 for 1 day (PMA + RGD + VitD3). Cells were harvested and DHRS9 expression was quantified using flow cytometry. (A) Blue, green, black, and red lines in representative flow cytometric histograms indicate PMA, PMA + RGD, PMA + VitD3, and PMA + RGD + VitD3 (Mregs), respectively. Gray shadows indicate control. (B) Bar graphs show means of relative DHRS9 expression ± SD. (C) and (D) Mregs were incubated with IFN-γ (25 ng/mL) for 24 h and DHRS9 expression was measured using flow cytometry. (C) Red and black lines in representative flow cytometric histograms indicate Mregs and Mregs + IFN-γ, respectively. Gray shadow indicates control. (D) Bar graphs show means of relative DHRS9 expression ± SD. *p < 0.05, **p < 0.005 vs. controls incubated with DMSO; #p < 0.05, ##p < 0.005.
Figure 5.
DHRS9 expression in Mregs. (A) and (B) cells were incubated with PMA (10 ng/mL) for 2 days, washed with DPBS, then maintained in RPMI-1640 media for 2 days (PMA group). Cells were incubated with PMA (10 ng/mL) for 2 days, followed by (RGD 1 μg/mL; PMA+ RGD group) or vitD3 (1 μM; PMA + VitD3 group) for 1 day, washed with DPBS, then maintained in RPMI-1640 media for 1 day. Cells were incubated with PMA (10 ng/mL) for 2 days, followed by RGD (1 μg/mL) for 1 day, then 1 μM vitD3 for 1 day (PMA + RGD + VitD3). Cells were harvested and DHRS9 expression was quantified using flow cytometry. (A) Blue, green, black, and red lines in representative flow cytometric histograms indicate PMA, PMA + RGD, PMA + VitD3, and PMA + RGD + VitD3 (Mregs), respectively. Gray shadows indicate control. (B) Bar graphs show means of relative DHRS9 expression ± SD. (C) and (D) Mregs were incubated with IFN-γ (25 ng/mL) for 24 h and DHRS9 expression was measured using flow cytometry. (C) Red and black lines in representative flow cytometric histograms indicate Mregs and Mregs + IFN-γ, respectively. Gray shadow indicates control. (D) Bar graphs show means of relative DHRS9 expression ± SD. *p < 0.05, **p < 0.005 vs. controls incubated with DMSO; #p < 0.05, ##p < 0.005.
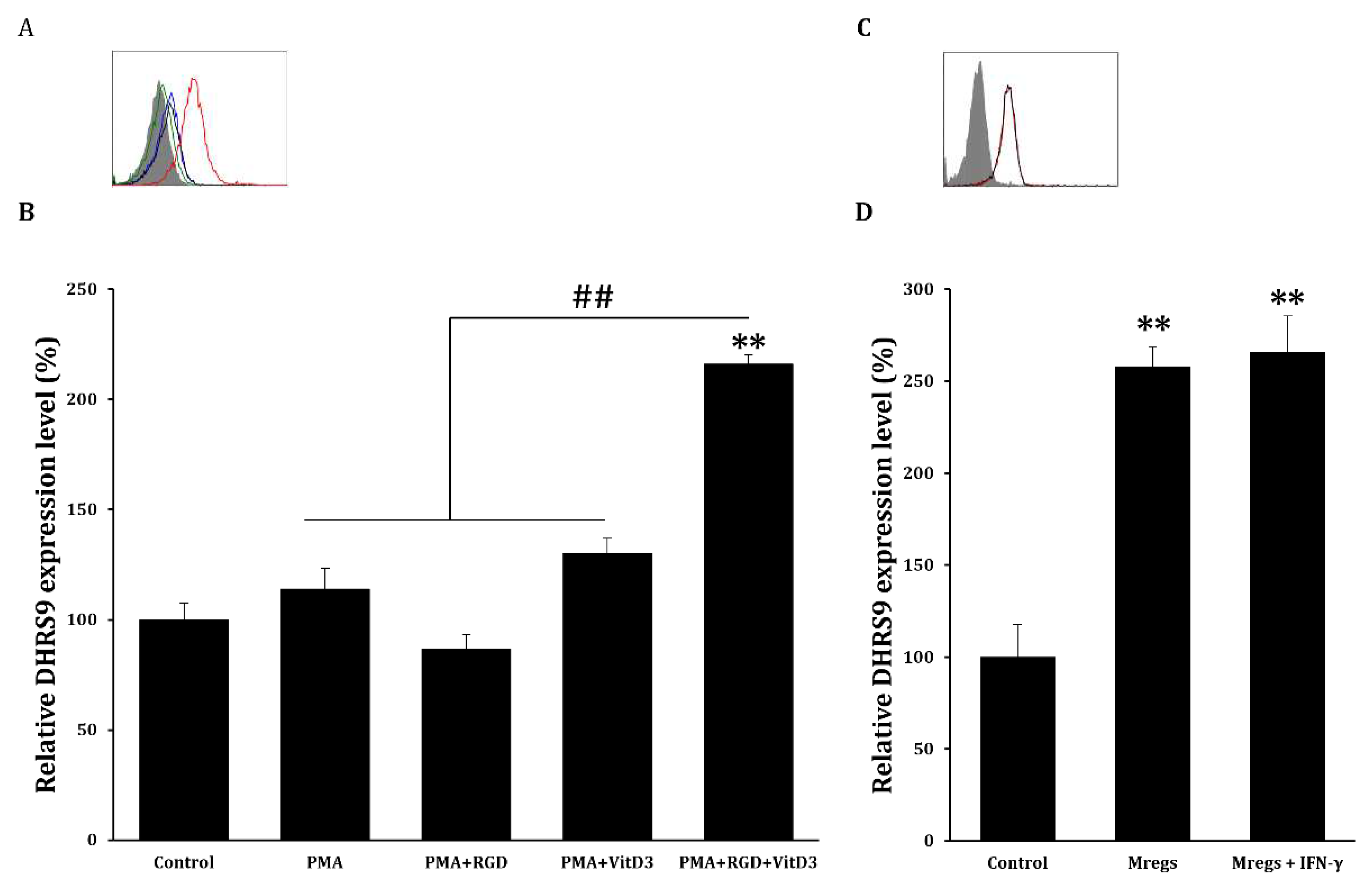
Figure 6.
Marker expression of Mregs does not change upon exposure to LPS. (A‒D) We generated Mregs as described in Figure 5. We stimulated THP-1 cells or Mregs with LPS (100 ng/mL) for 24 h, then quantified expression of cell surface markers using flow cytometry. (A) and (C), Red and black lines representative flow cytometric histograms, Mregs and Mregs + LPS, respectively. Pink lines and gray shadow, THP-1 + LPS and controls, respectively. (B) and (D), Bar graphs show means of relative protein expression ± SD. *p < 0.05, **p < 0.005 vs. controls incubated with DMSO; #p < 0.05, ## p < 0.005.
Figure 6.
Marker expression of Mregs does not change upon exposure to LPS. (A‒D) We generated Mregs as described in Figure 5. We stimulated THP-1 cells or Mregs with LPS (100 ng/mL) for 24 h, then quantified expression of cell surface markers using flow cytometry. (A) and (C), Red and black lines representative flow cytometric histograms, Mregs and Mregs + LPS, respectively. Pink lines and gray shadow, THP-1 + LPS and controls, respectively. (B) and (D), Bar graphs show means of relative protein expression ± SD. *p < 0.05, **p < 0.005 vs. controls incubated with DMSO; #p < 0.05, ## p < 0.005.
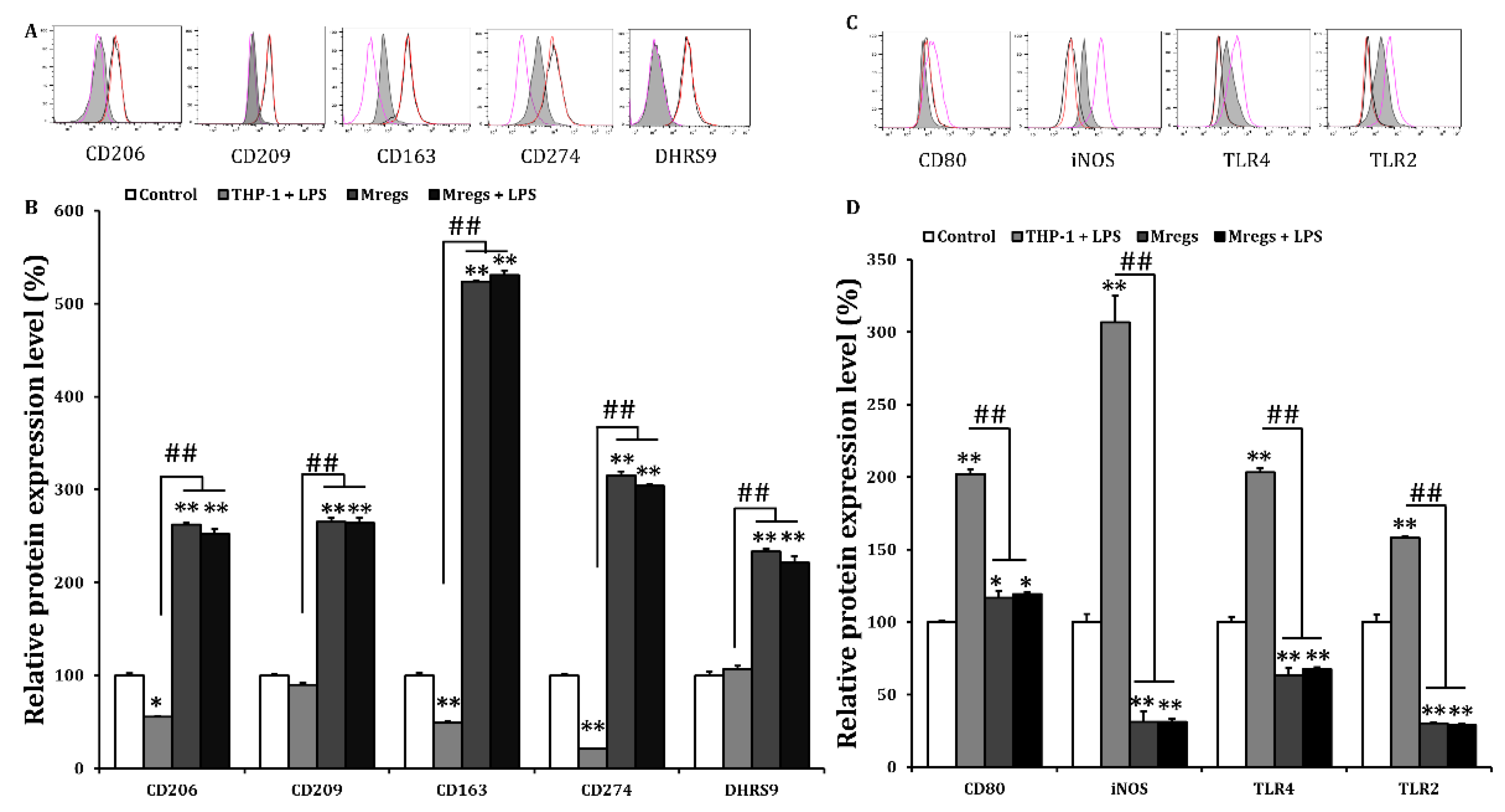
Figure 7.
Marker expression of Mregs does not change upon exposure to xenogeneic antigen. (A‒D) We generated Mregs as described in Figure 5. We stimulated THP-1 cells or Mregs with α-gal (100 ng/mL) for 24 h, then quantified expression of cell surface markers using flow cytometry. (A) and (C) Representative flow cytometric histograms; red and black lines, Mregs and Mregs + α-gal, respectively. Pink lines and gray shadow, THP-1 + α-gal and controls, respectively. (B) and (D) Means of relative protein expression ± SD. *p < 0.05, **p < 0.005 vs. controls incubated with DMSO; #p < 0.05, ##p < 0.005.
Figure 7.
Marker expression of Mregs does not change upon exposure to xenogeneic antigen. (A‒D) We generated Mregs as described in Figure 5. We stimulated THP-1 cells or Mregs with α-gal (100 ng/mL) for 24 h, then quantified expression of cell surface markers using flow cytometry. (A) and (C) Representative flow cytometric histograms; red and black lines, Mregs and Mregs + α-gal, respectively. Pink lines and gray shadow, THP-1 + α-gal and controls, respectively. (B) and (D) Means of relative protein expression ± SD. *p < 0.05, **p < 0.005 vs. controls incubated with DMSO; #p < 0.05, ##p < 0.005.
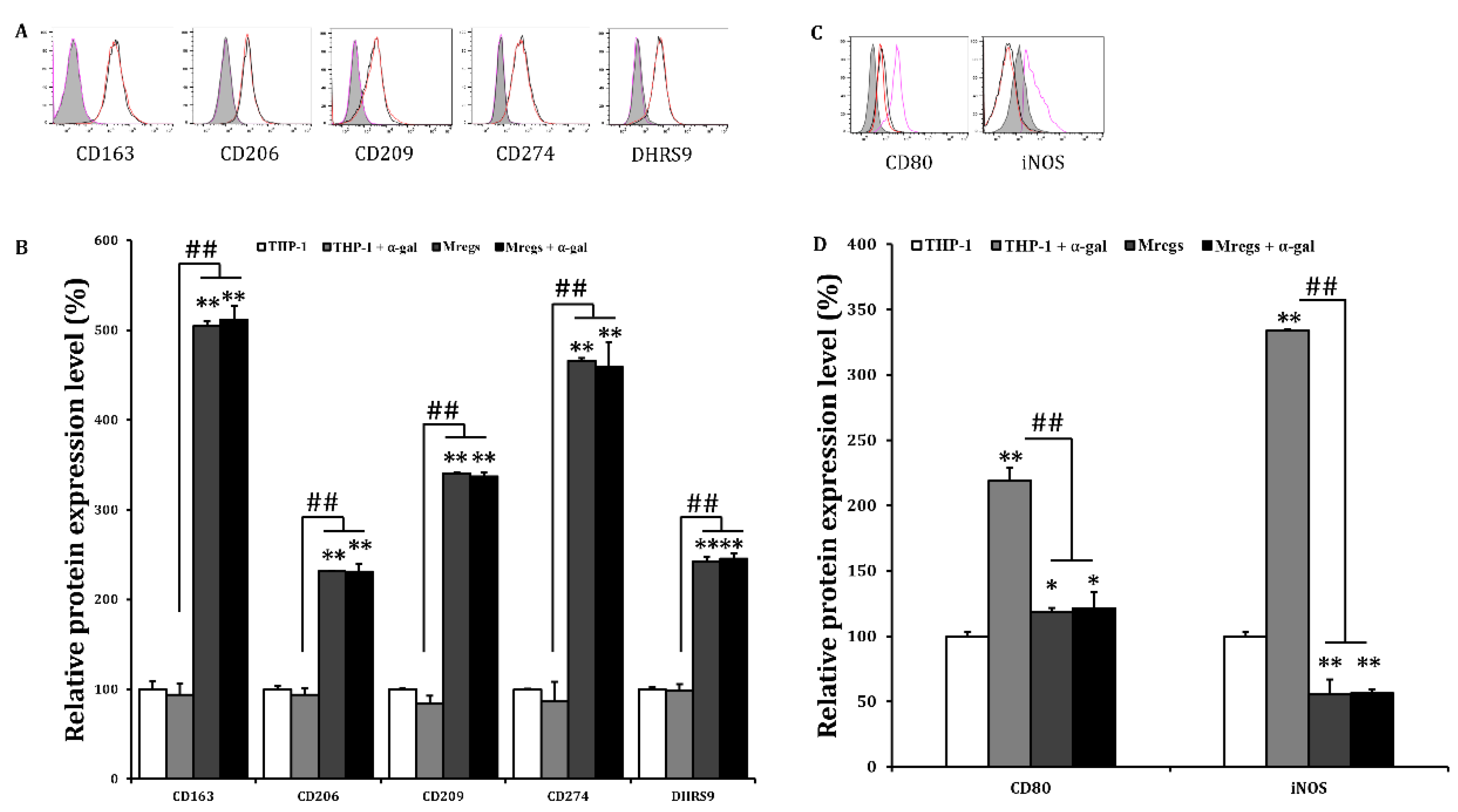
Figure 8.
Regulatory macrophages express less inflammatory and coagulation-related genes and more immunosuppressive genes in response to pig endothelial cell line MPN3. We generated M1 macrophages by incubating THP-1 cells with LPS (100 ng/mL) and IFN-γ (20 ng/mL) for 24 h, and generated Mregs as described in Figure 5. Thereafter, MPN-3 cells were co-cultured with THP-1, M1, or Mregs for 2 h (fgl-2, and TF), 6 h (IDO, IL-1β, IL-6, IL-10, IL-12, iNOS, and TNF-α), or 12 h (PAR-1 and MCP-1). Cells were harvested and mRNA expression of cytokines and chemokines (A), as well as coagulation-related genes (B) were evaluated using qRT-PCR. *p < 0.05, **p < 0.005 vs. MPN3 + THP-1; #p < 0.05, ##p < 0.005.
Figure 8.
Regulatory macrophages express less inflammatory and coagulation-related genes and more immunosuppressive genes in response to pig endothelial cell line MPN3. We generated M1 macrophages by incubating THP-1 cells with LPS (100 ng/mL) and IFN-γ (20 ng/mL) for 24 h, and generated Mregs as described in Figure 5. Thereafter, MPN-3 cells were co-cultured with THP-1, M1, or Mregs for 2 h (fgl-2, and TF), 6 h (IDO, IL-1β, IL-6, IL-10, IL-12, iNOS, and TNF-α), or 12 h (PAR-1 and MCP-1). Cells were harvested and mRNA expression of cytokines and chemokines (A), as well as coagulation-related genes (B) were evaluated using qRT-PCR. *p < 0.05, **p < 0.005 vs. MPN3 + THP-1; #p < 0.05, ##p < 0.005.
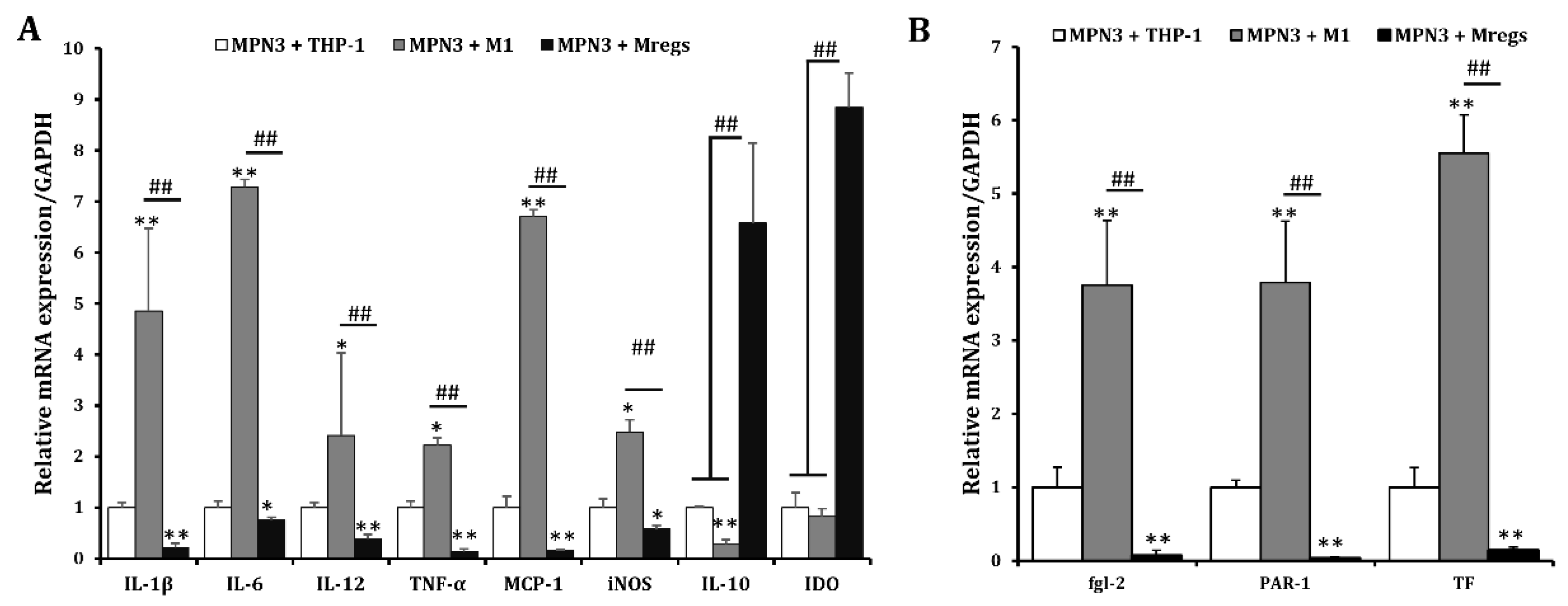
Figure 9.
Regulatory macrophages suppressed mRNA expression of pro-inflammatory and coagulation-related genes in M1 and MPN3 co-cultures according to ratios of Mregs and M1 cells. We generated M1 macrophages by incubating THP-1 cells with LPS (100 ng/mL) and IFN-γ (20 ng/mL) for 24 h, to generate Mregs as described in Figure 5. Thereafter, Mregs were cocultured them at indicated ratios for 1 day. Thereafter, THP-1, M1, or Mregs/M1 cocultures were incubated with MPN-3 cells for 2 h (fgl-2, and TF), 6 h (IDO, IL-1β, IL-6, IL-10, IL-12, iNOS, and TNF-α), or 12 h (PAR-1 and MCP-1). Cells were harvested and mRNA expression of cytokines and chemokines (A) and coagulation-related genes (B) were evaluated using qRT-PCR. *p < 0.05, **p < 0.005 vs. MPN3 + THP-1; #p < 0.05, ##p < 0.005.
Figure 9.
Regulatory macrophages suppressed mRNA expression of pro-inflammatory and coagulation-related genes in M1 and MPN3 co-cultures according to ratios of Mregs and M1 cells. We generated M1 macrophages by incubating THP-1 cells with LPS (100 ng/mL) and IFN-γ (20 ng/mL) for 24 h, to generate Mregs as described in Figure 5. Thereafter, Mregs were cocultured them at indicated ratios for 1 day. Thereafter, THP-1, M1, or Mregs/M1 cocultures were incubated with MPN-3 cells for 2 h (fgl-2, and TF), 6 h (IDO, IL-1β, IL-6, IL-10, IL-12, iNOS, and TNF-α), or 12 h (PAR-1 and MCP-1). Cells were harvested and mRNA expression of cytokines and chemokines (A) and coagulation-related genes (B) were evaluated using qRT-PCR. *p < 0.05, **p < 0.005 vs. MPN3 + THP-1; #p < 0.05, ##p < 0.005.
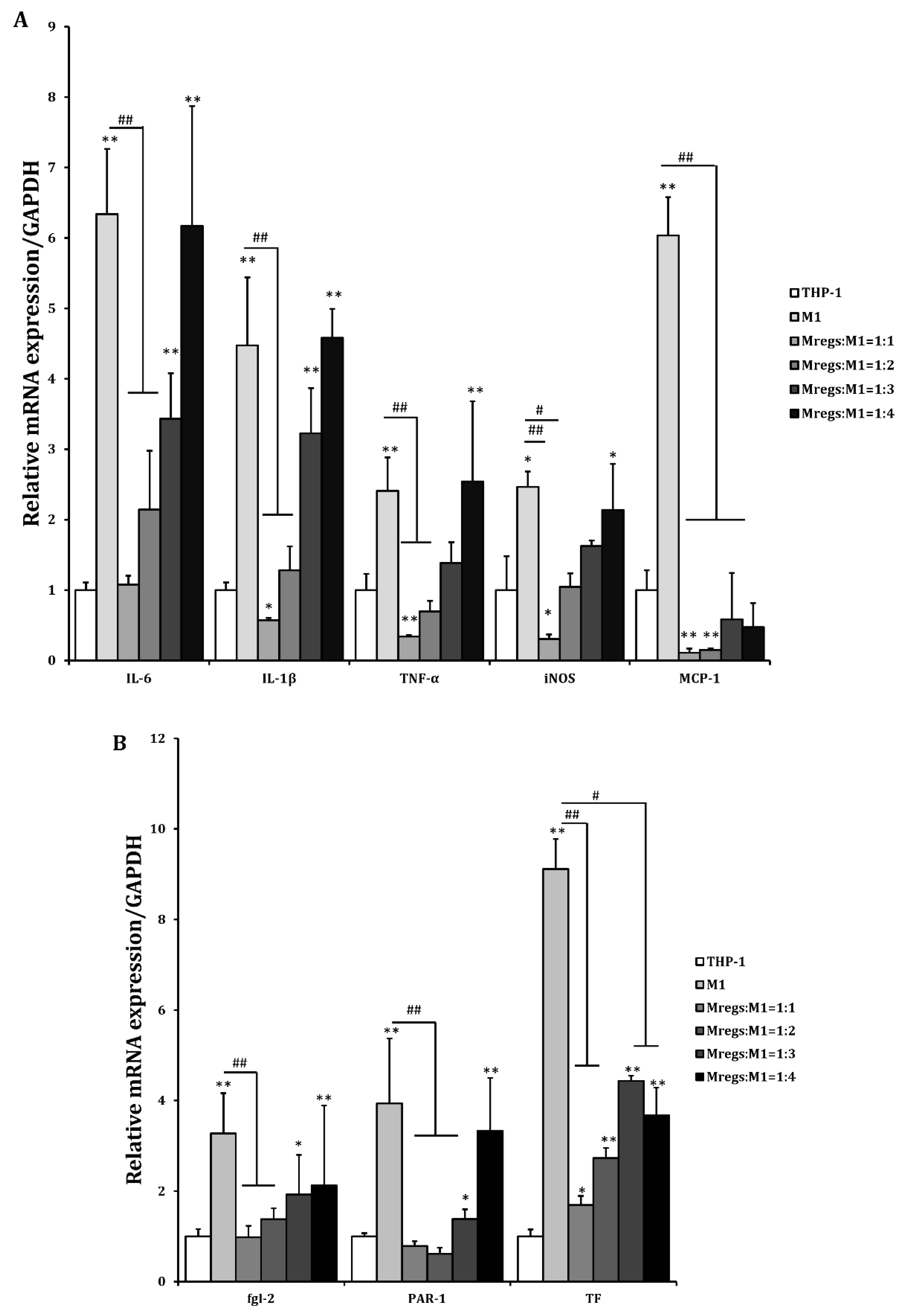
Figure 10.
Mregs suppress mitogen-induced proliferation of Jurkat T cells. We generated Mregs as described in Figure 5, then cocultured them with CFSE-labeled Jurkat T cells at ratio 1:1. Co-cultured and T cell groups (5 days) were stimulated with PHA (2 µg/mL) for 5 days, then harvested and quantified by flow cytometry. Gray shadow, black line, and red line in representative histograms show T cells on day 5, day 0, and co-cultured Mregs and T cells on day 5.
Figure 10.
Mregs suppress mitogen-induced proliferation of Jurkat T cells. We generated Mregs as described in Figure 5, then cocultured them with CFSE-labeled Jurkat T cells at ratio 1:1. Co-cultured and T cell groups (5 days) were stimulated with PHA (2 µg/mL) for 5 days, then harvested and quantified by flow cytometry. Gray shadow, black line, and red line in representative histograms show T cells on day 5, day 0, and co-cultured Mregs and T cells on day 5.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



