
Submitted:
18 May 2023
Posted:
19 May 2023
You are already at the latest version
Abstract
Forest vegetation dynamics (succession and dominance) is an ecological phenomenon that is still difficult to characterize and integrate into management practices. In this study, the understanding of forest dynamics is explored based on Grime’s theoretical triangular model of plant adaptive strategies using the example of Lithuania’s forest ecosystems. The idea behind this is the hypothesis that forest dynamics is linked to natural selection as an evolutionary process that exhibits differential species responses to competition, stress, and disturbance. The aim of this study is to explore the adaptive relationships in hemi-boreal forests. Grime’s and Pierce’s secondary CSR strategies, which describe various equilibria between competitiveness (C), stress tolerance (S), and ruderalism (R), were considered to reflect four establishment and development adaptive specialization characteristics of forest tree species. As a result of the study, four types of tree functional groups were identified: stress-resistant ruderals, competitive stress-sensitive ruderals, ruderal stress-sensitive competitors, and stress-resistant competitors. Based on this, we propose that reforestation move away from single species regeneration by implementing the maintenance of these four types of functional groups. In conclusion, forest management must consider the existence of the established equilibria between plant competitiveness, stress tolerance, and ruderalism. The formal concepts presented in this article can serve as a guide for future relevant research and development of appropriate methods for studying real forests. This study is unique in that no previous work has linked forest dynamics and natural selection in the context of Lithuania’s forest ecosystems.
Keywords:
adaptive strategies
; competitiveness
; stress tolerance
; ruderalism
; natural selection
; forest dynamics
; Lithuania
1. Introduction
Mimicking the natural forest disturbance and regeneration regimes can provide management guidelines towards sustainable forest management [1]. However, natural forest vegetation succession and dominance are an ecological phenomenon that is still difficult to characterize [2]. Integrating natural forest patterns and processes [3], such as natural disturbances, succession, and regeneration into traditional forest management adds an even greater level of complexity [4]. Understanding the manifests of natural forest disturbances and succession involves a range of stable and transitional social-ecological states which have different outcomes [5,6].
During natural disturbances, the succession of habitat patches allows the forest ecosystem to continually evolve so that it can maintain its functions or develop new ones [7,8]. Natural disturbances are usually pulse disturbances with various magnitude and frequency, but human activities tend to simplify them by through consistent pressure which leads to chronic disturbances [5]. In consequence, the disappearance of the natural forest landscape mosaics results in the loss of species that cannot cope with habitat change or do not have the capacity for long-range dispersal between suitable habitat patches. This leads to similar temporal dynamics of large areas, where the forest no longer has the necessary conditions for natural selection to occur [9].
In the process of forest stand succession, vegetation communities are transformed through a series of successional events: stand initiation after major disturbances, such as violent windthrow, fire, or clearcutting; stem exclusion, during which competition among trees is intense and density-dependent self-thinning is the major cause of mortality; understory re-initiation, when a stand undergoes demographic transition from one structural cohort to multi-cohort; and multi-aged tree community that completes the demographic transition to gap phase [10,11]. This is because plants acquire a distinctive functional organization that justifies their status as organisms through the process of natural selection [12,13,14]. Natural selection is the differential survival and reproduction due to differences in phenological type [15].
Lithuania’s hemi-boreal forests include a diverse spectrum of forest vegetation communities, but they are still at risk of being lost due to widespread even-aged clearcutting practices, i.e., the normal forest model i.e., maximum sustained yield [16], and early age harvesting by the forestry sector [8]. Subsequent re-establishment practices of deep mechanical scarification and planting of singular tree species continue to place increased pressure on Lithuania’s hemi-boreal forests as functionally integrated complex adaptive dynamical systems [7,17]. The most important potential threats of forestry to natural biodiversity in Lithuania are not only intensified cuttings, but also the loss of natural forest mosaics, the reduction in the average age of forest stands, and the distortion towards “main“ vs. “secondary“ forest types [18]. “High forestry” in Lithuania is still associated with the growing use of timber [19], the supply areas of which “depend on political decisions” [20,21].
In this study, we focus on the ecological aspects of mimicking natural hemi-boreal forest disturbances and succession to help develop knowledge for sustainable forest management by reviewing four forest dynamic characteristics: i) potential natural forest (climax) communities, ii) successional characteristics of hemi-boreal forest communities, iii) adaptive strategies of forest tree species, and iv) adaptive relationships in hemi-boreal tree communities. Adaptation affects all aspects of an organism’s life, so the relationships between plant life histories, generation times, reproductive behaviours, dispersal patterns and other emergent phenomena of forest vegetation dynamics are to be explored as life cycle events and adaptations [22]. Thus, the aim of the study is to explore the adaptive relationships in contiguous tree communities based on Grime’s theoretical triangular model of plant adaptive strategies using the example of Lithuania’s forest ecosystems.
2. Natural Forests in Lithuania
All though the tree species of the natural potential forests continue to exist in Lithuania current forest ecosystems have become simplified in both their patterns and processes due to anthropogenic forest use forest management. For instance, large-scale changes in forest cover, species survival, and composition throughout millennia have led to post-climaxes of favourable soils and sites rather than a true climatic climaxes [8,23]. Natural English oak dominant forest have become rarer due to high yield forest management. Nearly all European mesic sites of deciduous and mixed forest are unable to produce and sustain advance regeneration of oaks [24]. Even though 20% of Lithuanian forest are is suitable for English oaks, oak stands on cover a little more than 2% of the forest area [25]. The largest areas lost for other tree species are in European ash and English oak stands; Scots pine stands are the most stable, followed by black alder stands [20]. Norway spruce stands remain stable only due to the reforestation of spruce in a much larger area than it is felled.
2.1. Climate and Forest Zone
Based on the biogeographic division into districts applied in the European Union, Lithuania is in the southern periphery of the boreal biogeographic region. It occupies the transition zone between boreal and temperate forests of nemoral Europe which known as the hemi-boreal forest zone [26,27]. This zone includes the southern margin of the boreal zone, i.e., southern Scandinavia, the Baltic states, and the south-west of Finland, as well as Belarus with wedges extending eastwards into central Russia. Its current natural vegetation corresponds to the topographical (reflected in climate, soil and vegetation conditions), actual climatic (temperature, precipitation, and their seasonal distribution) and edaphic conditions (structure and texture, water balance, nutrient supply), and the native flora in the various landscapes.
Lithuania is situated in the northern part of the medium climate zone between 53°54‘ and 56°27‘ of the northern latitude. The climate in Lithuania is conditioned by zonal and azonal factors [18]. Zonal factors include the Lithuania’s territorial geographical situation and dominating carriage of air masses from the west covering the entire troposphere and the lower part of stratosphere. The features of Lithuania’s climate also depend on azonal factors: surrounding areas of land, situation of oceans and seas, absolute altitude of relief, soil characteristics, and cover of bed surface. The main local climate factors are as follows: relief and topography, surface and groundwater bodies, soils, vegetation, urbanisation elements as well as their physical status.
In view of the climatic conditions, Lithuania belongs to the zone of excessive moisture, i.e., the annual precipitation rate is higher than the evaporation rate, however, the country suffers dry seasons and even draughts almost each year [18]. Increasingly stronger and longer droughts usually repeating every 3.5 years have influence on the rapid decrease in the groundwater level. In Lithuania’s territory, moving from west to east, the continental feature of climate is increasing: the annual and daily amplitudes of temperature are increasing, winters become colder, the snow cover lasts for a longer period, and the air becomes drier.
Since for plant growth the critical factor is the water-supplying power of the soil in relation to water losses from transpiration, topographical-edaphic situation (as well as climate) play a large part in determining the drought limit of the forest. Moreover, there is a close analogy between soil development and the development of the potential natural forest (climax) formations. The distinction now commonly adopted between 'zonal' and 'azonal' soils may be recalled here. Zonal soils are the soil types of each climatic region, in the development of which climate and vegetation play the principal part; azonal soils show the predominant effect of parent material, age or topography [23].
2.2. Potential Natural Forest Communities
According to Bohn et al. [27] Europe’s forest can be characterized by their potential natural vegetation at two main levels. At level 1, Lithuania’s potential natural forests are broadly divided into two climatically based zonal formations: i) mesophytic and hygromesophytic coniferous and broadleaved-coniferous forests (D11, D12, D8), and ii) mesophytic deciduous broadleaved and coniferous-broadleaved forests (F3, F4) (Figure 1). At level 2, Lithuania’s hemi-boreal can be further categorized into eight main forest vegetation types: i) spruce forests (Picea abies) with broadleaved trees in first storey (Quercus robur, Tilia cordata, Ulmus glabra, Acer platanoides and other), where large amount of even aged birches and aspens (Betula pendula, Populus tremula) is referring to clearcutting and/or violent windthrow (D8); ii) boreal pine and hemi-boreal forests (Pinus sylvestris), partly with deciduous small-leaved tree species (Betula pendula, Betula pubescens, Populus tremula) and spruce (Picea abies) (D11); iii) hemi-boreal pine forests (Pinus sylvestris), partly with birch (Betula pendula) (D12); iv) species-rich oak-hornbeam forests (Carpinus betulus, Quercus robur, Picea abies, Betula pendula, Populus tremula, Tilia cordata, Acer platanoides, Ulmus glabra) (F3); v) lime-oak forests (Quercus robur, Tilia cordata) sometimes with maple (Acer platanoides) and elm (Ulmus glabra) (F4); vi) pine bog forests (Pinus sylvestris) (S12); vii) swamp and fen forests (Alnus glutinosa, Betula pubescens, Betula pendula, Fraxinus excelsior) (T); and viii) floodplain forests (Quercus robur, Fraxinus excelsior, Ulmus laevis, Ulmus minor, Salix fragilis, Salix alba as well as Alnus glutinosa) (U1). The last three forest vegetation types belong to azonal vegetation, determined by the specific properties of soils and water balances.
The relative distribution of Norway spruce in the hemi-boreal climate zone is driven mainly by climatic and edaphic conditions. Lithuania’s hemi-boreal spruce forests with broadleaved trees, which form the climax communities on relatively fresh to moist and base-richer soils, are characterized by varying degrees of participation of nemoral (e.g., Anemone nemorosa, Hepatica nobilis, Stellaria holostea etc.; Corylus avellana, Lonicera xylosteum, Daphne mezereum etc.) in combination with boreal herbaceous and shrub species (e.g., Oxalis acetosella, Vaccinium myrtillus, Vaccinium uliginosum, Maianthemum bifolium etc.; Sorbus aucuparia, Ribes spicatum etc.). In Oxalido-nemoroso-Piceetum/Quercetum/Fraxinetum/Populetum/Betuletum pendulae/Alnetum forest types (D8), the most important indicator species of the herb layer is Anemone nemorosa. Characteristic and widely distributed herbaceous species include Oxalis acetosella, Maianthemum bifolium, Lusula pilosa, Galeobdolon luteum and other. Corylus avellana prevails in the shrub layer; Sorbus aucuparia, Frangula alnus, Daphne mezereum, Lonicera xylosteum, Euonymus europaea are also common. In Oxalido-Piceetum/Pinetum/Populetum/Betuletum pendulae/Quercetum forest types (D8), the herb layer is dominated by Oxalis acetosella and Vaccinium myrtillus; Maianthemum bifolium, Luzula pilosa, Calamagrostis arundinacea, Solidago virgaurea, Convallaria majalis, Dryopteris carthusiana also occur very often. The Oxalido-Piceetum forest type forms the ecophysiologically optimal habitat for Norway spruce. The most important indicator species of the herb layer is Pteridium aquilinum. Sorbus aucuparia, Corylus avellana prevail in the shrub layer. The moss layer is dominated by Pleurozium schreberi and Hylocomium splendens. In Myrtillo-oxalido-Piceetum/Betuletum pendulae/Populetum/Pinetum forest types (D8), Vaccinium myrtillus and Oxalis acetosella are indicator species. The moss layer is dominated by Pleurozium schreberi and Hylocomium splendens, although Rhytidiadelphus triquetrus also occurs. Sorbus aucuparia, Frangula alnus are characteristic for the shrub layer.
Lithuania’s Scots pine forests that are distributed on the more edaphically extreme sites should be considered as edaphic climax formations dependent on special conditions of soil (e.g., very oligotrophic sand, peaty soils) or topography (e.g., steep slopes, permanent over moisture). A well-developed moss layer is characteristic for most forest types, particularly in edaphically poorer forests, such species as Hylocomium splendens, Pleurozium schreberi, Dicranum scoparium, Dicranum polysetum, Polytrichum juniperinum are commonly found. In Myrtillo-Pinetum/Piceetum/Betuletum pendulae/Populetum forest types (D11), the most constant species of the herb layer are Vaccinium myrtillus and Vaccinium vitis-idaea. In the shrub layer, Frangula alnus prevails, Sorbus aucuparia is common, Salix cinerea can also occur. In Vaccinio-myrtillo-Pinetum/Betuletum pendulae/Populetum/Piceetum forest types (D11), the herb layer is dominated by Vaccinium myrtillus and Vaccinium vitis-idaea; Festuca ovina, Calluna vulgaris, Pteridium aquilinum also occur very often. Shrub layer is sparse; Sorbus aucuparia prevails, Frangula alnus and Juniperus communis are rare. In Vaccinio-Pinetum/Betuletum pendulae forest types (D12), Vaccinium vitis-idaea as well as Calluna vulgaris are characteristic for the herb layer; they grow abundantly in exposed places. In Cladonio-Pinetum forest type (D12), lichens are dominant, especially Cladonia and Cetraria species. Vaccinium vitis-idaea and Hieracium umbellatum are characteristic for the usually weakly developed herb layer. Other important herbaceous species include Arctostaphylos uva-ursi and Calluna vulgaris. Shrub layer is very sparse; it consists of Juniperus communis.
Scots pine bogs develop on hummocks, have a thick peat layer and are very poor in species. Peat mosses form a contiguous layer with Sphagnum spp. On moist-acidic forest sites, Sphagnum magellanicum, Sphagnum recurvum, Polytrichum commune, Polytrichum strictum are common. Boreal floristic elements such as Ledum palustre, Vaccinium uliginosum and several other dwarf shrubs are frequent or even dominant [28]. In Myrtillo-sphagno-Pinetum/Betuletum pubescentis/Piceetum forest types (S12), the most constant species of the herb layer is Vaccinium myrtillus, although Vaccinium uliginosum, Vaccinium vitis-idaea, Carex lasiocarpa, Carex nigra also occur very often. The most constant moss species is Pleurozium schreberi. Shrub layer is absent; rare specimens of Frangula alnus and Salix cinerea occur. In Carico-sphagno-Pinetum/Betuletum pubescentis forest types (S12), the most important indicator species of the herb layer are Menyanthes trifoliata, Carex lasiocarpa, Vaccinium oxycoccus. Shrub layer is sparse; it consists of Frangula alnus, Salix cinerea and other. In Ledo-sphagno-Pinetum forest type (S12), the most important indicator species of the poorly developed herb layer are Ledum palustre, Eriophorum vaginatum, Calluna vulgaris, Andromeda polifolia, Vaccinium uliginosum, Vaccinium oxycoccus. Shrub layer is absent.
Lithuania’s species-rich oak-hornbeam forests, which can be regarded as climax vegetation, are common on moderately dry to moist areas [27]. In certain areas, the typical herb-rich oak-hornbeam forests can be similar in its site ecology to lowland beech forests. In Hepatico-oxalido-Quercetum/Piceetum/Carpinetum/Fagetum/Populetum/Betuletum pendulae forest types (F3), the most important indicator species of the herb layer is Hepatica nobilis; Oxalis acetosella occurs with a high frequency. Other characteristic herbaceous species include Maianthemum bifolium, Galeobdolon luteum, Stellaria holostea. In the shrub layer Corylus avellana prevails, Sorbus aucuparia is frequent. Lonicera xylosteum, Frangula alnus, Daphne mezereum, Euonymus europaea, Viburnum opulus, Rhamnus cathartica are present as well.
Lithuania’s lime-oak forests form an island in the zone of hemi-boreal spruce forests with broadleaved trees (Figure 1). In Aegopodio-Quercetum/Fraxinetum/Tilietum/Ulmetum/Populetum/Betuletum forest types (F4), Aegopodium podagraria dominates in the herb layer. The most important indicator species of the herb layer are Carex sylvatica, Ranunculus cassubicus, Paris quadrifolia, Asarum europaeum, Stachys sylvatica, Brachypodium sylvaticum. Mnium undulatum is characteristic for the sparsely developed moss layer. In the shrub layer, Corylus avellana prevails; other characteristic species include Lonicera xylosteum, Euonymus europaea, Sorbus aucuparia. Frangula alnus, Padus avium, Daphne mezereum are present as well. In Carico-mixtoherbo-Fraxinetum/Quercetum/Populetum/Betuletum/Alnetum forest types (F4), Cirsium oleraceum, Carex remota, Carex pallescens, Geum urbanum as well as Carex vaginata and Carex panicea are characteristic for the herb layer. Frangula alnus, Corylus avellana, Sorbus aucuparia, Padus avium prevail in the shrub layer.
Lithuania’s swamp and fen forests, i.e., black alder carrs as well as downy birch fen and swamp forests, are grouped together. All these forests have a single-staged tree layer, a poorly developed shrub layer and a luxuriant, usually closed floor vegetation. A characteristic feature of black alder swamp and fen forests is an uneven microrelief with hummocks around the bases of trees, among which seasonally flooded spaces stretch. This microrelief determines the existence of a distinctly mosaic pattern of vegetation with no mono dominating species in the herb and moss layers. In Urtico-Alnetum glutinosae/Fraxinetum/Betuletum forest types (T), the most constant species of the herb layer is Urtica dioica; further characteristic species include Chrysosplenium alternifolium, Filipendula ulmaria, Ranunculus repens, Galeobdolon luteum, Oxalis acetosella, Athyrium filix-femina and other. Padus avium, Ribes nigrum, Frangula alnus are present in the weakly developed shrub layer. In Filipendulo-mixtoherbo-Alnetum glutinosae/Fraxinetum/Betuletum forest types (T), the herb layer is abundant in species and mostly has a high coverage; Filipendula ulmaria dominates. Athyrium filix-femina, Calamagrostis canescens, Oxalis acetosella, Urtica dioica occur frequently; further typical species include Galium palustre, Impatiens noli-tangere, Ranunculus repens, Scutellaria galericulata, Caltha palustris, Lycopus europaeus and other. Frangula alnus, Sorbus aucuparia, Padus avium are characteristic for the usually weakly developed shrub layer. In Carico-Irido-Alnetum glutinosae/Betuletum pubescentis forest types (T), Carex acutiformis, Carex vesicaria, Iris pseudacorus, Thelypteris palustris, Peucedanum palustre, Naumburgia thyrsiflora, Solanum dulcamara and other hygrophytes prevail in the herb layer. Frangula alnus, Salix cinerea are present in the weakly developed shrub layer. Downy birch carrs and swamp forests naturally occupy a considerably smaller range with a much smaller expanse than do alder carrs. The moss layer of birch carrs and bog forests is highly characteristic with Sphagnum spp. In Carico-Betuletum pubescentis/Alnetum glutinosae forest types (T), Carex spp., Thelypteris palustris, are characteristic for the herb layer. Frangula alnus and Salix cinerea prevail in the shrub layer; Sorbus aucuparia is present. In Calamagrostido-Betuletum pubescentis/Alnetum glutinosae forest types (T), the most constant species of the herb layer are Calamagrostis canescens and Lysimachia vulgaris. Frangula alnus prevails in the sparsely developed shrub layer; Salix cinerea, Sorbus aucuparia are present as well.
Lithuania’s floodplain forests are species-rich often multi-layered communities characterised by different assemblages of deciduous broadleaved trees. In Fluviale-aegopodio-Quercetum/Fraxinetum/Ulmetum forest types (U1), Aegopodium podagraria dominates in the herb layer. Other characteristic herbaceous species include Pulmonaria obscura, Asarum europaeum, Galium rubioides, Hepatica nobilis, Lamiastrum galeobdolon, Stellaria holostea, Mercurialis perennis, Viola mirabilis, Equisetum arvense, Glechoma hederacea, Chaerophylum aromaticum, Urtica dioica etc. Mnium undulatum is characteristic for the sparsely developed moss layer. Corylus avellana prevails in the shrub layer, Padus avium is frequent as well. Fluviale-urtico-Alnetum glutinosae (U1) and Fluviale-hepatico-oxalido-Quercetum (U1) are two more types of vegetation belonging to floodplain forests, which require more detailed research.
2.3. Natural Forest Dynamics
Disturbances are key drivers of forest ecosystem dynamics. Disturbance regimes shape the physical template for future regeneration dynamics of the natural forest landscape. The development of natural forest communities have major adaptation traits and strategies to cope with the locally prevalent disturbance regime [5,11,29,30,31]. For example, the niche position of hornbeam is restricted to the gap dynamics caused by the death of individual trees or small groups of trees in mixed species forests (F3). In contrast, the niche position of Scots pine can be categorized as having successional development after repeated stand-replacing disturbances in mixed spruce forests (D8) and multi-cohort succession related to repeated partial disturbances in pine forests (D11, D12, S12). In general, Lithuania’s forests are subject to pulse disturbances at various spatial and temporal scales, and a stand that is subject to a larger-scale disturbance may also be subject to smaller-scale disturbances [5,32]. For example, in a mixed spruce forest (D8), disturbance can range from a light gap or small patch to a stand or large patch-sized disturbance. Moreover, the mode of vegetation succession and dominance is strongly dependent on the composition of the stand and site conditions [27]. For example, if a stand is a mixture of shade-intolerant (e.g., silver birch) and shade-tolerant (e.g., small-leaved lime) species, and occurs on a rich mesic site, any canopy gaps that occur through deaths of single trees are likely to be ‘captured’ by shade-tolerant species [32]. This is because shade-tolerant species are likely to be better represented in the reproduction layer and their growth rates are optimal on such sites [10]. On the other hand, if a similar stand develops on a drier, less fertile site, the less shade-tolerant species have a greater chance to establish themselves because on these sites their survival rates exceed those of the more moisture- and nutrient-demanding shade-tolerant species [33].
Therefore, we attempt to characterize Lithuanian forests using (1) forest site types, (2) potential natural vegetation, and (3) forest dynamics types (Table 1).
3. Successional Characteristics of Hemi-Boreal Forest Communities
It is generally accepted that many primary forest species repopulate reforested areas as soon as vegetation development after disturbance follows a deterministic path imposed on this process by higher order ecological constraints, the long-term evolution of the Earth's climate [35,36]. However, the recovery process, which follows a disturbance in an area where the primary communities of forest organisms existed begins if biological remnants (e.g., buried seeds) survive. The greater the soil deterioration and changes in micro climate during the bare or cultivated period, between primary forest destruction and the onset of succession, the more subsequent succession will resemble a primary sere [23]. In other words, forest disturbances trigger successional events that lead to functionally determined potential vegetation communities [8].Therefore, the phenomenon of vegetation climax was considered in this review as the smallest invariant set of successional events, the occurrence of which cannot be reduced to the properties of individual ecosystem components [35,37]. End communities, or so-called climax communities, sometimes referred to as the 'potential vegetation' of a site, and shaped primarily by the local climate, were regarded as a position of relative stability in vegetation succession [10,23,38,39] (Table 2).
Ecological succession is the regular change in the biotope of a natural ecosystem, where some species prevail, while others are displaced, that is an important condition for natural biodiversity to flourish. In all forest communities, the tendency of succession is towards the restoration of the climatically determined end communities, although windthrows, clearcut harvesting and soil deterioration due to overcultivation may deflect the succession and even prevent the development of climax vegetation [8,23]. Deflecting factors alter the course of succession by giving an advantage to certain species over others, as the genetic component of natural biodiversity is the most sensitive of all components to destruction because of reductions in effective population size and interruptions in gene flow [23,40]. For example, Norway spruce is fire-intolerant and thus is often eliminated together with its seed bank; in contrast, Scots pine is fire-tolerant, and fire creates multi-cohort pine stands [17]. Scots pine forest that appear to be dependent on recurrent fires is a biotic climax determined by fire (fire climax). By the way, in modern forestry, clearcutting and deep mechanical scarification is a substitute for low-intensity fire [17], which is primarily a human-caused factor [41].
In landscapes with little human impact, the dynamics of life cycles in different forests is unsynchronized, resulting in a mosaic of successional events and habitat patches [5]. The gap phase is of crucial importance in determining the floristic composition of the entire forest cycle in this mosaic [42]. Nevertheless, when speaking of forest dynamics, we usually envision some model of tree species turnover and replacement, where the mode of replacement is strongly dependent on the plant–environment adaptation to competition, stress, and disturbance. Moreover, there is growing evidence that “the genotypes of the majority of plants appear to represent compromises between the conflicting selection pressures resulting from particular combinations of competition, stress, and disturbance” [43,44,45]. A group of ruderal species is best adapted to low stress and highly disturbed sites, a group of stress-tolerant species—to high stress and low levels of disturbance, and a group of competitive species—to low levels of both stress and disturbance [46] (Table 2). Stress tolerance as a distinct strategy evolved in inherently unproductive habitats or in site conditions of extreme resource depletion induced by the plants themselves [43]. Each plant species has its own stress tolerance limits at different stages of ontogenesis [47].
4. Adaptive Strategies of Forest Tree Species
The variety of tree life cycle events and adaptations evolves around specific features which are caused by the species-specific rate of response of the developing organism [47,50,51]. Each tree species has its own pattern and timing for these events and adaptations, often known as its ontogeny [52]. The development of trees from one ontogenetic stage to another occurs as branches of new orders appear in their root and shoot systems. Differences in tree ontogeny lead to different species’ responses to competition, stress, and disturbance. Therefore, the comparison of traits at the corresponding stages of ontogeny, from the establishment and growth of seedlings to the development and survival of mature trees, is a key aspect of studying the adaptive specialization of forest tree species.
Grime’s theoretical triangular model of adaptive strategies, which can be reconciled with the existing theories of natural selection, provides an insight into the processes of vegetation dynamics (succession and dominance) [43]. Grime and Pierce [53] state, “a universal three-way trade-off constrains adaptive strategies throughout the life of a tree, with extreme strategies facilitating the survival of genes via: (C) the survival of the individual using traits that maximize resource acquisition and resource control in consistently productive niches, (S) individual survival via maintenance of metabolic performance in variable and unproductive niches, or (R) rapid gene propagation via rapid completion of the life cycle and regeneration in niches where events are frequently lethal to the individual.” This means the trade-offs that occur in species responses to competition, stress, and disturbance in the forest are related to natural selection, which does not act directly on the traits, but on the general fitness of a collection of individuals who share certain heritable traits [54,55].
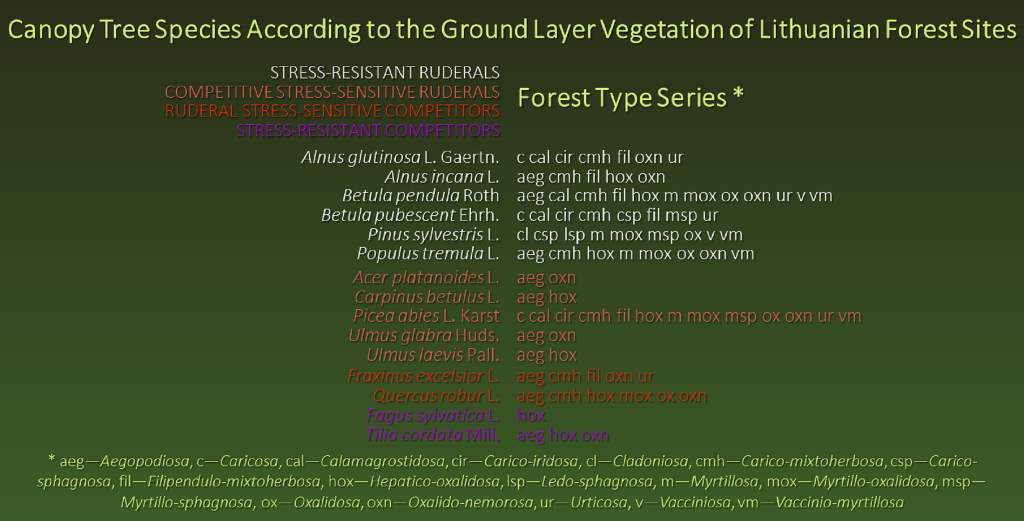
So, Grime’s and Pierce’s secondary CSR strategies, which describe various equilibria between competitiveness (C), stress tolerance (S), and ruderalism (R), can be considered to reflect four establishment and development adaptive specialization characteristics of forest tree species; i) stress-resistant ruderals, ii) competitive stress-sensitive ruderals, iii) ruderal stress-sensitive competitors, and iv) stress-resistant competitors represent the four types of trees in the forest.
Stress-resistant ruderals emerge as gap makers and grow only in forest sites that have been completely disturbed and damaged [56]. Their juveniles have the highest growth potential and colonize into large gaps (frequently with exposed mineral soils) after their formation and grow only in them as dominants [32,57]. Eurasian aspen, silver birch and Scots pine are characterized by a high light demand and low shade tolerance; the undergrowth of these species is absent under the dense canopy [58]. Gray alder is regarded as a more light-demanding species compared to black alder, which tends to be outcompeted by other species once the canopy closes [59]. Birches are seen as opportunists that take over abandoned or newly cleared areas [60]. Scots pine is usually replaced by Norway spruce on more nutrient-rich and less edaphically extreme sites where there is a lack of fire [27,61]. It is the least common admixture in stands of other tree species in Lithuania [20]. Eurasian aspen is usually found growing in small groups or stands in Norway spruce forest types [62], especially with species that allow sunlight through the canopy, such as Scots pine and birches [63].
Competitive stress-sensitive ruderals emerge as gap fillers [57] with their seeds germinating better in light gaps with medium canopy openness than in the understory or large gaps, and saplings can survive in closed forests [32,58]. In the stages from juveniles to adolescence, Norway maple, wych elm, and Norway spruce are characterized by a considerable shade tolerance and a high light demand. The undergrowth of the European hornbeam and Norway spruce is more common in forest sites with high light conditions [58]. Nonetheless, hornbeam and spruce are among the most shade-tolerant species. European hornbeam grows mostly in mixed stands below the canopy of other broadleaves, such as relatively high light-demanding English oak [58,64]. Norway spruce is the most common admixture in stands of other tree species in Lithuania [20].
Ruderal stress-sensitive competitors emerge as gap successors with advance regeneration [56,57]. Their already established juveniles survive in newly created light gaps [32]. European ash and English oak are very shade-tolerant in their juvenile stage, but at subsequent stages of ontogeny their need for light increases sharply; both species use full light, and their significant productivity is a result of high photosynthesis rates [58]. Mature ash stands are remarkable for the largest growing stock volume of wych elm in Lithuania [65].
Stress-resistant competitors regenerate before gap formation in the shade as gap advancers; juveniles have average growth rates [32,57]. Small-leaved lime is comparable to Norway spruce and European hornbeam in the juvenile stage in terms of the light minimum, but in the generative stage it has similar light demands as European aspen and European ash [58]. Exploitation of beech during the last two centuries is the main reason for the decline in the occurrence of lime in woodlands [66]. European beech is the most shade-tolerant broadleaved tree in its range and the strongest competitor among the trees in its range [67]. Its saplings often pre-exist in the understory before the canopy opening [68].
Based on these characteristics, we classified each of Lithuania’s main forest tree species into one of the four adaptive strategy types of tree establishment and development, i) stress-resistant ruderals, ii) competitive stress-sensitive ruderals, iii) ruderal stress-sensitive competitors, and iv) stress-resistant competitors (Table 3).
To understand the adaptive relationships in hemi-boreal forest tree communities we summarized chapters 2, 3 and 4 into Table 4. Adhering to previous research on European and Lithuanian forest types [27,28,34], we classified the tree species of each typical forest vegetation community according to four types of tree adaptive strategies. The classification was based on the characteristics of forest site types, potential natural vegetation, and forest dynamics.
The adaptive relationships in hemi-boreal forest trees (Table 4) are related to forest habitats and site conditions and indicate that forest management must consider the existence of the established equilibria between plant competitiveness, stress tolerance, and ruderalism. This is because of the disturbances in the established equilibria, which can have just as disastrous consequences, as is known from the history of agriculture, horticulture and silviculture [39]. Sudden outbreaks of diseases in clonally propagated trees such as poplars are proof of this; there are also examples from fruit trees and shrubs, former members of forest communities, that have recently been subjected to horticultural or agricultural cultivation. Due to modern forms of agricultural use, characterized by cloning of genetically homogeneous material and homogenization of the landscape, some of the ‘harmless’ species have even become serious pests/weeds.
Tree species’ life histories, generation times, reproductive behaviours, dispersal patterns and other emergent phenomena are linked in a vast and intricate network of adaptive relationships [70]. A sustainable forest ecosystem remains functional if there is functional redundancy, i.e., it contains functional groups, each of which has a number of substitutable insurance species [5,71]. This diversity of species leads to a variety of plant responses to competition, stress, and disturbance, which reduces the risk of complete loss of ecosystem functioning [29]. In this study, there are four types of such groups, i.e., stress-resistant ruderals, competitive stress-sensitive ruderals, ruderal stress-sensitive competitors, and stress-resistant competitors (Table 4). Based on this, we propose that reforestation move away from single species regeneration by implementing the maintenance of these four types of tree functional groups. Sustainable forest management based on the concept of vegetation climax is better at ensuring natural biodiversity survival and flourishing, because forests are the bedrock for a multitude of life forms and home to many communities, whose resilience and adaptive potential must replace productivity and stability, today's management goals [34]. In the context of global climate change and disturbances, the adaptive potential of forests lies in natural regeneration in the contiguous communities of plants. This requires the understanding and proposals to assisted natural regeneration strategies as an alternative adaptive model for forest management in Lithuania. The ultimate task is to create a scientific basis for maintaining or restoring the diversity of tree species adaptive relationships in forest ecosystems compared to that of monoculture forest stand in traditional high sustained wood yield forestry.
5. Concluding Remarks
Succession knowledge evolved with more in-depth assessments of the mechanisms of community assembly such as plant–environment adaptation, species performance strategies, and the niche complementarity hypothesis [46]. The recent idea of succession as a community assembly in progress has improved the applicability of this theory, which is one of the oldest ecological theories [50]. However, it is important not to overlook the foundational conceptual frameworks built on classic successional studies. Specifically, classic successional research has emphasized natural disturbance, community trajectories, and temporal dynamics, all of which are critical to understanding how communities assemble and disassemble in response to factors such as physical site conditions, initial stand composition and intermediate disturbance effects [77,78]. This study explored Grime’s plant adaptive strategies, each of which may be identified by reference to competitiveness, stress tolerance, and ruderalism, to determine the processes of vegetation succession and dominance in the light of natural selection and natural regeneration. It focused on differential species responses to competition, stress, and disturbance that alter species fitness through adaptive specialization. In general, adaptive strategies that explain the varying responses of tree species to competition, stress, and disturbance are a result of natural selection which is defined in terms of differential survival and reproduction [54,79,80,81,82]. Therefore, attention to the existence of the established equilibria between plant competitiveness, stress tolerance, and ruderalism can provide an insight into the adaptive processes of maintaining or restoring the diversity of plant communities and the productive capacity of ecological systems, the core elements of ecological sustainability. Participating in forest ecology means recognizing the inherent value of adaptive relationships in the forest and with the forest [32]. This could contribute to the continued survival and flourishing of natural biodiversity [83].
Author Contributions
Conceptualization, R.P. and M.M.; writing—original draft preparation, R.P. and M.M.; writing—review and editing, M.M. and R.P. All authors have read and agreed to the published version of the manuscript.
Funding
This review presents research findings that have been obtained through the long-term research program “Sustainable forestry and global changes”, implemented by the Lithuanian Research Centre for Agriculture and Forestry.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are grateful to Jean McCollister for proofreading the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kuuluvainen, T.; Angelstam, P.; Frelich, L.; Jõgiste, K.; Koivula, M.; Kubota, Y.; Lafleur, B.; Macdonald, E. Natural Disturbance-Based Forest Management: Moving Beyond Retention and Continuous-Cover Forestry. Frontiers in Forests and Global Change 2021, 4. [Google Scholar] [CrossRef]
- Spies, T.A.; Hessburg, P.F.; Skinner, C.N.; Puettmann, K.J.; Reilly, M.J.; Davis, R.J.; Kertis, J.A.; Long, J.W.; Shaw, D.C. Chapter 3: Old Growth, Disturbance, Forest Succession, and Management in the Area of the Northwest Forest Plan. In: Spies, T.A.; Stine, P.A.; Gravenmier, R.; Long, J.W.; Reilly, M.J., tech. coords. 2018. Synthesis of science to inform land management within the Northwest Forest Plan area. Gen. Tech. Rep. PNW-GTR-966. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 95-243. 2018, 966, 95–243.
- Turner, M.G. Landscape Ecology: The Effect of Pattern on Process. Annual Review of Ecology and Systematics 1989, 20, 171–197. [Google Scholar] [CrossRef]
- Angelstam, P.; Asplund, B.; Bastian, O.; Engelmark, O.; Fedoriak, M.; Grunewald, K.; Ibisch, P.L.; Lindvall, P.; Manton, M.; Nilsson, M.; et al. Tradition as Asset or Burden for Transitions from Forests as Cropping Systems to Multifunctional Forest Landscapes: Sweden as a Case Study. Forest Ecology and Management 2022, 505, 119895. [Google Scholar] [CrossRef]
- Bengtsson, J.; Angelstam, P.; Elmqvist, T.; Emanuelsson, U.; Folke, C.; Ihse, M.; Moberg, F.; Nyström, M. Reserves, Resilience and Dynamic Landscapes. ambi 2003, 32, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Millán, J.A.A.; Segarra, J.C.; Gómez, J.P.S.; Calzada, V.R.V.; Carmona, A.V. Problemas y perspectivas de la utilización de leñosas autóctonas en la restauración forestal. In Proceedings of the Restauración de ecosistemas mediterráneos, ISBN 84-8138-549-2; Universidad de Alcalá: Madrid, Spain, 2003; pp. 11–42. [Google Scholar]
- Filotas, E.; Parrott, L.; Burton, P.J.; Chazdon, R.L.; Coates, K.D.; Coll, L.; Haeussler, S.; Martin, K.; Nocentini, S.; Puettmann, K.J.; et al. Viewing Forests through the Lens of Complex Systems Science. Ecosphere 2014, 5, art1. [Google Scholar] [CrossRef]
- Petrokas, R.; Baliuckas, V.; Manton, M. Successional Categorization of European Hemi-Boreal Forest Tree Species. Plants 2020, 9, 1381. [Google Scholar] [CrossRef]
- Whisenant, S. Managing and Directing Natural Succession. In Forest Restoration in Landscapes: Beyond Planting Trees; Mansourian, S., Vallauri, D., Dudley, N., Eds.; Springer: New York, NY, 2005; pp. 257–261 . ISBN 978-0-387-29112-3. [Google Scholar]
- Kotar, J. Approaches to Ecologically Based Forest Management on Private Lands; USDA Forest Service, Minnesota Extension Service, University of Minnesota: St. Paul, Minnesota, USA, 1997. [Google Scholar]
- Kuuluvainen, T. Conceptual Models of Forest Dynamics in Environmental Education and Management: Keep It as Simple as Possible, but No Simpler. Forest Ecosystems 2016, 3, 18. [Google Scholar] [CrossRef]
- Wilson, D.S.; Sober, E. Reviving the Superorganism. Journal of Theoretical Biology 1989, 136, 337–356. [Google Scholar] [CrossRef]
- Dercole, F.; Ferriere, R.; Rinaldi, S. Chaotic Red Queen Coevolution in Three-Species Food Chains. Proc Biol Sci 2010, 277, 2321–2330. [Google Scholar] [CrossRef]
- Darwin, C. On the Origin of Species, 1859; 1st ed.; Routledge: London, 2004; ISBN 978-0-203-50910-4.
- Hall, B.K.; Strickberger, M.W.; Hallgrímsson, B. Strickberger’s Evolution; 4th ed.; Jones & Bartlett Learning: Sudbury, MA, USA, 2008; ISBN 978-0-7637-0066-9. [Google Scholar]
- Mozgeris, G.; Kazanavičiūtė, V.; Juknelienė, D. Does Aiming for Long-Term Non-Decreasing Flow of Timber Secure Carbon Accumulation: A Lithuanian Forestry Case. Sustainability 2021, 13, 2778. [Google Scholar] [CrossRef]
- Manton, M.; Ruffner, C.; Kibirkštis, G.; Brazaitis, G.; Marozas, V.; Pukienė, R.; Makrickiene, E.; Angelstam, P. Fire Occurrence in Hemi-Boreal Forests: Exploring Natural and Cultural Scots Pine Fire Regimes Using Dendrochronology in Lithuania. Land 2022, 11, 260. [Google Scholar] [CrossRef]
- Bukantis, A.; Ignatavičius, G.; Satkūnas, J.; Sinkevičius, S.; Šulijienė, G.; Vasarevičius, S.; Veteikis, D. Lithuania’s Environment: State, Processes and Trends; Kopa: Vilnius, Lietuva, 2013; ISBN 978-9955-772-58-3. [Google Scholar]
- Navasaitis, M.; Ozolinčius, R.; Smaliukas, D.; Balevičienė, J., M. Lietuvos dendroflora: monografija; Lutute: Kaunas, 2003; ISBN 978-9955-575-35-1. Kaunas.
- Kuliešis, A.; Kulbokas, G.; Kasperavičius, A.; Kazanavičiūtė, V.; Kvalkauskienė, M. Lithuanian National Forest Inventory, 1998–2017. From Measurements to Decision Making; Lututė: Kaunas, 2021; ISBN 978-9955-37-234-9. [Google Scholar]
- Bartašius, L. A Message from the Forest - “We Each Own the Forest”. Live Stories - Labanoras. Available online: https://www.youtube.com/watch?v=LH-3I9D3J6M (accessed on 26 March 2023).
- Hutchinson, G.E. The Ecological Theater and the Evolutionary Play; Yale University Press: New Haven, London, USA, 1965; ISBN 0-300-00586-5. [Google Scholar]
- Richards, P.W. The Tropical Rain Forest: An Ecological Study.; University Press: Cambridge [Eng.], 1952. [Google Scholar]
- Johnson, P.S.; Shifley, S.R.; Rogers, R.; Dey, D.C.; Kabrick, J.M. The Ecology and Silviculture of Oaks; 3rd ed.; CABI, 2019; ISBN 978-1-78064-708-1.
- National Environmental Protection Strategy. Ministry of Environment of the Republic of Lithuania. JSC ARX Reklama, Kaunas, Lithuania, 2016. Available online: https://www.fao.org/faolex/results/details/en/c/LEX-FAOC163665/ (accessed on 12 January 2023). :.
- Jõgiste, K.; Korjus, H.; Stanturf, J.A.; Frelich, L.E.; Baders, E.; Donis, J.; Jansons, A.; Kangur, A.; Köster, K.; Laarmann, D.; et al. Hemiboreal Forest: Natural Disturbances and the Importance of Ecosystem Legacies to Management. Ecosphere 2017, 8, e01706. [Google Scholar] [CrossRef]
- Bohn, U.; Gollub, G.; Hettwer, C.; Weber, H.; Neuhäuslová, Z.; Raus, T.; Schlüter, H. Karte Der Natürlichen Vegetation Europas / Map of the Natural Vegetation of Europe - Maßstab / Scale 1:2,500,000; Federal Agency for Nature Conservation: Bonn, Germany, 2000. [Google Scholar]
- Barbati, A.; Corona, P.; Marchetti, M. European Forest Types. Categories and Types for Sustainable Forest Management and Reporting; EEA Technical report; European Environment Agency: Copenhagen, Denmark, 2006; ISBN 92-9167-886-4. [Google Scholar]
- Seidl, R.; Rammer, W.; Spies, T.A. Disturbance Legacies Increase the Resilience of Forest Ecosystem Structure, Composition, and Functioning. Ecological Applications 2014, 24, 2063–2077. [Google Scholar] [CrossRef] [PubMed]
- Angelstam, P.; Kuuluvainen, T. Boreal Forest Disturbance Regimes, Successional Dynamics and Landscape Structures: A European Perspective. Ecological Bulletins 2004, 51. [Google Scholar]
- Petrokas, R.; Kavaliauskas, D. Concept for Genetic Monitoring of Hemiboreal Tree Dynamics in Lithuania. Land 2022, 11, 1249. [Google Scholar] [CrossRef]
- Petrokas, R.; Ibanga, D.-A.; Manton, M. Deep Ecology, Biodiversity and Assisted Natural Regeneration of European Hemiboreal Forests. Diversity 2022, 14, 892. [Google Scholar] [CrossRef]
- Walters, M.B.; Reich, P.B. Are Shade Tolerance, Survival, and Growth Linked? Low Light and Nitrogen Effects on Hardwood Seedlings. Ecology 1996, 77, 841–853. [Google Scholar] [CrossRef]
- Karazija, S. Forest types of Lithuania; Mokslas: Vilnius, 1988; ISBN 978-5-420-00421-0. [Google Scholar]
- Anand, M. Towards a Unifying Theory of Vegetation Dynamics, Faculty of Graduate Studies, University of Western Ontario: London, ON, Canada, 1997.
- Lamb, D. Restoring Tropical Moist Broad-Leaf Forests. In Forest Restoration in Landscapes: Beyond Planting Trees; Mansourian, S., Vallauri, D., Dudley, N., Eds.; Springer: New York, NY, 2005; pp. 291–297. ISBN 978-0-387-29112-3. [Google Scholar]
- Petrokas, R. Forest Climax Phenomenon: An Invariance of Scale. Forests 2020, 11, 56. [Google Scholar] [CrossRef]
- Whittaker, R.H. A Consideration of Climax Theory: The Climax as a Population and Pattern. Ecological Monographs 1953, 23, 41–78. [Google Scholar] [CrossRef]
- Stern, K.; Roche, L. Genetics of Forest Ecosystems; Chapman and Hall, 1974; ISBN 978-0-387-06095-8.
- Putz, F.E.; Redford, K.H.; Robinson, J.G.; Fimbel, R.; Blate, G.M. Biodiversity Conservation in the Context of Tropical Forest Management; World Bank: Washington, DC, 2000. [Google Scholar]
- Stewart, O.C. Fire as the First Great Force Employed by Man. In Man’s Role in Changing the Face of the Earth; Thomas, William L., Ed.; University of Chicago Press: Chicago, IL, USA, 1956; Vol. 2, pp. 115–133. [Google Scholar]
- Whitmore, T.C. Changes Over Twenty-One Years in the Kolombangara Rain Forests. Journal of Ecology 1989, 77, 469–483. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. The American Naturalist 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Chai, Y.; Yue, M.; Wang, M.; Xu, J.; Liu, X.; Zhang, R.; Wan, P. Plant Functional Traits Suggest a Change in Novel Ecological Strategies for Dominant Species in the Stages of Forest Succession. Oecologia 2016, 180, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Huang, J.; Xu, Y.; Ding, Y.; Zang, R. Plant Functional Niches in Forests Across Four Climatic Zones: Exploring the Periodic Table of Niches Based on Plant Functional Traits. Frontiers in Plant Science 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Anyomi, K.A.; Neary, B.; Chen, J.; Mayor, S.J. A Critical Review of Successional Dynamics in Boreal Forests of North America. Environ. Rev. 2022, 30, 563–594. [Google Scholar] [CrossRef]
- Evstigneev, O.I. Features of Undergrowth Development in Eastern European Forests. Russian Journal of Ecosystem Ecology 2019, 4, 1–23. [Google Scholar] [CrossRef]
- Král, K.; Shue, J.; Vrška, T.; Gonzalez-Akre, E.B.; Parker, G.G.; McShea, W.J.; McMahon, S.M. Fine-Scale Patch Mosaic of Developmental Stages in Northeast American Secondary Temperate Forests: The European Perspective. Eur J Forest Res 2016, 135, 981–996. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies, Vegetation Processes, and Ecosystem Properties; 2nd ed.; John Wiley & Sons, 2006; ISBN 978-0-470-85040-4.
- Cequinel, A.; Capellesso, E.S.; Marcilio-Silva, V.; Cardoso, F.C.G.; Marques, M.C.M. Determinism in Tree Turnover during the Succession of a Tropical Forest. Perspectives in Plant Ecology, Evolution and Systematics 2018, 34, 120–128. [Google Scholar] [CrossRef]
- Petrokas, R. Appropriate Measures for Retention Forestry. Baltic Forestry 2016, 22, 382–389. [Google Scholar]
- Holmberg, T.J. 4.2.3: Life Histories and Natural Selection. Available online: https://bio.libretexts.org/Sandboxes/tholmberg_at_nwcc.edu/BIOL_1213/04%3A_Unit_4%3A_-_Ecology/4.02%3A_Population_Ecology/4.2.03%3A_Life_Histories_and_Natural_Selection (accessed on 2 May 2023).
- Grime, J.P.; Pierce, S. The Evolutionary Strategies That Shape Ecosystems; John Wiley & Sons: Chichester, West Sussex, England, 2012; ISBN 978-0-470-67481-9. [Google Scholar]
- Lefèvre, F.; Boivin, T.; Bontemps, A.; Courbet, F.; Davi, H.; Durand-Gillmann, M.; Fady, B.; Gauzere, J.; Gidoin, C.; Karam, M.-J.; et al. Considering Evolutionary Processes in Adaptive Forestry. Annals of Forest Science 2014, 71, 723–739. [Google Scholar] [CrossRef]
- Metz, J.A.J.; Nisbet, R.M.; Geritz, S.A.H. How Should We Define ‘Fitness’ for General Ecological Scenarios? Trends in Ecology & Evolution 1992, 7, 198–202. [Google Scholar] [CrossRef]
- Tinner, W.; Ammann, B. Long-Term Responses of Mountain Ecosystems to Environmental Changes: Resilience, Adjustment, and Vulnerability. In Global Change and Mountain Regions: An Overview of Current Knowledge; Huber, U.M., Bugmann, H.K.M., Reasoner, M.A., Eds.; Advances in Global Change Research; Springer Netherlands: Dordrecht, 2005; pp. 133–143; ISBN 978-1-4020-3508-1. [Google Scholar]
- Yamamoto, S. Gap Regeneration of Major Tree Species in Different Forest Types of Japan. Vegetatio 1996, 127, 203–213. [Google Scholar] [CrossRef]
- Evstigneev, O.I. Ontogenetic Scales of Relation of Trees to Light (on the Example of Eastern European Forests). Russian Journal of Ecosystem Ecology 2018, 3, 1–18. [Google Scholar] [CrossRef]
- McVean, D.N. Alnus Glutinosa (L.) Gaertn. Journal of Ecology 1953, 41, 447–466. [Google Scholar] [CrossRef]
- Dickson, J.H. Pleistocene History of Betula with Special Reference to the British Isles. Proceedings of the Royal Society of Edinburgh, Section B: Biological Sciences 1984, 85, 1–11. [Google Scholar] [CrossRef]
- Steijlen, I.; Zackrisson, O. Long-Term Regeneration Dynamics and Successional Trends in a Northern Swedish Coniferous Forest Stand. Can. J. Bot. 1987, 65, 839–848. [Google Scholar] [CrossRef]
- Rogers, P.C.; Pinno, B.D.; Šebesta, J.; Albrectsen, B.R.; Li, G.; Ivanova, N.; Kusbach, A.; Kuuluvainen, T.; Landhäusser, S.M.; Liu, H.; et al. A Global View of Aspen: Conservation Science for Widespread Keystone Systems. Global Ecology and Conservation 2020, 21, e00828. [Google Scholar] [CrossRef]
- Savill, P.S. The Silviculture of Trees Used in British Forestry, 3rd Edition; CABI, 2019; ISBN 978-1-78639-392-0.
- Kuehne, C.; Nosko, P.; Horwath, T.; Bauhus, J. A Comparative Study of Physiological and Morphological Seedling Traits Associated with Shade Tolerance in Introduced Red Oak (Quercus Rubra) and Native Hardwood Tree Species in Southwestern Germany. Tree Physiology 2014, 34, 184–193. [Google Scholar] [CrossRef]
- Kuliesis, A.; Kasperavicius, A.; Kulbokas, G.; Kvalkauskiene, M. Lithuanian National Forest Inventory, 2003-2007. Forest Resources and Their Dynamic; Lututė: Kaunas, 2009. [Google Scholar]
- Boratyńska, K.; Dolatowski, J. Systematics and geographical distribution. In Limes–Tilia cordata Mill., Tilia platyphyllos Scop.; Białobok, S., Ed.; Monografie popularnonaukowe; PAN, Instytut Dendrologii: Poznań, Poland, 1991. [Google Scholar]
- Walter, H. Vegetation of the Earth and Ecological Systems of the Geo-Biosphere; Heidelberg Science Library; 2nd ed.; Springer: New York, NY, USA, 2012; ISBN 978-1-4684-0468-5.
- Wagner, S.; Collet, C.; Madsen, P.; Nakashizuka, T.; Nyland, R.D.; Sagheb-Talebi, K. Beech Regeneration Research: From Ecological to Silvicultural Aspects. Forest Ecology and Management 2010, 259, 2172–2182. [Google Scholar] [CrossRef]
- Franklin, J. Regeneration and Growth of Pioneer and Shade-tolerant Rain Forest Trees in Tonga. New Zealand Journal of Botany 2003, 41, 669–684. [Google Scholar] [CrossRef]
- Holbrook, M. Adventures in Complexity: An Essay on Dynamic Open Complex Adaptive Systems, Butterfly Effects, Self-Organizing Order, Coevolution, the Ecological Perspective, Fitness Landscapes, Market Spaces, Emergent Beauty at the Edge of Chaos, and All That Jazz. Academy of Marketing Science Review 2003, 2003, 1–181. [Google Scholar]
- Hillebrand, H.; Kunze, C. Meta-Analysis on Pulse Disturbances Reveals Differences in Functional and Compositional Recovery across Ecosystems. Ecology Letters 2020, 23, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Myking, T.; Bøhler, F.; Austrheim, G.; Solberg, E.J. Life History Strategies of Aspen (Populus Tremula L.) and Browsing Effects: A Literature Review. Forestry: An International Journal of Forest Research 2011, 84, 61–71. [Google Scholar] [CrossRef]
- Aakala, T.; Kuuluvainen, T.; Wallenius, T.; Kauhanen, H. Tree Mortality Episodes in the Intact Picea Abies-Dominated Taiga in the Arkhangelsk Region of Northern European Russia. Journal of Vegetation Science 2011, 22, 322–333. [Google Scholar] [CrossRef]
- Stobbe, A.; Gumnior, M. Palaeoecology as a Tool for the Future Management of Forest Ecosystems in Hesse (Central Germany): Beech (Fagus Sylvatica L.) versus Lime (Tilia Cordata Mill.). Forests 2021, 12, 924. [Google Scholar] [CrossRef]
- Plesa, I.M.; González-Orenga, S.; Al Hassan, M.; Sestras, A.F.; Vicente, O.; Prohens, J.; Sestras, R.E.; Boscaiu, M. Effects of Drought and Salinity on European Larch (Larix Decidua Mill.) Seedlings. Forests 2018, 9, 320. [Google Scholar] [CrossRef]
- Gabrilavičius, R.; Petrokas, R.; Danusevičius, J. Rare tree species in Lithuanian forests; Baltic Printing House: Klaipėda, 2013; ISBN 978-609-460-072-2. [Google Scholar]
- Taylor, A.R.J. Concepts, Theories and Models of Succession in the Boreal Forest of Central Canada. Thesis, 2009.
- Chang, C.; HilleRisLambers, J. Integrating Succession and Community Assembly Perspectives. F1000Res 2016, 5, F1000. [Google Scholar] [CrossRef]
- Bader, M.K.-F.; Leuzinger, S. Hydraulic Coupling of a Leafless Kauri Tree Remnant to Conspecific Hosts. iScience 2019, 19, 1238–1247. [Google Scholar] [CrossRef]
- Simard, S.W.; Perry, D.A.; Jones, M.D.; Myrold, D.D.; Durall, D.M.; Molina, R. Net Transfer of Carbon between Ectomycorrhizal Tree Species in the Field. Nature 1997, 388, 579–582. [Google Scholar] [CrossRef]
- Klein, T.; Siegwolf, R.T.W.; Körner, C. Belowground Carbon Trade among Tall Trees in a Temperate Forest. Science 2016, 352, 342–344. [Google Scholar] [CrossRef]
- Rosenau, J. Natural Selection / Survival of the Fittest. Available online: https://ncse.ngo/natural-selection-survival-fittest (accessed on 6 April 2023).
- Ibanga, D.-A. Is Deep Ecology Inapplicable in African Context: A Conversation with Fainos Mangena. Filosofia Theoretica: Journal of African Philosophy, Culture and Religions 2017, 6, 101–119. [Google Scholar] [CrossRef]
Figure 1.
The potential natural vegetation of Lithuania: D8—hemi-boreal spruce forests with broadleaved trees; D11—boreal and hemi-boreal pine forests, partly with birch and spruce; D12—hemi-boreal pine forests, partly with birch; F3— species-rich oak-hornbeam forests; F4—lime-oak forests; U1—floodplain forests. Source: Bohn et al. [27] and Barbati et al. [28].
Figure 1.
The potential natural vegetation of Lithuania: D8—hemi-boreal spruce forests with broadleaved trees; D11—boreal and hemi-boreal pine forests, partly with birch and spruce; D12—hemi-boreal pine forests, partly with birch; F3— species-rich oak-hornbeam forests; F4—lime-oak forests; U1—floodplain forests. Source: Bohn et al. [27] and Barbati et al. [28].
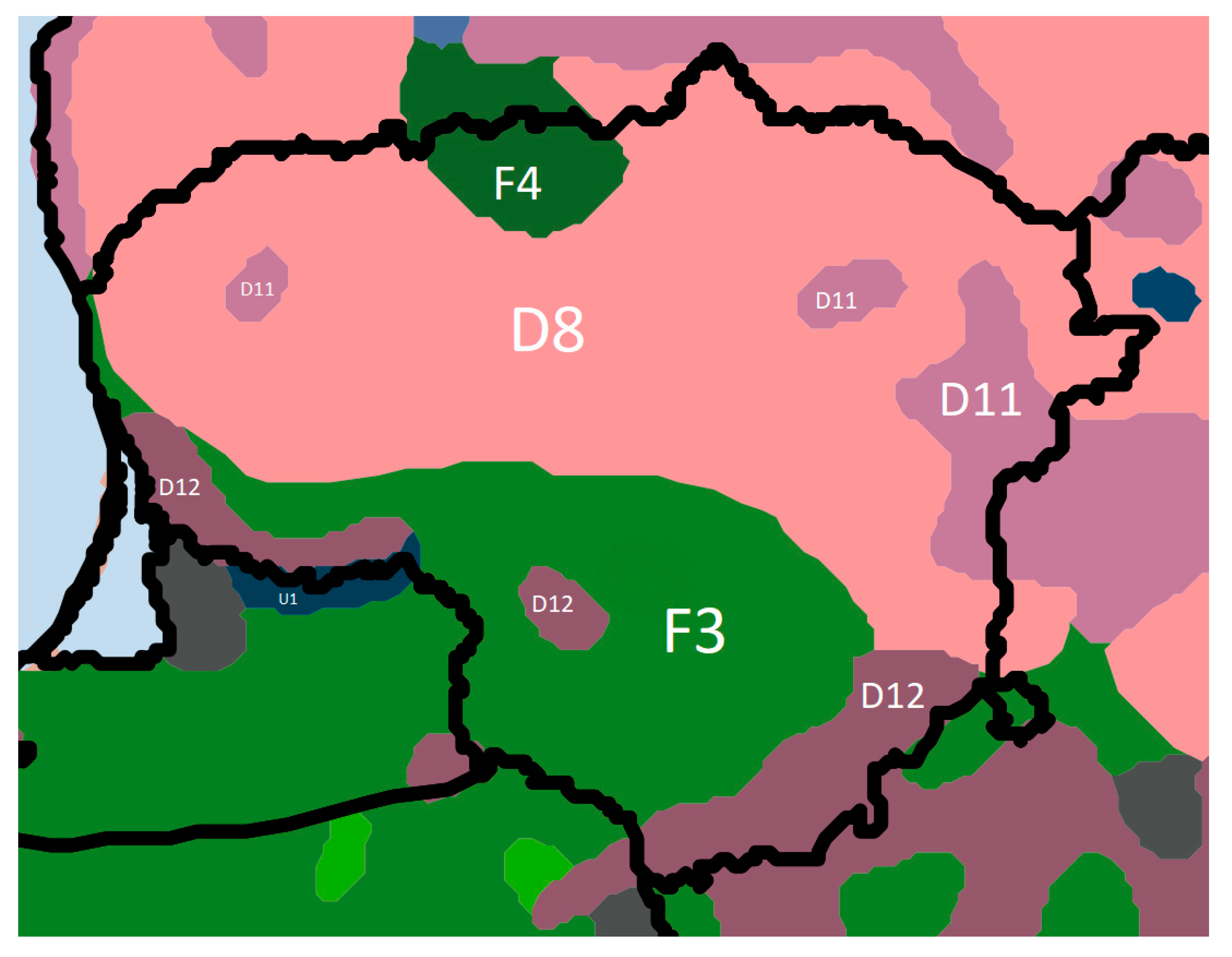

Table 1.
A summary of Lithuanian forest types based on forest site conditions, potential natural vegetation and forest dynamics [32,34].
| Forest Site Types | Dominant Ground Vegetation Types | Forest Stand Types | Forest Dynamics Types |
|---|---|---|---|
| Temporarily over moist eutrophic | Oxalido-nemorosa | Piceetum, Quercetum, Fraxinetum, Populetum, Betuletum pendulae, Alnetum | Successional development |
| Normally moist mesotrophic | Oxalidosa | Piceetum, Pinetum, Populetum, Betuletum pendulae, Quercetum | Successional development |
| Temporarily over moist mesotrophic | Myrtillo-oxalidosa | Piceetum, Betuletum pendulae, Populetum, Pinetum | Successional development |
| Temporarily over moist oligotrophic | Myrtillosa | Pinetum, Piceetum, Betuletum pendulae, Populetum | Multi-cohort succession |
| Normally moist oligotrophic | Vaccinio-myrtillosa | Pinetum, Betuletum pendulae, Populetum, Piceetum | Multi-cohort succession |
| Normally moist (very) oligotrophic | Vacciniosa | Pinetum, Betuletum pendulae | Multi-cohort succession |
| Normally moist very oligotrophic (sands) | Cladoniosa | Pinetum | Multi-cohort succession |
| Over moist oligotrophic | Myrtillo-sphagnosa | Pinetum, Betuletum pubescentis, Piceetum | Multi-cohort succession |
| Peatland oligotrophic | Carico-sphagnosa | Pinetum, Betuletum pubescentis | Multi-cohort succession |
| Peatland very oligotrophic | Ledo-sphagnosa | Pinetum | Multi-cohort succession |
| Normally moist eutrophic | Hepatico-oxalidosa | Quercetum, Piceetum, Carpinetum, Fagetum, Populetum, Betuletum pendulae | Gap dynamics |
| Normally moist very eutrophic | Aegopodiosa | Quercetum, Fraxinetum, Tilietum, Ulmetum, Populetum, Betuletum | Gap dynamics |
| Temporarily over moist very eutrophic | Carico-mixtoherbosa | Fraxinetum, Quercetum, Populetum, Betuletum, Alnetum | Gap dynamics |
| Over moist very eutrophic | Urticosa | Alnetum glutinosae, Fraxinetum, Betuletum | Gap dynamics |
| Over moist eutrophic | Filipendulo-mixtoherbosa | Alnetum glutinosae, Fraxinetum, Betuletum | Gap dynamics |
| Peatland eutrophic | Carico-iridosa | Alnetum glutinosae, Betuletum pubescentis | Gap dynamics |
| Peatland mesotrophic | Caricosa | Betuletum pubescentis, Alnetum glutinosae | Gap dynamics |
| Over moist mesotrophic | Calamagrostidosa | Betuletum pubescentis, Alnetum glutinosae | Gap dynamics |
| End Communities | Forest Dynamics | Plant Functional Groups |
|---|---|---|
| Biotic climax | Multi-cohort succession | Ruderals |
| Edaphic climax | Successional development | Stress tolerators |
| Climatic climax | Gap dynamics | Competitors |
Table 3.
Adaptive strategies of forest tree species: the four types of tree establishment and development in the forest resemble Grime’s [43,53] plant adaptive strategies, which describe the various equilibria between competitiveness, stress tolerance, and ruderalism. Modified from Franklin [69].
| Development | Establishment | |
|---|---|---|
| Forest | Gaps | |
| Forest | Stress-resistant competitors:Tilia cordata, Fagus sylvatica *. | Competitive stress-sensitive ruderals:Acer platanoides, Carpinus betulus, Picea abies, Ulmus glabra, Ulmus laevis. |
| Gaps | Ruderal stress-sensitive competitors:Fraxinus excelsior, Quercus robur. | Stress-resistant ruderals:Alnus glutinosa, Alnus incana, Betula pendula, Betula pubescens, Pinus sylvestris, Populus tremula. |
* European beech may be expanding its range into the Baltics through the introduction of forestry.5. Adaptive Relationships in Hemi-Boreal Forest Trees: A Synthesis.
| Forest Site Type | Canopy Tree Species * | Potential End Community | |||
|---|---|---|---|---|---|
| Stress-Resistant Ruderals | Competitive Stress-Sensitive Ruderals | Ruderal Stress-Sensitive Competitors | Stress-Resistant Competitors | ||
| Hemi-boreal spruce forests with broadleaved trees (D8) | |||||
| Temporarily over moist eutrophic | Be Bu Pt Ai Ag | Pa Ug Ap | Fe Qr | Tc | Climatic climax |
| Normally moist mesotrophic | Ps Be Pt | Pa | Qr | - | Edaphic climax |
| Temporarily over moist mesotrophic | Be Bu Pt Ps | Pa | Qr | - | Edaphic climax |
| Boreal and hemi-boreal pine forests, partly with birch and spruce (D11) | |||||
| Temporarily over moist oligotrophic | Ps Be Pt | Pa | - | - | Edaphic climax |
| Normally moist oligotrophic | Ps Be Pt | Pa | - | - | Edaphic climax |
| Hemi-boreal pine forests, partly with birch (D12) | |||||
| Normally moist (very) oligotrophic | Ps Be | - | - | - | Fire climax |
| Normally moist very oligotrophic (sands) | Ps | - | - | - | Edaphic climax |
| Pine bog forests (S12) | |||||
| Over moist oligotrophic | Ps Bu | Pa | - | - | Edaphic climax |
| Peatland oligotrophic | Ps Bu | - | - | - | Edaphic climax |
| Peatland very oligotrophic | Ps | - | - | - | Fire climax |
| Oak-hornbeam forests (F3) | |||||
| Normally moist eutrophic | Be Pt Ai | Pa Cb Ug Ul Ap | Qr | Tc Fs | Climatic climax |
| Lime-oak forests (F4) | |||||
| Normally moist very eutrophic | Pt Ai Be Bu Ag | Ug Ul Ap | Qr Fe | Tc | Climatic climax |
| Temporarily over moist very eutrophic | Be Bu Pt Ai Ag | Pa Ug | Fe Qr | - | Climatic climax |
| Swamp and fen forests (T) | |||||
| Over moist very eutrophic | Ag Bu Be | Pa | Fe | - | Climatic climax |
| Over moist eutrophic | Ag Bu Be Ai | Pa | Fe | - | Climatic climax |
| Peatland eutrophic | Ag Bu | Pa | - | - | Edaphic climax |
| Peatland mesotrophic | Bu Ag | Pa | - | - | Edaphic climax |
| Over moist mesotrophic | Bu Ag Be | Pa | - | - | Edaphic climax |
* Ag – Alnus glutinosa L. Gaertn., Ai – Alnus incana L. Moench, Ap – Acer platanoides L., Be – Betula pendula Roth, Bu – Betula pubescent Ehrh., Cb – Carpinus betulus L., Fs – Fagus sylvatica L., Fe – Fraxinus excelsior L., Pa – Picea abies L. Karst, Ps – Pinus sylvestris L., Pt – Populus tremula L., Qr – Quercus robur L., Tc – Tilia cordata Mill., Ug – Ulmus glabra Huds., Ul – Ulmus laevis Pall.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



