Submitted:
19 May 2023
Posted:
19 May 2023
You are already at the latest version
Abstract
Insulin resistance, as a common pathological process of many metabolic diseases, including diabetes and obesity, has attracted much attention due to its relevant influencing factors. To date, studies have mainly focused on the mechanisms between mitochondrial stress and insulin resistance, and they are now pursued as a very attractive therapeutic target due to their extensive involvement in many human clinical settings. In view of the complex pathogenesis of diabetes, natural drugs have become new players in diabetes prevention and treatment because of their wide targets and few side effects. In particular, plant phenolics have been paid more attention because of their close relationship with oxidative stress. In this review, we briefly review the mechanisms by which mitochondrial stress leads to insulin resistance. Moreover, we list some cytokines and genes that have recently been found to play roles in mitochondrial stress and insulin resistance. Furthermore, we describe several natural drugs that are currently widely used and give a brief overview of their therapeutic mechanisms. Finally, we suggest possible ideas for future research related to the unique role that natural drugs play in the treatment of insulin resistance through the above targets.
Keywords:
mitochondrial stress
; insulin resistance
; natural drugs
1. Introduction
Diabetes mellitus is associated with metabolic disorders, with 25% of patients progressing to several microvascular complications and macrovascular complications, followed by blindness, renal failure, myocardial infarction or stroke within 20-40 years [1]. Diabetes mellitus can also progress to diabetic ketoacidosis in 10% of cases, which is the leading cause of diabetes mellitus-related death worldwide. As diabetes mellitus is a major healthcare burden, the mechanism of type 2 diabetes (T2DM) has been widely discussed for many years, and the most important mechanism is insulin resistance [2]. The term insulin resistance refers to the decreased efficiency of insulin in promoting glucose uptake and utilization in tissues involved in glucose homeostasis for various reasons, for which the body compensates by secreting too much insulin to produce hyperinsulinemia to maintain the stability of blood glucose. Decades of extensive research results and clinical trials have provided a detailed protocol for the treatment of these diseases, but the specific mechanism remains unclear, greatly limiting early intervention and the prevention and treatment of complications.
A growing recognition of the integrality of metabolic physiology has led to research into the mechanisms and related influencing factors of insulin resistance. As early as 1936, Himsworth clarified the concept of insulin resistance. Initially, it was assumed that reduced insulin receptor binding was responsible for the typical obesity-related insulin resistance, but this hypothesis was soon replaced by a model centered on insulin signal transduction defects. Countless hypotheses have been proven or disproved, but there is certainly a basic consensus on the consequences that insulin resistance can cause. Insulin levels increase in the livers of humans with insulin resistance, leading to a series of diseases. Most of these diseases are caused by a combination of insulin resistance in skeletal muscle, liver and fat. Insulin-induced glycogen synthesis in the liver and skeletal muscle is significantly impaired by insulin resistance [3], and adipose insulin resistance can lead to excess triglyceride levels in the liver of patients with nonalcoholic fatty liver disease by promoting the reesterification of circulating fatty acids.
The liver is the starting point of insulin resistance and plays a central regulatory role in glucose and lipid metabolism [4]. Moreover, the liver is rich in mitochondria, which, as the "energy factory" of cells, are the main site where reactive oxygen species (ROS) are produced. When excessive ROS production exceeds the body's ability to remove them [5], mitochondrial stress and energy metabolism disorders occur, and they play an important role in the early onset of a variety of metabolic diseases [6,7]. This makes it possible to further explore the specific mechanisms of hepatic insulin resistance.
Various research groups have developed drugs based on mechanisms that have been discovered in recent years. For instance, in 2005, Burkey et al. [8] realized that peptidyl peptidase-IV can improve insulin resistance and help in the treatment of diabetes. Regarding the crosstalk between insulin-responsive tissues, which is one of the complex mechanisms associated with insulin resistance in vivo, Klymenko et al. [9] have carried out relevant expositions and experiments over the years. Traditional thiazolidinediones (TZDs) or metformin, sodium-dependent glucose transporter 2 (SGLT-2) inhibitors, and other medications can have undesirable side effects [10,11,12], while natural drugs have shown unique advantages due to their wide range of targets, few side effects and mild efficacy. A large number of studies have shown that natural drugs such as cassia semen, rhubarb, aloe vera and senna can improve insulin resistance [13,14,15].
Overall, in our review, we will introduce the newly discovered cytokines and genes identified by various research groups that are closely related to mitochondrial stress and insulin resistance. This review will also provide a unique summary of some natural drugs for the treatment of insulin resistance-related targets to pave the way for some future research directions (Figure 1).
Natural drugs are an important means of prevention and treatment of insulin resistance-related diseases such as diabetes. IR is closely related to many pathological processes, such as abnormal insulin signaling pathway conduction, energy metabolism regulation, and inflammation, and emodin, aloe-emodin, chrysol and beranin, as the main active components of many natural drugs, play a role by targeting one or more targets in the above pathological process.
2. Mitochondrial Stress, a Key Inducer of Insulin Resistance
As mentioned earlier, the pathological process of insulin resistance in the liver is closely related to mitochondrial stress. Mitochondrial stress and insulin resistance form a vicious cycle, which jointly promotes the development of the disease. In recent years, people have gradually turned their attention toward the mechanism of mitochondrial stress, hoping to treat metabolic diseases with this as a target. In Figure 2, we visualize some of the factors that play an important role in mitochondrial stress and insulin resistance and point out some possible bridging factors between the two.
2.1. Major Factors Leading to Mitochondrial Stress
As mentioned above, a large number of research groups around the world have discussed the mechanisms involved in mitochondrial stress. ROS, the mediators of mitochondrial stress, have long been thought to play an important role in neurodegenerative diseases and cardiovascular diseases. As early as 1994, Paolisso et al. [16] found a relationship between mitochondrial stress and metabolic diseases by measuring O2 levels in the bodies of non-insulin-dependent diabetic patients. Over the next two decades, various research teams have proposed several related factors that affect mitochondrial stress. Among them, the most representative and most likely therapeutic targets include peroxisome proliferator-activated receptors (PPARs), cyclophilin D (CypD), and advanced glycation end products (AGEs), which we will discuss in the following sections.
PPARs, activated receptor ligands of the nuclear hormone receptor family that have been found to have three subtypes and to control many intracellular metabolic processes, are ligand-inducible nuclear receptors. By reducing inflammation and insulin resistance induced by interleukin-6 [17], Lee realized that PPAR-β/δ inhibits mitochondrial stress, reduces the production of ROS, and delays the disease progression of obesity, T2DM, NASH and other conditions [18]. In 2018, Chen et al. [19] noted that PPAR-β/δ of the PPAR family has remarkable effects on improving hepatic insulin sensitivity and reducing insulin resistance and blood sugar. Knockout or low expression of PPAR-γ significantly reduced macrophage activation, putting mice at increased risk of obesity and insulin resistance. In summary, PPARs play a role in fatty acid decomposition, cholesterol transport and energy metabolism, suggesting that PPARs can be an important treatment option for diseases such as NASH [20].
Some studies have mentioned that the key molecule CypD plays an important role in regulating mitochondrial stress. The mitochondrial permeability transition pore (MPTP) is a nonselective channel in the mitochondrial intima. Under physiological conditions, periodic opening and closing are essential for maintaining cell homeostasis. However, abnormal increase or modification (acetylation) of the protein expression of the key mitochondrial stress protein CypD leads to excessive irreversible opening of the MPTP [21], causing mitochondrial swelling and dysfunction of the mitochondrial respiratory chain (impaired activity of mitochondrial complex enzymes I-V) and leading to mitochondrial stress and energy metabolism disorders. In 2019, Castillo et al. [22] showed that after CypD caused MPTP opening, the presence of nonesterified fatty acids led to proton leakage from the islets. Changes in CypD expression or acetylation are considered to be key factors in the regulation of mitochondrial stress; protein acetylation modification, as one of the important aspects of epigenetics, has been a hot research topic in recent years. Recently, we found that in an insulin resistance mouse model induced by a high-fat diet, the luciferase activity and protein expression of CypD in the liver were increased compared with those in the control group. Moreover, CypD gene knockout reduced liver mitochondrial stress, thereby alleviating insulin resistance.
Two molecules that affect mitochondrial stress have thus far been described. Therefore, let us take a look at the compounds involved. AGEs are stable covalent compounds generated by the spontaneous reaction of macromolecular substances such as proteins, lipids or nucleic acids with glucose or other reduced monosaccharides without the involvement of enzymes. A number of research teams have found that AGEs accelerate aging in the human body; that is, they are closely linked to the development of a number of chronic degenerative diseases [23]. AGEs induce the expression of their own receptors and bind with their receptors to activate NADPH, thereby promoting the formation of oxidative stress (OS), which in turn induces the expression of AGEs, thus aggravating the occurrence of mitochondrial stress [23]. The above process mainly occurs through the mitochondrial respiratory chain and occasionally occurs through stress-related signaling pathways, such as Jun N-terminal kinase and p38. NF-κB activated by AGEs also increases the expression of certain cytokines, such as those that promote inflammation, and the increase in inflammation in the body leads to the exacerbation of disease [4]. AGEs have long been used as biomarkers or predictors of diabetes complications; however, in 2020, Shen et al. [24] showed that the promotion of ROS by AGEs implies that AGEs have a role in the treatment of diabetes, NASH and other diseases.
In addition to these three factors, other molecules and compounds that cause mitochondria to produce too many ROS are being discovered. Cytochrome P450 family 2 subfamily E member 1 (CYP2E1), located on chromosome 10, is mainly involved in the metabolism of low-molecular-weight substances. High insulin levels in the body and other pathological changes, such as steatosis, can promote the body’s ability to increase the production of ROS [25]. The oxidizing capacity of iron also contributes to the production of ROS [26,27]. In addition, some recent studies have shown that activation of NADPH oxidase 4 (NOX4) promotes ROS production while prolonging the endoplasmic reticulum stress mediated by OS, which leads to apoptosis of liver cells and accelerates the progression of NASH and other diseases. In addition, many experiments have shown that liver-specific peroxisome proliferator-activated receptor-γ coactivator-1 (PGC-1) deficiency expands the influence of mitochondrial stress and accelerates the development of NASH in mice, suggesting that PGC-1 inhibits oxidative stress and inflammation, leading to the improvement in disease [28]. In terms of compounds, in 2016, Chakraborti et al. [26] showed that a diet rich in fructose greatly speeds up the production of ROS.
These factors usually regulate insulin resistance by affecting mitochondrial stress, but some factors not only mediate or inhibit mitochondrial stress but also aggravate or improve insulin resistance themselves. For example, many scholars have proven that microRNAs not only inhibit mitochondrial stress to improve insulin resistance but also directly reduce the effect of insulin resistance [29]. MicroRNAs, or miRNAs for short, are a class of endogenous small noncoding RNAs with regulatory functions found in eukaryotes. Several articles have described the mechanisms of these small RNAs in various types of diabetes and other metabolic diseases [30,31,32]. MiR-155-5p protects beta cells from insulin resistance by promoting insulin secretion by the body's beta cells in the presence of glucose [33]. Moreover, miR-126 and miR-30c are two important benign factors in patients with diabetic cardiomyopathy. The former indirectly activates sirtuin-1 (SIRT1) and superoxide dismutase (SOD) to induce resistance to oxidative stress, while the latter targets PGC-1, not only inducing PPAR-α but also playing a positive role in key mitochondrial regulation. Together, the two can reduce excessive ROS and myocardial lipid accumulation in patients with diabetic cardiomyopathy, thereby preventing pathological processes such as cardiac insufficiency [34]. MiRNAs have been applied in various fields because of their newly discovered wide use. At present, most in vitro experiments on reducing the effects of insulin resistance and treating diabetes with miRNAs have been successful, but the ability to exert the therapeutic effects of miRNAs in vivo remains to be further studied.
2.2. How does Mitochondrial Stress Affect Insulin Resistance?
2.2.1. HSP60
Mitochondrial stress disrupts the function of pancreatic beta cells, leading to impaired glucose tolerance and insulin resistance, but the relationship between mitochondrial stress and insulin resistance is certainly not through one factor, as many factors act as links. Therefore, research on how mitochondrial stress leads to insulin resistance has become the focus of many research teams. We mentioned endoplasmic reticulum stress, which is closely related to mitochondrial stress, in the previous paragraph. Taking heat shock protein 60 (HSP60) as an example, HSP60 is a chaperone protein that is overexpressed when mitochondrial stress occurs. This state not only impairs insulin signaling and insulin sensitivity in mouse liver cells but also promotes endoplasmic reticulum stress, which also induces liver adipogenesis and insulin resistance [35]. In 2020, Taddeo et al. [36] presented experimental evidence for this claim. In their experiment, HSP60 knockout inhibited mammalian target of rapamycin complex 1 – sterol-regulatory-element-binding protein 1 (mTORC1-SREBP1) signaling and hepatic adipogenesis, suggesting that HSP60 may be involved in endoplasmic reticulum stress-induced mTORC1-SREBP1 signal transduction related to adipogenesis and steatosis. Therefore, the researchers concluded that mitochondrial stress and endoplasmic reticulum stress coregulate the insulin resistance process in the body.
However, HSP60 acts differently in different parts of the body. The high serum level of HSP60 is often accompanied by the downregulation of HSP60 in the myocardium, which means that the inhibition of HSP60 in the myocardium will lead to the intensification of insulin resistance [37]. As described previously, miRNA plays an important role in influencing insulin resistance. In 2020, Wen et al. [38] found that miR-802-5p causes cardiac insulin resistance by reducing HSP60 expression. HSP60 is also an important therapeutic target for insulin resistance in the heart that researchers have recently discovered.
2.2.2. FGF21
Does mitochondrial stress under pathological conditions necessarily result in insulin resistance? In 2013, Kim argued against this. Autophagy-related 7 (ATG7) is a gene that is critical for autophagosome activity; in the ATG7 knockout mouse, Kim et al. [39] were surprised to observe that the mice were not fat but lean and were unaffected by obesity and insulin resistance induced by a high-fat diet. Inhibition of autophagy leads to mitochondrial stress; however, this mitochondrial stress improves insulin resistance. Therefore, what is the bridge between the two? In a series of experiments, they identified a factor called fibroblast growth factor 21 (FGF21), which once served as a marker for a variety of mitochondrial diseases caused by mutations in mitochondrial DNA (mtDNA) [40,41]. Researchers have found that FGF21 can be induced by mitochondrial stress, which inhibits the expression of mitochondrial oxidative phosphorylation genes, resulting in damage to the mitochondrial respiratory chain and a reduction in ATP, thereby activating the integrated stress response (ISR) and mediating the production of FGF21 [41]. Similar to the HSP60 knockout results, FGF21 also improves hepatic insulin sensitivity by inhibiting mTORC1. Moreover, FGF21 activation increases fatty acid oxidation, lipolysis, and browning of white adipose tissue in mice, which protects mice from obesity and insulin resistance. The researchers also observed that metformin administration increases serum levels of FGF21, suggesting that FGF21 is induced by metformin, providing indirect evidence of FGF21's therapeutic value [42].
Currently, FGF21 and its analogs have been used in laboratory animals and human subjects to significantly improve lipid distribution in the body and reduce the body weight of the subjects, which is why it has become a promising new drug for the treatment of obesity, diabetes and metabolic syndrome. Unfortunately, in addition to the mitochondrial stress mentioned above, FGF21 can also be induced by a variety of stresses, such as obesity, exercise or cold exposure, and the PPAR family. The connection between these obvious nonmitochondrial inducers of FGF21 and mitochondrial stress is still unclear and needs to be uncovered.
2.2.3. GDF-15
Stress response protein growth differentiation factor-15 (GDF-15) is secreted during mitochondrial stress or functional damage [43,44,45], and high levels of GDF-15 are closely associated with a variety of pathological diseases, including inflammation, cancer, and various mitochondrial diseases [46], which is why GDF-15 has been extensively explored as a biomarker of mitochondrial disease [47] and is closely related to the mitochondrial function of patients [48,49]. The increase in GDF-15 in the body has been proven to be induced by dysfunction of the mitochondrial respiratory chain rather than other metabolic dysfunctions [50]. Furthermore, Straub [51]suggested a potential connection between mitochondrial respiratory dysfunction, NADH/NAD ratio, impaired ATP levels and GDF-15. This result implies that GDF-15 regulates its own expression in response to impaired mitochondrial function through the ISR pathway, just as the ISR pathway also induces GDF-15 hyperexpression during metformin treatment [52]. In conclusion, the above results indicate that mitochondrial stress caused by mitochondrial dysfunction induces an increase in GDF-15, which means that GDF-15 can not only be used as a marker of some metabolic diseases but can also play an important role in the diagnosis and treatment of diseases.
The relationship between GDF-15 and insulin resistance has also been explored by some research groups. Several studies in recent years have found that GDF-15 can be used as a therapeutic target for obesity and prediabetic glucose tolerance disorders. Studies have shown that GDF-15 is associated with obesity and insulin resistance, abnormal blood glucose levels, and impaired blood glucose homeostasis in the body. In 2017, Chung et al. [53] used recombinant GDF-15 in ob/ob mice and found that the mice lost weight and exhibited improved insulin sensitivity. In 2020, Choi [44] found that GDF-15 and FGF21 secretion was significantly upregulated in adipocyte-specific Crif1 (also known as Gadd45gip1) knockout mice on a high-fat diet, which was able to inhibit weight gain and improve glucose tolerance. Recently, the relationship and the specific mechanism between GDF-15 and insulin resistance have received increasing attention.
GDF-15 shows complex anti-inflammatory or proinflammatory effects in different stages of disease [54]; however, its specific mechanism in the early onset of metabolic diseases, especially hepatic insulin resistance, is still unclear. As we mentioned earlier, CypD plays an important role in the regulation of mitochondrial stress, and there is a large body of evidence to suggest this; at the same time, our group's previous study suggested that GDF-15 is downstream of CypD. Therefore, in this review, we propose a possible mechanism that describes the possible relationship between CypD and GDF-15. Increased expression or activity of CypD causes mitochondrial stress and energy metabolism disorders, which may affect insulin resistance by regulating the stress response protein GDF-15. The specific mechanism remains to be further studied (Figure 3).
3. Complex Mechanisms of Natural Drugs in the Treatment of Diseases Related to Insulin Resistance
Plant-derived secondary metabolites [55] are organic compounds produced by plants and are commonly known as a class of natural drugs. Plant-derived secondary metabolites are more readily available and safer than conventional chemical drugs [55] and have been shown to have clinically meaningful and mild efficacy in cancer treatment, inflammation reduction, and diabetes treatment. To date, researchers have identified more than 400,000 secondary metabolites in nature [56], including allicin, quercetin, eugenol, lycorine, tea polyphenols and berberine, and more new plant-derived active compounds are being discovered [11]. Among them, these active compounds can be widely classified into terpenoids and their derivatives, alkaloids, steroids, amino acids, polysaccharide antimicrobial peptides, lignans, saponins and new structures [56]. Recently, an increasing number of research teams have begun to pay attention to the effect of secondary metabolites on insulin resistance and related mechanisms in the pathogenesis of diabetes.
A large number of studies have confirmed the good hypoglycemic ability of a variety of plant derivatives. Moreover, a number of clinical cases have verified the therapeutic effect of resveratrol on diabetic patients [60]. Fasting blood glucose and serum insulin levels were significantly reduced during the treatment, and insulin sensitivity and glucose and lipid metabolism of the body were also restored [61]. Similar to the way that resveratrol exerts its therapeutic effect, curcumin, as a new drug for obesity and diabetes, can also alleviate the progression of insulin resistance through adenosine 5‘-monophosphate-activated protein kinase (AMPK), Nuclear factor erythroid2-related factor 2 (Nrf2) and PPAR related molecular pathways [62,63], which may involve endoplasmic reticulum stress and other oxidative stress abnormalities [64]. And it is closely related to the regulation of intestinal flora [65]. However, anthocyanins (anthocyanin 3-glucoside) and proanthocyanidins (PCs), which are widely present in black bean seeds, have been shown in animal experiments to activate AMPK and Glucose transporter type 4 (GLUT4) related pathways, alleviate hyperglycemia, and restore insulin sensitivity in mice [66]. In addition to the AMPK-related pathway, targeting other important factors in the regulation of insulin resistance can achieve similar therapeutic effects. The abnormal activation of Insulin receptor substrate (IRS) often leads to the impairment of insulin signaling pathway, and then mediates the occurrence of insulin resistance through inflammatory factors such as Tumor Necrosis Factor (TNF)-α [67]. Carnosic acid and Rosmarinic acid can reduce the occurrence of insulin resistance by regulating AMPK pathway and the activation of IRS [68-70]. In addition, Carica papaya extracts are equally rich in plant polyphenols. It has been confirmed that they reduce blood glucose and lipid levels, and the mechanism may be related to the improvement of endothelial NO synthase (eNOS), which is related to the accumulation of ROS and energy imbalance [71]. In summary, plant polyphenols are beneficial in the treatment of insulin resistance, and the mechanism is often closely related to the abnormal insulin signaling mediated by oxidative stress.
Anthraquinone natural products, including emodin, aloe-emodin, rhein, chrysophanol, aurantio-obtusin, and alaternin, are the main active ingredients of many natural drugs. Existing studies have found that they can alleviate insulin resistance from many aspects, such as regulating the insulin signaling pathway, regulating energy homeostasis, and improving inflammation. Emodin has been shown to increase the phosphorylation of AMPK protein and Acetyl-CoA carboxylase (ACC) protein in the liver, upregulate the expression of Carnitine palmitoyltransferase 1 (CPT1), and downregulate the expression of SREBP-1c and fatty acid synthase (FAS), thereby improving energy metabolism disorders, reducing fat accumulation, and promoting cellular glucose absorption. Emodin can reduce the levels of fasting blood glucose and fasting insulin in mice and improve insulin resistance [72,73]. Moreover, emodin can promote M2 polarization of macrophages by increasing Triggering receptor expressed on myeloid cells 2 (TREM2) expression, significantly reduce local and systemic inflammatory responses in obese mice, inhibit weight gain and lipid accumulation, and reduce fasting blood glucose and fasting insulin levels [74]. Aloe-emodin has also been found to be effective in reducing the production of the inflammatory factors TNF-α and interleukin (IL)-6 and in suppressing the NF-κB signaling pathway, thereby restoring insulin signaling, lowering fasting blood glucose, regulating islet β-cell function, and inhibiting fat accumulation to reduce obesity and insulin resistance. Aloe-emodin has been shown to alleviate inflammation [15,75,76,77]. Rhein can also improve insulin resistance by reducing adipose tissue inflammation and liver triglyceride accumulation in mice [78]. Rhein can also promote macrophage polarization toward the M2 phenotype [79], which can reduce inflammation and thus alleviate insulin resistance [80]. Cassia seed, a natural drug rich in anthraquinone products such as aurantio-obtusin and alaternin, can reduce the fasting blood glucose and fasting insulin contents in obese mice and restore insulin sensitivity by increasing skeletal muscle glucose uptake in obese mice [81]. The targets of aurantio-obtusin are more diverse. On the one hand, it phosphorylates AMPK, an important pathway for energy homeostasis. On the other hand, the expression of FAS is inhibited to reduce fat synthesis and thus improve insulin resistance [82]. In addition, aurantio-obtusin can activate the Phosphoinositide 3-kinase (PI3K)-Protein kinase B (AKT) signaling pathway in liver and adipose tissue, reduce fasting blood glucose, and improve glucose tolerance [83]. Alaternin also plays a role in regulating insulin signaling. Alaternin regulates downstream signals of insulin and restores insulin signaling by competitive inhibition of protein tyrosine phosphatase 1B (PTP1B) [84].
Among natural drugs, other natural products besides plant polyphenols and anthraquinones have shown promising results in the treatment of insulin resistance. Allicin, an organic sulfur compound extracted from the bulb of Allium garlic, has been shown to be effective in the treatment of various diseases due to its specific actions, including antibacterial, antioxidative stress, regulation of cardiovascular and cerebrovascular diseases, reduction in blood lipids, and regulation of diabetes. One study showed that allicin had a hypoglycemic effect in type 2 diabetes patients and animal models in which it could activate pancreatic insulin secretion and reduce Total Cholesterol (TC), Triglyceride (TG) and Fast Blood Glucose (FBG) [85]. Zhai et al. [86] showed that the hypoglycemic activity of allicin on diabetic rats was similar to that of glibenclamide and insulin. In addition, similar to the way in which allicin acts, allyl propyl disulfide, cysteine sulfoxide and S-allyl cysteine sulfoxide prevent liver-induced insulin activation, increase the insulin production capacity of islet β-cells, separate insulin from its bound form, and increase the insulin sensitivity of cells. The subsequent effect is to reduce blood glucose levels [87,88]. Therefore, the use of bioactive compounds derived from natural sources has become a new direction for the treatment of diabetes due to the moderate and obvious therapeutic effects of these metabolites, which is very consistent with the idea of treating chronic diseases.
4. Conclusions and Future Perspectives
In this article, we introduced the mitochondrial stress and insulin resistance mechanisms and the high-profile inducing or inhibiting factors, and importantly, we proposed in this review that CypD may lead to abnormal mitochondrial stress and affect insulin resistance through GDF-15, which represents a direction for future research (Table 1). Currently, an increasing number of drugs have been applied to the treatment of diseases related to glucose and lipid metabolism disorders. Throughout years of exploring the efficacy and safety of various diabetes drugs, natural drugs have shown unique advantages. We thus put forward a good prospect: In the future, natural drugs will become the new favored approach in the process of diabetes prevention and treatment due to their characteristics of high safety and broad action targets. Plant polyphenols, in particular, are closely related to oxidative stress and alleviate the progression of diabetes caused by insulin resistance through the regulation of insulin signaling pathway. We look forward to more research teams discovering additional natural drugs, and we hope to further explore the specific mechanisms by which natural drugs work, which represents a research direction with great potential.
Author Contributions
Writing-Original draft preparation, P.W.; Writing-Review & editing, P.W. & X.W.; Supervision, X.W.; Funding acquisition, X.W.
Funding
This work was supported by the China National Natural Science Foundation, grant number 81900736; Project funded by China Postdoctoral Science Foundation; Shandong Province "Double-Hundred Talent Plan" on 100 Foreign Experts and 100 Foreign Expert Teams Introduction, grant number WST2018004 and the Innovation Project of Shandong Academy of Medical Sciences, grant number 2019-11.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
I would like to thank Dr. Liu for his guidance.
Conflicts of Interest
The authors declare no conflict of interest. All coauthors have seen and agree with the contents of the manuscript, and there are no financial interests to report.
References
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sundaram, K.; Mu, J.; Dryden, G.W.; Sriwastva, M.K.; Lei, C.; Zhang, L.; Qiu, X.; Xu, F.; Yan, J.; et al. High-fat diet-induced upregulation of exosomal phosphatidylcholine contributes to insulin resistance. Nat. Commun. 2021, 12, 213. [Google Scholar] [CrossRef] [PubMed]
- Perseghin, G.; Price, T.B.; Petersen, K.F.; Roden, M.; Cline, G.W.; Gerow, K.; Rothman, D.L.; Shulman, G.I. Increased glucose transport–phosphorylation and muscle glycogen synthesis after exercise training in insulin-resistant subjects. N. Engl. J. Med. 1996, 335, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Nowotny, K.; Jung, T.; Höhn, A.; Weber, D.; Grune, T. Advanced glycation end products and oxidative stress in type 2 diabetes mellitus. Biomolecules 2015, 5, 194–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Du, H.; Shao, S.; Bo, T.; Yu, C.; Chen, W.; Zhao, L.; Li, Q.; Wang, L.; Liu, X.; et al. Cyclophilin D deficiency attenuates mitochondrial perturbation and ameliorates hepatic steatosis. Hepatology 2018, 68, 62–77. [Google Scholar] [CrossRef]
- Nassir, F.; Ibdah, J.A. Role of mitochondria in nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 8713–8742. [Google Scholar] [CrossRef]
- Galloway, C.A.; Lee, H.; Brookes, P.S.; Yoon, Y. Decreasing mitochondrial fission alleviates hepatic steatosis in a murine model of nonalcoholic fatty liver disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G632–G641. [Google Scholar] [CrossRef]
- Burkey, B.F.; Li, X.; Bolognese, L.; Balkan, B.; Mone, M.; Russell, M.; Hughes, T.E.; Wang, P.R. Acute and chronic effects of the incretin enhancer vildagliptin in insulin-resistant rats. J Pharmacol Exp Ther. 2005, 315, 688–695. [Google Scholar] [CrossRef]
- Klymenko, O.; Brecklinghaus, T.; Dille, M.; Springer, C.; de Wendt, C.; Altenhofen, D.; Binsch, C.; Knebel, B.; Scheller, J.; Hardt, C.; et al. Histone deacetylase 5 regulates interleukin 6 secretion and insulin action in skeletal muscle. Mol. Metab. 2020, 42, 101062. [Google Scholar] [CrossRef]
- Beck, G.R., Jr.; Khazai, N.B.; Bouloux, G.F.; Camalier, C.E.; Lin, Y.; Garneys, L.M.; Siqueira, J.; Peng, L.; Pasquel, F.; Umpierrez, D.; et al. The effects of thiazolidinediones on human bone marrow stromal cell differentiation in vitro and in thiazolidinedione-treated patients with type 2 diabetes. Transl. Res. 2013, 161, 145–155. [Google Scholar] [CrossRef]
- Bonnet, F.; Scheen, A. Understanding and overcoming metformin gastrointestinal intolerance. Diabetes Obes. Metab. 2017, 19, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.I.; Blau, J.E.; Rother, K.I.; Beitelshees, A.L. SGLT2 inhibitors as adjunctive therapy for type 1 diabetes: Balancing benefits and risks. Lancet Diabetes Endocrinol. 2019, 7, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.; Um, M.Y.; Choi, M.; Han, T.; Kim, I.H.; Shin, S. Cassia tora seed improves pancreatic mitochondrial function leading to recovery of glucose metabolism. Am. J. Chin. Med. 2020, 48, 615–629. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.R.; Cui, H.X.; Fang, J.L.; Yuan, K.; Guo, Y. Ameliorative effect and mechanism of the purified anthraquinone-glycoside preparation from Rheum palmatum L. on type 2 diabetes mellitus. Molecules 2019, 24, 1454. [Google Scholar] [CrossRef] [PubMed]
- Deora, N.; Sunitha, M.M.; Satyavani, M.; Harishankar, N.; Vijayalakshmi, M.A.; Venkataraman, K.; Venkateshan, V. Alleviation of diabetes mellitus through the restoration of β-cell function and lipid metabolism by Aloe vera (L.) Burm. f. extract in obesogenic WNIN/GR-Ob rats. J. Ethnopharmacol. 2021, 272, 113921. [Google Scholar] [CrossRef] [PubMed]
- PAOLISSO, G.D.A.A.; VOLPE, C.; et al. Evidence for a relationship between oxidative stress and insulin action in non-insulin-dependent (type II) diabetic patients. Metabolism 1994, 43, 1426–1429. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.K.S.; Peixoto, C.A. Role of peroxisome proliferator-activated receptors in non-alcoholic fatty liver disease inflammation. Cell. Mol. Life Sci. 2018, 75, 2951–2961. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Yeon, J.E.; Ko, E.J.; Yoon, E.L.; Suh, S.J.; Kang, K.; Kim, H.R.; Kang, S.H.; Yoo, Y.J.; Je, J.; et al. Peroxisome proliferator-activated receptor-delta agonist ameliorated inflammasome activation in nonalcoholic fatty liver disease. World J. Gastroenterol. 2015, 21, 12787–12799. [Google Scholar] [CrossRef]
- Chen, J.; Montagner, A.; Tan, N.; Wahli, W. Insights into the role of PPARβ/δ in NAFLD. Int. J. Mol. Sci. 2018, 19, 1893. [Google Scholar] [CrossRef]
- Liu, X.; Du, H.; Chai, Q.; jia, Q.; Liu, L.; Zhao, M.; Li, J.; Tang, H.; Chen, W.; Zhao, L.; et al. Blocking mitochondrial cyclophilin D ameliorates TSH-impaired defensive barrier of artery. Redox Biol. 2018, 15, 418–434. [Google Scholar] [CrossRef]
- Hou, D. , Hu, F., Mao, Y., Yan, L., Zhang, Y., Zheng, Z., Wu, A., Forouzanfar, T., Pathak, J. L., & Wu, G. Cationic antimicrobial peptide NRC-03 induces oral squamous cell carcinoma cell apoptosis via CypD-mPTP axis-mediated mitochondrial oxidative stress. Redox Biol. 2022, 54, 102355. [Google Scholar] [CrossRef] [PubMed]
- Castillo, E.C.; Morales, J.A.; Chapoy-Villanueva, H.; Silva-Platas, C.; Trevino-Saldana, N.; Guerrero-Beltran, C.E.; Bernal-Ramirez, J.; Torres-Quintanilla, A.; Garcia, N.; Youker, K.; et al. Mitochondrial hyperacetylation in the failing hearts of obese patients mediated partly by a reduction in SIRT3: The involvement of the mitochondrial permeability transition pore. Cell. Physiol. Biochem. 2019, 53, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’nikova, T.I.; Porozov, Y.B.; Terentiev, A.A. Oxidative stress and advanced lipoxidation and glycation end products (ALEs and AGEs) in aging and age-related diseases. Oxidative Med. Cell. Longev. 2019, 3085756. [Google Scholar] [CrossRef] [PubMed]
- Shen, C. Y. , Lu, C. H., Wu, C. H., Li, K. J., Kuo, Y. M., Hsieh, S. C., & Yu, C. L. The Development of Maillard Reaction, and Advanced Glycation End Product (AGE)-Receptor for AGE (RAGE) Signaling Inhibitors as Novel Therapeutic Strategies for Patients with AGE-Related Diseases. Molecules 2020, 25, 5591. [Google Scholar] [CrossRef] [PubMed]
- Selenscig, D.; Ferreira, M.D.R.; Chicco, A.; Lombardo, Y.B. Dietary fish oil ameliorates adipose tissue dysfunction in insulin-resistant rats fed a sucrose-rich diet improving oxidative stress, peroxisome proliferator-activated receptor γ and uncoupling protein 2. Food Funct. 2018, 9, 2496–2507. [Google Scholar] [CrossRef] [PubMed]
- Sil, R.; Chakraborti, A.S. Oxidative inactivation of liver mitochondria in high fructose diet-induced metabolic syndrome in rats: Effect of glycyrrhizin treatment. Phytother. Res. 2016, 30, 1503–1512. [Google Scholar] [CrossRef]
- Besse-Patin, A.; Léveillé, M.; Oropeza, D.; Nguyen, B.N.; Prat, A.; Estall, J.L. Estrogen signals through peroxisome proliferator-activated receptor−γ coactivator 1α to reduce oxidative damage associated with diet-induced fatty liver disease. Gastroenterology 2017, 152, 243–256. [Google Scholar] [CrossRef]
- Qadir, M.M.F.; Klein, D.; Álvarez-Cubela, S.; Domínguez-Bendala, J.; Pastori, R.L. The role of MicroRNAs in diabetes-related oxidative stress. Int. J. Mol. Sci. 2019, 20, 5423. [Google Scholar] [CrossRef]
- Berry, C.; Lal, M.; Binukumar, B.K. Crosstalk between the unfolded protein response, MicroRNAs, and insulin signaling pathways: In search of biomarkers for the diagnosis and treatment of type 2 diabetes. Front. Endocrinol. 2018, 9, 210. [Google Scholar] [CrossRef]
- Dotta, F.; Ventriglia, G.; Snowhite, I.V.; Pugliese, A. MicroRNAs: Markers of beta-cell stress and autoimmunity. Curr. Opin. Endocrinol. Diabetes Obes. 2018, 25, 237–245. [Google Scholar] [CrossRef]
- LaPierre, M.P.; Stoffel, M. MicroRNAs as stress regulators in pancreatic beta cells and diabetes. Mol. Metab. 2017, 6, 1010–1023. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wei, Y.; Geißler, C.; Abschlag, K.; Campos, J.C.; Hristov, M.; Möllmann, J.; Lehrke, M.; Karshovska, E.; Schober, A. Hyperlipidemia-induced MicroRNA-155-5p improves β-cell function by targeting Mafb. Diabetes 2017, 66, 3072–3084. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Zhao, Y.; He, M.; Li, H.; Fan, J.; Nie, X.; Yan, M.; Chen, C.; Wang, D.W. MiR-30c/PGC-1β protects against diabetic cardiomyopathy via PPARα. Cardiovasc. Diabetol. 2019, 18, 7. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.; Liang, X.; Liu, H.; Zhang, F.; Meng, W.; Hu, F. Mitochondrial stress protein HSP60 regulates ER stress-induced hepatic lipogenesis. J. Mol. Endocrinol. 2020, 64, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Juwono, J.; Martinus, R.D. Does Hsp60 provide a link between mitochondrial stress and inflammation in diabetes mellitus? J. Diabetes Res. 2016, 2016, 8017571. [Google Scholar] [CrossRef]
- Taddeo, E.P.; Alsabeeh, N.; Baghdasarian, S.; Wikstrom, J.D.; Ritou, E.; Sereda, S.; Erion, K.; Li, J.; Stiles, L.; Abdulla, M.; et al. Mitochondrial proton leak regulated by cyclophilin D elevates insulin secretion in islets at nonstimulatory glucose levels. Diabetes 2020, 69, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S. Effect of mitochondrial stress on systemic metabolism. Ann. N. Y. Acad. Sci. 2015, 1350, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Li, J.; Fu, Y.; Zheng, Y.; Ma, M.; Wang, C. Hypertrophic adipocyte–derived exosomal miR-802-5p contributes to insulin resistance in cardiac myocytes through targeting HSP60. Obesity 2020, 28, 1932–1940. [Google Scholar] [CrossRef]
- Kim, K. H.; Jeong, Y. T.; Oh, H.; Kim, S. H.; Cho, J. M.; Kim, Y. N.; Kim, S. S.; Kim, D. H.; Hur, K. Y.; Kim, H. K.; et al. Autophagy deficiency leads to protection from obesity and insulin resistance by inducing Fgf21 as a mitokine. Nature medicine 2013, 19, 83–92. [Google Scholar] [CrossRef]
- Forsström, S.; Jackson, C.B.; Carroll, C.J.; Kuronen, M.; Pirinen, E.; Pradhan, S.; Marmyleva, A.; Auranen, M.; Kleine, I.-M.; Khan, N.A.; et al. Fibroblast growth factor 21 drives dynamics of local and systemic stress responses in mitochondrial myopathy with mtDNA deletions. Cell Metabol. 2019, 30, 1040–1054.e1047. [Google Scholar] [CrossRef]
- Lehtonen, J.M.; Auranen, M.; Darin, N.; Sofou, K.; Bindoff, L.; Hikmat, O.; Uusimaa, J.; Vieira, P.; Tulinius, M.; Lönnqvist, T.; et al. Diagnostic value of serum biomarkers FGF21 and GDF15 compared to muscle sample in mitochondrial disease. J. Inherit. Metab. Dis. 2021, 44, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Joe, Y.; Ryter, S.W.; Surh, Y.J.; Chung, H.T. Similarities and distinctions in the effects of metformin and carbon monoxide in immunometabolism. Mol. Cells 2019, 42, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.S.; Goeminne, L.J.E.; Kim, J.T.; Tian, J.W.; Kim, S.H.; Nga, H.T.; Kang, S.G.; Kang, B.E.; Byun, J.S.; Lee, Y.S.; et al. Growth differentiation factor 15 protects against the aging-mediated systemic inflammatory response in humans and mice. Aging Cell 2020, 19, e13195. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.J.; Jung, S.B.; Lee, S.E.; Kang, S.G.; Lee, J.H.; Ryu, M.J.; Chung, H.K.; Chang, J.Y.; Kim, Y.K.; Hong, H.J.; et al. An adipocyte-specific defect in oxidative phosphorylation increases systemic energy expenditure and protects against diet-induced obesity in mouse models. Diabetologia 2020, 63, 837–852. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.S. Implications of mitochondrial unfolded protein response and mitokines: A perspective on fatty liver diseases. Endocrinol. Metab. 2019, 34, 39–46. [Google Scholar] [CrossRef]
- Hariyanto, T.I.; Kurniawan, A. Appetite problem in cancer patients: Pathophysiology, diagnosis, and treatment. Cancer Treat. Res. Commun. 2021, 27, 100336. [Google Scholar] [CrossRef]
- Rosenberg, B.J.; Hirano, M.; Quinzii, C.M.; Colantuoni, E.; Needham, D.M.; Lederer, D.J.; Baldwin, M.R. Growth differentiation factor-15 as a biomarker of strength and recovery in survivors of acute respiratory failure. Thorax 2019, 74, 1099–1101. [Google Scholar] [CrossRef]
- Dominguez-Gonzalez, C.; Badosa, C.; Madruga-Garrido, M.; Martí, I.; Paradas, C.; Ortez, C.; Diaz-Manera, J.; Berardo, A.; Alonso-Pérez, J.; Trifunov, S.; et al. Growth differentiation factor 15 is a potential biomarker of therapeutic response for TK2 deficient myopathy. Sci. Rep. 2020, 10, 10111. [Google Scholar] [CrossRef]
- Poulsen, N.S.; Madsen, K.L.; Hornsyld, T.M.; Eisum, A.-S.V.; Fornander, F.; Buch, A.E.; Stemmerik, M.G.; Ruiz-Ruiz, C.; Krag, T.O.; Vissing, J. Growth and differentiation factor 15 as a biomarker for mitochondrial myopathy. Mitochondrion 2020, 50, 35–41. [Google Scholar] [CrossRef]
- Montero, R.; Yubero, D.; Villarroya, J.; Henares, D.; Jou, C.; Rodríguez, M.A.; Ramos, F.; Nascimento, A.; Ortez, C.I.; Campistol, J.; et al. GDF-15 is elevated in children with mitochondrial diseases and is induced by mitochondrial dysfunction. PLoS One 2016, 11, e0148709. [Google Scholar] [CrossRef]
- Straub, I. R.; Weraarpachai, W.; Shoubridge, E. A. Multi-OMICS study of a CHCHD10 variant causing ALS demonstrates metabolic rewiring and activation of endoplasmic reticulum and mitochondrial unfolded protein responses. Hum Mol Genet 2021, 30, 687–705. [Google Scholar] [CrossRef] [PubMed]
- Steffen, J.; Ngo, J.; Wang, S. P.; Williams, K.; Kramer, H. F.; Ho, G.; Rodriguez, C.; Yekkala, K.; Amuzie, C.; Bialecki, R.; et al. The mitochondrial fission protein Drp1 in liver is required to mitigate NASH and prevents the activation of the mitochondrial ISR. Mol Metab 2022, 64, 101566. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.K.; Ryu, D.; Kim, K.S.; Chang, J.Y.; Kim, Y.K.; Yi, H.-S.; Kang, S.G.; Choi, M.J.; Lee, S.E.; Jung, S.-B.; et al. Growth differentiation factor 15 is a myomitokine governing systemic energy homeostasis. J. Cell Biol. 2017, 216, 149–165. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, H.; Zhong, Y. Hepatic GDF15 is regulated by CHOP of the unfolded protein response and alleviates NAFLD progression in obese mice. Biochem. Biophys. Res. Commun. 2018, 498, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Akazawa, S.; Sun, F.; Ito, M.; Kawasaki, E.; Eguchi, K. Efficacy of troglitazone on body fat distribution in type 2 diabetes. Diabetes Care 2000, 23, 1067–1071. [Google Scholar] [CrossRef] [PubMed]
- Ensuring drug safety: Lessons from the thiazolidinediones. Lancet 2007, 370, 1101. [CrossRef]
- Hou, CY.; Tain, YL.; Yu, HR.; Huang, LT. The Effects of Resveratrol in the Treatment of Metabolic Syndrome. Int J Mol Sci 2019, 20, 535. [Google Scholar] [CrossRef]
- Tamaki, N.; Cristina Orihuela-Campos, R.; Inagaki, Y.; Fukui, M.; Nagata, T.; Ito, H.-O. Resveratrol improves oxidative stress and prevents the progression of periodontitis via the activation of the Sirt1/AMPK and the Nrf2/antioxidant defense pathways in a rat periodontitis model. Free Radic. Biol. Med. 2014, 75, 222–229. [Google Scholar] [CrossRef]
- Canto, C.; Auwerx, J. Targeting sirtuin 1 to improve metabolism: All you need is NAD(+)? Pharm. Rev. 2012, 64, 166–187. [Google Scholar] [CrossRef]
- Barber, T.M.; Kabisch, S.; Randeva, H.S.; Pfeiffer, A.F.H.; Weickert, M.O. Implications of Resveratrol in Obesity and Insulin Resistance: A State-of-the-Art Review. Nutrients 2022, 14, 2870. [Google Scholar] [CrossRef]
- Zhu, X.; Wu, C.H.; Qiu, S.H.; Yuan, X.L.; Li, L. Effects of resveratrol on glucose control and insulin sensitivity in subjects with type 2 diabetes: Systematic review and meta-analysis. Nutr. Metab. 2017, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, Z.; Hekmatdoost, A.; Mirmiran, P. Anti-hyperglycemic and insulin sensitizer effects of turmeric and its principle constituent curcumin. Int J Endocrinol Metab 2014, 12, e18081. [Google Scholar] [CrossRef] [PubMed]
- Na, L. X.; Zhang, Y. L.; Li, Y.; Liu, L. Y.; Li, R.; Kong, T.; Sun, C. H. Curcumin improves insulin resistance in skeletal muscle of rats. Nutr Metab Cardiovasc Dis 2011, 21, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Pirola, L.; Balcerczyk, A.; Okabe, J.; El-Osta, A. Epigenetic phenomena linked to diabetic complications. Nat. Rev. Endocrinol. 2010, 6, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Xiao, Y.; Gao, J.; Zheng, Z.; Zhang, Z.; Yao, L.; Li, D. Curcumin improves insulin sensitivity in high-fat diet-fed mice through gut microbiota. Nutr Metab (Lond) 2022, 19, 76. [Google Scholar] [CrossRef]
- Kim, Y.; Keogh, JB.; Clifton, PM. Polyphenols and Glycemic Control. Nutrients 2016, 8, 17. [Google Scholar] [CrossRef]
- Thirone, A.C.; Huang, C.; Klip, A. Tissue-specific roles of IRS proteins in insulin signaling and glucose transport. Trends Endocrinol Metab 2006, 17, 72–78. [Google Scholar] [CrossRef]
- Jayanthy, G. ; Roshana, Devi. V.; Ilango, K.; Subramanian, S.P. Rosmarinic Acid Mediates Mitochondrial Biogenesis in Insulin Resistant Skeletal Muscle Through Activation of AMPK. J Cell Biochem 2017, 118, 1839–1848. [Google Scholar] [CrossRef]
- Den Hartogh, D.J.; Vlavcheski, F.; Tsiani, E. Muscle Cell Insulin Resistance Is Attenuated by Rosmarinic Acid: Elucidating the Mechanisms Involved. Int J Mol Sci 2023, 24, 5094. [Google Scholar] [CrossRef]
- Mimura, J.; Inose-Maruyama, A.; Taniuchi, S.; Kosaka, K.; Yoshida, H.; Yamazaki, H.; Kasai, S.; Harada, N.; Kaufman, R. J.; Oyadomari, S.; et al. Concomitant Nrf2- and ATF4-activation by Carnosic Acid Cooperatively Induces Expression of Cytoprotective Genes. Int J Mol Sci 2019, 20, 1706. [Google Scholar] [CrossRef]
- Santana, L.F.; Inada, A.C.; Espirito Santo, B.L.S.D.; Filiú, W.F.O.; Pott, A.; Alves, F.M.; Guimarães, R.C.A.; Freitas, K.C.; Hiane, P.A. Nutraceutical Potential of Carica papaya in Metabolic Syndrome. Nutrients 2019, 11, 1608. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, R.H.; Kamel, E.M.; Mahmoud, A.M.; El-Bassuony, A.A.; Bin-Jumah, M.; Lamsabhi, A.M.; Ahmed, S.A. Rumex dentatus L. phenolics ameliorate hyperglycemia by modulating hepatic key enzymes of carbohydrate metabolism, oxidative stress and PPARγ in diabetic rats. Food Chem. Toxicol. 2020, 138, 111202. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Kim, J.H.; Ghim, J.; Yoon, J.H.; Lee, A.; Kwon, Y.; Hyun, H.; Moon, H.Y.; Choi, H.S.; Berggren, P.O.; et al. Emodin regulates glucose utilization by activating AMP-activated protein kinase. J. Biol. Chem. 2013, 288, 5732–5742. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Yu, N.; Peng, J.; Zhao, Y.; Zhang, L.; Wang, X.; Xu, X.; Zhou, J.; Wang, F. Emodin inhibits lipid accumulation and inflammation in adipose tissue of high-fat diet-fed mice by inducing M2 polarization of adipose tissue macrophages. FASEB J. 2021, 35, e21730. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Dang, Y.; Xu, H.; Li, A.; Zhou, X.; Gao, X.; Li, Z. Aloe vera-fermented beverage ameliorates obesity and gut dysbiosis in high-fat-diet mice. Foods 2022, 11, 3728. [Google Scholar] [CrossRef] [PubMed]
- Dou, F.; Liu, Y.; Liu, L.; Wang, J.; Sun, T.; Mu, F.; Guo, Q.; Guo, C.; Jia, N.; Liu, W.; et al. Aloe-emodin ameliorates renal fibrosis via inhibiting PI3K/Akt/mTOR signaling pathway in vivo and in vitro. Rejuvenation Res. 2019, 22, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Quan, Y.; Gong, L.; He, J.; Zhou, Y.; Liu, M.; Cao, Z.; Li, Y.; Peng, C. Aloe emodin induces hepatotoxicity by activating NF-κB inflammatory pathway and P53 apoptosis pathway in zebrafish. Toxicol. Lett. 2019, 306, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Régnier, M.; Rastelli, M.; Morissette, A.; Suriano, F.; Le Roy, T.; Pilon, G.; Delzenne, N.M.; Marette, A.; Van Hul, M.; Cani, P.D. Rhubarb supplementation prevents diet-induced obesity and diabetes in association with increased Akkermansia muciniphila in mice. Nutrients 2020, 12, 2932. [Google Scholar] [CrossRef]
- Zhou, Y.; Gao, C.; Vong, C.T.; Tao, H.; Li, H.; Wang, S.; Wang, Y. Rhein regulates redox-mediated activation of NLRP3 inflammasomes in intestinal inflammation through macrophage-activated crosstalk. Br. J. Pharmacol. 2022, 179, 1978–1997. [Google Scholar] [CrossRef]
- Genua, M.; Rutella, S.; Correale, C.; Danese, S. The triggering receptor expressed on myeloid cells (TREM) in inflammatory bowel disease pathogenesis. J. Transl. Med. 2014, 12, 293. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Tong, A.H.; Pan, Y.Y.; Zhang, X.D.; Ding, W.Y.; Xiong, W. The effect of cassia seed extract on the regulation of the LKB1-AMPK-GLUT4 signaling pathway in the skeletal muscle of diabetic rats to improve the insulin sensitivity of the skeletal muscle. Diabetol. Metab. Syndr. 2019, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Ding, M.; Gu, Y.; Fan, G.; Liu, C.; Li, Y.; Sun, R.; Wu, J.; Li, J.; Xue, X.; et al. Aurantio-obtusin attenuates non-alcoholic fatty liver disease through AMPK-mediated autophagy and fatty acid oxidation pathways. Front. Pharmacol. 2021, 12, 826628. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.Y.; Liao, W.T.; Qiu, R.J.; Zhou, D.S.; Ni, W.J.; Yu, C.P.; Zeng, Y. Aurantio-obtusin improves obesity and insulin resistance induced by high-fat diet in obese mice. Phytother. Res. 2021, 35, 346–360. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Ali, M.Y.; Choi, J.S. Promising inhibitory effects of anthraquinones, naphthopyrone, and naphthalene glycosides, from Cassia obtusifolia on α-glucosidase and human protein tyrosine phosphatases 1B. Molecules 2016, 22, 28. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.K.; Prasad, S.K.; Kumar, R.; Hemalatha, S. An overview on antidiabetic medicinal plants having insulin mimetic property. Asian Pac. J. Trop. Biomed. 2012, 2, 320–330. [Google Scholar] [CrossRef]
- Zhai, B.; Zhang, C.; Sheng, Y.; Zhao, C.; He, X.; Xu, W.; Huang, K.; Luo, Y. Hypoglycemic and hypolipidemic effect of S-allyl-cysteine sulfoxide (alliin) in DIO mice. Sci. Rep. 2018, 8, 3527. [Google Scholar] [CrossRef]
- El-Saber Batiha, G.; Beshbishy, A.M.; Wasef, L.G.; Elewa, Y.H.A.; Al-Sagan, A.A.; Abd El-Hack, M.E.; Taha, A.E.; Abd-Elhakim, Y.M.; Devkota, H.P. Chemical constituents and pharmacological activities of garlic (Allium sativum L.): A review. Nutrients 2020, 12, 872. [Google Scholar] [CrossRef]
- Faroughi, F.; Charandabi, S.; Javadzadeh, Y.; Mirghafourvand, M. Effects of garlic pill on blood glucose level in borderline gestational diabetes mellitus: A randomized controlled trial. Iran. Red Crescent Med. J. 2018, 20, e60675. [Google Scholar] [CrossRef]
Figure 1.
Extensive use and specific mechanisms of natural drugs in insulin resistance-related diseases.
Figure 1.
Extensive use and specific mechanisms of natural drugs in insulin resistance-related diseases.
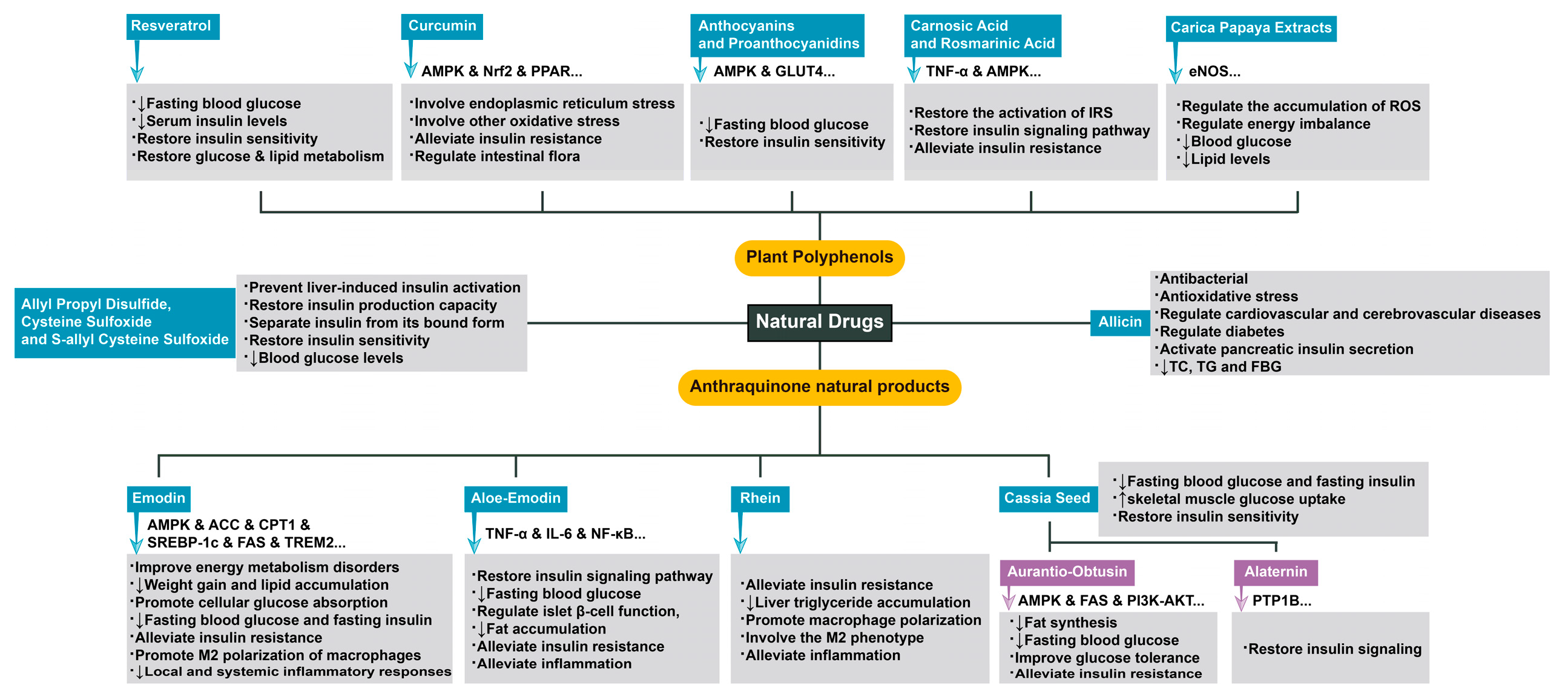
Figure 2.
The related factors mentioned in this review that affect mitochondrial stress and insulin resistance or connect the two. Some pathological factors lead to increased mitochondrial stress in the body. This process can be induced or aggravated by factors such as cyclophilin D (CypD) and advanced glycation end products (AGEs), but newly discovered factors such as peroxisome proliferator-activated receptors (PPARs) and proliferator-activated receptor-γ coactivator-1 (PGC-1) can improve this pathological process. Among miRNAs, miR-30c can inhibit mitochondrial stress and insulin resistance by targeting PGC-1, miR-126 can protect the body from the effects of reactive oxygen species (ROS) imbalance, and miR-155-5p can improve the adaptation of cells to insulin resistance. Mitochondrial stress leads to an increase in ROS, which aggravates insulin resistance in many ways, such as the overexpression of heat shock protein 60 (HSP60). However, mitochondrial stress does not only cause purely negative effects. Its activation of fibroblast growth factor 21 (FGF21) can inhibit the progression of insulin resistance to a certain extent, and we hypothesize that its activation of growth differentiation factor-15 (GDF-15) may do the same.
Figure 2.
The related factors mentioned in this review that affect mitochondrial stress and insulin resistance or connect the two. Some pathological factors lead to increased mitochondrial stress in the body. This process can be induced or aggravated by factors such as cyclophilin D (CypD) and advanced glycation end products (AGEs), but newly discovered factors such as peroxisome proliferator-activated receptors (PPARs) and proliferator-activated receptor-γ coactivator-1 (PGC-1) can improve this pathological process. Among miRNAs, miR-30c can inhibit mitochondrial stress and insulin resistance by targeting PGC-1, miR-126 can protect the body from the effects of reactive oxygen species (ROS) imbalance, and miR-155-5p can improve the adaptation of cells to insulin resistance. Mitochondrial stress leads to an increase in ROS, which aggravates insulin resistance in many ways, such as the overexpression of heat shock protein 60 (HSP60). However, mitochondrial stress does not only cause purely negative effects. Its activation of fibroblast growth factor 21 (FGF21) can inhibit the progression of insulin resistance to a certain extent, and we hypothesize that its activation of growth differentiation factor-15 (GDF-15) may do the same.
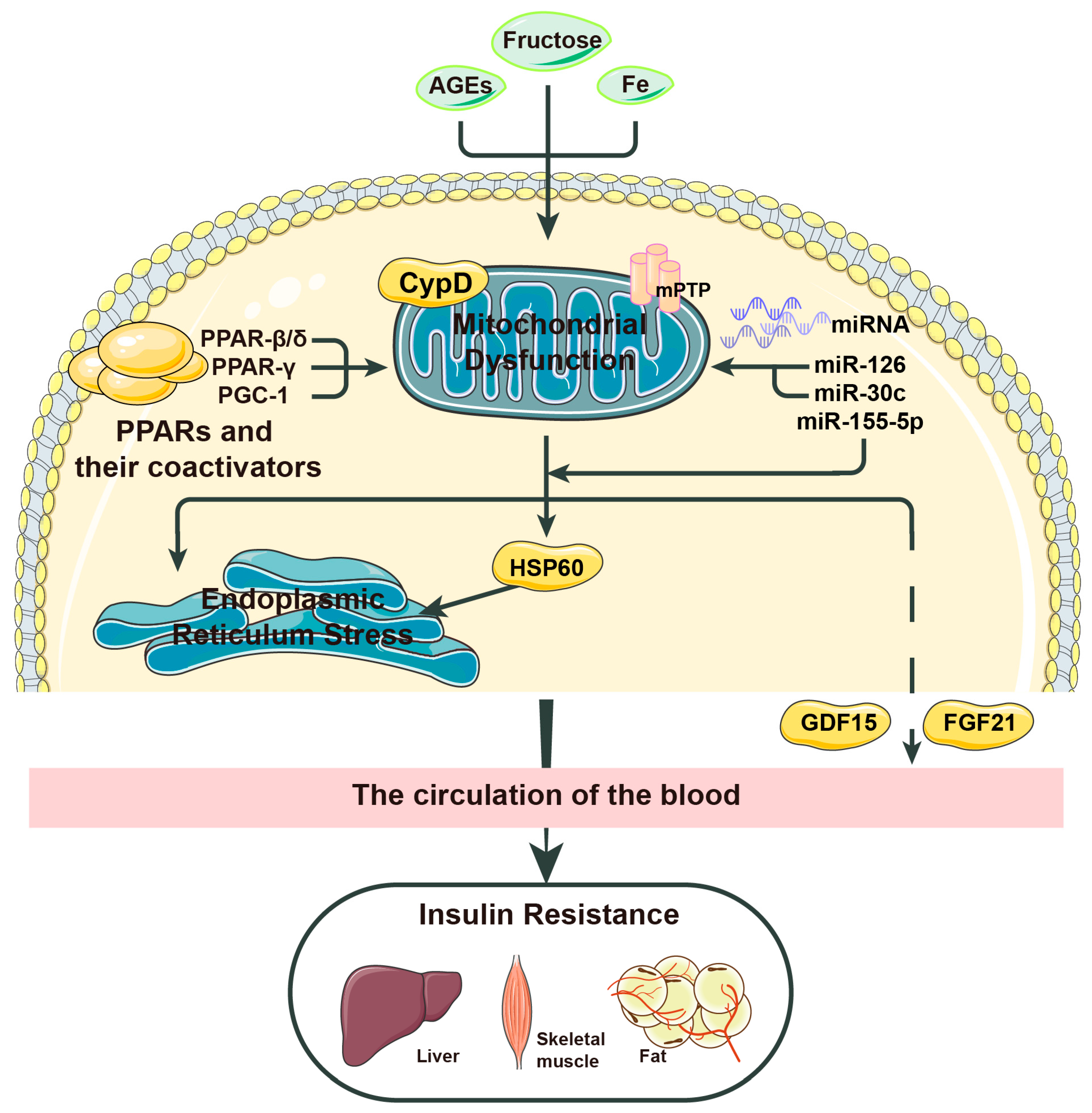
Figure 3.
The current main applications of GDF-15 and its role in mitochondrial stress and insulin resistance. GDF-15 is an important marker in the diagnosis of many diseases. In addition, it is a drug that can be used to treat the abnormal pathological state of mitochondrial stress, which is elevated to varying degrees in a variety of diseases, and its expression can effectively improve insulin resistance. For example, by activating ATF4, metformin effectively alleviates insulin resistance by inducing GDF-15. Notably, CypD may aggravate the progression of diseases related to insulin resistance by inhibiting the expression of GDF-15.
Figure 3.
The current main applications of GDF-15 and its role in mitochondrial stress and insulin resistance. GDF-15 is an important marker in the diagnosis of many diseases. In addition, it is a drug that can be used to treat the abnormal pathological state of mitochondrial stress, which is elevated to varying degrees in a variety of diseases, and its expression can effectively improve insulin resistance. For example, by activating ATF4, metformin effectively alleviates insulin resistance by inducing GDF-15. Notably, CypD may aggravate the progression of diseases related to insulin resistance by inhibiting the expression of GDF-15.
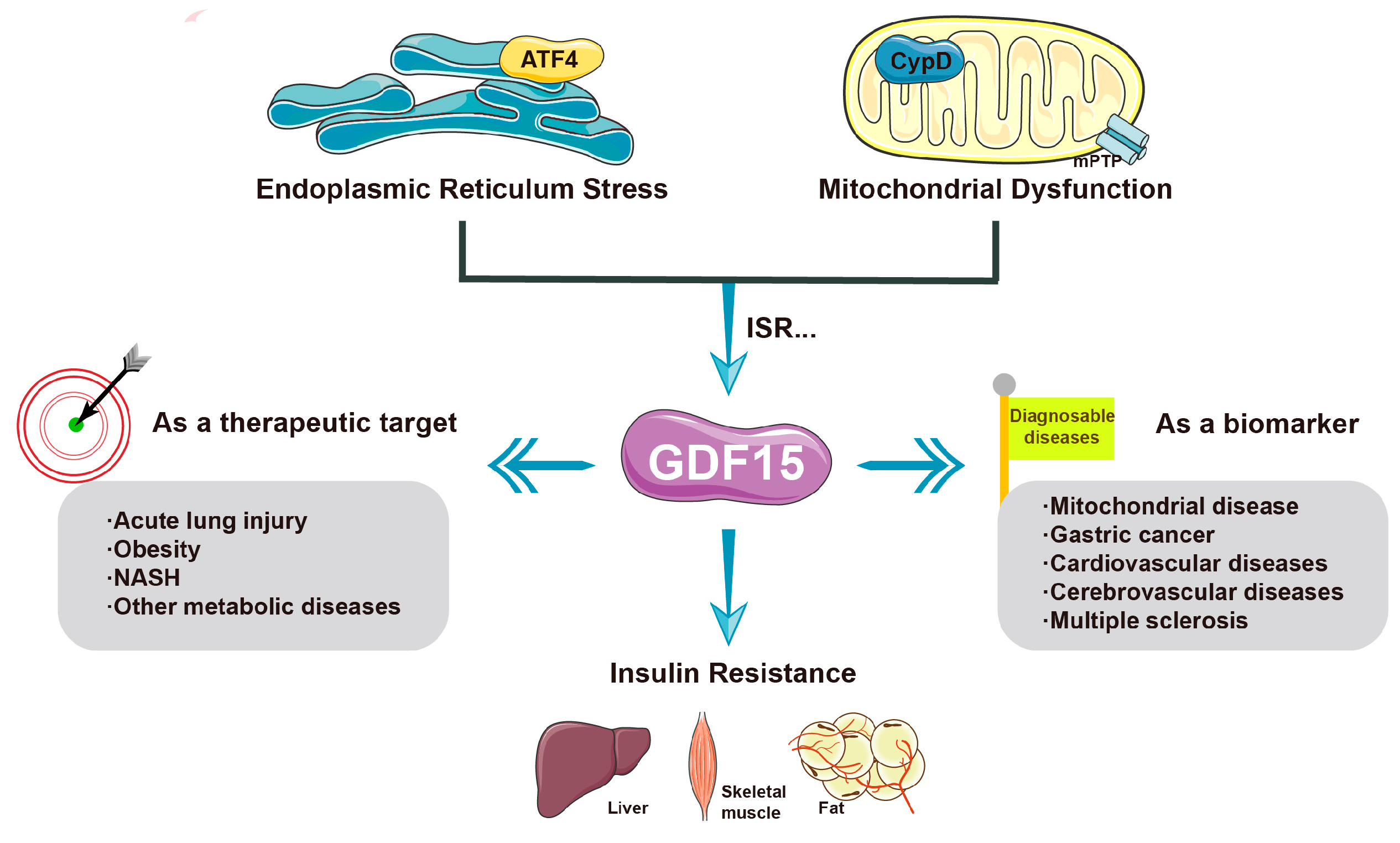

Table 1.
A brief summary of the relevant mechanisms mentioned in the review.
| Factor | Mechanism |
|---|---|
| AGEs | Promotes mitochondrial stress by activating NADPH |
| CypD | Promotes mitochondrial stress when the MPTP is open and NEFAs are present |
| PPARs | Inhibit mitochondrial stress by inhibiting IL-6 |
| miR-126 | Inhibits mitochondrial stress by activating SIRT1 and SOD |
| miR-30c | Inhibits mitochondrial stress by activating PGC-1 and PPAR-α |
| HSP60 | Mitochondrial stress causes overexpression of HSP60; HSP60 promotes mTORC1-SREBP1 signal transduction and promotes insulin resistance together with endoplasmic reticulum stress |
| FGF21 | Metformin induces mitochondrial stress through the Perk-eIF2α-ATF4 axis; mitochondrial stress promotes FGF21 expression by activating the comprehensive stress response (ISR); FGF21 improves insulin resistance |
| GDF-15 | GDF-15 improves insulin resistance; CYPD may cause insulin resistance by inhibiting GDF-15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



