
Submitted:
19 May 2023
Posted:
22 May 2023
You are already at the latest version
Abstract
Since Darwin’s “Power of movement in plants” the precise mechanism of oscillatory plant growth remains elusive. Hence the search continues for the hypothetical growth oscillator that regulates a huge range of growth phenomena ranging from circumnutation to pollen tube tip growth and stomatal movements. Oscillators are essentially simple devices with few components. A universal growth oscillator with only four major components became apparent recently with the discovery of a missing component, notably arabinogalactan glycoproteins (AGPs) that store dynamic Ca2+ at the cell surface. Demonstrably, auxin-activated proton pumps, AGPs, Ca2+ channels and auxin efflux “PIN” proteins, embedded in the plasma membrane, combine to generate cytosolic Ca2+ oscillations that ultimately regulate oscillatory growth: Hechtian adhesion of the plasma membrane to the cell wall and auxin-activated proton pumps trigger the release of dynamic Ca2+ stored in periplasmic AGP monolayers. These four major components represent a molecular PINball machine a strong visual metaphor that also recognizes auxin efflux “PIN” proteins as an essential component. Proton “pinballs” dissociate Ca2+ ions bound by paired glucuronic acid residues of AGP glycomodules, hence reassesses the role of proton pumps. It shifts the prevalent paradigm away from the recalcitrant “acid growth” theory that proposes direct action on cell wall properties, with an alternative explanation that connects proton pumps to Ca2+ signaling with dynamic Ca2+ storage by AGPs, auxin transport by auxin-efflux PIN proteins and Ca2+ channels. The extensive Ca2+ signalling literature of plants ignores arabinogalactan proteins (AGPs). Such scepticism leads us to reconsider the validity of the universal growth oscillator proposed here with some exceptions that involve marine plants and perhaps the most complex stress-test, stomatal regulation.
Keywords:
calcium homeostasis
; AGPs
; proton pump
; cell wall
Introduction
About four hundred million years ago bryophytes evolved an upright sporophyte with a terminal sporogenous capsule that enhanced aerial spore dissemination but created the opposing demands of water retention while permitting carbon dioxide entry. Development of a waterproof cuticle saved water but restricted CO2 absorption; that was solved by the evolution of sporophyte stomata. Pairs of kidney-shaped guard cells form stomatal pores that respond to ambient conditions by opening and closing and thus allow a trade-off between water retention and CO2 entry. A critical balance depends on the osmotic generation of turgor pressure and the elasticity of guard cell walls with anisotropic reinforcement. Guard cell turgor depends on the regulated influx and efflux of specific osmolytes. Opening and closing are distinct processes; both depend on Ca2+ signalling. However, a simple model remains elusive. Even after fifty years the extensive literature shows that major problems remain stubbornly unresolved. That implies a major piece of the puzzle is missing. Certainly, the glaring omission of AGPs from current discussion is a strong hint of their possible relevance especially considering their location and Ca2+ binding properties. The following discussion outlines how AGPs may contribute to the Ca2+homeostasis that underlies the mechanical dynamics of stomatal function. This may resolve the apparent paradox of Ca2+ required for both opening and closing stomata [1,2,3,4].
This brief personal historical perspective began in 1958 in Cambridge in the late fifties then the epicentre of biochemical energetics represented by David Keilin, Robin Hill, Peter Mitchell, A.F.Huxley and Alan Hodgkin. Cambridge was also the epicentre of structural biochemistry spearheaded by Frances Crick’s elucidation of the DNA double helix, Fred Sanger’s breakthrough in sequencing protein (then DNA) and protein X-Ray diffraction by Perutz and Kendrew. Peter Mitchell’s discovery of chemiosmosis and the proton motif force generated by proton gradients across a membrane is one of the great Nobel Prizes [5]. It paved the way to a profound understanding of biochemical energetics that powers all life on planet earth. It solved the great biochemical mystery of ATP generation and beautifully integrates David Keilin’s electron transport chain. It was also apparent that while proton gradients in one direction generate ATP, in the reverse direction ATP can pump protons out of the cell to establish a membrane electrostatic potential that enables cation import. Those were the two major roles of membrane proton pumps until the recent discovery of its third role, notably in Ca2+ homeostasis that was quite unanticipated. The Plant Kingdom has recruited reverse proton gradients; a novel extrapolation of Mitchell’s proton pump integrates auxin-controlled growth with cell surface glycoproteins and Ca2+ signalling as a major regulator of plant growth (Figure 1). This contrasts with the Animal Kingdom where reverse proton gradients contribute mainly to indigestion and medication with proton pump inhibitors.
A Venn diagram: four components embedded in the plasma membrane generate cytosolic Ca2+.
- Auxin efflux PIN proteins transport auxin
- Auxin activates proton pump to generate protons
- Protons release Ca2+ from AGP-Ca2+
- Open Ca2+ channels supply cytosolic Ca2+
Cell surface arabinogalactan glycoproteins (AGPs) were for many years with only modest evidence [6] considered signalling proteins until comprehensive structural carbohydrate analyses of Li Tan and Marcia Kieliszewski at Ohio University elucidated a consensus structure [7] of the repetitive arabinogalactan glycomodule: A 3D-molecular modelling Eureka moment revealed AGP function in one fell swoop! Two glycomodule sidechains terminated by glucuronic acid residues were potential Ca2+-binding sites [8]. Further direct assay and molecular dynamics simulations confirmed that observation. Quite unlike Ca2+ stores of animal cells, plants locate their dynamic Ca2+ storage at the most strategic point of entry, the plasma membrane! Release of dynamic Ca2+ then identifies a novel function of proton pump activity.
Another centre of activity involved National Service at Number two Radio School, RAF Yatesbury where two years as an instructor in radio theory, a focus on oscillators became the essential backdrop to biological oscillators that later identified the AGP-Ca2+-dependent Ca2+ oscillator [8]. Over 25 years ago oscillatory growth was correlated with oscillations in the pollen tube tip focussed Ca2+ gradient; this suggested Ca2+ influx could originate from external stores in the cell wall [9]. However, AGPs at the pollen tube tip [10] became significant when the recent discovery of AGP-Ca2+ and the cell surface Ca2+ oscillator explained its crucial role in pollen tube growth. The wide ramifications include not only tip growth of pollen tubes but numerous other plant processes such as phyllotaxis and even stomatal movements![11] .
To some these ramifications may appear contentious. They merit further consideration. How does this simple pinball hypothesis involve such profound interactions? Mitchell’s proton pump and ATP generation drives virtually all biochemical processes. In both plants and animals a dual role of proton pumps generates ATP and also the negative potential that drives cation influx across the plasma membrane. However, a third major role unique to plants recruits proton pumps that generate reverse proton gradients to enable Ca2+ homeostasis that regulates plant growth and differentiation thus exemplifying the parsimony of nature (Figure 1).
These broad generalisations might appear as extravagant claims! The following sections outline the historical background and discuss the pro and cons of the pinball hypothesis particularly in the light of recent papers. It concludes with stomatal regulation as a stress-test of stomatal dynamics as the most complex example:
Historical Back Ground
Sydney Ringer’s serendipitous discovery of Ca2+-dependent muscle contraction [12] was followed somewhat later by identification of Ca2+ as essential for pollen germination [13]. These discoveries stimulated huge interest in both animals and plants and generated a prolific literature over many years. Animal cells have two sources of dynamic Ca2+, a plentiful external supply and a more limited internal store within the endoplasmic reticulum (ER) released to the cytosol by inositol triphosphate IP3 of the Ca2+-signalling phosphoinositide (PI) pathway [14]. Many consider that this pathway also operates in plants [15] reminiscent of T.H. Huxley’s famous but misleading aphorism that “A plant cell is an animal in a wooden box.” However, profound differences between animal and plant cells include AGPS unique to plants and the missing link in Ca2+ homeostasis: Indeed, it is now clear that plants store dynamic Ca2+ externally as periplasmic AGP-Ca2+ rather than in the ER. One hardly expects immediate acceptance of this model. Time sifts. Paradigms shift. Indeed our inference extrapolates Peter Mitchell’s chemiosmotic model that couples proton gradients across mitochondrial membranes with ATP generation [5] (Nobel Prize 1978). Across plasma membranes such gradients also involve proton secretion. Plants have uniquely recruited that property to regulate Ca2+homeostasis and growth. Somehow, plants have bypassed the enormous complexities of hormonal regulation in animals by an elegant strategy that regulates plant growth by unifying an auxin-activated proton pump with AGPs and Ca2+ homeostasis.
Simplicity
A friendly critic ignoring Occam’s razor considers the four component oscillator as “too simple”. However, each component has many forms, notably eleven proton pumps in Arabidopsis AHA1 to AHA11, and nine PIN auxin efflux proteins that solve the long standing problem of polar auxin transport. Together with multiple Ca2+ channels [16] and AGPs the oscillator is superbly adaptable.
The Mathematical Basis
Previous attempts to impose mathematical rules on plant growth [17] lack the essential biochemical details described here. Although one might expect some mathematical insight, the detailed structure of the crucial AGP glycomodule with paired glucuronic acid residues based on impeccable NMR analyses [18] and molecular simulations of Ca2+-binding [8] all depend on sophisticated mathematical technique.
Cytosolic Ca2+: is AGP Glucuronic Acid the Source?
It is generally assumed that Ca2+ freely available in the in the plant apoplast is the immediate source of dynamic cytosolic Ca2+ as well as internal stores [15] thus similar to the classical animal model [14] that involves storage in the ER and vacuole with release mediated by inositol triphosphate (IP3), but fundamentally quite different from the model described here which is specific to plants.
The crucial role of the proton pump in generating dynamic cytosolic Ca2+ in plant cells is clearly evidenced by pollen tube tip-focussed Ca2+ influx, summarised by >30 years work of [19]. However, many papers ignore the proposed role of AGPs as a precursor of cytosolic Ca2+ despite the evidence of AGP periplasmic location [20] and detailed structure of AGP glycomodules.... paired sidechains with terminal glucuronic acid residues that enable stoichiometric Ca2+-binding essential to the Ca2+ storage function of AGPs.
Every model has a weak link: Are glucuronic acid residues essential? Theory predicts but experiment decides: The 2:1 glucuronic : Ca2+ stoichiometry [8] suggests a critical test by generating classical AGPs that lack glucuronic acid. Although only a “minor” component, glucuronic acid is invariably present in classical AGPs, but more abundant in AGPs of plants like Eelgrass, monocot Zostera marina that have returned to the high salt levels of a marine environment. [21]. Presumably, the increased glucuronic acid content is an evolutionary adaptation that enhances Ca 2+ binding, signifying a pivotal ecological role for AGPs [22].
However, multiple glucuronosyl transferases (GlcATs) of the Arabidopsis genome create significant genetic redundancy and therefore a challenge to functional analysis.
GlcAT Knockouts Affect Ca2+ Homeostasis
A critical test of the AGP-Ca2+ hypothesis by two groups with substantial contributions to AGP structural chemistry generated multiple mutants that lack AGP glucuronic acid, the Dupree group at Cambridge UK and the Showalter group at Athens Ohio. The genetic redundancy of glucuronosyl transferases that add GLcA to AGP polysaccharides involved complementary approaches to the generation of multiple GlcAT gene knockouts: Firstly, T-DNA insertion lines involving Ti plasmid transformation mediated by Agrobacterium tumefaciens generated Arabidopsis mutants in four glucuronosyl transferases (GlcAT14A, -B, -D, and -E) [23].
Secondly, similar Arabidopsis mutants generated by the CRISPRCas9 multiplexing approach for GLcAT14A, GLcAT14B, and GLcAT14C [24].
A comprehensive and detailed study of plant growth by the Dupree group showed that multiple glucuronosyl knockouts severely impaired Ca2+ homeostasis. This was consistent with greatly decreased glucuronidation and Ca2+-binding capacity of AGPs isolated from leaves and roots of glcat14a/b/d mutants [23], while growth of the glcat14a/b/e triple mutant was poor. Other mutants showed numerous pleiotropic growth effects: shorter plants, decreased leaf expansion (including cell shape formation and expansion), shorter hypocotyls lacking apical hook, trichomes with decreased branching and abnormal Ca2+ transients in roots with attenuated Ca2+ wave propagation. Significantly, exogenous Ca2+ supressed/rescued these AGP glucuronidation mutant phenotypes!
A parallel study by the Showalter group focussed on sexual reproduction and the role of multiple glucuronosyl transferase (Glcat) mutants in Arabidopsis. Over many years significant contributions to AGP biology by the Showalter group have demonstrated widespread involvement of AGPs encoded by 85 genes and eleven GLCAT genes. Their current work demonstrates that multiple Glcat mutants significantly impair sexual reproduction, particularly pollen development, polytubey block, and successful fertilisation that requires cytoplasmic calcium oscillations in synergid cells after physical contact with the pollen tube tip followed by normal embryo development in Arabidopsis.
Taken together the Dupree and Showalter groups corroborate the pinball model and support a unified theory of plant growth based on the properties of membrane glycoproteins.
Multiple glucuronosyl transferase knockouts severely impair Ca2+ homeostasis, therefore Glucuronic-bound Ca2+is the essential source of cytosolic Ca2+! The novel idea of a Ca2+ store strategically located at the surface of the plant plasma membrane was previously overlooked because Mother Nature keeps some of her best secrets well-hidden!
Pinball Machine is Random?
In a conventional pinball machine pinball trajectories are largely random. Thus, although a strong visual metaphor a molecular pinball machine (Figure 1) has limitations but clear advantages: All four are components of the plasma membrane that emphasises their close physical proximity, hence a single entity integral to their overall function. Indeed, while the trajectory of physical pinballs may appear random the plasma membrane is far from random consisting of a highly organised hydrophobic lipid surface covered by a hydrophilic glycoprotein layer of AGPs [20]. At such a hydrophobic-hydrophilic interface nanoconfined water may behave as an interfacial layer where protons diffuse within a matrix with high ionic conductivity that occurs through surface hopping of protons [25]. Thus proton conduction connects the proton pump with AGP glycomodules and may explain very rapid responses. The glycomodule structure itself may facilitate proton conduction to the Ca2+-binding centre.
What Connects Auxin with Ca2+ Signaling?
Although a connection between auxin and Ca2+ has been apparent for many years [26], both auxin and fusicoccin activate the proton pump [27] with a concomitant rapid elevation of cytosolic Ca2+. While Vanneste & Friml concluded [28] that “the role of auxin-induced Ca2+ signaling is poorly understood” Dindas and colleagues confirmed [29] a close connection, now described by the pinball model where periplasmic AGPs are a sine qua non of Ca2+-mediated metabolic regulation. An alternative explanation postulates complex auxin activated Ca2+ channels [30]: Auxin (IAA) enters cells through the protein AUX1 and binds to a protein of the TIR1/AFB family. This leads to rapid ion flow (within seconds) through the protein AHA and a Ca2+ channel. While they demonstrate Ca2+ influx within seconds of IAA application, it completely ignores the source of dynamic Ca2+ bound by cell surface AGP-Ca2+ stores, hence overlooks the direct role of the proton pump.
Origin of Growth Oscillations and Ion Fluxes
Oscillations in growth and cytosolic Ca2+ fluxes of pollen tube tips [31,32] and root hairs [33] suggest a fundamental growth oscillator; but what activates the oscillator to initiate these oscillations? The pinball model identifies the proton pump as a prime candidate triggered by auxin and mechanostress. While Ca2+ oscillations have been described as inherent in root hairs and pollen tubes [31] ubiquitous auxin efflux “PIN” proteins transport auxin throughout the plant [34,35].
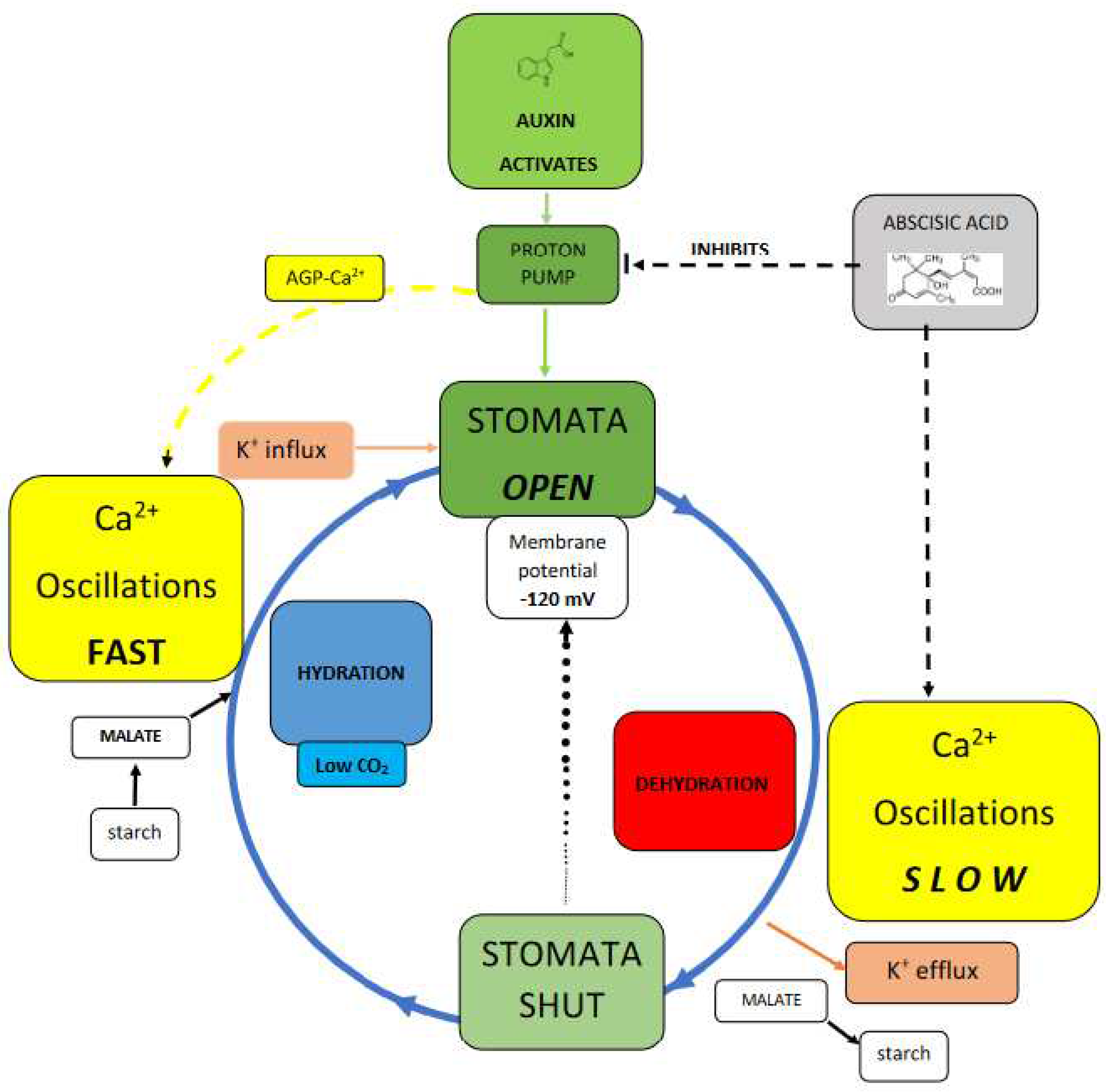
Stomata Test the Pinball Hypothesis
Proton pumps drive a pinball machine that generates cytosolic Ca2+.
Auxin is the accelerator and abscisic acid is the brake:
In hydrated cells an auxin-activated pump dissociates AGP-Ca2+ that triggers rapid Ca2+ oscillations thus increasing osmolytes K+, malate and turgor.
In stressed cells abscisic acid inactivates the proton pump thus decreasing Ca2+ oscillations, osmolytes and turgor.
A contribution to current descriptions of stomatal regulation runs the risk of over simplification. (Figure 2). Balancing opposing demands of water conservation and CO2 uptake involves a complex guard cell wall structure and numerous signals mediating subtle control of osmolytes and turgor pressure; they include the Ca2+signal and its oscillations discussed in a recent comprehensive review [36] that assumes the source of cytosolic Ca2+ is largely from endomembrane stores, now almost an article of faith, although at the growing tip of the pollen tube the cytosolic Ca2+ source is quite clearly external! Nevertheless, “Best estimates indicate that endomembrane release accounts for more than 95% of the Ca2+ entering the cytosol” [37]. That seems inaccurate as it ignores AGPs the missing link connecting AGP-Ca2+ with the proton pump whose primary role maintains a large negative membrane potential essential for stomatal opening via influx of K+ osmolyte and concomitant malate synthesis that increase turgor pressure. Significantly, treatment with auxin and fusicoccin also open stomata similar to the constitutively fully active proton pump mutant AHA1 [38].
Arguably, the proton pump regulates Ca2+ homeostasis. This is a more subtle role than maintaining membrane potential but of hitherto unappreciated significance. Thus, immunolocalisation of AGPs and detection of their abundance in guard cells by the Yariv reagent [39] confirm Ca2+ involvement in stomatal regulation. However, precisely how plants achieve specificity in intracellular calcium signaling is largely speculative. [38]. Some suggest [40] that “Oscillations in cytosolic Ca2+ allow for information to be encoded in both the amplitude and frequency.” This “calcium code” has spawned a voluminous literature dealing with stomatal closure and associated Ca2+ oscillations; their frequency and amplitude trigger opening and closure by cation osmolyte influx and efflux respectively. Not surprisingly metabolic activity highest in turgid cells triggers highest cytosolic Ca2+ oscillations [2]. In open stomata the high malate concentration [25 µM cytosol; 464 µM vacuole [36] is of particular significance as malate chelates excess Ca2+ that might otherwise overwhelm the cytosol.
The proton pump acts as a central hub regulated by virtually all factors known to regulate plant growth, hormones, and classical plant hormones [41] generating the complexity summarised by [42].
Figure 2 compares turgid and flaccid stomatal guard cells:
In turgid guard cells the proton pump activated by auxin releases Ca2+ from AGP-Ca2+ and also generates the membrane potential. Thus AGP-Ca2+ dissociation initiates cytosolic Ca2+ oscillations while a large membrane potential enables K+ influx increasing cytosolic osmolyte levels including malate derived from starch. These Ca2+ oscillations provide the cytosolic Ca2+ that triggers vesicle exocytosis: [43,44]. Indeed, the pivotal metabolic role of Ca2+ is evidenced by the over 200 EF-hand Ca2+-binding proteins as key transducers mediating Ca2+ action encoded in the Arabidopsis thaliana genome [45].
In flaccid cells osmotic stress and dehydration generate abscisic acid that triggers stomatal closure [46] by inactivating proton pump activity [47]. This results in membrane depolarisation that enables guard cells to jettison K+ osmolyte via the specific K+ efflux channel GORK [48] and recycle malate as starch. Thus transition between the turgid and flaccid cell states depend on Ca2+ oscillations that regulate osmolyte influx and efflux respectively; increased cytosolic Ca2+ opens stomata [2,26,49] }. On the other hand the flaccid guard cells of closed stomata are virtually quiescent with low metabolic demands just sufficient to open K+ efflux channels and convert malate to starch. Presumably such low demands reflect the much weaker Ca2+ oscillations hence a low level of metabolic activity. This resolves the paradox, depicted in Figure 2 that represents a simplified scenario.
Final Conclusions
This review recapitulates recent significant progress in AGP biology represented by the molecular pinball machine of the plasma membrane that unifies proton pumps, AGPs and Ca2+ signalling [50].
Here we presented a critical test of the pinball hypothesis and suggested how it resolves the paradox that Ca2+ can both open and close stomata, with AGPs as the missing link and source of cytosolic Ca2+. Arguably, the proton motive force controls both opening and closing of stomata. Thus positive and negative regulation of the proton pump involve an auxin accelerator and an abscisic acid brake. That simple analogy explains how low Ca2+ levels close stomata, emphasizing elegant control of both water loss and carbon dioxide gain!
Recent work validates the pinball hypothesis and is a suitable closure to a research career extending over sixty years initiated by the discovery of hydroxyproline rich cell wall proteins [51] in 1960. Plants have uniquely recruited hydroxyproline-rich proteins, extensins, AGPS and their allies, to serve dynamic functions that range from direct involvement in cell division [52] and gravitropism [53] to Ca2+ homeostasis and its related tropisms. It has taken many years to appreciate the versatility of these cell surface glycoproteins.
References
- Ng, C.K.-Y.; Mcainsh, M.R.; Gray, J.E.; Hunt, L.; Leckie, C.P.; Mills, S.; Hetherington, A.M. Calcium-based signalling systems in guard cells. New Phytol. 2001, 151, 109–120. [Google Scholar]
- Young, J.J.; Mehta, S.; Israelsson, M.; Godoski, J.; Grill, E.; Schroeder, J.I. CO2 signaling in guard cells: Calcium sensitivity response modulation, a Ca2+-independent phase, and CO2 insensitivity of the gca2 mutant. Proc. Natl. Acad. Sci. USA 2006, 163, 7506–7511. [Google Scholar]
- Kim, T.-H.; Bohmer, M.; Hu, H.; Nishimura, N.; Schroeder, J.I. Guard Cell Signal Transduction Network:Advances in Understanding Abscisic Acid, CO2, and Ca2+ Signaling. Annu. Rev. Plant Biol. 2010, 61, 561–591. [Google Scholar]
- Parramona, C.M.; Wang, Y.; Hills, A.; Vialet-Chabrand, S.; Griffiths, H.; Rogers, S.; Lawson, T.; Lew, V.L.; Blatt, M. An Optimal Frequency in Ca2+ Oscillations for Stomatal Closure Is an Emergent Property of Ion Transport in Guard Cells. Plant Physiol. 2016, 170, 33–42. [Google Scholar]
- Mitchell, P. David Keilin's Respiratory Chain Concept and Its Chemiosmotic Consequences. Nobel Lect. Chem. 1978, 1971–1980, 295–330. [Google Scholar]
- Silva, J.; Ferraz, R.; Dupree, P.; Showalter, A.M.; Coimbra, S. Three Decades of Advances in Arabinogalactan-Protein Biosynthesis. Front. Plant Sci. 2020, 11, 610377. [Google Scholar]
- Tan, L.; Varnai, P.; Lamport, D.T.A.; Yuan, C.; Xu, J.; Qiu, F.; Kieliszewski, M.J. Plant O-Hydroxyproline Arabinogalactans Are Composed of Repeating Trigalactosyl Subunits with Short Bifurcated Side Chains. J. Biol. Chem. 2010, 285, 24575–24583. [Google Scholar]
- Lamport, D.T.A.; Varnai, P. Periplasmic arabinogalactan glycoproteins act as a calcium capacitor that regulates plant growth and development. New Phytol. 2013, 197, 58–64. [Google Scholar]
- Holdaway-Clarke, T.; Feijo, A.; Hackett, G.R.; Kunkel, J.G.; Hepler, P.K. Pollen Tube Growth and the lntracellular Cytosolic Calcium Gradient Oscillate in Phase while Extracellular Calcium Influx is Delayed. Plant Cell 1997, 9, 1999–2010. [Google Scholar]
- Mollet, J.C.; Kim, S.; Jauh, G.Y.; Lord, E.M. Arabinogalactan proteins, pollen tube growth, and the reversible effects of Yariv phenylglycoside. Protoplasma 2002, 219, 89–98. [Google Scholar]
- Lamport, D.T.A.; Tan, L.; Held, M.A.; Kieliszewksi, M.J. The Role of the Primary Cell Wall in Plant Morphogenesis. Int. J. Mol. Sci. 2018, 19, 2674. [Google Scholar]
- Ringer, S. A further Contribution regarding the influence of the different Constituents of the Blood on the Contraction of the Heart. J. Physiol. 1883, 4, 29–42. [Google Scholar]
- Brewbaker, J.L.; Kwack, B.H. The essential role of calcium ion in pollen germination and pollen tube growth. Am. J. Bot. 1963, 50, 859–865. [Google Scholar]
- Berridge, M.J. Elementary and global aspects of calcium signalling. J. Physiol. 1997, 499, 291–306. [Google Scholar]
- Franklin-Tong, V.E. Signaling and the modulation of pollen tube growth. Plant Cell 1999, 11, 727–738. [Google Scholar]
- Very, A.-A.; Sentenac, H. Cation channels in the Arabidopsis plasma membrane. Trends Plant Sci. 2002, 7, 168–175. [Google Scholar]
- Turing, A.M. The chemical basis of morphogenesis. Phil Trans. R. Soc. Lond. B 1952, 237, 37–72. [Google Scholar]
- Tan, L.; Xu j Held, M.A.; Lamport, D.T.A.; Kieliszewski, M. Arabinogalactan Structures of Repetitive Serine-Hydroxyproline Glycomodule Expressed by Arabidopsis Cell Suspension Cultures. Plants 2023, 12, 1036. [Google Scholar]
- Hepler, P.K. The Cytoskeleton and Its Regulation by Calcium and Protons. Plant Physiol. 2016, 170, 3–22. [Google Scholar]
- Lamport, D.T.A.; Kieliszewski, M.J.; Showalter, A.M. Salt stress upregulates periplasmic arabinogalactan proteins: Using salt stress to analyze AGP function. New Phytol. 2006, 169, 479–492. [Google Scholar]
- Pfeifer, L.; Shafee, T.; Johnson, K.L.; Bacic, A.; Classen, B. Arabinogalactan-proteins of Zostera marina L. contain unique glycan structures and provide insight into adaption processes to saline environments. Nat. Sci. Rep. 2020, 10, 8232. [Google Scholar]
- Pfeifer, L.; Classen, B. The Cell Wall of Seagrasses: Fascinating, Peculiar and a Blank Canvas for Future Research. Front. Plant Sci. 2020, 11, 588754. [Google Scholar]
- Lopez-Hernandez, F.; Tryfona, T.; Rizza, A.; Yu, X.L.; Harris, M.O.B.; Webb, A.A.R.; Kotake, T.; Dupree, P. Calcium Binding by Arabinogalactan Polysaccharides Is Important for Normal Plant Development. Plant Cell 2020, 32, 3346–3369. [Google Scholar]
- Ajayi, O.O.; Held, M.A.; Showalter, A.M. Three β-Glucuronosyltransferase Genes Involved in Arabinogalactan Biosynthesis Function in Arabidopsis Growth and Development. Plants 2021, 10, 1172. [Google Scholar]
- Kapil, V.; Schran, C.; Zen, A.; Chen, J.; Pickard, C.J.; Michaelides, A. The first-principles phase diagram of monolayer nanoconfined water. Nature 2022, 609, 512–517. [Google Scholar]
- Irving, H.R.; Gehring, C.A.; Parish, R.W. Changes in cytosolic pH and calcium of guard cells precede stomatal movements. Proc. Natl. Acad. Sci. USA 1992, 89, 1790–1794. [Google Scholar]
- Chater, C.; Kamisugi, Y.; Movahedi, M.; Fleming, A.; Cuming, A.C.; Gray, J.E.; Beerling, D.J. Regulatory Mechanism Controlling stomatal Behavior Conserved across 400 Million Years of Land Plant Evolution. Curr. Biol. 2011, 21, 1025–1029. [Google Scholar]
- Vanneste, S.; Friml, J. Calcium: The missing link in auxin action. Plants 2013, 2, 650–675. [Google Scholar]
- Dindas, J.; Scherzer, S.; Roelfsema, M.R.G.; von Meyer, K.; Muller, H.M.; Al-Rasheid, K.A.S.; Palme, K.; Dietrich, P.; Becker, D.; Bennett, M.J.; Hedrich, R. AUX1-mediated root hair auxin influx governs SCFTIR1/AFB-type Ca2+ signaling. Nat. Commun. 2018, 9, 1174. [Google Scholar]
- Murphy, A.S.; Peer, W.A. An auxin-binding protein resurfaces after deep dive. Nature 2022, 609, 475–476. [Google Scholar]
- Baluska, F.; Mancuso, S. Root apex transition zone as oscillatory zone. Front. Plant Sci. 2013, 4, 354. [Google Scholar]
- Hepler, P.K. The Cytoskeleton and Its Regulation by Calcium and Protons. Plant Physiol. 2016, 170, 3–22. [Google Scholar]
- Steger, A.; Palmgren, M. Root hair growth from the pH point of view. Front. Plant Sci. 2022, 13, 949672. [Google Scholar]
- Habets, M.E.; Offringa, R. PIN-driven polar auxin transport in plant developmental plasticity: A key target for environmental and endogenous signals. New Phytol. 2014, 203, 362–377. [Google Scholar]
- Lamport, D.T.A.; Tan, L.; Held, M.A.; Kieliszewksi, M.J. Phyllotaxis Turns Over a New Leaf-A New Hypothesis. Int. J. Mol. Sci. 2020, 21, 1–15. [Google Scholar]
- Jezek, M.; Blatt, M. The Membrane Transport System of the Guard Cell and Its Integration for Stomatal Dynamics. Plant Physiol. 2017, 174, 487–519. [Google Scholar]
- Parramona, C.M.; Wang, Y.; Hills, A.; Vialet-Chabrand, S.; Griffiths, H.; Rogers, S.; Lawson, T.; Lew, V.L.; Blatt, M. An Optimal Frequency in Ca2+ Oscillations for Stomatal Closure Is an Emergent Property of Ion Transport in Guard Cells. Plant Physiol. 2016, 170, 33–42. [Google Scholar]
- Kim, T.-H.; Bohmer, M.; Hu, H.; Nishimura, N.; Schroeder, J.I. Guard Cell Signal Transduction Network:Advances in Understanding Abscisic Acid, CO2, and Ca2+ Signaling. Annu. Rev. Plant Biol. 2010, 61, 561–591. [Google Scholar]
- Giannoutsou, E.; Apostolakos, P.; Galatis, B. Spatio-temporal diversification of the cell wall matrix materials in the developing stomatal complexes of Zea mays. Planta 2016, 244, 1125–1143. [Google Scholar]
- Ng, C.K.-Y.; Mcainsh, M.R.; Gray, J.E.; Hunt, L.; Leckie, C.P.; Mills, S.; Hetherington, A.M. Calcium-based signalling systems in guard cells. New Phytol. 2001, 151, 109–120. [Google Scholar]
- Falhof, J.; Pedersen, J.T.; Fuglsang, J.T.; Palmgren, M. Plasma Membrane H+ATPase Regulation in the Center of Plant Physiology. Mol. Plant 2016, 9, 323–337. [Google Scholar] [PubMed]
- Peak, D.; Hogan, M.T.; Mott, K.A. Stomatal patchiness and cellular computing. Proc. Natl. Acad. Sci. USA 2023, 120, e2220270120. [Google Scholar] [PubMed]
- Battey, N.H.; James, N.C.; Greenland, A.J.; Brownlee, C. Exocytosis and Endocytosis. Plant Cell 1999, 11, 643–659. [Google Scholar] [PubMed]
- Campanoni, P.; Blatt, R. Membrane trafficking and polar growth in root hairs and pollen tubes. J. Exp. Bot. 2007, 58, 65–74. [Google Scholar]
- Day, I.S.; Reddy, V.S.; Ali, G.S.; Reddy, A.S.N. Analysis of EF-hand-containing proteins in Arabidopsis. Genome Biol. 2002, 3, 0056–1. [Google Scholar]
- Chen, Z.-H.; Hills, A.; Batz, U.; Amtmann, A.; Lew, V.L.; Blatt, M. Systems Dynamic Modeling of the Stomatal Guard Cell Predicts Emergent Behaviors in Transport, Signaling, and Volume Control. Plant Physiol. 2020, 159, 1235–1251. [Google Scholar]
- Hayashi, Y.; Takahashi, K.; Inoue, S.; Kinoshita, T. Abscisic Acid Suppresses Hypocotyl Elongation by Dephosphorylating Plasma Membrane H+-ATPase in Arabidopsis thaliana. Plant Cell Physiol. 2014, 55, 845–853. [Google Scholar]
- Adem, G.D.; Chen, G.; Shabala, L.; Chen, Z.-H.; Shabala, S. GORK Channel: A Master Switch of Plant Metabolism? Trends Plant Sci. 2020, 25, 434–445. [Google Scholar]
- Brandt, B.; Munemasa, S.; Wang, C.; Nguyen, D.; Yong, T.; Yang, P.G.; Poretsky, E.; Waadt, R.; Aleman, F.; Schroeder, J.I. Calcium specificity signaling mechanisms in abscisic acid signal transduction in Arabidopsis guard cells. eLife 2015, 4, e03599. [Google Scholar]
- Lamport, D.T.A.; Tan, L.; Kieliszewksi, M.J. A Molecular Pinball Machine of the Plasma Membrane Regulates Plant Growth—A New Paradigm. Cells 2021, 10, 10081935. [Google Scholar]
- Lamport, D.T.A.; Northcote, D.H. Hydroxyproline in primary cell walls of higher plants. Nature 1960, 188, 665–666. [Google Scholar]
- Cannon, M.C.; Terneus, K.A.; Hall, Q.; Tan, L.; Wang, Y.; Wegenhart, B.; Chen, L.; Lamport, D.T.A.; Chen, Y.; Kieliszewksi, M.J. Self-assembly of the plant cell wall requires an extensin scaffold. Proc. Natl. Acad. Sci. USA 2008, 105, 2226–2231. [Google Scholar] [PubMed]
- Lamport, D.T.A.; Tan, L.; Held, M.A.; Kieliszewski, M.J. Root-shoot gravitropism paradox resolved. Acad. Lett. 2022, 2022, 4998. [Google Scholar]
Figure 1.
The pinball machine encapsulates a role of the proton pump in Ca2+ homeostasis.
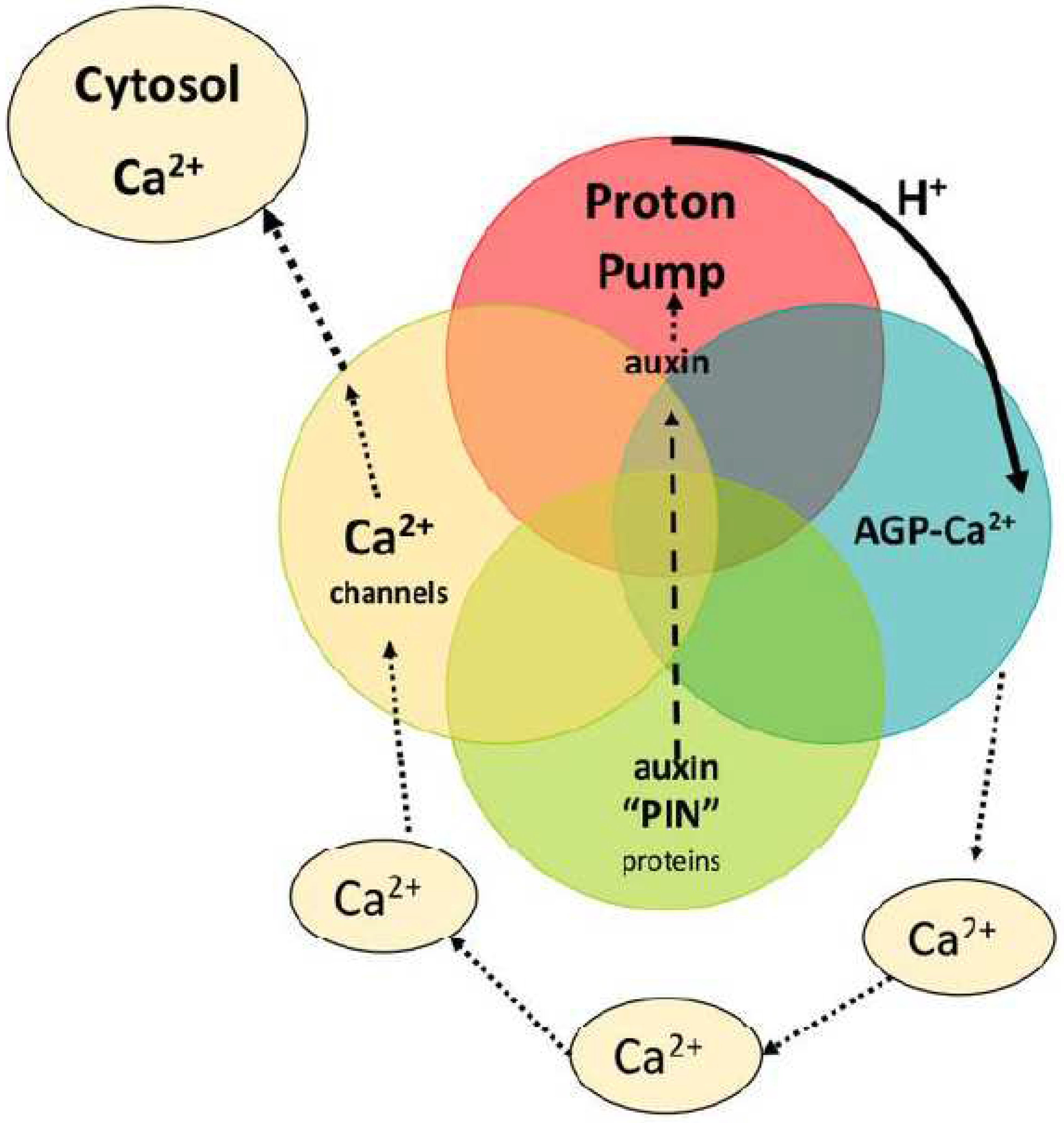
Figure 2.
Stomatal dynamics summarized.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



