
Submitted:
19 May 2023
Posted:
22 May 2023
You are already at the latest version
Abstract
The genus Amphichorda has been recently re-erected as an independent linage from Beauveria, circumscribed within Cordycipitaceae. However, its phylogenetic relationships with other members of this family remain obscure. In our on-going efforts to expand the knowledge on the diversity of culturable ascomycetes from the Mediterranean Sea, we isolated several specimens of Amphichorda. Preliminary sequence analyses revealed great phylogenetic distance with accepted Amphichorda species and a close relation to Onychophora coprophila. Onychophora is a monotypic genus of enteroblastic conidiogenous cells, presumably related to Acremonium (Bionectriaceae); while Amphichorda exhibits holoblastic conidiogenesis. Here, we examine representative strains of Amphichorda species to resolve the taxonomy of the genus and the above-mentioned fungi combining morphological, ultrastructure and multi-locus phylogenetic analyses (ITS, LSU, tef1, BenA). The results revealed Amphichorda as a member of the Bionectriaceae, where its asexual morphs represent a transition between enteroblastic and holoblastic conidiogenesis for this group of fungi. We also describe and illustrate Amphichorda littoralis sp. nov., and propose the new combination Amphichorda coprophila. In addition, we stablish key phenotypic features to distinguish Amphichorda species and demonstrate the higher salt tolerance degree of A. littoralis, consistent with its marine origin. This work provides a comprehensive framework for future studies in the genus.
Keywords:
Ascomycota
; asexual fungi
; marine fungi
; multi-locus phylogeny
; new taxa
; taxonomy
; ultrastructure
1. Introduction
The order Hypocreales (Sordariomycetes, Pezizomycotina, Ascomycota) currently comprises around 300 genera distributed across 17 families. Their species inhabit a wide range of substrates in terrestrial and aquatic (marine and freshwater) environments, and they show a great variety of lifestyles, such as saprobic, endophytic, and pathogenic fungi for plants and animals, including humans [1,2]. The Cordycipitaceae is one of the most complex families in the order due to the pathogenic behavior of most of its species, which includes a wide range of invertebrate hosts and results in a variety of morphological features of the sexual morph, primarily associated with its ascomata (stroma and perithecia). The asexual morph, however, are very similar, most frequently showing phialidic conidiogenous cells. Therefore, genera like Amphichorda, Beauveria, Cordyceps or Isaria have been difficult to circumscribe and recent works have dealt with numerous taxonomical problems [3,4]. In particular, the genus Amphichorda has been traditionally accepted as a member of Cordycipitaceae based on the taxonomical history of its type species, Amphichorda felina (= Beauveria felina). Despite this, the most recent multi-locus phylogeny of the family has resolved Amphichorda as a sister linage to the main Cordycipitaceae clade [4]. Indeed, the taxonomic status of Amphichorda has been controversial since its original description.
The genus Amphichorda was described by Fries in 1825 and typified by A. felina, a fungus isolated from cat dung in France and previously classified in the genus Clavaria [5,6]. The morphological description stated the white farinaceous color of the colonies and the production of filiform conidiogenous cells, which inspired the name of the genus. In 1832, A. felina was transferred to the genus Isaria, a genus that at this time lacked a type species and, therefore, comprised morphologically heterogenous fungi [7,8]. In 1972, de Hoog re-defined Isaria based on the production of synnemata and, following von Arx, accepted Isaria felina as the lectotype of the genus [9]. This circumscription, however, was rejected due to the previous lectotypification with the species Isaria farinosa [8,10]. Isaria felina was then transferred to the genus Beauveria based on the morphological resemblance of their holoblastic conidiogenous cells. Although, early phylogenetic analyses suggested great dissimilarity between B. felina and other Beauveria species [8]. Recently, the genus Amphichorda has been re-erected for the description of two novel species, Amphichorda cavernicola and Amphichorda guana, by Zhang et al. [11,12]. Zhang’s studies represent the first phylogenetic backbone for the genus Amphichorda and demonstrate the great phylogenetic distance between Amphichorda and Beauveria. Despite this, morphological differences between Amphichorda species are confusing and the phylogenetic relationships between Amphichorda and other genera remain obscure. Moreover, the type material of A. felina seems to be lost. Zhang et al. considered the strain CBS 250.34, the type of Isaria cretaceae, as the type strain of A. felina [12]. Isaria cretaceae was synonymized with A. felina by de Hoog [9], however, so far this strain has not been designated as the epitype of the species. Nevertheless, the type strain of I. cretacea was isolated from a package of moldy pressed yeast from Epsom, England [13] and, according to the criteria of substrate and geographical similarity required for fungal epityfication [14], the strain CBS 250.34 could not represent A. felina on the basis of its coprophilous origin from France [5,6]. Thus, the taxonomical status and the phylogenetic relationships of Amphichorda need to be revised.
In our latest efforts to expand the knowledge on the diversity of culturable ascomycetes from the Mediterranean Sea, we isolated several interesting specimens of an amphichorda-like fungus. A preliminary sequence analysis of the nuclear ribosomal operon (i.e., the 28S large ribosomal subunit-LSU, and the internal transcribed spacer -ITS, including the 5.8S rDNA gene) revealed that these specimens belong to the genus Amphichorda, but they did not fit into any of the described species. This preliminary sequence analysis also revealed that the marine strains were closely related to Onychophora coprophila. Onychophora is a monotypic genus, which conidiogenous apparatus morphologically resembles Amphichorda species. However, the conidiogenous cells of the former were described as enteroblastic (phialidic), while Amphichorda exhibits holoblastic conidiogenesis [11,12,15]. In addition, according to Mycobank and Index Fungorum databases, O. coprophila is a fungus of uncertain position among Ascomycota, although, the original authors suggested a possible relation to Acremonium (Bionectriaceae) [15].
The aim of the present study was, therefore, to clarify the taxonomy of the above-mentioned fungi based on morphological features, including ultrastructure, and multi-locus phylogenetic analyses inferred with sequences of the nuclear markers available for Amphichorda species. These are the ITS and LSU regions of the rDNA and partial fragments of the translation elongation factor 1-α (tef1) and the β-tubulin (BenA) regions. Then, we used these nuclear markers to determine the strains phylogenetically related to our strains and examined their available living cultures in order to assess the diversity within Amphichorda. In this work, we provide and update on the morphological and molecular diversity of Amphichorda, determine its phylogenetic relationships with other genera in Hypocreales and discuss the type strain for A. felina.
2. Materials and Methods
2.1. Sampling and strains
Sediment samples were collected from the Mediterranean Sea in two points of the Tarragona coast, the Miracle and Arrabassada beaches, through 2021 and 2022. Each beach was sampled twice, the Miracle beach on June and October 2021 and the Arrabassada beach on February and June 2022. These beaches are located in the southern part of Catalonia, right next to the port of Tarragona, the fifth most important harbor in Spain and an important stop for tourism cruise ships [16].
Marine sediments were collected following the same methodology at both locations. We stablished four collection points based on the sediment grain size and depth in the column of water. The first point was at 6 meters of depth (sand sediment), the second at 13 m (sand sediment), the third at 20 m (transition between sand and silt sediment) and the last point was at 30 m of depth (silt sediment). Four sub-samples were collected at each point ca 15 cm below the surface of the sea bed, using 50 mL sterile plastic containers, which were transported in a refrigerated container to the laboratory and processed immediately. For each sampling point, sediment sub-samples were mixed and vigorously shaken in a container; then, after 1 min at rest, the water was decanted and the sediment was poured into plastic trays with several layers of sterile filter paper to remove excess water.
Three agar media were used to achieve a greater ascomycetous diversity in culture and restrict the growth of certain fungal groups: dichloran rose-bengal-chloramphenicol agar (DRBC; 5 g peptone, 10 g glucose, 1 g KH2PO4, 0.5 g MgSO4, 25 mg rose-bengal, 200 mg chloramphenicol, 2 mg dichloran, 15 g agar, 1 L distilled water); 3% malt extract agar supplemented with sea water (SWMEA3%; 30 g malt extract, 5 g mycological peptone, 15 g agar, 1L sea water) was used as a suitable medium for the isolation of marine fungi [17] and potato dextrose agar (PDA; Condalab, Madrid, Spain) supplemented with 2 g/L of cycloheximide (PDA+A) was used to isolate strains resistant to this protein synthesis inhibitor, a frequent trait among fungi [18]. Both SWMEA3% and PDA+A culture media were supplemented with 5 mL of chloramphenicol (15 g/L ethanol) to prevent bacterial growth.
The culture methodology was as follows: 1 g of sediment from each sampling point was distributed across two Petri dishes and mixed with melted SWMEA3% at 45 °C; the same procedure was used to mix the sediment with PDA+A. In the case of the sediment mixed with DRBC, only 0.5 g of sediment from each sampling point was distributed across two Petri dishes to deal with fast growing fungi. A set of the plates of the different culture media was incubated at 22–24 °C and the other set at 15 °C to enable the detection of slow-growing fungi. Plates were stored in darkness and examined weekly by stereomicroscope for 5–8 weeks. Pure cultures were obtained from colony fragments or conidia of the fungi growing on primary plate cultures using a sterile dissection needle. These fragments were cultured on PDA and incubated at 25 °C in darkness. These PDA cultures were used for a preliminary morphological identification before DNA extraction.
Living cultures of putative novel or rare fungi were preserved and deposited in the culture collection of the Faculty of Medicine in Reus (FMR, Spain) for further studies. Taxonomic information and nomenclature for the new species were deposited in MycoBank. Cultures from ex-type strains and holotypes, which consisted of dry colonies on the most appropriate media for their sporulation, were also deposited at the Westerdijk Fungal Biodiversity Institute in Utrecht (CBS, The Netherlands).
In addition to the strains from marine sediments, we revived another amphichorda-like fungus from our fungal culture collection (FMR 17952). This strain was isolated from a fragment of a rubber tire floating in the seawater of the Miracle beach in July 2020. The tire fragment was washed three times with 10% NaClO (bleach) for 30 s, cut in small pieces and cultured on DRBC at 25 °C in darkness. We also examined several strains of A. felina, available in the CBS culture collection and labelled as Beauveria felina (i.e., CBS 110.08, CBS 250.34, CBS 312.50, CBS 648.66 and CBS 173.71), to study the morphological and molecular variability of this species and for epityfication purposes based on the lack of a type strain for this species. Furthermore, the ex-type strain and a reference strain of O. coprophila (i.e., CBS 247.82 and CBS 424.88) were also added to the study due to the above-mentioned reasons (Table 1).
2.2. Phenotypic analysis
Microscopic characterization was performed from strains growing on oatmeal agar (OA; 30 g oatmeal, 15 g agar, 1 L distilled water) after 14 days at 25 °C in darkness. Microscopic slides were mounted with lactic acid and observed with an Olympus BH-2 bright field microscope (Olympus Corporation, Tokyo, Japan). In species descriptions, size ranges of relevant structures derived from at least 30 measurements. Photomicrographs were obtained using a Zeiss Axio-Imager M1 light microscope (Zeiss, Oberkochen, Germany) with a DeltaPix Infinity digital camera. Due to the difficulties in observing the conidiogenic patterns in Amphichorda and Onychophora under light microscope, representative strains of species of both genera were examined under Scanning Electron Microscopy (SEM) using the Quanta 600 FEG Scanning Electron Microscope (Thermo Fisher Scientific, Waltham, Massachusetts, USA). The specimens were processed in accordance to Figueras & Guarro [19].
Macroscopic characterization of the colonies was made on PDA, OA and synthetic nutrient-poor agar (SNA; 1 g KH2PO4, 1 g KNO3, 0.5 g MgSO47·H2O, 0.5 g KCl, 0.2 g glucose, 0.2 g sucrose, 15 g agar, 1 L distilled water) after 14 days at 25 °C in darkness. Color notations in descriptions followed Kornerup and Wanscher [20]. Photoplates were assembled using GIMP v.2.10.34 (GNU Image Manipulation Program).
In addition, we assessed the ability of Amphichorda species to grow at different temperatures by culturing the strains on PDA from 5 to 40 °C, at intervals of 5 °C. Colony diameter was measured after 14 days in darkness. Moreover, the marine strains were compared with their phylogenetically related taxa to test a possible adaptation to the marine environment (salt tolerance). Cultures were carried out on solid agar plates of malt extract agar (MEA; 20 g malt extract, 15 g agar, 1 L distilled water) and MEA supplemented with 3.5% (35 ppt, the salt concentration in the marine environment), 5%, 10% and 15% NaCl. We used the strain Aspergillus chevalieri FMR 19829 (obtained from the marine sediments of this study) as a positive control of the ability to grow at high NaCl concentrations. Colony diameter was measured after 14 days (data not shown) and after 28 days at 25 °C. All tests were performed in duplicate, and the results represent the mean of the colony diameter between duplicate plates.
2.3. DNA extraction, PCR amplification and sequencing
Total genomic DNA was extracted through the modified protocol of Müller et al. [21] and quantified using Nanodrop 2000 (Thermo Scientific, Madrid, Spain). Four loci were used and amplified with the following primer pairs: ITS and LSU barcodes of the nrDNA with ITS5/LR5 [22,23], partial fragments of the BenA gene with T10/Bt2b [24] and the tef1 gene with EF-983F/EF-2218R [25], respectively. Briefly, PCR conditions for ITS, LSU, BenA and tef1 were set as follows: an initial denaturation at 95 °C for 5 min, followed by 35 cycles of 30 s at 95 °C, 45 s at 56 °C, and 1 min at 72 °C, and a final extension step at 72 °C for 10 min. PCR products were purified and sequenced at Macrogen Corp. Europe (Madrid, Spain) with the same primers used for amplification. Consensus sequences were assembled using SeqMan v. 7.0.0 (DNAStar Lasergene, Madison, Wisconsin, USA).
The preliminary species identification of the strains was performed by comparing their ITS regions with those available at the National Center for Biotechnology Information (NCBI) using the Basic Local Alignment Search Tool (BLAST; https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 6 October 2022). A maximum similarity level of ≥ 99% with ≥ 90% of sequence cover was used for species-level identification. Lower similarity values were considered as putative unknown fungi, and their taxonomic position was assessed with the loci mentioned above.
The sequences used for species-level identification were obtained from GenBank. In Table 1 are listed the strains of Amphichorda and Onychophora and their GenBank accession numbers. In Table S1 (supplementary material) is included the information about representative strains of Bionectriaceae, Cordycipitaceae and outgroups used in the phylogenetic analyses.
2.4. Phylogenetic analyses
The phylogenetic relationships of the strains examined in the present study were assessed using the ITS, LSU and tef1 regions. Sequences from each gene region were aligned individually in MEGA (Molecular Evolutionary Genetics Analysis) software v.6.0 [26], using the ClustalW algorithm [27] and, when necessary, refined with MUSCLE [28] or adjusted manually. Before combining the regions, the phylogenetic concordance between each individual phylogeny was tested through visual comparison to assess incongruent results among clades with high statistical support. When the concordance was confirmed, individual alignments were concatenated into a single data matrix. The partial fragments of the BenA gene were excluded from the phylogenetic analyses due to the limited availability of this region within the Bionectriaceae and most notably on taxa phylogenetically close to Amphichorda. However, BenA sequences were used to assess the similarity between Amphichorda species.
Maximum-likelihood (ML) and Bayesian analysis (BA) were used for phylogenetic inference of individual sequence alignments and the concatenated alignment (ITS-LSU and ITS-tef1). ML analyses were conducted using the CIPRES Science Gateway portal v. 3.3 (https://www.phylo.org/; [29]) and RAxML-HPC2 on XSEDE v. 8.2.12 [30] with default GTR substitution matrix and 1000 rapid bootstrap replications. Additional ML analyses were performed using IQ-TREE v. 2.1.2 [31,32] with ultrafast bootstrapping for the estimation of branch support [33]. The most suitable evolutionary model for each partition was estimated using ModelFinder [34,35], implemented in IQ-TREE. Bootstrap support (bs) ≥ 70 was considered significant [36]. Bayesian analyses were performed using MrBayes v. 3.2.6 [37]. The best substitution model for each locus was estimated using jModelTest v.2.1.3 following the Akaike criterion [38,39]. Markov Chain Monte Carlo sampling (MCMC) was performed for 10 million generations using four simultaneous chains (one cold chain and three heated chains) starting from a random tree topology. Trees were sampled every 1000 generation or until the run was stopped automatically when the average standard deviation of split frequencies fell below 0.01. The first 25 % of the trees were discarded as the burn-in phase of each analysis, and the remaining trees were used to calculate posterior probabilities (pp). A pp value of ≥ 0.95 was considered significant [40]. The resulting trees were plotted using FigTree v.1.3.1 (http://tree.bio.ed.ac.uk/software/figtree/, accessed on 5 December 2022). The DNA sequences generated in this study were deposited in GenBank (Table 1) and the alignments were submitted to Zenodo (https://doi.org/10.5281/zenodo.7937438).

Table 1.
GenBank accesions of the Amphichorda strains included in the present study.
| Species | Strain number | Substrate (country) | GenBank accesion numbers1 | Citation | |||
| ITS | LSU | tef1 | BenA | ||||
| A. cavernicola | CGMCC3.19571T | Bird faeces (China) | MK329056 | MK328961 | MK335997 | NA | [12] |
| LC 12560 | Animal faeces (China) | MK329061 | MK328966 | MK336002 | NA | [12] | |
| LC 12674 | Plant debris (China) | MK329065 | MK328970 | MK336006 | NA | [12] | |
| A. coprophila | CBS 247.82T (ex-type of O. coprophila) | Rabbit dung (England) | MH861494 | MH873238 | OQ954487 | pending | [41]; this study |
| CBS 424.88 (received as O. coprophila) | Chipmunk dung (Canada) | OQ942929 | OQ943166 | OQ954488 | pending | This study | |
| CBS 173.71 (received as B. felina) | Porcupine dung (Canada) | AY261368 | MH871833 | OQ954489 | pending | [41]; this study | |
| A. felina | CBS 250.34 (ex-type of I. cretacea) | Pressed yeast (England) | MH855498 | OQ943167 | OQ954490 | pending | [41]; this study |
| CBS 648.66 (received as B. felina) | Unknown, (Argentina) | OQ942930 | MH870575 | OQ954491 | pending | [41]; this study | |
| CBS 110.08 (received as B. felina) | Unknown | MH854578 | OQ943168 | OQ954492 | pending | [41]; this study | |
| A. guana | CGMCC3.17908T | Bat guano (China) | KU746665 | KU746711 | KX855211 | NA | [11] |
| CGMCC3.17909 | Bat guano (China) | KU746666 | KU746712 | KX855212 | NA | [11] | |
| CBS 312.50 (received as B. felina) | Rabbit dung (Unknown) | MH856641 | MH868150 | OQ954493 | pending | [41]; this study | |
| A. littoralis | FMR 17952 | Floating rubber tire (Spain) | OQ942925 | OQ943162 | OQ954483 | pending | This study |
| FMR 19404T | Marine sediment (Spain) | OQ942924 | OQ943161 | OQ954482 | pending | This study | |
| FMR 19611 | Marine sediment (Spain) | OQ942926 | OQ943163 | OQ954484 | pending | This study | |
| FMR 20067 | Marine sediment (Spain) | OQ942927 | OQ943164 | OQ954485 | pending | This study | |
| FMR 20149 | Marine sediment (Spain) | OQ942928 | OQ943165 | OQ954486 | pending | This study | |
CBS: Culture Collection of the Westerdijk Fungal Biodiversity Institute, Utrecht, the Netherlands; CGMCC: China General Microbiological Culture Collection Center, China; FMR: Facultat de Medicina i Ciències de la Salut, Reus, Spain. T indicates ex-type strains. “NA” indicates sequences not used in this study. 1ITS: Internal transcribed spacer region of the rDNA and 5.8S gene; LSU: 28S large ribosomal subunit; tef 1: translation elongation factor 1α; BenA: tubulin. Sequences generated in this study are highlighted in bold.
3. Results
Among the fungi detected from marine sediments collected at different depths, we recovered four strains (FMR 19404, FMR 19611, FMR 20067 and FMR 20149) exclusively from samples collected at 20 m of depth in both the Miracle and Arrabassada beaches, using SWMEA3% and DRBC culture media. These strains and FMR 17952, the latter isolated from a rubber tire floating in seawater, were morphologically identified as Amphichorda sp. However, although they showed the typical morphological features of the genus (i.e. synnematous and mononematous conidiophores, flask-shaped with a strongly bent neck conidiogeous cells and solitary conidia that remain attached to the apex of the conidiogenous cell), they exhibited some morphological traits that did not exactly fit into any of the accepted species of Amphichorda.
3.1. Phylogeny
The molecular identification based on the BLAST search of our five unidentified strains revealed a high percentage of similarity with species of the genus Amphichorda using ITS sequences. Specifically, the percentage of identity was of 98% with A. cavernicola (CGMCC 3.19571) and between 95-97% with other species of this genus. The molecular comparison using the LSU region revealed a 99% of similarity to O. coprophila (CBS 247.82), A. cavernicola (CGMCC 3.19571) and between 98-99% of similarity with other species of the genus Amphichorda. Other taxa closely related to our strains with a 97% of identity with this locus were Nigrosabulum globosum (CBS 512.70), Acremonium (Ac.) curvum (GZUIFR 22.035) and Ac. alternatum (CBS 407.66). Members of the genera Beauveria and Cordyceps did not match with our sequences in the BLAST results. Despite this, we included them in the phylogenetic analysis due to the traditional placement of Amphichorda as a member of Cordycipitaceae. In addition, sequence analyses of these two gene markers allowed us to confirm or reidentify the CBS reference strains of A. felina as following: CBS 110.08 and CBS 648.66 as A. felina, CBS 312.50 as A. guana, and CBS 173.71 as O. coprophila.
Based on the BLAST results, we assessed the phylogenetic relationships among genera phylogenetically related to Amphichorda with the ITS and LSU regions. The resulting tree topologies from the individual analyses of these two gene markers were similar and did not show incongruences. Therefore, both alignments were concatenated into a single matrix. The final alignment of the concatenated ITS and LSU regions comprised 65 taxa that included two representative strains from each Amphichorda species, as well as, two representative strains from O. coprophila and the strains recovered from the marine environment to prevent branch imbalance; together with representative species belonging to the families Bionectriaceae and Cordycipitaceae. The tree was rooted with Pochonia chlamydospora (CBS 504.66) and Metapochonia suchlasporia (CBS 251.83) as outgroup. The total length comprised 1435 characters including gaps (ITS: 624, LSU: 811 characters). Among these, 906 characters were conserved sites (ITS: 255, LSU: 651), 529 characters were variable sites (ITS: 369, LSU: 160) and 417 characters were parsimony informative (ITS: 290, LSU: 127). For the ML analyses, the best fit models were TIM2+F+I+G4 for the ITS region and TIM2e+I+G4 for the LSU region. For the BI analysis, the best fit models were GTR+I+G for both the ITS and LSU region. Here, we represented the Maximum Likelihood (RAxML) tree with the bootstraps support values of the ML analyses (RAxML and IQ-TREE) and Bayesian posterior probabilities at the nodes. The resulting phylogenetic tree resolved the genus Amphichorda as a monophyletic linage within the family Bionectriaceae (Figure 1), being closely related with a well-supported clade that comprised two accepted genera in the family, Nigrosabulum and Hapsidospora, together with a recently described Acremonium species, Ac. curvum [42]. This latter species was, however, placed very distantly from the genus Acremonium s. str. The concatenated analysis defined five terminal clades within Amphichorda, where two marine strains (FMR 19404 and FMR 17952), representatives of our unidentified Amphichorda species, and those strains of O. coprophila (CBS 173.71 and CBS 247.82) represented two independent Amphichorda linages. However, these molecular markers lacked resolution to determine the phylogenetic relationships among Amphichorda species. Therefore, we performed a phylogenetic analysis combining the ITS and LSU regions and the elongation factor (tef1) gene in order to delineate Amphichorda species with precision.
The individual ITS, LSU and tef1 alignments were concatenated into a single matrix, because the resulting individual trees represented similar topologies. The final ITS, LSU and tef1 alignment comprised the five unidentified Amphichorda strains, nine strains representatives of the known Amphichorda species, and three strains identified as O. coprophila. Acremonium curvum (GZUIFR 22.035), Ac. globosisporium (GZUIFR 22.036) and Ac. sclerotigenum (A101) were used as outgroup. The total length comprised 2168 characters including gaps (ITS: 506, LSU: 779, tef1: 883 characters). Among these, 1865 characters were conserved sites (ITS: 393, LSU: 727, tef1: 745), 303 characters were variable sites (ITS: 113, LSU: 52, tef1: 138), and 166 characters were parsimony informative (ITS: 55, LSU: 30, tef1: 81). For the ML analyses, the best fit models were TNe+G4 for the ITS region, TNe+I for the LSU region and TN+F+G4 for the tef1 region. For the BI analysis the best fit models were K80+I for both the ITS and LSU regions and GTR+G for the tef1 region. Here, we represented the Maximum Likelihood (RAxML) tree with the bootstraps support values of the ML analyses (RAxML and IQ-TREE) and Bayesian posterior probabilities at the nodes. The resulting phylogenetic tree resolved the three species currently accepted in Amphichorda (A. cavernicola, A. felina and A. guana) as independent linages (Figure 2). The five marine strains delineated an undescribed linage within Amphichorda, closely related to the clade representative of O. coprophila. The marine strains are proposed below as Amphichorda littoralis and O. coprophila is accepted as an Amphichorda species. A detailed morphological characterization of the novel fungi is provided in the taxonomy section.
3.2. Morphological analysis
In order to perform a morphological comparison of our strains, we reviewed the existing literature on Amphichorda and examined living cultures of the following species: A. coprophila (CBS 247.82, CBS 424.88 and CBS 173.71), A. felina (CBS 250.34, CBS 110.08 and CBS 648.66) and A. guana (CBS 312.50). Unfortunately, strains of A. cavernicola were not available for comparison. The colony color displayed across PDA, OA and SNA culture media represented the most accurate character to distinguish species. Microscopically, the reproductive structures were almost similar between species and consisting in flask-shaped conidiogenous cells that produced subglobose and smooth conidia. Although, there were subtle differences on the size range of these structures, they overlapped between species. In Table 2 we provide a synopsis of the key morphological characters that allow discrimination among species of Amphichorda.
However, the pattern of conidiogenesis could not be properly determined under light microscopy. Therefore, we selected representative strains of A. coprophila, A. felina, A. guana and A. littoralis for examination under SEM (Figure 3).
Almost all strains of Amphichorda species mentioned above showed a holoblastic pattern of conidiogenesis, producing few conidia in an apparent sympodial proliferation. However, the strains of A. coprophila differed in that, while the ex-type strain CBS 247.82 showed exclusively holoblastic conidiogenesis (Figure 3D), CBS 424.88 exhibited both holoblastic (Figure 3F) and enteroblastic (phialidic) conidiogenous cells even showing a small collaret in the apex (Figure 3E). Another difference observed in these latter strains was the presence of both smooth or roughened conidiogenous cells. In the rest of the species examined the conidiogenous cells were smooth.
3.3. Salt tolerance test
Considering the marine origin of A. littoralis, we compared its ability to grow under different NaCl concentrations with other species of the genus, predominantly isolated from terrestrial environments, to ascertain a possible preference for the marine environment. Although, all Amphichorda strains managed to grow up to 10% NaCl, each species showed different colony diameters across media. The colony diameter of A. coprophila, A. felina and A. guana decreased in an inverse proportion to the addition of NaCl. The only exception was the strain CBS 247.82 of A. coprophila, which growth was restricted and nearly similar across media with different salt concentrations. Amphichorda littoralis achieved similar maximum colony diameters across MEA and MEA supplemented with 3.5% and 5% NaCl. In particular, the strains FMR 19404, FMR 19611 and FMR 20067 reached the maximum colony diameter in MEA supplemented with 3.5% and 5% NaCl (Figure 4).
3.4. Taxonomy
Amphichorda Fries, Systema Orbis vegetalis 1:170 (1825)
= Onychophora W. Gams, P.J. Fisher & J. Webster, Transactions of the British Mycological Society 82 (1): 174 (1984)
Type species. Amphichorda felina (DC) Fries, Systema Orbis vegetalis 1: 170 (1825).
For synonyms of the species see Mycoblank database (https://www.mycobank.org/).
Emended description
Asexual morph with conidiophores synnematous or mononematous, semi-macronematous, erect, straight or flexuous, bearing lateral or terminal conidiogenous cells, arranged single or in whorls, sometimes micronematous and reduced to conidiogenous cells growing directly from vegetative hyphae. Conidiogenous cells flask-shaped, usually with a strongly bent neck, holoblastic, rarely enteroblastic, phialidic, hyaline, smooth-walled or roughened. Conidia solitary, often remaining attached to the apex of the conidiogenous cell, subglobose, hyaline, smooth-walled. Sexual morph not observed.
Amphichorda coprophila (W. Gams, P.J. Fisher & J. Webster) Guerra-Mateo, Cano & Gené, comb. nov.
Mycobank: MB848789.
Basionym. Onychophora coprophila W. Gams, P.J. Fisher & J. Webster, Transactions of the British Mycological Society 82 (1): 174 (1984)
Type. ENGLAND, Devon, Dawlish Warren, from rabbit dung incubated at relative humidity of 95% for several weeks, Dec. 1981, J. Webster (holotype CBS H-1740 = IMI 275663, ex-type culture CBS 247.82).
Asexual morph described in Gams et al. [15].
Culture characteristics (after 14 days at 25 °C). Colonies on PDA attaining 22-24 mm diam., slightly raised, irregularly sulcated, glabrous and brownish orange (7C5) at center (CBS 424.88 and CBS 173.71 orange (5B5)), velvety and white at periphery, margin crenate; reverse brownish orange at center and white at periphery; diffusible pigment greyish orange (6B5). On OA, colonies reaching 34–40 mm diam., flat, velvety, pale yellow (4A3) at center to white at periphery, margin entire and slightly lobated; reverse pale yellow. On SNA, colonies reaching 5–10 mm diam., glabrous, pale yellow, margin slightly lobated; reverse pale yellow.
Additional specimens examined. CANADA, Ontario, Landmark County, along Clyde River, from chipmunk dung, K.A. Seifert (CBS 424.88); ibid., Stoneleigh, from porcupine dung, Sep. 1969, R.F. Cain and D.W. Malloch (CBS 173.71).
Notes. Amphichorda coprophila is a well-supported species that represents a distant independent linage in the genus Amphichorda (Figs 1, 2). It can be morphologically distinguished by its orange to brownish orange colonies on PDA (Table 2), the production of conidia through both holoblastic and phialidic conidiogenous cells (Figure 3 D-F) and the occasional rough ornamentation of the conidiogenous cells under SEM.
Amphichorda littoralis Guerra-Mateo, Torres-Garcia, Cano & Gené, sp. nov. Figure 5.
Mycobank: MB 848035.
Etymology. Name refers to the area where this species was isolated, Mediterranean coast (Tarragona, Spain).
Type. SPAIN, Catalonia, Mediterranean coast, Tarragona, Platja del Miracle, N 41º6´19´´, E 1º15´37´´, from sediments at 20 m of depth, Jun. 2021, G. Quiroga-Jofre and D. Guerra-Mateo (holotype CBS H-25254, ex-type culture FMR 19404, CBS 149935).
Asexual morph on OA. Mycelium composed of smooth-walled, branched, septate, hyaline, 1–1.5 µm wide hyphae. Conidiophores monomematous, rarely synnematous, arising directly from superficial mycelium, micronematous and reduced to conidiogenous cells growing directly or on a short lateral protrusion from vegetative hyphae, or semi-macronematous, erect, straight or flexuous, commonly unbranched, bearing lateral or terminal conidiogenous cells, arranged single or in whorls of 2–4, hyaline and smooth-walled; synnematous conidiophores only observed in FMR 20067 on PDA at the margin of the colony, yellowish white, cylindrical with tomentose apex. Conidiogenous cells flask-shaped, usually with a strongly bent neck, 6–10(–11.5) × 1.5–2 µm, hyaline, smooth-walled. Conidia solitary, often remaining attached to the apex of the conidiogenous cell, subglobose, 3–4 x 2.5–3 µm, hyaline, smooth-walled. Sexual morph not observed.
Culture characteristics (after 14 days at 25 °C). Colonies on PDA attaining 20 mm diam., slightly raised, irregularly sulcated, glabrous and greenish yellow (1A7) at center, velvety (fasciculate in FMR 20067) and white at periphery, margin crenate; reverse greenish yellow (1A7) at center and white at periphery; diffusible pigment light yellow (4A4) produced after 21 days. On OA, colonies reaching 30–32 mm diam., flat, velvety, greenish yellow at center to greyish yellow at periphery, margin entire and slightly lobated; reverse greenish yellow (1A7). On SNA, colonies reaching 9–14 mm diam., glabrous, greenish yellow, margin slightly lobated; reverse greenish yellow (1A7).
Cardinal temperatures for growth. Minimum at 10 °C, optimum at 25 °C, maximum at 30 °C.
Additional specimens examined. SPAIN, Catalonia, Mediterranean coast, Tarragona, Platja del Miracle, N 41°6´19´´, E 1°15´37´´, from sediments at 20 m of depth, Oct. 2021, G. Quiroga-Jofre and D. Guerra-Mateo (FMR 19611); ibid., Platja de la Arrabassada, N 41°6´53´´, E 1°16´48´´, from sediments at 20 m of depth, Jun. 2022, G. Quiroga-Jofre and D. Guerra-Mateo (FMR 20149); ibid., from sediments at 20 m of depth, Jun. 2022, G. Quiroga-Jofre and D. Guerra-Mateo (FMR 20067); ibid., Mediterranean coast, Tarragona, from a fragment of floating rubber tire, Jul. 2020, D. Torres-García (FMR 17952).
Notes. Amphichorda littoralis is phylogenetically related to A. coprophila (Figs 1, 2). Macroscopically, they can be distinguished by the color of the colony (Table 2). In the novel species, colonies are consistently greenish yellow across PDA, OA and SNA, while in A. coprophila, colony color ranges from brown orange to pale yellow. Microscopically, the conidiogenous cells of A. littoralis are consistently smooth, while A. coprophila can show a rough ornamentation. Moreover, the phylogenetic distance between this novel species and other members of Amphichorda is around 96% for the tef1 region and 95% for the BenA region.
4. Discussion
In previous morphological and phylogenetic studies, the taxonomic circumscription of the genus Amphichorda has been controversial and its phylogenetic relationships with other taxa have remained obscure. The most recent multi-locus phylogenetic tree assessing the diversity within Cordycipitaceae, included the type strain of A. guana, analysed its LSU and tef1 sequences and resolved Amphichorda as a distant independent linage sister to Cordycipitaceae [4]. Our phylogenetic tree combining the ITS and LSU regions determined the close phylogenetic relationships of Amphichorda with the genera Hapsidospora and Nigrosabulum (Figure 1), two accepted members of the family Bionectriaceae. These results allow to recognize Amphichorda as a member of this family, despite it is the only representative genus with members producing conidia holoblastically. In the most recent review of the family 41 genera were accepted, composed exclusively of fungi showing phialidic conidiogenous cells [2]. However, the order Hypocreales comprises members with asexual morphs producing both enteroblastic (phialidic) and holoblastic conidiogenous cells. Although most of the families accepted in the order, such as Clavicipitaceae, Ijuhyaceae, Myrotheciomycetaceae, Nectriaceae, Niessliaceae, Ophiocordycipitaceae, Sarocladiaceae, Stachybotriaceae, Stromatonectriaceae, Tilachlidiaceae and Xanthonectriaceae only show phialidic conidiogenesis, other families like Calcarisporiaceae, Cordycipitaceae and Hypocreaceae show both types of conidiogenesis[1,2,4,43,44,45,46,47]. Only genera like Beauveria and Calcarisporium exhibit holoblastic conidiogenous cells [2,48]. Thus, it seems that phialides represent the ancestral way of asexual reproduction in Hypocreales. In this sense, blastic conidiogenesis would have appeared independently as a secondary trait across different families. In particular, Amphichorda seems to represent a transition between both types of conidiogenesis for the Bionectriaceae. The type species of the genus, A. felina, and the rest of the species accepted show holoblastic conidiogenesis. The exception is A. coprophila, which can produce both types of condiogenous cells depending on the strain studied, but in particular in the strain CBS 424.88. Here, we propose this species as a novel combination of the genus Amphichorda. However, as mentioned before, the original authors of the species already suggested the possible relation of this species with Acremonium (Bionectriaceae) [15]. They observed the conidiogenous cells of the species and concluded a phialidic conidiogenous pattern. Based on the close phylogenetic relationship of A. coprophila with holoblastic species, we can conclude with confidence that it can produce both types of conidiogenous cells. This trait, although odd, has already been described in Bionectriaceae with the holoblastic mesoconidia described on some Fusarium species [49,50].
Amphichorda represents a group of morphologically cryptic species. The microscopic reproductive structures show subtle variations on their size range (Table 2). Thus, the best morphological character to distinguish species is the colony color across different culture media. We have found this trait consistent across several strains on PDA, OA and SNA. However, colony color may be of little use in culture media like MEA (Figure 4). For this reason, phylogenetic analyses represent the most accurate way to identify Amphichorda species. In particular, the ITS region, the fungal barcode, is able to distinguish species of Amphichorda with precision, but other structural genes like tef1 and BenA can be used as secondary barcodes with similar results.
Correct identification of species based on phylogenetic analyses needs for DNA sequences obtained from type strains. This is frequently a limitation when working with fungi described before the development of DNA sequencing techniques. In our particular case, the type material of A. felina seems to be lost. We therefore selected representative strains identified as A. felina from the CBS culture collection, including some of coprophilous origin like the protologue of this species, in order to determine a suitable candidate for epitypification and study the morphological and genetic variability of the species. However, the former goal was not feasible because the strains of coprophilous origin represented A. guana and A. coprophila (Table 1), and the strains that phylogenetically matched with A. felina did not correspond with the origin of the protologue, preventing the epitypification [14]. Despite this, the strain CBS 250.34 fits the morphological description of A. felina and it has been extensively used to characterize the species in phylogenetic analyses. Therefore, we accept the strain CBS 250.34 as reference to stabilize the nomenclature of A. felina and, consequently, the genus Amphichorda, but its representation as type strain should be avoided.
Finally, we propose the novel species A. littoralis, the first species of the genus described from the marine environment. Marine fungi are described as those that can grow and sporulate under marine-like conditions [51]. All the strains that represent this species have been isolated from the marine habitat and all strains managed to grow and sporulate up to MEA 10% NaCl. We understand this as an indicative for a preference for the marine environment and consider this species may represent a marine fungus. This species updates the ecological range of the genus. Amphichorda was previously recognized as a group of parasitic and coprophilous fungi. Although, a pathogenic behavior has been described for A. felina [52], we conclude that it is predominantly composed of saprotrophic fungi with preference for substrates with abundant organic matter like dung or marine sediments.
5. Concluding remarks
Our phylogenetic analyses combining the ITS and LSU regions revealed the genus Amphichorda as a member of the family Bionectriaceae, where it represents the only holoblastic group. The combination of morphological and phylogenetic analyses determined our marine strains as a novel species, A. littoralis, and resolved the taxonomic position of O. coprophila as a new member of the genus Amphichorda. The current study is the largest sampling of Amphichorda ever subjected to multi-locus sequence analyses, provides a comprehensive phylogenetic backbone and represents a framework for future studies on the genus.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: GenBank accessions of representative taxa from Bionectriaceae, Cordycipitaceae and outgroups included in the phylogenetic analyses.
Author Contributions
Conceptualization, D.G.-M., J.F.C.-L. and J.G.; methodology, D.G.-M., J.G. and J.F.C.-L.; software, D.G.-M. and J.F.C.-L.; validation, J.F.C.-L. and J.G.; formal analysis, D.G.-M., J.F.C.-L and J.G.; investigation, D.G.-M., J.F.C.-L and J.G.; resources, V.B., J.F.C.-L and J.G.; data curation, J.F.C.-L and J.G.; writing original draft preparation, D.G.-M. and J.G.; writing review and editing, D.G.-M., J.F.C.-L. and J.G.; visualization, J.F.C.-L and J.G.; supervision, J.F.C.-L and J.G.; project administration, V.B. and J.G.; funding acquisition, J.G. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the grant PID2021-128068NB-100 funded by MCIN/AEI/10. 13039/501100011033/ and by “ERDF A way of making Europe”.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank to the CBS culture collection (The Netherlands) and its curators for providing some fungal strain included in the study and to Gabriel Quiroga-Jofre (Tarraco Diving Center) for the services in the collection of the samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined Families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Perera, R.H.; Hyde, K.D.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Bundhun, D.; Camporesi, E.; Akulov, A.; Liu, J.K.; Liu, Z.Y. Profile of Bionectriaceae, Calcarisporiaceae, Hypocreaceae, Nectriaceae, Tilachlidiaceae, Ijuhyaceae fam. nov., Stromatonectriaceae fam. nov. and Xanthonectriaceae fam. nov. Fungal Divers. 2023, 118, 95–271. [Google Scholar] [CrossRef]
- Kepler, R.M.; Luangsa-ard, J.J.; Hywel-Jones, N.L.; Quandt, C.A.; Sung, G.H.; Rehner, S.A.; Aime, M.C.; Henkel, T.W.; Sanjuan, T.; Zare, R.; et al. A Phylogenetically-Based Nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 2017, 8, 335–353. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.B.; Wang, Y.; Fan, Q.; Duan, D.E.; Zhang, G.-D.; Dai, R.-Q.; Dai, Y.-D.; Zeng, W.B.; Chen, Z.-H.; Li, D.D.; et al. Multigene Phylogeny of the Family Cordycipitaceae (Hypocreales): New Taxa and the New Systematic Position of the Chinese Cordycipitoid Fungus Paecilomyces hepiali. Fungal Divers. 2020, 103, 1–46. [Google Scholar] [CrossRef]
- Lamarck, J.B. De Flore françoise ou descriptions succinctes de toutes les plantes qui croissent naturellement en France, disposées selon une nouvelle méthode d’analyse, et précédées par un exposé des principes élémentaires de la botanique; Paris, 1815; Volume 6. [Google Scholar]
- Fries, E. Systema orbis vegetabilis: Primas lineas novae constructionis periclitatur; e Typographia academica: Lunde, 1825. [Google Scholar]
- Fries, E.M. Systema mycologicum: sistens fungorum ordines, genera et species, huc usque cognitas, quas ad normam methodi naturalis determinavit; Sumtibus Ernesti Mauritii; 1832; Volume 3. [Google Scholar]
- Hodge, K.T.; Gams, W.; Samson, R.A.; Korf, R.P.; Seifert, K.A. Lectotypification and Status of Isaria Pers. : Fr. Taxon 2005, 54, 485–489. [Google Scholar] [CrossRef]
- De Hoog, G.S. The Genera Beauveria, Isaria, Tritirachium and Acrodontium gen. nov. Stud. in Mycol. 1972, 1, 1–41. [Google Scholar]
- Clements, F.F.; Shear, C.L. The Genera of Fungi.; The H. W. Wilson Company: New York, 1931. [Google Scholar]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable Mycobiota from Karst Caves in China, with Descriptions of 20 New Species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-F.; Zhou, S.-Y.; Eurwilaichitr, L.; Ingsriswang, S.; Raza, M.; Chen, Q.; Zhao, P.; Liu, F.; Cai, L. Culturable Mycobiota from Karst Caves in China II, with Descriptions of 33 New Species. Fungal Divers. 2021, 106, 29–136. [Google Scholar] [CrossRef]
- Beyma, F.H. van. Beschreibung Einiger Neuer Pilzarten Aus Dem Centraalbureau Voor Schimmelcultures Baarn (Holland). Zentralblatt für Bakteriologie und Parasitenkunde 1935, 2, 345–355. [Google Scholar]
- Lendemer, J.C. Epitypes Are Forever: Best Practices for an Increasingly Misused Nomenclatural Action. Taxon 2020, 69, 849–850. [Google Scholar] [CrossRef]
- Gams, W.; Fisher, P.J.; Webster, J. Onychophora, a New Genus of Phialidic Hyphomycetes from Dung. TBMS 1984, 82, 174–177. [Google Scholar] [CrossRef]
- Expósito, N.; Rovira, J.; Sierra, J.; Folch, J.; Schuhmacher, M. Microplastics Levels, Size, Morphology and Composition in Marine Water, Sediments and Sand Beaches. Case Study of Tarragona Coast (Western Mediterranean). Sci. Total Environ. 2021, 786, 147453. [Google Scholar] [CrossRef] [PubMed]
- Kossuga, M.H.; Romminger, S.; Xavier, C.; Milanetto, M.C.; Valle, M.Z. do; Pimenta, E.F.; Morais, R.P.; Carvalho, E. de; Mizuno, C.M.; Coradello, L.F.C.; et al. Evaluating Methods for the Isolation of Marine-Derived Fungal Strains and Production of Bioactive Secondary Metabolites. Rev. Bras. Farmacogn. 2012, 22, 257–267. [Google Scholar] [CrossRef]
- Wingfield, B.D.; Wingfield, M.J.; Duong, T.A. Molecular Basis of Cycloheximide Resistance in the Ophiostomatales Revealed. Curr. Genet. 2022, 68, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Figueras, M.J.; Guarro, J. A Scanning Electron Microscopic Study of Ascoma Development in Chaetomium malaysiense. Mycol. 1988, 80, 298–306. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Methuen: London, UK, 1978. [Google Scholar]
- Müller, F.M.C.; Werner, K.E.; Kasai, M.; Francesconi, A.; Chanock, S.J.; Walsh, T.J. Rapid Extraction of Genomic DNA from Medically Important Yeasts and Filamentous Fungi by High-Speed Cell Disruption. J. Clin. Microbiol. 1998, 36, 1625–1629. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid Genetic Identification and Mapping of Enzymatically Amplified Ribosomal DNA from Several Cryptococcus Species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; New York, 1990; pp. 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of Primer Sets Designed for Use with the PCR to Amplify Conserved Genes from Filamentous Ascomycetes. AEM 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria Phylogeny Inferred from Nuclear ITS and EF1-α Sequences: Evidence for Cryptic Diversification and Links to Cordyceps Teleomorphs. Mycol. 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES Science Gateway: Enabling High-Impact Science for Phylogenetics Researchers with Limited Resources. In Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the eXtreme to the Campus and Beyond, Chicago, IL, USA, 16–20 July 2012; Association for Computing Machinery: New York, NY, USA, 2012; pp. 1–8. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace Aware Data Structure for Phylogenomic Inference from Supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Hillis, D.M.; Bull, J.J. An Empirical Test of Bootstrapping as a Method for Assessing Confidence in Phylogenetic Analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat Methods 2012, 9, 772–772. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Hespanhol, L.; Vallio, C.S.; Costa, L.M.; Saragiotto, B.T. Understanding and Interpreting Confidence and Credible Intervals around Effect Estimates. BJPT 2019, 23, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-Scale Generation and Analysis of Filamentous Fungal DNA Barcodes Boosts Coverage for Kingdom Fungi and Reveals Thresholds for Fungal Species and Higher Taxon Delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Z.-Y.; Ren, Y.-L.; Chen, W.-H.; Liang, J.-D.; Pan, J.-M.; Huang, J.-Z.; Liang, Z.-Q.; Han, Y.-F. Morphological characteristics and phylogenetic evidence reveal two new species of Acremonium (Hypocreales, Sordariomycetes). MycoKeys 2022, 91, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Bao, D.-F.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Perera, R.H.; Thiyagaraja, V.; Hongsanan, S.; Wanasinghe, D.N.; Shen, H.-W.; Tian, X.; Yang, L.-Q.; et al. Taxonomy, Phylogeny and Evolution of Freshwater Hypocreomycetidae (Sordariomycetes), In Review. 2023.
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.St.J.; Gené, J.; Guarro, J.; Baseia, I.G.; García, D.; Gusmão, L.F.P.; Souza-Motta, C.M.; et al. Fungal Planet Description Sheets: 716–784. Persoonia 2018, 40, 240–393. [Google Scholar] [CrossRef]
- Lombard, L.; Houbraken, J.; Decock, C.; Samson, R.A.; Meijer, M.; Réblová, M.; Groenewald, J.Z.; Crous, P.W. Generic Hyper-Diversity in Stachybotriaceae. Persoonia 2016, 36, 156–246. [Google Scholar] [CrossRef]
- Huang, S.-K.; Hyde, K.; Maharachchikumbura, S.; Mckenzie, E.; Wen, T.-C. Taxonomic Studies of Coronophorales and Niessliaceae (Hypocreomycetidae). Mycosphere 2021, 12, 875–992. [Google Scholar] [CrossRef]
- Quandt, C.A.; Kepler, R.M.; Gams, W.; Araújo, J.P.M.; Ban, S.; Evans, H.C.; Hughes, D.; Humber, R.; Hywel-Jones, N.; Li, Z.; et al. Phylogenetic-Based Nomenclatural Proposals for Ophiocordycipitaceae (Hypocreales) with New Combinations in Tolypocladium. IMA Fungus 2014, 5, 121–134. [Google Scholar] [CrossRef]
- Rehner, S.A.; Minnis, A.M.; Sung, G.-H.; Luangsa-ard, J.J.; Devotto, L.; Humber, R.A. Phylogeny and Systematics of the Anamorphic, Entomopathogenic Genus Beauveria. Mycol. 2011, 103, 1055–1073. [Google Scholar] [CrossRef]
- Pascoe, I.G. Fusarium Morphology. I. Identification and Characterization of a Third Conidial Type, the Mesoconidium. Mycotaxon 1990, 37, 121–160. [Google Scholar]
- De Hoog, G.S.; Guarro, J.; Gené, J.; Ahmed, S.; Al-Hatmi, A.M.S.; Figueras, M.J.; Vitale, R.G. Atlas of Clinical Fungi: The Ultimate Benchtool for Diagnostics. Introductions, Lower Fungi, Basidiomicetes, Yeasts, Filamentous Ascomycetes, 4th ed.; Foundation Atlas: Hilversum, The Netherlands, 2020. [Google Scholar]
- Gonçalves, M.F.M.; Esteves, A.C.; Alves, A. Marine Fungi: Opportunities and Challenges. Encyclopedia 2022, 2, 559–577. [Google Scholar] [CrossRef]
- Ramanujam, B.; Poornesha, B.; Kandan, A.; Mohan, M.; Sivakumar, G. Natural Occurrence of Entomopathogenic Fungus Beauveria Felina (DC.) J.W. Carmich on Fall Armyworm, Spodoptera frugiperda (J. E. Smith). J. Entomol. Zool. Stud. 2021, 9, 140–143. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree inferred from a Maximum Likelihood (RAxML) analysis based on a concatenated alignment of ITS and LSU sequences of 65 strains representing Bionectriaceae, Cordycipitaceae and outgroups. Numbers at the branches indicate support values (RAxML-BS / IQ-TREE-BS / BI-PP) above 70 % / 90 % / 0.95. The genus Amphichorda is printed in bold and the other strains are collapsed based on their genera. Bold branches indicate full support values (100/100/1). T indicates ex-type strains. The tree is rooted to Metapochonia suchlasporia CBS 251.83 and Pochonia chlamydosporia CBS 504.66. Quote marks indicate strains with unresolved taxonomy. The scale bar represents the expected number of changes per site.
Figure 1.
Phylogenetic tree inferred from a Maximum Likelihood (RAxML) analysis based on a concatenated alignment of ITS and LSU sequences of 65 strains representing Bionectriaceae, Cordycipitaceae and outgroups. Numbers at the branches indicate support values (RAxML-BS / IQ-TREE-BS / BI-PP) above 70 % / 90 % / 0.95. The genus Amphichorda is printed in bold and the other strains are collapsed based on their genera. Bold branches indicate full support values (100/100/1). T indicates ex-type strains. The tree is rooted to Metapochonia suchlasporia CBS 251.83 and Pochonia chlamydosporia CBS 504.66. Quote marks indicate strains with unresolved taxonomy. The scale bar represents the expected number of changes per site.
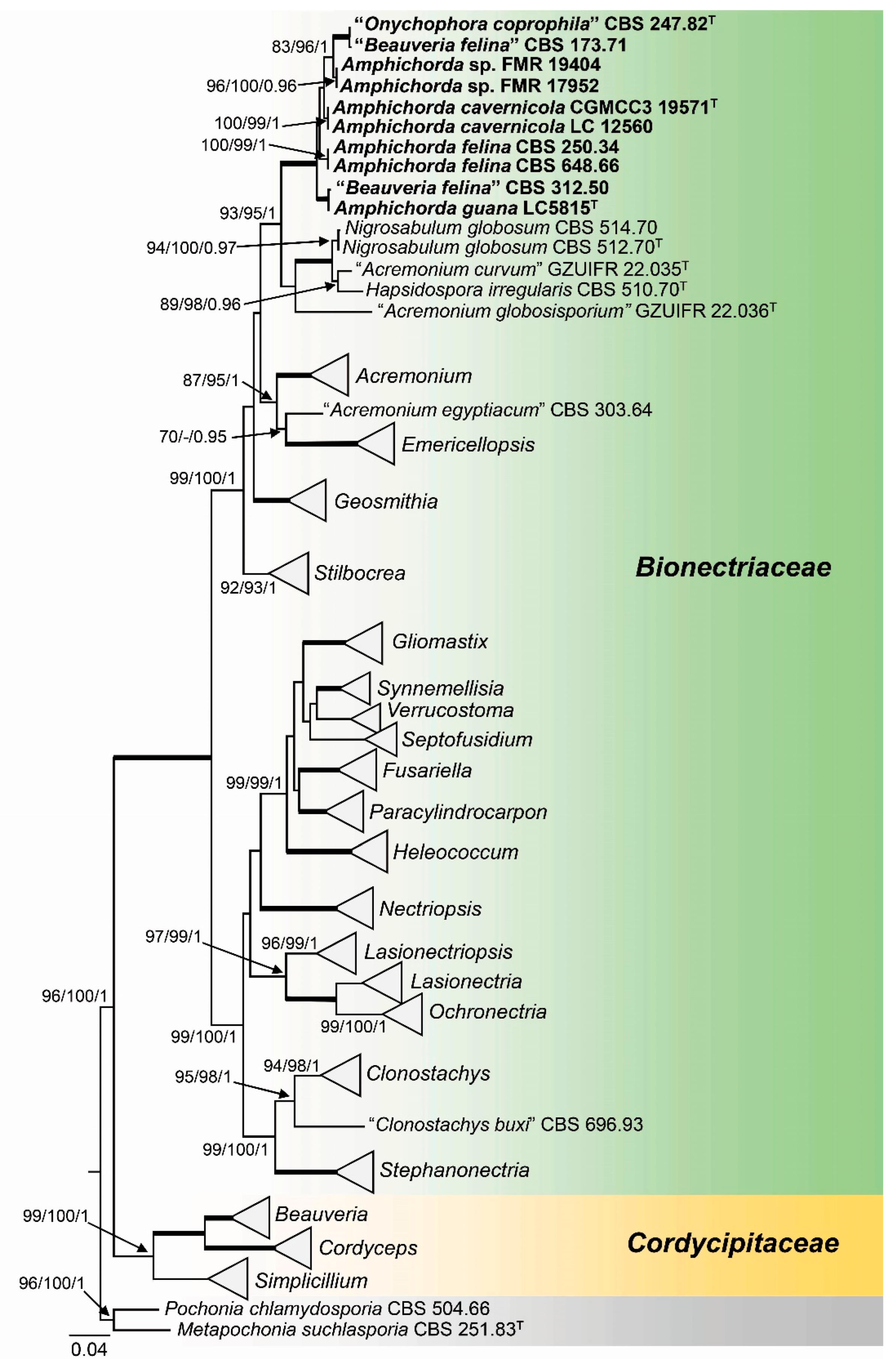
Figure 2.
Phylogenetic tree inferred from a Maximum Likelihood (RAxML) analysis based on a concatenated alignment of ITS, LSU and tef1 sequences of 20 strains representing Amphichorda and outgroups. Numbers at the branches indicate support values (RAxML-BS / IQ-TREE-BS / BI-PP) above 70 % / 90 % / 0.95. Bold branches indicate full support values (100/100/1). The novel species and combination are printed in bold. T indicates ex-type strains. The tree is rooted to Acremonium curvum (GZUIFR 22.035), Acremonium globosisporium GZUIFR 22.036 and Acremonium sclerotigenum A101. The scale bar represents the expected number of changes per site.
Figure 2.
Phylogenetic tree inferred from a Maximum Likelihood (RAxML) analysis based on a concatenated alignment of ITS, LSU and tef1 sequences of 20 strains representing Amphichorda and outgroups. Numbers at the branches indicate support values (RAxML-BS / IQ-TREE-BS / BI-PP) above 70 % / 90 % / 0.95. Bold branches indicate full support values (100/100/1). The novel species and combination are printed in bold. T indicates ex-type strains. The tree is rooted to Acremonium curvum (GZUIFR 22.035), Acremonium globosisporium GZUIFR 22.036 and Acremonium sclerotigenum A101. The scale bar represents the expected number of changes per site.
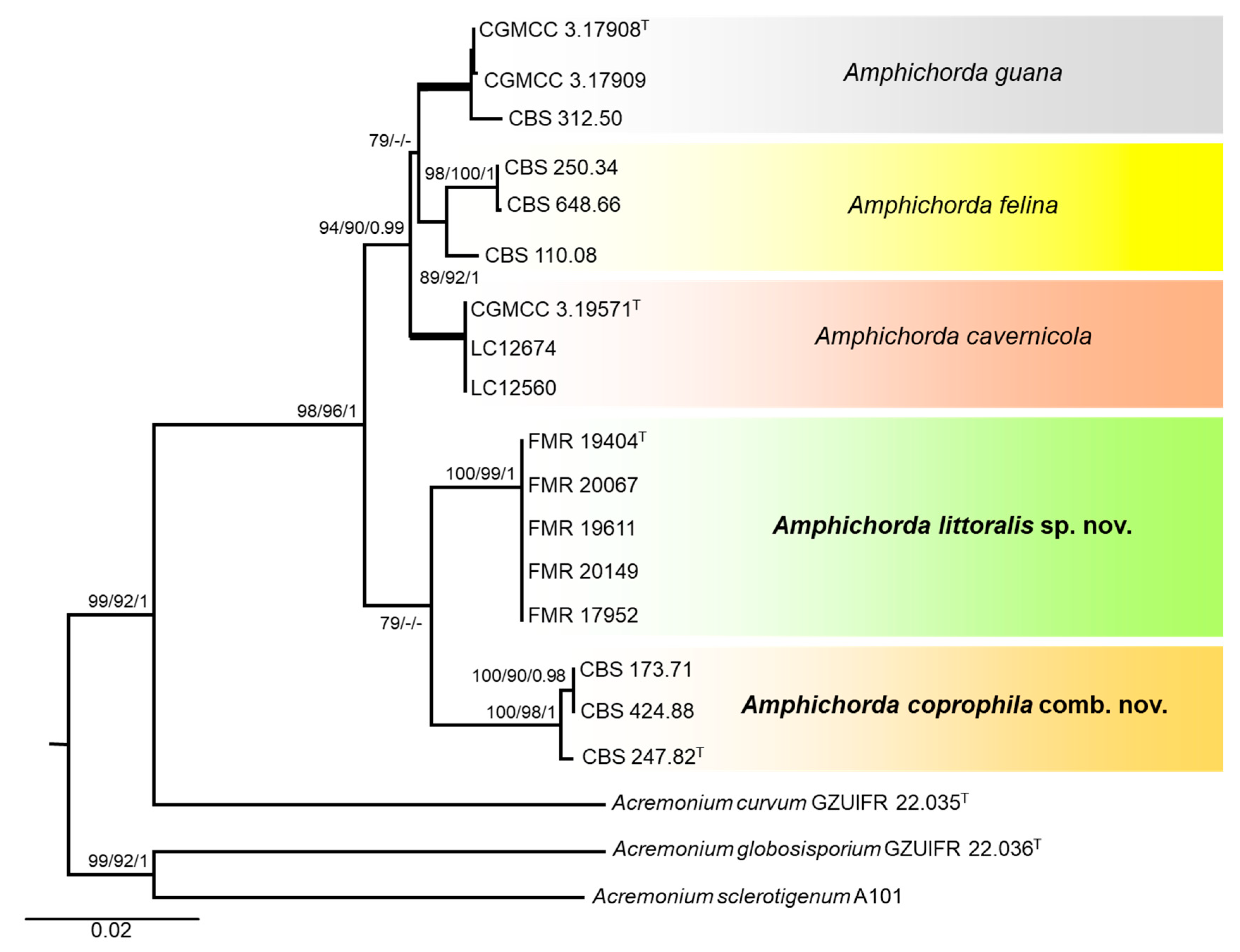
Figure 3.
Scanning electron microscopy (SEM) of the reproductive structures from representative Amphichorda species. (A) Conidiophores, holoblastic conidiogenous cells and conidia in Amphichorda felina (CBS 250.34). (B) Holoblastic conidiogenous cell in Amphichorda guana (CBS 312.50). (C) Conidiophores, holoblastic conidiogenous cells and conidia in Amphichorda littoralis (FMR 20067). (D) Conidiophores, holoblastic conidiogenous cells and conidia from Amphichorda coprophila (ex-type CBS 247.82). (E, F) Enteroblastic (phialidic−arrows) and holoblastic conidiogenous cells in Amphichorda coprophila (CBS 424.88), respectively. The black arrow points at the enteroblastic collarette. Scale bars: (A, C, D) = 5 µm; (B, E, F) = 2.5 µm.
Figure 3.
Scanning electron microscopy (SEM) of the reproductive structures from representative Amphichorda species. (A) Conidiophores, holoblastic conidiogenous cells and conidia in Amphichorda felina (CBS 250.34). (B) Holoblastic conidiogenous cell in Amphichorda guana (CBS 312.50). (C) Conidiophores, holoblastic conidiogenous cells and conidia in Amphichorda littoralis (FMR 20067). (D) Conidiophores, holoblastic conidiogenous cells and conidia from Amphichorda coprophila (ex-type CBS 247.82). (E, F) Enteroblastic (phialidic−arrows) and holoblastic conidiogenous cells in Amphichorda coprophila (CBS 424.88), respectively. The black arrow points at the enteroblastic collarette. Scale bars: (A, C, D) = 5 µm; (B, E, F) = 2.5 µm.
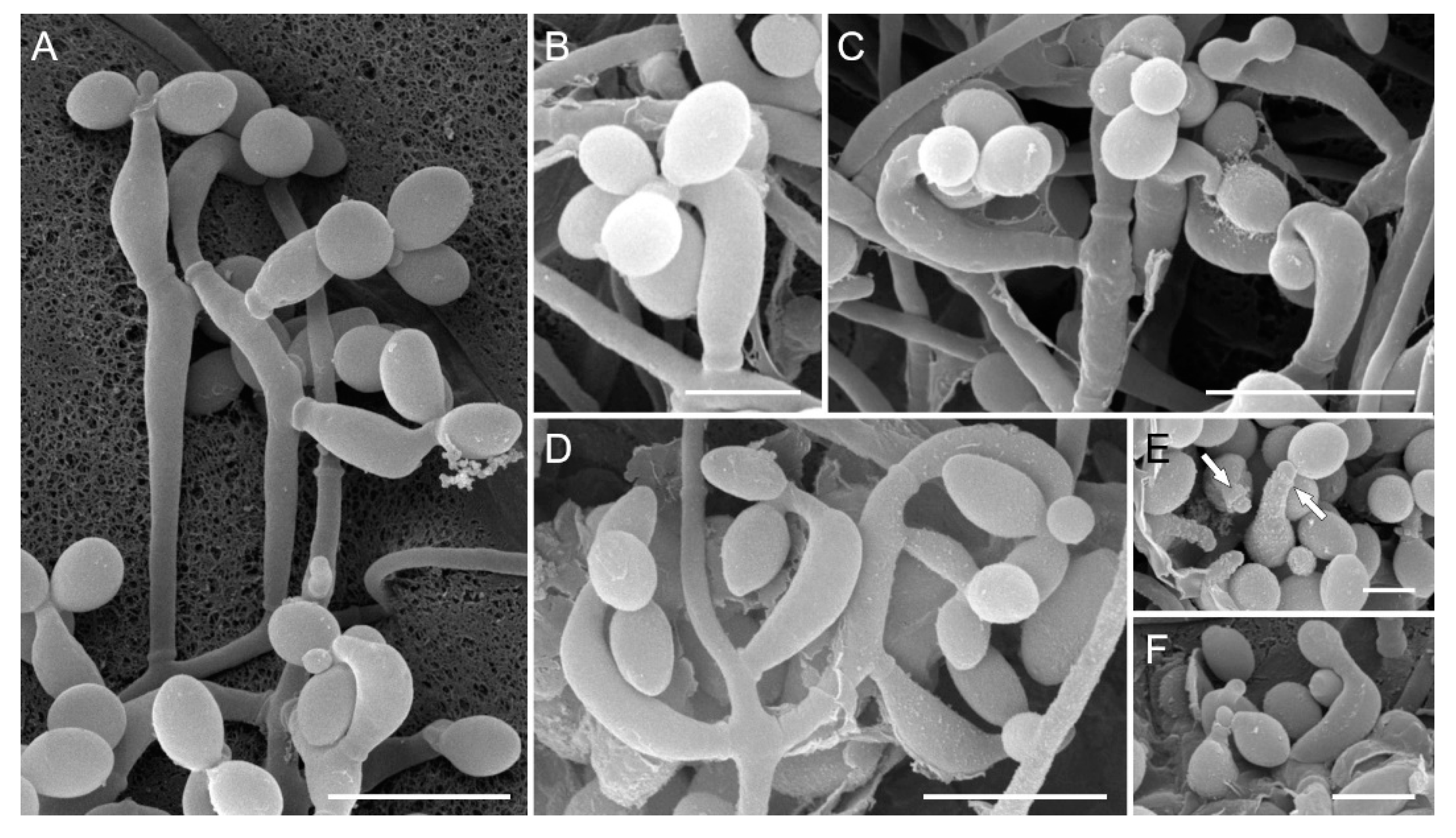
Figure 4.
Mean colony diameter achieved by representative Amphichorda strains on malt extract agar (MEA) supplemented with different concentrations of NaCl after 4 weeks at 25 °C. The strain Aspergillus chevalieri (FMR 19829) was used as a positive control of growth in the different culture media. On the right side, culture plates on MEA5% and MEA10% are represented for representative strains from each species.
Figure 4.
Mean colony diameter achieved by representative Amphichorda strains on malt extract agar (MEA) supplemented with different concentrations of NaCl after 4 weeks at 25 °C. The strain Aspergillus chevalieri (FMR 19829) was used as a positive control of growth in the different culture media. On the right side, culture plates on MEA5% and MEA10% are represented for representative strains from each species.
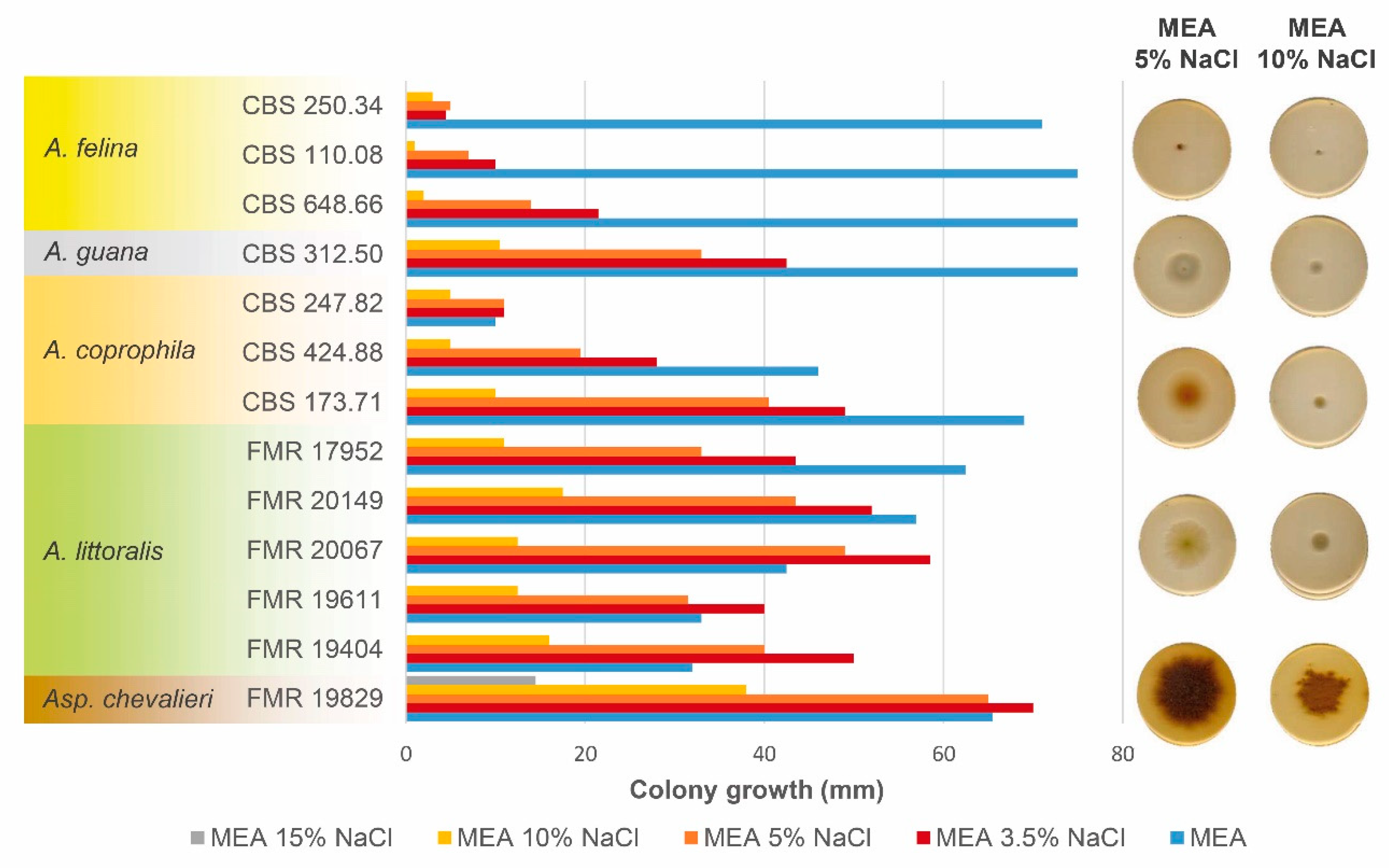
Figure 5.
Amphichorda littoralis (ex-type FMR 19404). (A) Colony on PDA. (B) Colony on OA. (C) Colony on SNA, after 14 d. at 25 °C. (D) Synnema from strain FMR 20067. (E) Semi-macronematous conidiophore. (F, G) Micronematous conidiophores with attached conidia. (H, I) Conidiogenous cells growing directly from hyphae with attached conidia. (J) Conidia. Scale bars: 10 µm.
Figure 5.
Amphichorda littoralis (ex-type FMR 19404). (A) Colony on PDA. (B) Colony on OA. (C) Colony on SNA, after 14 d. at 25 °C. (D) Synnema from strain FMR 20067. (E) Semi-macronematous conidiophore. (F, G) Micronematous conidiophores with attached conidia. (H, I) Conidiogenous cells growing directly from hyphae with attached conidia. (J) Conidia. Scale bars: 10 µm.
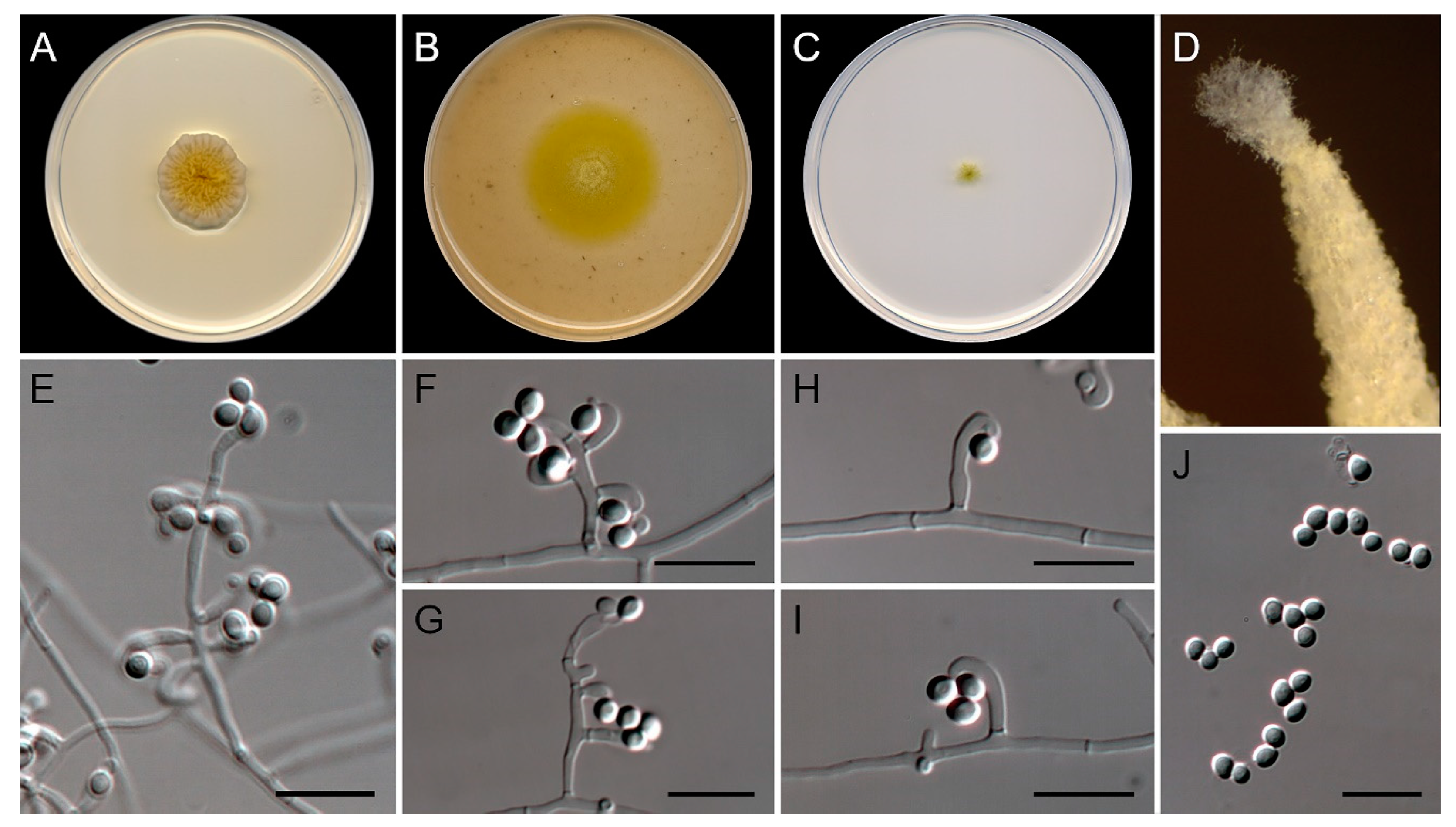
Table 2.
Synopsis of the morphological characters defining Amphichorda species.
| Species | Colony on PDA* | Colony on OA/SNA* | Microscopic features | Citation | ||
| Color | Diffusible pigment | Color | Conidiogenous cells size (µm) | Conidia size (µm) | ||
| A. cavernicola | Cream yellow to sea shell | Not observed | White | 4.5-8 x 2-3 | 2.5-4 x 2-3.5 | [12]; this study |
| A. coprophila | Orange to brownish orange | Greyish orange | Light yellow | 6-10 x 2-2.5 | 3.5-5.5 x 2-2.5 | [15]; this study |
| A. felina | White | Greyish orange | White | 3-8.5 x 2-2.5 | 2.5-4.5 x 2-3.5 | [9]; this study |
| A. guana | White to yellowish | Yellowish | White | 7-10 x 2-3 | 4.5-5.5 x 3.5-5 | [11]; this study |
| A. littoralis | Greenish yellow | Light yellow | Greenish yellow | 5.5-11.5 x 1.5-2.5 | 2.5-4 x 2.5-3 | This study |
* Obverse side of the colony.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



