Submitted:
22 May 2023
Posted:
23 May 2023
You are already at the latest version
Abstract
Melanocortins play crucial roles in regulating the stress response, inflammation, and skin pigmentation. In this review, we focus on the melanocortin 1 receptor (MC1R), a G protein-coupled receptor primarily known for regulating skin pigmentation and exhibiting anti-inflammatory effects. First, we provide an overview of the structure, signaling pathways, and related diseases of MC1R. Next, we discuss the potential therapeutic use of synthetic peptides and small molecule modulators of MC1R, highlighting the development of various drugs that enhance stability through amino acid sequence modifications and small molecule drugs to overcome limitations associated with peptide characteristics. Notably, MC1R-targeted drugs have applications beyond skin pigmentation-related diseases, which predominantly affect MC1R in melanocytes. These drugs can also be useful in treating inflammatory diseases with MC1R expression present in various cells. Our review underscores the potential of MC1R-targeted drugs to treat a wide range of diseases and encourages further research in this area.
Keywords:
Melanocortin
; receptor
; peptide
; modulator
; disease
1. Introduction
Melanocortins are a group of hormones that play crucial roles in regulating various physiological processes, including stress response, inflammation, and skin pigmentation. These hormones are polypeptides derived from pro-opiomelanocortin (POMC) and include adrenocorticotropin hormone (ACTH), α-, β-, and γ-melanocyte stimulating hormone (α-, β-, and γ-MSH) [1,2]. While the functions of ACTH and α-MSH are well-established [3] in skin pigmentation, anti-inflammatory and microbicidal characteristics [4,5,6], the role of β-MSH and γ-MSH are less well understood. Some studies suggested that they possess anti-inflammatory properties [7,8,9,10].
The functions of melanocortin peptides are mediated by G protein-coupled receptors (GPCRs), primarily the stimulatory G protein (Gs) [11]. The melanocortin receptor family is the smallest member of class A, a rhodopsin-like family of GPCRs, and consists of five isotypes (MC1R, MC2R, MC3R, MC4R, and MC5R) with varied tissue expression and functions [12]. MC1R, mainly expressed in both melanocytes and leukocytes, enhances UV resistance and anti-inflammatory signaling when activated. MC2R is found in the adrenal cortex, and both MC3R and MC4R, which are reported to control food intake and sexual function, are largely present in the CNS. MC5R, located in the brain and skeletal muscle, has an exocrine role. The low sequence homology (40–60%) among the five receptors explains the lack of ligand selectivity [13,14,15,16].
The MC1R is a well-known receptor for α-MSH expressed in the skin and hair follicles, where it controls pigmentation. However, recent studies have revealed that MC1R is also expressed in various other cell types that are susceptible to the anti-inflammatory effects of melanocortins [17,18]. Thus, understanding the regulation of MC1R could potentially lead to the development of novel therapeutic strategies. This review article provides a brief overview of the structure, signaling pathway, and related diseases associated with MC1R and discusses the potential of synthetic peptide modulators and small molecule modulators of MC1R as therapeutic agents. Through a detailed examination of MC1R and its modulators, this review aims to provide insights into the potential clinical implications of modulating MC1R activity.
2. Melanocortin 1 receptor (MC1R)
2.1. Structure
The human MC1R is primarily found on melanocytes and malignant melanoma cells and consists of 317 amino acids [19,20,21]. While the normal expression of MC1R protein is low, melanocytes express approximately 700 protein units, with slightly higher amounts found on melanoma cells [22,23]. MC1R is a GPCR with seven α-helical transmembrane (TM) domains, an N-linked glycosylation site at the external N-terminus, a palmitoylation site at the intracellular C-terminus, and a DRY motif at the junction of the third TM domain. Unlike other GPCRs, the first and second extracellular domains of the MC receptor subfamily lack one or two cysteines, while the fourth and fifth TM domains lack proline. The intracellular and transmembrane domains of MC1R regulate adenylyl cyclase connections and signaling, while the extracellular and transmembrane domains interact with MC1R ligands [11,12].
The extracellular N-terminal tail serves as a signal anchor and plays a crucial role in ligand affinity. A conserved cysteine residue at the intersection of the N-terminus and the first TM domain is crucial for receptor function [24,25,26,27,28]. The C-terminus is involved in receptor interactions with the G protein at the plasma membrane, as well as protein trafficking from the endoplasmic reticulum to the plasma membrane. Also, C-terminus affects desensitization, internalization, and the plasma membrane location of the receptor [29,30,31,32,33].
The intracellular and extracellular loops (ils and els, respectively) lie between the transmembrane domains of the melanocortin 1 receptor. These loops share conserved sequences with other MC receptors. Despite being smaller than most GPCRs, MC1R els are essential for constitutive basal signaling activity. Mutations in this area affect binding affinity, as the els of the MC1R interact with ligands. Due to conserved proline and cysteine residues, els appears to be essential for melanocortin affinity. Similarly, MC1R ils plays a crucial role in Gs protein binding and have phosphorylation sites that impact signal modulation, internalization, and receptor cycling [25,27,28,31,34].
Recently, the Cryo-electron microscopy (Cryo-EM) structure of MC1R and the MC1R–Gs complexes bound to the endogenous hormone α-MSH, the marketed drug afamelanotide, and a synthetic agonist called SHU9119 were determined [35]. These findings may pave the way for resolving the lack of selectivity in MC drug discovery.
2.2. Signaling pathway
The melanocortin 1 receptor (MC1R) is a receptor that forms a complex with heterotrimeric G proteins. When agonistic ligands bind to MC1R, the Gαs protein is separated, and MC1R activates adenylyl cyclase, which leads to the production of cAMP, a crucial second messenger that regulates many cellular processes. In melanocytes, cAMP activates protein kinase A (PKA) and triggers downstream signaling pathways that activate different effector pathways, including the CREB and MITF networks. These pathways lead to the increased expression of tyrosinase and dopachrome tautomerase, two enzymes that are involved in melanin synthesis, resulting in the production of melanin. The melanin produced is then transmitted to nearby keratinocytes, creating a protective layer that improves the skin's ability to prevent further UV damage. Moreover, the increase in cAMP levels in melanocytes enhances antioxidant defenses and accelerates nucleotide excision repair (NER), which is vital for safeguarding the skin from UV damage (Figure 1) [36,37,38,39,40,41,42,43].
Moreover, it is also reported that there is a cAMP-independent pathway mediated by cKIT, which is not discussed in this review. This pathway activates the extracellular signal-regulated protein kinases 1 and 2 (ERK 1/2) by triggering the NRAS-BRAF-MEK-ERK cascade and active ERKs can phosphorylate MITF. Furthermore, it has been suggested that the activation of AKT downstream of α-MSH may also occur via cKIT [44].
Furthermore, the MC1R signaling axis has an anti-inflammatory effect through downstream pathways. When α-MSH binds to MC1R, it triggers the production of intracellular cAMP, which activates protein kinases C and A, leading to the activation of the MAPK and JAK-STAT pathways. These pathways prevent IκB degradation and activate CREB, a transcription factor that regulates anti-inflammatory mediators such as IB and IL-10. In addition, protein kinase activation enhances the levels of cytoplasmic IκB, inhibiting the expression of downstream pro-inflammatory genes, including IL-1, TNF-a, IL-6, IL-8, and IL-12, iNOS, and adhesion molecules (ICAM-1, VCAM-1, and MMPs) (Figure 2) [45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60].
3. MC1R related diseases
3.1. Melanoma
Melanoma is a highly aggressive and resistant cancer that results from the malignant progression of melanocytes, which are pigment cells located in the skin's epidermis [61]. The traditional chemotherapy treatments have demonstrated limited efficacy in treating melanoma, which highlights the need for innovative therapeutic approaches that can take advantage of the distinctive features of these tumors [62]. One such characteristic is the overexpression of the MC1R cell surface endocytic receptor on the surface of human melanomas, which makes it a crucial tumor marker [63,64]. The activation of MC1R is necessary to regulate melanocyte cell division and melanin production, which provides the skin with protection from UV radiation. Additionally, MC1R has potential applications in diagnostics and targeted drug delivery [65,66]. Innovative treatment approaches are currently being developed based on MC1R for melanoma. MC1R expression is frequently elevated in both melanoma cell lines and melanomas, and contrary to earlier claims that MC1R activation could cause melanoma, recent scientific evidence demonstrates that this receptor not only does not cause melanoma but that its activation can actually stimulate DNA repair processes that have the potential to prevent the disease [67,68,69].
3.2. Systemic sclerosis
Systemic sclerosis (SSc), or scleroderma, is an autoimmune disorder characterized by dysregulation of the immune system and inflammation, microvascular dysfunction, and widespread fibrosis in various organs [70,71]. While no approved medications exist for SSc apart from those for SSc-associated ILD (interstitial lung disease), there is a pressing need for innovative treatments that can effectively target fibrosis in multiple organs, including the skin [72,73,74]. Activating the melanocortin 1 receptor has been shown to have broad anti-inflammatory and antifibrotic effects. Thus, a recent study investigated the potential of a novel oral MC1R agonist, dersimelagon (MT-7117), as a treatment for SSc. In preclinical SSc models, MT-7117 exhibited disease-modifying effects. Target expression analysis and research into its mode of action indicated that MT-7117 has a favorable effect on inflammation, vascular dysfunction, and fibrosis, all of which are key pathologies in SSc. These findings suggest that MT-7117 may have potential as a treatment for SSc. A phase 2 clinical trial is currently underway to evaluate the efficacy and tolerability of MT-7117 in patients with early, progressive diffuse cutaneous SSc [74]. More information about dersimelagon will be discussed in the section on small molecule modulators.
3.3. Neuroinflammation
Neuroinflammation is a critical factor in the progression of neurological damage caused by hypoxic-ischemic (HI) events, and microglia play a significant role in this process [75,76]. In various neurological disorders, MC1R activation has been shown to have anti-inflammatory and neuroprotective effects [77,78]. Recently, researchers investigated the potential of MC1R activation to mitigate neuroinflammation and repair neurological impairments in neonatal rats with hypoxic-ischemic neurological damage [79]. The results showed that BMS-470539, an MC1R activator, reduced neuroinflammation and repaired neurological impairments in these rats. The study also revealed that the anti-inflammatory and neuroprotective effects of MC1R activation were mediated through the MC1R/cAMP/PKA/Nurr1 signaling pathway. The findings suggest that MC1R activation could be a promising therapeutic target for the treatment of hypoxic-ischemic encephalopathy (HIE) in newborns. Additional details regarding BMS-470539 will be provided in the small molecule modulators section.
3.4. Atherosclerosis
Monocytes and macrophages are known to express the MC1R, which mediates anti-inflammatory effects and helps prevent macrophage foam cell production by increasing cholesterol efflux via ABCA1 and ABCG1 transporters [80,81,82]. Recently, a study investigated whether a systemic deficiency of MC1R signaling affects the development of atherosclerosis. The study highlighted the significant role of MC1R in the development of atherosclerosis. The findings suggested that a lack of MC1R signaling may exacerbate atherosclerosis by disrupting cholesterol transport and increasing arterial monocyte deposition [83]. Furthermore, MC1R plays a role in immunomodulation. Specifically, α-MSH has been shown to decrease pro-inflammatory cytokines in various pulmonary inflammatory disorders, including asthma, sarcoidosis, and acute respiratory distress syndrome. Animal models of pulmonary fibrosis have also demonstrated that α-MSH can reduce fibrogenesis [84]. In addition to the diseases previously mentioned, MC1R has also been implicated in other diseases, such as intestinal and ocular inflammation [85], and Parkinson's disease [86]. Recent research has shown that modulating MC1R may have therapeutic potential for the diseases previously mentioned.
4. Modulators
4.1. Endogenous ligands
After UV damage, melanocortins such as α-MSH and ACTH are induced in the skin, and these ligands protect the skin by binding to MC1R and triggering changes in melanocytes that enhance their resistance to UV (Figure 3) [87,88,89]. However, the MC1R can also be affected by other ligands, such as agouti signaling protein (ASIP) and β-defensin 3 (βD3), which can have a significant impact on MC1R signaling [90,91,92,93]. Melanocortins stimulate MC1R signaling, while ASIP inhibits MC1R signaling and reduces cAMP levels [94,95]. In contrast, βD3 has little effect on signaling but can act as a competitive inhibitor, interfering with the binding of α-MSH or ASIP [96,97]. Synthetic modulators of MC1R, including both peptides and small molecules, have been reported, and now some of these modulators will be introduced.
4.2. Synthetic Peptide modulators
As previously explained, the endogenous ligands for melanocortin receptors are ACTH and α-, β-, γ-MSH. The most important finding in early drug development was that the amino acid sequence, His6-Phe-Arg-Trp9, was the key sequence needed to activate the melanocortin receptor [98]. And the sequence was modified to solve the instability, which was a problem of α-MSH, leading to the development of [Nle4, D-Phe7]-α-MSH (Melanotan I) [99,100]. Additionally, studies on the cyclization of peptides were conducted to obtain Melanotan II using NDP-MSH as a scaffold [101,102]. With these findings, efforts are currently underway to produce selective and stable peptides on the melanocortin 1 receptor, such as PL-8177 [67]. Among them, we would like to introduce Melanotan I and II, which are key substances in the development of peptide drugs (Figure 4).
4.2.1. Melanotan I (MT-I)
Melanotan I is an early analog of α-MSH that acts as a non-selective agonist of melanocortin receptors and stimulates melanin production. During the development of peptide-based drugs, researchers focused on the common sequence of MSH, namely His6-Phe-Arg-Trp9, and further studies were conducted on this sequence (Figure 4) [98]. Eventually, in the frog skin bioassay, it was observed that Ac(Acetyl)-α-MSH7-10-NH2 and Ac-α-MSH6-8-NH2 had no activity, while Ac-α-MSH6-9-NH2 did. This led to the discovery that the minimal sequence required for the biological activity of α-MSH is His6-Phe-Arg-Trp9 [99]. Furthermore, to increase the stability of the initial α-MSH analog, Sawyer et al replaced Phe at position 7 with D-Phe and Methionine at position 4 with the amino acid norleucine to prevent oxidation of methionine. These changes resulted in the production of [Nle4, D-Phe7]-α-MSH (NDP-MSH, Melanotan I), a peptide with higher potency and stability than α-MSH [100].
When Melanotan I activates MC1R, cAMP is produced and it activates microphthalmia transcription factor (MITF) expression, which induces the expression of enzymes for eumelanin production. This process increases the production of eumelanin in melanocytes. In addition, Melanotan I activates tyrosinase, and induces an increase in eumelanin content in melanocytes [103,104,105].
Afamelanotide, well-known as the international nonproprietary name of Melanotan I, has been used in patients with erythropoietic protoporphyria (EPP) since 2019. EPP is a disease that causes abnormal hemoglobin synthesis in red blood cells, which can cause skin damage even with a little sunlight. When Afamelanotide binds to MC1R, it activates melanocytes and stimulates eumelanin synthesis. Eumelanin protects against UV light by absorbing and scattering it, scavenging free radicals and reactive oxygen species, and acting as a neutral density filter capable of decreasing transmission of all wavelengths of light [106,107]. Recent studies have shed light on the binding structure of MC1R and ligands, while no information has yet been disclosed about the binding structure of receptors and peptides. Afamelanotide, the compound that was switched from Phe7 to D-Phe7 in α-MSH, has an extra hydrogen bond with the TM2 domain in D-Phe7. It has been demonstrated that Afamelanotide has a higher affinity for receptors than α-MSH, as evidenced by its capacity to make cAMP, which is superior to α-MSH, as determined by various mutants (Figure 5) [35].
4.2.2. Melanotan II (MT-II)
Melanotan II is a cyclic peptide that is derived from Melanotan I (Figure 6). In particular, the peptide is modified by replacing Glu5 with Asp and Gly10 with Lys. Previous studies by Obeidi and Hadley have demonstrated that cyclic peptides are more stable and potent than linear peptides. Their research investigated the efficacy and stability of a linear form of Ac-[Nle4, D-Phe7, Lys10]-α-MSH4-10-NH2 and a cyclic form, in which Asp5 and Lys10 were linked. The findings of their bioassay demonstrated that cyclic peptides exhibited greater potency and stability than linear peptides [101,102]. Melanotan II has been shown to activate melanocytes as an MC1R agonist, but it is weaker than Melanotan I. However, Melanotan II was proven to have a stronger sexual effect [108]. It has been reported to induce penile erections in male rats and sexual inspiration [109,110], and increases the sexual activities of female rats [111]. Bremelanotide (PT-141), derived from Melanotan II, acts as an agonist of the MC1R and MC4R and has been demonstrated to increase erectile function and sexual desire (Figure 6) [112,113].
In 2020, Zhou et al reported an attractive study in which melanotan II was utilized as a probe for selective drug delivery [114,115]. The authors synthesized and designed ligand-drug conjugates with the melanocortin 1 receptor (MC1R) agonist melanotan-II (MT-II) to couple a cytotoxic drug, camptothecin with low cancer resistance (Figure 7). The drug-MT-II conjugates efficiently bound to MC1R and selectively delivered drugs to A375 melanoma cells in vitro. Camptothecin-MT-II was used to study the inhibitory activity on A375 melanoma cells and yielded an IC50 of 16 nM. This approach of drug-MT-II conjugates offers more options in cytotoxic drug selection and is noteworthy for potentially overcoming the cancer resistant problem of melanoma.
4.3. Small molecule modulators
During the development of peptide-based drugs, challenges such as low stability of enzymes and selectivity for receptors were encountered. For example, the peptide agent, Melanotan I (Afamelanotide) is a non-selective agonist of melanocortin receptors except for MC2R, and its binding to MC3R in the gut may cause nausea [116]. To address the limitations of peptide-based therapeutics, small molecule development has been rapidly evolving, utilizing peptides as starting scaffolds and screening compound libraries [117]. As a result, BMS-470539, derived from an MC4R agonist, has been developed [118]. Additionally, various agonists, including AP1189 and CD08108, are under development [119,120]. Recently, Dersimelagon (MT-7117), which selectively acts on MC1R, has been approved in Europe for the treatment of EPP and also has an anti-inflammatory effect [74,116]. Furthermore, JNJ-10229570, an antagonist of the melanocortin 1, 5 receptors, has been developed, and it has been reported to affect sebaceous lipid production [121].
4.3.1. BMS-470539
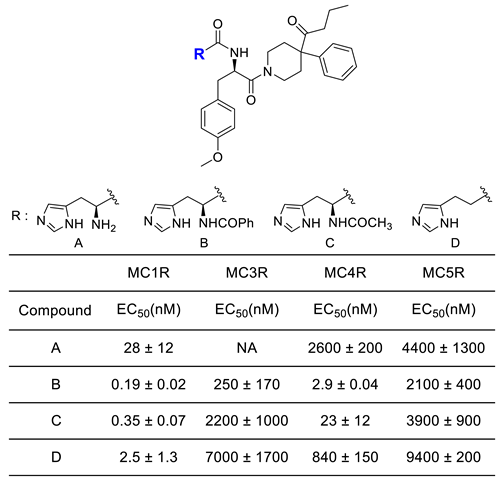
BMS-470539 is a small molecule agonist selective for MC1R, which was synthesized by modifying the structure of a known MC4R agonist (Figure 8). In a study conducted by Timothy et al., the selectivity for receptors of several compounds was measured by comparing their activity using EC50 values of other isotypes. The EC50 (nM) value for MC1R of Compound A (BMS-470539) was 28 ± 12 nM, which was higher than Compound B, Compound C, and Compound D, indicating relatively low activity compared to other compounds (Table 1). However, when expanded to other receptors, it was found that Compound 1 did not activate MC3R, and the selectivity for MC4R and MC5R was significantly lower than that of MC1R. Also, the other compounds had receptors with relatively similar efficacy to MC1R and had relatively high intrinsic activity (IA) for all receptors. This suggests that other compounds may have higher activity but lower selectivity than compound 1 [118]. Therefore, Compound 1, known as BMS-470539, was identified as having much better selectivity than other compounds (Table 1).
BMS-470539 is also known for its anti-inflammatory effects. BMS-470539 was administered to mice in which an immune response was induced by LPS, and the amount of TNF-α, an inflammatory factor, was measured. The results showed a rapid decrease of up to 92 percent at 100μmol/kg of BMS-470539 [118]. Kang et al investigated the activity of NF-κB associated with inflammation to prove the anti-inflammatory effect of BMS-470539 and found a noticeable decrease in the activity of NF-κB. Moreover, it induced a decrease in the infiltration of white blood cells in the lungs of mice by LPS [78]. Recently, BMS-470539 was found to decrease the phosphorylation of p38, ERK1/2, and JNK involved in the MAPK pathway associated with inflammation in neutrophils stimulated by LPS [122].
4.3.2. MT-7117 (Dersimelagon)
MT-7117, or Dersimelagon, is a selective agonist of MC1R that has been found to have a much higher selectivity for MC1R than NDP-MSH (Afamelanotide), a peptide effective in treating erythropoietic protoporphyria (EPP). In binding affinity and agonistic activity measurement experiments conducted by Suzuki et al, MT-7117 was demonstrated to have a Ki value of 2.26 nM for hMC1R (human MC1R), with much higher selectivity ranging from at least 15 to as much as 700 times compared to other receptors (Figure 9). On the other hand, NDP-MSH showed only a 10-fold difference in selectivity for MC1R compared to other receptors which suggested that NDP-MSH has lower selectivity than MT-7117 [123]. Due to issues with enzyme stability associated with peptide drugs and higher selectivity for MC1R, MT-7117 has been recently approved in Europe as an alternative treatment for EPP [116].
A recent study revealed that MT-7117 exhibits anti-inflammatory effects [74]. The study showed that daily oral administration of MT-7117 to rats with fibrosis induced by bleomycin significantly delayed skin fibrosis and lung inflammation. In addition, it was found to significantly reduce the expression of ACTA2 (α-smooth muscle actin) mRNA in human skin fibroblasts that had been increased by TGF-β. This effect was attributed to the inhibition of inflammatory signaling pathways such as IL-6 signaling. Based on these effects, a clinical trial is currently underway to investigate the association of MT-7117 with SSc patients.
4.3.3. AP1189 (Resomelagon)
A series of pyrrole aminoguanidine derivatives were investigated as ligands in the melanocortin receptors by Lundstedt et al. In order to assess the activity of melanocortin receptors, researchers measured the binding affinities (Ki) of pyrrole aminoguanidine derivatives for MC1R, MC3R, MC4R, and MC5R. While most of the compounds didn’t displayed significant activity or selectivity, the phenylpyrrole allylidene derivatives exhibited enhanced binding affinities towards MC1R and therefore, meaningful selectivities. The phenyl group was found to be particularly important for binding to MC1R. AP1189 ([1-(4-chlorophenyl)-1H-pyrrol-2-yl-allylideneamino]guanidinium acetate) was identified as an agonist for melanocortin receptors MC1R and MC3R. (Figure 10) [119,124].
AP1189 has a unique mechanism that distinguishes it from other compounds. It does not stimulate cAMP production and is not involved in melanogenesis. Instead, it induces phosphorylation of ERK1/2, resulting in a decrease in cytokine levels in macrophages. , This may help resolve acute inflammation and potentially alleviate arthritis in mice [125]. Based on these findings, AP1189 is currently being evaluated in clinical trials for the treatment of rheumatoid arthritis and idiopathic membranous nephropathy [67].
4.3.4. CD08108
Selective novel MC1R agonists were discovered by the research group in GALDERMA research and development in 2013 [120]. Imidazolyl-linked azetidine derivatives were synthesized and evaluated for activity for human melanocortin receptors. Among them, CD08108 exhibited a significant hMC1R agonistic activity with EC50 of 70 nM and showed the selectivity of MC1R over MC4R up to 64 times (Figure 11). In 2015, Boiteau et al. reported an efficient synthetic route for CD08108 in a kilogram scale [126]. CD08108 has been implicated in the process of melanogenesis, indicating its potential as a therapeutic target for the treatment of skin conditions such as hypodermic or photosensitive diseases.
4.3.5. JNJ-10229570
2,3-Diphenyl-5-anilino[1,2,4]thiadiazole has been confirmed to act as an agonist for MC4R, thereby inhibiting food intake and affecting eating behavior in mice. JNJ-10229570 is a compound with 2,3-(2-methoxyphenyl) at 2,3-Diaryl, and reported to function as an antagonist for MC1R and MC5R, which are melanocortin receptors expressed in sebocytes (Figure 12). Melanocortin receptors in sebocytes are known to play a crucial role in producing sebaceous lipids. Studies on the relationship between MC5R activation and sebocytes indicate that the activation of MC5R increases the production of cAMP, inducing the production of sebaceous lipids [127,128,129]. Sebaceous lipids are essential for skin integrity and inflammatory processes. However, excessive sebum production is shown to be a significant contributor to the pathophysiology of Acne vulgaris [130,131,132]. Eisinger et al. demonstrated that JNJ-10229570 acts as an antagonist for MC1R and MC5R present in human sebaceous cells, inhibiting sebaceous secretion and reducing sebaceous gland size [121]. The IC50 measurements for MC1R and MC5R, using 125I-NDP-α-MSH, were found to be 270 ± 120 and 200 ± 50nM (Mean ± S.D.), respectively, indicating that JNJ-10229570 binds to both receptors. Furthermore, the reduction in the amount of cAMP, elevated by NDP-MSH after the sebocyte was exposed to 0.6 nM of JNJ-10229570, suggests that this compound acts as an antagonist for MC1R and MC5R. In addition, human skin transplanted into SCID mice and treated with JNJ-10229570 showed a decrease in squalene, wax esters, and triglyceride, suggesting new information about the effect of JNJ-10229570 on surface lipid secretion in vivo.
5. Conclusions
The discovery of the melanocortin 1 receptor several decades ago has led to extensive research into its potential as a therapeutic target. Studies have revealed that the MC1R signaling pathway is crucial in treating melanoma, as it affects various physiological changes in melanocytes. However, recent research has also shown a growing interest in investigating the molecular mechanisms responsible for the non-pigmentary effects of MC1R, particularly in the treatment of inflammatory diseases. Ongoing investigations suggest that drugs targeting MC1R may hold promise in treating patients with high-need conditions such as systemic sclerosis, neuroinflammation, rheumatoid arthritis, fibrosis, and others. With numerous innovative molecules and clinical trials currently underway, it is possible that these drugs may soon improve the quality of life for those with chronic inflammatory conditions. Furthermore, the development of MC1R ligands using multiple chemistry approaches and knowledge of the receptor's crystal structure may lead to the creation of additional drugs to treat new conditions in the future.
Author Contributions
Conceptualization, D.S.; methodology, Y.M. W.K. and D.S.; writing—original draft preparation, Y.M. and W.K.; writing—review and editing, D.S. All authors have read and agreed to publishing this version of the manuscript.
Funding
This research was funded by the National Research Foundation of Korea (NRF-2020R1A6A1A03043708 and NRF-2021R1A2C1012280)
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
References
- Cawley, N.X.; Li, Z.; Loh, Y.P. 60 YEARS OF POMC: Biosynthesis, trafficking, and secretion of pro-opiomelanocortin-derived peptides. J Mol Endocrinol 2016, 56, T77–97. [Google Scholar] [CrossRef]
- Dores, R.M.; Liang, L.; Davis, P.; Thomas, A.L.; Petko, B. 60 YEARS OF POMC: Melanocortin receptors: evolution of ligand selectivity for melanocortin peptides. J Mol Endocrinol 2016, 56, T119–133. [Google Scholar] [CrossRef]
- Gallo-Payet, N. 60 YEARS OF POMC: Adrenal and extra-adrenal functions of ACTH. J Mol Endocrinol 2016, 56, T135–156. [Google Scholar] [CrossRef]
- Lipton, J.M.; Ceriani, G.; Macaluso, A.; McCoy, D.; Carnes, K.; Biltz, J.; Catania, A. Antiinflammatory effects of the neuropeptide alpha-MSH in acute, chronic, and systemic inflammation. Ann N Y Acad Sci 1994, 741, 137–148. [Google Scholar] [CrossRef]
- Catania, A.; Cutuli, M.; Garofalo, L.; Carlin, A.; Airaghi, L.; Barcellini, W.; Lipton, J.M. The neuropeptide alpha-MSH in host defense. Ann N Y Acad Sci 2000, 917, 227–231. [Google Scholar] [CrossRef]
- Cutuli, M.; Cristiani, S.; Lipton, J.M.; Catania, A. Antimicrobial effects of alpha-MSH peptides. J Leukoc Biol 2000, 67, 233–239. [Google Scholar] [CrossRef]
- Madeira, M.F.; Queiroz-Junior, C.M.; Montero-Melendez, T.; Werneck, S.M.; Correa, J.D.; Soriani, F.M.; Garlet, G.P.; Souza, D.G.; Teixeira, M.M.; Silva, T.A.; et al. Melanocortin agonism as a viable strategy to control alveolar bone loss induced by oral infection. FASEB J 2016, 30, 4033–4041. [Google Scholar] [CrossRef]
- Gomez-SanMiguel, A.B.; Martin, A.I.; Nieto-Bona, M.P.; Fernandez-Galaz, C.; Villanua, M.A.; Lopez-Calderon, A. The melanocortin receptor type 3 agonist d-Trp(8)-gammaMSH decreases inflammation and muscle wasting in arthritic rats. J Cachexia Sarcopenia Muscle 2016, 7, 79–89. [Google Scholar] [CrossRef]
- Gomez-SanMiguel, A.B.; Villanua, M.A.; Martin, A.I.; Lopez-Calderon, A. D-TRP(8)-gammaMSH Prevents the Effects of Endotoxin in Rat Skeletal Muscle Cells through TNFalpha/NF-KB Signalling Pathway. PLoS One 2016, 11, e0155645. [Google Scholar] [CrossRef]
- Kadiri, J.J.; Thapa, K.; Kaipio, K.; Cai, M.; Hruby, V.J.; Rinne, P. Melanocortin 3 receptor activation with [D-Trp8]-gamma-MSH suppresses inflammation in apolipoprotein E deficient mice. Eur J Pharmacol 2020, 880, 173186. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y. Structure, function and regulation of the melanocortin receptors. Eur J Pharmacol 2011, 660, 125–130. [Google Scholar] [CrossRef]
- Wolf Horrell, E.M.; Boulanger, M.C.; D'Orazio, J.A. Melanocortin 1 Receptor: Structure, Function, and Regulation. Front Genet 2016, 7, 95. [Google Scholar] [CrossRef]
- Gantz, I.; Shimoto, Y.; Konda, Y.; Miwa, H.; Dickinson, C.J.; Yamada, T. Molecular cloning, expression, and characterization of a fifth melanocortin receptor. Biochem Biophys Res Commun 1994, 200, 1214–1220. [Google Scholar] [CrossRef]
- Gantz, I.; Miwa, H.; Konda, Y.; Shimoto, Y.; Tashiro, T.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular cloning, expression, and gene localization of a fourth melanocortin receptor. J Biol Chem 1993, 268, 15174–15179. [Google Scholar] [CrossRef]
- Desarnaud, F.; Labbe, O.; Eggerickx, D.; Vassart, G.; Parmentier, M. Molecular cloning, functional expression and pharmacological characterization of a mouse melanocortin receptor gene. Biochem J 1994, 299 ( Pt 2) Pt 2, 367–373. [Google Scholar] [CrossRef]
- Mountjoy, K.G.; Robbins, L.S.; Mortrud, M.T.; Cone, R.D. The cloning of a family of genes that encode the melanocortin receptors. Science 1992, 257, 1248–1251. [Google Scholar] [CrossRef]
- Schaible, E.V.; Steinstrasser, A.; Jahn-Eimermacher, A.; Luh, C.; Sebastiani, A.; Kornes, F.; Pieter, D.; Schafer, M.K.; Engelhard, K.; Thal, S.C. Single administration of tripeptide alpha-MSH(11-13) attenuates brain damage by reduced inflammation and apoptosis after experimental traumatic brain injury in mice. PLoS One 2013, 8, e71056. [Google Scholar] [CrossRef]
- Bhardwaj, R.; Becher, E.; Mahnke, K.; Hartmeyer, M.; Schwarz, T.; Scholzen, T.; Luger, T.A. Evidence for the differential expression of the functional alpha-melanocyte-stimulating hormone receptor MC-1 on human monocytes. J Immunol 1997, 158, 3378–3384. [Google Scholar] [CrossRef]
- Siegrist, W.; Stutz, S.; Eberle, A.N. Homologous and heterologous regulation of alpha-melanocyte-stimulating hormone receptors in human and mouse melanoma cell lines. Cancer Res 1994, 54, 2604–2610. [Google Scholar] [PubMed]
- Siegrist, W.; Solca, F.; Stutz, S.; Giuffre, L.; Carrel, S.; Girard, J.; Eberle, A.N. Characterization of receptors for alpha-melanocyte-stimulating hormone on human melanoma cells. Cancer Res 1989, 49, 6352–6358. [Google Scholar] [PubMed]
- Ghanem, G.E.; Comunale, G.; Libert, A.; Vercammen-Grandjean, A.; Lejeune, F.J. Evidence for alpha-melanocyte-stimulating hormone (alpha-MSH) receptors on human malignant melanoma cells. Int J Cancer 1988, 41, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.W.; Newton, R.A.; Beaumont, K.A.; Helen Leonard, J.; Sturm, R.A. Quantitative analysis of MC1R gene expression in human skin cell cultures. Pigment Cell Res 2006, 19, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Donatien, P.D.; Hunt, G.; Pieron, C.; Lunec, J.; Taieb, A.; Thody, A.J. The expression of functional MSH receptors on cultured human melanocytes. Arch Dermatol Res 1992, 284, 424–426. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Laorden, B.L.; Sanchez-Mas, J.; Turpin, M.C.; Garcia-Borron, J.C.; Jimenez-Cervantes, C. Variant amino acids in different domains of the human melanocortin 1 receptor impair cell surface expression. Cell Mol Biol (Noisy-le-grand) 2006, 52, 39–46. [Google Scholar] [PubMed]
- Frandberg, P.A.; Doufexis, M.; Kapas, S.; Chhajlani, V. Cysteine residues are involved in structure and function of melanocortin 1 receptor: Substitution of a cysteine residue in transmembrane segment two converts an agonist to antagonist. Biochem Biophys Res Commun 2001, 281, 851–857. [Google Scholar] [CrossRef]
- Wallin, E.; von Heijne, G. Properties of N-terminal tails in G-protein coupled receptors: a statistical study. Protein Eng 1995, 8, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Chhajlani, V.; Xu, X.; Blauw, J.; Sudarshi, S. Identification of ligand binding residues in extracellular loops of the melanocortin 1 receptor. Biochem Biophys Res Commun 1996, 219, 521–525. [Google Scholar] [CrossRef]
- Garcia-Borron, J.C.; Sanchez-Laorden, B.L.; Jimenez-Cervantes, C. Melanocortin-1 receptor structure and functional regulation. Pigment Cell Res 2005, 18, 393–410. [Google Scholar] [CrossRef]
- Pitcher, J.A.; Freedman, N.J.; Lefkowitz, R.J. G protein-coupled receptor kinases. Annu Rev Biochem 1998, 67, 653–692. [Google Scholar] [CrossRef]
- Luttrell, L.M.; Lefkowitz, R.J. The role of beta-arrestins in the termination and transduction of G-protein-coupled receptor signals. J Cell Sci 2002, 115, 455–465. [Google Scholar] [CrossRef]
- Strader, C.D.; Fong, T.M.; Tota, M.R.; Underwood, D.; Dixon, R.A. Structure and function of G protein-coupled receptors. Annu Rev Biochem 1994, 63, 101–132. [Google Scholar] [CrossRef] [PubMed]
- Qanbar, R.; Bouvier, M. Role of palmitoylation/depalmitoylation reactions in G-protein-coupled receptor function. Pharmacol Ther 2003, 97, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Schulein, R.; Hermosilla, R.; Oksche, A.; Dehe, M.; Wiesner, B.; Krause, G.; Rosenthal, W. A dileucine sequence and an upstream glutamate residue in the intracellular carboxyl terminus of the vasopressin V2 receptor are essential for cell surface transport in COS.M6 cells. Mol Pharmacol 1998, 54, 525–535. [Google Scholar] [CrossRef]
- Holst, B.; Schwartz, T.W. Molecular mechanism of agonism and inverse agonism in the melanocortin receptors: Zn(2+) as a structural and functional probe. Ann N Y Acad Sci 2003, 994, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Chen, Y.; Dai, A.; Yin, W.; Guo, J.; Yang, D.; Zhou, F.; Jiang, Y.; Wang, M.W.; Xu, H.E. Structural mechanism of calcium-mediated hormone recognition and Gbeta interaction by the human melanocortin-1 receptor. Cell Res 2021, 31, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, S.G.; Horrell, E.M.W.; Christian, P.A.; Vanover, J.C.; Boulanger, M.C.; Zou, Y.; D'Orazio, J.A. PKA-mediated phosphorylation of ATR promotes recruitment of XPA to UV-induced DNA damage. Mol Cell 2014, 54, 999–1011. [Google Scholar] [CrossRef]
- Kadekaro, A.L.; Leachman, S.; Kavanagh, R.J.; Swope, V.; Cassidy, P.; Supp, D.; Sartor, M.; Schwemberger, S.; Babcock, G.; Wakamatsu, K.; et al. Melanocortin 1 receptor genotype: an important determinant of the damage response of melanocytes to ultraviolet radiation. FASEB J 2010, 24, 3850–3860. [Google Scholar] [CrossRef]
- Song, X.; Mosby, N.; Yang, J.; Xu, A.; Abdel-Malek, Z.; Kadekaro, A.L. alpha-MSH activates immediate defense responses to UV-induced oxidative stress in human melanocytes. Pigment Cell Melanoma Res 2009, 22, 809–818. [Google Scholar] [CrossRef]
- Kokot, A.; Metze, D.; Mouchet, N.; Galibert, M.D.; Schiller, M.; Luger, T.A.; Bohm, M. Alpha-melanocyte-stimulating hormone counteracts the suppressive effect of UVB on Nrf2 and Nrf-dependent gene expression in human skin. Endocrinology 2009, 150, 3197–3206. [Google Scholar] [CrossRef]
- Abdel-Malek, Z.A.; Swope, V.B.; Starner, R.J.; Koikov, L.; Cassidy, P.; Leachman, S. Melanocortins and the melanocortin 1 receptor, moving translationally towards melanoma prevention. Arch Biochem Biophys 2014, 563, 4–12. [Google Scholar] [CrossRef]
- Suzuki, I.; Tada, A.; Ollmann, M.M.; Barsh, G.S.; Im, S.; Lamoreux, M.L.; Hearing, V.J.; Nordlund, J.J.; Abdel-Malek, Z.A. Agouti signaling protein inhibits melanogenesis and the response of human melanocytes to alpha-melanotropin. J Invest Dermatol 1997, 108, 838–842. [Google Scholar] [CrossRef] [PubMed]
- Dorsam, R.T.; Gutkind, J.S. G-protein-coupled receptors and cancer. Nat Rev Cancer 2007, 7, 79–94. [Google Scholar] [CrossRef]
- Neves, S.R.; Ram, P.T.; Iyengar, R. G protein pathways. Science 2002, 296, 1636–1639. [Google Scholar] [CrossRef] [PubMed]
- Herraiz, C.; Garcia-Borron, J.C.; Jimenez-Cervantes, C.; Olivares, C. MC1R signaling. Intracellular partners and pathophysiological implications. Biochim Biophys Acta Mol Basis Dis 2017, 1863, 2448–2461. [Google Scholar] [CrossRef] [PubMed]
- Bohm, M.; Schulte, U.; Kalden, H.; Luger, T.A. Alpha-melanocyte-stimulating hormone modulates activation of NF-kappa B and AP-1 and secretion of interleukin-8 in human dermal fibroblasts. Ann N Y Acad Sci 1999, 885, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb Perspect Biol 2014, 6, a016295. [Google Scholar] [CrossRef]
- Bonfiglio, V.; Camillieri, G.; Avitabile, T.; Leggio, G.M.; Drago, F. Effects of the COOH-terminal tripeptide alpha-MSH(11-13) on corneal epithelial wound healing: role of nitric oxide. Exp Eye Res 2006, 83, 1366–1372. [Google Scholar] [CrossRef]
- Becher, E.; Mahnke, K.; Brzoska, T.; Kalden, D.H.; Grabbe, S.; Luger, T.A. Human peripheral blood-derived dendritic cells express functional melanocortin receptor MC-1R. Ann N Y Acad Sci 1999, 885, 188–195. [Google Scholar] [CrossRef]
- Scholzen, T.E.; Sunderkotter, C.; Kalden, D.H.; Brzoska, T.; Fastrich, M.; Fisbeck, T.; Armstrong, C.A.; Ansel, J.C.; Luger, T.A. Alpha-melanocyte stimulating hormone prevents lipopolysaccharide-induced vasculitis by down-regulating endothelial cell adhesion molecule expression. Endocrinology 2003, 144, 360–370. [Google Scholar] [CrossRef]
- Kalden, D.H.; Scholzen, T.; Brzoska, T.; Luger, T.A. Mechanisms of the antiinflammatory effects of alpha-MSH. Role of transcription factor NF-kappa B and adhesion molecule expression. Ann N Y Acad Sci 1999, 885, 254–261. [Google Scholar] [CrossRef]
- Zhang, L.; Dong, L.; Liu, X.; Jiang, Y.; Zhang, L.; Zhang, X.; Li, X.; Zhang, Y. alpha-Melanocyte-stimulating hormone protects retinal vascular endothelial cells from oxidative stress and apoptosis in a rat model of diabetes. PLoS One 2014, 9, e93433. [Google Scholar] [CrossRef]
- Galimberti, D.; Baron, P.; Meda, L.; Prat, E.; Scarpini, E.; Delgado, R.; Catania, A.; Lipton, J.M.; Scarlato, G. Alpha-MSH peptides inhibit production of nitric oxide and tumor necrosis factor-alpha by microglial cells activated with beta-amyloid and interferon gamma. Biochem Biophys Res Commun 1999, 263, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Delgado, R.; Carlin, A.; Airaghi, L.; Demitri, M.T.; Meda, L.; Galimberti, D.; Baron, P.; Lipton, J.M.; Catania, A. Melanocortin peptides inhibit production of proinflammatory cytokines and nitric oxide by activated microglia. J Leukoc Biol 1998, 63, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Taherzadeh, S.; Sharma, S.; Chhajlani, V.; Gantz, I.; Rajora, N.; Demitri, M.T.; Kelly, L.; Zhao, H.; Ichiyama, T.; Catania, A.; et al. alpha-MSH and its receptors in regulation of tumor necrosis factor-alpha production by human monocyte/macrophages. Am J Physiol 1999, 276, R1289–1294. [Google Scholar] [CrossRef] [PubMed]
- Ottani, A.; Giuliani, D.; Neri, L.; Calevro, A.; Canalini, F.; Vandini, E.; Cainazzo, M.M.; Ruberto, I.A.; Barbieri, A.; Rossi, R.; et al. NDP-alpha-MSH attenuates heart and liver responses to myocardial reperfusion via the vagus nerve and JAK/ERK/STAT signaling. Eur J Pharmacol 2015, 769, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Buggy, J.J. Binding of alpha-melanocyte-stimulating hormone to its G-protein-coupled receptor on B-lymphocytes activates the Jak/STAT pathway. Biochem J 1998, 331 Pt 1, 211–216. [Google Scholar] [CrossRef]
- Mandrika, I.; Muceniece, R.; Wikberg, J.E. Effects of melanocortin peptides on lipopolysaccharide/interferon-gamma-induced NF-kappaB DNA binding and nitric oxide production in macrophage-like RAW 264.7 cells: evidence for dual mechanisms of action. Biochem Pharmacol 2001, 61, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Gerlo, S.; Kooijman, R.; Beck, I.M.; Kolmus, K.; Spooren, A.; Haegeman, G. Cyclic AMP: a selective modulator of NF-kappaB action. Cell Mol Life Sci 2011, 68, 3823–3841. [Google Scholar] [CrossRef]
- Manna, S.K.; Aggarwal, B.B. Alpha-melanocyte-stimulating hormone inhibits the nuclear transcription factor NF-kappa B activation induced by various inflammatory agents. J Immunol 1998, 161, 2873–2880. [Google Scholar] [CrossRef]
- Wang, W.; Guo, D.Y.; Lin, Y.J.; Tao, Y.X. Melanocortin Regulation of Inflammation. Front Endocrinol (Lausanne) 2019, 10, 683. [Google Scholar] [CrossRef]
- Lo, J.A.; Fisher, D.E. The melanoma revolution: from UV carcinogenesis to a new era in therapeutics. Science 2014, 346, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, A.A.; Slastnikova, T.A.; Durymanov, M.O.; Sobolev, A.S. Malignant melanoma and melanocortin 1 receptor. Biochemistry (Mosc) 2013, 78, 1228–1237. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Onfray, F.; Lopez, M.; Lundqvist, A.; Aguirre, A.; Escobar, A.; Serrano, A.; Korenblit, C.; Petersson, M.; Chhajlani, V.; Larsson, O.; et al. Tissue distribution and differential expression of melanocortin 1 receptor, a malignant melanoma marker. Br J Cancer 2002, 87, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Ogbah, Z.; Puig-Butille, J.A.; Simonetta, F.; Badenas, C.; Cervera, R.; Mila, J.; Benitez, D.; Malvehy, J.; Vilella, R.; Puig, S. Molecular characterization of human cutaneous melanoma-derived cell lines. Anticancer Res 2012, 32, 1245–1251. [Google Scholar] [PubMed]
- Eggermont, A.M. Advances in systemic treatment of melanoma. Ann Oncol 2010, 21 Suppl 7, vii339–344. [Google Scholar] [CrossRef]
- Iriyama, N.; Yoshino, Y.; Yuan, B.; Horikoshi, A.; Hirabayashi, Y.; Hatta, Y.; Toyoda, H.; Takeuchi, J. Speciation of arsenic trioxide metabolites in peripheral blood and bone marrow from an acute promyelocytic leukemia patient. J Hematol Oncol 2012, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Montero-Melendez, T.; Boesen, T.; Jonassen, T.E.N. Translational advances of melanocortin drugs: Integrating biology, chemistry and genetics. Semin Immunol 2022, 59, 101603. [Google Scholar] [CrossRef]
- Chen, S.; Zhu, B.; Yin, C.; Liu, W.; Han, C.; Chen, B.; Liu, T.; Li, X.; Chen, X.; Li, C.; et al. Palmitoylation-dependent activation of MC1R prevents melanomagenesis. Nature 2017, 549, 399–403. [Google Scholar] [CrossRef]
- Castejon-Grinan, M.; Herraiz, C.; Olivares, C.; Jimenez-Cervantes, C.; Garcia-Borron, J.C. cAMP-independent non-pigmentary actions of variant melanocortin 1 receptor: AKT-mediated activation of protective responses to oxidative DNA damage. Oncogene 2018, 37, 3631–3646. [Google Scholar] [CrossRef]
- Denton, C.P.; Ong, V.H. Targeted therapies for systemic sclerosis. Nat Rev Rheumatol 2013, 9, 451–464. [Google Scholar] [CrossRef]
- Allanore, Y.; Simms, R.; Distler, O.; Trojanowska, M.; Pope, J.; Denton, C.P.; Varga, J. Systemic sclerosis. Nat Rev Dis Primers 2015, 1, 15002. [Google Scholar] [CrossRef] [PubMed]
- Khanna, D.; Lin, C.J.F.; Furst, D.E.; Goldin, J.; Kim, G.; Kuwana, M.; Allanore, Y.; Matucci-Cerinic, M.; Distler, O.; Shima, Y.; et al. Tocilizumab in systemic sclerosis: a randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Respir Med 2020, 8, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Distler, O.; Highland, K.B.; Gahlemann, M.; Azuma, A.; Fischer, A.; Mayes, M.D.; Raghu, G.; Sauter, W.; Girard, M.; Alves, M.; et al. Nintedanib for Systemic Sclerosis-Associated Interstitial Lung Disease. N Engl J Med 2019, 380, 2518–2528. [Google Scholar] [CrossRef]
- Kondo, M.; Suzuki, T.; Kawano, Y.; Kojima, S.; Miyashiro, M.; Matsumoto, A.; Kania, G.; Blyszczuk, P.; Ross, R.L.; Mulipa, P.; et al. Dersimelagon, a novel oral melanocortin 1 receptor agonist, demonstrates disease-modifying effects in preclinical models of systemic sclerosis. Arthritis Res Ther 2022, 24, 210. [Google Scholar] [CrossRef]
- Dixon, B.J.; Reis, C.; Ho, W.M.; Tang, J.; Zhang, J.H. Neuroprotective Strategies after Neonatal Hypoxic Ischemic Encephalopathy. Int J Mol Sci 2015, 16, 22368–22401. [Google Scholar] [CrossRef]
- McRae, A.; Gilland, E.; Bona, E.; Hagberg, H. Microglia activation after neonatal hypoxic-ischemia. Brain Res Dev Brain Res 1995, 84, 245–252. [Google Scholar] [CrossRef]
- Wu, X.; Fu, S.; Liu, Y.; Luo, H.; Li, F.; Wang, Y.; Gao, M.; Cheng, Y.; Xie, Z. NDP-MSH binding melanocortin-1 receptor ameliorates neuroinflammation and BBB disruption through CREB/Nr4a1/NF-kappaB pathway after intracerebral hemorrhage in mice. J Neuroinflammation 2019, 16, 192. [Google Scholar] [CrossRef]
- Kang, L.; McIntyre, K.W.; Gillooly, K.M.; Yang, Y.; Haycock, J.; Roberts, S.; Khanna, A.; Herpin, T.F.; Yu, G.; Wu, X.; et al. A selective small molecule agonist of the melanocortin-1 receptor inhibits lipopolysaccharide-induced cytokine accumulation and leukocyte infiltration in mice. J Leukoc Biol 2006, 80, 897–904. [Google Scholar] [CrossRef]
- Yu, S.; Doycheva, D.M.; Gamdzyk, M.; Yang, Y.; Lenahan, C.; Li, G.; Li, D.; Lian, L.; Tang, J.; Lu, J.; et al. Activation of MC1R with BMS-470539 attenuates neuroinflammation via cAMP/PKA/Nurr1 pathway after neonatal hypoxic-ischemic brain injury in rats. J Neuroinflammation 2021, 18, 26. [Google Scholar] [CrossRef]
- Rinne, P.; Rami, M.; Nuutinen, S.; Santovito, D.; van der Vorst, E.P.C.; Guillamat-Prats, R.; Lyytikainen, L.P.; Raitoharju, E.; Oksala, N.; Ring, L.; et al. Melanocortin 1 Receptor Signaling Regulates Cholesterol Transport in Macrophages. Circulation 2017, 136, 83–97. [Google Scholar] [CrossRef]
- Catania, A.; Gatti, S.; Colombo, G.; Lipton, J.M. Targeting melanocortin receptors as a novel strategy to control inflammation. Pharmacol Rev 2004, 56, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Rinne, P.; Silvola, J.M.; Hellberg, S.; Stahle, M.; Liljenback, H.; Salomaki, H.; Koskinen, E.; Nuutinen, S.; Saukko, P.; Knuuti, J.; et al. Pharmacological activation of the melanocortin system limits plaque inflammation and ameliorates vascular dysfunction in atherosclerotic mice. Arterioscler Thromb Vasc Biol 2014, 34, 1346–1354. [Google Scholar] [CrossRef]
- Rinne, P.; Kadiri, J.J.; Velasco-Delgado, M.; Nuutinen, S.; Viitala, M.; Hollmen, M.; Rami, M.; Savontaus, E.; Steffens, S. Melanocortin 1 Receptor Deficiency Promotes Atherosclerosis in Apolipoprotein E(-/-) Mice. Arterioscler Thromb Vasc Biol 2018, 38, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Moscowitz, A.E.; Asif, H.; Lindenmaier, L.B.; Calzadilla, A.; Zhang, C.; Mirsaeidi, M. The Importance of Melanocortin Receptors and Their Agonists in Pulmonary Disease. Front Med (Lausanne) 2019, 6, 145. [Google Scholar] [CrossRef] [PubMed]
- Spana, C.; Taylor, A.W.; Yee, D.G.; Makhlina, M.; Yang, W.; Dodd, J. Probing the Role of Melanocortin Type 1 Receptor Agonists in Diverse Immunological Diseases. Front Pharmacol 2018, 9, 1535. [Google Scholar] [CrossRef]
- Cai, W.; Srivastava, P.; Feng, D.; Lin, Y.; Vanderburg, C.R.; Xu, Y.; McLean, P.; Frosch, M.P.; Fisher, D.E.; Schwarzschild, M.A.; et al. Melanocortin 1 receptor activation protects against alpha-synuclein pathologies in models of Parkinson's disease. Mol Neurodegener 2022, 17, 16. [Google Scholar] [CrossRef]
- Chakraborty, A.K.; Funasaka, Y.; Slominski, A.; Ermak, G.; Hwang, J.; Pawelek, J.M.; Ichihashi, M. Production and release of proopiomelanocortin (POMC) derived peptides by human melanocytes and keratinocytes in culture: regulation by ultraviolet B. Biochim Biophys Acta 1996, 1313, 130–138. [Google Scholar] [CrossRef]
- Schauer, E.; Trautinger, F.; Kock, A.; Schwarz, A.; Bhardwaj, R.; Simon, M.; Ansel, J.C.; Schwarz, T.; Luger, T.A. Proopiomelanocortin-derived peptides are synthesized and released by human keratinocytes. J Clin Invest 1994, 93, 2258–2262. [Google Scholar] [CrossRef]
- Abdel-Malek, Z.; Scott, M.C.; Suzuki, I.; Tada, A.; Im, S.; Lamoreux, L.; Ito, S.; Barsh, G.; Hearing, V.J. The melanocortin-1 receptor is a key regulator of human cutaneous pigmentation. Pigment Cell Res 2000, 13 Suppl 8, 156–162. [Google Scholar] [CrossRef]
- Ollmann, M.M.; Lamoreux, M.L.; Wilson, B.D.; Barsh, G.S. Interaction of Agouti protein with the melanocortin 1 receptor in vitro and in vivo. Genes Dev 1998, 12, 316–330. [Google Scholar] [CrossRef]
- Blanchard, S.G.; Harris, C.O.; Ittoop, O.R.; Nichols, J.S.; Parks, D.J.; Truesdale, A.T.; Wilkison, W.O. Agouti antagonism of melanocortin binding and action in the B16F10 murine melanoma cell line. Biochemistry 1995, 34, 10406–10411. [Google Scholar] [CrossRef] [PubMed]
- Candille, S.I.; Kaelin, C.B.; Cattanach, B.M.; Yu, B.; Thompson, D.A.; Nix, M.A.; Kerns, J.A.; Schmutz, S.M.; Millhauser, G.L.; Barsh, G.S. A -defensin mutation causes black coat color in domestic dogs. Science 2007, 318, 1418–1423. [Google Scholar] [CrossRef] [PubMed]
- Harder, J.; Bartels, J.; Christophers, E.; Schroder, J.M. Isolation and characterization of human beta -defensin-3, a novel human inducible peptide antibiotic. J Biol Chem 2001, 276, 5707–5713. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.D.; Ollmann, M.M.; Kang, L.; Stoffel, M.; Bell, G.I.; Barsh, G.S. Structure and function of ASP, the human homolog of the mouse agouti gene. Hum Mol Genet 1995, 4, 223–230. [Google Scholar] [CrossRef]
- Lu, D.; Willard, D.; Patel, I.R.; Kadwell, S.; Overton, L.; Kost, T.; Luther, M.; Chen, W.; Woychik, R.P.; Wilkison, W.O.; et al. Agouti protein is an antagonist of the melanocyte-stimulating-hormone receptor. Nature 1994, 371, 799–802. [Google Scholar] [CrossRef]
- Nix, M.A.; Kaelin, C.B.; Ta, T.; Weis, A.; Morton, G.J.; Barsh, G.S.; Millhauser, G.L. Molecular and functional analysis of human beta-defensin 3 action at melanocortin receptors. Chem Biol 2013, 20, 784–795. [Google Scholar] [CrossRef]
- Swope, V.B.; Jameson, J.A.; McFarland, K.L.; Supp, D.M.; Miller, W.E.; McGraw, D.W.; Patel, M.A.; Nix, M.A.; Millhauser, G.L.; Babcock, G.F.; et al. Defining MC1R regulation in human melanocytes by its agonist alpha-melanocortin and antagonists agouti signaling protein and beta-defensin 3. J Invest Dermatol 2012, 132, 2255–2262. [Google Scholar] [CrossRef]
- Harris, J.I.; Lerner, A.B. Amino-acid sequence of the alpha-melanocyte-stimulating hormone. Nature 1957, 179, 1346–1347. [Google Scholar] [CrossRef]
- Hruby, V.J.; Wilkes, B.C.; Hadley, M.E.; Al-Obeidi, F.; Sawyer, T.K.; Staples, D.J.; de Vaux, A.E.; Dym, O.; Castrucci, A.M.; Hintz, M.F.; et al. alpha-Melanotropin: the minimal active sequence in the frog skin bioassay. J Med Chem 1987, 30, 2126–2130. [Google Scholar] [CrossRef]
- Sawyer, T.K.; Sanfilippo, P.J.; Hruby, V.J.; Engel, M.H.; Heward, C.B.; Burnett, J.B.; Hadley, M.E. 4-Norleucine, 7-D-phenylalanine-alpha-melanocyte-stimulating hormone: a highly potent alpha-melanotropin with ultralong biological activity. Proc Natl Acad Sci U S A 1980, 77, 5754–5758. [Google Scholar] [CrossRef]
- Hadley, M.E.; Marwan, M.M.; al-Obeidi, F.; Hruby, V.J.; Castrucci, A.M. Linear and cyclic alpha-melanotropin [4-10]-fragment analogues that exhibit superpotency and residual activity. Pigment Cell Res 1989, 2, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Al-Obeidi, F.; Castrucci, A.M.; Hadley, M.E.; Hruby, V.J. Potent and prolonged acting cyclic lactam analogues of alpha-melanotropin: design based on molecular dynamics. J Med Chem 1989, 32, 2555–2561. [Google Scholar] [CrossRef] [PubMed]
- Hunt, G.; Todd, C.; Cresswell, J.E.; Thody, A.J. Alpha-melanocyte stimulating hormone and its analogue Nle4DPhe7 alpha-MSH affect morphology, tyrosinase activity and melanogenesis in cultured human melanocytes. J Cell Sci 1994, 107 ( Pt 1) Pt 1, 205–211. [Google Scholar] [CrossRef]
- Hunt, G.; Kyne, S.; Ito, S.; Wakamatsu, K.; Todd, C.; Thody, A. Eumelanin and phaeomelanin contents of human epidermis and cultured melanocytes. Pigment Cell Res 1995, 8, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Dorr, R.T.; Dvorakova, K.; Brooks, C.; Lines, R.; Levine, N.; Schram, K.; Miketova, P.; Hruby, V.; Alberts, D.S. Increased eumelanin expression and tanning is induced by a superpotent melanotropin [Nle4-D-Phe7]-alpha-MSH in humans. Photochem Photobiol 2000, 72, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Langendonk, J.G.; Balwani, M.; Anderson, K.E.; Bonkovsky, H.L.; Anstey, A.V.; Bissell, D.M.; Bloomer, J.; Edwards, C.; Neumann, N.J.; Parker, C.; et al. Afamelanotide for Erythropoietic Protoporphyria. N Engl J Med 2015, 373, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.M.; McKay, J.T.; Bonkovsky, H.L. Advances in the management of erythropoietic protoporphyria - role of afamelanotide. Appl Clin Genet 2016, 9, 179–189. [Google Scholar] [CrossRef]
- Hadley, M.E.; Hruby, V.J.; Blanchard, J.; Dorr, R.T.; Levine, N.; Dawson, B.V.; al-Obeidi, F.; Sawyer, T.K. Discovery and development of novel melanogenic drugs. Melanotan-I and -II. Pharm Biotechnol 1998, 11, 575–595. [Google Scholar] [CrossRef]
- Wessells, H.; Hruby, V.J.; Hackett, J.; Han, G.; Balse-Srinivasan, P.; Vanderah, T.W. Ac-Nle-c[Asp-His-DPhe-Arg-Trp-Lys]-NH2 induces penile erection via brain and spinal melanocortin receptors. Neuroscience 2003, 118, 755–762. [Google Scholar] [CrossRef]
- Wessells, H.; Levine, N.; Hadley, M.E.; Dorr, R.; Hruby, V. Melanocortin receptor agonists, penile erection, and sexual motivation: human studies with Melanotan II. Int J Impot Res 2000, 12 Suppl 4, S74–79. [Google Scholar] [CrossRef]
- Rossler, A.S.; Pfaus, J.G.; Kia, H.K.; Bernabe, J.; Alexandre, L.; Giuliano, F. The melanocortin agonist, melanotan II, enhances proceptive sexual behaviors in the female rat. Pharmacol Biochem Behav 2006, 85, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Diamond, L.E.; Earle, D.C.; Garcia, W.D.; Spana, C. Co-administration of low doses of intranasal PT-141, a melanocortin receptor agonist, and sildenafil to men with erectile dysfunction results in an enhanced erectile response. Urology 2005, 65, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Shadiack, A.M.; Sharma, S.D.; Earle, D.C.; Spana, C.; Hallam, T.J. Melanocortins in the treatment of male and female sexual dysfunction. Curr Top Med Chem 2007, 7, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Hugo, W.; Zaretsky, J.M.; Sun, L.; Song, C.; Moreno, B.H.; Hu-Lieskovan, S.; Berent-Maoz, B.; Pang, J.; Chmielowski, B.; Cherry, G.; et al. Genomic and Transcriptomic Features of Response to Anti-PD-1 Therapy in Metastatic Melanoma. Cell 2016, 165, 35–44. [Google Scholar] [CrossRef]
- Zhou, Y.; Mowlazadeh Haghighi, S.; Liu, Z.; Wang, L.; Hruby, V.J.; Cai, M. Development of Ligand-Drug Conjugates Targeting Melanoma through the Overexpressed Melanocortin 1 Receptor. ACS Pharmacol Transl Sci 2020, 3, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Minder, E.I. Afamelanotide, an agonistic analog of alpha-melanocyte-stimulating hormone, in dermal phototoxicity of erythropoietic protoporphyria. Expert Opin Investig Drugs 2010, 19, 1591–1602. [Google Scholar] [CrossRef] [PubMed]
- Haskell-Luevano, C.; Rosenquist, A.; Souers, A.; Khong, K.C.; Ellman, J.A.; Cone, R.D. Compounds that activate the mouse melanocortin-1 receptor identified by screening a small molecule library based upon the beta-turn. J Med Chem 1999, 42, 4380–4387. [Google Scholar] [CrossRef]
- Herpin, T.F.; Yu, G.; Carlson, K.E.; Morton, G.C.; Wu, X.; Kang, L.; Tuerdi, H.; Khanna, A.; Tokarski, J.S.; Lawrence, R.M.; et al. Discovery of tyrosine-based potent and selective melanocortin-1 receptor small-molecule agonists with anti-inflammatory properties. J Med Chem 2003, 46, 1123–1126. [Google Scholar] [CrossRef]
- Boman, A., T.E.N. Jonassen, and T. Lundstedt, Phenyl pyrrole aminoguanidine derivatives. 2007, WO2007141343.
- Bouix-Peter, C., Oxazetidine derivatives, process for preparing them and use in human medicine and in cosmetics. 2013, WO2013001030.
- Eisinger, M.; Li, W.H.; Anthonavage, M.; Pappas, A.; Zhang, L.; Rossetti, D.; Huang, Q.; Seiberg, M. A melanocortin receptor 1 and 5 antagonist inhibits sebaceous gland differentiation and the production of sebum-specific lipids. J Dermatol Sci 2011, 63, 23–32. [Google Scholar] [CrossRef]
- Lee, S.; Ju, W.; Tin, T.D.; Kim, J.; Lee, J.S.; Park, C.H.; Kwak, S.H. Effect of BMS-470539 on lipopolysaccharide-induced neutrophil activation. Korean J Anesthesiol 2020, 73, 151–157. [Google Scholar] [CrossRef]
- Suzuki, T.; Kawano, Y.; Matsumoto, A.; Kondo, M.; Funayama, K.; Tanemura, S.; Miyashiro, M.; Nishi, A.; Yamada, K.; Tsuda, M.; et al. Melanogenic effect of dersimelagon (MT-7117), a novel oral melanocortin 1 receptor agonist. Skin Health Dis 2022, 2, e78. [Google Scholar] [CrossRef]
- Lundstedt, T., et al., N-phenylpyrrole guanidine derivatives as melanocortin receptor ligands. 2008, US7442807.
- Montero-Melendez, T.; Gobbetti, T.; Cooray, S.N.; Jonassen, T.E.; Perretti, M. Biased agonism as a novel strategy to harness the proresolving properties of melanocortin receptors without eliciting melanogenic effects. J Immunol 2015, 194, 3381–3388. [Google Scholar] [CrossRef]
- Boiteau, J.G.; Rodeville, N.; Martin, C.; Tabet, S.; Moureou, C.; Muller, F.; Jetha, J.C.; Cardinaud, I. Development of a Kilogram-Scale Synthesis of a Novel MC1R Agonist. Organic Process Research & Development 2015, 19, 646–651. [Google Scholar] [CrossRef]
- Zhang, L.; Li, W.H.; Anthonavage, M.; Eisinger, M. Melanocortin-5 receptor: a marker of human sebocyte differentiation. Peptides 2006, 27, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Rosenfield, R.L.; Wu, P.P.; Ciletti, N. Sebaceous epithelial cell differentiation requires cyclic adenosine monophosphate generation. In Vitro Cell Dev Biol Anim 2002, 38, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Anthonavage, M.; Huang, Q.; Li, W.H.; Eisinger, M. Proopiomelanocortin peptides and sebogenesis. Ann N Y Acad Sci 2003, 994, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C. Acne and sebaceous gland function. Clin Dermatol 2004, 22, 360–366. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Schagen, S.; Alestas, T. The sebocyte culture: a model to study the pathophysiology of the sebaceous gland in sebostasis, seborrhoea and acne. Arch Dermatol Res 2008, 300, 397–413. [Google Scholar] [CrossRef]
- Thiboutot, D.; Gollnick, H.; Bettoli, V.; Dreno, B.; Kang, S.; Leyden, J.J.; Shalita, A.R.; Lozada, V.T.; Berson, D.; Finlay, A.; et al. New insights into the management of acne: an update from the Global Alliance to Improve Outcomes in Acne group. J Am Acad Dermatol 2009, 60, S1–50. [Google Scholar] [CrossRef]
Figure 1.
Signaling pathway of MC1R in melanocytes.
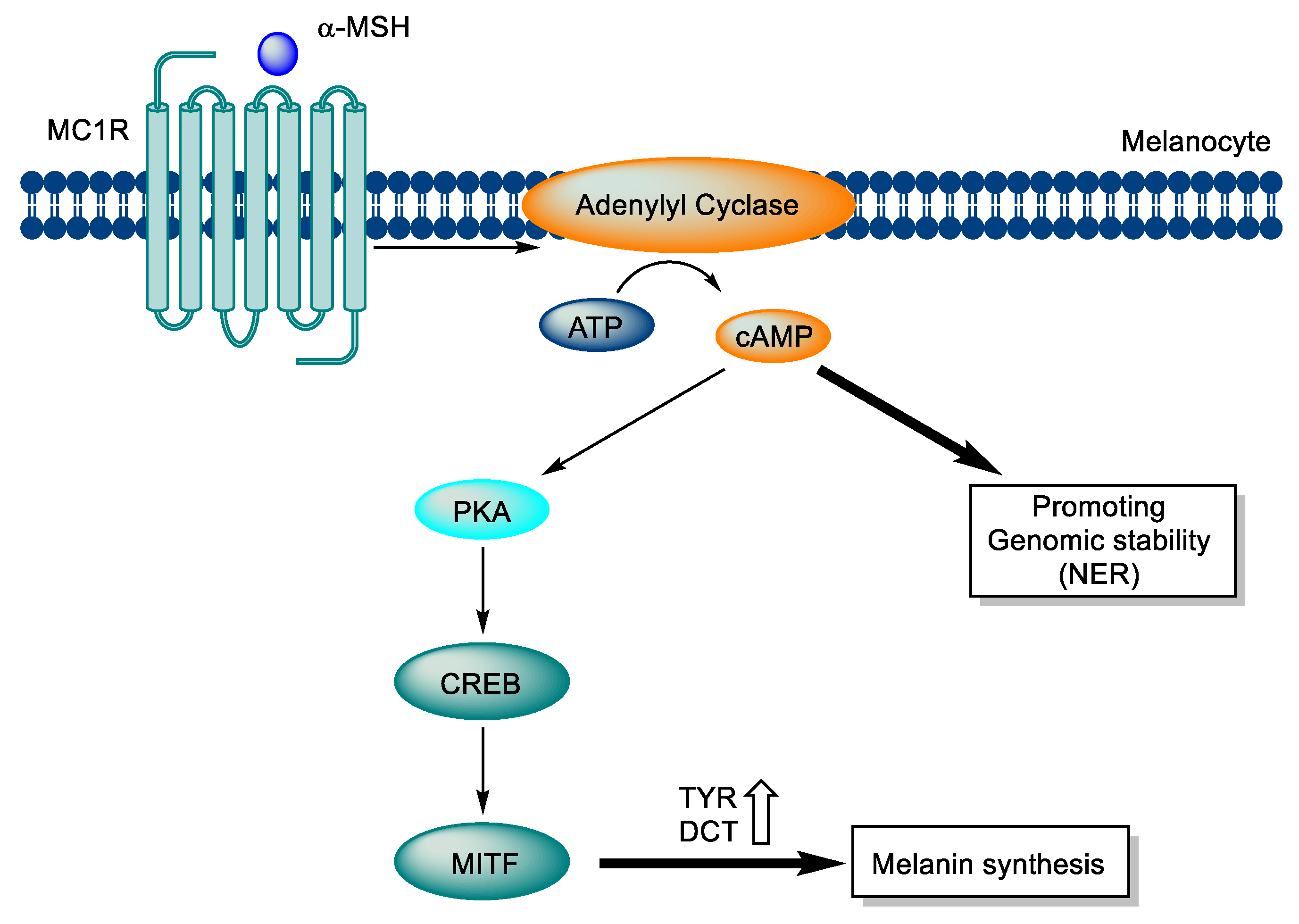
Figure 2.
The role of MC1R signaling in anti-inflammatory effects.
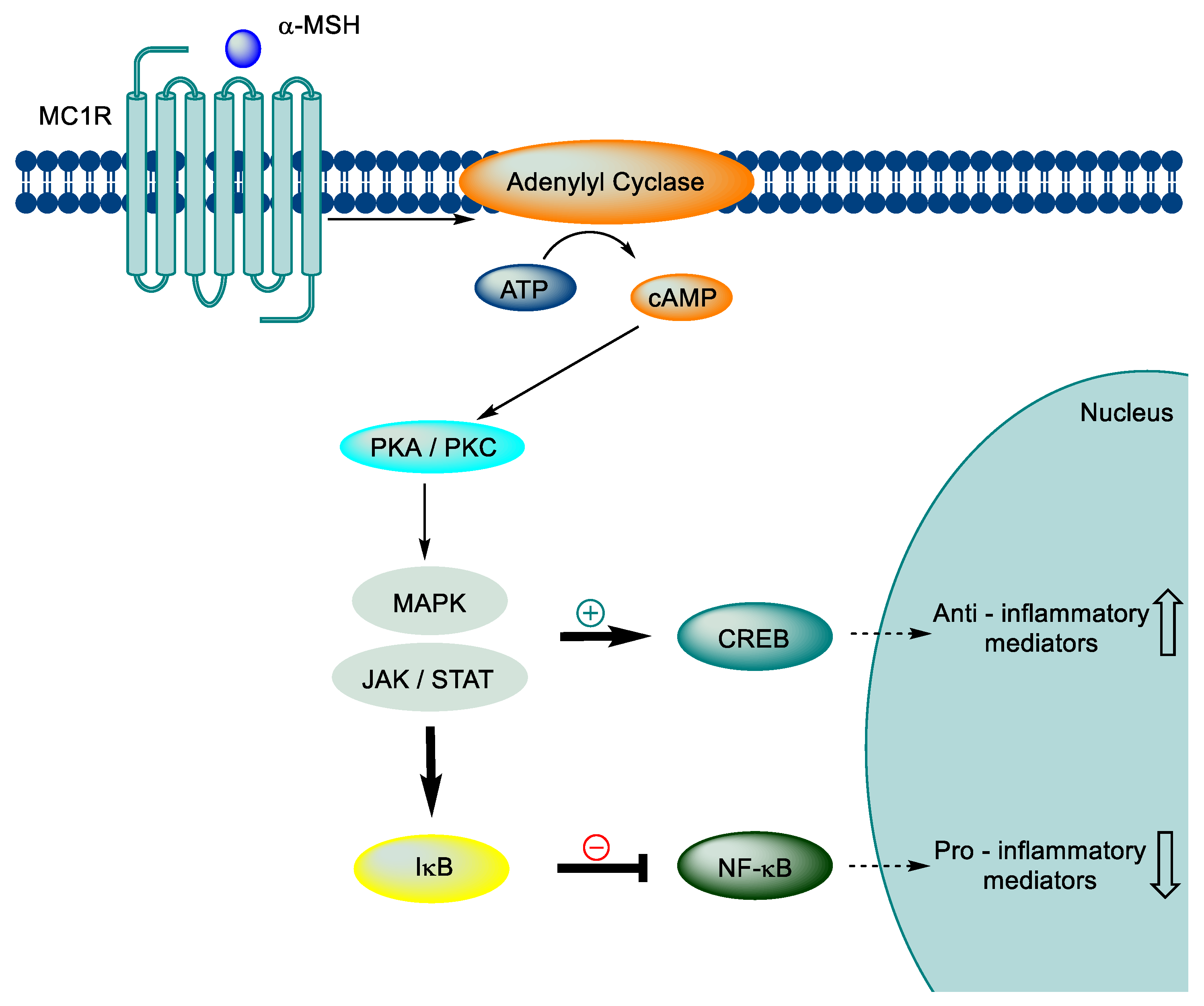
Figure 3.
Structure of α-MSH.
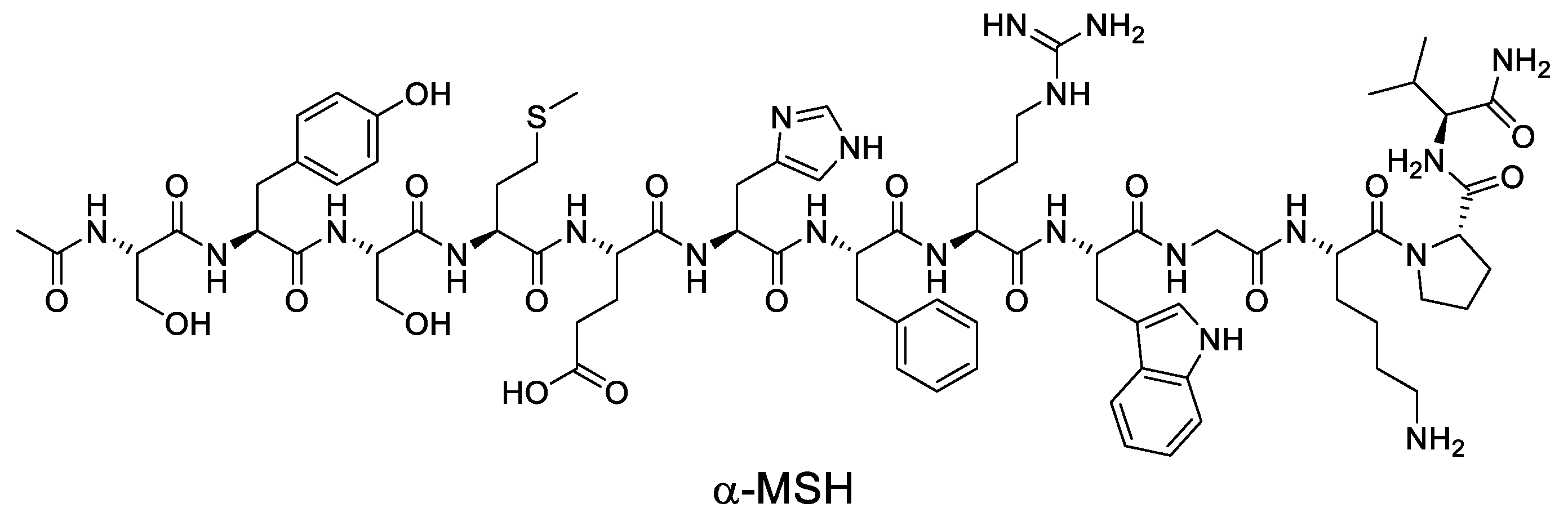
Figure 4.
Structure of Melanotan I.
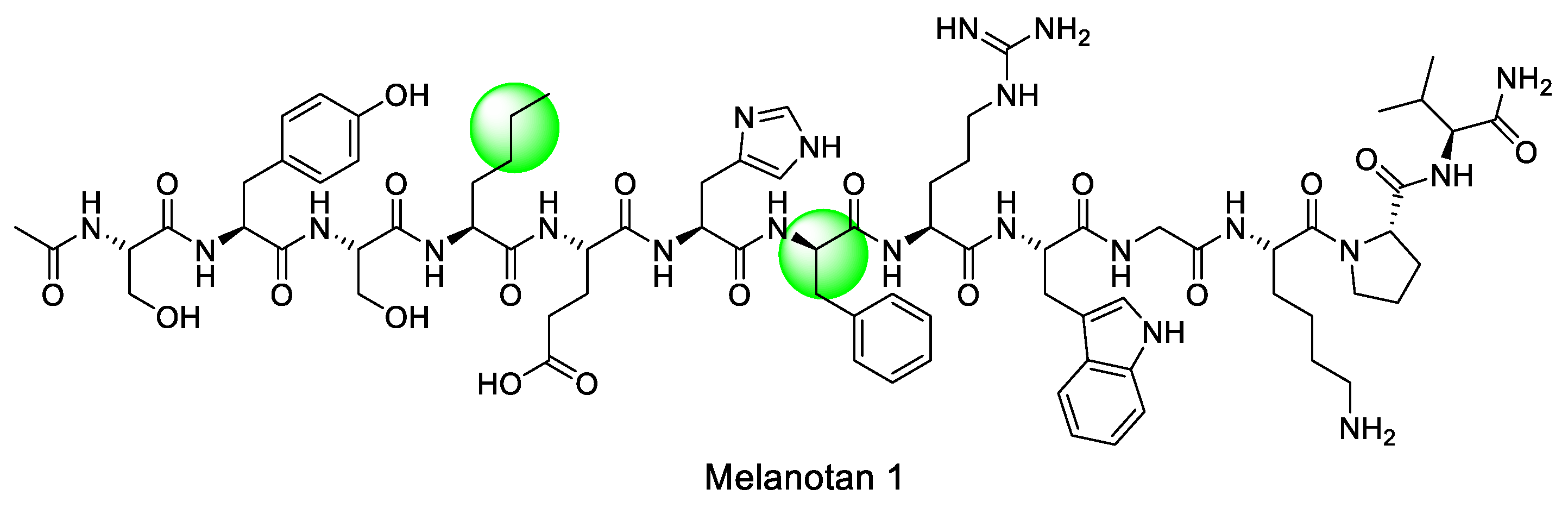
Figure 5.
Interactions of Afamelanotide (Melanotan I) with MC1R domains.
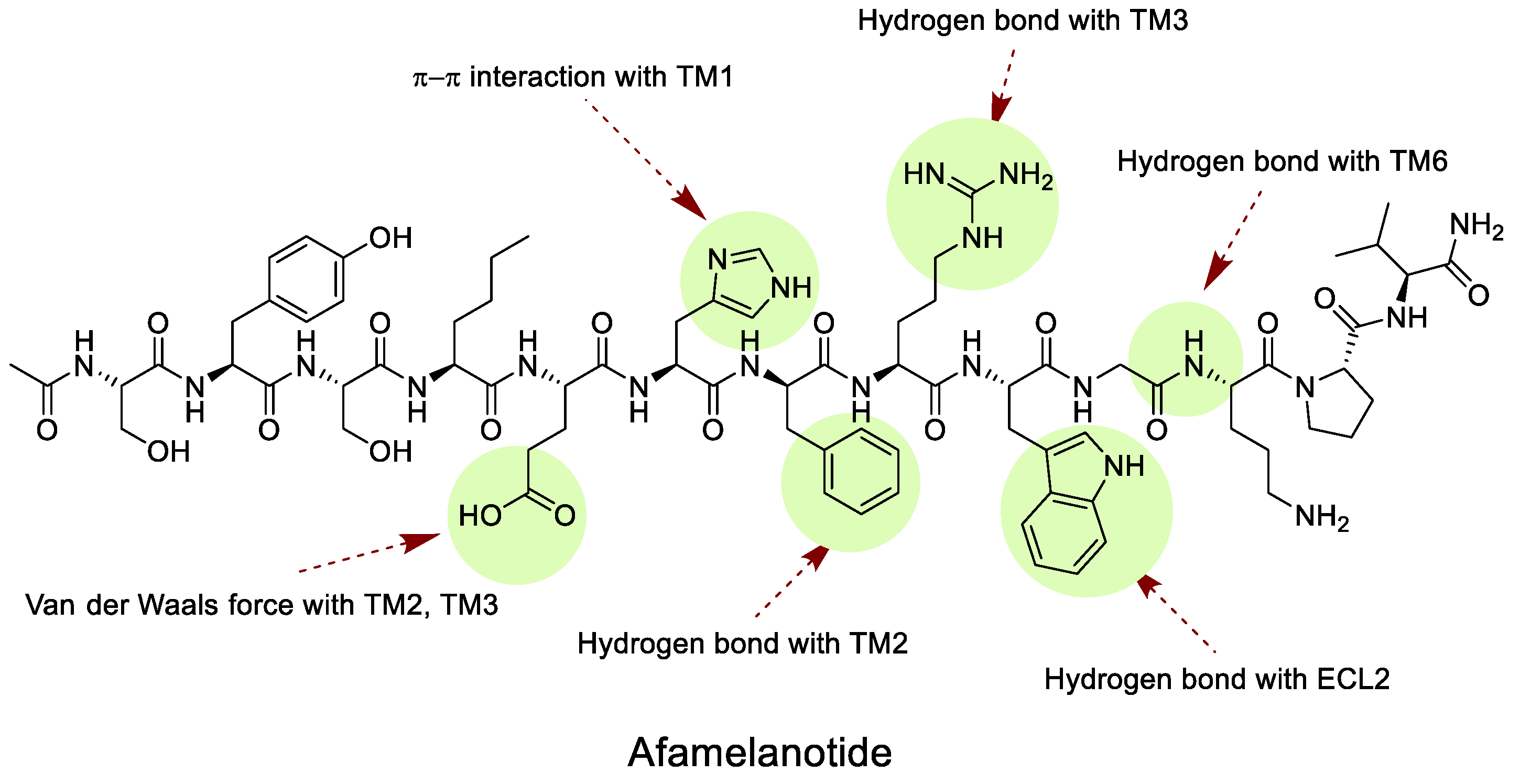
Figure 6.
Structure of Melanotan II and bremelanotide.
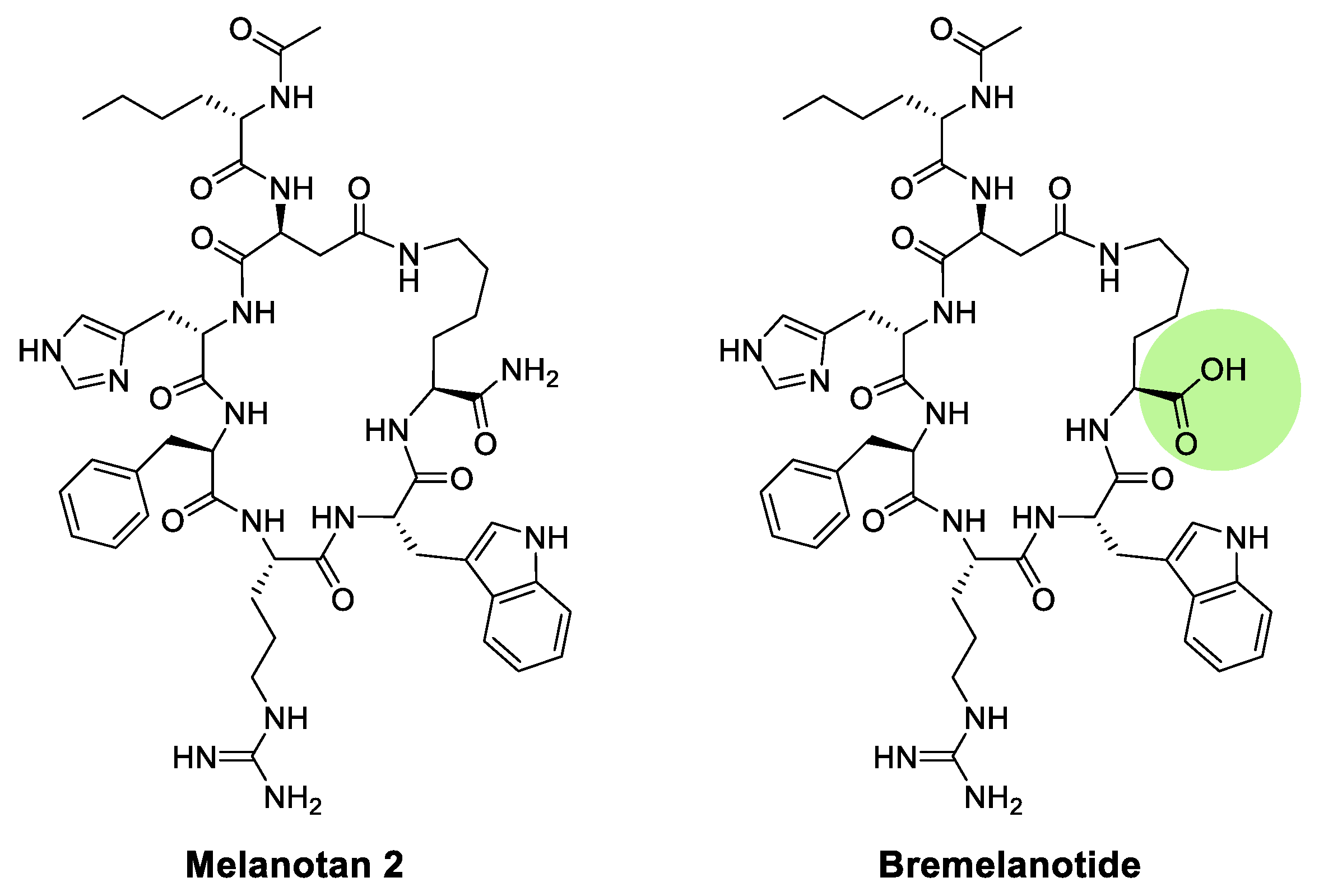
Figure 7.
Structure of Camptothecin-MT-II conjugate.
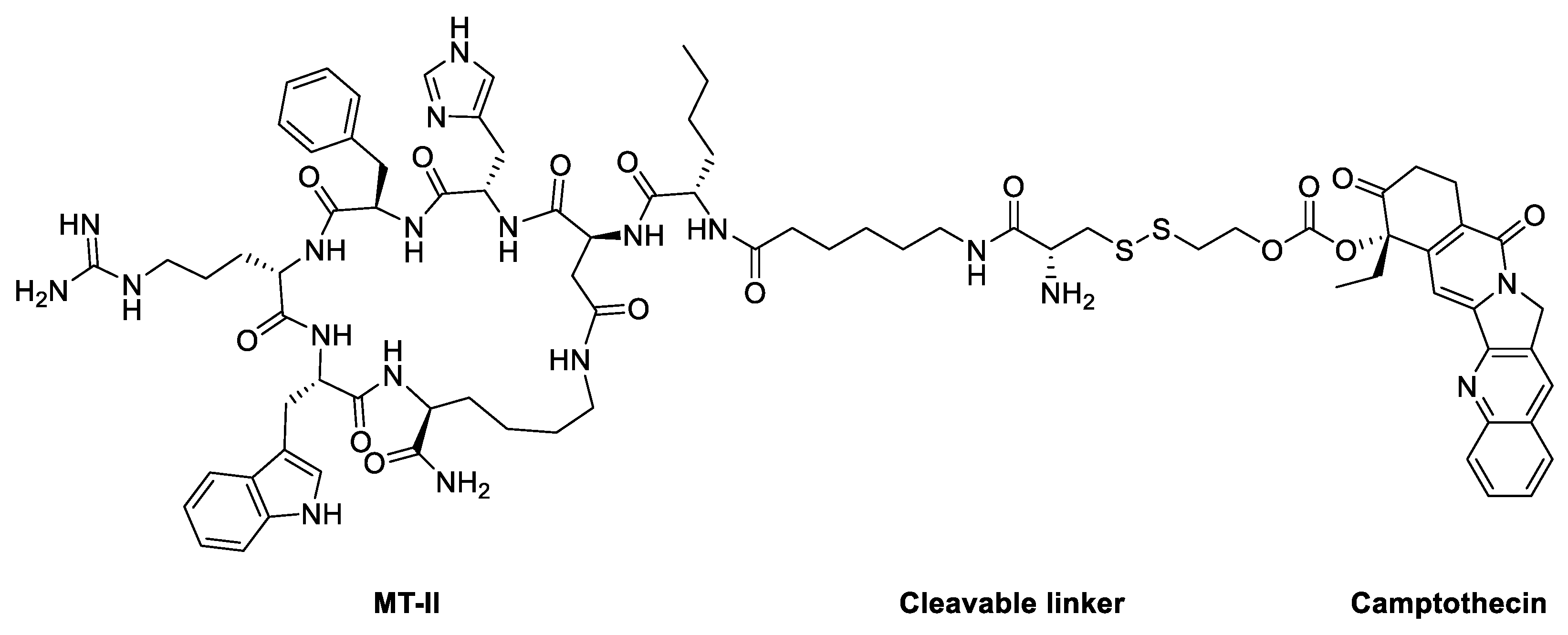
Figure 8.
Structure of BMS-470539 and MC4R agonist.
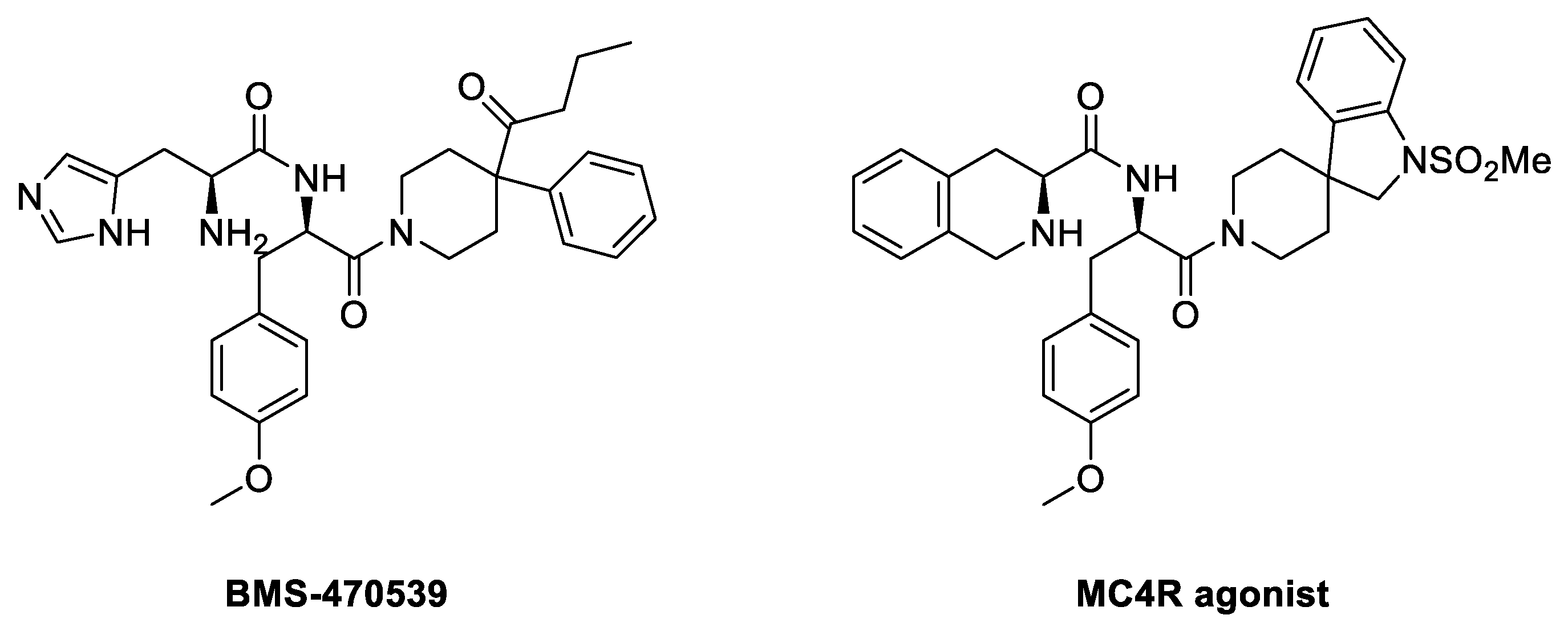
Figure 9.
Binding affinities (Ki) of MT-7117 (Dersimelagon).
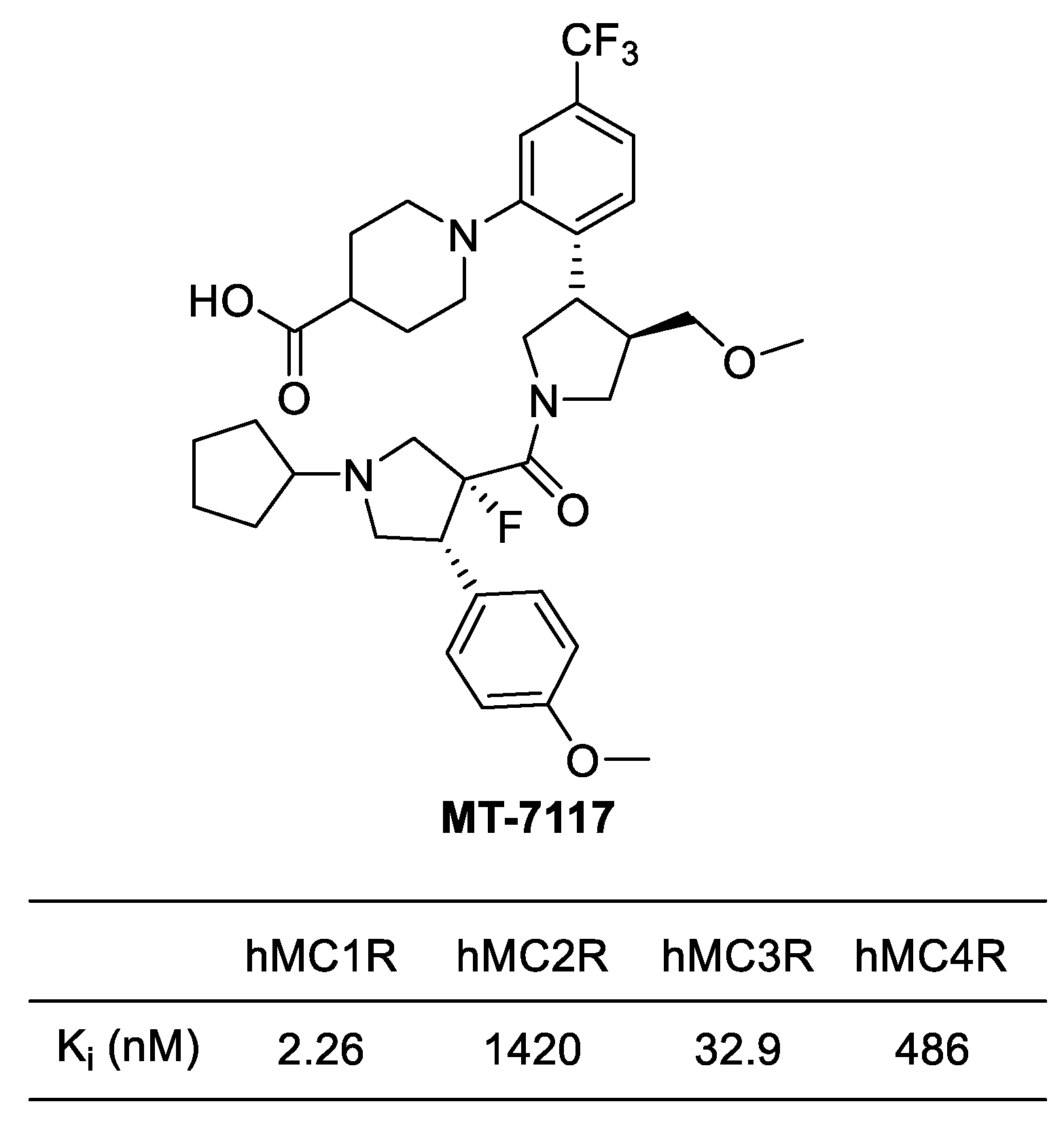
Figure 10.
Structure and Binding affinity of AS1189.
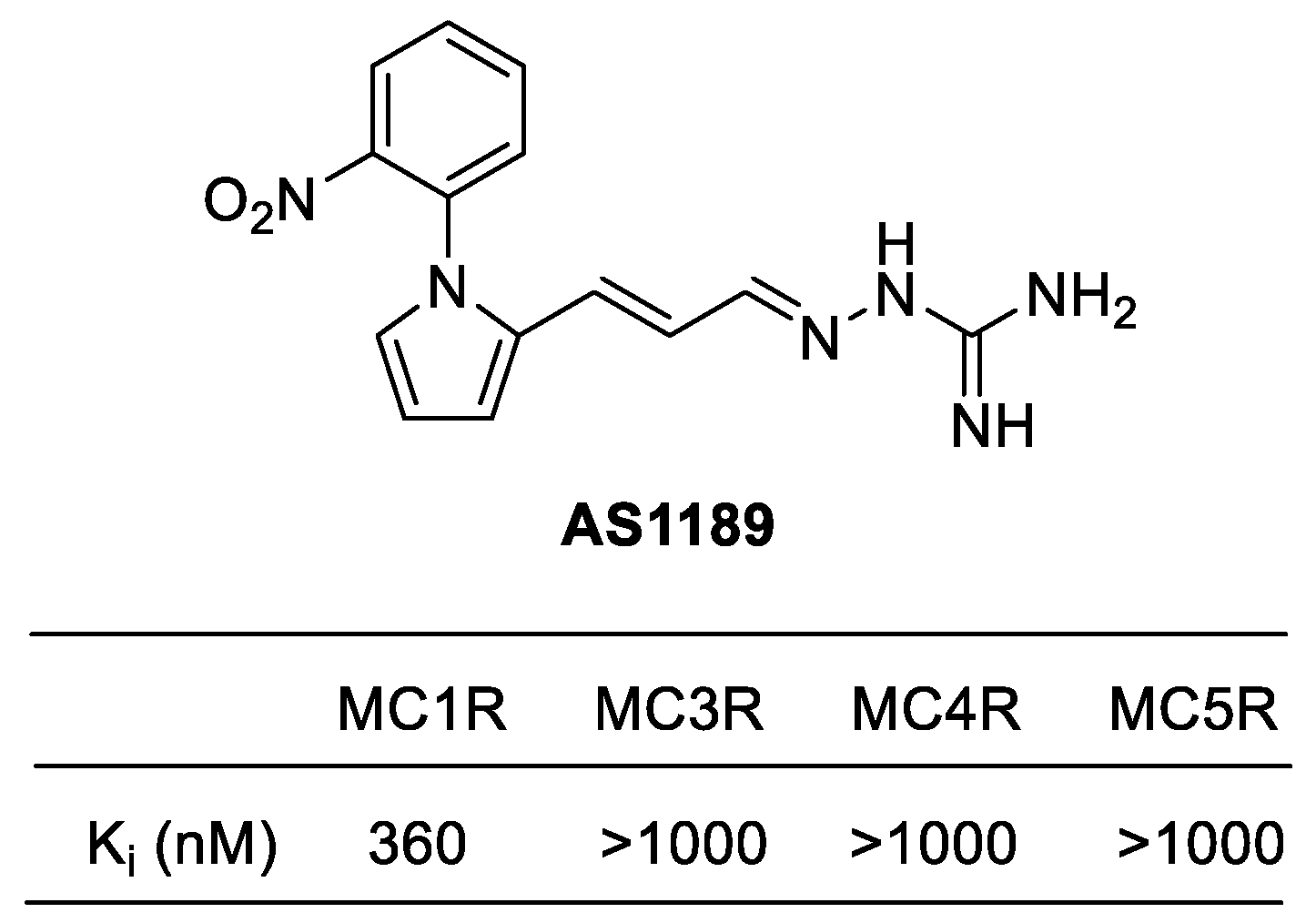
Figure 11.
Structure of CD08108.
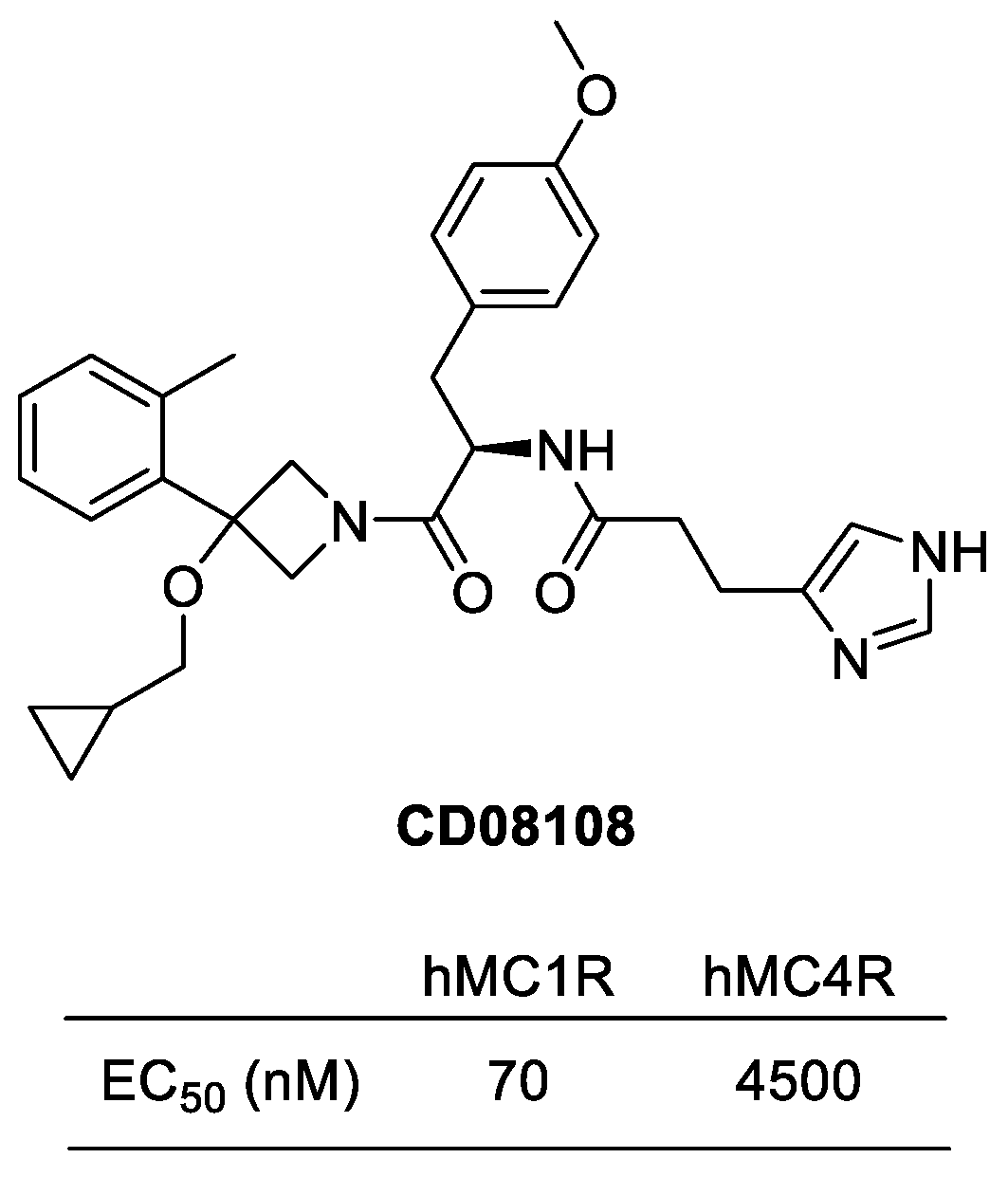
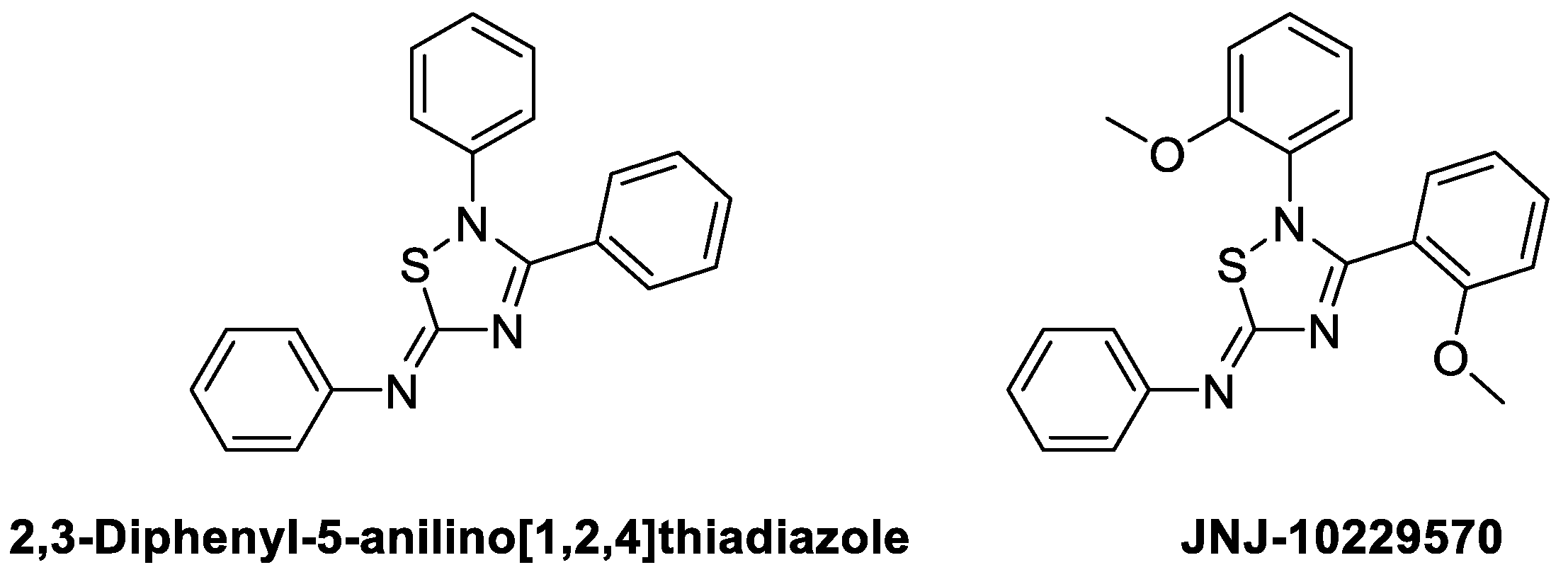

Table 1.
SAR of side chain of BMS-470539.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



