
Submitted:
22 May 2023
Posted:
23 May 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Rapid emergence of evasive SARS-CoV-2 variants is an on-going challenge for COVID-19 vaccinology. Traditional virus neutralization tests provide detailed datasets of neutralization titers against the viral variants. Such datasets are difficult to interpret and do not immediately inform on the sufficiency of the breadth of antibody response. Some of these issues could be tackled using the antigenic cartography approach. In this study, we created antigenic maps using neutralization titers of sera from donors who received Sputnik V booster vaccine after primary Sputnik V vaccination and compared them with the antigenic maps based on serum neutralization titers of Comirnaty-boosted donors. Traditional analysis of neutralization titers against WT, Alpha, Beta, Delta, Omicron BA.1 and BA.4/BA.5 showed a significant booster humoral response after both homologous (Sputnik V) and heterologous (Comirnaty) revaccinations against all the studied viral variants. However, despite this, a more in-depth analysis using antigenic cartography revealed that Omicron variants remain antigenically distant from WT, which is indicative of the formation of insufficient levels of cross-neutralizing antibodies. The implications of these findings may be significant when developing a new vaccine regimen.
Keywords:
SARS-CoV-2
; COVID-19
; revaccination
; virus-neutralization
; antigenic cartography
1. Introduction
The rapid evolution of SARS-CoV-2 has resulted in a range of antigenic variants. The currently dominant Omicron lineage has over 30 substitutions in its Spike (S) protein and features an increased transmissibility allowing the virus to evade neutralizing antibody responses. This makes a waning immunity against the emerging viral variants a significant challenge in COVID-19 vaccinology.
To date, over 50 COVID-19 vaccines have been approved (https://covid19.trackvaccines.org/). Most of them are based on the Spike sequences derived from the original, ancestral SARS-CoV-2 strain. Some vaccines have been quickly adapted for producing a Spike protein from the Omicron subvariants, such as BNT162b2 Bivalent, mRNA-1273.214, and -222. However, developing and approving these vaccines for widespread use is slower than the new viral variants emerge. Evaluating virus-neutralizing antibody (NAb) responses is crucial for understanding the existing vaccines' efficacy against immune-evasive strains of the virus.
Various assays have been developed for determining virus-neutralization titers (VNTs) of serum samples in laboratories [1,2]. These assays provide extensive but rather fragmented data which are difficult to visualize and interpret when considering multiple variants of concern (VOCs). A more in-depth analysis also requires quantifying the breadth of the antibody response (recognizing several antigenically distinct mutant variants). To address these issues, the method of antigenic cartography has proven itself well. This approach is based on multidimensional scaling [3], simplifies data visualization and quantifies parameters such as antigenic distances and breadth of virus neutralization.
Initially, antigenic maps were created for SARS-CoV-2 using sera from donors who had recovered from COVID-19 with PCR-confirmed SARS-CoV-2 variant as well as from animals immunized with known viral variants [4,5,6,7]. Subsequently, it was used to evaluate the effect of mRNA revaccination. This method is particularly useful for monitoring the antibody maturation process after multiple vaccine doses [8,9,10]. Shortening of antigenic distances between ancestral and mutant variants after antigen reexposure indicates the emergence of cross-reactive serum antibodies [8]. So far, limited data are available for adenovirus-based COVID-19 vaccines [5,11], and none for other platforms.
In this study, we compared antigenic maps before and after homologous Sputnik V revaccination (Sputnik Light, which uses the first, Ad26-based, dose, of a two-component GAM-COVID-Vac), including the individuals with hybrid immunity, and after the heterologous Sputnik V/Comirnaty (BNT162b2, BioNTech/Pfizer) revaccination.
2. Results
2.1. Study design
Our study included 58 individuals who were primed with two doses of Sputnik V vaccine in 2021. After six months, 46 individuals received Sputnik V as a booster vaccine while 12 received Comirnaty. Donors recruited to the homologous arm of the study received only the first dose of Spuntik V (Ad26-based) as a booster. In this group, 39 individuals did not have COVID-19 at any time throughout the study (negative for Nucleocapsid-specific antibodies, subgroup “Non-exposed donors, Sputnik V booster”), while 7 individuals had a hybrid immunity due to a prior infection (subgroup “Convalescent donors, Sputnik V booster”). The “Convalescent” subgroup was determined based on three parameters: clinical data, the levels of anti-Nucleocapsid IgG antibodies, and the level of anti-S IgG antibodies before revaccination (Table 1).
The study measured the neutralizing antibody titers in sera collected immediately before and one month after the booster using a panel of 6 pseudoviruses: WT, Alpha, Beta, Delta, Omicron BA.1 and BA.4/BA.5.
2.2. The level of post-boost SARS-CoV-2 NAbs depends on the type of booster vaccine and the history of exposure to SARS-CoV-2
Pre-boost levels of NAbs against WT, Alpha, Beta, Delta, Omicron BA.1, BA.4/BA.5 were above the baseline in 97%, 100%, 97%, 100%, 82%, 87% of virus non-exposed donors (Sputnik V booster), respectively (Figure 1A). Thus, the majority of Sputnik-V vaccinated donors maintain high levels NAbs for 6 months after the primary vaccination. Only one sample at the post-boost timepoint remained on the baseline for the neutralization of BA.4/BA.5, while the rest of the samples showed the titers significantly above the baseline (Figure 1B).
Sera from non-exposed donors revaccinated with (A-C) Sputnik V or (D-F) Comirnaty. Sera from (G-I) convalescent donors revaccinated with Sputnik V. Asterisks indicate significant difference when comparing ID50 values against WT with variants. Connecting lines indicate serum from the same individual. (C, F, I) Summary of virus-neutralizing activities of serum samples against the WT and variants before and after Sputnik/Comirnaty boost. Asterisks indicate significant difference when comparing ID50 values before and after boost for each variant. Medians ± IQR of ID50 values are shown. Statistical analysis was performed using the Friedman test and Dunn's posthoc test, *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns = not significant.
We observed that pre-boost neutralization titers against Beta, Delta, and Omicron variants were significantly lower than those against WT (p < 0.05 for Beta and Delta, p < 0.0001 for Omicron variants). The activity of NAbs present in the post-boost sera was comparable to the pre-boost sera (p < 0.01, p < 0.001, p < 0.0001 for Beta, Delta, and Omicron variants respectively).
Next, we examined serum samples from naïve donors who received a heterologous Comirnaty booster (Figure 1D,E). 92% of pre-boost serum samples exceeded the neutralization titer baseline against WT, Alpha, Beta, Delta and only 58% and 83% exceeded the baseline against BA.1, BA.4/BA.5 respectively (Figure 1D). After Comirnaty boost, all VNT values were above the baseline (Figure 1E).
Both Beta and Omicron variants demonstrated significantly lower neutralization than WT (p < 0.01, p < 0.0001, p < 0.001 for Beta, BA.1, and BA.4/BA.5 respectively) in pre-boost sera (Figure 1D). Following the heterologous booster, serum antibodies had lower neutralizing activity against Beta, Omicron variants, and also, in contrast to pre-boost sera, against Delta compared to WT (p < 0.05, p < 0.001, p < 0.0001 for Delta, Beta and BA.1, BA.4/BA.5 respectively) (Figure 1E).
The impact of booster on the convalescent donors was distinctly different from that on the non-exposed donors (Figure 1G,H,I). First of all, the level of pre-boost NAbs in this subgroup was higher than in the non-exposed subgroup, and 100% samples exceeded the baseline of virus neutralization across all the studied variants (Figure 1G). The significant difference in neutralization levels compared to WT was observed only for the Omicron variants in both pre- and post- boost sera (p < 0.01 for both time points and both BA.1 and BA.4/BA.5) (Figure 1G,H).
Finally, we compared pre- and post-boost level of NAbs with each other for each variant (Figure 1C,F,I). Sputnik V booster significantly increased neutralization titers to all studied variants in the non-exposed subgroup (Figure 1C). We observed the largest increase ID50 for Alpha (5.6-fold change, p < 0.0001), and the least increase for the WT (2.9-fold change, p < 0.0001) variant. The titers against Beta, Delta, BA.1, and BA.4/BA.5 increased by 3.6, 4.7, 3.2, and 5.3 times, respectively (p < 0.01 for BA.4/BA.5 and p < 0.0001 for others). These results suggest that Sputnik V can boost neutralizing antibody levels in non-exposed donors against various variants of SARS-CoV-2, including Omicron.
The Comirnaty booster induced strong increase in VNT values against all the analyzed variants (Figure 1F). Fold changes were higher than those observed with the homologous booster, with increases of 12.6, 26.2, 29.1, 16.3, 34.5, and 26.5 times for WT, Alpha, Beta, Delta, BA.1, and BA.4/BA.5 respectively.
No changes in VNTs were observed among the recovered donors who received the Sputnik V booster (Figure 1I), suggesting that it was the infection rather than boosting that contributed the most to the neutralization profiles observed.
2.3. Antigenic maps reveal the emergence of booster-induced cross-neutralizing antibodies
Antigenic maps were created to visualize the relationships between the sera and the VNTs of SARS-CoV-2 Spike-pseudotyped lentiviral particles (Figure 2). The positioning of each serum sample on the maps is indicative of its efficacy in neutralizing a particular variant. One grid line on the antigenic maps corresponds to a two-fold dilution in the virus neutralization assay (one arbitrary unit, AU). We used this scale to examine the changes in antigenic distances between WT and other studied variants to reveal cross-neutralization activity of serum samples.
Serum samples are shown in open (A, B, C, before boost) or solid (B, D, E, after boost) diamonds as follows: non-exposed donors with Sputnik V boost (black), non-exposed donors with Comirnaty boost (olive), and convalescent donors with Sputnik V boost (blue). Each grid square (one antigenic unit) corresponds to a twofold dilution in the neutralization assay. Antigens are shown as circles and labeled.
(G, H. I) Hat graphs represent changes in antigenic distances between the WT and Spike variants during revaccination. The brim of the hat corresponds to the antigenic distance for pre-boost samples. The hat height indicates the change in antigenic distance between pre- and post-boost samples. Green/red hats indicate increase/decrease in antigenic distance. The numbers indicate the corresponding changes in the antigenic units.
(J) Breadth gain plots of serum samples from individuals primarily vaccinated with Sputnik V and who received homologous with Sputnik V or heterologous with Comirnaty revaccination. The x axis represents the antigenic distances of Alpha, Beta, Delta, and Omicron BA.1 and BA.4/BA.5 variants from WT, which were taken from [6,9]. The y axis represents fold increase in neutralization titer after Sputnik V or Comirnaty revaccination.
In the subgroup of non-exposed donors before Sputnik V revaccination, serum samples clustered near the WT- and Alpha-based Spikes and less so near the Beta variant (Figure 2A). Sputnik V booster led to tighter clustering near Alpha, which corresponds to the maximum increase in ID50 against Alpha, and in a smaller increase near the WT Spike (Figure 2B).
Sera from the “Comirnaty booster” group before revaccination likewise clustered around WT and Alpha. Two sera were close to Delta (Figure 2D) which could be due to the asymptomatic or undetected infection with the Delta variant, which was spread in Russia in 2021 [12]. To control for potential errors related to these two sera, we constructed additional maps, where the data from these two donors were excluded (Figure S1). These maps did not show significant differences of antigenic distances between WT and the variants, so our next analysis was based on the full subgroup (n=12). No sera were observed to cluster near the Beta variant, in contrast to the “Sputnik V booster” subgroup. Post-boost sera, again, clustered close to WT and Alpha (Figure 2E).
In the subgroup of the convalescent individuals, both before and after revaccination, sera clustered close to WT and Alpha (Figure 2G,H).
Initially, in the sample collected before revaccination, the vaccine-resistant variants BA.1 and BA.4/BA.5 were positioned the farthest from WT across all three study subgroups (Figure 2A,D,G), consistent with the previous studies [4,6,11]. Hat graphs showed that BA.1 after Sputnik V became 1.1 AU closer to WT in non-exposed donors’ subgroup, unlike the BA.4/BA.5 that became more distant by 0.4 AU (Figure 2C). Beta, Delta, BA.1, and BA.4/BA.5 became approximately equidistant to WT after Comirnaty revaccination, also, BA.1 became closer to WT (1.0 AU) (Figure 2F). The change in antigenic distance between WT and BA.4/BA.5 was not significant in this subgroup. In the convalescent subgroup, BA.1 and BA.4/BA.5 did not appreciably move relatively to the WT (Figure 2I).
Next, our analysis uncovered the opposite trends in the antigenic distance shifts between WT and Delta Spike variants, depending on the nature of the booster. Specifically, Sputnik V booster led to a decrease by 0.6 AU in the non-exposed and convalescent subgroups. In contrast, after Comirnaty booster, the antigenic distance increased by 0.6 AU.
Finally, we created a breadth gain plot [9] for all three study subgroups. Here, x axis shows the antigenic distance between WT and the variants, which were taken from [6,9]. Y axis corresponds to the boost-induced fold change in VNTs. Area under the curve in Comirnaty boost subgroup is visually larger than for other two subgroups. The gain in titer was smaller for Delta and BA.4/BA.5 compared to the earlier viral variants (Alpha and Beta, respectively) than reported earlier [9]. That study demonstrated that fold increase in VNT values corresponds to larger antigenic distance between the vaccine strain (WT) and the variant. The non-exposed subgroup was deviated from this trend because of smaller fold changes for Delta, Beta, and BA.1 compared to the variants that emerged earlier. The graph for the convalescent donors was essentially parallel and very close to the x axis.
3. Discussion
The most common method for assessing humoral anti-viral immunity induced by vaccination or booster vaccination is through the measurements of VNTs. In early days of SARS-CoV-2 research, when the ancestral strain or a small number of early viral lineages dominated, the traditional method of evaluating virus neutralizing activity was appropriate. However, as SARS-CoV-2 evolved rapidly, the large amount of neutralization data against multiple variants has made the traditional graphs difficult to interpret.
Under these circumstances, the approach referred to as antigenic cartography provided a more comprehensive understanding of the antigenic diversity of the viral Spike. This approach was initially developed for human seasonal influenza virus but more recently it has also been successfully applied to lyssaviruses, flaviviruses and other viruses, as well [13,14,15]. Antigenic cartography is now also being used to analyze the antigenic relationships between SARS-CoV-2 variants [6].
The crucial task of serological studies is to figure out the effectiveness of vaccine-induced humoral response against a broad range of viral antigens, including those of the emerging and immune-evading viral variants. However, although the term “breadth of virus neutralization” applied to serum response is widely spread in the scientific literature, there is no quantitative assessment for this key parameter. In the case of influenza infection, antigenic cartography is employed to estimate the antigenic distances between the variants and to inform decisions about vaccine strain updates. It is recommended to update the vaccine strain when the antigenic distances between the previous vaccine strain and the new VOCs exceed approximately 2 AU. This recommendation is based on the immunological understanding that the breadth of virus neutralization in humoral response is insufficient for protection against new variants. Conventionally, the standard reference set of ferret or human sera following primary infection is used to create antigenic maps [6,7]. Because of the fact that SARS-CoV-2-specific human immunity is currently a composite of the previous infection and/or vaccination history, SARS-CoV-2 antigenic maps are now also being constructed using vaccine-elicited sera.
In this study, to quantify and interpret the antigenic relationships between SARS-CoV-2 variants after booster, we used both the traditional method for assessing the virus-neutralizing activity of sera and the antigenic cartography. Some peculiarities of the immune response were revealed via traditional approach (Figure 1). Both homologous and heterologous regimens of revaccination successfully boosted antibody response in non-exposed donors against all the studied variants. Despite the fact that both vaccines (Sputnik V and Comirnaty) were based on the ancestral strain, we did not observe the maximum increase in titers against the WT Spike-pseudotyped lentiviral particles. This is in agreement with the theory of saturation of high-affinity antibodies [5,16], in our example against WT, and biased selection towards low-affinity antibodies that can neutralize virus variants. In non-exposed individuals VNTs against the Beta and Omicron variants, but not the Alpha variant, were significantly lower than those against WT both before and after revaccination. Heterologous booster elicited a more robust immune response, consistent with the previously published data on booster with Comirnaty after prime vaccination with ChAdOx1-nCoV-19 [17,18,19].
Sputnik V booster did not result in increased antibody levels in a subgroup of individuals who had recovered from COVID-19 1-4 months prior to booster. Previously, for Sputnik V revaccination it was shown that the magnitude of boost-induced increase in antibody levels depended on the initial level of antibodies at the time of revaccination [20]. This effect may have been related to the presence of high levels of specific anti-S antibodies and T-cells, which were sufficient to neutralize the antigen induced by the booster. It is also possible that longer intervals between recovery from infection and subsequent vaccination might be needed to achieve improvements in antibody levels and better cross-neutralization of viral variants [21].
Constructing the antigenic maps allowed us to uncover subtle features of the homologous and heterologous boosters. Despite the fact that both boosters were based on the ancestral Spike sequence, a decreased antigenic distance was observed between some of the viral variants, which is probably best explained by the emergence of cross-reactivity in the sera.
Our results also indicate that antigenic distances between WT and Delta shifted in opposite directions following the different booster vaccination regimens. Thus, our data agree with the previously published studies demonstrating that booster with mRNA COVID-19 vaccines leads to the increased distance between WT and Delta [8,9]. Here, we demonstrate that the homologous adenovirus-based vaccine booster reduces the antigenic distance between WT and Delta for both non-exposed and convalescent individuals. The exact mechanism behind this difference remains to be discovered.
The most significant changes between WT and the studied variants were observed for BA.1 in non-exposed donors for both regimens. In contrast to Delta, these changes were unidirectional, i.e. the distances decreased by nearly 1 AU. Previous research on mRNA vaccines demonstrated that a third booster reduces the antigenic distance between WT and BA.1 by approximately 2.6 AU [8,9]. Our findings highlight that a single Comirnaty booster, following a priming dose of adenovirus-based vaccine, also led to shorter antigenic distances between WT and BA.1 but to a lesser extent.
We observed only minor increase following Comirnaty booster, which contrasts with the previous study, where antigenic distance between WT and BA.4/BA.5 decreased by nearly 1 AU following the third Comirnaty booster [9]. After Sputnik V booster, the antigenic distances between WT and BA.4/BA.5 in our study increased by 0.4 AU. As a result, after the homologous booster, sera were less effective in recognizing BA.4/BA.5 compared to BA.1. However, following the heterologous booster, both Omicron variants became nearly antigenically equivalent.
To explain the mechanisms responsible for the observed differences following boosting regimens, a more detailed examination of the antigen (S-protein) epitope characteristics induced by the vaccine is required. We hypothesize that the different conformation of the S-protein on target cells may influence the recognition patterns during antibody maturation. mRNA-based vaccines encode prefusion-stabilized conformation of the Spike protein with two proline substitutions, while Gam-COVID-Vac encodes an unmodified full-length Spike [22,23,24].
Our study has several limitations. First, the number of study participants in the convalescent and Comirnaty-revaccinated groups was relatively small. A second limitation is that some of donors could have had asymptomatic COVID-19 that remained undetected.
4. Materials and Methods
4.1. Ethics
Before carrying out any study procedures, written informed consent was given by all the study participants. The Institute of Immunology's Medical Ethical Committee assessed and approved the study protocol (#12-1, December 29, 2020).
4.2. Volunteers and samples collection
A cohort of 59 volunteers was enrolled at the National Research Center Institute of Immunology of The Federal Medical Biological Agency of Russia. Between January and March 2021, all subjects received two doses of Gam-COVID-Vac vaccine 21 days apart. Approximately 6 months after the prime, the booster vaccine dose was given. We divided study cohort in 3 subgroups: 1) non-exposed donors, who received Sputnik Light as a booster (n = 39), 2) non-exposed donors, who received Pfizer/BioNTech’s as a booster (n = 12), 3) donors who recovered from COVID-19 and subsequently received Sputnik Light as a booster (n = 7); Sputnik Light representing the first dose of Gam-COVID-Vac (Sputnik V). Non-exposed donors did not have a history of COVID-19, anti-N antibodies were absent from their serum samples before the revaccination. Convalescent donors were infected by SARS-CoV-2 in the period between prime and booster vaccination (Table 1).
Serum samples were collected one day before the booster vaccination and about one month after. Sera were aliquoted and frozen at -70°C for ELISA and neutralization tests.
4.3. SARS-CoV-2 pseudoviral particles production
Three plasmids were utilized to generate lentiviral particles pseudotyped with SARS-CoV-2 Spike: HIV-1 packaging pCMVΔ8.2R (Addgene), transfer pUCHR-GFP (Addgene), and envelope pCAGGS-Swt- Δ1932 [25]. Several plasmids encoding variants of the SARS-CoV-2 Spike were obtained via site-specific mutagenesis or gene synthesis (Genewiz) using the pCAGGS-Swt-Δ19 plasmid as a template. This plasmid encoded an ancestral Wuhan-Hu-1 Spike(Δ19) that lacked 19 C-terminal amino acid residues and was codon-optimized.
Specifically, these Spike variants had the following substitutions: del69-70, del144, N501Y, A570D, D614G, P681H, T716L, S982A, D1118A (Alpha); D80A, D215G, del241/243, K417N, E484K, N501Y, D614G, A701V (Beta); T19R, T95I, del156, del157, R158G, L452R, T478K, D614G, P681R, D950N (Delta); A67V, Δ69–70, T95I, G142D, Δ143–145, del211, L212I, ins214EPE, G339D, S371L, S373P, S375F, K417N, N440K, G446S, S477N, T478K, E484A, Q493R, G496S, Q498R, N501Y, Y505H, T547K, D614G, H655Y, N679K, P681H, N764K, D796Y, N856K, Q954H, N969K, L981F (Omicron BA.1); T19I, L24S, del25/27, del69/70, G142D, V213G, G339D, S371F, S373P, S375F, T376A, D405N, R408S, K417N, N440K, L452R, S477N, T478K, E484A, F486V, Q498R, N501Y, Y505H, D614G, H655Y, N679K, P681H, N764K, D796Y, N969K (Omicron BA.4/BA.5). Lentiviral particles were pseudotyped with the SARS-CoV-2 Spike protein from either the WT strain or the variant strains as previously described [26]. Pseudoviral particles were frozen at -70°C for later use in neutralization assays. Titers of the particles obtained were determined based on the HEK293-hACE2 cells.
4.4. Pseudotyped virus neutralization assay
Pseudotyped virus neutralization assay was performed as previously described [24]. Briefly, serial diluted sera (1:4, 1:16, 1:64, 1:256, 1:1024, 1:4096) were incubated with pseudoviral particles for 1 hour at room temperature, added to HEK293-hACE2 cells seeded in a 96-well plate and placed into a CO2 incubator for 4 days (37 °C, 5% CO2). Neutralization titers (ID50) were calculated by non-linear regression curve fit with Sigmoidal, 5PL (GraphPad Software, San Diego, CA, USA, version 8.0.1).
4.5. Antigenic cartography
The antigenic cartography approach is based on modified multidimensional scaling [3]. «Racmacs» package (https://acorg.github.io/Racmacs/, version 1.1.35) was used to create antigenic maps with 1000 optimizations. Antigenic maps represent geometric interpretation of relationships between neutralization titers of sera and viral antigens in a 2D space. One grid line on the maps (antigenic unit, AU) corresponds to a two-fold dilution of sera in the virus neutralization assay.
4.6. Statistical analysis
Non-parametric analogous of ANOVA for matched data (Friedman test) with Dunn’s multiple comparisons (titers of variants compared to WT or titers compared to each other before and after the booster vaccination) was performed using GraphPad Prism software. Data on the graphs are presented as medians and interquartile ranges (IQR). P values below 0.05 were considered statistically significant.
5. Conclusions
Our study contributes to the discussion on vaccine regimens and methods of determining cross-reactivity of the antibody repertoires. We show that heterologous booster with Comirnaty following the primary vaccination with Sputnik V broadens the neutralization activity of sera to a greater extent than the homologous Sputnik V/Sputnik V booster. However, regardless of the nature of the booster used, the most recent of the studied variants, BA.1 and BA.4/BA.5, are still antigenically distant from the ancestral variant, which is used in both vaccines. The implications of these findings may be significant when considering a new vaccine regimen.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Antigenic maps of (A) pre- and (B) post-boost serum samples against wild-type (WT), Alpha, Beta, Delta, and Omicron BA.1 and BA.4/BA.5 variants from non-exposed donors with Comirnaty boost.
Author Contributions
“Conceptualization, A.V.F.; methodology, E.A.A.; software, A.A.M.; validation, E.A.A. and M.G.B.; formal analysis, A.A.M.; investigation, E.A.A., A.A.M., M.G.B., M.M.S., and A.A.M; resources, E.A.A.; data curation, A.A.M.; writing—original draft preparation, E.A.A.; writing—review and editing, E.A.A., A.A.G., and A.V.F.; visualization, E.A.A. and A.A.M.; supervision, A.V.F.; project administration, A.V.F.; funding acquisition, E.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant number 23-25-00472.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki. The study protocol was reviewed and approved by the Medical Ethical Committee of the Institute of Immunology (protocol code #12-1, 29 December 2020).
Informed Consent Statement
Written informed consent was obtained from each of the study participants before performing any study procedures.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Alexander Taranin for helpful discussion and providing reagents.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gattinger, P.; Ohradanova-Repic, A.; Valenta, R. Importance, Applications and Features of Assays Measuring SARS-CoV-2 Neutralizing Antibodies. Int J Mol Sci 2023, 24, 5352. [Google Scholar] [CrossRef] [PubMed]
- Sholukh, A.M.; Fiore-Gartland, A.; Ford, E.S.; Miner, M.D.; Hou, Y.J.; Tse, L. V.; Kaiser, H.; Zhu, H.; Lu, J.; Madarampalli, B.; et al. Evaluation of Cell-Based and Surrogate SARS-CoV-2 Neutralization Assays. J Clin Microbiol 2021, 59, e00527–21. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.J.; Lapedes, A.S.; De Jong, J.C.; Bestebroer, T.M.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Mapping the Antigenic and Genetic Evolution of Influenza Virus. Science 2004, 305, 371–376. [Google Scholar] [CrossRef]
- Mykytyn, A.Z.; Rissmann, M.; Kok, A.; Rosu, M.E.; Schipper, D.; Breugem, T.I.; van den Doel, P.B.; Chandler, F.; Bestebroer, T.; de Wit, M.; et al. Antigenic Cartography of SARS-CoV-2 Reveals That Omicron BA.1 and BA.2 Are Antigenically Distinct. Sci Immunol 2022, 7, eabq4450. [Google Scholar] [CrossRef] [PubMed]
- Rössler, A.; Netzl, A.; Knabl, L.; Schäfer, H.; Wilks, S.H.; Bante, D.; Falkensammer, B.; Borena, W.; von Laer, D.; Smith, D.J.; et al. BA.2 and BA.5 Omicron Differ Immunologically from Both BA.1 Omicron and Pre-Omicron Variants. Nat Commun 2022, 13, 7701. [Google Scholar] [CrossRef]
- Wilks, S.H.; Mühlemann, B.; Shen, X.; Türeli, S.; LeGresley, E.B.; Netzl, A.; Caniza, M.A.; Chacaltana-Huarcaya, J.N.; Corman, V.M.; Daniell, X.; et al. Mapping SARS-CoV-2 Antigenic Relationships and Serological Responses. bioRxiv Prepr Serv Biol 2022, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, J.; Zou, J.; Kalveram, B.; Machado, R.R.G.; Ren, P.; Türeli, S.; Smith, D.J.; Weaver, S.C.; Xie, X.; et al. Cross-Neutralization and Cross-Protection among SARS-CoV-2 Viruses Bearing Different Variant Spikes. Signal Transduct Target Ther 2022, 7, 2020–2023. [Google Scholar] [CrossRef]
- Lusvarghi, S.; Pollett, S.D.; Neerukonda, S.N.; Wang, W.; Wang, R.; Vassell, R.; Epsi, N.J.; Fries, A.C.; Agan, B.K.; Lindholm, D.A.; et al. SARS-CoV-2 BA.1 Variant Is Neutralized by Vaccine Booster–Elicited Serum but Evades Most Convalescent Serum and Therapeutic Antibodies. Sci Transl Med 2022, 14, 8543. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lusvarghi, S.; Subramanian, R.; Epsi, N.J.; Wang, R.; Goguet, E.; Fries, A.C.; Echegaray, F.; Vassell, R.; Coggins, S.A.; et al. Antigenic Cartography of Well-Characterized Human Sera Shows SARS-CoV-2 Neutralization Differences Based on Infection and Vaccination History. Cell Host Microbe 2022, 30, 1745–1758.e7. [Google Scholar] [CrossRef] [PubMed]
- Neerukonda, S.N.; Vassell, R.; Lusvarghi, S.; Wang, R.; Echegaray, F.; Bentley, L.; Eakin, A.E.; Erlandson, K.J.; Katzelnick, L.C.; Weiss, C.D.; et al. SARS-CoV-2 Delta Variant Displays Moderate Resistance to Neutralizing Antibodies and Spike Protein Properties of Higher Soluble ACE2 Sensitivity, Enhanced Cleavage and Fusogenic Activity. Viruses 2021, 13, 2485. [Google Scholar] [CrossRef] [PubMed]
- van der Straten, K.; Guerra, D.; van Gils, M.J.; Bontjer, I.; Caniels, T.G.; van Willigen, H.D.G.; Wynberg, E.; Poniman, M.; Burger, J.A.; Bouhuijs, J.H.; et al. Antigenic Cartography Using Sera from Sequence-Confirmed SARS-CoV-2 Variants of Concern Infections Reveals Antigenic Divergence of Omicron. Immunity 2022, 55, 1725–1731. [Google Scholar] [CrossRef] [PubMed]
- Gushchin, V.A.; Pochtovyi, A.A.; Kustova, D.D.; Ogarkova, D.A.; Tarnovetskii, I.Y.; Belyaeva, E.D.; Divisenko, E. V.; Vasilchenko, L.A.; Shidlovskaya, E. V.; Kuznetsova, N.A.; et al. Dynamics of SARS-CoV-2 Major Genetic Lineages in Moscow in the Context of Vaccine Prophylaxis. Int J Mol Sci 2022, 23, 14670. [Google Scholar] [CrossRef] [PubMed]
- Horton, D.L.; McElhinney, L.M.; Marston, D.A.; Wood, J.L.N.; Russell, C.A.; Lewis, N.; Kuzmin, I. V.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Fooks, A.R.; et al. Quantifying Antigenic Relationships among the Lyssaviruses. J Virol 2010, 84, 11841–11848. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, A.; Marini, V.; Di Gennaro, A.; Ronchi, G.F.; Casaccia, C.; Carelli, G.; Passantino, G.; D’Alterio, N.; D’Innocenzo, V.; Savini, G.; et al. Antigenic Relationship among Zoonotic Flaviviruses from Italy. Infect Genet Evol 2019, 68, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Katzelnick, L.C.; Fonville, J.M.; Gromowski, G.D.; Arriaga, J.B.; Green, A.; James, S.L.; Lau, L.; Montoya, M.; Wang, C.; Vanblargan, L.A.; et al. Dengue Viruses Cluster Antigenically but Not as Discrete Serotypes. Science 2015, 349, 1338–1343. [Google Scholar] [CrossRef] [PubMed]
- Schaefer-Babajew, D.; Wang, Z.; Muecksch, F.; Cho, A.; Loewe, M.; Cipolla, M.; Raspe, R.; Johnson, B.; Canis, M.; DaSilva, J.; et al. Antibody Feedback Regulates Immune Memory after SARS-CoV-2 mRNA Vaccination. Nature 2023, 613, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Bekliz, M.; Adea, K.; Vetter, P.; Eberhardt, C.S.; Hosszu-Fellous, K.; Vu, D.L.; Puhach, O.; Essaidi-Laziosi, M.; Waldvogel-Abramowski, S.; Stephan, C.; et al. Neutralization Capacity of Antibodies Elicited through Homologous or Heterologous Infection or Vaccination against SARS-CoV-2 VOCs. Nat Commun 2022, 13, 1–10. [Google Scholar] [CrossRef]
- Fabricius, D.; Ludwig, C.; Scholz, J.; Rode, I.; Tsamadou, C.; Jacobsen, E.M.; Winkelmann, M.; Grempels, A.; Lotfi, R.; Janda, A.; et al. mRNA Vaccines Enhance Neutralizing Immunity against SARS-CoV-2 Variants in Convalescent and ChAdOx1-Primed Subjects. Vaccines 2021, 9, 918. [Google Scholar] [CrossRef]
- Hillus, D.; Schwarz, T.; Tober-Lau, P.; Vanshylla, K.; Hastor, H.; Thibeault, C.; Jentzsch, S.; Helbig, E.T.; Lippert, L.J.; Tscheak, P.; et al. Safety, Reactogenicity, and Immunogenicity of Homologous and Heterologous Prime-Boost Immunisation with ChAdOx1 NCoV-19 and BNT162b2: A Prospective Cohort Study. Lancet Respir Med 2021, 9, 1255–1265. [Google Scholar] [CrossRef]
- Godkov, M.A.; Ogarkova, D.A.; Gushchin, V.A.; Kleymenov, D.A.; Mazunina, E.P.; Bykonia, E.N.; Pochtovyi, A.A.; Shustov, V. V.; Shcheblyakov, D. V.; Komarov, A.G.; et al. Revaccination in Age-Risk Groups with Sputnik V Is Immunologically Effective and Depends on the Initial Neutralizing SARS-CoV-2 IgG Antibodies Level. Vaccines 2023, 11, 90. [Google Scholar] [CrossRef]
- Bates, T.A.; Leier, H.C.; McBride, S.K.; Schoen, D.; Lyski, Z.L.; Lee, D.X.; Messer, W.B.; Curlin, M.E.; Tafesse, F.G. An Extended Interval between Vaccination and Infection Enhances Hybrid Immunity against SARS-CoV-2 Variants. JCI Insight 2023, 8, e165265. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.J.; Rouphael, N.G.; Widge, A.T.; Jackson, L.A.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Safety and Immunogenicity of SARS-CoV-2 mRNA-1273 Vaccine in Older Adults. N Engl J Med 2020, 383, 2427–2438. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.E.; Frenck, R.W.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based COVID-19 Vaccine Candidates. N Engl J Med 2020, 383, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Logunov, D.Y.; Dolzhikova, I. V.; Zubkova, O. V.; Tukhvatullin, A.I.; Shcheblyakov, D. V.; Dzharullaeva, A.S.; Grousova, D.M.; Erokhova, A.S.; Kovyrshina, A. V.; Botikov, A.G.; et al. Safety and Immunogenicity of an rAd26 and rAd5 Vector-Based Heterologous Prime-Boost COVID-19 Vaccine in Two Formulations: Two Open, Non-Randomised Phase 1/2 Studies from Russia. Lancet 2020, 396, 887–897. [Google Scholar] [CrossRef]
- Byazrova, M.G.; Kulemzin, S. V.; Astakhova, E.A.; Belovezhets, T.N.; Efimov, G.A.; Chikaev, A.N.; Kolotygin, I.O.; Gorchakov, A.A.; Taranin, A. V.; Filatov, A. V. Memory B Cells Induced by Sputnik V Vaccination Produce SARS-CoV-2 Neutralizing Antibodies Upon Ex Vivo Restimulation. Front Immunol 2022, 13, 840707. [Google Scholar] [CrossRef]
- Astakhova, E.A.; Byazrova, M.G.; Yusubalieva, G.M.; Kulemzin, S. V.; Kruglova, N.A.; Prilipov, A.G.; Baklaushev, V.P.; Gorchakov, A.A.; Taranin, A. V.; Filatov, A. V. Functional Profiling of In Vitro Reactivated Memory B Cells Following Natural SARS-CoV-2 Infection and Gam-COVID-Vac Vaccination. Cells 2022, 11. [Google Scholar] [CrossRef]
Figure 1.
Neutralization antibody titers (ID50 values) against SARS-CoV-2 variant pseudoviruses for pre- and post-boost sera from individuals primarily vaccinated with Sputnik V.
Figure 1.
Neutralization antibody titers (ID50 values) against SARS-CoV-2 variant pseudoviruses for pre- and post-boost sera from individuals primarily vaccinated with Sputnik V.
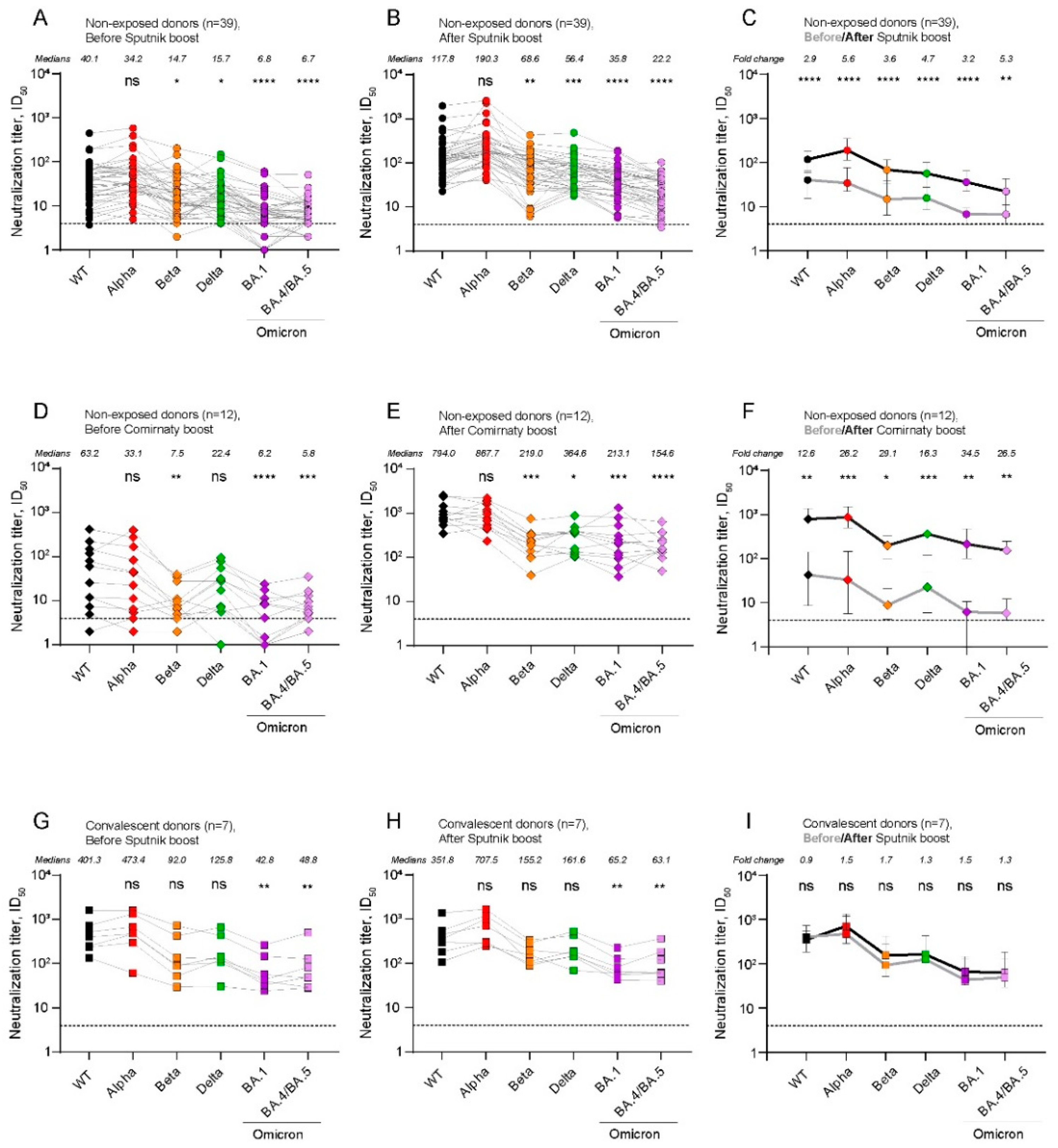
Figure 2.
Antigenic cartography of pre- and post-boost serum samples against wild-type (WT), Alpha, Beta, Delta, and Omicron BA.1 and BA.4/BA.5 variants.
Figure 2.
Antigenic cartography of pre- and post-boost serum samples against wild-type (WT), Alpha, Beta, Delta, and Omicron BA.1 and BA.4/BA.5 variants.
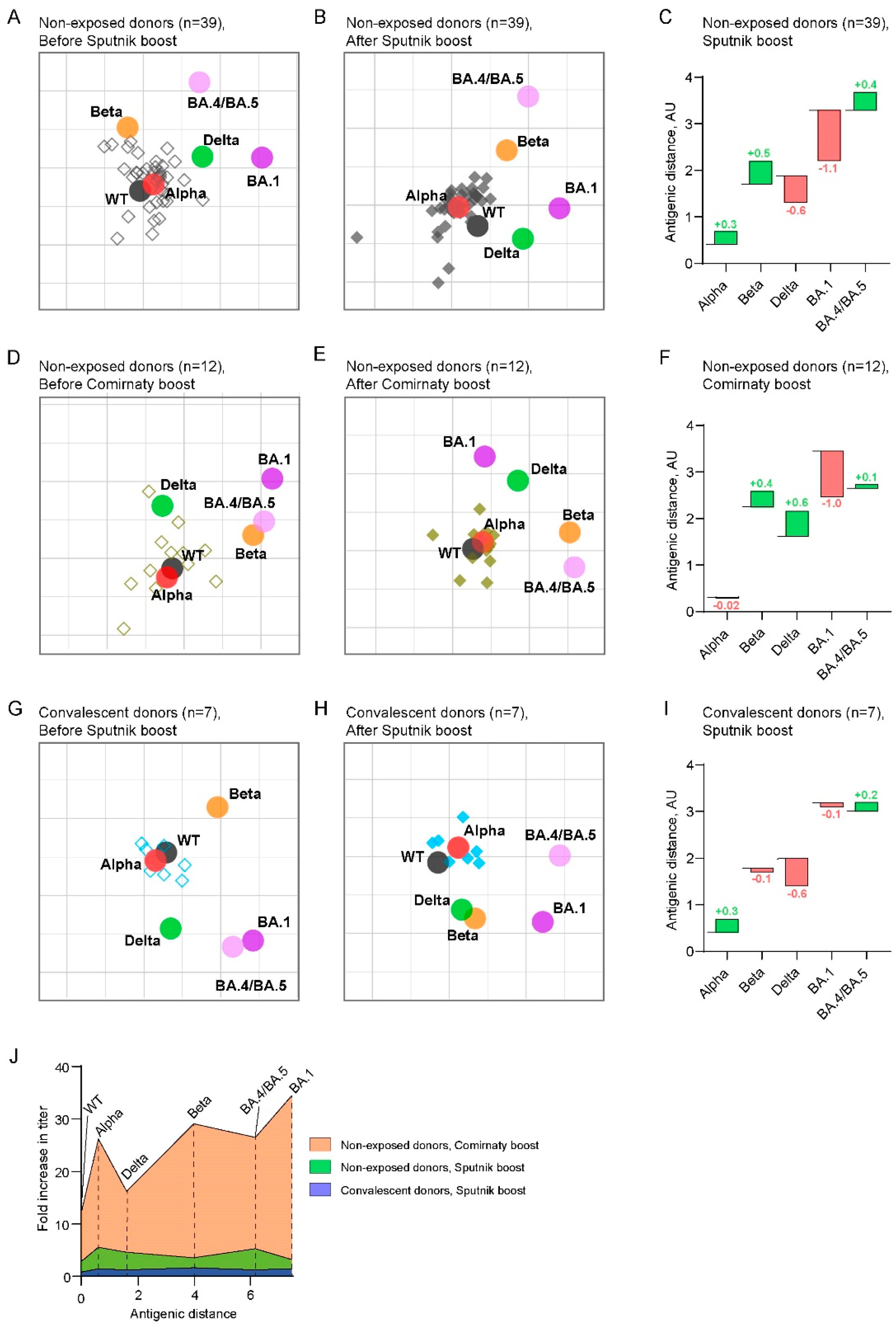

Table 1.
Participant characteristics.
| Name of subgroup | Non-exposed donors, Sputnik V booster |
Non-exposed donors, Comirnaty booster |
Convalescent donors, Sputnik V booster |
|
|---|---|---|---|---|
| Number of participants | 39 | 12 | 7 | |
| Age | Years, median (range) | data data |
data data |
|
| Sex | Female | 19 | 6 | 3 |
| Male | 20 | 6 | 4 | |
| Booster vaccine | Spuntik V | Comirnaty | Spuntik V | |
| Probe sampling after second dose of the first vaccination | 195 (176 - 270) | 275 (261 - 335) | 207 (183 - 282) | |
| Probe sampling after boost | 33 (29 - 39) | 38 (30 - 46) | 31 (30 - 36) | |
| Anti-N IgG antibodies before boost | 0/39 | 0/12 | 5/7 | |
| Anti-S IgG antibodies before boost | 6.0 (0 – 10.4) | 7.3 (0 – 11.8) | 12.7 (11.2 – 14.4) | |
| PCR-confirmed COVID-19 | ||||
| Time between recovery and boost | 0/39 | 0/12 | 7/7 | |
| Infection period | - | - | May – August 2021 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



