
Submitted:
23 May 2023
Posted:
24 May 2023
You are already at the latest version
Abstract
The response of boreal vegetation to global warming has shown a weakening trend over the last three decades. However, in previous studies, models of vegetation activity responses to temperature change have often only considered changes in mean daily temperature (Tmean), with the diurnal temperature range (DTR) being neglected. The goal of this study is to evaluate the temporal trends of the relationships between two temperature factors (Tmean and DTR) and vegetation activity across the boreal regions on both annual and seasonal timescales, by simultaneously employing satellite and climate datasets. We found that the interannual partial correlation between growing season (GS) NDVI and Tmean (RNDVI−Tmean) showed a significant decreasing trend over the last 34 years. At the seasonal scale, RNDVI−Tmean shows a significant upward trend in the spring, while in the summer and autumn, RNDVI−Tmean exhibits a significant downward trend. The temporal trend characteristics of the partial correlation between NDVI and DTR (RNDVI−DTR), at both the GS and seasonal scales, are fully consistent with RNDVI−Tmean. The area with a significant decrease in GS RNDVI−Tmean and RNDVI−DTR accounted for approximately 44.40% and 41.21% of the boreal region with the 17-year moving window, respectively. In stark contrast, the area exhibiting a significant increasing trend in GS RNDVI−Tmean and RNDVI−DTR accounted for only approximately 14.65% and 13.59% of the boreal region with the 17-year moving window, respectively. With respect to the seasonal patterns of RNDVI−Tmean and RNDVI−DTR, the area with a significant upward trend in spring was greater than that with a significant downward trend. Nevertheless, more areas had a significant downward trend of RNDVI−Tmean and RNDVI−DTR in summer and autumn than had a significant upward trend. Overall, our research reveals a weakening trend in the impact of temperature on vegetation activity in the boreal regions, and contributes to a deeper understanding of the vegetation response to global warming.
Keywords:
Temperature factors
; Vegetation activity
; Diurnal temperature range
; Boreal forest
; Climate change
1. Introduction
Boreal forest is the largest terrestrial biome in the world and stores more than 30% of global forest carbon (Bonan et al., 1992; Sulla-Menashe et al., 2018; Zhao et al., 2021). Recent studies investigating the responses of vegetation to climatic factors in northern boreal ecosystems (Deng et al., 2022; Piao et al., 2014; Tan et al., 2015) involving satellite-derived data and manipulative experiments, have all indicated that vegetation productivity in northern high latitudes is sensitive to temperature change (Huang et al., 2017; Wolkovich et al., 2012; Wu et al., 2011). The results show that boreal forests are closely linked to mean daily temperature (Tmean) and diurnal temperature range (DTR; the difference between daily maximum and daily minimum temperature) (Gauthier et al., 2015). However, most of this research was static and therefore unable to clarify the temporal variations in the relationships between vegetation greenness and temperature factors [e.g. maximum daily temperature (Tmax), minimum daily temperature(Tmin) and DTR] over the last few decades (Du et al., 2022).
In fact, the relationships between temperature factors and vegetation activity may change over time due to the limitations of other environmental factors at regional and global scales (Piao et al., 2014; Zhao et al., 2022). For instance, Piao et al. (2014) found that the strength of the relationship between the interannual variability of vegetation greenness and air temperature in northern ecosystems declined substantially over the last three decades. This decline may be related to an increase of days with extreme heat and the nonlinear response of photosynthesis to air temperature in these northern high-latitude ecosystems (Piao et al., 2014). Zhao et al. (2022) investigated the inter-annual responses of vegetation greenness to the diurnal asymmetric warming at the global scale during the last 34 years. They found that the decline in the vegetation greenness response to Tmax occurred mainly in the high latitudes of the northern hemisphere, whereas the decline in the vegetation activity response to Tmin was primarily concentrated at low latitudes.
Although the response of vegetation activity to Tmean, Tmax, and Tmin during the last three decades has been extensively investigated, the effect of DTR on vegetation activity remains largely unknown. Previous observational analyses show that the land surface DTR has significantly decreased, at a rate of − 0.036 K decade− 1, over the last century (Sun et al., 2018). Huang et al. (2020) examined the impact of preseason DTR on spring vegetation phenology in the Northern Hemisphere, and recommend that more studies should be carried out to help understand the physiological mechanisms governing the response of the start of the growing season to preseason DTR. Variations in the link between DTR and boreal vegetation activity over time currently remain unclear.
The main objective of this study was to explore the temporal dynamics of the relationships between temperature factors (Tmean and DTR) and vegetation activity across boreal North America over the last three decades, by simultaneously employing gridded meteorological data and the Normalized Difference Vegetation Index (NDVI) obtained from National Oceanic and Atmospheric Administration (NOAA) satellites. The insights derived from the findings are expected to have important implications for studying climate change and its effect on the boreal forests.
2. Data and methods
2.1. Data sources
2.1.1. Remote sensing data
The third-generation NDVI dataset for the period 1982–2015 used in this study was produced by the Global Inventory Modelling and Mapping Studies group (GIMMS) from the NOAA/AVHRR, which have been corrected for sensor degradation, cloud cover, solar zenith angle, and viewing angle effects due to volcanic aerosols and satellite drift (Tucker et al., 2005). The dataset is the longest sequence of NDVI data released to date and has been widely used in the investigation of large-scale vegetation activity and dynamics (Lian et al., 2021; Sulla-Menashe et al., 2018) .The biweekly NDVI data with a spatial resolution of 8km × 8 km were further aggregated to 0.5° × 0.5° to match the resolution of the meteorological data (Chen and Yang, 2020; Shen et al., 2018).
Monthly Tmean, Tmax, Tmin and precipitation data with a spatial resolution of 0.5° × 0.5° were compiled from the Climate Research Unit, University of East Anglia (CRU TS 4.01) (Harris et al., 2020). This dataset was produced using angular-distance weighting interpolation (Zhao et al., 2022), and has been wildly used in studies of the relationship between regional and global vegetation activity and climate change (Peng et al., 2013; Piao et al., 2011; Wen et al., 2018). DTR was calculated as the difference between the Tmax and Tmin values obtained from the CRU TS 4.01 product.
2.3. Methods
The study covered the boreal (as defined by the Köppen-Geiger climate classification, www.gloh2o.org/koppen, accessed on 20 March 2023) vegetated areas (defined as areas with a mean growing season (GS) NDVI during 1982–2015 larger than 0.1) north of 40°N (Piao et al., 2011). The study period, which was based on the availability of satellite observations, was 1982–2015. GS was defined as April to October, while spring, summer, and autumn, were defined as April to May, June to August, and September to October, respectively (Piao et al., 2014; Piao et al., 2011).
To determine temporal trends in the relationships between vegetation activity and temperature factors (Tmean and DTR), we first calculated the first-order partial correlation coefficients between averaged GS NDVI and GS Tmean (RNDVI−Tmean) and DTR (RNDVI−DTR) for each of the 17-year moving windows from 1982–1998 to 1999–2015 (i.e., 1982–1998, 1983–1999, . . . ,1999–2015), with the sum of precipitation as the control variable. Specifically, the first-order partial correlation coefficient was calculated by the correlation coefficient. The correlation coefficient was calculated by equation 1.
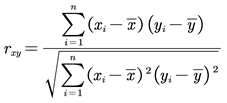 where x and y are the variables for which the correlation coefficient needs to be calculated.
where x and y are the variables for which the correlation coefficient needs to be calculated.
Therefore, the first-order correlation coefficient can be calculated by equation 2.
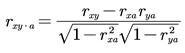 where rxy, rxa and rya are the correlation coefficients and a is the control variable.
where rxy, rxa and rya are the correlation coefficients and a is the control variable.
Accordingly, there were 18 moving windows with centers ranging from 1990 to 2007, and 18 corresponding values for RNDVI-Tmean and RNDVI-DTR. These values were then regressed, using a unary linear regression model (equation 2), against the centers of the moving windows to determine their respective temporal trends (Zhao et al., 2022).
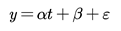 where α is the regression coefficient; t is a year in the time series; β is the regression constant, and ε is the fitted residual.
where α is the regression coefficient; t is a year in the time series; β is the regression constant, and ε is the fitted residual.
Least squares fitting was employed to determine trends at both the inter-annual and gridded scales, with statistical significance considered at the 5% (or 1%) level.
A similar approach was used to calculate temporal changes in the relationships between vegetation activity and temperature during the different seasons. In addition, to ensure the robustness of the research results, the analysis was also conducted with the 15-year and 19-year sliding windows as described above.
3. Results
3.1. Trends of correlations between vegetation activity and temperature factors
3.1.1. Inter-annual changes in RNDVI-Tmean and RNDVI-DTR
For the entire period 1982–2015, the inter-annual variation of the growing season (April to October, GS) NDVI across the boreal regions of the Northern Hemisphere is significantly correlated with the corresponding GS Tmean (RNDVI-Tmean = 0.45, p < 0.01), but not with the corresponding GS DTR (R = 0.37, p = 0.07). At the regional scale, however, the partial correlation coefficients between the GS averaged NDVI and mean daily temperature showed a substantial temporal trend during the past 34 years (Figure 1). The RNDVI−Tmean was about 0.61 (p < 0.05) for period 1982-1997, and then generally decreased to about 0.31 (p > 0.05) for period 1999-2015 (Figure 1c). Similarly, the RNDVI−DTR was about 0.44 (p > 0.05) for period 1982-1997, and then decreased to about -0.20 (p > 0.05) for period 1999-2015 (Figure 1d).
Specifically, RNDVI−Tmean decreased significantly at rates of −0.09/10a (p < 0.01, R2 = 0.44), −0.15/10a (p < 0.01, R2 = 0.47) and −0.08/10a (p < 0.01, R2 = 0.45) with the 15-year, 17-year and 19-year moving windows, respectively (Figure 1a, c, d). In addition, the RNDVI−DTR also decreased significantly at rates of −0.31/10a (p < 0.01, R2 = 0.63), −0.34/10a (p < 0.01, R2 = 0.47) and −0.24/10a (p < 0.01, R2 = 0.45) with the 15-year, 17-year and 19-year moving windows, respectively (Figure 1b, d, f). Overall, at the regional scale, both RNDVI−Tmean and RNDVI−DTR showed a weakening trend during the last 34 years.
3.1.2. Intra-annual changes in RNDVI-Tmean and RNDVI-DTR
At the regional scale, both RNDVI−Tmean and RNDVI−DTR showed distinct patterns among spring, summer, and autumn. During spring, the annual variation of the mean NDVI for the entire period of 1982–2015 is significantly correlated with the corresponding Tmean (RNDVI-Tmean = 0.50, p < 0.01) and the corresponding DTR (RNDVI-DTR = 0.51, p < 0.01). The RNDVI−Tmean was about 0.29 (p > 0.05) for period 1982-1997, and then generally increased to about 0.76 (p < 0.01) for the final window (1999–2015) (Figure 2c). Similarly, we also found that the partial correlation coefficient between spring mean NDVI and DTR significantly increased from 0.34 (p > 0.05) in the first half of the period (1982–1998) to about 0.55 (p < 0.01) in the second half (1999–2015) (Figure 2d).
The RNDVI−Tmean for spring increased significantly at rates of 0.24/10a (p < 0.01, R2 = 0.66), 0.22/10a (p < 0.01, R2 = 0.57) and 0.20/10a (p < 0.01, R2 = 0.45) with the 15-year, 17-year and 19-year moving windows, respectively (Figure 2a, c, d). In addition, the RNDVI−DTR also increased significantly at rates of 0.17/10a (p < 0.01, R2 = 0.61), 0.16/10a (p < 0.01, R2 = 0.52) and 0.18/10a (p < 0.01, R2 = 0.56) with the 15-year, 17-year and 19-year moving windows, respectively (Figure 1b, d, f). Overall, at the regional scale, both RNDVI−Tmean and RNDVI−DTR showed a strengthening trend during the last 34 years.
During summer, the annual variation of the mean NDVI for the entire period of 1982–2015 is significantly correlated with the corresponding Tmean (RNDVI-Tmean = 0.77, p < 0.01), but not with the corresponding DTR (RNDVI-DTR = 0.19, p = 0.30). The RNDVI−Tmean was about 0.29 (p > 0.05) for the period 1982–1998, and then generally decreased to about 0.76 (p < 0.01) for the final window (1999–2015). Similarly, the summer RNDVI-DTR decreased from 0.10 (p > 0.05) in the first half of the period (1982–1998) to about −0.13 (p > 0.05) in the second half (1999–2015) (Figure 2d).
Figure 3 shows that the RNDVI−Tmean for summer decreased significantly at rates of −0.26/10a (p < 0.01, R2 = 0.67), −0.22/10a (p < 0.01, R2 = 0.69) and −0.36/10a (p < 0.01, R2 = 0.74) with the 15-year, 17-year and 19-year moving windows, respectively (Figure 3a, c, d). In addition, the RNDVI−DTR also decreased significantly at rates of −0.24/10a (p < 0.01, R2 = 0.44), −0.28/10a (p < 0.01, R2 = 0.44) and −0.35/10a (p < 0.01, R2 = 0.48) with the 15-year, 17-year and 19-year moving windows, respectively (Figure 1b, d, f). Therefore, both summer RNDVI−Tmean and RNDVI−DTR showed a weakening trend during the last 34 years at the regional scale.
During autumn, the annual variation of the mean NDVI for the entire period of 1982–2015 are significantly correlated with the corresponding Tmean (RNDVI-Tmean = 0.38, p < 0.05), but not with the corresponding DTR (RNDVI-DTR = 0.15, p > 0.05). The autumn RNDVI−Tmean was about 0.51 (p < 0.05) for period 1982-1997, and then generally decreased to about 0.24 (p > 0.05) for period 1999-2015 (Figure 4c). Similarly, the RNDVI−DTR was about 0.10 (p > 0.05) for period 1982-1997, and then decreased to about −0.13 (p > 0.05) for period 1999-2015 (Figure 4d).
We also found that both autumn RNDVI−Tmean and RNDVI−DTR showed a weakening trend during the last 34 years at the regional scale. Figure4 shows that the autumn RNDVI−Tmean decreased significantly at rates of −0.18/10a (p < 0.01, R2 = 0.36), −0.21/10a (p < 0.01, R2 = 0.37) and −0.22/10a (p < 0.05, R2 = 0.35) with the 15-year, 17-year and 19-year moving windows, respectively (Figure 4a, c, d). In addition, the RNDVI−DTR also decreased significantly at rates of −0.16/10a (p < 0.01, R2 = 0.44) and −0.15/10a (p < 0.05, R2 = 0.34) with the 15-year and 17-year moving windows, respectively (Figure 4b, d).
3.2. Spatial patterns of the trends in the correlations between vegetation activity and temperature factors
3.2.1. Inter-annual patterns of RNDVI-Tmean and RNDVI-DTR
Our results indicate that the spatial pattern of the temporal dynamics in the partial correlation coefficients between NDVI and both Tmean and DTR exhibits a high level of consistency when using three different sliding windows. For the 15-year moving window, in most areas (59.77%), RNDVI−Tmean for the GS showed a downward trend, with statistical significance (p < 0.05) for 41.22% of the boreal regions (Figure 5a). Similarly, in most areas (59.10 % of boreal regions), RNDVI−DTR for the GS showed a downward trend, with statistical significance (p < 0.05) for 40.35% of the boreal regions (Figure 5b). For the 17-year moving window, in most areas (62.41%), RNDVI−Tmean for the GS showed a downward trend, with statistical significance (p < 0.05) for 44.40% of the boreal regions (Figure 5c). Similarly, in most areas (59.06 % of boreal regions), RNDVI−DTR for the GS showed a downward trend, with statistical significance (p < 0.05) for 41.21% of the boreal regions (Figure 5d). For the 19-year moving window, in most areas (60.03%), RNDVI−Tmean for the GS showed a downward trend, with statistical significance (p < 0.05) for 40.16% of the boreal regions (Figure 5e). Similarly, in most areas (58.60% of boreal regions), RNDVI−DTR for the GS showed a downward trend, with statistical significance (p < 0.05) for 39.01% of the boreal regions (Figure 5f).
Overall, regarding RNDVI−Tmean, pixels with a significant decreasing trend were primarily distributed in eastern Eurasia and the high latitudes of North America (Figure 5a, c, e). However, regions where RNDVI−Tmean exhibited a significant upward trend were concentrated in the southern region of North America and the eastern region of Europe (Figure 5a, c, e). Moreover, the pixels of RNDVI−DTR with significant decreasing trend were mainly distributed in western Eurasia and the northwestern part of North America (Figure 5b, d, f). In contrast, regions where RNDVI−DTR exhibited a significant upward trend were concentrated in the southern region of North America and the eastern region of Eurasia (Figure 5b, d, f).
3.2.2. Intra-annual patterns of RNDVI-Tmean and RNDVI-DTR
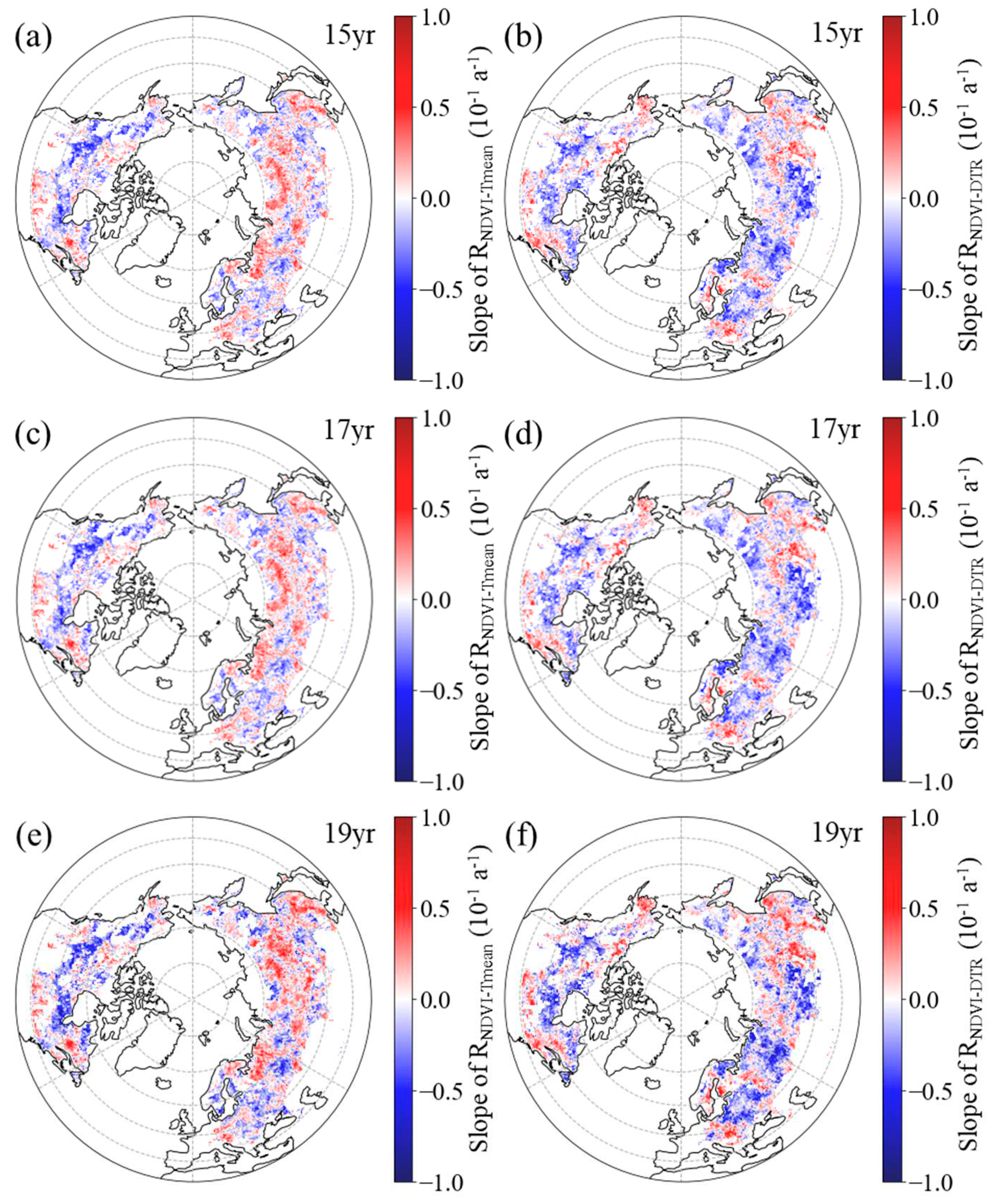
In spring, the RNDVI−Tmean exhibited a positive temporal trend in 63.28%, 61.66%, and 61.83% of boreal regions for the 15-year, 17-year, and 19-year moving windows, respectively (Figure 6a, c, e). Within boreal regions, 45.67%, 43.01%, and 42.04% of pixels showed a significant positive trend for the corresponding moving windows, mainly distributed across the southern regions of Eurasia and the central part of North America (Figure 6a, c, e). For the 15-year, 17-year, and 19-year moving windows, negative trends of RNDVI−Tmean were found in the remaining boreal regions, comprising 36.72%, 38.34%, and 38.17%, respectively (Figure 6a, c, e). Among these regions, the trend was found to be significant (p < 0.05) in 22.09%, 23.24%, and 22.14% of the areas, respectively, with these pixels primarily located in the central and eastern parts of Eurasia and the northern regions of North America (Figure 6a, c, e).
However, in most areas (53.70%, 53.93% and 55.34% of the boreal regions for the 15-year, 17-year, and 19-year moving windows, respectively), the spring RNDVI−DTR had a downward trend (Figure 6b, d, f). Specifically, for the 15-year, 17-year, and 19-year moving windows, the downward trend was found to be statistically significant (p < 0.05) in 33.99%, 34.98%, and 34.53% of the boreal regions, respectively, with these pixels primarily located in the central and western parts of Eurasia and the northern regions of North America (Figure 6b, d, f). Positive trends of RNDVI−DTR were found in the other 46.30%, 46.07% and 44.66% of the boreal regions for the 15-year, 17-year, and 19-year moving windows, respectively (Figure 6b, d, f). Additionally, across the boreal regions, 45.67%, 43.01%, and 42.04% of pixels showed a significant positive trend for the corresponding moving windows, with these pixels mainly distributed across the eastern parts of Eurasia and the northeastern regions of North America (Figure 6b, d, f).
In summer, the RNDVI−Tmean exhibited a negative temporal trend in 60.16%, 62.39%, and 60.16% of boreal regions for the 15-year, 17-year, and 19-year moving windows, respectively (Figure 7a, c, e). Additionally, across the boreal regions, 41.66%, 45.18%, and 40.62% of pixels showed a significant negative trend for the corresponding moving windows, with these pixels primarily located in the central and western parts of Eurasia and the northeastern regions of North America (Figure 7a, c, e). For the 15-year, 17-year, and 19-year moving windows, positive trends of RNDVI−Tmean were found in the remaining boreal regions, comprising 39.84%, 37.61%, and 39.84%, respectively (Figure 7a, c, e). Among these regions, the positive trend was found to be significant (p < 0.05) in 23.35%, 21.98%, and 22.24% of the areas, respectively, with these pixels mainly located in the eastern parts of Eurasia and the southern regions of North America (Figure 7a, c, e).
Similarly, in most areas (54.26%, 54.78% and 53.49% of the boreal regions for the 15-year, 17-year, and 19-year moving windows, respectively), the spring RNDVI−DTR had a downward trend (Figure 7b, d, f). Within boreal regions, 36.02%, 36.93%, and 34.67% of pixels showed a significant downward trend for the corresponding moving windows, mainly located in the western regions of Eurasia and the northwestern regions of North America (Figure 7b, d, f). Positive trends of RNDVI−DTR were found in the other 45.74%, 45.22% and 46.51% of the boreal regions for the 15-year, 17-year, and 19-year moving windows, respectively (Figure 7b, d, f). Additionally, across the boreal regions, 28.77%, 28.71%, and 28.21% of pixels showed a significant positive trend for the corresponding moving windows, with these pixels mainly located in the eastern regions of Eurasia and the southern regions of North America (Figure 7b, d, f).
In autumn, the RNDVI−Tmean exhibited a negative temporal trend in 50.58%, 51.25%, and 52.19% of boreal regions for the 15-year, 17-year, and 19-year moving windows, respectively (Figure 8a, c, e). Additionally, across the boreal regions, 31.54%, 32.29%, and 32.46% of pixels showed a significant negative trend for the corresponding moving windows, with these pixels mainly located in the western regions of Eurasia and the central regions of North America (Figure 8a, c, e). For the 15-year, 17-year, and 19-year moving windows, positive trends of RNDVI−Tmean were found in the remaining boreal regions, comprising 49.42%, 48.75%, and 47.81%, respectively (Figure 8a, c, e). Among these regions, the positive trend was found to be significant (p < 0.05) in 30.41%, 30.01%, and 29.35% of the areas, respectively, with these pixels primarily located in the central and eastern regions of Eurasia and the southern regions of North America (Figure 8a, c, e).
Similarly, in most areas (60.09%, 60.16% and 57.52% of the boreal regions for the 15-year, 17-year, and 19-year moving windows, respectively), the spring RNDVI−DTR had a downward trend (Figure 8b, d, f). Within boreal regions, 40.77%, 40.93%, and 37.24% of pixels showed a significant downward trend for the corresponding moving windows, mainly located in the central and western regions of Eurasia and the northern regions of North America (Figure 8b, d, f). Positive trends of RNDVI−DTR were found in the other 39.91%, 39.84% and 42.48% of the boreal regions for the 15-year, 17-year, and 19-year moving windows, respectively (Figure 8b, d, f). Additionally, across the boreal regions, 23.04%, 23.47%, and 24.41% of pixels showed a significant positive trend for the corresponding moving windows, with these pixels mainly located in the eastern regions of Eurasia and the southern regions of North America (Figure 8b, d, f).
4. Discussion
Our results show considerable temporal trends in the partial correlation coefficients between GS NDVI and daily mean temperature across the boreal regions over the last 34 years. This variability indicates a weakening relationship between interannual temperature variability and boreal vegetation activity. Varying responses of vegetation activity to global warming can be also found in the results of previous studies of high-latitude or high-altitude areas (Cong et al., 2017; Piao et al., 2014). For example, Cong et al. (2017) found that the relationship between summer NDVI and air temperature for alpine meadow across the Tibetan Plateau showed a decreasing trend. Piao et al. (2014) found that in the Northern Hemisphere, the positive influence of the growing season air temperature on vegetation growth exhibited a weakening trend over the last three decades. Overall, our finding of an interannual change of the response of boreal vegetation activity to air temperature is either a verification of, or a supplement to, previous studies at different scales.
Here, we also analyzed the temporal variation in boreal vegetation response to air temperature on the seasonal scale, which has been neglected in previous studies. We found a significant weakening trend in the relationship between NDVI and Tmean, mainly in the summer and autumn seasons. This timing means that the weakening of the influence of air temperature on vegetation activity during the summer and autumn seasons dominates the overall weakening of the correlation during the GS. In contrast, during the spring, we found an increasing trend in the relationship between NDVI and temperature, suggesting that in the spring the relationship between temperature changes and vegetation growth may have become more closely linked.
The reasons for the changes in vegetation activity response to temperature changes are still difficult to verify due to the complexity and variability of the factors involved. Possible explanations for the weakening trend in vegetation response to temperature during summer are that, under the background of global warming, the temperature in the region may have approached the optimal temperature or physiological and ecological threshold of some plants, or that the vegetation has gradually adapted to the warming environment. Piao et al. (2014) have suggested that the weakening relationship between vegetation activity and temperature changes can be attributed to the increasing frequency of drought in the Northern Hemisphere. Qin et al. (2022) supported this idea and found that the relationship between vegetation activity and temperature during warm years exhibited a significant decreasing trend. However, the temperature rise in spring has led to an earlier start to the growing season for vegetation in the northern high latitudes (Piao et al., 2007). For instance, Piao et al. (2007) found an earlier onset of spring (0.16 days yr-1) during the last two decades across the Northern Hemisphere. The continuous increase in the vegetation GS caused by global warming is almost certainly one reason for the growing enhancement of vegetation response to temperature.
In addition, we found that the effect of the GS diurnal temperature range on vegetation activity also showed a weakening trend. This finding has important implications for refining the understanding of the effects of diurnal asymmetric warming on terrestrial ecosystems (Qin et al., 2022; Zhao et al., 2022). For example, Zhao et al. (2022) found that the strength of the relationship between the greenness of high-latitude vegetation and daytime warming exhibited a weakening trend at the inter-annual scale. However, in most of the boreal regions of North America, there was an increasing trend in the correlation between nighttime warming and vegetation activity (Zhao et al., 2022).
Multiple uncertainties still remain in understanding the temporal dynamics of boreal vegetation activity response to temperature. A few indirect factors, such as fire disturbance (Zhao et al., 2021; Zheng et al., 2021), insect damage (Kurz et al., 2008), spring frosts, land use and land management changes (Edith and Xue, 2020; Epstein et al., 2013) and changes in nutrient use efficiency also have potential to weaken the observed relationship between vegetation activity and temperature change. These factors could also explain some of the differences in the spatial patterns of temporal trends of NDVI–temperature correlations.
5. Conclusions
Our study analyzes the temporal trends in boreal vegetation activity responses to mean daily temperature and diurnal temperature range temperature changes at the regional and seasonal scales over the past 34 years. We found that the GS RNDVI−Tmean showed a significant decreasing (-0.15/10a) trend during the last three decades at the regional scale, which was mainly due to decreasing RNDVI−Tmean in summer (-0.31/10a) and autumn (-0.21/10a). In contrast, RNDVI-Tmean for spring increased significantly at the rate of 0.22/10a, suggesting that the relationship between mean daily temperature changes and vegetation activity may have become more closely linked in the spring. Similarly to RNDVI−Tmean, the GS RNDVI−DTR also showed a significant decreasing (-0.34/10a) trend during the last three decades at the regional scale. This was mainly due to decreasing RNDVI−DTR in summer (-0.28/10a) and autumn (-0.12/10a). In terms of spatial patterns, the areas showing a significantly negative trend in GS RNDVI−Tmean and RNDVI−DTR accounted for approximately 44.40% and 41.21% of the boreal region, respectively. In contrast, the areas that exhibited a significant increasing trend in GS RNDVI−Tmean and RNDVI−DTR accounted for only approximately 14.65% and 13.59% of the boreal region, respectively. At the seasonal scale, the area showing a significantly negative trend in the partial correlation coefficient between vegetation activity and temperature factors was bigger than the area with a significant positive trend for both summer and autumn. In spring, however, the area with a significant upward trend in RNDVI−Tmean and RNDVI−DTR was greater than that with a significant downward trend.
Author Contributions
Zhao H drafted the article; Jin N and Fu G performed data analysis and interpreted the results; Zhao J and Xiang K designed the statistical analysis; Wang X and Zhao J revised the article; all authors contributed to further drafts.
Acknowledgments
We sincerely appreciate the editors and anonymous reviewers for their valuable comments. This work was supported by the Natural Science Foundation of Guangdong Province (2020A1515110423), the Science and Technology Research Project of Guangdong Meteorological Administration, (GRMC2021M15) and the Open Research Fund of the National Engineering Research Center for Agro-Ecological Big Data Analysis & Application, Anhui University (AE202211).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bonan, G.B. , Pollard, D., Thompson, S.L. Effects of boreal forest vegetation on global climate. Nature 1992, 359, 716–718. [Google Scholar]
- Chen, X.; Yang, Y. Observed earlier start of the growing season from middle to high latitudes across the Northern Hemisphere snow-covered landmass for the period 2001–2014. Environmental Research Letters 2020, 15. [Google Scholar] [CrossRef]
- Cong, N.; Shen, M.; Yang, W.; Yang, Z.; Zhang, G.; Piao, S. Varying responses of vegetation activity to climate changes on the Tibetan Plateau grassland. Int J Biometeorol 2017, 61, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.; Zhang, H.; Zhao, J.; Guo, X.; Biniyaz, E.; Zhao, H.; Guo, D.; Wu, R.; Li, H. Diverse variations in middle and high latitudes of the Northern Hemisphere spring phenology sensitivity to diurnal temperature during 1982–2015. International Journal of Climatology 2022, 42, 9385–9403. [Google Scholar] [CrossRef]
- Du, Z.; Liu, X.; Wu, Z.; Zhang, H.; Zhao, J. Responses of Forest Net Primary Productivity to Climatic Factors in China during 1982-2015. Plants (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Edith, B.A.I.; Xue, B. A review of influences of land use and land cover change on ecosystems. Chinese Journal of Plant Ecology 2020, 44, 543–552. [Google Scholar] [CrossRef]
- Epstein, H.E.; Myers-Smith, I.; Walker, D.A. Recent dynamics of arctic and sub-arctic vegetation. Environmental Research Letters 2013, 8. [Google Scholar] [CrossRef]
- Gauthier, S.; Bernier, P.; Kuuluvainen, T.; Shvidenko, A.Z.; Schepaschenko, D.G. Boreal forest health and global change. Science 2015, 349, 819–822. [Google Scholar] [CrossRef]
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci Data 2020, 7, 109. [Google Scholar] [CrossRef]
- Huang, M.; Piao, S.; Janssens, I.A.; Zhu, Z.; Wang, T.; Wu, D.; Ciais, P.; Myneni, R.B.; Peaucelle, M.; Peng, S.; et al. Velocity of change in vegetation productivity over northern high latitudes. Nat Ecol Evol 2017, 1, 1649–1654. [Google Scholar] [CrossRef]
- Huang, Y.; Jiang, N.; Shen, M.; Guo, L. Effect of preseason diurnal temperature range on the start of vegetation growing season in the Northern Hemisphere. Ecological Indicators 2020, 112. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain pine beetle and forest carbon feedback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.; Piao, S.; Chen, A.; Wang, K.; Li, X.; Buermann, W.; Huntingford, C.; Penuelas, J.; Xu, H.; Myneni, R.B. Seasonal biological carryover dominates northern vegetation growth. Nat Commun 2021, 12, 983. [Google Scholar] [CrossRef]
- Peng, S.; Piao, S.; Ciais, P.; Myneni, R.B.; Chen, A.; Chevallier, F.; Dolman, A.J.; Janssens, I.A.; Penuelas, J.; Zhang, G.; et al. Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature 2013, 501, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Global Biogeochemical Cycles 2007, 21, GB3018. [Google Scholar] [CrossRef]
- Piao, S.; Nan, H.; Huntingford, C.; Ciais, P.; Friedlingstein, P.; Sitch, S.; Peng, S.; Ahlstrom, A.; Canadell, J.G.; Cong, N.; et al. Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nat Commun 2014, 5, 5018. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Wang, X.; Ciais, P.; Zhu, B.; Wang, T.A.O.; Liu, J.I.E. Changes in satellite-derived vegetation growth trend in temperate and boreal Eurasia from 1982 to 2006. Global Change Biology 2011, 17, 3228–3239. [Google Scholar] [CrossRef]
- Qin, M.; Jin, N.; Zhao, J.; Feng, M.; Wang, C. Dynamics of the Response of Vegetation Activity to Air Temperature Change in Temperate China. Atmosphere 2022, 13. [Google Scholar] [CrossRef]
- Shen, X.; Liu, B.; Henderson, M.; Wang, L.; Wu, Z.; Wu, H.; Jiang, M.; Lu, X. Asymmetric effects of daytime and nighttime warming on spring phenology in the temperate grasslands of China. Agricultural and Forest Meteorology 2018, 259, 240–249. [Google Scholar] [CrossRef]
- Sulla-Menashe, D.; Woodcock, C.E.; Friedl, M.A. Canadian boreal forest greening and browning trends: an analysis of biogeographic patterns and the relative roles of disturbance versus climate drivers. Environmental Research Letters 2018, 13. [Google Scholar] [CrossRef]
- Sun, X.; Ren, G.; You, Q.; Ren, Y.; Xu, W.; Xue, X.; Zhan, Y.; Zhang, S.; Zhang, P. Global diurnal temperature range (DTR) changes since 1901. Climate Dynamics 2018, 52, 3343–3356. [Google Scholar] [CrossRef]
- Tan, J.; Piao, S.; Chen, A.; Zeng, Z.; Ciais, P.; Janssens, I.A.; Mao, J.; Myneni, R.B.; Peng, S.; Penuelas, J.; et al. Seasonally different response of photosynthetic activity to daytime and night-time warming in the Northern Hemisphere. Glob Chang Biol 2015, 21, 377–387. [Google Scholar] [CrossRef]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. International Journal of Remote Sensing 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Wen, Y.; Liu, X.; Pei, F.; Li, X.; Du, G. Non-uniform time-lag effects of terrestrial vegetation responses to asymmetric warming. Agricultural and Forest Meteorology 2018, 252, 130–143. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Cook, B.I.; Allen, J.M.; Crimmins, T.M.; Betancourt, J.L.; Travers, S.E.; Pau, S.; Regetz, J.; Davies, T.J.; Kraft, N.J.; et al. Warming experiments underpredict plant phenological responses to climate change. Nature 2012, 485, 494–497. [Google Scholar] [CrossRef]
- Wu, Z.; Dijkstra, P.; Koch, G.W.; PeÑUelas, J.; Hungate, B.A. Responses of terrestrial ecosystems to temperature and precipitation change: a meta-analysis of experimental manipulation. Global Change Biology 2011, 17, 927–942. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, L.; Hou, X.; Li, G.; Tian, Q.; Chan, E.; Ciais, P.; Yu, Q.; Yue, C. Fire Regime Impacts on Postfire Diurnal Land Surface Temperature Change Over North American Boreal Forest. Journal of Geophysical Research: Atmospheres 2021, 126. [Google Scholar] [CrossRef]
- Zhao, J.; Xiang, K.; Wu, Z.; Du, Z. Varying Responses of Vegetation Greenness to the Diurnal Warming across the Global. Plants (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Zheng, B.; Ciais, P.; Chevallier, F.; Chuvieco, E.; Chen, Y.; Yang, H. Increasing forest fire emissions despite the decline in global burned area. Science Advances 2021, 7, eabh2646. [Google Scholar] [CrossRef]
Figure 1.
Variations in the sliding partial correlation coefficients between growing season mean NDVI and daily mean temperature (a, c, e, RNDVI-Tmean) and diurnal temperature range (b, d, f, RNDVI-DTR) after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows. The black lines represent the inter-annual change trend in the partial correlation coefficients. R2 represents the coefficient of determination. Shading denotes 95% prediction intervals.
Figure 1.
Variations in the sliding partial correlation coefficients between growing season mean NDVI and daily mean temperature (a, c, e, RNDVI-Tmean) and diurnal temperature range (b, d, f, RNDVI-DTR) after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows. The black lines represent the inter-annual change trend in the partial correlation coefficients. R2 represents the coefficient of determination. Shading denotes 95% prediction intervals.
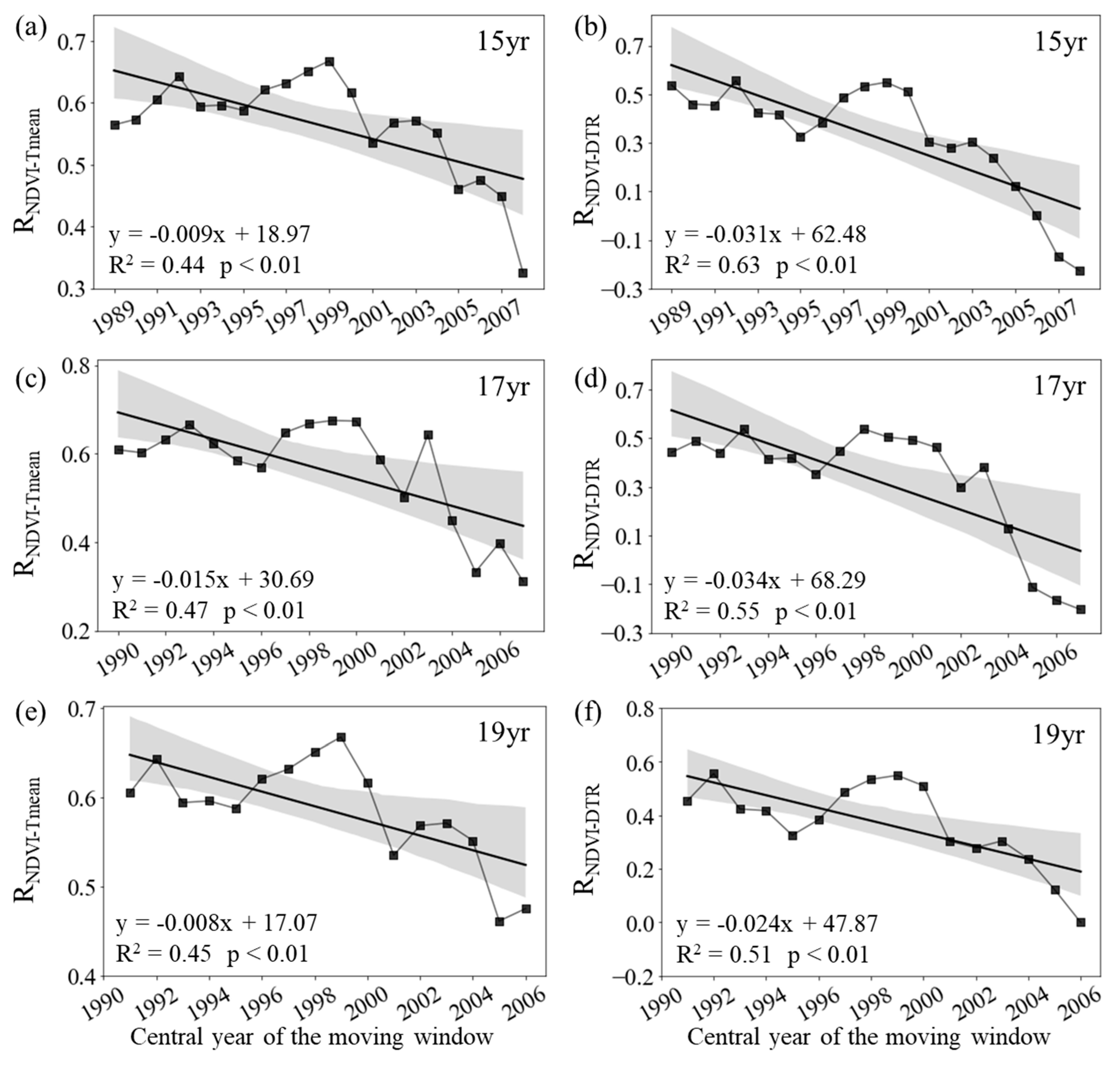
Figure 2.
Variations in the sliding partial correlation coefficients between spring mean NDVI and daily mean temperature (a, RNDVI-Tmean) and diurnal temperature range (b, RNDVI-DTR) after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows. The black lines represent the inter-annual change trend in partial correlation coefficients. R2 represents the coefficient of determination. Shading denotes 95% prediction intervals.
Figure 2.
Variations in the sliding partial correlation coefficients between spring mean NDVI and daily mean temperature (a, RNDVI-Tmean) and diurnal temperature range (b, RNDVI-DTR) after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows. The black lines represent the inter-annual change trend in partial correlation coefficients. R2 represents the coefficient of determination. Shading denotes 95% prediction intervals.
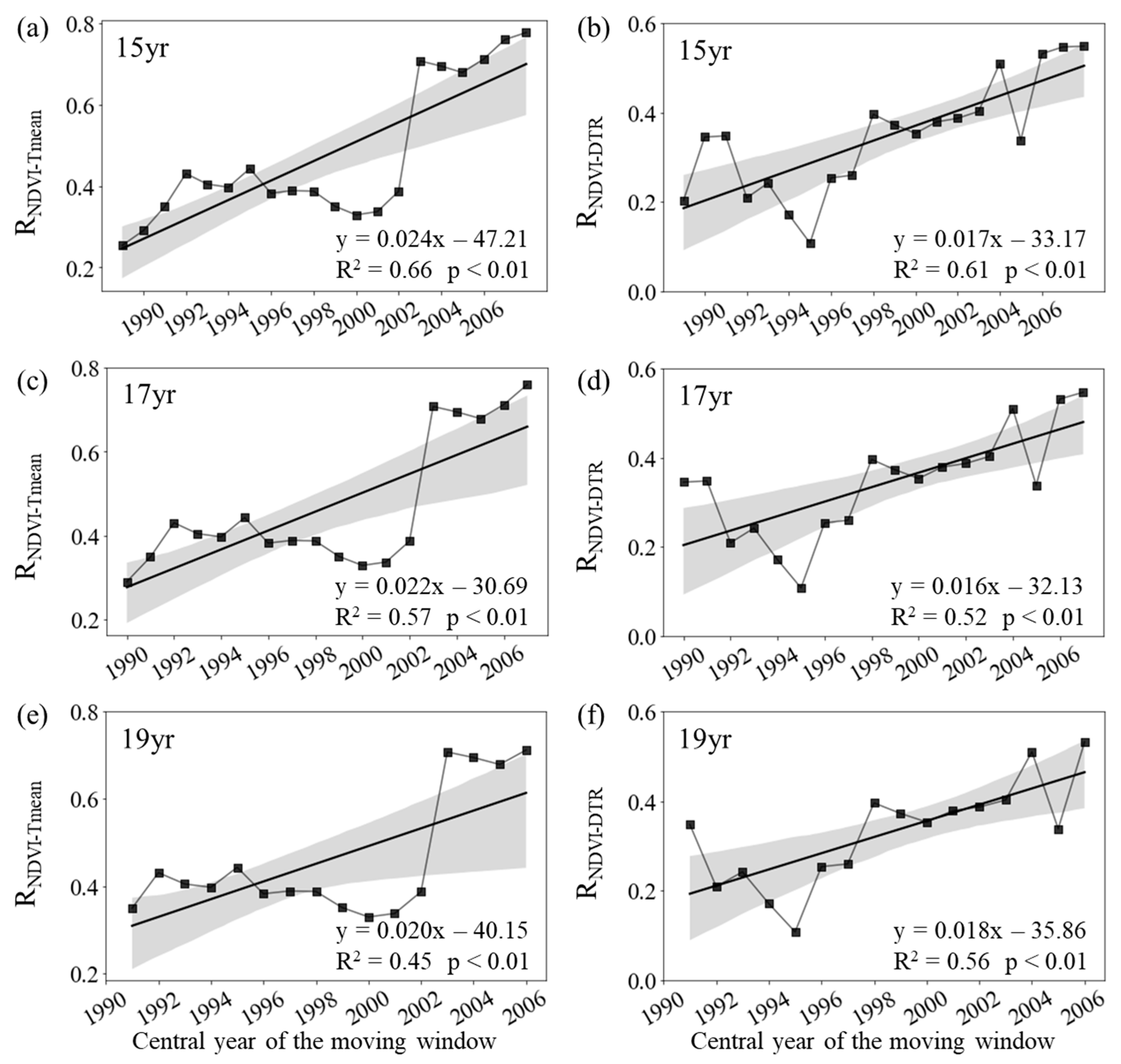
Figure 3.
Variations in the sliding partial correlation coefficients between summer mean NDVI and daily mean temperature (a, RNDVI-Tmean) and diurnal temperature range (b, RNDVI-DTR) after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows. The black lines represent the inter-annual change trend in the partial correlation coefficients. R2 represents the coefficient of determination. Shading denotes 95% prediction intervals.
Figure 3.
Variations in the sliding partial correlation coefficients between summer mean NDVI and daily mean temperature (a, RNDVI-Tmean) and diurnal temperature range (b, RNDVI-DTR) after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows. The black lines represent the inter-annual change trend in the partial correlation coefficients. R2 represents the coefficient of determination. Shading denotes 95% prediction intervals.
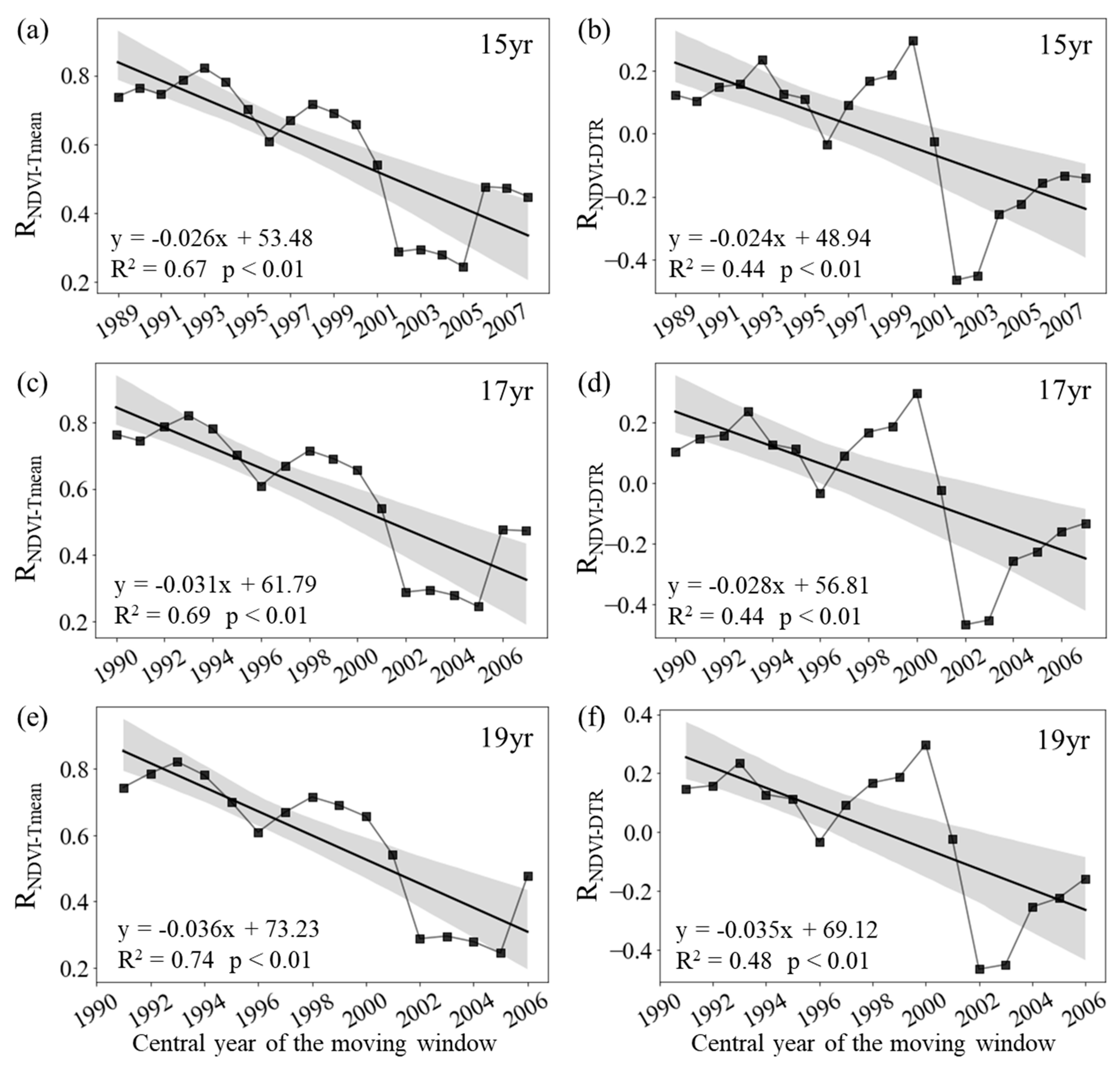
Figure 4.
Variations in the sliding partial correlation coefficients between autumn mean NDVI and daily mean temperature (a, c, e, RNDVI-Tmean) and diurnal temperature range (b, d, f, RNDVI-DTR) after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows. The black lines represent the inter-annual change trend in the partial correlation coefficients. R2 represents the coefficient of determination. Shading denotes 95% prediction intervals.
Figure 4.
Variations in the sliding partial correlation coefficients between autumn mean NDVI and daily mean temperature (a, c, e, RNDVI-Tmean) and diurnal temperature range (b, d, f, RNDVI-DTR) after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows. The black lines represent the inter-annual change trend in the partial correlation coefficients. R2 represents the coefficient of determination. Shading denotes 95% prediction intervals.
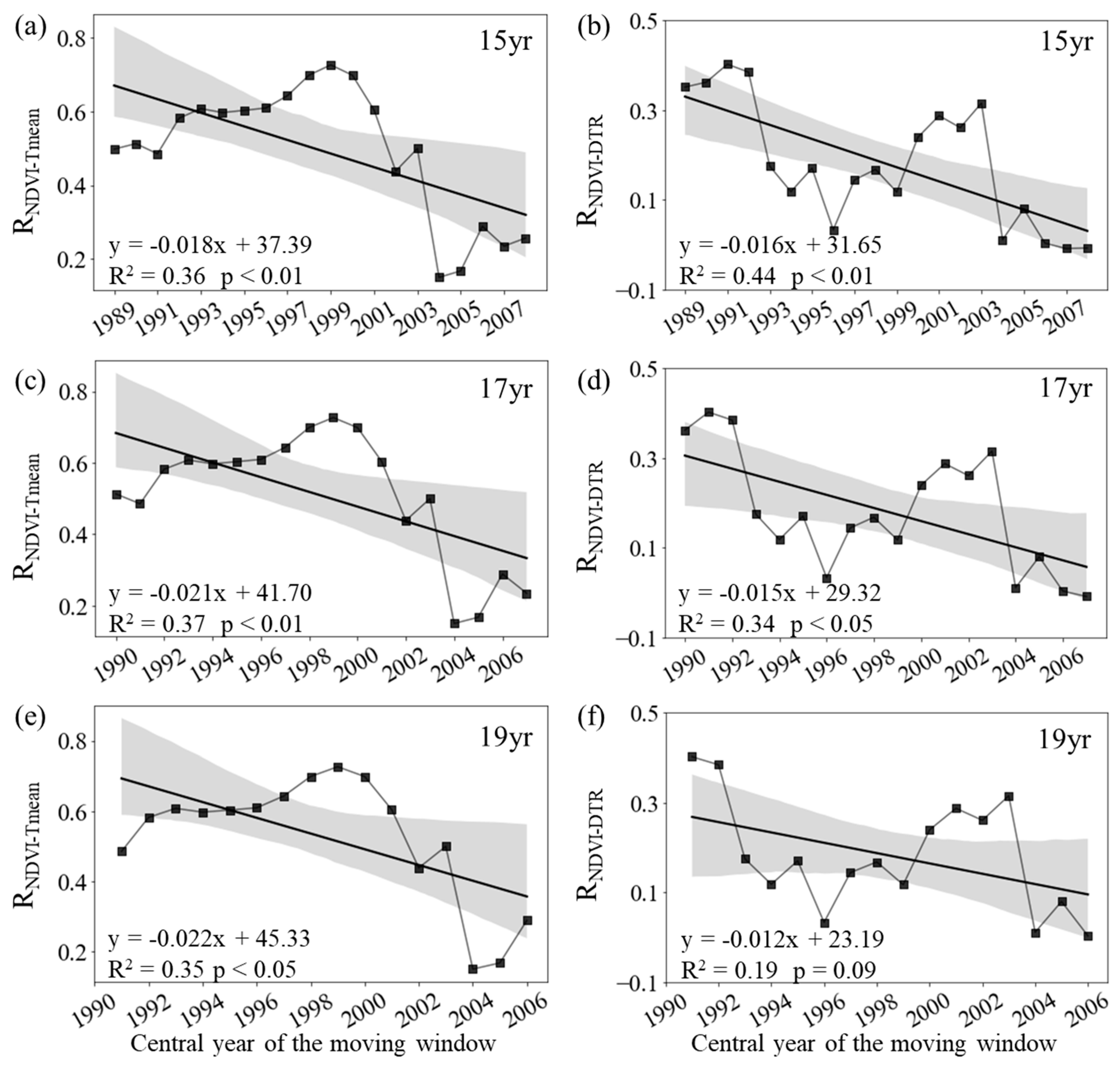
Figure 5.
Spatial distribution of the temporal trend of partial correlation coefficient between growing season mean NDVI and daily mean temperature (a, c, e, RNDVI−Tmean) and diurnal temperature range (b, d, f, RNDVI-DTR) during 1982–2015 after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows.
Figure 5.
Spatial distribution of the temporal trend of partial correlation coefficient between growing season mean NDVI and daily mean temperature (a, c, e, RNDVI−Tmean) and diurnal temperature range (b, d, f, RNDVI-DTR) during 1982–2015 after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows.
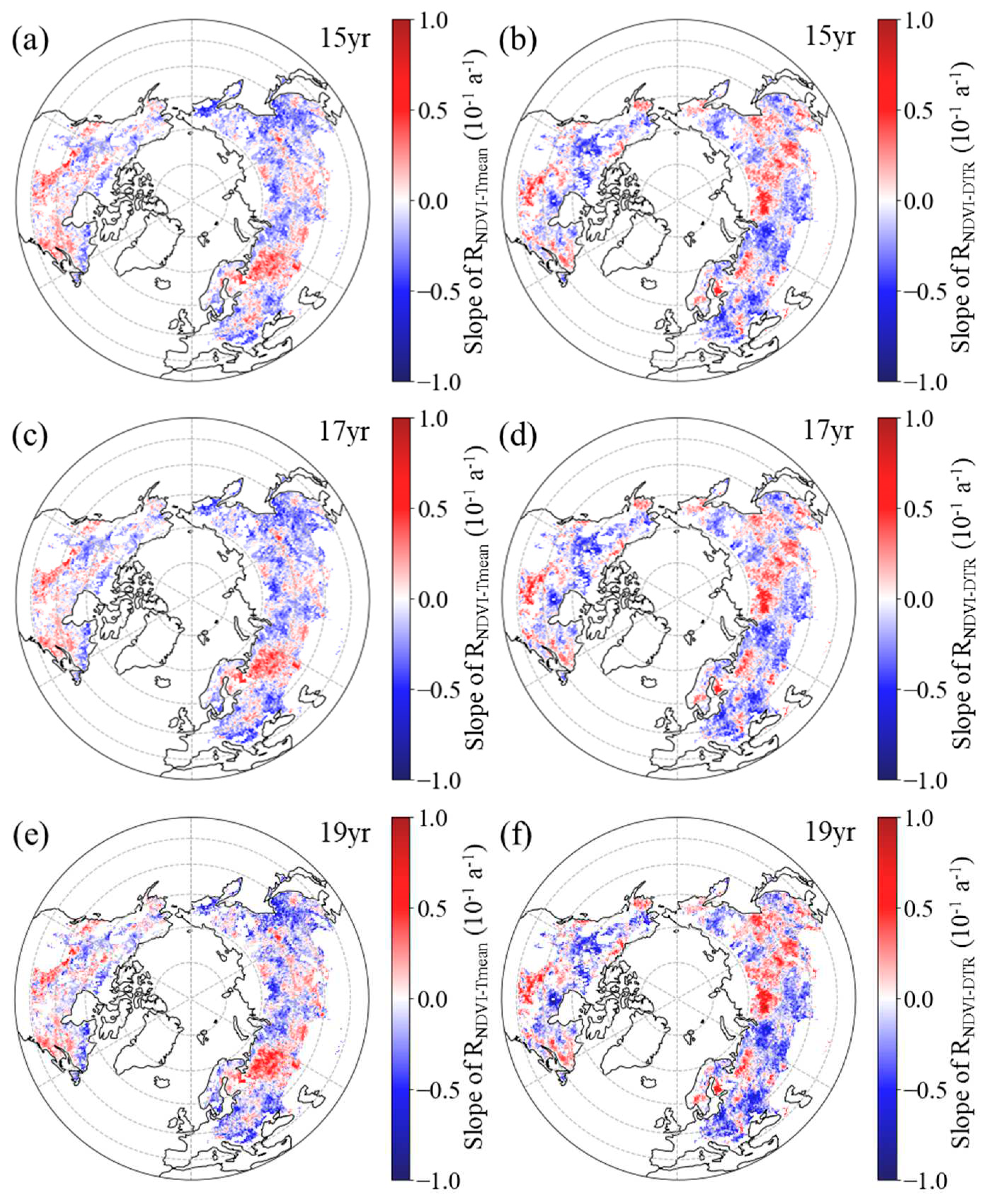
Figure 6.
Spatial distribution of the temporal trend of partial correlation coefficient between spring NDVI and daily mean temperature (a, c, e, RNDVI−Tmean) and diurnal temperature range (b, d, f, RNDVI-DTR) for the period 1982–2015 after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows.
Figure 6.
Spatial distribution of the temporal trend of partial correlation coefficient between spring NDVI and daily mean temperature (a, c, e, RNDVI−Tmean) and diurnal temperature range (b, d, f, RNDVI-DTR) for the period 1982–2015 after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows.
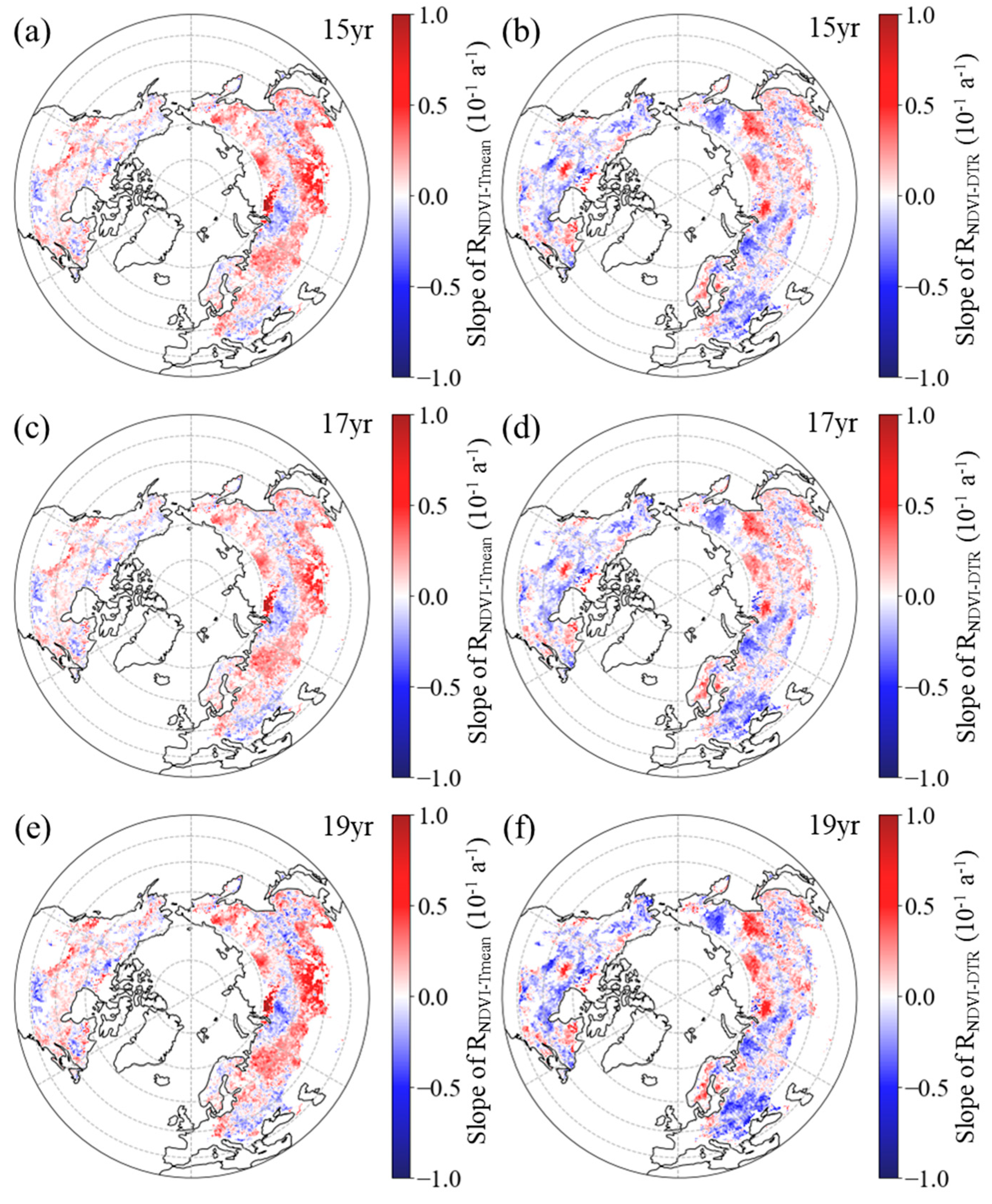
Figure 7.
Spatial distribution of the temporal trend of partial correlation coefficient between summer NDVI and daily mean temperature (a, c, e, RNDVI−Tmean) and diurnal temperature range (b, d, f, RNDVI-DTR) during 1982–2015 after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows.
Figure 7.
Spatial distribution of the temporal trend of partial correlation coefficient between summer NDVI and daily mean temperature (a, c, e, RNDVI−Tmean) and diurnal temperature range (b, d, f, RNDVI-DTR) during 1982–2015 after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows.
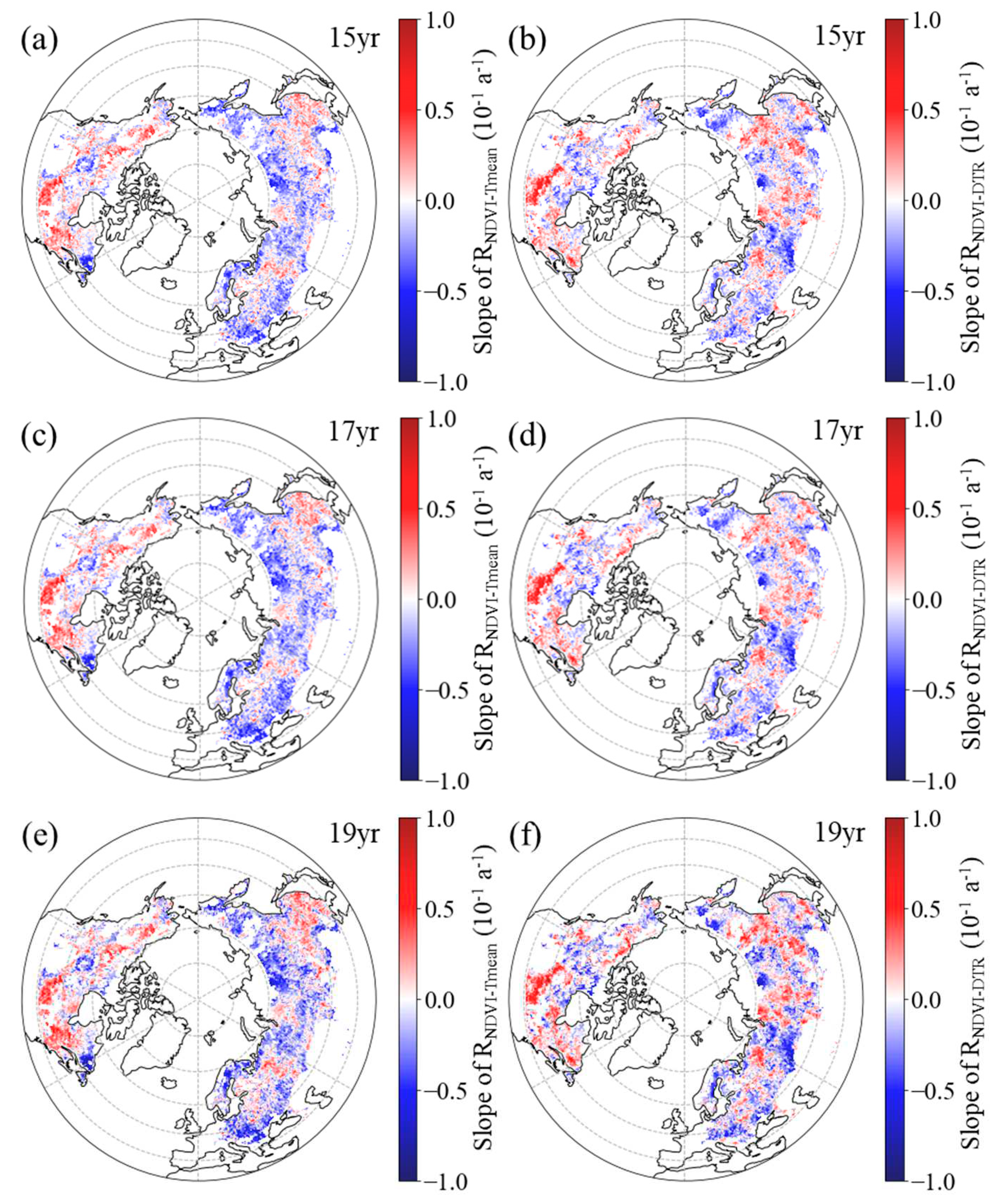
Figure 8.
Spatial distribution of the temporal trend of partial correlation coefficient between autumn NDVI and daily mean temperature (a, c, e, RNDVI−Tmean) and diurnal temperature range (b, d, f, RNDVI-DTR) during 1982–2015 after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows.
Figure 8.
Spatial distribution of the temporal trend of partial correlation coefficient between autumn NDVI and daily mean temperature (a, c, e, RNDVI−Tmean) and diurnal temperature range (b, d, f, RNDVI-DTR) during 1982–2015 after applying 15-year (a, b), 17-year (c, d) and 19-year (e, f) moving windows.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



