Submitted:
22 May 2023
Posted:
24 May 2023
You are already at the latest version
Abstract
The cytoplasmic membrane is the fundamental component of all living cells, which participates in various physiological processes, such as material exchange, stress response, cell recognition, signal transduction, cellular immunity, apoptosis, pathogenicity, etc. The normal function of a cytoplasmic membrane requires stable organization of transmembrane protein-lipid microdomains, transmembrane protein-cell wall microdomains, and cytoskeleton-transmembrane protein microdomains. Here, we review the mechanisms and functions of various membrane lipid components, fatty acid content and saturation, membrane curvature, and cell wall and cytoskeleton in plasma membrane homeostasis affecting the pathogenicity of pathogenic fungi. Pathogenic fungi maintains plasma membrane homeostasis and contributes to fungal virulence by maintaining plasma membrane assembly, structural and functional integrity of pathogenic fungi at various stages of cell development through interactions among lipid components of cytoplasmic membranes, transmembrane proteins, cytoskeleton and cell wall components, etc.
Keywords:
Cytoplasmic membrane homeostasis
; Pathogenicity
; Pathogenic fungi
; Membrane lipid metabolism
; Membrane curvature
; Transmembrane protein
; Cell wall components
; Cytoskeleton
; Growth and development
; Host infection
1. Introduction
The cell plasma membrane is the defining feature of all cell life forms [1]. It is mainly composed of lipids and proteins, and the amphiphilic lipid bilayer forms the overall framework of the cell plasma membrane [2,3]. The cytoplasmic membrane determines both the physical properties and structural integrity of the cell [4].
The plasma membrane is the key barrier for conidial survival in nature, and any damage to the membrane will lead to the loss of conidial viability [5]. Fungal plasma membranes help early recognition of nutrient availability, impending stress, or pathogen attack [6]. Although the mechanism and function of the plasma membrane in yeast have been reviewed [7], the relevance of cytoplasmic membrane homeostasis has not been discussed much. In pathogenic fungi, the status of cytoplasmic membrane homeostasis contributes to fungi virulence. At present, there is no review about the involvement of cytoplasmic membrane homeostasis in the pathogenicity of pathogenic fungi.
The cell plasma membrane not only determines the cell structure but also affects various cell activities based on cytoplasmic membrane homeostasis, which refers to the structural and functional integrity and fluidity of the cytoplasmic membrane. In this review, we summarize the literature related to cytoplasmic membrane homeostasis and its effect on fungi pathogenicity by discussing the role of fungal cytoplasmic membrane constituents including transmembrane protein-lipid microdomains, cell wall-lipid microdomains, cytoskeleton-transmembrane protein microdomains, and membrane curvature.
2. Effect of lipid metabolism on the pathogenicity of fungi
2.1. cytoplasmic membrane lipid components
The rich and diverse lipid composition of the plasma membrane affects the cytoplasmic membrane homeostasis. Lipid diversity is more prevalent in eukaryotes, indicating the complex role of plasma membrane lipids in various cell functions [8]. The cytoplasmic membrane lipids are usually arranged as lipid bilayers and the membrane lipids or lipid structures can directly participate in various cellular events as messengers or regulators of signal transduction [9,10]. Some lipids, such as phosphatidylserine [11], Phosphatidylcholine [12], ergosterol [13], Phosphatidylinositol-4-phosphate (PI(4)P) [8], C8-desaturated and C9-methylated glycosylceramide (GlcCer) [14], phosphoinositol diphosphate [15] and ceramide [16], etc., tend to accumulate in specific regions (domains) of the cytoplasmic membrane forming membrane/lipid rafts (MLR) [17], which have evolutionarily conserved roles in the integrity and repair of the plasma membrane.
The host mechanism of pathogenic fungi infection is complex. The membrane lipids of cytoplasmic membrane are very important components in the regulation of asexual spore germination mycelium growth and pathogenicity of pathogenic fungi. In Pestalotiopsis microspora, the absence of the choC or choA gene affects the formation of phosphatidyl ethanolamine into phosphatidylcholine, which impedes the development of fungal spores and conidia and destroys the cell wall integrity [12]. Similarly, choline, a precursor of membrane lipids, plays an important role in the mycelial morphology of Aspergillus nidulans [18]. In Candida albicans, PI(4,5) phosphoinositol diphosphate (P-2), a cytoplasmic membrane lipid molecule, influences the polarization and host infection ability of the fungi [15]. Ergosterol in Aspergillus fumigatus directly affects its growth and infection ability, and therefore, it is used as a very effective drug target against fungal infection[13]. In Aspergillus nidulans, C8-desaturated and C9-methylated glycosylceramide (GlcCer), a fungus-specific sphingolipid, affects the growth and virulence of the fungi [14]. Acyl-coA-dependent ceramide synthetase (BarA) regulates cytoskeletal organization and mycelial morphogenesis by controlling ceramide synthesis to form specialized lipid microdomains [16]. Phospholipid phosphatidylinositol 4-phosphate (PI(4)P), which participates in the transport of substances through the membrane, was shown to regulate conidial germination and mycelial growth of the human pathogenic fungus Candida albicans affecting its pathogenicity [19]. In addition, some domains of membrane lipids can recruit specific proteins regulating membrane-protein participation in important cytological events such as cell signal transduction [10]. For example, spectrin, which forms filamentous networks on the cytoplasmic surface of the fungal plasma membrane, participates in the structural integrity of the membrane and controls the lateral mobility of membrane proteins [20]. The structural integrity of the plasma membrane is closely related to the pathogenicity of fungi.
As an indispensable component of the plasma membrane, the above membrane lipid components can participate in maintaining the structural and functional integrity of the plasma membrane, and can also respond to substance exchange, stress response, cell recognition, signal transduction, cellular immunity, and cell apoptosis of the plasma membrane. It makes an important contribution to the biological processes such as vegetative growth, morphological development and host infection of pathogenic fungi. Membrane lipid components are not only an important part of maintaining the structural integrity of the plasma membrane of cells, but also provide energy for the growth and development of pathogenic fungi, making an irreplaceable contribution to the virulence of fungi. Therefore, the membrane lipid composition is one of the important factors affecting cytoplasmic membrane homeostasis and in turn the pathogenicity of pathogenic fungi.
2.2. Degree of unsaturated fatty acids
The content and degree of unsaturated fatty acids (UFA) are one of the important factors determining cytoplasmic membrane integrity and permeability in fungi, which affects cytoplasmic membrane homeostasis and in turn the pathogenicity of pathogenic fungi. Higher unsaturated fatty acids content in the plasma membrane increases its "fluidity", which is conducive to the rotation and movement of functional proteins in the membrane [21]. Most fungi contain both saturated (such as palmitic acid and stearic acid) and unsaturated (such as oleic acid, linoleic acid, and linolenic acid) fatty acids. Ole1 introduces the double bond into saturated acyl-coA to produce monounsaturated fatty acids [22,23]. The anabolic fatty acid biosynthesis pathway is conserved in filamentous fungi [24]. For example, the Ole1 enzyme participates in the synthesis of ultra-long fatty acid chains and plays a crucial role in cytoplasmic membrane homeostasis [25,26]. Ole1 is regulated by upstream transcription factors, such as ER membrane proteins MGA2/SPT23 (yeast), HapX (Beauveria bassiana), etc [27,28,29,30]. In Aspergillus flavus, FarA (a Zn2-Cys6 transcription factor) positively controls stearic acid desaturase genes required for oleic acid synthesis [31]. In addition, Ubx2p is a key upstream regulator of the essential fatty acid desaturase Ole1p. The absence of Ubx2p affects the transcription of OLE1, causing fatty acid desaturation and abnormal nuclear membrane morphology [32]. In Aspergillus giganteus, 2-hydroxyfat n-acyl-δ3(E) -desaturase controls saturation levels of fatty acid chains and interferes with plasma membrane integrity [33]. The absence of BbSAY1 decreases the content of oleic acid and membrane permeability, leading to a significant reduction in the fungal virulence of Bassiana bassiana [34].
Studies have shown that UFA contributes to plasma membrane integrity and determines the ability of cells to respond to environmental stresses such as high salt stress [35]. The fatty acid synthetase β-subunit dehydrase (FAS1) gene in Magnaporthe oryzae plays an important role in fungal germination and host penetration by providing recombinant turgor pressure for fungal appressors through lipid synthesis[36]. In addition, in the filamentous pathogenic fungus M. oryzae, hydrolysis of stored lipids to FA and glycerol facilitates conidial germination, which is critical for fungal toxicity [37]. In conclusion, the length and saturation of fatty acid chains can provide reserve energy for pathogenic fungi by responding to more diverse signaling pathways on the one hand [38,39,40]. On the other hand, unsaturated fatty acids provide turgor pressure for fungal growth, germ tube germination and spore maturation, so that the spore can contribute in the early stage of infection.
3. Plasma membrane curvature affects membrane homeostasis and the pathogenicity of fungi
Membrane curvature is a major player in the growth, division, and movement of cells; curvature and curvature-sensing proteins help vesicle budding and vesicle transport [41]. The shape of the cytoplasmic membrane is a conservatively controlled phenotype. The membrane shape/curvature has a significant effect on the biological function of the cell [42]. Certain intracellular proteins bend lipid bilayers to generate membrane bending required for transport [43]. cytoplasmic membranes are the flexible barrier that surrounds cells and their compartments. To perform locomotion or receptor conversion, cells need to control the shape of the plasma membrane. The proteins are inserted into the cytoplasmic membrane like wedges, altering the membrane curvature beyond the plasma membrane [41]. For example, the BAR-domain proteins help other proteins recruit to the location of the membrane that needs to be bent [44].
In fungi, membrane curvature is closely related to the cytoskeleton and lipid content. Septin plays an important role in fungi to allow shape changes and organize the f-actin cytoskeleton to promote aggressive growth [45,46,47]. The septins in Magnaporthe grisea support the F-actin protein via the ezrin-radixin-moesin protein Tea1 and interact with the phosphatidyl side on the appressorium plasma membrane, providing cortical rigidity and membrane curvature to form a pressure dome cell named appressorium. The fungi physically destroy the cuticle of the leaf to enter the plant tissue [48]. Studies on the lipidation of eukaryotic cells showed that lipids accumulation increases the curvature of the cytoplasmic membrane, which affects cell growth [49]. In filamentous pathogenic fungi, the curvature of the cytoplasmic membrane not only affects the cell transformation into mycelia but also affects the growth and spore formation. Overall, the curvature of the cytoplasmic membrane affects the fungi virulence (Figure 1).
4. Effect of membrane proteins on cytoplasmic membrane homeostasis of pathogenic fungi
Fungal transmembrane proteins, such as G protein-coupled receptors (GPCRs), glycosylphosphatidylinositol-anchored proteins (GPI-APs), GTPases, MCCS (Can1 membrane compartment as arginine/H (+) transporter), etc., form a novel membrane domain with the plasma membrane, which helps regulate the various physiological functions of the membrane [50,51]. These membrane proteins regulate membrane transport and interact with membrane lipids and cell wall and cytoskeleton components to maintain cytoplasmic membrane homeostasis.
Transmembrane proteins exist in almost all areas of life, and they have a variety of biological roles, including defense and attack, biological development, cell adhesion, and signaling [52]. The plasma membrane, as a selective osmotic barrier for living cells, is essential for cell survival and function [53]. The transmembrane protein family maintains the concentration of various substances inside and outside of the cytoplasmic membrane within the normal range, ensuring the normal operation of the membrane function. GPCRs are known to be involved in a wide range of membrane transport and are capable of initiating or modulating signal transduction responses at multiple membrane sites [54]. GPI-APs become attached to the outer plasma membrane through post-translational modification and play important roles in cell signaling, cell adhesion, cell wall metabolism, immune response, and cytoplasmic membrane homeostasis [55]. The FgDnf family of genes encoding invertase, which mediates transmembrane transport of substances and is involved in vesicle budding, charge, and intramembrane protein diffusion, is critical for vegetative growth, development, and pathogenicity of Fusarium graminearum [56].
Several fungal membrane proteins interact with chitin, lipids and actin, widely involved in substance transport, and play an important role in cell function and structure. Fungal membrane proteins can form dynamic and stable structures with the cytoskeleton to maintain cytoplasmic membrane homeostasis. In fungi, two main mechanisms, turgor and the actin cytoskeleton, regulate plasma membrane tension (PMTS) that helps the connection between the protoplasts and cell wall. These systems work with the plasma membrane to help adhesion to the cell wall. Depolymerization of F-actin leads to the rapid formation of endocytic pits while inhibiting their division from the plasma membrane [57]. Actin filaments can be organized into a variety of structures that are capable of exerting or resisting mechanical forces in the cellular environment [58]. In Aspergillus fumigatus, transmembrane GTPases protein combines with the septin (AspA-E), a kind of cytoskeleton, to regulate chitin deposition, separation, cytokinesis, and sporulation of fungal cell wall and contributes to host invasion and virulence of mycogenic fungi [59,60].
The cytoskeleton not only helps stabilize the cell structure through the cytoplasmic membrane but also regulates mycelium and spore formation to a certain extent. Phosphatidylinositol-(4,5)-bisphosphate [PI(4,5)P2] and septin in Candida albicans regulate the caspofungin response and the development of fungal mycelia in early fungal stage[61,62]. Septin also colocalizes with membrane sterol-rich regions and promotes the recruitment of cell wall synthases during cell wall remodeling[63]. In addition, a protein kinase associated with septin ring assembly, Gin4, is an important regulator of fungal plasma membrane asymmetry[64].
Fungal membrane proteins maintain cytoplasmic membrane homeostasis by forming dynamic and stable structures with cell walls. The cell walls of filamentous fungi are mainly composed of chitin and glucan. Fungal chitin synthetase is a large family of proteins in filamentous fungi that contain transmembrane domains and are transported to the plasma membrane by secretory vesicles to synthesize new cell walls [65]. In Trichoderma atroviride, LysM domain protein TAL6 binds to certain forms of polymerized chitin, inhibiting plant defense responses and protecting plants from chitinase [66]. Furthermore, Sur7, one of the many proteins that constitute MCC (the membrane compartment of Can1 acting as an arginine/H(+) transporter), is involved in membrane-related cellular events and regulates fungal spore formation, cell wall integrity, stress tolerance, and virulence [67]. We speculate that transmembrane proteins help in cell wall synthesis by transporting cell wall components. Therefore, fungal membrane proteins might be playing crucial roles in cell wall homeostasis.
5. Cell wall affect cytoplasmic membrane homeostasis of the pathogenic fungi
Cell growth, membrane tension, and cell wall synthesis are interdependent. The interplay between membrane tension and cell wall mechanical stress enables cells to rapidly rectify mismatches between the rates of the membrane and wall synthesis during cell growth, which ensures balanced and normal growth [68]. Cell walls isolate cells from their surroundings and protect them from environmental pressures. Cell wall integrity is maintained by highly regulated processes of cell wall biosynthesis [69]. The structural integrity between the cell wall and membrane is one of the important conditions for cell wall homeostasis. Studies in Candida albicans showed that cell growth is regulated by two complementary powerful inducible forces: membrane tension inhibits the expansion of the cell wall by interfering with the peptide glycan synthesis, while mechanical stress within the cell wall promotes the irreversible expansion of the cell wall. Only optimal membrane tension and cell wall stress ensure balanced cytoplasmic membrane and wall synthesis [68]. The mechanical cell wall stress sensor Wsc1 (MCW) participates in the regulation of cytoplasmic membrane and wall homeostasis, apical growth of fungal cells, and the protection of membrane domains in stressed environments [70].
Fungal cell wall components can provide mechanical protection for the integrity of fungal cytoplasmic membrane and cell wall. The fungal cell wall is an important barrier against pathogen attack [71]. Filamentous fungi regulate cell wall and membrane growth and pathogenicity through endocytosis and exemesis [72]. BbLec1, which binds to chitosan and chitin in fungal cell walls, plays a wide range of physiological roles in Beauveria bassiana including in defense responses and host-pathogen interactions [73]. MoVps13, an important virulence factor in Magnaporthe oryzae, plays an important role in cell wall integrity and plasma membrane homeostasis by participating in cell wall and sphingolipid synthesis [74]. Hex1 regulates to osmotic stress and influences the integrity of the cell wall and plasma membrane; it was shown to affect the colonization and pathogenicity of Verticillium dahliae fungi [75]. Chitosan can inhibit conidial germination by destroying the integrity of conidial membrane [76].
Fungal cell wall components also affect the tension of cytoplasmic membrane and mechanical tension of cell wall, providing suitable turgor pressure for fungal germination and even host infection. In Metarhizium acridum, the absence of MaPmt1 makes fungi more sensitive to cell wall disruptors, and the cell wall becomes thinner and the composition of the cell wall changes, thus reducing the appressorium dilatation pressure and unable to maintain the normal appressorium dilatation pressure through the host cuticle [77].
6. Discussion
Pathogenic fungi are widely present in nature and infect diverse host species including higher animals, plants, and lower animals, which indicates extensive interaction between the pathogenic fungi and the host [78]. cytoplasmic membrane homeostasis is closely related to the pathogenicity of both plant and animal pathogenic fungi. Fatty acid desaturase BbOle1 and BbSAY1 affect the lipid components of cytoplasmic membranes by regulating the content of unsaturated fatty acids through the lipid synthesis and catabolic pathways, respectively, ultimately regulating cytoplasmic membrane integrity and permeability in Beauveria bassiana [34,73]. Membrane integrity and permeability are important indicators of plasma membrane homeostasis. cytoplasmic membrane homeostasis is not only closely related to membrane lipid and cell wall homeostasis but also depends on cytoplasmic membrane proteins and cytoskeleton. These organelles or components together contribute to cytoplasmic membrane homeostasis, regulating fungal pathogenicity. For example, the lipid bilayer plays a central role in the physicochemical environment and shape of the membrane by influencing the arrangement and function of membrane proteins [79]. Lipids, especially membrane lipids, are important for the integrity of the cell plasma membrane. In addition, many fungal membrane lipid components, such as sphingolipids, ergosterol, and phospholipids, participate in signaling pathways, which regulate the integrity of fungal cell walls [80]. The cell wall and membrane barrier are closely related to lipids. The actin skeleton exerts mechanical forces on the cytoplasmic membrane and thereby regulates various cellular and subcellular morphogenetic processes such as cell migration, cytoplasmic division, and membrane transport [81]. This suggests that the cytoskeleton can independently determine the homeostasis of fungal cytoplasmic membranes. In all, the microdomains formed between membrane proteins, cytoskeleton, cell wall, and membrane lipid components are critical for plasma membrane homeostasis and the pathogenicity of fungal cells.
The cytoplasmic membrane homeostasis is directly related to the growth and development of fungi, which largely determines fungal virulence and pathogenicity. cytoplasmic membrane homeostasis is a unified complex system that also involves the homeostasis of other membrane-structured organelles such as endoplasmic reticulum [82,83], mitochondrial membrane [84], Golgi apparatus, vacuolar membrane, and vesicle membrane system [85]. However, here, we only focused on plasma membrane homeostasis and its effects on fungal pathogenicity. In terms of composition, membrane lipids and transmembrane proteins are indispensable factors determining the complexity and diversity of all cell organelles and membrane systems. The contribution of transmembrane proteins and lipid components to cytoplasmic membrane homeostasis has been widely reported, however, it is not clear how the micromembrane domains formed by transmembrane proteins and various lipid components contribute to cytoplasmic membrane homeostasis and pathogenicity of fungi (Figure2).
Besides, membrane curvature is another important factor affecting the homeostasis of the fungal cytoplasmic membrane [86]. Transmembrane proteins, lipid content, cytoskeleton, and protein-forming microdomains all determine the degree of membrane curvature, which then affect the dimorphic transformation of fungi and the formation of invasive organs (such as appressorium) that are important constituents of fungi virulence. However, most of the research work on these topics has been biophysical simulations, and only a small amount of research focused on the mechanism of host disease due to changes in the membrane curvature of fungi. Especially for filamentous pathogenic fungi, many important questions remain to be answered. For instance, how do transmembrane proteins, membrane lipids, cell walls, and cytoskeletal components regulate plasma membrane-cytoskeleton? What is plasma membrane-cell wall involvement in conidia or blastospore (yeast cell) conversion to filamentous cells? How do they mediate fungal infection of the host? Nonetheless, unsaturated non-polar fatty acids also play an important role in determining fungal membrane curvature. However, no related genes have been reported to date that may influence the curvature of the fungal plasma membrane. The effect of membrane curvature on the homeostasis and pathogenicity of fungal cytoplasmic membranes could be a promising research direction.
Acknowledgments
This work was financially supported by the Natural Science Foundation of Yunnan Province (202201AT070976) and Scientific Research Fund Project of Education Department of Yunnan Province (2023Y0902) grants.
References
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: where they are and how they behave. Nat Rev Mol Cell Bio 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, G.L. The Fluid-Mosaic Model of Membrane Structure: still relevant to understanding the structure, function and dynamics of biological membranes after more than 40 years. Biochim Biophys Acta 2014, 1838, 1451–1466. [Google Scholar] [CrossRef] [PubMed]
- Sonnino, S.; Prinetti, A. Membrane domains and the "lipid raft" concept. Curr Med Chem 2013, 20, 4–21. [Google Scholar] [PubMed]
- Horn, A.; Jaiswal, J.K. Structural and signaling role of lipids in plasma membrane repair. Curr Top Membr 2019, 84, 67–98. [Google Scholar] [CrossRef] [PubMed]
- Cerioni, L.; Volentini, S.I.; Prado, F.E.; etc. Cellular damage induced by a sequential oxidative treatment on Penicillium digitatum. J Appl Microbiol 2010, 109, 1441–1449. [CrossRef]
- Malinsky, J.; Opekarova, M. New Insight Into the Roles of Membrane Microdomains in Physiological Activities of Fungal Cells. Int Rev Cel Mol Bio 2016, 325, 119–180. [Google Scholar] [CrossRef]
- Ziolkowska, N.E.; Christiano, R.; Walther, T.C. Organized living: formation mechanisms and functions of plasma membrane domains in yeast. Trends Cell Biol 2012, 22, 151–158. [Google Scholar] [CrossRef]
- Harayama, T.; Riezman, H. Understanding the diversity of membrane lipid composition. Nat Rev Mol Cell Bio 2018, 19, 281–296. [Google Scholar] [CrossRef]
- Escriba, P.V.; Busquets, X.; Inokuchi, J.; etc. Membrane lipid therapy: Modulation of the cell membrane composition and structure as a molecular base for drug discovery and new disease treatment. Prog Lipid Res 2015, 59, 38–53. [CrossRef]
- Epand, R.M. Introduction to membrane lipids. Methods Mol Biol 2015, 1232, 1–6. [Google Scholar] [CrossRef]
- Thomas, F.B.; Omnus, D.J.; Bader, J.M.; etc. Tricalbin proteins regulate plasma membrane phospholipid homeostasis. Life Sci Alliance 2022, 5. [CrossRef]
- Akhberdi, O.; Zhang, Q.; Wang, H.; etc. Roles of phospholipid methyltransferases in pycnidia development, stress tolerance and secondary metabolism in the taxol-producing fungus Pestalotiopsis microspore. Microbiol Res 2018, 210, 33–42. [CrossRef] [PubMed]
- Beauvais, A.; Latge, J.P. Membrane and cell wall targets in Aspergillus fumigatus. Drug Resist Update 2001, 4, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.M.; de Castro, P.A.; Singh, A.; etc. Functional characterization of the Aspergillus nidulans glucosylceramide pathway reveals that LCB Delta8-desaturation and C9-methylation are relevant to filamentous growth, lipid raft localization and Psd1 defensin activity. Mol Microbiol 2016, 102, 488–505. [CrossRef]
- Vernay, A.; Schaub, S.; Guillas, I.; etc. A steep phosphoinositide bis-phosphate gradient forms during fungal filamentous growth. J Cell Biol 2012, 198, 711–730. [CrossRef] [PubMed]
- Li, S.; Du L; Yuen, G.; etc. Distinct ceramide synthases regulate polarized growth in the filamentous fungus Aspergillus nidulans. Mol Biol Cell 2006, 17, 1218–1227. [CrossRef]
- Head, B.P.; Patel, H.H.; Insel, P.A. Interaction of membrane/lipid rafts with the cytoskeleton: impact on signaling and function: membrane/lipid rafts, mediators of cytoskeletal arrangement and cell signaling. Biochim Biophys Acta 2014, 1838, 532–545. [Google Scholar] [CrossRef]
- Markham, P.; Robson, G.D.; Bainbridge, B.W.; etc. Choline: its role in the growth of filamentous fungi and the regulation of mycelial morphology. Fems Microbiol Rev 1993, 10, 287–300. [CrossRef]
- Ghugtyal, V.; Garcia-Rodas, R.; Seminara, A.; etc. Phosphatidylinositol-4-phosphate-dependent membrane traffic is critical for fungal filamentous growth. P Natl Acad Sci Usa 2015, 112, 8644–8649. [CrossRef]
- Cotado-Sampayo, M.; Ramos, P.O.; Perez, R.O.; etc. Specificity of commercial anti-spectrin antibody in the study of fungal and Oomycete spectrin: cross-reaction with proteins other than spectrin. Fungal Genet Biol 2008, 45, 1008–1015. [CrossRef]
- Zhang, L.; Cao, X.; Wang, Z.; etc. Brassinolide alleviated chilling injury of banana fruit by regulating unsaturated fatty acids and phenolic compounds. Sci Hortic-Amsterdam 2022, 297, 110922. [CrossRef]
- Resnick, M.A.; Mortimer, R.K. Unsaturated fatty acid mutants of Saccharomyces cerevisiae. J Bacteriol 1966, 92, 597–600. [Google Scholar] [CrossRef] [PubMed]
- Wisnieski, B.J.; Kiyomoto, R.K. Fatty acid desaturase mutants of yeast: growth requirements and electron spin resonance spin-label distribution. J Bacteriol 1972, 109, 186–195. [Google Scholar] [CrossRef]
- Reich, M.; Gobel, C.; Kohler, A.; etc. Fatty acid metabolism in the ectomycorrhizal fungus Laccaria bicolor. New Phytol 2009, 182, 950–964. [CrossRef] [PubMed]
- Micoogullari, Y.; Basu, S.S.; Ang, J.; etc. Dysregulation of very-long-chain fatty acid metabolism causes membrane saturation and induction of the unfolded protein response. Mol Biol Cell 2020, 31, 7–17. [CrossRef] [PubMed]
- Ballweg, S.; Ernst, R. Control of membrane fluidity: the OLE pathway in focus. Biol Chem 2017, 398, 215–228. [Google Scholar] [CrossRef]
- Zhang, S.; Skalsky, Y.; Garfinkel, D.J. MGA2 or SPT23 is required for transcription of the delta9 fatty acid desaturase gene, OLE1, and nuclear membrane integrity in Saccharomyces cerevisiae. Genetics 1999, 151, 473–483. [Google Scholar] [CrossRef]
- Ballweg, S.; Ernst, R. Control of membrane fluidity: the OLE pathway in focus. Biol Chem 2017, 398, 215–228. [Google Scholar] [CrossRef]
- Ogasawara, Y.; Kira, S.; Mukai, Y.; etc. Ole1, fatty acid desaturase, is required for Atg9 delivery and isolation membrane expansion during autophagy in Saccharomyces cerevisiae. Biol Open 2017, 6, 35–40. [CrossRef]
- Covino, R.; Ballweg, S.; Stordeur, C.; etc. A Eukaryotic Sensor for Membrane Lipid Saturation. Mol Cell 2016, 63, 49–59. [CrossRef]
- Luo, X.; Affeldt, K.J.; Keller, N.P. Characterization of the Far Transcription Factor Family in Aspergillus flavus. G3-Genes Genom Genet 2016, 6, 3269–3281. [Google Scholar] [CrossRef] [PubMed]
- Surma, M.A.; Klose, C.; Peng, D.; etc. A lipid E-MAP identifies Ubx2 as a critical regulator of lipid saturation and lipid bilayer stress. Mol Cell 2013, 51, 519–530. [CrossRef] [PubMed]
- Paege, N.; Warnecke, D.; Zauner, S.; etc. Species-Specific Differences in the Susceptibility of Fungi to the Antifungal Protein AFP Depend on C-3 Saturation of Glycosylceramides. Msphere 2019, 4. [CrossRef]
- Peng, Y.J.; Zhang, H.; Feng, M.G.; etc. SterylAcetyl Hydrolase 1 (BbSay1) Links Lipid Homeostasis to Conidiogenesis and Virulence in the Entomopathogenic Fungus Beauveria bassiana. J Fungi 2022, 8. [CrossRef]
- Turk, M.; Abramovic, Z.; Plemenitas, A.; etc. Salt stress and plasma-membrane fluidity in selected extremophilic yeasts and yeast-like fungi. Fems Yeast Res 2007, 7, 550–557. [CrossRef]
- Sangappillai, V.; Nadarajah, K. Fatty Acid Synthase Beta Dehydratase in the Lipid Biosynthesis Pathway Is Required for Conidiogenesis, Pigmentation and Appressorium Formation in Magnaporthe oryzae S6. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Soanes, D.M.; Kershaw, M.J.; etc. Functional analysis of lipid metabolism in Magnaporthe grisea reveals a requirement for peroxisomal fatty acid beta-oxidation during appressorium-mediated plant infection. Mol Plant Microbe in 2007, 20, 475–491. [CrossRef]
- Goodrich-Tanrikulu, M.; Howe, K.; Stafford, A.; etc. Changes in fatty acid composition of Neurospora crassa accompany sexual development and ascospore germination. Microbiol-Sgm, 1998; 144, Pt 7, 1713–1720. [CrossRef]
- Khunyoshyeng, S.; Cheevadhanarak, S.; Rachdawong, S.; etc. Differential expression of desaturases and changes in fatty acid composition during sporangiospore germination and development in Mucor rouxii. Fungal Genet Biol 2002, 37, 13–21. [CrossRef]
- Thines, E.; Weber, R.W.; Talbot, N.J. MAP kinase and protein kinase A-dependent mobilization of triacylglycerol and glycogen during appressorium turgor generation by Magnaporthe grisea. Plant Cell 2000, 12, 1703–1718. [Google Scholar] [CrossRef]
- McMahon, H.T.; Gallop, J.L. Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature 2005, 438, 590–596. [Google Scholar] [CrossRef]
- Kunding, A.H.; Mortensen, M.W.; Christensen, S.M.; etc. Intermembrane Docking Reactions Are Regulated by Membrane Curvature. Biophys J 2011, 101, 2693–2703. [CrossRef] [PubMed]
- Roux, A.; Koster, G.; Lenz, M.; etc. Membrane curvature controls dynamin polymerization. Proceedings of the National Academy of Sciences 2010, 107, 4141–4146. [Google Scholar] [CrossRef] [PubMed]
- Mim, C.; Unger, V.M. Membrane curvature and its generation by BAR proteins. Trends Biochem Sci 2012, 37, 526–533. [Google Scholar] [CrossRef]
- Momany, M.; Talbot, N.J. Septins Focus Cellular Growth for Host Infection by Pathogenic Fungi. Front Cell Dev Biol 2017, 5, 33. [Google Scholar] [CrossRef]
- Bridges, A.A.; Jentzsch, M.S.; Oakes, P.W.; etc. Micron-scale plasma membrane curvature is recognized by the septin cytoskeleton. J Cell Biol 2016, 213, 23–32. [CrossRef]
- Cannon, K.S.; Woods, B.L.; Crutchley, J.M.; etc. An amphipathic helix enables septins to sense micrometer-scale membrane curvature. J Cell Biol 2019, 218, 1128–1137. [CrossRef]
- Dagdas, Y.F.; Yoshino, K.; Dagdas, G.; etc. Septin-mediated plant cell invasion by the rice blast fungus, Magnaporthe oryzae. Science 2012, 336, 1590–1595. [CrossRef] [PubMed]
- Nath, S.; Dancourt, J.; Shteyn, V.; etc. Lipidation of the LC3/GABARAP family of autophagy proteins relies on a membrane-curvature-sensing domain in Atg3. Nat Cell Biol 2014, 16, 415–424. [CrossRef]
- Guna, A.; Hegde, R.S. Transmembrane Domain Recognition during Membrane Protein Biogenesis and Quality Control. Curr Biol 2018, 28, R498–R511. [Google Scholar] [CrossRef]
- Douglas, L.M.; Konopka, J.B. Fungal membrane organization: the eisosome concept. Annu Rev Microbiol 2014, 68, 377–393. [Google Scholar] [CrossRef]
- Ota, K.; Butala, M.; Viero, G.; etc. Fungal MACPF-like proteins and aegerolysins: bi-component pore-forming proteins? Subcell Biochem 2014, 80, 271–291. [CrossRef]
- Wang, F.; Wang, Y.; Zhang, X.; etc. Recent progress of cell-penetrating peptides as new carriers for intracellular cargo delivery. J Control Release 2014, 174, 126–136. [CrossRef]
- Eichel, K.; von Zastrow, M. Subcellular Organization of GPCR Signaling. Trends Pharmacol Sci 2018, 39, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, S.; Zurzolo, C.; Paladino, S. Organization of GPI-anchored proteins at the cell surface and its physiopathological relevance. Crit Rev Biochem Mol 2018, 53, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.; Guo, P.; Zhang, J.; etc. Flippases play specific but distinct roles in the development, pathogenicity, and secondary metabolism of Fusarium graminearum. Mol Plant Pathol 2020, 21, 1307–1321. [CrossRef] [PubMed]
- Mazheika, I.; Voronko, O.; Kamzolkina, O. Early endocytosis as a key to understanding mechanisms of plasma membrane tension regulation in filamentous fungi. Biol Cell 2020, 112, 409–426. [Google Scholar] [CrossRef]
- Blanchoin, L.; Boujemaa-Paterski, R.; Sykes, C.; etc. Actin dynamics, architecture, and mechanics in cell motility. Physiol Rev 2014, 94, 235–263. [CrossRef]
- Juvvadi, P.R.; Fortwendel, J.R.; Rogg, L.E.; etc. Differential localization patterns of septins during growth of the human fungal pathogen Aspergillus fumigatus reveal novel functions. Biochem Bioph Res Co 2011, 405, 238–243. [CrossRef]
- Vargas-Muniz, J.M.; Renshaw, H.; Richards, A.D.; etc. The Aspergillus fumigatus septins play pleiotropic roles in septation, conidiation, and cell wall stress, but are dispensable for virulence. Fungal Genet Biol 2015, 81, 41–51. [CrossRef]
- Badrane, H.; Nguyen, M.H.; Blankenship, J.R.; etc. Rapid redistribution of phosphatidylinositol-(4,5)-bisphosphate and septins during the Candida albicans response to caspofungin. Antimicrob Agents Ch 2012, 56, 4614–4624. [CrossRef]
- Badrane, H.; Nguyen, M.H.; Clancy, C.J. Highly Dynamic and Specific Phosphatidylinositol 4,5-Bisphosphate, Septin, and Cell Wall Integrity Pathway Responses Correlate with Caspofungin Activity against Candida albicans. Antimicrob Agents Ch 2016, 60, 3591–3600. [Google Scholar] [CrossRef] [PubMed]
- Mela, A.; Momany, M. Septins coordinate cell wall integrity and lipid metabolism in a sphingolipid-dependent process. J Cell Sci 2022, 135. [Google Scholar] [CrossRef] [PubMed]
- Roelants, F.M.; Su, B.M.; von Wulffen, J.; etc. Protein kinase Gin4 negatively regulates flippase function and controls plasma membrane asymmetry. J Cell Biol 2015, 208, 299–311. [CrossRef] [PubMed]
- Takeshita, N. Control of Actin and Calcium for Chitin Synthase Delivery to the Hyphal Tip of Aspergillus. Curr Top Microbiol 2020, 425, 113–129. [Google Scholar] [CrossRef]
- Seidl-Seiboth, V.; Zach, S.; Frischmann, A.; etc. Spore germination of Trichoderma atroviride is inhibited by its LysM protein TAL6. Febs J 2013, 280, 1226–1236. [CrossRef]
- Zhang, L.B.; Tang, L.; Guan, Y.; etc. Subcellular localization of Sur7 and its pleiotropic effect on cell wall integrity, multiple stress responses, and virulence of Beauveria bassiana. Appl Microbiol Biot 2020, 104, 6669–6678. [CrossRef]
- Rojas, E.R.; Huang, K.C.; Theriot, J.A. Homeostatic Cell Growth Is Accomplished Mechanically through Membrane Tension Inhibition of Cell-Wall Synthesis. Cell Syst 2017, 5, 578–590. [Google Scholar] [CrossRef]
- Radeck, J.; Lautenschlager, N.; Mascher, T. The Essential UPP Phosphatase Pair BcrC and UppP Connects Cell Wall Homeostasis during Growth and Sporulation with Cell Envelope Stress Response in Bacillus subtilis. Front Microbiol 2017, 8, 2403. [Google Scholar] [CrossRef]
- Athanasopoulos, A.; Andre, B.; Sophianopoulou, V.; etc. Fungal plasma membrane domains. Fems Microbiol Rev 2019, 43, 642–673. [CrossRef]
- Kubicek, C.P.; Starr, T.L.; Glass, N.L. Plant cell wall-degrading enzymes and their secretion in plant-pathogenic fungi. Annu Rev Phytopathol 2014, 52, 427–451. [Google Scholar] [CrossRef]
- Commer, B.; Shaw, B.D. Current views on endocytosis in filamentous fungi. Mycology 2020, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.J.; Ding, J.L.; Lin, H.Y.; etc. A virulence-related lectin traffics into eisosome and contributes to functionality of cytomembrane and cell-wall in the insect-pathogenic fungus Beauveria bassiana. Fungal Biol-Uk 2021, 125, 914–922. [CrossRef]
- Zhu, X.; Li, L.; Wang, J.; etc. acuolar Protein-Sorting Receptor MoVps13 Regulates Conidiation and Pathogenicity in Rice Blast Fungus Magnaporthe oryzae. J Fungi 2021, 7. [CrossRef]
- Vangalis, V.; Papaioannou, I.A.; Markakis, E.A.; etc. Hex1, the Major Component of Woronin Bodies, Is Required for Normal Development, Pathogenicity, and Stress Response in the Plant Pathogenic Fungus Verticillium dahliae. J Fungi 2020, 6. [CrossRef]
- Palma-Guerrero, J.; Huang, I.C.; Jansson, H.B.; etc. Chitosan permeabilizes the plasma membrane and kills cells of Neurospora crassa in an energy dependent manner. Fungal Genet Biol 2009, 46, 585–594. [CrossRef] [PubMed]
- Wen, Z.; Tian, H.; Xia, Y.; etc. MaPmt1, a protein O-mannosyltransferase, contributes to virulence through governing the appressorium turgor pressure in Metarhizium acridum. Fungal Genet Biol 2020, 145, 103480. [CrossRef]
- Klein, D.A.; Paschke, M.W. Filamentous fungi: the indeterminate lifestyle and microbial ecology. Microb Ecol 2004, 47, 224–235. [Google Scholar] [CrossRef]
- Phillips, R.; Ursell, T.; Wiggins, P.; etc. Emerging roles for lipids in shaping membrane-protein function. Nature 2009, 459, 379–385. [CrossRef]
- Fabri, J.; Rocha, M.C.; Malavazi, I. Overview of the Interplay Between Cell Wall Integrity Signaling Pathways and Membrane Lipid Biosynthesis in Fungi: Perspectives for Aspergillus fumigatus. Curr Protein Pept Sc 2020, 21, 265–283. [Google Scholar] [CrossRef]
- Pedersen, R.; Drubin, D.G. Type I myosins anchor actin assembly to the plasma membrane during clathrin-mediated endocytosis. J Cell Biol 2019, 218, 1138–1147. [Google Scholar] [CrossRef]
- Christianson, J.C.; Carvalho, P. Order through destruction: how ER-associated protein degradation contributes to organelle homeostasis. Embo J 2022, 41, e109845. [Google Scholar] [CrossRef] [PubMed]
- Christianson, J.C.; Ye, Y. Cleaning up in the endoplasmic reticulum: ubiquitin in charge. Nat Struct Mol Biol 2014, 21, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Kepp, O.; Trojel-Hansen, C.; etc. Mitochondrial control of cellular life, stress, and death. Circ Res 2012, 111, 1198–1207. [CrossRef] [PubMed]
- Nakatogawa, H. Mechanisms governing autophagosome biogenesis. Nat Rev Mol Cell Bio 2020, 21, 439–458. [Google Scholar] [CrossRef]
- Larsen, J.B.; Jensen, M.B.; Bhatia, V.K.; etc. Membrane curvature enables N-Ras lipid anchor sorting to liquid-ordered membrane phases. Nat Chem Biol 2015, 11, 192–194. [CrossRef]
Figure 1.
Formation of membrane curvature in pathogenic fungi. A. Early spore germination morphology of pathogenic fungi. B. The dividing or germinating germ tube in yeast cell of pathogenic fungus. C. Hypha of filamentous pathogenic fungi. The red box lines indicate where pathogenic fungi may form membrane curvature at different developmental stages. D. Several interaction forms of membrane curvature formation. a. Formation of membrane curvature of the plasma membrane caused by insertion of mosaic membrane proteins. b. Formation of membrane curvature of lipid membrane is caused by the combination of polar fatty acid chains and various lipid components. c. Membrane bending caused by interaction between membrane proteins and cytoskeletal proteins.
Figure 1.
Formation of membrane curvature in pathogenic fungi. A. Early spore germination morphology of pathogenic fungi. B. The dividing or germinating germ tube in yeast cell of pathogenic fungus. C. Hypha of filamentous pathogenic fungi. The red box lines indicate where pathogenic fungi may form membrane curvature at different developmental stages. D. Several interaction forms of membrane curvature formation. a. Formation of membrane curvature of the plasma membrane caused by insertion of mosaic membrane proteins. b. Formation of membrane curvature of lipid membrane is caused by the combination of polar fatty acid chains and various lipid components. c. Membrane bending caused by interaction between membrane proteins and cytoskeletal proteins.
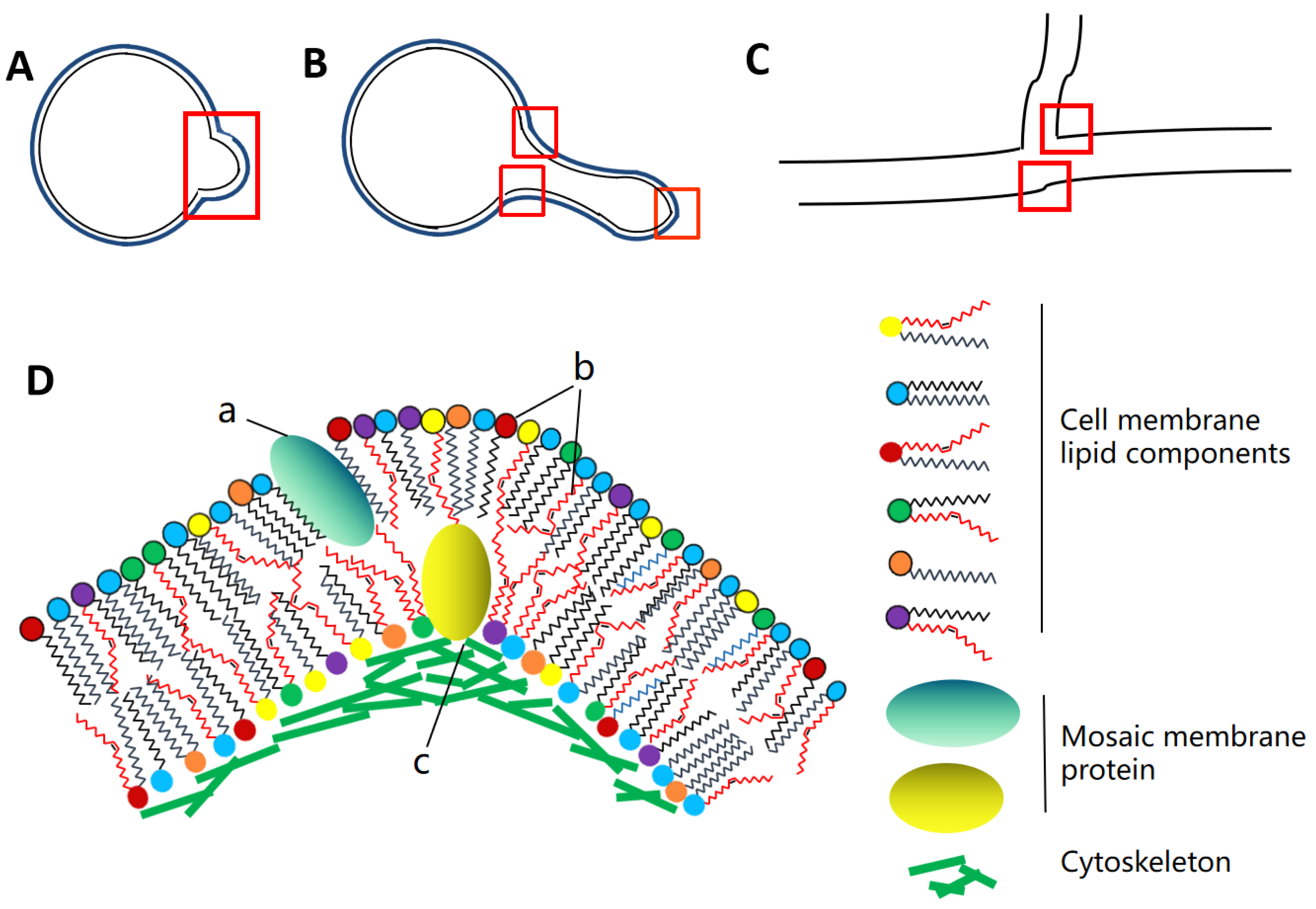
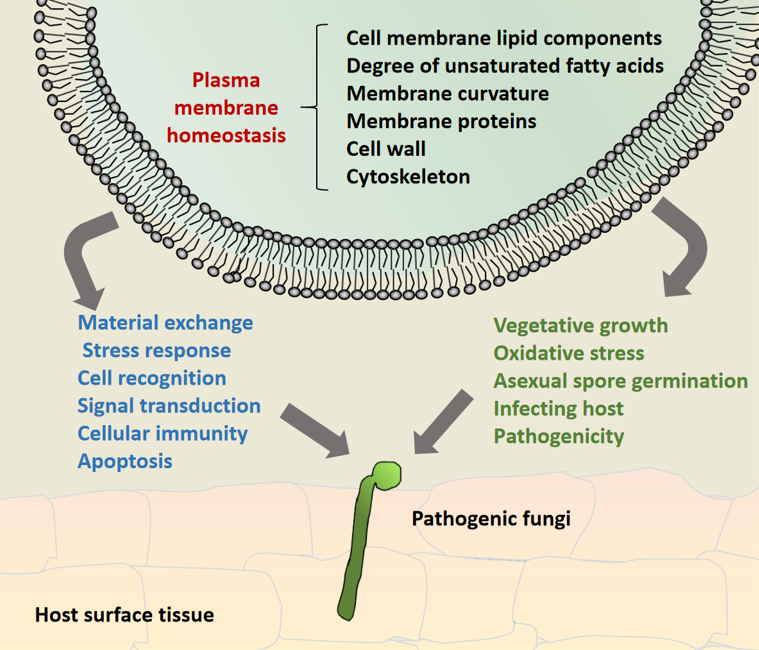
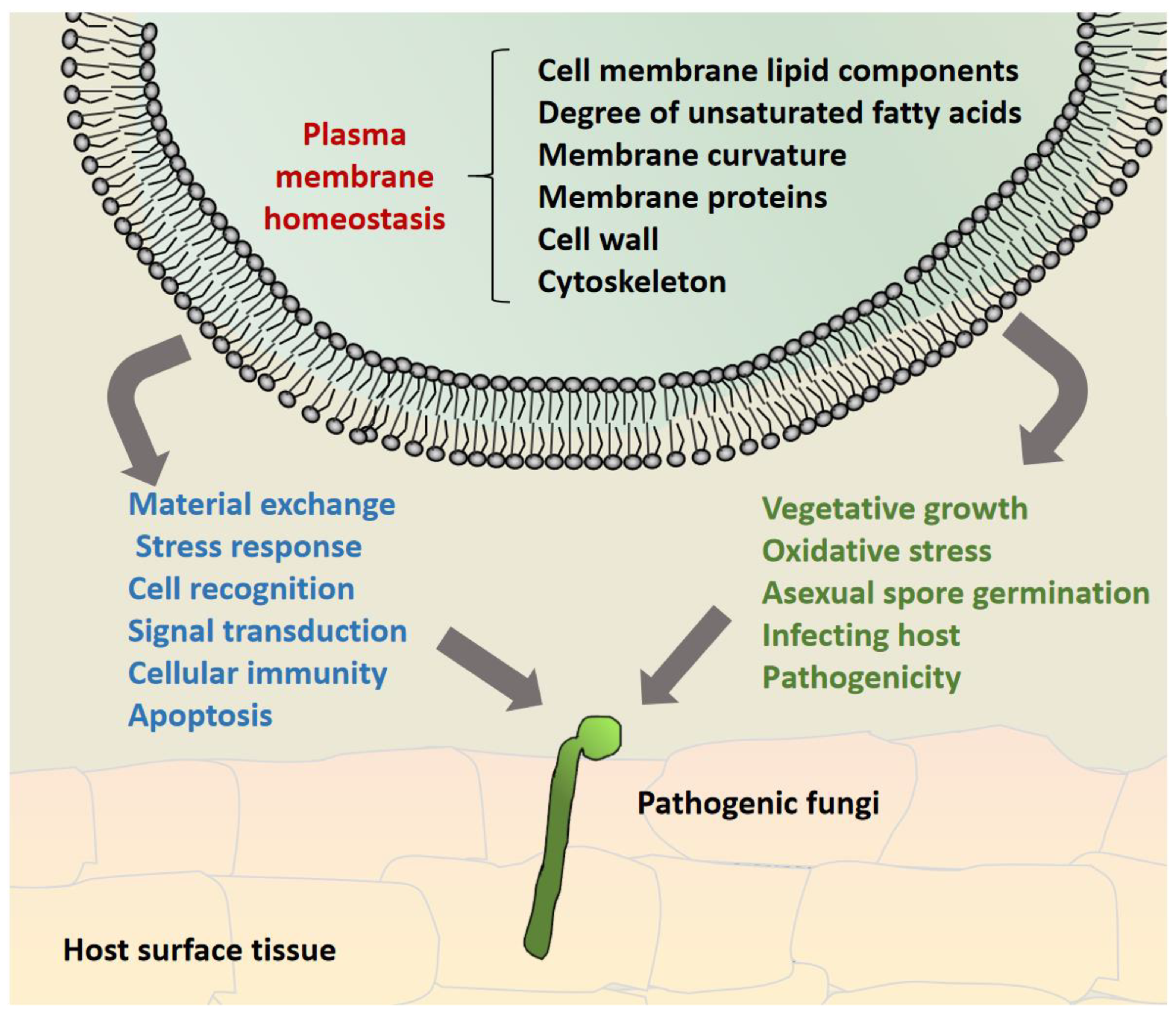
Figure 2.
Model diagram of pathogenic fungi participating in fungal growth and development and pathogenicity by maintaining cytoplasmic membrane homeostasis. It mainly relies on the interaction between membrane lipid components, transmembrane proteins, cytoskeleton and cell wall components, as well as the formation of membrane curvature, to maintain the integrity of plasma membrane assembly, structure and function of pathogenic fungi at various stages of cell development, so as to maintain cytoplasmic membrane homeostasis and contribute to fungal virulence.
Figure 2.
Model diagram of pathogenic fungi participating in fungal growth and development and pathogenicity by maintaining cytoplasmic membrane homeostasis. It mainly relies on the interaction between membrane lipid components, transmembrane proteins, cytoskeleton and cell wall components, as well as the formation of membrane curvature, to maintain the integrity of plasma membrane assembly, structure and function of pathogenic fungi at various stages of cell development, so as to maintain cytoplasmic membrane homeostasis and contribute to fungal virulence.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



