
Submitted:
24 May 2023
Posted:
25 May 2023
You are already at the latest version
Abstract
Caseinolytic peptidase P (CLPP) plays a central role in mitochondrial unfolded protein response (mtUPR) by promoting the breakdown of misfolded proteins and setting in motion a cascade of reactions to re-establish protein homeostasis. Global germline deletion of Clpp in mice results in female infertility and accelerated follicular depletion. Telomeres, on the other hand, are tandem repeats of 5’-TTAGGG-3’ sequences found at the ends of the chromosomes. Telomeres are essential for maintaining chromosome stability during somatic cell division and their shortening is associated with cellular senescence and aging. In this study, we asked whether the infertility and ovarian aging phenotype caused by global germline deletion of Clpp is associated with somatic aging, and tested telomere length in tissues of young and aging mice. We found that impaired mtUPR caused by the lack of CLPP is associated with accelerated telomere shortening in both oocytes and somatic cells of aging mice. In addition, expression of several genes that maintain telomere integrity were decreased, and double strand DNA breaks were increased in telomeric regions. Our results highlight how impaired mtUPR can affect telomere integrity and demonstrate a link between loss of mitochondrial protein hemostasis, infertility, and somatic aging.
Keywords:
Telomere length
; aging
; Clpp
; mitochondrial dysfunction
; unfolded protein response
Introduction
Human longevity has increased rapidly since the beginning of the 20th century due to medical advances, societal changes, and improved quality of life, which subsequently has led to delayed childbearing [1,2]. Consequently, the past two decades witnessed an increased interest in the investigation of the aging process. However, mechanisms regulating germ cell aging, and how they relate to somatic aging remain incompletely characterized.
Several mechanisms have been implicated in promoting cellular senescence, including telomere shortening [3] and mitochondrial dysfunction [4,5]. Telomeres consist of tandem repeats of 5′-TTAGGG-3′ sequences that are located at the end of the chromosomes and play a crucial role in preserving chromosome stability. During each cell division, 20-50 telomeric base pairs cannot be replicated and are lost at the 5’-end of the newly synthesized DNA strand, leading to telomere shortening [6]. Decreasing telomere length acts as a “mitotic clock” for cellular senescence and aging. This is because telomere shortening that occurs with each round of cell division ultimately leads to chromosome ends becoming exposed and activates a DNA damage response, which results in a permanent mitotic arrest known as replicative senescence [7]. Telomere damage may also occur in non-dividing cells, such as oocytes, and may result from oxidative DNA damage to guanine-rich telomeric repeats when exposed to reactive oxygen species (ROS), as well as epigenetic, environmental, dietary, and lifestyle variables [8,9].
Mitochondria on the other hand serve a crucial role in energy production, cellular metabolism, regulation of membrane potential, and apoptosis [10]. More recently, mitochondrial unfolded protein response (mtUPR), which ensures mitochondrial protein hemostasis by sensing mitochondrial (unfolded protein) stress, has been implicated in aging [11]. Activation of mtUPR contributes to enhanced longevity in experimental models [12-14], while a dysfunctional mtUPR may result in age-related accumulation of damaged proteins, decreased oxidative phosphorylation, and increased ROS (reviewed in [11]). Multiple mouse models of mitochondrial dysfunction also result in female infertility, further highlighting the the complex relationship between mitochondrial function, aging and reproductive potential [5,15,16].
Caseinolytic peptidase P (CLPP) is a key regulator of mtUPR [17-20]. CLPP cleaves misfolded mitochondrial proteins that are then exported to the cytoplasm where they activate transcription factors that induce mtUPR (reviewed in [11]). Previous research has shown that global germline Clpp loss in female mice impairs oocyte maturation and two-cell embryo formation and causes blastocyst development failure, which eventually results in infertility [5,21]. In addition, the absence of CLPP causes accelerated ovarian follicle depletion and results in a phenotype similar to diminished ovarian reserve and ovarian aging [5].
In this study, we aimed to investigate whether the infertility and ovarian follicular depletion phenotype observed in mice with global deletion of Clpp is associated with changes in somatic tissues that suggest accelerated aging. Our findings demonstrate that lack of Clpp causes shorter telomeres in oocytes and a number of somatic tissues of Clpp-/- mice. These changes are associated with decreased expression of genes that regulate telomere length and stability.
Materials and methods
Animals
Clpp+/- mice in a C57BL/6J background (Founder Line # IST13563G11) generated by Texas A&M Institute for Genomic Medicine (TIGM), an institute of AgriLife, and obtained from Georg Auburger, PhD (Goethe University Medical School, Frankfurt am Main, Germany) [21] were crossbred to obtain Clpp-/- mice. Mice care, breeding, and experimental procedures were conducted according to the Yale University Animal Research requirements, by using protocols approved by Institutional Animal Care and Use Committee (Protocol #2020-11207). Genotyping was carried out using the methods previously described [22].
Collection of oocytes and cumulus cells
To obtain immature (germinal vesicle-stage, GV) oocytes, 2-, 6-, and 9-month-old Clpp-/- and wild type (WT) female mice were injected intraperitoneally with 10IU pregnant mare’s serum gonadotropin (PMSG, Sigma, St. Louis, MO). After 44 hours of PMSG injection, ovaries were removed and punctured with a 26-gauge needle in M2 medium (Sigma, St. Louis, MO) supplemented by 10µM milrinone (Sigma, St. Louis, MO) to prevent meiotic resumption.
To collect mature oocytes arrested in the metaphase of the second meiotic divisio (MII), 10IU of human chorionic gonadotropin (hCG; Sigma, St. Louis, MO) was injected 48h hours after the PMSG (Sigma, St. Louis, MO). After 14-16 hours of hCG injection, unfertilized MII oocytes were collected from oviducts. Oocytes were stripped from cumulus cells with a mouth pipette and collected in individual tubes.
Blood and tissue collection
Blood samples (0.5 ml) were collected from each mice (n=5) through intracardiac puncture with a 18-gauge needle. After blood collection, mice were perfused with normal saline through a needle in the left ventricle. After perfusion, tissues (heart, liver, spleen, lung, kidney, uterus, and ovaries) were dissected and washed in Dulbecco’s Phosphate Buffer Saline (DPBS) for 10 seconds. Tissues were stored at -80°C until further experiments.
Telomere length measurement
GV and MII stage oocytes (collected from 10 mice for each genotype at each timepoint, and pooled as 2 mice per sample) were collected from 2-, 6-, and 9-month-old Clpp-/- mice and compared to WT. DNA extraction from oocytes and cumulus cells was performed using the QIAmp DNA Micro Kit (Qiagen, Valencia, CA), and DNA extraction from white blood cells (WBC) was conducted with DNA Isolation Kit for Mammalian Blood (Roche, Basel, Switzerland) according to manufacturer’s protocol and both concentrations were quantified using Qubit 3.0 (Life Technologies).
Average telomere length was measured from total genomic mouse DNA using a real-time quantitative PCR method previously described [16]. The average telomere length ratio was obtained by quantifying telomeric DNA with specially designed primer sequences and dividing that amount by the quantity of a single-copy gene, acidic ribosomal phosphoprotein PO (36B4) gene. Forward and reverse telomere and 36B4 primers are shown in Supplementary Table 1. Each reaction included 10 μl iQ™ SYBR® Green Supermix, (Bio-Rad), 400 nM of each primer, 1 ng genomic DNA, and enough double-distilled H2O to complete the volume in 20-μl reaction. PCR reactions were performed on the iCycler iQ real-time PCR detection system (Bio-Rad, Hercules, CA). For each PCR reaction, a standard curve was generated by serial dilutions of known amounts of DNA from the same tissues. The telomere signal was normalized to the signal from the single-copy gene to generate a T/S ratio indicative of relative telomere length. The relative input amount of the telomere PCR then was divided by the relative input amount of the 36B4 PCR of the same sample. Each real-time PCR experiment was repeated a minimum of three times.
Quantitative reverse-transcription polymerase chain reaction (qRT-PCR)
Total RNA was obtained from 20 oocytes per mouse using RNAqueous Microkit (Ambion, Austin, TX) and was treated for genomic DNA contamination using DNase I (Ambion). Reverse transcription was performed using the RETROscript kit (Ambion) in two steps: first, template RNA and oligo(dT) primers were incubated at 850C for 3 min to eliminate any secondary structures, and then the buffer and enzyme were added and the reaction was carried out at 420C for 1 h. qPCR was carried out on an iCycler (Bio-Rad Laboratories). Each 10-µl PCR reaction contained 5µl of iQ™ SYBR® Green Supermix (Bio-Rad Laboratories), 3µl of H2O, 0.5µl of each primer, and 1µl of cDNA. The 2-∆∆CT (cycle threshold) method was used to calculate relative expression levels after normalization to β-actin or Gapdh levels. Samples were assayed in triplicate and each experiment was repeated at least three times using individual animals from each genotype. The primers used for real-time PCR reactions are reported in Supplementary Table 1.
Immunofluorescent staining
For immunofluorescent staining, cumulus oophorus complexes (COCs) containing GV stage oocytes were collected and fixed with 4% paraformaldehyde (Sigma, St. Louis, MO) in Dulbecco’s Phosphate Buffer Saline (DPBS) for 5 min, then washed three times in 1X DPBS. They were placed in 0.5% Triton X-100 in DPBS at room temperature for 10 minutes, then washed in DPBS for 5 minutes. After blocking in 3% BSA (Sigma, St. Louis, MO) at room temperature for 1h, COCs were incubated overnight at 4°C with rat anti-TRF1 monoclonal antibody (Abcam, Cambridge, UK Cat# ab192629, 1:100) or mouse anti-H2A.X monoclonal antibody (Sigma-Aldrich, St. Louis, MO Cat# 05-636-25UG 1:100) as a primary antibody. After three washes with 1X DPBS for 10 minutes, COCs were incubated with Alexa Fluor 488-conjugated goat anti-mouse antibody (1:400) or Alexa Fluor 568-conjugated goat anti-rat antibody (1:400) for 1h at room temperature. Finally, they were mounted with 4′, 6-diamidino-2-phenylindole (DAPI;1:1000) (Life Technologies, Carlsbad, CA) on glass slides and stored at 4°C until imaging. Images were captured on Leica SP8 spectral scanning confocal microscope and Image J software was used to quantify the fluorescence intensity.
Statistical analysis
Quantitative data are expressed as mean ± SEM. The student’s t-test was used to analyze the statistical significance between the two groups and ANOVA was used for multiple groups. Data are representative of at least three independent experiments unless otherwise specified. All statistical analyses were done using Graph Pad Prism software version 9 and significance was assessed at p < 0.05.
Results
Telomere length in Clpp-/- mice oocytes, ovaries and somatic cells and tissues
In young (2-month-old) Clpp-/- mice, telomere length in GV and MII oocytes was similar to WT (1.16 ± 0.14 vs 0.88 ± 0.42, p = 0.08 and 1.08 ± 0.05 vs 1.06 ± 0.26, p = 0.52, respectively) (Figure 1A,B). Telomere length of cumulus cells (CCs) and other somatic cells in 2-month-old Clpp-/- mice were also found to be similar compared to WT (Figure 1B,C).
To assess the impact of aging, samples from 6-, and 9-month-old Clpp-/- and WT mice were analyzed. Telomere length of GV and MII oocytes from 6-month-old Clpp-/- mice were significantly shorter compared to WT (0.98 ± 0.01 vs 0.80 ± 0.02, p < 0.0001 and 0.99 ± 0.03 vs 0.69 ± 0.05, p = 0.001, respectively) (Figure 2A,B). Telomere length was also shorter in the uterus, ovary, and liver of 6-month-old Clpp-/- mice compared to WT (0.99 ± 0.44 vs 0.89 ± 0.31, p = 0.04; 0.99 ± 0.03 vs 0.87 ± 0.01, p = 0.002 and 1.02 ± 0.007 vs 0.86 ± 0.05, p = 0.04, respectively) (Figure 2C). In CCs, white blood cells (WBCs), lung, heart and spleen, mean telomere length was shorter in Clpp-/-, however the difference was not statistically signficant compared to WT mice (Figure 2B,C).
In 9-month-old mice, telomere length in GV and MII oocytes was also significantly shorter in Clpp-/- mice compared to WT (1.0 ± 0.09 vs 0.84 ± 0.04, p = 0.01 and 0.99 ± 0.04 vs 0.89 ± 0.03, p = 0.01, respectively) (Figure 3A,B). In somatic tissues, telomere length was signficantly shorter in the uterus (1.14 ± 0.03 vs 1.01 ± 0.03, p = 0.01), ovary (1.06 ± 0.04 vs 0.79 ± 0.01, p = 0.004), liver (1.03 ± 0.03 vs 0.90 ± 0.07, p = 0.01) and spleen (0.98 ± 0.01 vs 0.88 ± 0.03, p = 0.01). Conversely, while the mean telomere length was aslo lower in CCs, WBCs, lung, and heart in Clpp-/- mice compared to WT, the difference was not statistically signficant (Figure 3B,C).
Expression of telomere-associated genes in Clpp -/- oocytes
TRF1, TRF2 and POT1a are shelterin protein complex molecules involved in maintaining telomere DNA integrity and stability. Their deficiency causes DNA damage response and the accumulation of DNA repair factors [23,24]. H2AX, is a minor nucleosomal histone protein [25,26]. H2AX becomes phosphorylated at sites of double stranded DNA breaks and antibodies against the serine-139 phosphorylated form of H2AX allows microscopic detection of individual double stranded breaks in the DNA [27].
We first used qRT-PCR to quantify Trf1, Trf2, Pot1a, and H2Ax expression in our model. In 2-month-old mice, there was no significant difference between Clpp-/- and WT mice for Trf1, Trf2, Pot1a, or H2Ax expression (Figure 4A). Conversely, in the 6-month-old mice there was a significant decrease in Clpp-/- compared to WT mice for Trf1 (0.58 ± 0.07 vs 1.12 ± 0.03, p = 0.03), Trf2 (0.59 ± 0.08 vs 1.11 ± 0.02, p = 0.008), Pot1a (0.61 ± 0.29 vs 1.13 ± 0.04, p = 0.05), and H2Ax (0.67 ± 0.15 vs 1.11 ± 0.02, p = 0.03) (Figure 4B).
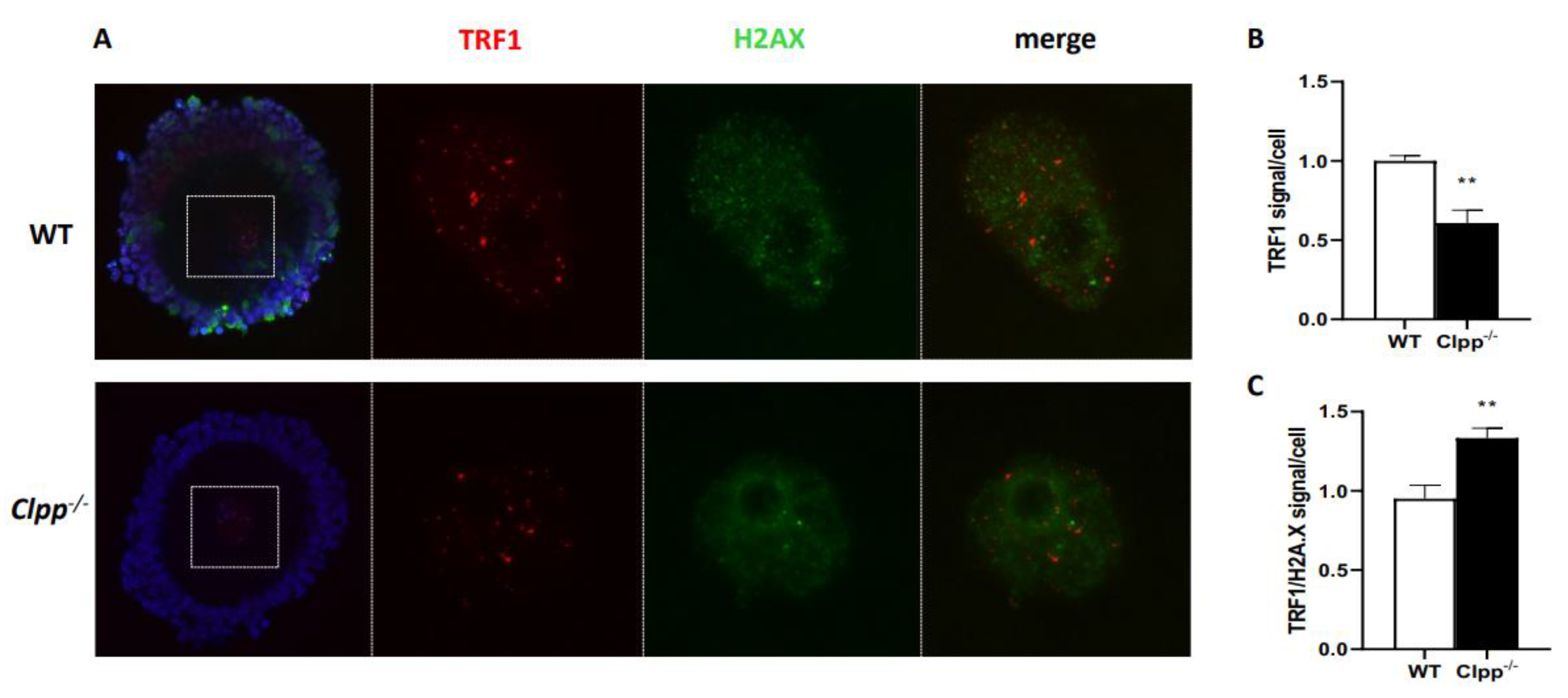
To further investigate the accelerated telomere damage in GV oocytes, we performed co-immunofluorescence staining of TRF1 and H2AX in 2- and 6-month-old Clpp-/- and WT mice. In the nuclei of 2-month-old Clpp-/- GV oocytes, there was no significant difference in TRF1 fluorescence compared to WT (1.0 ± 0.19 vs 1.19 ± 0.12, p = 0.23) (Figure 5A,B). In addition, TRF1 did not co-localize with H2AX in Clpp-/- oocytes, indicating absence of telomere damage (1.0 ± 0.21 vs 1.33 ± 0.26, p = 0.16) (Figure 5C). In 6-month-old mice, we found the fluorescence of TRF1 was significantly decreased in oocytes’ nuclei compared to WT (1.0 ± 0.08 vs 0.60 ± 0.18, p = 0.005) (Figure 6A,B). Additionally, there was an increased co-localization of TRF1 with H2AX (0.98 ± 0.21 vs 1.33 ± 0.13, p = 0.006) indicating telomeric damage in the Clpp-/- oocytes (Figure 6C).
Discussion
CLPP is essential for mtUPR as it promotes the breakdown of misfolded proteins within the mitochondria and initiates a series of reactions to eliminate the detrimental impact of mitochondrial stressors and to re-establish protein homeostasis [11]. Studies using knockout mouse models demonstrated that CLPP is required for female fertility [5,21]. In addition, global germline deletion of Clpp results in the activation of the mTOR (mammalian target of rapamycin) pathway and causes accelerated loss of ovarian follicular reserve, highlighting the role of CLPP in female reproductive competence and senescence [5]. In this study, we asked whether the infertility and ovarian aging phenotype caused by global germline deletion of Clpp is associated with somatic aging, and tested telomere length in young and aging mice gametes, gonads and somatic tissues. We found shortening of telomeres in both oocytes and somatic tissues at 6 and 9 months. In addition, expression of several genes associated with telomere integrity were decreased, and double strand DNA breaks were increased in telomeric regions. Our findings demonstrate how loss of mitochondrial protein homeostasis may accelerate telomere shortening in oocytes and somatic cells, and provide a link between reproductive and somatic aging.
A number of plausible mechanisms could be linking mitochondrial dysfunction and telomere shortening. Oxidative stress causes cell death and/or senescence, and as a result, rapid cell divions occur in the surrounding essential cells as a protective strategy, which leads to telomere shortening [28,29]. Reciprocally, in tissues with short telomeres, mitochondrial quantity and oxidative phosphorylation capacity are impaired, resulting in decreased ATP synthesis, decreased metabolic capacity, impaired gluconeogenesis, and elevated ROS levels [30,31]. We have previously reported that global knockout of Clpp resulted in increased ROS levels in GV oocytes [5], strengthening the association between increased ROS and telomere shortening in this model.
In 6- and 9-month-old Clpp-deficient mice, we found shorter telomere length in the liver when compared with WT, suggesting that the impaired mitrochondrial function within the liver in Clpp-/- mice could be triggering hepatic cellular changes that culminate in telomere shortening. Highly proliferative tissues, including the hematopoietic and immune systems exhibit impaired proliferative capacity in later generations of the telomerase RNA component (TERC)-deficient mice [32,33]. Telomerase complex gene mutations have been linked to rare human diseases such as Dyskeratosis Congenita (DKC) and Idiopathic Pulmonary Fibrosis [34], both of which are characterized by accelerated telomere shortening and organ failure. Patients with such diseases had an increased frequency of liver pathologies such as fibrosis and cirrhosis [35,36]. Similarly, two studies investigated the frequency of telomerase mutations in patients with sporadic cirrhosis compared to healthy controls and demonstrated mutation missense mutations in the Telomerase Reverse Transcriptase (TERT) and TERC genes in diseased patients [37].
Global deletion of Clpp resulted in mitochondrial dysfunction in oocytes by decrasing ATP production, expression levels of Electron Transport Chain (ETC) enzymes and increased levels of ROS [5]. Besides, the lack of Clpp affected the reproductive phenotype. Mice demonstrated a decrease in the number of mature oocytes and 2-cell embryos, and no blastocysts, resulting in infertility. In addition, they had accelerated follicular depletion and a phenotype consistent with dimished ovarian reserve [5]. In the current study, oocyte telomeres are shortened in at 6- and 9-month-old mice models with global deletion of Clpp. Consistent with our findings, when guanine-rich telomeric repeats are exposed to oxidative stress including ROS, they undergo oxidative DNA damage that shortens telomeres in non-dividing cells like oocytes, thereby potentially accelerating aging [38].
Global deletion of Clpp is also associated with spindle abnormalities and decreased ability to complete in vitro maturation of GV stage oocytes [5]. In the current study, shorter telomeres in 6- and 9-month-old Clpp-/- mice oocytes suggest that the reproductive phenotype of Clpp-/- mice may be, in part, due to oocyte telomere shortening. Indeed, telomeres play an important role in the regulation of chromosomal motions during meiosis, including bouquet creation at the leptotene stage, homologous pairing, and interaction with microtubules and spindles [39,40]. Short telomeres are linked to infertility, aberrant spindles, and misaligned metaphase chromosomes in oocytes of telomerase-deficient mice [41]. Similarly, telomere length in human unfertilized oocytes was associated with the morphological quality of the embryos generated from sibling oocytes from the same cohort, and subsequent pregnancy outcomes [42,43].
The impact of telomere length on oocyte function has been explored in animal and human studies [42-44]. Meanwhile, the role telomere length in cumulus/granulosa cell function remains to be further characterized, and exisiting data does not suggest this as a key regulatory mechanism. In the context of mtUPR, while global Clpp knockout was associated with infertility [5], granulosa cell-specific targeted deletion of Clpp did not affect reproduction [45]. In keeping with these findings, we did not find cumulus/granulosa cell telomere length to be shortened in Clpp-/- mice. This is not surprising as recent studies investigating the telomere length and epigenetic aging markers in human cumulus/granulosa cells did not see a difference when comparing younger versus older reproductive age women or those with good versus poor ovarian response [46-48].
A number of telomere proteins, including TRF1, TRF2, and POT1a, regulate telomere stability and length [49]. Zhang et al. revealed that mice with targeted deletion of mitochondrial fusion protein Mitofusin 2 (MFN2) have defective oocyte maturation, subfertility, shortened telomeres, and decreased TRF1 expression in oocytes [16]. They also showed co-localization of TRF1 with DNA repair factor 53BP1, suggesting DNA damage. Our findings are similar in that Trf1, Trf2, and Pot1a expression is decreased in 6-month-old Clpp-/- mice oocytes, and TRF1 co-localizes with DBA repair factor H2AX. Collectively these two studies show a consistent effect of mitochondrial dysfunction impacting dysregulated telomere mechanisms, thereby leading to DNA damage and telomere shortening.
In this study, we expanded our understanding of the role of Clpp and mtUPR in female reproduction and aging by characterizing telomere shortening and damage in a mouse model with global deletion of Clpp. As reported previously, the lack of Clpp results in functional mitochondrial abnormalities, infertility, and ovarian follicular depletion/aging. The current study demonstrates how this mitochondrial pathway, when impaired, may also promote somatic aging. In addition, our findings provide a preliminary understanding of how mitochondrial and telomeric aging mechansims may interact to accelerate reproductive and somatic aging. Further studies are needed to delineate the intricate interactions between these two aging pathways and to determine whether these could be exploited to delay or reverse ovarian (or somatic) aging.
Funding
This study was supported by a grant from Foundation for Embryonic Competence to ES.
Conflicts of Interest
M.C., Y.E, E.R., A.G. and G.I. declare no conflict of interests concerning this research. E.S. is a consultant for and receives research funding from the Foundation for Embryonic Competence.
References
- Esencan, E.; Simsek, B.; Seli, E. Analysis of female demographics in the United States: life expectancy, education, employment, family building decisions, and fertility service utilization. Curr Opin Obstet Gynecol 2021, 33, 170–177. [Google Scholar] [CrossRef]
- Sweeney, M.M.; Raley, R.K. Race, ethnicity, and the changing context of childbearing in the United States. Annu Rev Sociol 2014, 40, 539–558. [Google Scholar] [CrossRef]
- Bernadotte, A.; V. M., M.; Spivak, I.M. Markers of cellular senescence. Telomere shortening as a marker of cellular senescence. Aging 2016, 8, 3–11. [Google Scholar] [CrossRef]
- Kasapoğlu, I.; Seli, E. Mitochondrial dysfunction and ovarian aging. Endocrinology 2020, 161. [Google Scholar] [CrossRef]
- Wang, T.; Babayev, E.; Jiang, Z.; Li, G.; Zhang, M.; Esencan, E.; Horvath, T.; Seli, E. Mitochondrial unfolded protein response gene Clpp is required to maintain ovarian follicular reserve during aging, for oocyte competence, and development of pre-implantation embryos. Aging Cell 2018, 30, e12784. [Google Scholar] [CrossRef]
- Barnes, R.P.; Fouquerel, E.; Opresko, P.L. The impact of oxidative DNA damage and stress on telomere homeostasis. Mech Ageing Dev 2019, 177, 37–45. [Google Scholar] [CrossRef]
- Victorelli, S.; Passos, J.F. Telomeres and Cell Senescence - Size Matters Not. EBioMedicine 2017, 21, 14–20. [Google Scholar] [CrossRef]
- Derevyanko, A.; Skowronska, A.; Skowronski, M.T.; Kordowitzki, P. The Interplay between telomeres, mitochondria, and chronic stress exposure in the aging egg. Cells 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- van der Reest, J.; Nardini Cecchino, G.; Haigis, M.C.; Kordowitzki, P. Mitochondria: Their relevance during oocyte ageing. Ageing Res Rev 2021, 70, 101378. [Google Scholar] [CrossRef] [PubMed]
- Babayev, E.; Seli, E. Oocyte mitochondria function and reproduction. Curr Opin Obstet Gynecol 2015, 27, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Seli, E.; Wang, T.; Horvath, T.L. Mitochondrial unfolded protein response: a stress response with implications for fertility and reproductive aging. Fertil Steril 2019, 111, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Houtkooper, R.H.; Mouchiroud, L.; Ryu, D.; Moullan, N.; Katsyuba, E.; Knott, G.; Williams, R.W.; Auwerx, J. Mitonuclear protein imbalance as a conserved longevity mechanism. Nature 2013, 497, 451–457. [Google Scholar] [CrossRef]
- Durieux, J.; Wolff, S.; Dillin, A. The cell-non-autonomous nature of electron transport chain-mediated longevity. Cell.
- Moehle, E.A.; Shen, K.; Dillin, A. Mitochondrial proteostasis in the context of cellular and organismal health and aging. J Biol Chem 2019, 294, 5396–5407. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Bener, M.B.; Jiang, Z.; Wang, T.; Esencan, E. ; Scott R III; Horvath, T. ; Seli, E. Mitofusin 1 is required for female fertility and to maintain ovarian follicular reserve Cell Death Dis 2019, 10, 560. [Google Scholar] [PubMed]
- Zhang, M.; Bener, M.B.; Jiang, Z.; Wang, T.; Esencan, E.; Scott, R., III; Horvath, T.; Seli, E. Mitofusin 2 plays a role in oocyte and follicle development, and is required to maintain ovarian follicular reserve during reproductive aging. Aging 2019, 11, 3919–3938. [Google Scholar] [CrossRef]
- Zhao, Q.; Wang, J.; Levichkin, I.V.; Stasinopoulos, S.; Ryan, M.T.; Hoogenraad, N.J. A mitochondrial specific stress response in mammalian cells. Embo J 2002, 21, 4411–4419. [Google Scholar] [CrossRef] [PubMed]
- Haynes, C.M.; Petrova, K.; Benedetti, C.; Yang, Y.; Ron, D. ClpP mediates activation of a mitochondrial unfolded protein response in C. elegans. Dev Cell 2007, 13, 467–480. [Google Scholar] [CrossRef]
- Al-Furoukh, N.; Ianni, A.; Nolte, H.; Hölper, S.; Krüger, M.; Wanrooij, S.; al. , e. ClpX stimulates the mitochondrial unfolded protein response (UPRmt) in mammalian cells. Biochim Biophys Acta 2015, 1853, 2580–2591. [Google Scholar] [CrossRef]
- Nargund, A.M.; Fiorese, C.J.; Pellegrino, M.W.; Deng, P.; Haynes, C.M. Mitochondrial and nuclear accumulation of the transcription factor ATFS-1 promotes OXPHOS recovery during the UPR(mt). Mol Cell 2015, 58, 123–133. [Google Scholar] [CrossRef]
- Gispert, S.; Parganlija, D.; Klinkenberg, M.; Dröse, S.; Wittig, I.; Mittelbronn, M.; Grzmil, P.; Koob, S.; Hamann, A.; Walter, M. , et al. Loss of mitochondrial peptidase Clpp leads to infertility, hearing loss plus growth retardation via accumulation of CLPX, mtDNA and inflammatory factors. Hum Mol Genet 2013, 22, 4871–4887. [Google Scholar] [CrossRef]
- Seli, E.; Lalioti, M.D.; Flaherty, S.M.; Sakkas, D.; Terzi, N.; Steitz, J.A. An embryonic poly(A)-binding protein (ePAB) is expressed in mouse oocytes and early preimplantation embryos. Proc Natl Acad Sci USA 2005, 102, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Martínez, P.; Flores, J.M.; Blasco, M.A. 53BP1 deficiency combined with telomere dysfunction activates ATR-dependent DNA damage response. J Cell Biol 2012, 197, 283–300. [Google Scholar] [CrossRef] [PubMed]
- Smogorzewska, A.; Steensel, B.; Bianchi, A.; Oelmann, S.; Schaefer, M.R.; Schnapp, G.; Lange, T. Control of Human Telomere Length by TRF1 and TRF2. Mol Cell Biol 2000, 20, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Bonner, W.M.; West, M.H.; Stedman, J.D. Two-dimensional gel analysis of histones in acid extracts of nuclei, cells, and tissues. Eur J Biochem 1980, 109, 17–23. [Google Scholar] [CrossRef]
- Celeste, A.; Petersen, S.; Romanienko, P.J.; Fernandez-Capetillo, O.; Tang Chen, H.; Sedelnikova, O.A.; Reina-San-Martin, B.; Coppola, V.; Meffre, E.; Difilippantonio, M.J. , et al. Genomic instability in mice lacking histone H2AX. Science 2002, 296, 922–927. [Google Scholar] [CrossRef]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J Biol Chem 1998, 273, 5858–5868. [Google Scholar] [CrossRef]
- von Zglinicki, T. Oxidative stress shortens telomeres. Trends Biochem Sci 2002, 27, 339–344. [Google Scholar] [CrossRef]
- Kordowitzki, P.; Hamdi, M.; Derevyanko, A.; Rizos, D.; Blasco, M. The effect of rapamycin on bovine oocyte maturation success and metaphase telomere length maintenance. Aging 2020, 12, 7576–7584. [Google Scholar] [CrossRef]
- Chang, A.C.Y., Ong. Telomere shortening and metabolic compromise underlie dystrophic cardiomyopathy. Proc Natl Acad Sci U S A 2016, 113, 13120–13125. [Google Scholar] [CrossRef]
- Amano, H.; Chaudhury, A.; Rodriguez-Aguayo, C.; Lu, L.; Akhanov, V.; Catic, A.; Popov, Y.V.; Verdin, E.; Johnson, H.; Stossi, F. , et al. Telomere dysfunction induces sirtuin repression that drives telomere-dependent disease. Cell Metab 2019, 29, 1274–1290. [Google Scholar] [CrossRef]
- Butts, S.; Riethman, H.; Ratcliffe, S.; Shaunik, A.; Coutifaris, C.; Barnhart, K. Correlation of telomere length and telomerase activity with occult ovarian insufficiency. J Clinl Endocrinol Metab 2009, 94, 4835–4843. [Google Scholar] [CrossRef] [PubMed]
- Hande, M.; Samper, E.; Lansdorp, P.; Blasco, M.T. Telomere length dynamics and chromosomal instability in cells derived from telomerase null mice. J Cell Biol 1999, 144, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.K.; Wright, W.E.; Shay, J.W. Human diseases of telomerase dysfunction: insights into tissue aging. . Nucleic Acids Res 2007, 35, 7406–7416. [Google Scholar] [CrossRef]
- Calado, R.T.; Young, N.S. Telomere diseases. N Engl J Med 2009, 361, 2353–2365. [Google Scholar] [CrossRef] [PubMed]
- Carulli, L. Telomere shortening as genetic risk factor of liver cirrhosis. World J Gastroenterol 2015, 21, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Calado, R.T.; Brudno, J.; Mehta, P.; Kovacs, J.J.; Wu, C.; Zago, M.A.; Chanock, S.J.; Boyer, T.D.; Young, N.S. Constitutional telomerase mutations are genetic risk factors for cirrhosis. Hepatology 2011, 53, 1600–1607. [Google Scholar] [CrossRef]
- Kordowitzki, P. Oxidative stress induces telomere dysfunction and shortening in human oocytes of advanced age donors. Cells 2021, 10, 1866. [Google Scholar] [CrossRef] [PubMed]
- de La Roche Saint-André, C. Alternative ends: telomeres and meiosis. Biochimie 2008, 90, 181–189. [Google Scholar] [CrossRef]
- Morell, M.A.; Werling, U.; Edelmann, W.; Roberson, M.S.; Cohen, P.E. Analysis of meiotic prophase I in live mouse spermatocytes. Chromosome Res 2008, 6, 743–760. [Google Scholar] [CrossRef]
- Liu, L.; Franco, S.; Spyropoulos, B.; Moens, P.B.; Blasco, M.A.; Keefe, D.L. Irregular telomeres impair meiotic synapsis and recombination in mice. Proc Natl Acad Sci U S A 2004, 101, 6496–6501. [Google Scholar] [CrossRef]
- Keefe, D.L.; Franco, S.; Liu, L.; Trimarchi, J.; Cao, B.; Weitzen, S.; Agarwal, S.; Blasco, M.A. Telomere length predicts embryo fragmentation after in vitro fertilization in women--toward a telomere theory of reproductive aging in women. Am J Obstet Gynecol 2005, 192, 1256–1260. [Google Scholar] [CrossRef] [PubMed]
- Keefe, D.L.; Liu, L.; Marquard, K. Telomeres and aging-related meiotic dysfunction in women. Cell Mol Life Sci 2007, 64, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Keefe, D.L. Telomeres, reproductive aging, and genomic instability during early development. Reprod Sci 2016, 23, 1612–1615. [Google Scholar] [CrossRef] [PubMed]
- Esencan, E.; Cozzolino, M.; Imamoglu, G.; Seli, E. Mitochondrial stress response gene Clpp is not required for granulosa cell function. Antioxidants 2020, 10. [Google Scholar] [CrossRef]
- Morin, S.J.; Tao, X.; Marin, D.; Zhan, Y.; Landis, J.; Bedard, J.; Scott, R.T.; Seli, E. DNA methylation-based age prediction and telomere length in white blood cells and cumulus cells of infertile women with normal or poor response to ovarian stimulation. Aging 2018, 10, 3761–3773. [Google Scholar] [CrossRef]
- Hanson, B.M.; Tao, X.; Zhan, Y.; Jenkins, T.G.; Morin, S.J.; Scott, R.T.; Seli, E.U. Young women with poor ovarian response exhibit epigenetic age acceleration based on evaluation of white blood cells using a DNA methylation-derived age prediction model. Hum Reprod 2020 35, 2579–2588. [CrossRef]
- Hanson, B.M.; Tao, X.; Zhan, Y.; Kim, J.G.; Klimczak, A.M.; Herlihy, N.S.; Scott, R.T.J.; Seli, E. Shorter telomere length of white blood cells is associated with higher rates of aneuploidy among infertile women undergoing in vitro fertilization. Fertil Steril 2021, 115, 957–965. [Google Scholar] [CrossRef]
- Lin, X.; Gu, J.; Lu, C.; Spitz, M.R.; Wu, X. Expression of telomere-associated genes as prognostic markers for overall survival in patients with non–small cell lung cancer. Clin Cancer Res 2006, 12, 5720–5725. [Google Scholar] [CrossRef]
Figure 1.
Telomere length in oocytes, ovaries, and somatic tissues of 2-month-old Clpp-/- and WT mice. (A) The standard curve was generated by serial dilution of known amounts of DNA to calculate relative DNA concentrations (log DNA) from Ct values of the qPCR products. Blue dots: telomere; orange dots: 36B4 single copy gene (control). The correlation regression equation and coefficients (R2) of Ct versus log DNA are shown. (B,C) The relative telomere lengths of GV and MII oocytes, ovaries and somatic cells and tissues are represented as ratios of T/S. GV: Germinal vesicle, CC: Cumulus cells, WBC: White blood cells. Data presented as mean ± SEM. **p < 0.01, *p < 0.05 using t-test.
Figure 1.
Telomere length in oocytes, ovaries, and somatic tissues of 2-month-old Clpp-/- and WT mice. (A) The standard curve was generated by serial dilution of known amounts of DNA to calculate relative DNA concentrations (log DNA) from Ct values of the qPCR products. Blue dots: telomere; orange dots: 36B4 single copy gene (control). The correlation regression equation and coefficients (R2) of Ct versus log DNA are shown. (B,C) The relative telomere lengths of GV and MII oocytes, ovaries and somatic cells and tissues are represented as ratios of T/S. GV: Germinal vesicle, CC: Cumulus cells, WBC: White blood cells. Data presented as mean ± SEM. **p < 0.01, *p < 0.05 using t-test.
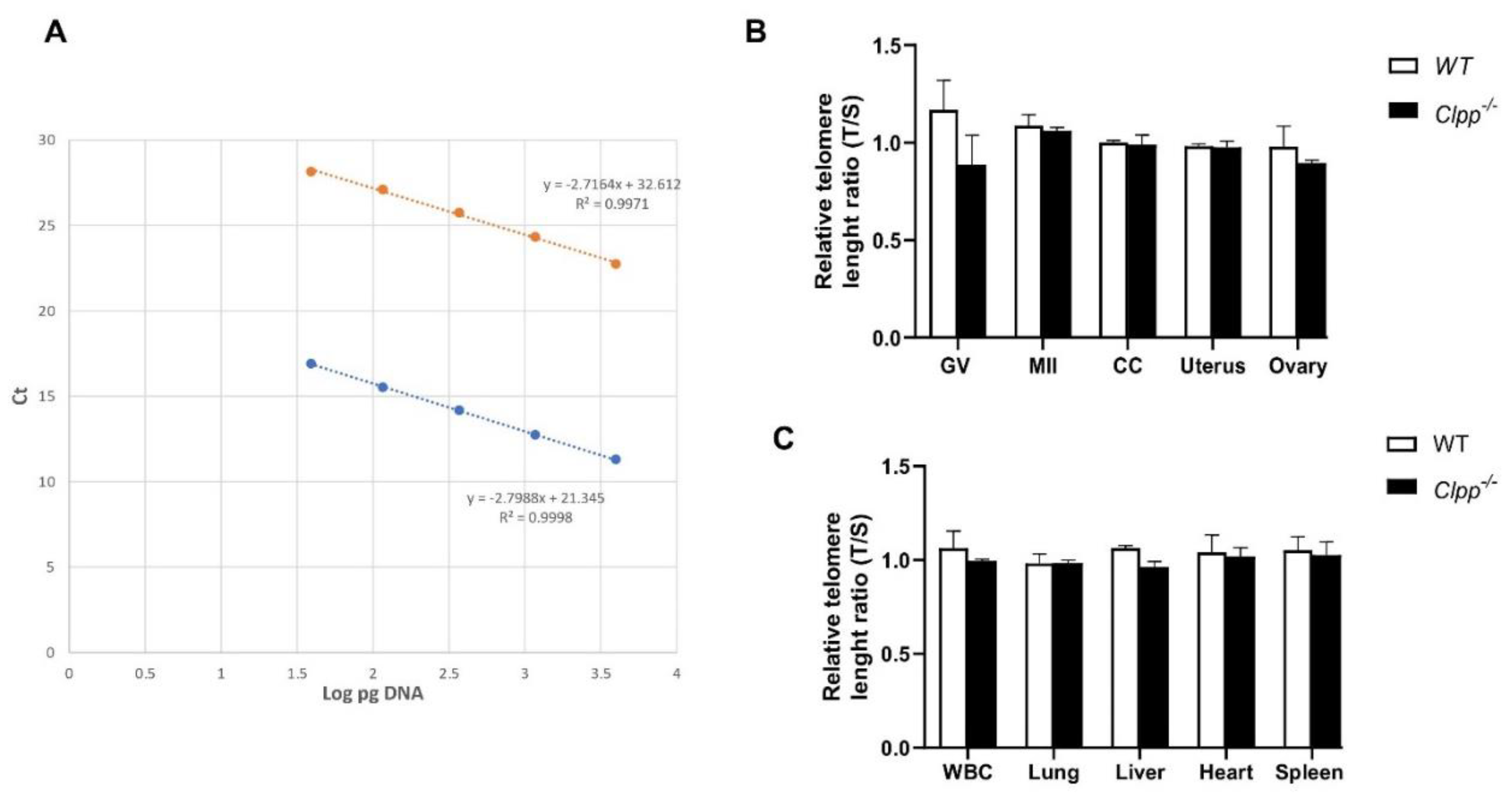
Figure 2.
Telomere length in oocytes, ovaries, and somatic tissues of 6-month-old Clpp-/- and WT mice. (A) The standard curve was generated by serial dilution of known amounts of DNA to calculate relative DNA concentrations (log DNA) from Ct values of the qPCR products. Blue dots: telomere; orange dots: 36B4 single copy gene (control). The correlation regression equation and coefficients (R2) of Ct versus log DNA are shown. (B,C) The relative telomere lengths of GV and MII oocytes, ovaries and somatic cells and tissues are represented as ratios of T/S. GV: Germinal vesicle, CC: Cumulus cells, WBC: White blood cells. Data presented as mean ± SEM. **p < 0.01, *p < 0.05 using t-test.
Figure 2.
Telomere length in oocytes, ovaries, and somatic tissues of 6-month-old Clpp-/- and WT mice. (A) The standard curve was generated by serial dilution of known amounts of DNA to calculate relative DNA concentrations (log DNA) from Ct values of the qPCR products. Blue dots: telomere; orange dots: 36B4 single copy gene (control). The correlation regression equation and coefficients (R2) of Ct versus log DNA are shown. (B,C) The relative telomere lengths of GV and MII oocytes, ovaries and somatic cells and tissues are represented as ratios of T/S. GV: Germinal vesicle, CC: Cumulus cells, WBC: White blood cells. Data presented as mean ± SEM. **p < 0.01, *p < 0.05 using t-test.

Figure 3.
Telomere length in oocytes, ovaries, and somatic tissues of 9-month-old Clpp-/- and WT mice. (A) The standard curve was generated by serial dilution of known amounts of DNA to calculate relative DNA concentrations (log DNA) from Ct values of the qPCR products. Blue dots: telomere; orange dots: 36B4 single copy gene (control). The correlation regression equation and coefficients (R2) of Ct versus log DNA are shown. (B,C) The relative telomere lengths of GV and MII oocytes, ovaries and somatic cells and tissues are represented as ratios of T/S. GV: Germinal vesicle, CC: Cumulus cells, WBC: White blood cells. Data presented as mean ± SEM. **p < 0.01, *p < 0.05 using t-test.
Figure 3.
Telomere length in oocytes, ovaries, and somatic tissues of 9-month-old Clpp-/- and WT mice. (A) The standard curve was generated by serial dilution of known amounts of DNA to calculate relative DNA concentrations (log DNA) from Ct values of the qPCR products. Blue dots: telomere; orange dots: 36B4 single copy gene (control). The correlation regression equation and coefficients (R2) of Ct versus log DNA are shown. (B,C) The relative telomere lengths of GV and MII oocytes, ovaries and somatic cells and tissues are represented as ratios of T/S. GV: Germinal vesicle, CC: Cumulus cells, WBC: White blood cells. Data presented as mean ± SEM. **p < 0.01, *p < 0.05 using t-test.
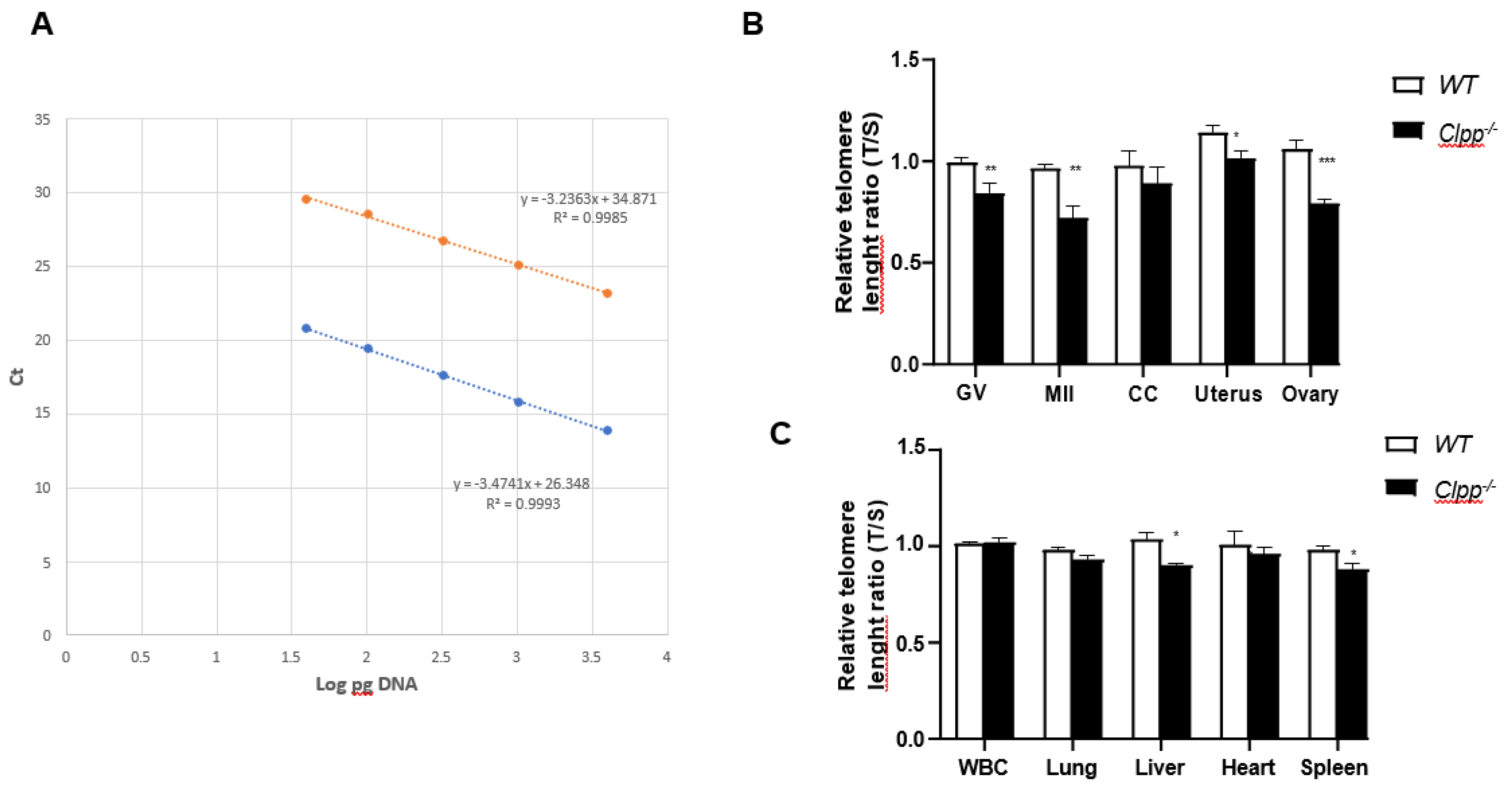
Figure 4.
Expression of telomere-associated genes in Clpp -/- and WT oocytes. Expression of telomere-associated genes was assessed using qRT-PCR in GV oocytes collected from 2-month-old (A) and 6-month-old (B) Clpp-/- and WT mice. Data presented as mean ± SEM with t-test (**p < 0.01, *p < 0.05).
Figure 4.
Expression of telomere-associated genes in Clpp -/- and WT oocytes. Expression of telomere-associated genes was assessed using qRT-PCR in GV oocytes collected from 2-month-old (A) and 6-month-old (B) Clpp-/- and WT mice. Data presented as mean ± SEM with t-test (**p < 0.01, *p < 0.05).
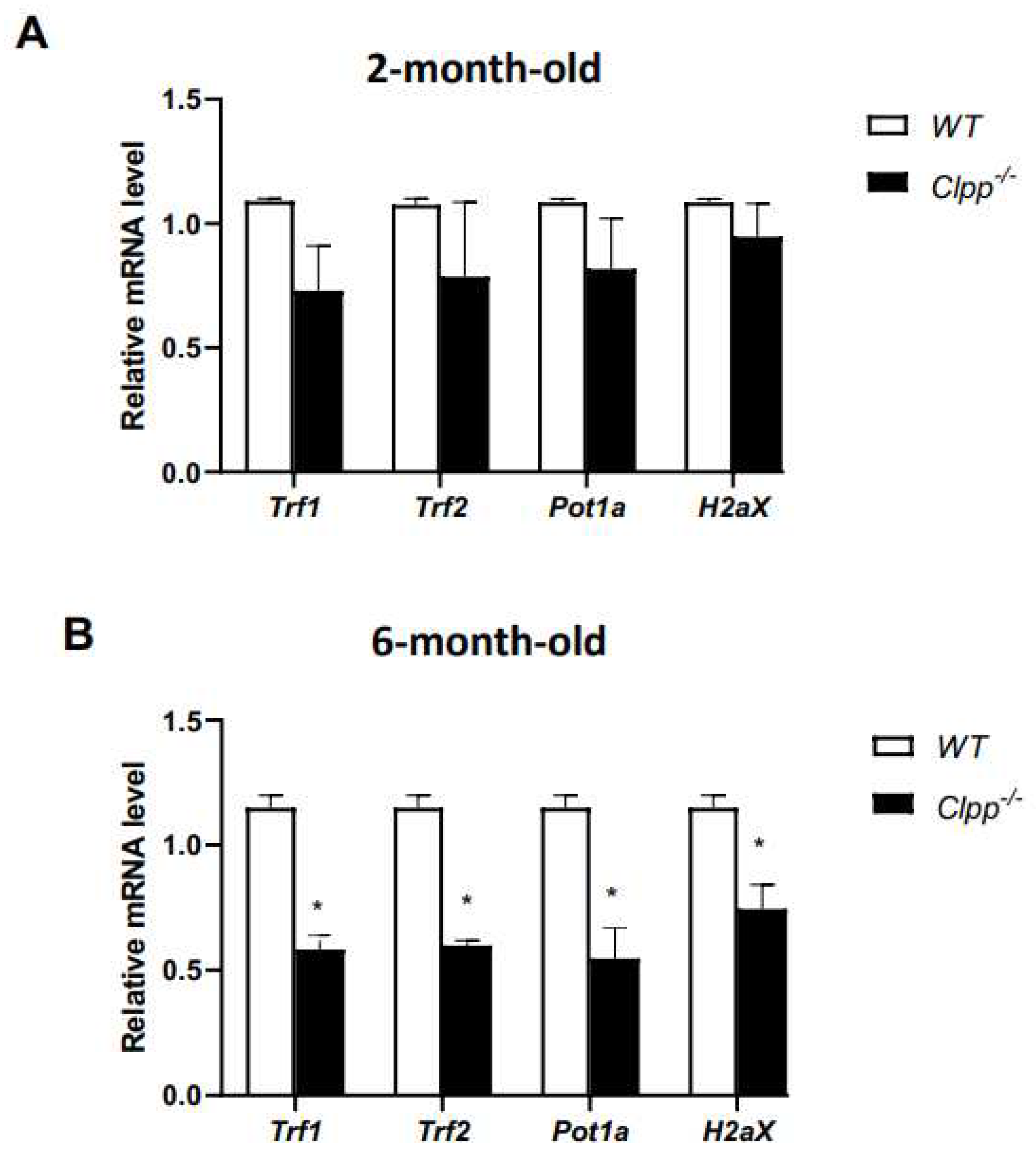
Figure 5.
TRF1 expression and TRF/H2AX co-localization in 2-month-old Clpp -/- and WT mice oocytes. (A) Immunofluorescence double staining of TRF1 (red) and H2AX (green) in cumulus oophorus complexes of 2-month-old Clpp-/- and WT mice. DAPI was used to stain nuclei (blue). (B) Quantitative analysis of TRF1 immunofluorescence in Clpp-/- and WT GV oocytes. (C) Quantitative analysis of co-localization of H2AX and TRF1 in Clpp-/- and WT GV oocytes. Data presented as mean ± SEM with t-test (**p < 0.01, *p < 0.05).
Figure 5.
TRF1 expression and TRF/H2AX co-localization in 2-month-old Clpp -/- and WT mice oocytes. (A) Immunofluorescence double staining of TRF1 (red) and H2AX (green) in cumulus oophorus complexes of 2-month-old Clpp-/- and WT mice. DAPI was used to stain nuclei (blue). (B) Quantitative analysis of TRF1 immunofluorescence in Clpp-/- and WT GV oocytes. (C) Quantitative analysis of co-localization of H2AX and TRF1 in Clpp-/- and WT GV oocytes. Data presented as mean ± SEM with t-test (**p < 0.01, *p < 0.05).
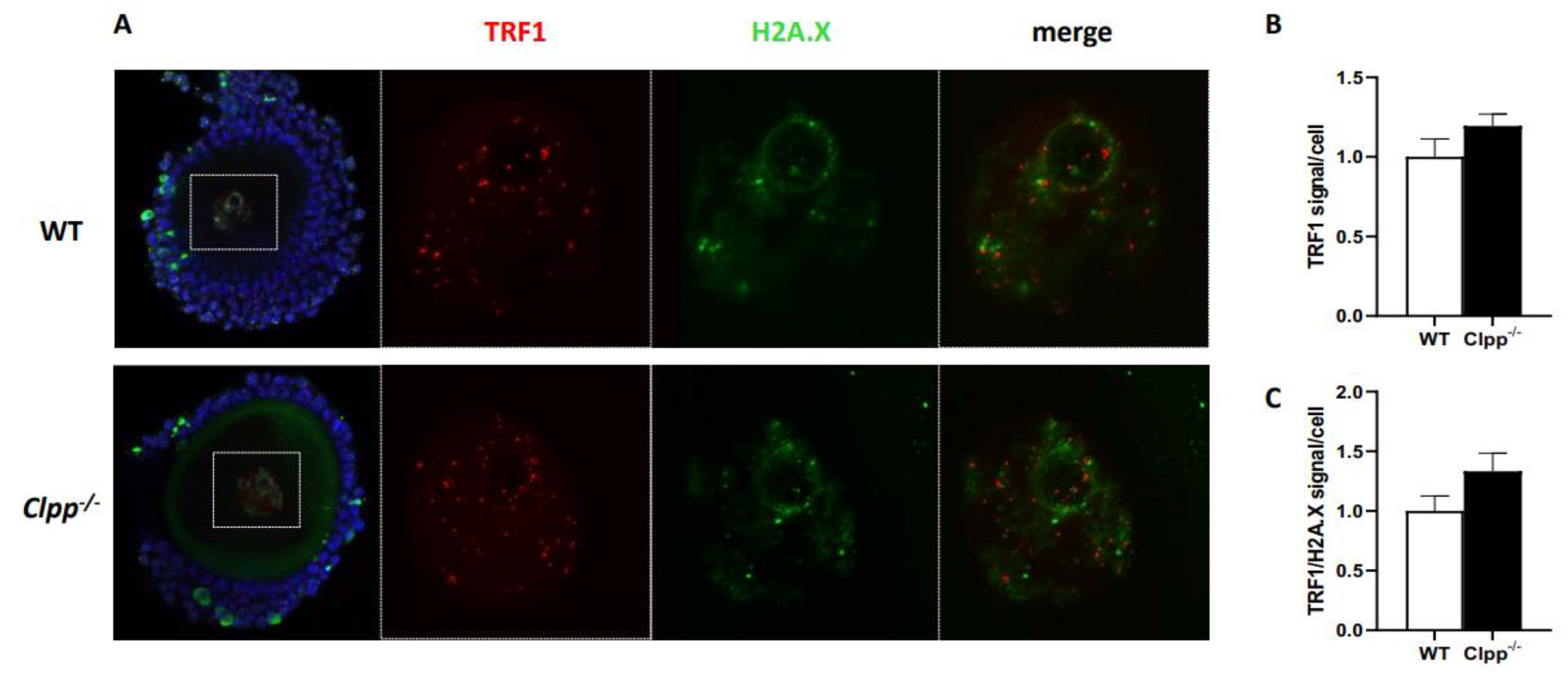
Figure 6.
TRF1 expression and TRF/H2AX co-localization in 6-month-old Clpp -/- and WT mice oocytes. (A) Immunofluorescence double staining of TRF1 (red) and H2AX (green) in cumulus oophorus complexes of 6-month-old Clpp-/- and WT mice. DAPI was used to stain nuclei (blue). (B) Quantitative analysis of TRF1 immunofluorescence in Clpp-/- and WT GV oocytes. (C) Quantitative analysis of co-localization of H2AX and TRF1 in Clpp-/- and WT GV oocytes. Data presented as mean ± SEM with t-test (**p < 0.01, *p < 0.05).
Figure 6.
TRF1 expression and TRF/H2AX co-localization in 6-month-old Clpp -/- and WT mice oocytes. (A) Immunofluorescence double staining of TRF1 (red) and H2AX (green) in cumulus oophorus complexes of 6-month-old Clpp-/- and WT mice. DAPI was used to stain nuclei (blue). (B) Quantitative analysis of TRF1 immunofluorescence in Clpp-/- and WT GV oocytes. (C) Quantitative analysis of co-localization of H2AX and TRF1 in Clpp-/- and WT GV oocytes. Data presented as mean ± SEM with t-test (**p < 0.01, *p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



