
Submitted:
24 May 2023
Posted:
26 May 2023
You are already at the latest version
Abstract
Among the five human senses, light, sound, and force perceived by eye, ear, and skin, respectively are physical actions, and therefore can be easily measured and expressed as objective, univocal, and simple digital data with physical quantity. However, taste and odor molecules perceived by tongue and nose are chemical actions, it has been difficult to express them as objective and univocal digital data, since no reference chemicals can be defined. Therefore, while the recording, saving, transmitting to remote locations, and replaying human visual, auditory, and tactile information as digital data in digital devices has been realized (this series of data flow is defined as DX (digital transformation) in this review), the DX of human taste and odor information is not yet in the realization stage. Particularly, since there are at least 400,000 types of odor molecules and an infinite number of complex odors that are mixtures of these molecules, it has been considered extremely difficult to realize "human olfactory DX" by converting all odors perceived by the human olfaction into digital data. In this review, we discuss the current status and future prospects of the development of "human olfactory DX," which we believe can be realized by utilizing odor sensors that employ the olfactory receptors (ORs) that support human olfaction as sensing molecules (i.e., human OR sensor).
Keywords:
olfactory receptor
; odor sensor
; cell array sensor
; odor matrix
; odor matrix library
1. Introduction
For implementing "recording, saving, transmitting to remote locations, and replaying " of human olfactory information (defined as human olfactory DX; Figure 1) in the next generation of information devices, it is necessary to represent all odors (whether simple or complex) perceived by the human olfaction as objective and univocal digital data in a simple common format as much as possible. The conventional odor quality evaluating methods used for this purpose include metal oxide-based or organic polymer-based semiconductor sensors that can only detect a limited number of odor molecules, GC-MS (gas chromatography-mass spectrometry) that detects all gas molecules, even those that do not smell, and sensory tests that depend on each individual's olfactory characteristics, but these methods are not necessarily sufficient for the human olfactory DX. In human olfaction, olfactory receptors (ORs; the number of all human ORs is defined as 388 in this review) are expressed on olfactory sensory neurons (OSNs) in the olfactory epithelium. Each OR is activated at different intensities for each odor molecule, and human olfaction recognizes odors with an overall activation pattern [1] (Figure 2). It is thought that if the activation intensity of each OR is used as an index, almost all odors (simple and complex) perceived by human olfaction will be capable of representing 388 dimensional parameters.
2. Establishment of Heterologous Cells Expressing ORs
Many attempts have been so far made to identify OR groups that respond to any odor molecule by using OSNs isolated from mouse olfactory epithelium (i.e., deorphanization). Recently, a robot with built-in time-lapse single-cell array cytometry has been developed to automatically pick up only one OSN that responds to odor molecule from the cell array containing mouse olfactory epithelium-derived cells [2], but it is still necessary to isolate olfactory epithelium from mouse neonates with complicated procedures. And it is ethically difficult to isolate human olfactory epithelium. Thus, the recent major method is to create a library of all human OR-expressing cells by recombinant DNA technology and screen odor-responsive OR-expressing cells by using any given odor [3]. However, when ORs are expressed in heterologous cells, they often aggregate and accumulate in the endoplasmic reticulum (ER) and are degraded [4], making it difficult to express all human ORs on the cell surface and enable them to respond to odor molecules. Matsunami et al. found that human embryonic kidney-derived HEK293 cells introduced with the chaperones RTP1 (receptor-transporting protein 1), RTP2, and REEP1 (receptor expression-enhancing protein 1), also called as Hana3A cells, greatly improved cell surface expression of ORs with an N-terminal Rho-tag (rhodopsin-derived signal peptide) [5] (Figure 3). In particularly, RTP1S, a C-terminal part shortened RTP1, more strongly improved the cell surface expression of ORs and odor molecule responses [6]. The Lucy-tag [7] and the IL-6-Halo-tag [8] also enabled cell surface expression of a wider range of ORs than the Rho-tag. Furthermore, co-expression of non-OR GPCRs (G protein-coupled receptors) (e.g., β2-adrenergic receptor, M3 muscarinic acetylcholine receptor) formed heterodimers with ORs and improved their sorting to the cell surface [9,10]. Co-expression of M3 muscarinic acetylcholine receptor has been found to suppress β-arrestin 2-mediated OR internalization [11,12]. Other improvements in second messenger (cAMP) generation and detection systems have also been achieved through the co-expression of olfactory-specific G protein α GNAL/Gαolf [13], which has high affinity for ORs, Ric-8B [14], a chaperone of Gα protein, and GloSensorTM [15], a highly sensitive luciferase for cAMP detection. The above improvements have made the majority of human ORs presentable on the cell surface of HEK293 cells, increasing the likelihood of identifying a group of ORs that respond to any given odor molecule [16]. More recently, common structural features of ORs that are expressed on the cell surface independent of RTPs have been found [17,18]. These structural features, unfortunately, cannot be applied to the human ORs for comprehensive deorphanization, since the mutant ORs harboring these structural features may alter the OR's inherent ability to recognize odor molecules.
3. Importance of Measuring OR Response in Real Time
The attempts to improve cell surface expression and odor molecule responsiveness of ORs as described in Section 2 are very important, but most of these improved cells have been used exclusively for the endpoint measurements of cAMP generation after prolonged odor stimulation (30 min to several days). Continuous exposure of odor to OR-expressing cells for a long period of time should be avoided because some odor molecules are easily denatured even at room temperature (e.g., oxidation, hydrolysis) or cytotoxic, thereby OR responses to these odor molecules will be different from the original OR responses.
In addition, human olfaction responds immediately after odor stimulation and adapts within a few minutes. This process over time is highly dependent on the OSN’s OR response to odor. While the best way to realize human olfactory DX is to measure OSN action potential changes in real time, at present it is difficult to measure all membrane potential changes in many OR-expressing cells at once in real time. Therefore, as a practical method, it is important to measure intracellular Ca2+ influx in real time, since CNG (cyclic nucleotide-activated channel) induces membrane potential changes in OSNs by mobilizes Ca2+ ion into the cell simultaneously with Na+ ion [19].
4. Human Olfactory Receptor-expressing Cell Array Sensor (Human OR Sensor)
For comprehensive measurement of the real-time response of all human ORs to various odor molecules simultaneously, CNGs and Ca2+-dependent fluorescent protein (GCaMP) were stably expressed in HEK293T cells optimized for OR expression as described in Section. 2, followed by expression of each of the 388 human ORs [20] (Figure 4). Next, 0.5 mm square microwells were printed in 20 rows by 20 columns on a glass slide (total 400 microwells) with hydrophobic ink, and a dispensing robot was used to make a cell array with approximately 400-500 cells of each OR-expressing cell in each microwell. After 48 h incubation, the cell array was set in a reflux apparatus under a fluorescence microscope equipped with a video camera. This system for simultaneous and real-time measurement of the response of all human ORs to odor is called a human olfactory receptor-expressing cell array sensor (hereafter abbreviated as human OR sensor; Figure 5). Ringer's solution dissolving the odor of interest (whether simple or complex) was refluxed continuously, and a fluorescent image of the entire array was captured on video to record the fluorescence emitted by cells expressing ORs responsive to the odor molecules for 7 min. Then, all fluorescence intensity changes were datamined at a single cell basis (4 pixels per cell) by ImageJ program ver. 1.52u (Image processing and analysis in Java; Wayne Rasband, National Institute of Health, Bethesda, MD, U.S.A.). Each cell emits spontaneously numerous nonspecific fluorescence due to the intracellular Ca2+ mobilization by normal cellular activity. Therefore, using an AI program that machine-learned a large number of temporal changes in fluorescence emitted by cells expressing odor-responded ORs, the actual changes in fluorescence intensity due to odor-responded ORs were extracted. Dozens of the actual changes in fluorescence intensity per OR were extracted and averaged. By this method, variation of fluorescence intensity changes due to biological fluctuations in OR-expressing cells could be suppressed to around 10% CV (coefficient of variation). Various odors, both simple and complex, can be represented by the response intensity of 388 human ORs (called the odor matrix; see Figure 5). Note that this odor matrix also includes changes over time for each OR, since the same OR showed different changes in fluorescence intensity for different odor molecules [20]. For easier understanding, a snapshot of the time when the fluorescence intensity reached its maximum is usually used. Although a similar real-time measurement of OR response can be performed with FLIPR (fluorescent imaging plate reader) [21], which is used by many pharmaceutical companies to screen GPCR agonists and antagonists, this human OR sensor based on cell array technology does not require many microplates or reagents, making it much faster and cheaper to measure different conditions sequentially without replacing the cell array sensor in the same measurement set.
5. Towards Human Olfactory DX Realization
Comprehensive deorphanization to determine exactly how all ORs recognize various odors in human olfaction remains challenging for current human OR-expressing cells. First, when the Gα protein in the cells used for OR expression is different from the OSN, the odor molecular response of some ORs may be altered [22]. Second, single nucleotide polymorphism (SNP) is found in many human OR genes, which sometimes alters the odor molecular response of OR [23,24,25]. Third, almost all odor sensors using OR-expressing cells measure the OR response to odors in the liquid phase. In human olfaction, the ORs of OSNs respond to odors in the gas phase via a very small amount of the nasal discharge, resulting in the detection threshold is much lower (typically 100- to 1000-fold) compared to the OR-based sensors. Currently, we cannot correctly explain this large difference in OR detection thresholds for odors. Indeed, these three issues are major obstacles to unraveling the scientific proposition of how the entire human ORs discriminates various odors with high sensitivity. However, for realizing human olfactory DX (especially in the parts of recording, saving, transmitting to remote locations), an odor sensor does not necessary perfectly reproduce the way human olfaction perceives odors. It is sufficient to always measure odors with representative OR set (i.e., 388 ORs) under fixed conditions and define them in a 388-dimensional odor matrix.
6. Preliminary Odor Reconstitution
So far, the original odor has been reconstituted by analyzing its components by GC-MS and mixing the major odor molecules in the same proportions. However, odor molecules that cannot be detected by GC-MS often contribute significantly to the odor quality, so perfumers had to spend a lot of time for formulation with many odor molecules by trial and error. For the achievement of the reconstitution part of the human olfactory DX, this process would be virtually impossible without preparing in advance all the approximately 400,000 types of odor molecules that exist in the world [26]. In other words, it is imperative to reduce the number of odor molecules used for the odor reconstitution of human olfactory DX. There is an attempt to reduce the number of odor molecules by GC-MS analysis of essential oils to sort out odor molecules that exhibit the same odor quality and reconstitute the odor quality of any given essential oil [27], but this method is applicable only to specific categories of odors (e.g., essential oils) and cannot be used to reconstitute a wide range of all odors.
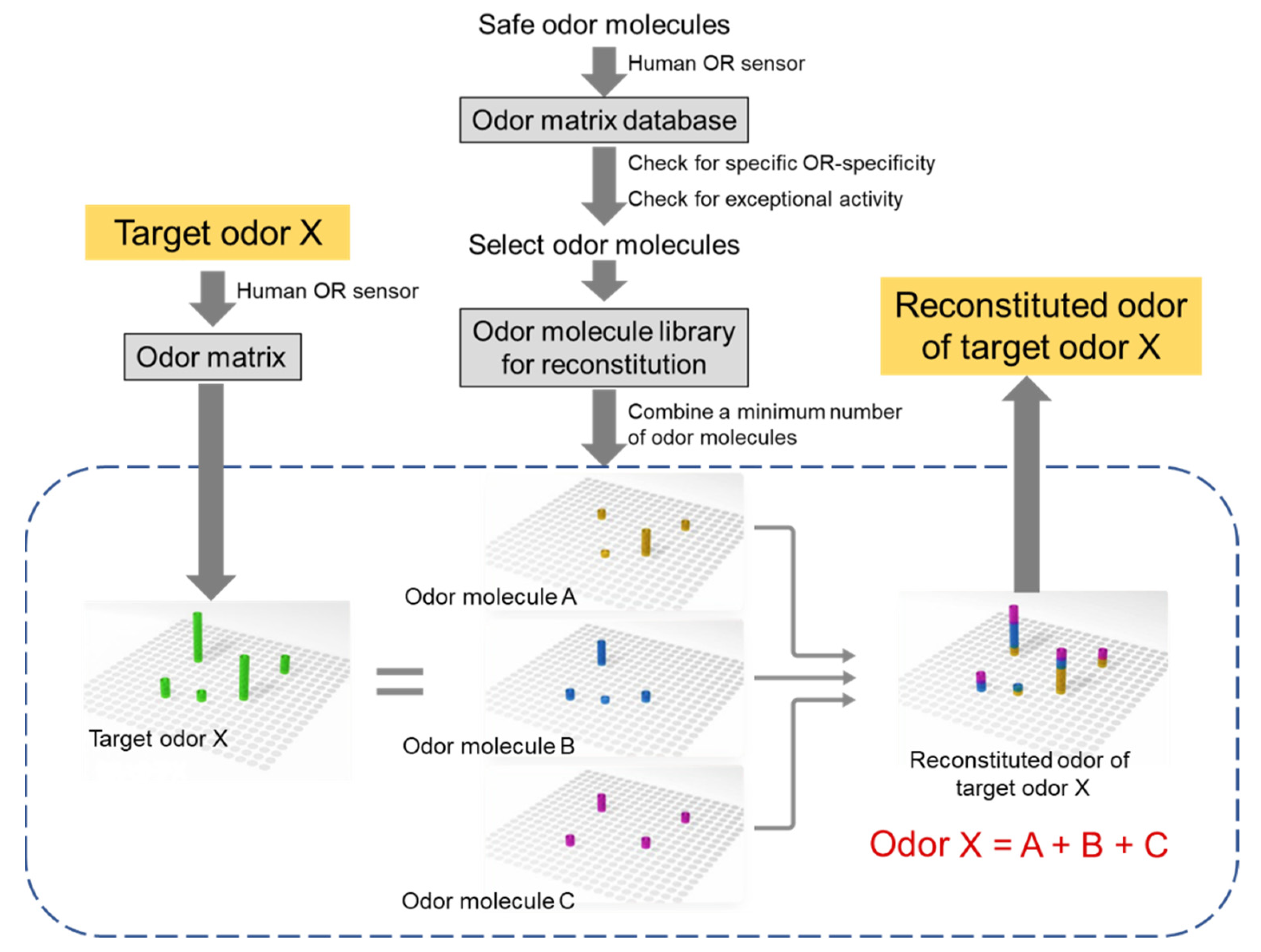
Thus, by taking advantage of human OR sensor, we preliminarily reconstituted the odor of dried bonito flakes, rose essential oil, lavender essential oil, and vanilla flavor. Even though each sample was found to contain numerous types of odor molecules (>500, >50, >50, and >50, respectively) by GC-MS, 10 human ORs responded significantly to dried bonito flakes, 4 ORs to rose essential oil, 3 ORs to lavender essential oil, and 7 ORs to vanilla flavor were identified by human OR sensor. In addition, several of the responsive ORs were common. Therefore, odor molecules that pinpointly stimulate these ORs were selected from odor molecule library and mixed in a ratio that could reproduce the odor matrix. Finally, the odor of dried bonito flakes, rose essential oil, lavender essential oil, and vanilla flavor were reconstituted with 7, 4, 3, and 5 odor molecules that were not included in the respective original samples, of which the odor quality was determined by sensory tests on a scale of several dozen people to be nearly reproduced [28]. This result strongly suggested that no matter how complex an odor is, if an odor matrix is obtained by a human OR sensor and mixed with odor molecules that stimulate each response OR with pinpoint accuracy, it is possible to reconstitute the odor with far fewer types of odor molecules (Figure 6).
In general, the OSN responses to complex odors is essentially a linear relationship with the OSN response of each odor molecule [29]. Nevertheless, there is also intermolecular interference between odor molecules when multiple odor molecules bind to an OR [30]. This competitive binding result in a nonlinear relationship between the OR responses of multiple odor molecules [31]. These are also explained by a model in which odor molecules bind to allosteric pockets on the OR. In addition, there are inverse agonists in which the same odor molecules promote or inhibit OR responses in a concentration-dependent manner [32]. Human OR sensor capable of measuring even complex odors can detect odor molecules eliciting these exceptional OR responses. Only odor molecules that pinpointly stimulate limited number of ORs without the exceptional activities to ORs should be used to reconstitute odors that have the almost same odor quality as any odor.
7. Future Improvements of Human OR Sensor
The main issue with human OR sensors is that their detection threshold for odor molecule is higher than that of human olfaction. To ameliorate this issue, we should consider to suppress mechanisms that inhibit intracellular cAMP generation (PDE (phosphodiesterase)) and intracellular Ca2+ mobilization (CaMKII (Ca2+-calmodulin-dependent protein kinase II), CaM (calmodulin), NCX (Na+/Ca2+ exchanger), PMCA (plasma membrane calcium pump)) [33,34,35], utilize extracellular OBPs (odorant-binding proteins) that promote odor molecule presentation to the ORs [36], utilize intracellular OMPs (olfactory marker proteins) that increase intracellular cAMP levels [37,38], and utilize GRK2 (G-protein-coupled receptor kinase 2) inhibitors that suppress the binding of β-arrestins that promote OR internalization [39].
Recently, there have been some reports that conventional ORs are not enough to detect all odors, human OR sensor should also be equipped with OSN-specific TAARs (trace amine-associated receptors; TAAR1, TAAR2, TAAR5, TAAR6, TAAR8, TAAR9) [40,41,42] to detect amine compounds and TRPs (transient receptor potential channels; 6 subfamily, 27 TRPs) [43,44] to detect odor molecules such as capsaicin and menthol. Since the second messenger of TAARs and TRPs is Ca2+ ion, the degree of activation of both TAARs and TRPs can be measured by simply diverting the cells used in the OR-expressing cells of human OR sensors.
The sensitivity of human olfaction is known to decline with age [45], but it is not a problem for daily life. This fact presents the proposition if all ORs is really necessary or not. Recently, not all OR mRNAs have found in human olfactory epithelium by RNAseq analysis [46]. All OR mRNAs were not equally expressed and 26 major OR mRNAs accounted for more than 90% of the total OR mRNAs [47]. These facts indicate that the number of ORs in current human OR sensors may be overrepresented.
8. Possible Alternatives for Human OR Sensor
So far, it has been time-consuming and labor-intensive to perform comprehensive deorphanization of all human ORs by using conventional OR-based sensors, thus OR databases (ODRactor, OlfactionBase DB) have been built to predict which ORs respond to any given odor molecules from OR structures [48,49,50]. Recently, deorphanization has been made possible by using a machine-learned proteochemometric model based on the structure of odor molecules and the orthosteric pocket sequences of responded OR [51]. In addition, an AI that learned the experimental results of interactions between odor molecules and OR structures succeeded in predicting the responsive ORs to any given odor molecules with over 90% correct response rate [52]. These predictions are promising, but they need to be biologically validated in any case. In the future, these methods will certainly complement the human OR sensor method for the human OR deorphanization.
Finally, because human OR sensors utilize living cells, odor measurement must always be performed in the laboratory after the sample has been collected elsewhere. Thus, it is not possible to use human OR sensors easily and readily in all settings. Recently, electrical odor sensors using only human OR proteins as the sensing molecule has been experimentally fabricated as eNose, which will compensate for the low portability of the human OR sensor [53]. For example, FET (field effect transistor) sensor in which liposome-based mini-cells incorporating ORs, adenylate cyclase, and CNG are immobilized via CNT (carbon nanotube) [54], multichannel FET sensor in which multiple ORs are directly immobilized via CNTs [55], and FET sensor immobilized with ORs embedded in a lipid membrane (nanodisc) [56] have been developed. These FET sensors using only human OR proteins surprisingly showed very low detection thresholds for odor molecules, comparable to human olfaction. This fact strongly suggests that the high detection threshold of the current human OR sensors is not caused by the OR molecules themselves, but by the intracellular signal transduction mechanism elicited by the ORs. These FET sensors, albeit highly sensitive, only detect interactions between odor molecules and ORs, and it may be difficult to correlate them with the odor matrix, which reflects the temporal changes of second messengers inside OSNs.
9. Conclusions
The emergence of human OR sensor has largely solved the technical issues that hampered the realization of human olfactory DX, though not sufficient for a comprehensive understanding of the odor recognition mechanism by the human olfaction. From now on, it is necessary to build up an “odor matrix database” of a huge number of odors while improving the human OR sensor (Figure 6). In this process, instead of randomly measuring odors everywhere, the odor molecules that are exemplified as covering the sensory perception of human olfaction [57] should be measured first in priority. Thereby, the human OR sensor must show that it can indeed detect almost all odors perceived by the human olfaction. Then, a lot of basically safe odor molecules (e.g., natural flavors and fragrances, synthetic fragrances, food additives) are measured by human OR sensor to obtain odor matrix to enrich the database.
The first step in odor reconstitution is to find ORs that are preferentially used by the human olfaction from the odor matrix database. Then, odor molecules that pinpointly stimulate limited number of ORs (1 OR if possible) without the exceptional activities (see Section 6) should be found from the odor matrix database to create an “odor molecule library for reconstitution.” When reconstituting an odor of interest, the target odor is measured with a human OR sensor to obtain an odor matrix (including changes in each human OR response over time). The minimum group of odor molecules necessary to reproduce this target odor matrix is selected from “the odor molecule library for reconstitution.” The mixing ratio of each odor molecule are then optimized and sprayed from a diffuser or the like to achieve odor reconstitution. The most significant challenge for human olfactory DX is real-time odor reconstitution, which will have a great impact on the world if it is utilized in video equipment that releases the desired odors on demand.
Author Contributions
Conceptualization, S.K.; writing, S.K.; reviewing Y.H.K., K.T., and S.H. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support of this work by KAKENHI (Grant-in-Aid for Challenging Research (Pioneering) from Japan Society for the Promotion of Science (JSPS) Grant Number 18H05359 (20K20370), 22K18343), and Adaptable and Seamless Technology transfer Program through Target-driven R&D (A-STEP) from Japan Science and Technology Agency (JST) Grant Number JPMJTR194C.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is available from the authors on request.
Acknowledgments
The authors are grateful to Mr. Kenji Kubo for his encouragement, Mr. Koji Abe for preparing figures, and the member of R&D Center, Komi Hakko Corp. for their technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Malnic, B. : Hirono, J.; Sato, T.; Buck, L.B. Combinatorial receptor codes for odors. Cell 1999, 96, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Yoshimoto, N.; Shimono, K.; Kuroda. S. Deciphering the receptor repertoire encoding specific odorants by time-lapse single-cell array cytometry. Sci. Rep. 2016, 6, 19934. [CrossRef]
- Peterlin, Z.; Firestein, S.; Rogers, M.E. The state of the art of odorant receptor deorphanization: a report from the orphanage. J. Gen. Physiol. 2014, 143, 527–542. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Echeverri, F.; Moyer, B.D. Endoplasmic reticulum retention, degradation, and aggregation of olfactory G-protein coupled receptors. Traffic 2003, 4, 416–433. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Kubota, M.; Roberts, R.W.; Chi, Q.; Matsunami, H. RTP family members induce functional expression of mammalian odorant receptors. Cell 2004, 119, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Pan, Y.; Chen, G.Q.; Matsunami, H.; Zhuang, H. Receptor-transporting protein 1 short (RTP1S) mediates translocation and activation of odorant receptors by acting through multiple steps. J. Biol. Chem. 2012, 287, 22287–22294. [Google Scholar] [CrossRef] [PubMed]
- Shepard, B.D.; Natarajan, N.; Protzko, R.J.; Acres, O.W.; Pluznick, J.L. A cleavable N-terminal signal peptide promotes widespread olfactory receptor surface expression in HEK293T cells. PLoS One 2013, 8, e68758. [Google Scholar] [CrossRef]
- Noe, F.; Frey, T.; Fiedler, J.; Geithe, C.; Nowak, B.; Krautwurst, D. IL-6-HaloTag® enables live-cell plasma membrane staining, flow cytometry, functional expression, and de-orphaning of recombinant odorant receptors. J. Biol. Methods 2017, 4, e81. [Google Scholar] [CrossRef]
- Hague, C.; Uberti, M.A.; Chen, Z.; Bush, C.F.; Jones, S.V.; Ressler, K.J.; Hall, R.A.; Minneman, K.P. Olfactory receptor surface expression is driven by association with the β2-adrenergic receptor. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 13672–13676. [Google Scholar] [CrossRef]
- Li, Y.R.; Matsunami, H. Activation state of the M3 muscarinic acetylcholine receptor modulates mammalian odorant receptor signaling. Sci. Signal 2011, 4, ra1. [Google Scholar] [CrossRef]
- Mashukova, A.; Spehr, M.; Hatt, H.; Neuhaus, E.M. β-arrestin 2-mediated internalization of mammalian odorant receptors. J. Neurosci. 2006, 26, 9902–9912. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, Y.R.; Tian, H.; Ma, M.; Matsunami, H. Muscarinic acetylcholine receptor M3 modulates odorant receptor activity via inhibition of β-arrestin-2 recruitment. Nat. Commun. 2015, 6, 6448. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Reed, R.R. Golf: an olfactory neuron specific-G protein involved in odorant signal transduction. Science 1989, 244, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Papasergi, M.M.; Patel, B.R.; Tall, G.G. The G protein α chaperone Ric-8 as a potential therapeutic target. Mol. Pharmacol. 2015, 87, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Kida, H.; Fukutani, Y.; Mainland, J.D.; de March, C.A.; Vihani, A.; Li, Y.R.; Chi, Q.; Toyama, A.; Liu, L.; Kameda, M.; Yohda, M.; Matsunami, H. Vapor detection and discrimination with a panel of odorant receptors. Nat. Commun. 2018, 9, 4556. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Chi, Q.; Zhuang, H.; Matsunami, H.; Mainland, J.D. Odor coding by a Mammalian receptor repertoire. Sci. Signal 2009, 2, ra9. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, K.; de March, C.A.; Nagai, M.H.; Ghosh, S.; Do, M.; Sharma, R.; Bruguera, E.S.; Lu, Y.E.; Fukutani, Y.; Vaidehi, N.; Yohda, M.; Matsunami, H. Structural instability and divergence from conserved residues underlie intracellular retention of mammalian odorant receptors. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 2957–2967. [Google Scholar] [CrossRef]
- Fukutani, Y.; Nakamura, Y.; Muto, N.; Miyanaga, S.; Kanemaki, R.; Ikegami, K.; Noguchi, K.; Ohsawa, I.; Matsunami, H.; Yohda, M. Hot spot mutagenesis improves the functional expression of unique mammalian odorant receptors. Int. J. Mol. Sci. 2021, 23, 277. [Google Scholar] [CrossRef]
- Menini, A. Calcium signalling and regulation in olfactory neurons. Curr. Opin. Neurobiol. 1999, 9, 419–426. [Google Scholar] [CrossRef]
- Yasunaga, M.; Takai, E.; Hattori, S.; Tatematsu, K.; Kuroda, S. Effects of 3-octen-2-one on human olfactory receptor responses to vanilla flavor. Biosci. Biotechnol. Biochem. 2022, 86, 1562–1569. [Google Scholar] [CrossRef]
- Emkey, R.; Rankl, N.B. Screening G protein-coupled receptors: measurement of intracellular calcium using the fluometric imaging plate reader. Methods Mol. Biol. 2009, 565, 145-158. [CrossRef]
- Shirokova, E.; Schmiedeberg, K.; Bedner, P.; Niessen, H.; Willecke, K.; Raguse, J.D.; Meyerhof, W.; Krautwurst, D. Identification of specific ligands for orphan olfactory receptors. G protein-dependent agonism and antagonism of odorants. J. Biol. Chem. 2005, 280, 11807-11815. [CrossRef]
- Mainland, J.D.; Keller, A.; Li, Y.R.; Zhou, T.; Trimmer, C.; Snyder, L.L.; Moberly, A.H.; Adipietro, K.A.; Liu, W.L.; Zhuang, H.; Zhan, S.; Lee, S.S.; Lin, A.; Matsunami, H. The missense of smell: functional variability in the human odorant receptor repertoire. Nat. Neurosci. 2014, 17, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, S.R.; McRae, J.F.; Bava, C.M.; Beresford, M.K.; Hunter, D.; Jia, Y.; Chheang, S.L.; Jin, D.; Peng, M.; Gamble, J.C.; Atkinson, K.R.; Axten, L.G.; Paisley, A.G.; Tooman, L.; Pineau, B.; Rouse, S.A.; Newcomb, R.D. A mendelian trait for olfactory sensitivity affects odor experience and food selection. Curr. Biol. 2013, 23, 1601–1605. [Google Scholar] [CrossRef] [PubMed]
- Trimmer, C.; Keller, A.; Murphy, N.R.; Snyder, L.L.; Willer, J.R.; Nagai, M.H.; Katsanis, N.; Vosshall, L.B.; Matsunami, H.; Mainland, J.D. Genetic variation across the human olfactory receptor repertoire alters odor perception. Proc. Natl. Acad. Sci. U. S. A. 2019, 116, 9475–9480. [Google Scholar] [CrossRef] [PubMed]
- Le Magnen, J. Odeurs et Parfums. Collection Que Sais-je 2e ed.; Presses universitaires de France, Paris, France, 1961; pp.128-137.
- Debnath, T.; Nakamoto, T. Extraction of sensing data for desired scent impressions using mass spectra of odorant molecules. Sci. Rep. 2022, 12, 16297. [Google Scholar] [CrossRef]
- Home page of Komi Hakko Corp.: https://komi-hakko.co.jp/archives/393 (accessed on 24/05/2023).
- Gupta, P.; Albeanu, D.F.; Bhalla, U.S. Olfactory bulb coding of odors, mixtures and sniffs is a linear sum of odor time profiles. Nat. Neurosci. 2015, 18, 272–281. [Google Scholar] [CrossRef]
- Reddy, G.; Zak, J.D.; Vergassola, M.; Murthy, V.N. Antagonism in olfactory receptor neurons and its implications for the perception of odor mixtures. Elife 2018, 7, e34958. [Google Scholar] [CrossRef]
- Singh, V.; Murphy, N.R.; Balasubramanian, V,; Mainland, J.D. Competitive binding predicts nonlinear responses of olfactory receptors to complex mixtures. Proc. Natl. Acad. Sci. U. S. A. 2019, 116, 9598-9603. [CrossRef]
- Inagaki, S.; Iwata, R.; Iwamoto, M.; Imai, T. Widespread inhibition, antagonism, and synergy in mouse olfactory sensory neurons in vivo. Cell Rep. 2020, 31, 107814. [Google Scholar] [CrossRef] [PubMed]
- Cygnar, K.D.; Zhao, H. Phosphodiesterase 1C is dispensable for rapid response termination of olfactory sensory neurons. Nat. Neurosci. 2009, 12, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhao, A.Z.; Chan, G.C.; Baker, L.P.; Impey, S.; Beavo, J.A.; Storm, D.R. Phosphorylation and inhibition of olfactory adenylyl cyclase by CaM kinase II in Neurons: a mechanism for attenuation of olfactory signals. Neuron 1998, 21, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Castillo, K.; Delgado, R.; Bacigalupo, J. ;Plasma membrane Ca(2+)-ATPase in the cilia of olfactory receptor neurons: possible role in Ca(2+) clearance. Eur. J. Neurosci. 2007, 26, 2524–2531. [Google Scholar] [CrossRef]
- Matarazzo, V.; Zsürger, N.; Guillemot, J.C.; Clot-Faybesse, O.; Botto, J.M.; Dal Farra, C.; Crowe, M.; Demaille, J.; Vincent, J.P.; Mazella, J.; Ronin, C. Porcine odorant-binding protein selectively binds to a human olfactory receptor. Chem. Senses 2002, 27, 691–701. [Google Scholar] [CrossRef]
- Nakashima, N.; Nakashima, K.; Taura, A.; Takaku-Nakashima, A.; Ohmor,i H.; Takano, M. Olfactory marker protein directly buffers cAMP to avoid depolarization-induced silencing of olfactory receptor neurons. Nat. Commun. 2020, 11, 2188. [CrossRef]
- Nakashima, N.; Nakashima, K.; Nakashima, A.; Takano, M. Olfactory marker protein elevates basal cAMP concentration. Biochem. Biophys. Res. Commun. 2020, 531, 203–208. [Google Scholar] [CrossRef]
- Mayberry, C.L.; Wilczek, M.P.; Fong, T.M.; Nichols, S.L.; Maginnis, M.S. GRK2 mediates β-arrestin interactions with 5-HT2 receptors for JC polyomavirus endocytosis. J. Virol. 2021, 95, e02139–20. [Google Scholar] [CrossRef]
- Wallrabenstein, I.; Kuklan, J.; Weber, L.; Zborala, S.; Werner, M.; Altmüller, J.; Becker, C.; Schmidt, A.; Hatt, H.; Hummel, T.; Gisselmann, G. Human trace amine-associated receptor TAAR5 can be activated by trimethylamine. PLoS One 2013, 8, e54950. [Google Scholar] [CrossRef]
- Xu, Z.; Li, Q. TAAR agonists. Cell. Mol. Neurobiol. 2020, 40, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Gisladottir, R.S.; Ivarsdottir, E.V.; Helgason, A.; Jonsson, L.; Hannesdottir, N.K.; Rutsdottir, G.; Arnadottir, G.A.; Skuladottir, A.; Jonsson, B.A.; Norddahl, G.L.; Ulfarsson, M.O.; Helgason, H.; Halldorsson, B.V.; Nawaz, M.S.; Tragante, V.; Sveinbjornsson, G.; Thorgeirsson, T.; Oddsson, A.; Kristjansson, R.P.; Bjornsdottir, G.; Thorgeirsson, G.; Jonsdottir, I.; Holm, H.; Gudbjartsson, D.F.; Thorsteinsdottir, U.; Stefansson, H.; Sulem, P.; Stefansson, K. Sequence variants in TAAR5 and other loci affect human odor perception and naming. Curr. Biol. 2020, 30, 4643-4653.e3. [CrossRef]
- Voets, T.; Droogmans, G.; Wissenbach, U.; Janssens, A.; Flockerzi, V.; Nilius, B. The principle of temperature-dependent gating in cold- and heat-sensitive TRP channels. Nature 2004, 430, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Koivisto, A.P.; Belvisi, M.G.; Gaudet, R.; Szallasi, A. Advances in TRP channel drug discovery: from target validation to clinical studies. Nat. Rev. Drug Discov. 2022, 21, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Verbeurgt, C.; Wilkin, F.; Tarabichi, M.; Gregoire, F.; Dumont, J.E.; Chatelain, P. Profiling of olfactory receptor gene expression in whole human olfactory mucosa. PLoS One 2014, 9, e96333. [Google Scholar] [CrossRef] [PubMed]
- Olender, T.; Keydar, I.; Pinto, J.M.; Tatarskyy, P.; Alkelai, A.; Chien, M.S.; Fishilevich, S.; Restrepo, D.; Matsunami, H.; Gilad, Y.; Lancet, D. The human olfactory transcriptome. BMC Genomics 2016, 17, 619. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, L.R.; Riveros-McKay, F.; Mezzavilla, M.; Abou-Moussa, E.H.; Arayata, C.J.; Makhlouf, M.; Trimmer, C.; Ibarra-Soria, X.; Khan, M.; Van Gerven, L.; Jorissen, M.; Gibbs, M.; O'Flynn, C.; McGrane, S.; Mombaerts, P.; Marioni, J.C.; Mainland, J.D.; Logan, D.W. A transcriptomic atlas of mammalian olfactory mucosae reveals an evolutionary influence on food odor detection in humans. Sci. Adv. 2019, 5, eaax0396. [Google Scholar] [CrossRef]
- Marenco, L.; Wang, R.; McDougal, R.; Olender, T.; Twik, M.; Bruford, E.; Liu, X.; Zhang, J.; Lancet, D.; Shepherd, G.; Crasto, C. ORDB, HORDE, ODORactor and other on-line knowledge resources of olfactory receptor-odorant interactions. Database (Oxford) 2016, 2016, baw132. [Google Scholar] [CrossRef]
- Liu, X.; Su, X.; Wang, F.; Huang, Z.; Wang, Q.; Li, Z.; Zhang, R.; Wu, L.; Pan, Y.; Chen, Y.; Zhuang, H.; Chen, G.; Shi, T.; Zhang, J. ODORactor: a web server for deciphering olfactory coding. Bioinformatics 2011, 27, 2302–2303. [Google Scholar] [CrossRef]
- Sharma, A.; Saha, B.K.; Kumar, R.; Varadwaj, P.K. OlfactionBase: a repository to explore odors, odorants, olfactory receptors and odorant-receptor interactions. Nucleic Acids Res. 2022, 50, D678–D686. [Google Scholar] [CrossRef]
- Cong, X.; Ren, W.; Pacalon, J.; Xu, R.; Xu, L.; Li, X.; de March, C.A.; Matsunami, H.; Yu, H.; Yu, Y.; Golebiowski, J. Large-scale G protein-coupled olfactory receptor-ligand pairing. ACS Cent. Sci. 2022, 8, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kumar, R.; Semwal, R.; Aier, I.; Tyagi, P.; Varadwaj, P.K. DeepOlf: deep neural network based architecture for predicting odorants and their interacting olfactory receptors. IEEE/ACM Trans. Comput. Biol. Bioinform. 2022, 19, 418-428. [CrossRef]
- Qin, C.; Wang, Y.; Hu, J.; Wang, T.; Liu, D.; Dong, J.; Lu, Y. Artificial olfactory biohybrid system: an evolving sense of smell. Adv. Sci. (Weinh) 2023, 10, e2204726. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Cho, D.G.; Lim, J.H.; Park, J.; Hong, S.; Ko, H.J.; Park, T.H. Real-time monitoring of geosmin and 2-methylisoborneol, representative odor compounds in water pollution using bioelectronic nose with human-like performance. Biosens. Bioelectron. 2015, 74, 199–206. [Google Scholar] [CrossRef]
- Son, M.; Kim, D.; Ko, H.J.; Hong, S.; Park, T.H. A portable and multiplexed bioelectronic sensor using human olfactory and taste receptors. Biosens. Bioelectron. 2017, 87, 901–907. [Google Scholar] [CrossRef]
- Lee, M.; Yang, H.; Kim, D.; Yang, M.; Park, T.H.; Hong, S. Human-like smelling of a rose scent using an olfactory receptor nanodisc-based bioelectronic nose. Sci. Rep. 2018, 8, 13945. [Google Scholar] [CrossRef]
- Castro, J.B.; Ramanathan, A.; Chennubhotla, C.S. Categorical dimensions of human odor descriptor space revealed by non-negative matrix factorization. PLoS One 2013, 8, e73289. [Google Scholar] [CrossRef]
Figure 1.
Conceptual diagram of human olfactory DX. All odors perceived by the human olfaction are recorded as univocal digital data in a simple common format, saved in portable memory device, transmitted to remote locations through radio waves or internet, and replayed by diffuser capable of mixing odor molecules in real time.
Figure 1.
Conceptual diagram of human olfactory DX. All odors perceived by the human olfaction are recorded as univocal digital data in a simple common format, saved in portable memory device, transmitted to remote locations through radio waves or internet, and replayed by diffuser capable of mixing odor molecules in real time.
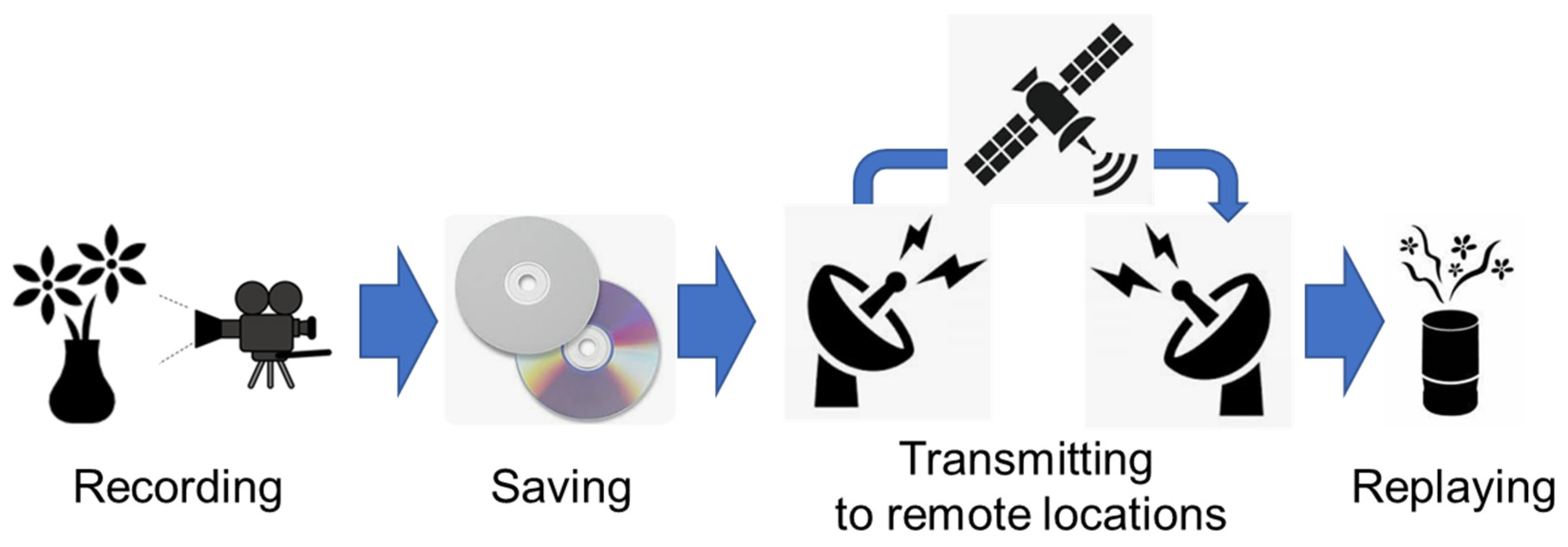
Figure 2.
Olfactory sensory neuron (OSN) and olfactory receptor (OR) in the human olfactory epithelium. Odor molecules entering the nasal cavity are recognized by 388 types of OR on OSN in the olfactory epithelium. At this point, various odor molecules bind to each OR with different intensities, and through pattern recognition by entire ORs, a vast number of odors (whether simple or complex odors) are discriminated.
Figure 2.
Olfactory sensory neuron (OSN) and olfactory receptor (OR) in the human olfactory epithelium. Odor molecules entering the nasal cavity are recognized by 388 types of OR on OSN in the olfactory epithelium. At this point, various odor molecules bind to each OR with different intensities, and through pattern recognition by entire ORs, a vast number of odors (whether simple or complex odors) are discriminated.
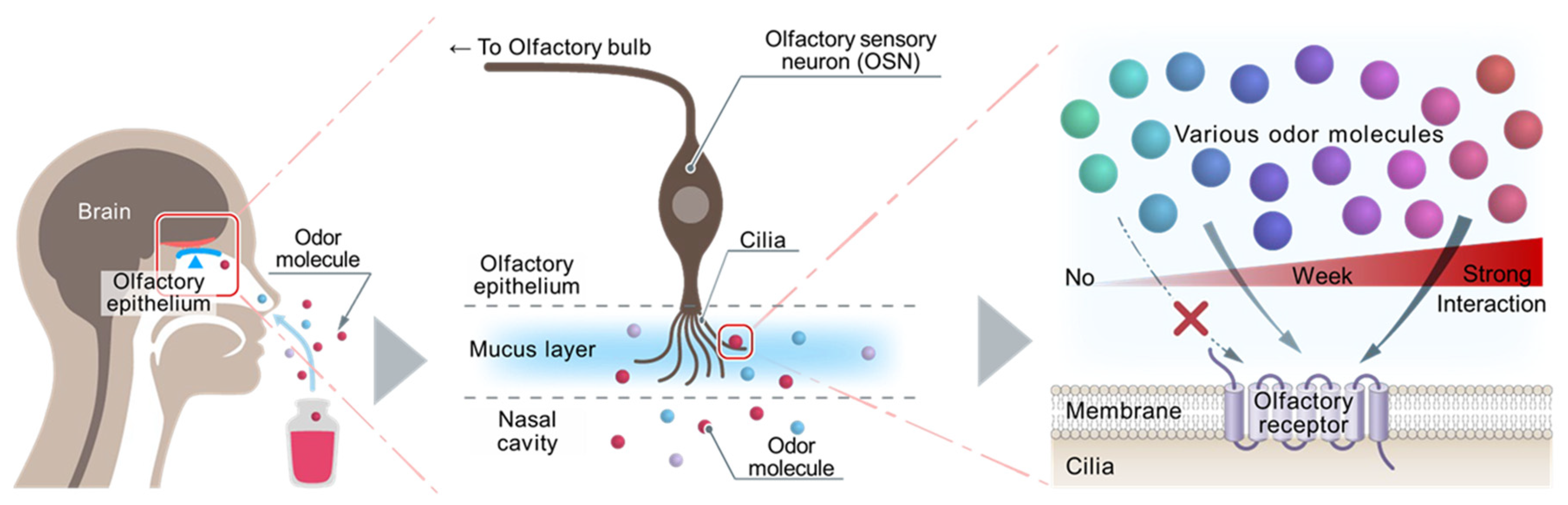
Figure 3.
Endpoint measurement of the binding affinity of odor molecule to OR expressed on heterologous cells. To promote OR expression on the surface of heterologous cells, a signal peptide (Rho-tag) is added to the N-terminus and the OR chaperones RTPs (RTP1, RTP1S, RTP2) and REEP1 and Gα protein chaperone Ric-8B are coexpressed. Binding of odor molecule causes OR to activate adenylate cyclase (AC) via trimeric G proteins to produce cAMP. The amount of cAMP is assayed by endpoint measurement of chemiluminescence using GloSensorTM and is determined as the binding affinity between the odor molecules and OR.
Figure 3.
Endpoint measurement of the binding affinity of odor molecule to OR expressed on heterologous cells. To promote OR expression on the surface of heterologous cells, a signal peptide (Rho-tag) is added to the N-terminus and the OR chaperones RTPs (RTP1, RTP1S, RTP2) and REEP1 and Gα protein chaperone Ric-8B are coexpressed. Binding of odor molecule causes OR to activate adenylate cyclase (AC) via trimeric G proteins to produce cAMP. The amount of cAMP is assayed by endpoint measurement of chemiluminescence using GloSensorTM and is determined as the binding affinity between the odor molecules and OR.
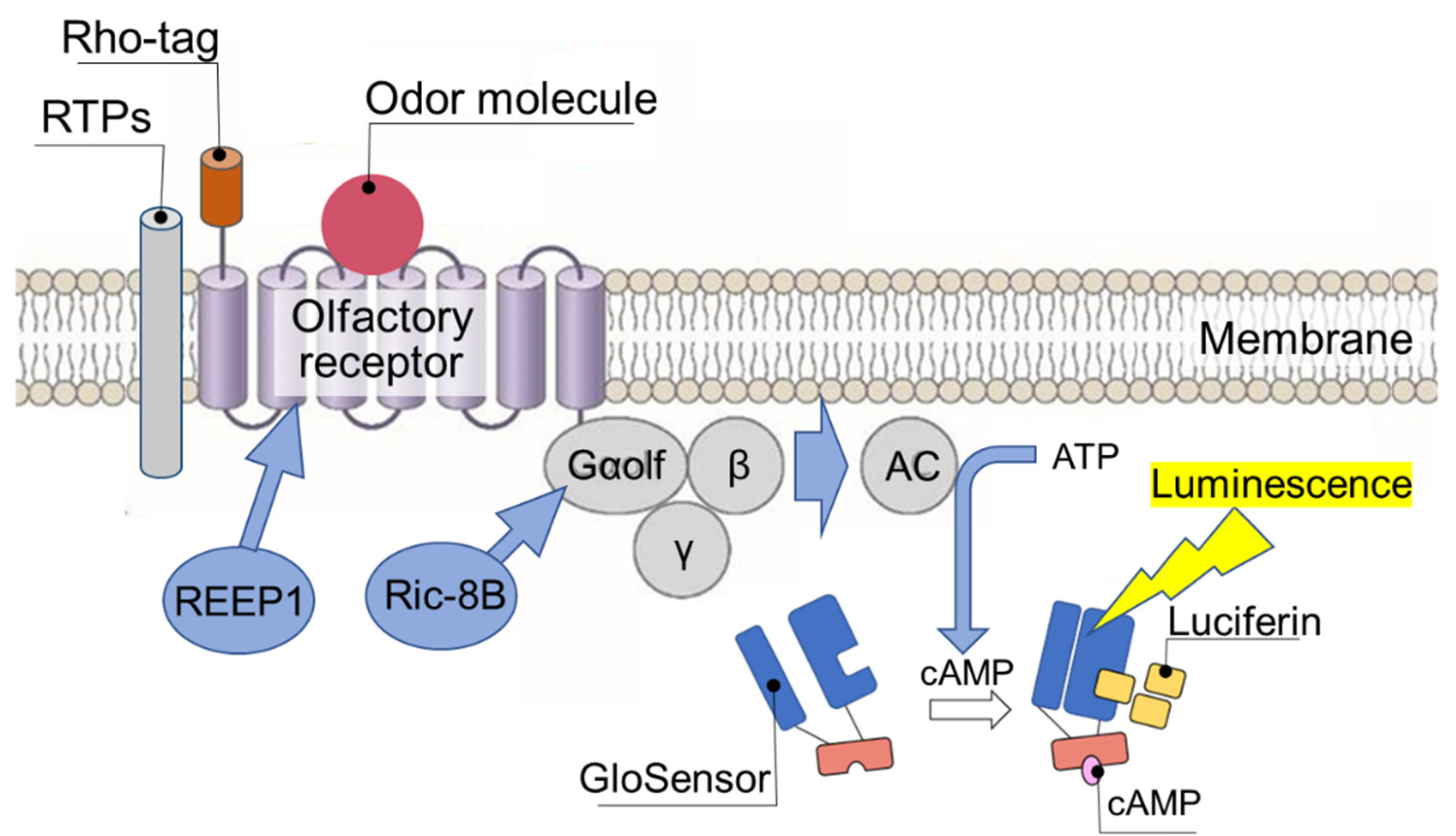
Figure 4.
Real-time measurement of the binding affinity of odor molecule to OR expressed on heterologous cells. Binding of odor molecule causes OR to activate adenylate cyclase (AC) via trimeric G proteins to produce cAMP, which activates and opens cyclic nucleotide-activated channel (CNG) to promote intracellular influx of Na+ ion and Ca2+ ion. Real-time changes in the amount of intracellular Ca2+ influx is measured by the fluorescence intensity change of Ca2+-dependent fluorescent protein (GCaMP) and are determined as real-time changes in the binding affinity between odor molecules and OR.
Figure 4.
Real-time measurement of the binding affinity of odor molecule to OR expressed on heterologous cells. Binding of odor molecule causes OR to activate adenylate cyclase (AC) via trimeric G proteins to produce cAMP, which activates and opens cyclic nucleotide-activated channel (CNG) to promote intracellular influx of Na+ ion and Ca2+ ion. Real-time changes in the amount of intracellular Ca2+ influx is measured by the fluorescence intensity change of Ca2+-dependent fluorescent protein (GCaMP) and are determined as real-time changes in the binding affinity between odor molecules and OR.
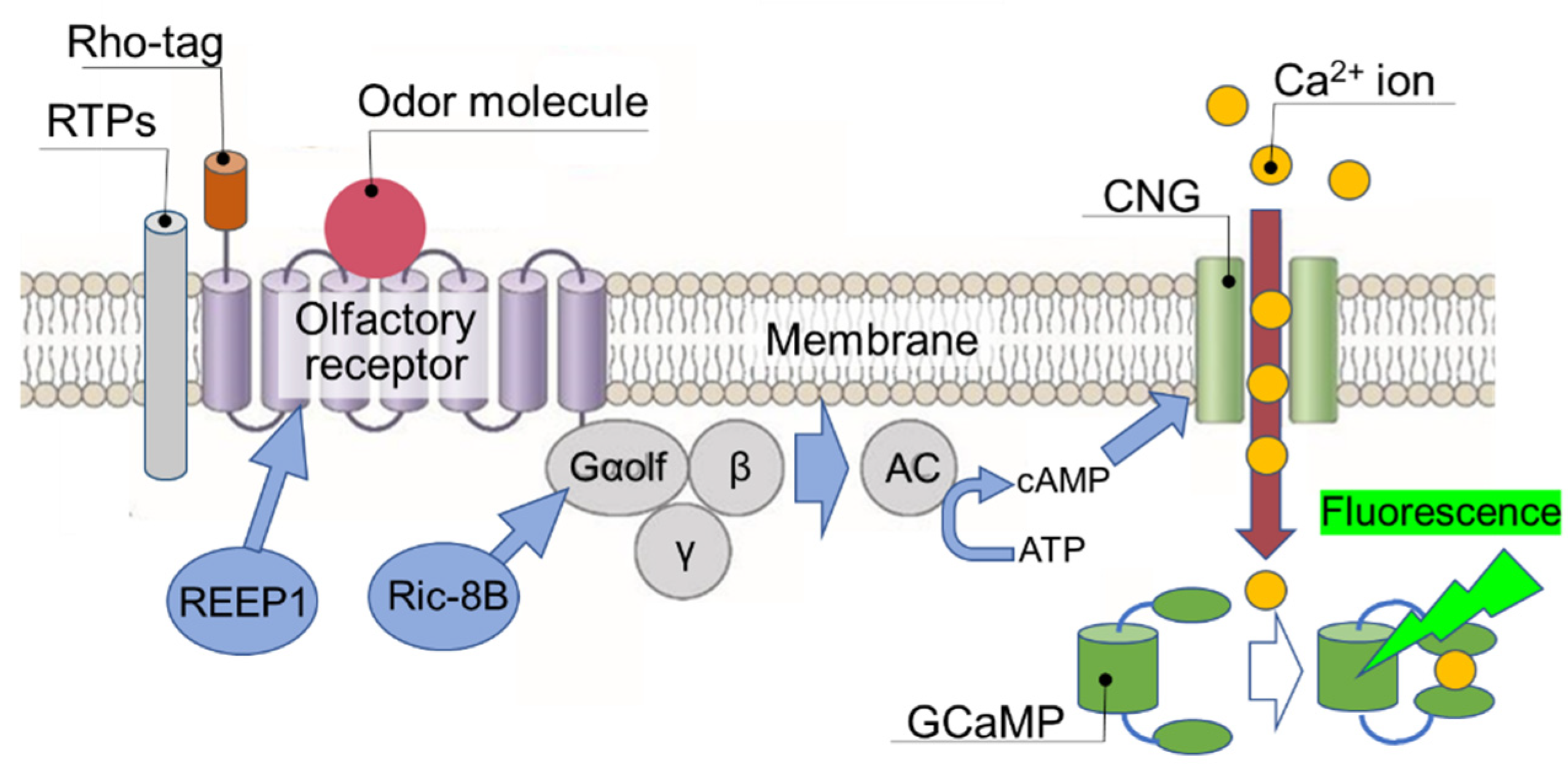
Figure 5.
Workflow from odor measurement with human OR sensor, single cell-based analysis of fluorescence intensity changes, and odor matrix generation. Note that these odor matrixes also include changes over time for each OR, for easier understanding, a snapshot of the time when the fluorescence intensity reached its maximum is usually used.
Figure 5.
Workflow from odor measurement with human OR sensor, single cell-based analysis of fluorescence intensity changes, and odor matrix generation. Note that these odor matrixes also include changes over time for each OR, for easier understanding, a snapshot of the time when the fluorescence intensity reached its maximum is usually used.
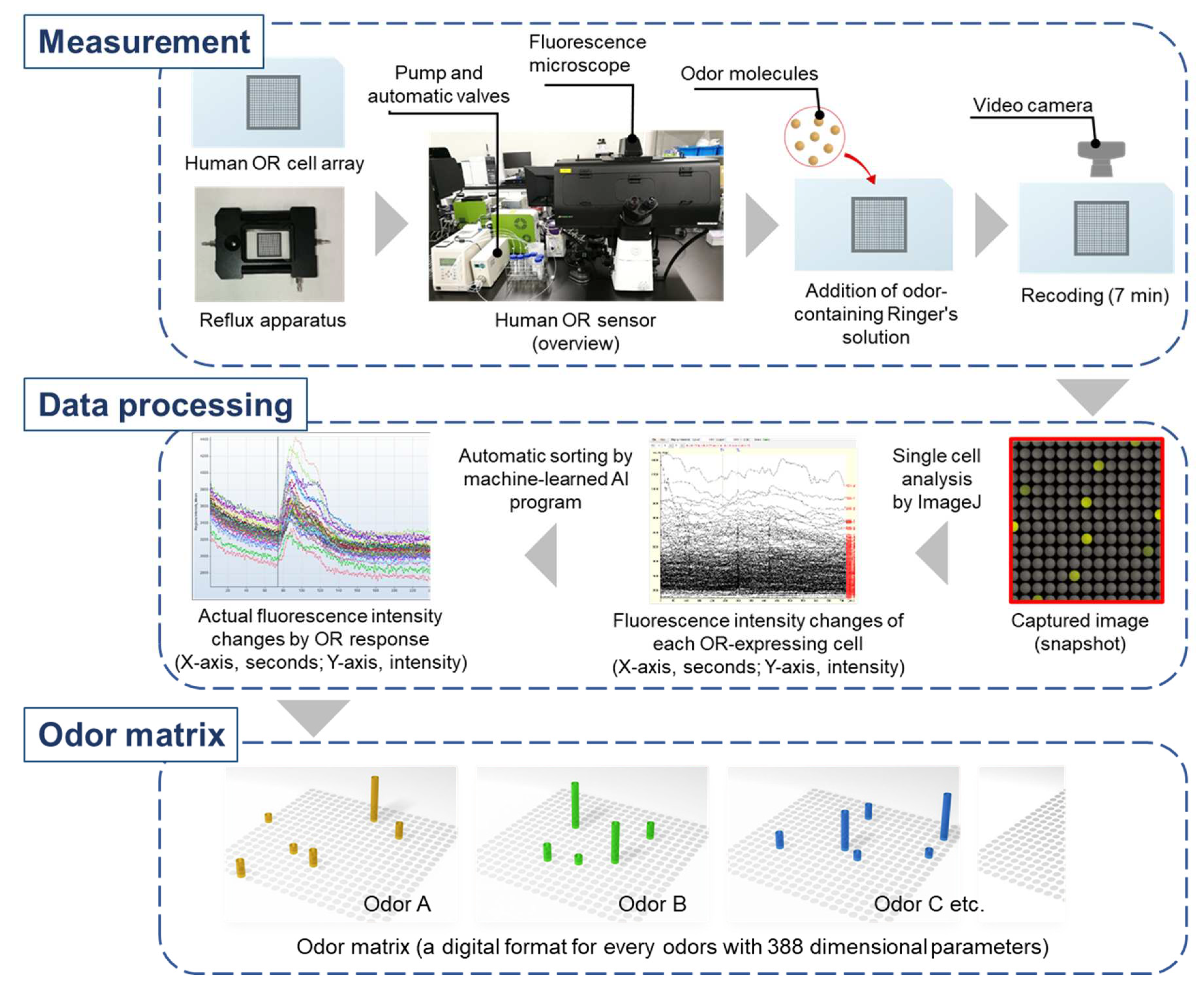
Figure 6.
Conceptual diagram of odor reconstitution. First, a lot of basically safe odor molecules (e.g., natural flavors and fragrances, synthetic fragrances, food additives) are measured by human OR sensor to build an “odor matrix database.” Incorporate odor molecules that pinpointly stimulate limited number of ORs in the database, exclude odor molecules showing the exceptional activities described in section 6 from database, and generate an “odor molecule library for reconstitution.” The minimum group of odor molecules necessary to reproduce this target odor matrix (including changes in each human OR response over time) is selected from the library.
Figure 6.
Conceptual diagram of odor reconstitution. First, a lot of basically safe odor molecules (e.g., natural flavors and fragrances, synthetic fragrances, food additives) are measured by human OR sensor to build an “odor matrix database.” Incorporate odor molecules that pinpointly stimulate limited number of ORs in the database, exclude odor molecules showing the exceptional activities described in section 6 from database, and generate an “odor molecule library for reconstitution.” The minimum group of odor molecules necessary to reproduce this target odor matrix (including changes in each human OR response over time) is selected from the library.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



