
Submitted:
25 May 2023
Posted:
26 May 2023
You are already at the latest version
Abstract
The calmodulin-binding transcription activators (CAMTAs) mediate transcriptional regulation of development, growth, and responses to various environmental stresses in plants. To characterize the biological processes of soybean CAMTA (GmCAMTA) family members in response to abiotic stress, we identified 15 GmCAMTA genes from soybean (Glycine max L.) and investigated their roles in the abiotic stress response. The transcriptions of GmCAMTAs exhibited distinct circadian regulation patterns and were expressed differently in response to salt, drought, and cold stresses, except ABA. Interestingly, the expression levels of GmCAMTA2, GmCAMTA8, and GmCAMTA12 were higher in stem tissue than in other soybean tissues. To determine the roles of GmCAMTAs in the regulation of developmental processes and stress responses, we isolated GmCAMTA2 and GmCAMTA8 cDNAs from soybean and generated Arabidopsis overexpressing transgenic plants. The GmCAMTA2-OX and GmCAMTA8-OX plants showed hypersensitivity to drought stress. The water in the leaves of GmCAMTA2-OX and GmCAMTA8-OX plants was lost faster than that in WT plants under drought stress conditions. In addition, stress-responsive genes in the GmCAMTA2-OX and GmCAMTA8-OX plants were down-regulated under drought stress conditions. Our results suggest that GmCAMTA2 and GmCAMTA8 genes are regulated by circadian rhythms and function as negative regulators in development and drought stress responses.
Keywords:
GmCAMTA
; circadian rhythm
; development
; drought stress
1. Introduction
In plants, the calcium (Ca2+) ion is an important element for maintaining the intracellular ion homeostasis of plant cells, as well as for regulating structural processes, metabolism, and signaling [1,2]. The major function of the Ca2+ ion is to enhance the plant’s tolerance to stabilize cell walls and membranes against various plant stresses [2,3,4]. The Ca2+ ion regulates the biochemical reactions in the plant cell by controlling the activity of ion-channel-related proteins, such as cyclic nucleotide-gated channels (CNGCs), ionotropic glutamate receptors, two-pore channel 1 (TPC1), annexins, and mechanosensitive channels [2]. The Ca2+ ion is a versatile secondary messenger in biotic and abiotic stress responses [3,4]. Increased Ca2+ concentrations through various abiotic stresses are recognized, amplified, and transmitted through Ca2+-binding proteins [4]. These Ca2+-binding proteins play a role as Ca2+ sensors, including calmodulin (CaM) and calmodulin-like proteins (CMLs), calcineurin B-like proteins (CBLs), and calcium-dependent protein kinases (CDPKs) [5]. These signal cascades in plants physiologically regulate the division and elongation of cells, stomatal movement, and developmental processes in plant tolerance or adaptation responses to abiotic stresses [5,6].
The CaMs with one or more EF-hand Ca2+-binding motifs are associated with a multitude of biochemical reactions, such as the control of transcriptional expression and enzyme activity [6,7]. Although the Ca2+ ion lacks cellular levels to activate CaM under natural conditions, when exposed to abiotic stresses, the Ca2+ levels are rapidly increased, generating the Ca2+ signals, and there is a boost in the activity of CaM [8]. CaM is the best characterized of the Ca2+ receptors, and the Ca2+–CaM complex regulates the ion channels, transporters, transcription factors (TFs), and various enzymes, including protein kinase, phosphatases, and metabolic enzymes [9,10]. The CaM-driven TFs such as calmodulin-binding transcription activators (CAMTAs), DREB, MYB, WRKY, NAC, bZIP, bHLH, and MADS-box play a pivotal role in multiple cellular processes, including plant stress responses and developmental processes [5,10].
A total of 465 CAMTA genes have been identified from 112 plant species, including dicots, monocots, basal angiosperms, gymnosperms, lycophyta, monilophytes, bryophytes, and chlorophytes [10]. Of the total, 302 CAMTAs were obtained from 49 dicots among plant species; in addition, 15 CAMTAs are the best isolated from soybean (Glycine max L.), respectively [10]. CAMTA proteins in soybean have been known to comprise conserved functional domains, including the DNA-binding (CG-1) domain, transcription-associated immunoglobulin (TIG) domain, calmodulin-binding (CaMB) domain, ankyrin (ANK) repeats, and IQ motifs [11]. The CG-1 domain recognizes and binds to specific DNA sequences of the CGCG box for activating gene transcription [12,13]. The ANK repeats and the TIG domain with binding to the non-specific DNA are involved in protein–protein interactions with either homo- or heterodimers [12,14]. A diverse number of IQ motifs in the C-terminal regions of CAMTAs contain amino acid sequences, such as IQXXXRGXXX, as Ca2+-independent calmodulin-binding (CaMB) sites [5,12,15]. The CaMB domain with its α-helix structure is an important Ca2+-dependent CaMB domain for determining characteristics of CAMTAs in plants [12,16].
The expression levels of CAMTAs show significant differences in the extensive developmental stages and plant stress responses [5,10]. In Arabidopsis, the expression of AtCAMTA6 (AT3G16940) shows a high level in mature leaves and siliques compared with that in other tissues [10]. In addition, the expression levels of AtCAMTA2 (AT5G64220) in seeds and young leaves are significantly lower than in other tissues [10]. In tomato (Solanum lycopersicum), PtCAMTA4 (JN566050) shows high levels of transcriptional expression in turning and orange fruit stages, while PtCAMTA2 (JN566047) is highly expressed in all developmental fruit stages except the mature green and breaker stages [16]. In tea (Camellia sinensis) plant, the expression of CsCAMTA1 (TEA025813.1) is at higher levels in old leaves than that in both other tissues and other CsCAMTA genes [17]. In black cottonwood (Populus trichocarpa), PtCAMTA1 (POPTR_0001s13700) is highly expressed in the mature leaves, while PtCAMTA2 (POPTR_0005s07660) and PtCAMTA3 (POPTR_0007s05410) are more weakly expressed in the roots than in different tissues [18]. In rice (Oryza sativa), the expression levels of all seven OsCAMTA genes are displayed more in leaves after flowering compared with those before flowering [10]. The MuCAMTA1 (Musa acuminate; LOC1039776) and phavuCAMTA1 (Phaseolus vulgaris; PHAVU_006G206400g) genes are highly expressed in plant responses under drought stress conditions [19,20]. In the root of maize (Zea mays L.), the expression of ZmCAMTA1 (GRMZM2G171600) is significantly decreased to cold stress, but increases to salt stress and jasmonic acid (JA) treatment [21]. In Medicago truncatula, the expression of MtCAMTA7 (Medtr8g090205) is induced by ABA treatment compared with other MtCAMTA genes [22]. Therefore, the expression of CAMTA genes presents substantial specificity in the developmental stages and tissues, as well as in stress responses.
CAMTAs in plants are known to be positive or negative regulators in plant development processes, as well as in biotic and abiotic stress responses [23,24,25]. AtCAMTA1 (AT5G09410) positively regulates the ABA-dependent response for enhancing plant tolerance to drought stress [23]. atcamta1 mutants showed hypersensitivity to drought stress by regulating stress-responsive genes, such as RD26, ERD7, RAB18, LTPs, and COR78 [23]. AtCAMTA3 (AT2G22300) functions as a negative regulator in plant defense responses against bacteria [26] and fungi [27]. atcamta3 mutants showed enhanced resistance to Pseudomonas syringae pv. tomato DC3000 (PstDC3000) and Botrytis cinerea by the high expression of pathogenesis-related (PR) genes [26,27]. In contrast, AtCAMTA3 acts as a positive regulator in plant tolerance to freezing stress [24]. A mutation of atcamta3 weakened the freezing tolerance caused by reducing the cold-induced accumulation of CBF genes transcript levels during exposure to low temperature [24]. AtCAMTA1 and AtCAMTA5 (AT4G16150) positively regulate the pollen developmental process by increasing the expression of Arabidopsis V-PPase 1 (AVP1) [28]. Transcriptome profiling using a camta6 mutant under salt stress showed that 638 up-regulated and 1,242 down-regulated genes were involved in AtCAMTA6-dependent manners [29]. In rice, OsCBT/OsCAMTA5 (LOC_Os07g30774) plays a negative role in the plant defense response against fungal pathogens by the high expression of PR genes, such as PR1, PR4, PR10, and PBZ [30]. Rice oscbt-1 mutants showed enhanced plant disease resistance against both rice blast fungus (Magnaporthe grisea) and bacteria (Xanthomonas oryzae) [30]. Soybean GmCAMTA12 (Glyma.17G031900) regulated the expression of its regulatory network-related genes with the CGCG/CGTTG motif under drought stress in Arabidopsis and soybean hairy roots [31]. In peach (Prunus persica L. Batsch), PpCAMTA1 (Prupe.1G108700) in atcamta2/3 double mutants suppressed salicylic acid (SA) biosynthesis and the expression of SA-related genes in the plant defense response against PstDC3000 [32]. Thus, CAMTAs in plants play crucial roles as transcription factors in both biotic and abiotic stress responses.
Various environmental changes have many implications for growth, developmental processes, and yield in crop plants. Drought stress is an important restricting factor for crop growth and the developmental tissue processes, holding both expansion and elongation down [33]. Since drought stress triggers low turgor pressure in the cell, the osmotic maintenance of cell turgor in crops is of great importance for plant growth and crop survival [33]. In many crops, soybean is highly sensitive to drought stress responses through various morphological changes [34]. When exposed to drought stress, soybean shows momentous changes in most of the tissue morphology, such as a reduction in new branches and trifoliate leaves per plant, growth inhibition of initiation leaves, and reductions in leaf area via repression of lamina expansion [34]. Drought stress has a negative effect on the developmental processes of stems and leaves by decreasing their elongation and expansion [33].
Soybean (Glycine max L.) is one of the most economical crops with a nutritional balance of protein, fat, and carbohydrate. It is mainly used as a portion in important foods, feeds, and bio-feedstocks [35]. The Williams 82 cultivar is a typical soybean breed that was used to be established the reference genome sequence [36]. The 15 GmCAMTAs were identified in soybean by the SoyBase database (http://soybase.org/), and the analysis of gene expression showed that the expression patterns of GmCAMTAs were altered during abiotic stress (salt, drought, cold, and oxidative) and hormone (ABA, SA, and JA) responses (Supplementary Table S1) [11]. Recently, it was reported that GmCAMTA12 plays an important role in the tolerance responses of Arabidopsis and soybean to drought stress [31]. However, GmCAMTAs still do not have enough study for the functional characterization in plant development processes and stress tolerance responses. In this study, GmCAMTA2 and GmCAMTA8 characterized the substantial specificity and reliability in the developmental stages and abiotic stress responses. The results of our study can be effective for further functional analysis of GmCAMTAs in soybean.
2. Results
2.1. Circadian rhythms affected the expression patterns of GmCAMTAs under long-day conditions
The stress-responsive genes in plants are rhythmically expressed in stress signaling, thereby producing daily cellular, metabolic, and physiological rhythms [37]. To investigate the influence on the transcripts of GmCAMTAs by daily circadian rhythms, the expression patterns of GmCAMTAs under long-day conditions (16h light/8h dark) were measured. Quantitative RT-PCR analyses showed that the expression of 15 GmCAMTAs accumulated with the onset of light and reached maximum access levels of their expression at ZT9 to ZT15 (Figure 1). After the light was turned off (ZT18 to ZT20), the expression levels of most GmCAMTAs significantly decreased more than approximately two-fold under light conditions (Figure 1). However, the expression levels of GmCAMTA3, GmCAMTA5, GmCAMTA6, GmCAMTA7, and GmCAMTA13 at ZT18 and ZT20 were subtly reduced less than two-fold than between ZT9 and ZT15 (Figure 1). In addition, in silico analysis using the Diurnal database tool showed the diurnal expression of GmCAMTA2, GmCAMTA5, GmCAMTA8, GmCAMTA12, and GmCAMTA13 (Supplementary Figure S1). Unfortunately, the diurnal expression changes of other GmCAMTAs were not available in the Diurnal web database. Therefore, the transcripts of GmCAMTAs were regulated according to circadian rhythms in the daily clock.
2.2. Various abiotic stresses affected the expression of GmCAMTAs
Previously, a few studies reported that the expression of GmCAMTAs is regulated by abiotic stresses and hormone treatment in soybean [11,31]. However, previous results had not considered the rhythmical changes of GmCAMTAs expression in abiotic stress responses. To investigate the expression patterns of GmCAMTAs under abiotic stress conditions in view of the rhythmical changes, we performed quantitative RT-PCR (qRT-PCR) analysis. After measuring the relative expression values of genes by GmTUBULIN as an internal control under abiotic stress conditions, we carried out the normalization according to the relative expression values of genes at each time point under natural conditions. When soybean plants were exposed to salt stress, all of the GmCAMTAs were highly expressed in early times, and their expression levels increased gradually as time passed (Figure 2A). The expression levels of most GmCAMTAs in drought stress responses were significantly increased compared to those under natural conditions, except GmCAMTA13 and GmCAMTA15 (Figure 2B). Interestingly, GmCAMTA1 and GmCAMTA2 were highly expressed to cold stress; however, the expression levels of other GmCAMTAs were reduced during cold stress responses (Figure 2C). ABA did not influence the expression of most GmCAMTAs in soybean, although GmCAMTA1, GmCAMTA5, GmCAMTA6, and GmCAMTA13 were weakly expressed under ABA stress (Figure 2D). Thus, even if GmCAMTAs seem invariably to be reactions to most abiotic stresses, the transcripts of GmCAMTAs were influenced differently by the specificity of plant stresses.
2.3. The transcripts of GmCAMTAs had substantial specificity in developmental processes
Plant developmental processes are associated with abiotic stress responses through impressive biochemical, physiological, and morphological changes [38]. We tested the changes in GmCAMTA expression levels in different tissues using qRT-PCR analysis. Many GmCAMTAs were expressed in the roots; however, GmCAMTA2, GmCAMTA7, GmCAMTA9, GmCAMTA14, and GmCAMTA15 were weakly expressed (Figure 3A). In the leaves, the expression of most GmCAMTAs was highly induced, except for that of GmCAMTA2, GmCAMTA8, and GmCAMTA12 (Figure 3B). Interestingly, GmCAMTA2, GmCAMTA8, and GmCAMTA12 were relatively highly expressed in the stem tissues as compared to other GmCAMTAs (Figure 3C). Unlike the expression patterns of GmCAMTAs in roots, leaves, and stem tissues, the expression levels of GmCAMTAs were similar in the flower (Figure 3D). Our results indicated that GmCAMTAs were selectively expressed in each of the plant tissues, thereby showing substantial specificity in the developmental stages.
2.4. Overexpression of GmCAMTA2 and GmCAMTA8 reduced drought tolerance in Arabidopsis
GmCAMTA2, GmCAMTA8, and GmCAMTA12 were highly expressed in stem (Figure 3). The overexpression of GmCAMTA12 in Arabidopsis and soybean enhances plant tolerance to drought stress [31]. To investigate the functions of GmCAMTA2 and GmCAMTA8 in drought stress responses, we generated Arabidopsis transgenic plants with overexpressed GmCAMTA2 and GmCAMTA8 under the 35S promoter. To establish the expression levels of GmCAMTA2 and GmCAMTA8 in Arabidopsis transgenic plants, we performed RT-PCR analysis. According to the results of RT-PCR, we selected three individual lines, each with different expression levels of GmCAMTA2 (GmCAMTA2-OX #1, #4, and #8 lines; Supplementary Figure S2A) and GmCAMTA8 (GmCAMTA8-OX #5, #8, and #9 lines; Supplementary Figure S2B). We examined the plant tolerance by overexpressing GmCAMTA2 and GmCAMTA8 during exposure to drought stress (Figure 4). In natural conditions, GmCAMTA2-OX and GmCAMTA8-OX plants were similar in growth and development compared with WT and empty vector-overexpressing (Vector-OX) Arabidopsis (Figure 4A). GmCAMTA2-OX and GmCAMTA8-OX plants showed hypersensitivity compared with WT and Vector-OX under drought stress and re-watering conditions (Figure 4A). The survival rates after re-watering under drought stress conditions were lower in the GmCAMTA2-OX (approximately 2.78 to 13.89 %) and GmCAMTA8-OX (approximately 2.78 to 11.11 %) than the WT (approximately 72.22 to 79.17 %) and Vector-OX (approximately 70.83 to 80.56 %) (Figure 4B). To confirm the drought phenotypes in the GmCAMTA2-OX and GmCAMTA8-OX plants, we measured the water loss from detached leaves during exposure to drought stress. The water loss of the GmCAMTA2-OX and GmCAMTA8-OX plants was significantly higher than that of the WT and Vector-OX plants in drought stress responses (Figure 4C). These results suggested that the overexpression of GmCAMTA2 and GmCAMTA8 reduced plant tolerance to drought stress.
2.5. Overexpression of GmCAMTA2 and GmCAMTA8 regulated the transcripts of stress-responsive genes in drought stress responses
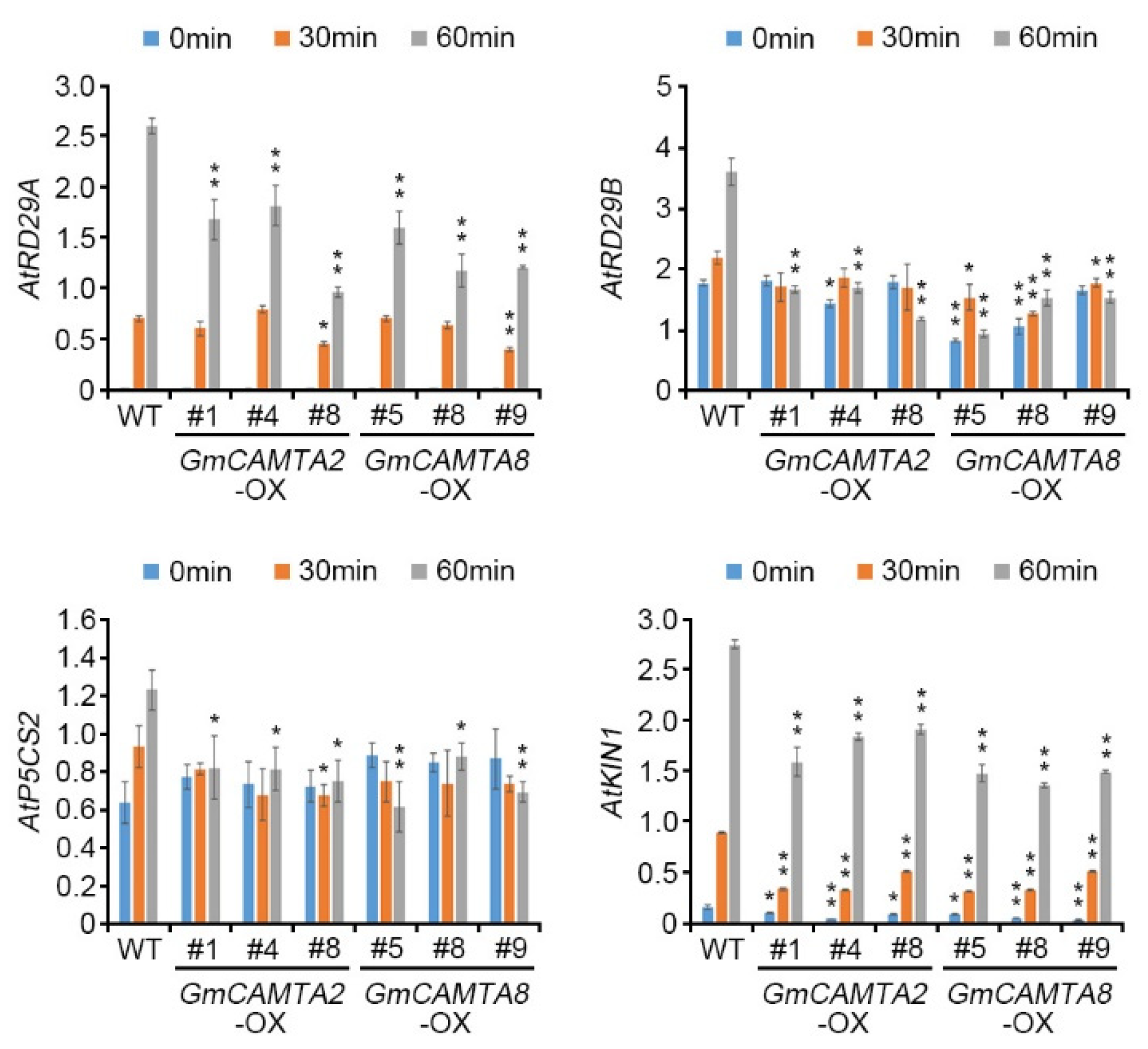
To investigate whether GmCAMTA2 and GmCAMTA8 affected the expression patterns of stress-responsive genes during drought stress responses, we performed qRT-PCR analysis of various stress-responsive genes (Figure 5). The expression levels of ABA-induced genes AtRD29A and AtRD29B were significantly decreased in GmCAMTA2-OX (AtRD29A; approximately 0.97- to 1.67-fold, AtRD29B; approximately 1.18- to 1.69-fold) and GmCAMTA8-OX (AtRD29A; approximately 1.18- to 1.61-fold, AtRD29B; approximately 0.94- to 1.53-fold) compared with those of WT plants (AtRD29A; approximately 2.61-fold, AtRD29B; approximately 3.60-fold) at the 60 min time point during drought stress. The Pyrroline-5-Carboxylate Synthase 2 (P5CS2) gene was regulated in the biosynthesis of proline, which is an antioxidant in water stress response [39]. The expression of AtP5CS2 was weakly decreased in GmCAMTA2-OX (approximately 0.75- to 0.82-fold) and GmCAMTA8-OX (approximately 0.62- to 0.88-fold) compared with that of WT plants (approximately 1.23-fold) at the 60 min time point during drought stress. In addition, the expression of the stress-related marker gene AtKIN1 was decreased in GmCAMTA2-OX (approximately 1.60- to 1.92-fold) and GmCAMTA8-OX (approximately 1.37- to 1.50-fold) compared with that of WT plants (approximately 2.76-fold) at the 60 min time point during drought stress. These results suggested that the overexpression of GmCAMTA2 and GmCAMTA8 down-regulated the expression of stress-responsive genes during drought stress. Thus, GmCAMTA2 and GmCAMTA8 act as negative regulators in drought stress responses.
3. Discussion
The characterization of various CAMTA genes, as core transcription factors with CaM binding sites, has been reported in recent years by genome-wide analysis and expression profiles [10,11,16,17,18,19,20,21,22,25]. Although 15 GmCAMTA genes in soybean have been identified, the functions of GmCAMTAs remained almost unknown, except for GmCAMTA12 [31]. In this study, we identified 15 GmCAMTA genes and established that GmCAMTA transcripts were controlled by circadian rhythms (Figure 1). Previous studies of functions of CAMTA genes were little known about circadian regulation in crops. Plant cellular responses, such as gene expression, development, and metabolic processes, were circadian regulated by circadian oscillators [40,41]. Circadian responses in plants were attributable to cellular oscillations in the intracellular Ca2+ concentration ([Ca2+]) in the surrounding environment, including photoperiod and light intensity [42,43]. The calmodulin-like 24 (CML24), one of the Ca2+-binding proteins, regulated the circadian period in Ca2+-dependent signaling [41]. The most important properties of CAMTAs in plants have a high correlation with CaM proteins, as a Ca2+ sensor to regulate the gene expression in Ca2+-dependent plant responses [5]. These facts were important to explain that the circadian regulation of GmCAMTAs transcripts was mediated by the cellular oscillation of [Ca2+].
Many CAMTAs in crops have been directly or indirectly associated with various developmental and cellular processes, whether through autonomously regulating a major transcription factor or targeting other transcription factors [5]. The fifteen GmCAMTAs in soybean [11], seven PtCAMTAs in Populus trichocarpa [18], nine ZmCAMTAs in maize [21], seven MtCAMTAs in Medicago truncatula [22], and fifteen VvCAMTAs in Vitis vinifera [44] were endowed with substantial specificity in the developmental stages by highly tissue-specific expression. Root-specific-induced PtCAMTA2 and PtCAMTA3 genes were negatively regulated under cold stress in a short time [18]. In this study, GmCAMTA2, GmCAMTA8, and GmCAMTA12 were especially highly expressed in stem tissues compared to other GmCAMTAs (Figure 3). The growth of plant tissues, such as flowers, stems, and leaves, were greatly affected by abiotic stress [43]. The specificity in tissues is an important characteristic for investigating the functions of GmCMATAs in diverse molecular processes against environmental stresses. Since plants are constantly exposed to environmental stimuli during their lifetime, they have developed various adaptive mechanisms between the developmental process and stress responses [44,45].
Drought stress is an important restricting factor for crop growth and developmental tissue processes, holding both expansion and elongation down [46]. Since drought stress triggers low turgor pressure in the cell, the osmotic maintenance of cell turgor in crops is of great importance for plant growth and crop survival [46]. In many crops, soybean is highly sensitive to drought stress responses through various morphological changes [47]. When exposed to drought stress, soybean showed momentous changes in most of the tissue morphology, such as a reduction in new branches and trifoliate leaves per plant, growth inhibition of initiation leaves, and reduction in leaf area through repression of lamina expansion [47]. Drought stress has a negative effect on the developmental processes of stems and leaves by decreasing their elongation and expansion [46]. In particular, drought stress often leads to the expression of common morphogenesis- and stress-responsive genes through molecular mechanisms, including reactive oxygen species (ROS)- and phytohormone-dependent signaling [45]. ATP-dependent metalloprotease 4 (AtFtsH4) played a tissue-specific role in maintaining the stem cell activity of shoot apical meristem (SAM) stages by ROS accumulation under high-temperature conditions [48]. MscS-like 2 (MSL2) and MSL3 were mediated to cell proliferation by regulation of WUSCHEL (WUS) gene expression in osmotic stress responses [49]. The mutation of NADPH genes showed unstable shoot development through decreasing the stem cell population [50]. In case of CAMTA genes, the relationships between development and stress responses have been studied more in fruit species than in crops. The tomato SISR2 and SISR3L transcripts were affected by ethylene signaling during fruit development and ripening [16]. The expression of peach PpCAMTA1 was induced in cold stress, but PpCAMTA3 was repressed with UV-B irradiation during fruit development [32]. Our results indicated that GmCAMTA2, GmCAMTA8, and GmCAMTA12 were significantly expressed in both stem tissues and drought stress responses (Figure 2 and 3). When GmCAMTA2 and GmCAMTA8 were overexpressed in Arabidopsis, GmCAMTA2-OX and GmCAMTA8-OX displayed the sensitive phenotype under drought stress conditions by losing faster water than the WT plant (Figure 4). These results suggest that GmCAMTA2 and GmCAMTA8 act as negative regulators in drought stress responses. Considering the tolerant phenotype produced by GmCAMTA12 overexpression [31] and sensitive phenotypes by GmCAMTA2 and GmCAMTA8 overexpression (Figure 4) under drought stress conditions, GmCAMTA2, GmCAMTA8, and GmCAMTA12 may be associated with drought tolerance by different regulating mechanisms. Our results suggest that GmCAMTA2 and GmCAMTA8 play an important role in both the development process and drought stress responses by circadian regulation.
4. Materials and Methods
4.1. Plant materials and growth conditions
We used the soybean (Glycine max cv. Williams 82) and Arabidopsis (Arabidopsis thaliana Col-0 ecotype) plants in this study. The soybean plants were cultured in a growth chamber at 28 °C under long-day conditions (16h Light / 8h Dark) and Arabidopsis plants in a growth chamber at 22 °C under long-day conditions (16h Light / 8h Dark). To test the expression levels of GmCAMTAs, the two-week-old soybean plants were hydroponically cultured in the 1/2 MS medium with circadian rhythms. For abiotic stress treatments, the two-week-old soybean plants were transferred uniformly in the 1/2 MS medium containing 150mM NaCl (salt stress) and 100 µM ABA. For cold stress treatment, the two-week-old soybean plants in the 1/2 MS medium were placed on ice (approximately 2 °C). In the treatment of drought stress, the two-week-old soybean or 10-day-old Arabidopsis plants after removing the medium were placed in a growth chamber under long-day conditions (16h light / 8h dark). For the expression analysis of GmCAMTAs in different tissues, the soybean plants after sowing seeds were grown in the greenhouse under the seasonal cultivation period.
4.2. Identification of GmCAMTA genes in soybean
The GmCAMTA genes were identified from the soybean genome database at the Phytozome version 13 (https://phytozome-next.jgi.doe.gov/). The specific primers for the qRT-PCR and RT-PCR analyses were designed with the IDT (Integrated DNA Technologies) service website (https://sg.idtdna.com/pages), based on the sequence information of GmCAMTAs (Supplementary Table S2). The full-length coding sequences of GmCAMTA2 (3,309 bp) and GmCAMTA8 (2,736 bp) were amplified by PCR using cDNA from soybean plants, and then cloned into the pMLBart binary vector under the control of the CaMV 35S promoter, which contained a BASTA resistance gene (Supplementary Figure S2). We confirmed the full-length coding sequences through DNA sequencing analysis.
4.3. Generation of GmCMATA-overexpressing Arabidopsis transgenic plants
The pMLBart-GmCAMTA2 and pMLBart-GmCAMTA8 plasmids were transformed into Arabidopsis plants using the Agrobacterium-mediated floral-dipping methods. The GmCAMTA2- and GmCAMTA8-overexpressing transgenic plants were selected on the soil by spraying with 0.2 % BASTA herbicide. The expression levels of GmCAMTA2 and GmCAMTA8 in Arabidopsis transgenic plants were analyzed using reverse transcription PCR (RT-PCR) with gene-specific primers (Supplementary Table S2).
4.4. Physiological assay of drought stress
To test the physiological phenotypes under drought stress conditions, two-week-old plants grown in soil with sufficient water were not watered for 13 days. After re-watering, the recovery of the drought-treated plants was monitored. The assays for drought sensitivity were thrice-replicated experiments using at least 12 plants for each line in each experiment. To measure the transpirational water loss, leaves from 4-week-old plants in soil were detached and placed on Petri dishes. The fresh weights of detached leaves were measured periodically at the indicated times, and the percentages of water loss were calculated. The water loss assays were replicated three times, using at least four plants for each line in each experiment.
4.5. Analysis of gene expression
The total RNA was isolated from each plant sample using the RNeasy Plant Mini Kit (Qiagen, Hilden, Germany), according to the manufacturer’s instructions. To remove any genomic DNA contaminants, purified total RNA was treated with DNaseI (Sigma-Aldrich, St. Louis, MO, USA). For the RT-PCR and quantitative RT-PCR (qRT-PCR) analyses, 1 µg of total RNA was used for cDNA synthesis using the RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, MA, USA), in accordance with the manufacturer’s protocol. The qRT-PCR analysis was performed using the QuantiSpeed SYBR No-ROX Kit (PhileKorea, Seoul, Republic of Korea), and the relative gene expression levels were automatically calculated using the CFX384 real-time PCR detection system (Bio-Rad Laboratories, Hercules, CA, USA). The expression levels of soybean GmTUBULIN and Arabidopsis AtTUBULIN2 were used as the internal control. The qRT-PCR experiments were performed in three independent replicates. The gene-specific primers used are listed in Supplementary Table S2.
5. Conclusions
In summary, we identified 15 GmCAMTA genes in soybean (Glycine max cv. Williams 82) plants. A total of 15 GmCAMTAs were significantly expressed in accordance with circadian rhythms in the daily clock. The expression levels of most GmCAMTAs were induced during various abiotic stresses, including salt, drought, and cold stress, except ABA. Interestingly, GmCAMTA2, GmCAMTA8, and GmCAMTA12 were more highly expressed in stem tissues than other GmCAMTAs. The overexpression of GmCAMTA2 and GmCAMTA8 in Arabidopsis suppressed plant tolerance under drought stress conditions. The expression levels of stress-responsive genes were dramatically reduced by overexpressing GmCAMTA2 and GmCAMTA8 in drought stress responses. These results suggest that GmCAMTAs function in tissue-specific and plant stress responses through rhythmic regulation under drought stress conditions.
Supplementary Materials
Supplementary Figure S1. Diurnal expression of GmCAMTAs in the Diurnal web database. Supplementary Figure S2. RT-PCR analysis in Arabidopsis transgenic plants overexpressing GmCAMTA2 and GmCAMTA8. Supplementary Table S1. Lists of 15 GmCAMTA genes in Phytozome database. Supplementary Table S2. Lists of primers in this study.
Author Contributions
D.B., H.M.C., Y.J.C., H.J.C. and M.C.K. designed and performed the experiments, analyzed data, and wrote the manuscript. B.J.J., S.H.L. and M.S.P. performed some experiments. M.C.K. discussed and commented on the results and revised the manuscript. All authors approved the final manuscript.
Funding
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2015R1A6A1A03031413 (M.C.K.), 2020R1I1A1A01067256 (H.J.C.), 2020R1A6A1A03044344 (D.B.) and 2022R1I1A1A01072247 (D.B.).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Demidchik, V.; Maathuis, F.J.M. Physiological roles of nonselective cation channels in plants: From salt stress to signalling and development. New Phytol. 2007, 175, 387–404. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Shabala, S.; Isayenkov, S.; Cuin, T.A.; Pottosin, I. Calcium transport across plant membranes: Mechanisms and functions. New Phytol. 2018, 220, 49–69. [Google Scholar] [CrossRef] [PubMed]
- Aldon, D.; Mbengue, M.; Mazars, C.; Galaud, J.P. Calcium Signalling in Plant Biotic Interactions. Int. J. Mol. Sci. 2018, 19, 665. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Y.; Jin, L.; Peng, R. Crosstalk between Ca2+ and Other Regulators Assists Plants in Responding to Abiotic Stress. Plants 2022, 11, 1351. [Google Scholar] [CrossRef]
- Iqbal, Z.; Shariq Iqbal, M.; Singh, S.P.; Buaboocha, T. Ca2+/Calmodulin Complex Triggers CAMTA Transcriptional Machinery Under Stress in Plants: Signaling Cascade and Molecular Regulation. Front. Plant Sci. 2020, 11, 598327. [Google Scholar] [CrossRef]
- Kudla, J.; Becker, D.; Grill, E.; Hedrich, R.; Hippler, M.; Kummer, U.; Parniske, M.; Romeis, T.; Schumacher, K. Advances and current challenges in calcium signaling. New Phytol. 2018, 218, 414–431. [Google Scholar] [CrossRef]
- Day, I.S.; Reddy, V.S.; Shad Ali, G.; Reddy, A.S. Analysis of EF-hand-containing proteins in Arabidopsis. Genome Biol. 2002, 3, RESEARCH0056. [Google Scholar] [CrossRef]
- Sun, Q.P.; Guo, Y.; Sun, Y.; Sun, D.Y.; Wang, X.J. Influx of extracellular Ca2+ involved in jasmonic-acid-induced elevation of [Ca2+]cyt and JR1 expression in Arabidopsis thaliana. J Plant Res. 2006, 119, 343–350. [Google Scholar] [CrossRef]
- Poovaiah, B.W.; Du, L. Calcium signaling: Decoding mechanism of calcium signatures. New Phytol. 2018, 217, 1394–1396. [Google Scholar] [CrossRef]
- Xiao, P.; Feng, J.W.; Zhu, X.T.; Gao, J. Evolution Analyses of CAMTA Transcription Factor in Plants and Its Enhancing Effect on Cold-tolerance. Front Plant Sci. 2021, 12, 758187. [Google Scholar] [CrossRef]
- Wang, G.; Zeng, H.; Hu, X.; Zhu, Y.; Chen, Y.; Shen, C.; Wang, H.; Poovaiah, B.W.; Du, L. Identification and expression analyses of calmodulin-binding transcription activator genes in soybean. Plant Soil. 2015, 386, 205–221. [Google Scholar] [CrossRef]
- Bouché, N.; Scharlat, A.; Snedden, W.; Bouchez, D.; Fromm, H. A novel family of calmodulin-binding transcription activators in multicellular organisms. J Biol Chem. 2002, 277, 21851–21861. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Han, J.; Reddig, K.; Li, H.S. A potential dimerization region of dCAMTA is critical for termination of fly visual response. J. Biol. Chem. 2007, 282, 21253–21258. [Google Scholar] [CrossRef] [PubMed]
- Finkler, A.; Ashery-Padan, R.; Fromm, H. CAMTAs: Calmodulin-binding transcription activators from plants to human. FEBS Lett. 2007, 581, 3893–3898. [Google Scholar] [CrossRef]
- Rhoads, A.R.; Friedberg, F. Sequence motifs for calmodulin recognition. FASEB J. 1997, 11, 331–340. [Google Scholar] [CrossRef]
- Yang, T.; Peng, H.; Whitaker, B.D.; Conway, W.S. Characterization of a calcium/calmodulin-regulated SR/CAMTA gene family during tomato fruit development and ripening. BMC Plant Biol. 2012, 12, 19. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhao, M.; Xing, F.; Mao, G.; Wang, Y.; Dai, Y.; Niu, M.; Yuan, H. Identification and Expression Analysis of CAMTA Genes in Tea Plant Reveal Their Complex Regulatory Role in Stress Responses. Front Plant Sci. 2022, 13, 910768. [Google Scholar] [CrossRef]
- Wei, M.; Xu, X.; Li, C. Identification and expression of CAMTA genes in Populus trichocarpa under biotic and abiotic stress. Sci. Rep. 2017, 7, 17910. [Google Scholar] [CrossRef]
- Meer, L.; Mumtaz, S.; Labbo, A.M.; Khan, M.J.; Sadiq, I. Genome-wide identification and expression analysis of calmodulin-binding transcription activator genes in banana under drought stress. Sci. Hortic. 2019, 244, 10–14. [Google Scholar] [CrossRef]
- Saeidi, K.; Zare, N.; Baghizadeh, A.; Asghari-Zakaria, R. Phaseolus vulgaris genome possesses CAMTA genes, and phavuCAMTA1 contributes to the drought tolerance. J Genet. 2019, 98, 31. [Google Scholar] [CrossRef]
- Yue, R.; Lu, C.; Sun, T.; Peng, T.; Han, X.; Qi, J.; Yan, S.; Tie, S. Identification and expression profiling analysis of calmodulin-binding transcription activator genes in maize (Zea mays L.) under abiotic and biotic stresses. Front. Plant Sci. 2015, 6, 576. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sun, T.; Xu, L.; Pi, E.; Wang, S.; Wang, H.; Shen, C. Genome-wide identification of CAMTA gene family members in Medicago truncatula and their expression during root nodule symbiosis and hormone treatments. Front. Plant Sci. 2015, 6, 459. [Google Scholar] [CrossRef] [PubMed]
- Pandey, N.; Ranjan, A.; Pant, P.; Tripathi, R.K.; Ateek, F.; Pandey, H.P.; Patre, U.V.; Sawant, S.V. CAMTA 1 regulates drought responses in Arabidopsis thaliana. BMC Genom. 2013, 14, 216. [Google Scholar] [CrossRef] [PubMed]
- Doherty, C.J.; Van Buskirk, H.A.; Myers, S.J.; Thomashow, M.F. Roles for Arabidopsis CAMTA transcription factors in cold-regulated gene expression and freezing tolerance. Plant Cell. 2009, 21, 972–984. [Google Scholar] [CrossRef] [PubMed]
- Noman, M.; Aysha, J.; Ketehouli, T.; Yang, J.; Du, L.; Wang, F.; Li, H. Calmodulin binding transcription activators: An interplay between calcium signalling and plant stress tolerance. J Plant Physiol. 2021, 256, 153327. [Google Scholar] [CrossRef]
- Du, L.; Ali, G.S.; Simons, K.A.; Hou, J.; Yang, T.; Reddy, A.S.; Poovaiah, B.W. Ca2+/calmodulin regulates salicylic-acid-mediated plant immunity. Nature 2009, 457, 1154–1158. [Google Scholar] [CrossRef]
- Galon, Y.; Nave, R.; Boyce, J.M.; Nachmias, D.; Knight, M.R.; Fromm, H. Calmodulin-binding transcription activator (CAMTA) 3 mediates biotic defense responses in Arabidopsis. FEBS Lett. 2008, 582, 943–948. [Google Scholar] [CrossRef]
- Mitsuda, N.; Isono, T.; Sato, M.H. Arabidopsis CAMTA family proteins enhance V-PPase expression in pollen. Plant Cell Physiol. 2003, 44, 975–981. [Google Scholar] [CrossRef]
- Shkolnik, D.; Finkler, A.; Pasmanik-Chor, M.; Fromm, H. CALMODULIN-BINDING TRANSCRIPTION ACTIVATOR 6: A Key Regulator of Na+ Homeostasis during Germination. Plant Physiol. 2019, 180, 1101–1118. [Google Scholar] [CrossRef]
- Koo, S.C.; Choi, M.S.; Chun, H.J.; Shin, D.B.; Park, B.S.; Kim, Y.H.; Park, H.M.; Seo, H.S.; Song, J.T.; Kang, K.Y.; Yun, D.J.; Chung, W.S.; Cho, M.J.; Kim, M.C. The calmodulin-binding transcription factor OsCBT suppresses defense responses to pathogens in rice. Mol. Cells. 2009, 27, 563–570. [Google Scholar] [CrossRef]
- Noman, M.; Jameel, A.; Qiang, W.D.; Ahmad, N.; Liu, W.C.; Wang, F.W.; Li, H.Y. Overexpression of GmCAMTA12 Enhanced Drought Tolerance in Arabidopsis and Soybean. Int. J. Mol. Sci. 2019, 20, 4849. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, Z.; Cao, X.; Duan, W.; Wei, C.; Zhang, C.; Jiang, D.; Li, M.; Chen, K.; Qiao, Y.; Liu, H.; Zhang, B. Genome-Wide Analysis of Calmodulin Binding Transcription Activator (CAMTA) Gene Family in Peach (Prunus persica L. Batsch) and Ectopic Expression of PpCAMTA1 in Arabidopsis camta2,3 Mutant Restore Plant Development. Int. J. Mol. Sci. 2022, 23, 10500. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.; Khare, D. Molecular approaches for genetic improvement of seed quality and characterization of genetic diversity in soybean: A critical review. Biotechnol. Lett. 2016, 38, 1645–1654. [Google Scholar] [CrossRef] [PubMed]
- Haun, W.J.; Hyten, D.L.; Xu, W.W.; Gerhardt, D.J.; Albert, T.J.; Richmond, T.; Jeddeloh, J.A.; Jia, G.; Springer, N.M.; Vance, C.P.; Stupar, R.M. The composition and origins of genomic variation among individuals of the soybean reference cultivar Williams 82. Plant Physiol. 2011, 155, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yuan, L.; Xie, Q. The circadian clock ticks in plant stress responses. Stress Biol. 2022, 2, 15. [Google Scholar] [CrossRef]
- Ma, X.; Su, Z.; Ma, H. Molecular genetic analyses of abiotic stress responses during plant reproductive development. J. Exp. Bot. 2020, 71, 2870–2885. [Google Scholar] [CrossRef]
- Sabbioni, G.; Funck, D.; Forlani, G. Enzymology and Regulation of δ1-Pyrroline-5-Carboxylate Synthetase 2 From Rice. Front. Plant Sci. 2021, 12, 672702. [Google Scholar] [CrossRef]
- Zhou, L.; Zhou, J.; Xiong, Y.; Liu, C.; Wang, J.; Wang, G.; Cai, Y. Overexpression of a maize plasma membrane intrinsic protein ZmPIP1;1 confers drought and salt tolerance in Arabidopsis. PLoS ONE 2018, 13, e0198639. [Google Scholar] [CrossRef]
- Más, P. Circadian clock function in Arabidopsis thaliana: Time beyond transcription. Trends Cell Biol. 2008, 18, 273–281. [Google Scholar] [CrossRef]
- Martí Ruiz, M.C.; Hubbard, K.E.; Gardner, M.J.; Jung, H.J.; Aubry, S.; Hotta, C.T.; Mohd-Noh, N.I.; Robertson, F.C.; Hearn, T.J.; Tsai, Y.C.; Dodd, A.N.; Hannah, M.; Carré, I.A.; Davies, J.M.; Braam, J.; Webb, A.A.R. Circadian oscillations of cytosolic free calcium regulate the Arabidopsis circadian clock. Nat. Plants 2018, 4, 690–698. [Google Scholar] [CrossRef]
- Love, J.; Dodd, A.N.; Webb, A.A. Circadian and diurnal calcium oscillations encode photoperiodic information in Arabidopsis. Plant Cell 2004, 16, 956–966. [Google Scholar] [CrossRef] [PubMed]
- Tataroglu, O.; Zhao, X.; Busza, A.; Ling, J.; O'Neill, J.S.; Emery, P. Calcium and SOL Protease Mediate Temperature Resetting of Circadian Clocks. Cell 2015, 163, 1214–1224. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Li, Y.; Xie, Q.; Wu, Y. Loss of CDKC;2 increases both cell division and drought tolerance in Arabidopsis thaliana. Plant J. 2017, 91, 816–828. [Google Scholar] [CrossRef]
- Shangguan, L.; Wang, X.; Leng, X.; Liu, D.; Ren, G.; Tao, R.; Zhang, C.; Fang, J. Identification and bioinformatic analysis of signal responsive/calmodulin-binding transcription activators gene models in Vitis vinifera. Mol. Biol. Rep. 2014, 41, 2937–2949. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundararam, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Mangena, P. Water stress: Morphological and anatomic changes in soybean (Glycine max L.) plants. In Plant, Abiotic Stress and Responses to Climate Change, ed V. Andjelkovic; Intech Open: London, UK, 2018; pp. 9–31. [Google Scholar] [CrossRef]
- Lee, H. Stem Cell Maintenance and Abiotic Stress Response in Shoot Apical Meristem for Developmental Plasticity. J. Plant Biol. 2018, 61, 358–365. [Google Scholar] [CrossRef]
- Dolzblasz, A.; Smakowska, E.; Gola, E.M.; Sokołowska, K.; Kicia, M.; Janska, H. The mitochondrial protease AtFTSH4 safeguards Arabidopsis shoot apical meristem function. Sci. Rep. 2016, 6, 28315. [Google Scholar] [CrossRef]
- Wilson, M.E.; Mixdorf, M.; Berg, R.H.; Haswell, E.S. Plastid osmotic stress influences cell differentiation at the plant shoot apex. Development 2016, 143, 3382–3393. [Google Scholar] [CrossRef]
Figure 1.
Transcriptional influence of GmCAMTAs in circadian rhythms. The expression levels of GmCAMTAs were analyzed for circadian regulation using qRT-PCR. The two-week-old seedlings of wild-type (WT) soybeans (Glycine max cv. Williams 82) were grown under long-day conditions (16h light / 8h dark). Zeitgeber time (ZT) means a time of turning on a light. The white and black areas mark the approximate division of light and dark. qRT-PCR analysis was performed with each GmCAMTA-specific primer (Supplementary Table S2). GmTUBULIN was used as an internal control for normalization. Error bars represent the SD of three biological replicates, each with three technical replicates.
Figure 1.
Transcriptional influence of GmCAMTAs in circadian rhythms. The expression levels of GmCAMTAs were analyzed for circadian regulation using qRT-PCR. The two-week-old seedlings of wild-type (WT) soybeans (Glycine max cv. Williams 82) were grown under long-day conditions (16h light / 8h dark). Zeitgeber time (ZT) means a time of turning on a light. The white and black areas mark the approximate division of light and dark. qRT-PCR analysis was performed with each GmCAMTA-specific primer (Supplementary Table S2). GmTUBULIN was used as an internal control for normalization. Error bars represent the SD of three biological replicates, each with three technical replicates.
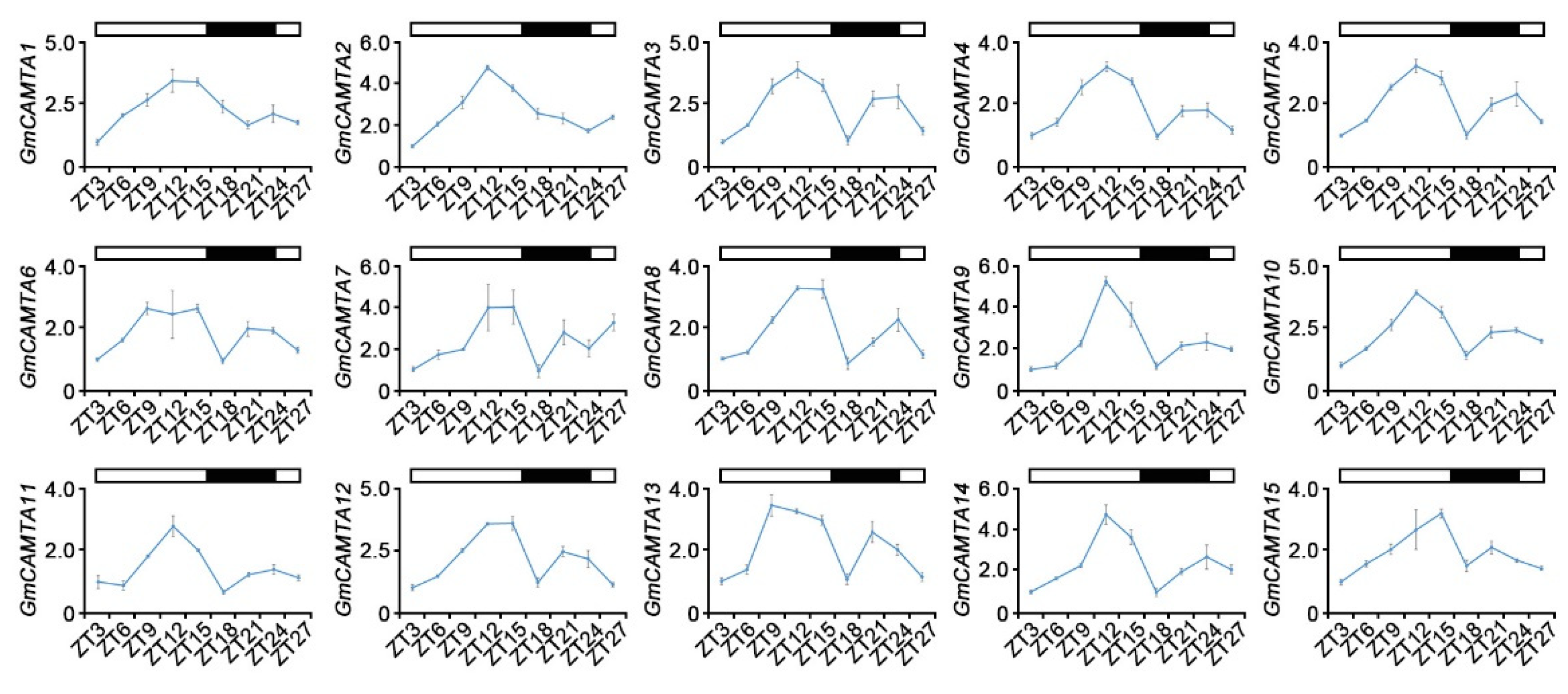
Figure 2.
Effects of various abiotic stresses on the transcriptional expression of GmCAMTAs. The expression levels of GmCAMTAs were analyzed in soybean plants during exposure to various abiotic stresses, using qRT-PCR. The total RNA was extracted from two-week-old plants of WT soybeans under different stresses: (A) salt (150 mM NaCl), (B) drought, (C) cold (approximately 2 °C), and (D) 100 µM ABA. qRT-PCR analysis was performed with each GmCAMTA-specific primer (Supplementary Table S2). GmTUBULIN was used as an internal control, and then the values of the expression levels were calculated by expression values of GmCAMTAs under natural conditions for normalization. Error bars represent the SD of three biological replicates, each with three technical replicates. Asterisks indicate significant differences compared with the expression of the zero indicated time point by Student’s t-test (* p-value < 0.05, ** p-value < 0.01).
Figure 2.
Effects of various abiotic stresses on the transcriptional expression of GmCAMTAs. The expression levels of GmCAMTAs were analyzed in soybean plants during exposure to various abiotic stresses, using qRT-PCR. The total RNA was extracted from two-week-old plants of WT soybeans under different stresses: (A) salt (150 mM NaCl), (B) drought, (C) cold (approximately 2 °C), and (D) 100 µM ABA. qRT-PCR analysis was performed with each GmCAMTA-specific primer (Supplementary Table S2). GmTUBULIN was used as an internal control, and then the values of the expression levels were calculated by expression values of GmCAMTAs under natural conditions for normalization. Error bars represent the SD of three biological replicates, each with three technical replicates. Asterisks indicate significant differences compared with the expression of the zero indicated time point by Student’s t-test (* p-value < 0.05, ** p-value < 0.01).
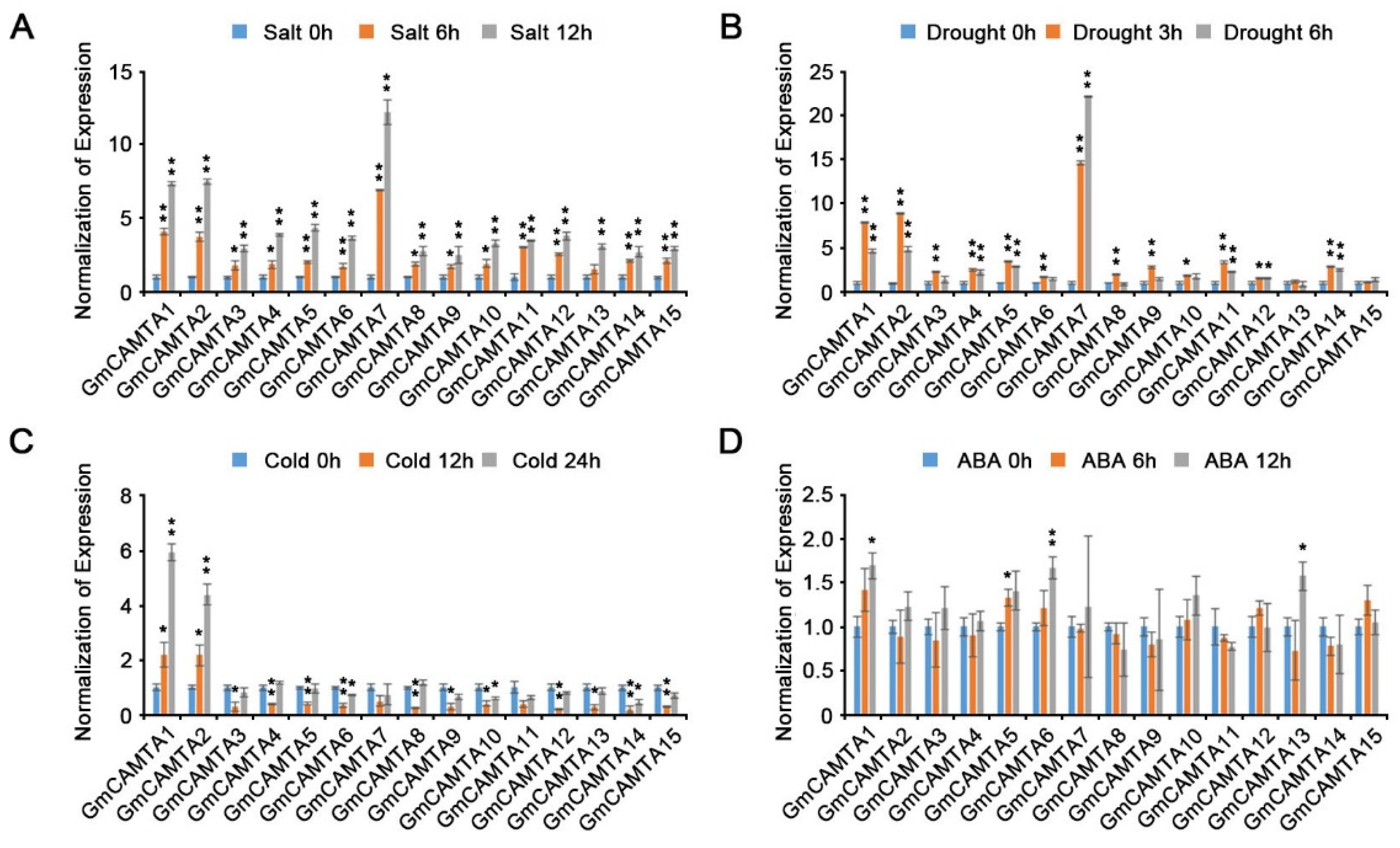
Figure 3.
Transcriptional expression of GmCAMTAs in soybean tissues. The expression levels of GmCAMTAs were analyzed in different tissues of soybean plants using qRT-PCR. The total RNA was extracted from various tissues, including (A) roots, (B) leaves, (C) stem, and (D) flower, at the V4 stages of WT soybeans grown under long-day conditions (16h light / 8h dark). qRT-PCR analysis was performed with each GmCAMTA-specific primer (Supplementary Table S2). GmTUBULIN was used as an internal control for normalization. Error bars represent the SD of three biological replicates, each with three technical replicates.
Figure 3.
Transcriptional expression of GmCAMTAs in soybean tissues. The expression levels of GmCAMTAs were analyzed in different tissues of soybean plants using qRT-PCR. The total RNA was extracted from various tissues, including (A) roots, (B) leaves, (C) stem, and (D) flower, at the V4 stages of WT soybeans grown under long-day conditions (16h light / 8h dark). qRT-PCR analysis was performed with each GmCAMTA-specific primer (Supplementary Table S2). GmTUBULIN was used as an internal control for normalization. Error bars represent the SD of three biological replicates, each with three technical replicates.
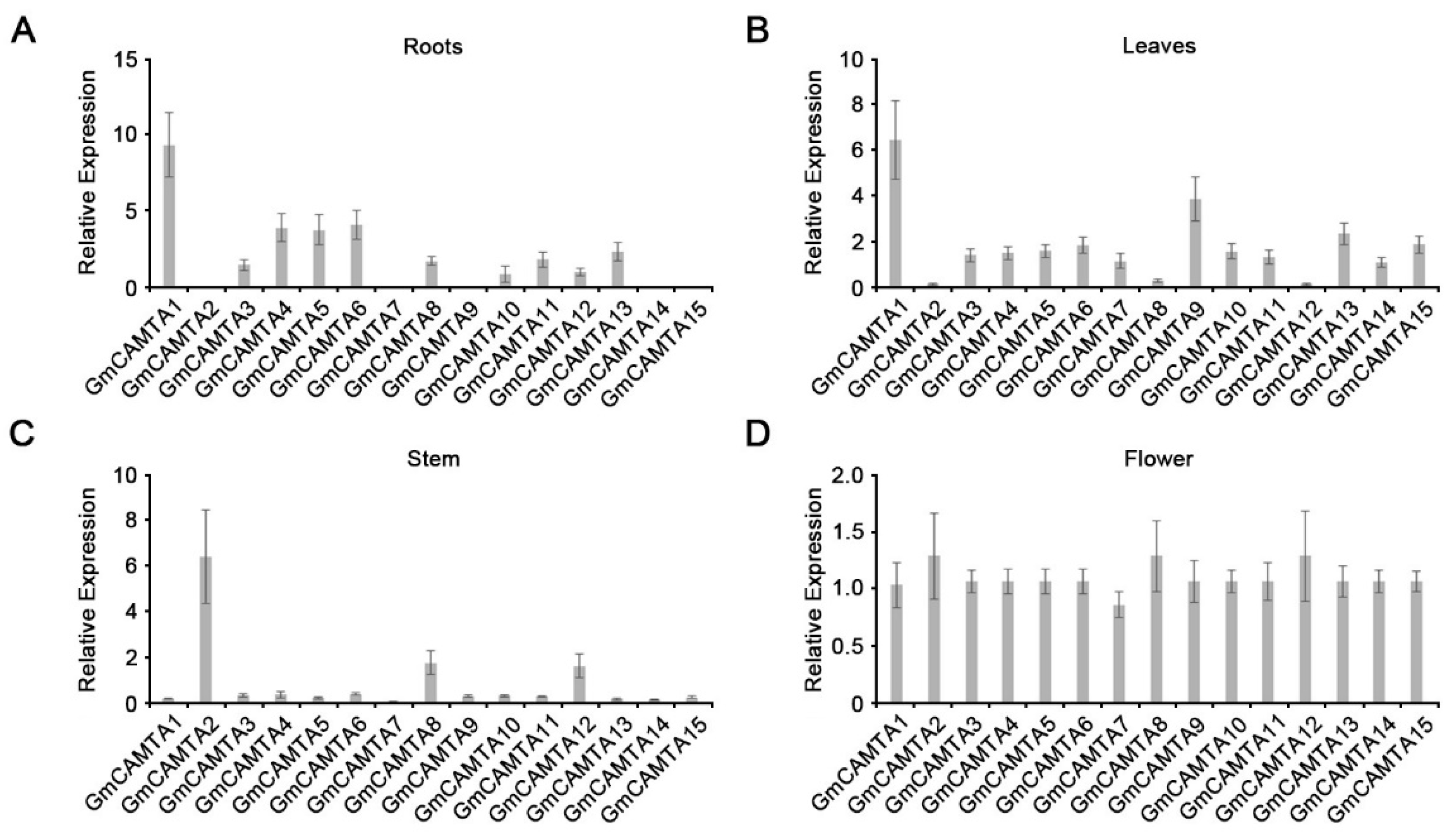
Figure 4.
Effect of drought stress on GmCAMTA2 and GmCAMTA8 overexpressing transgenic Arabidopsis. (A) Phenotypes of GmCAMTA2-OX and GmCAMTA8-OX compared with WT and Vector-OX plants in response to drought stress and re-watering. The plants were grown in soil with sufficient water for two weeks (upper panels), then water was withheld for thirteen days (middle panels). The drought-stressed plants were then re-watered for one day (bottom panels). Each experiment comprised at least 12 plants, and three replicates were performed. (B) Comparison of survival rates (%) among GmCAMTA2-OX, GmCAMTA8-OX, Vector-OX, and WT plants against drought stress. Each experiment comprised at least 12 plants, and error bars represent the SD of three biological replicates. Asterisks indicate significant differences compared with WT plants by Student’s t-test (* p-value < 0.05, ** p-value < 0.01). (C) Analysis of water loss from detached leaves of three-week-old GmCAMTA2-OX, GmCAMTA8-OX, Vector-OX, and WT plants under drought stress conditions. Average values of water loss were measured at the indicated time points. Each experiment comprised at least four plants, and error bars represent the SD of three biological replicates. Asterisks indicate significant differences compared with WT plants by Student’s t-test (* p-value < 0.05, ** p-value < 0.01).
Figure 4.
Effect of drought stress on GmCAMTA2 and GmCAMTA8 overexpressing transgenic Arabidopsis. (A) Phenotypes of GmCAMTA2-OX and GmCAMTA8-OX compared with WT and Vector-OX plants in response to drought stress and re-watering. The plants were grown in soil with sufficient water for two weeks (upper panels), then water was withheld for thirteen days (middle panels). The drought-stressed plants were then re-watered for one day (bottom panels). Each experiment comprised at least 12 plants, and three replicates were performed. (B) Comparison of survival rates (%) among GmCAMTA2-OX, GmCAMTA8-OX, Vector-OX, and WT plants against drought stress. Each experiment comprised at least 12 plants, and error bars represent the SD of three biological replicates. Asterisks indicate significant differences compared with WT plants by Student’s t-test (* p-value < 0.05, ** p-value < 0.01). (C) Analysis of water loss from detached leaves of three-week-old GmCAMTA2-OX, GmCAMTA8-OX, Vector-OX, and WT plants under drought stress conditions. Average values of water loss were measured at the indicated time points. Each experiment comprised at least four plants, and error bars represent the SD of three biological replicates. Asterisks indicate significant differences compared with WT plants by Student’s t-test (* p-value < 0.05, ** p-value < 0.01).
Figure 5.
Effect on the transcripts of Arabidopsis stress-responsive genes by the overexpression of GmCAMTA2 and GmCAMTA8 in drought stress responses. The expression levels of stress-responsive genes were analyzed using qRT-PCR in GmCAMTA2-OX and GmCAMTA8-OX plants under drought stress conditions. Total RNA was extracted from 10-day-old GmCAMTA2-OX and GmCAMTA8-OX Arabidopsis at indicated time points to drought stress. The qRT-PCR analysis was performed with each specific primer of the Arabidopsis stress-responsive genes, AtRD29A, AtRD29B, AtP5CS2, and AtKIN1 (Supplementary Table S2). The expression of Arabidopsis AtTUBULIN2 was used as an internal control for normalization. Error bars represent the SD of two biological replicates, each with three technical replicates. Asterisks indicate significant differences compared with expression levels in WT plants by Student’s t-test (* p-value < 0.05, ** p-value < 0.01).
Figure 5.
Effect on the transcripts of Arabidopsis stress-responsive genes by the overexpression of GmCAMTA2 and GmCAMTA8 in drought stress responses. The expression levels of stress-responsive genes were analyzed using qRT-PCR in GmCAMTA2-OX and GmCAMTA8-OX plants under drought stress conditions. Total RNA was extracted from 10-day-old GmCAMTA2-OX and GmCAMTA8-OX Arabidopsis at indicated time points to drought stress. The qRT-PCR analysis was performed with each specific primer of the Arabidopsis stress-responsive genes, AtRD29A, AtRD29B, AtP5CS2, and AtKIN1 (Supplementary Table S2). The expression of Arabidopsis AtTUBULIN2 was used as an internal control for normalization. Error bars represent the SD of two biological replicates, each with three technical replicates. Asterisks indicate significant differences compared with expression levels in WT plants by Student’s t-test (* p-value < 0.05, ** p-value < 0.01).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



