
Submitted:
25 May 2023
Posted:
26 May 2023
You are already at the latest version
Abstract
Purpureocillium lilacinum PL11 (formerly Paecilomyces lilacinus) is a ubiquitous fungus found in several habitats, mainly in the soil, and belongs to the Ophiocordycipitaceae family. This fungus was shown to have potential applications in agriculture as a biocontrol agent and biofertilizer. P. lilacinum can inhibit plant‒pathogenic fungi and nematodes, including root-knot and potato cyst nematodes. Additionally, P. lilacinum produces siderophores and indole-3-acetic acid (IAA), enhancing plant development and increasing soil nutrient availability. P. lilacinum has been extensively studied in various commodities, including pineapple, maize, soybean, and legume. Most studies have evaluated P. lilacinum as a biocontrol agent, especially for nematode control. Other studies have evaluated the production of secondary metabolites and bioremediation, and few studies have used this fungus as a plant growth promoter. This review addressed important aspects of using this fungus. P. lilacinum is a promising fungus that can be used for agricultural production, reducing environmental impact, and thus collaborating for a sustainable agriculture production system.
Keywords:
Sustainability
; plant production
; fungus
; crop production
Introduction
Utilizing biological inputs to enhance plant growth is highly advantageous in promoting sustainable agriculture [1]. Understanding the advantages of microbial isolate applications on plant growth remains inadequate. Consequently, it is imperative to understand the impact of soil inoculation to establish a comprehensive repository of information that can be utilized to devise sustainable agricultural methodologies [2]. Healthy soil for growing crops is influenced by biological agriculture farming techniques, weather factors, and nutritional and biological parts. One possible way to improve soil nutrients and plant health is using biofertilizers. Biofertilizers can be made of bacteria and fungi, making nutrients available to plants through organic processes [3]. Some fungal isolates change the soil by making nutrients more soluble and helping plants grow. However, when used, little is known about how these isolates affect soil biology and nutrition. The most noticeable effect of fungi that helps plants grow is that roots and shoots gain more weight. These benefits may come from how the fungus makes and controls the production of phytohormones. Fungal treatments can help control root growth factors such as ethylene, indole-3-acetic acid, abscisic acid, salicylic acid, and jasmonic acid [4]. Fungi can also be used for nematode control [5], insect control [6], and the production of secondary metabolites [7], which affect the growth of several phytopathogens, and this fungus can be used to ameliorate soil contamination through the process named bioremediation [8] and can stimulate plant growth due to several mechanisms of action [9]. This review addresses current studies regarding the utilization of the fungi P. lilacinum for these skills.
Purpureocillium lilacinum for biocontrol
Entomopathogenic fungi may reduce nematode and insect populations [10]. These fungi can be used as biocontrol agents, reducing the damage caused by these nematodes, including Meloidogyne incognita, which causes significant damage to several crops [11]. The effectiveness of using these kinds of fungi has been investigated and has been promising for several important agricultural crops worldwide [2,9,10,12] . The use of P. lilacinum has been combined with some chemical nematicides; for instance, a chemical nematicide named Velum and a biological nematicide produced by P. lilacinum combined to regulate the root-knot worm M. incognita in tomato plants have been investigated. Their results showed that combining both nematicides was more successful than using either nematicide separately in decreasing worm numbers and increasing tomato output [13]. The nematicide Velum successfully reduced the worm population at sowing and sustained P. lilacinum biological activity throughout the growing season. The combination of P. lilacinum and the nematicide Velum resulted in the greatest decrease in the second-stage juvenile population, followed closely by Velum alone, which resulted in a 92% reduction in second-stage juveniles. The second-stage juvenile population was decreased by 61% by P. lilacinum. The fungus P lilacinum is able to control M. incognita worms. During a two-year indoor tomato-cucumber cycle, researchers tested the effectiveness of using both vigorous tomato plants and the fungus P. lilacinum to control M. incognita worms. Worm populations and crop losses might be reduced with just plant resilience [5]. This type of study is significant because it demonstrates the possibility of combining the effectiveness of chemicals and fungi, which improves the results compared to using them separately. Additionally, this study showed that some chemical nematicides are compatible with P. lilacinum. Fungal inoculation causes a reduction in egg viability, which leads to nematode control. In vitro, the fungus parasitized 94.5% of eggs, but not embryos, and had protease, lipase, and chitinase activities [14]. An important aspect to consider for nematode control is the impact of water stress and temperature on the growth and enzymatic activity of P. lilacinum. This fungus can combat Nacobbus aberrans, a type of plant-parasitic nematode. The fungus exhibited the ability to thrive under varying levels of water stress, but its growth rate decreased as water stress levels intensified. In addition, under inductive conditions, the fungus produced chitinases, proteases, and leucinostatins. The study concluded that P. lilacinum was a useful biocontrol agent for phytoparasitic nematodes in tomatoes under various agroecological conditions. Additionally, P. lilacinum demonstrated saprophytic soil colonization ability and displayed endophytic potential in tomato plants [15]. The formulation of biological products is important because it can significantly impact the effectiveness of fungi in controlling nematodes. Therefore, extensive studies focusing on the compatibility between the fungus and the formulation are necessary before creating a new product. Such studies are crucial to ensure that the new product will be effective in controlling nematodes and to prevent any reduction in efficacy due to compatibility issues between the fungus and the formulation. A new bioformulation of P. lilacinum for managing plant-parasitic nematodes has been tested. Karanja deoiled cake (KDG) and sundried biogas sludge (BGS) were evaluated as a mixed base to create a bioformulation for plant-parasitic nematodes. P. lilacinum spores with improved virulence were mass-produced using optimum food sources. KDC-BGS base with P. lilacinum controlled worms in wheat better than P. lilacinum alone. Controlling plant-parasitic nematodes with the KDC-BGS base bioformulation with P. lilacinum was eco-friendly and cost-effective [16]. This study has shown the importance of the bioformulation for the effectiveness and efficiency of the use of P. lilacinum for biocontrol. Another possibility to inoculate P. lilacinum is using liquid formulations to apply biological agents for agriculture: liquid formulations are easy to handle and apply. They can be easily sprayed onto crops using conventional spraying equipment, making them a convenient option for farmers. Liquid formulations can provide better crop coverage than other formulations, such as powders or granules. This is because they can be applied evenly and can penetrate the plant tissue more effectively, increasing the efficacy of the biological agent. Liquid formulations can have a longer shelf life than others, as they are less susceptible to environmental conditions such as humidity and temperature. This can result in a more stable and effective product, reducing the need for frequent replacements and saving costs for farmers. Finally, liquid formulations can be easily mixed with other agricultural inputs, such as fertilizers, herbicides, or insecticides, allowing farmers to save time and reduce the number of applications needed. This also reduces the potential for phytotoxicity or negative interactions between products [17]. Liquid formulations have been evaluated using three biocontrol agents, Pseudomonas fluorescens, P. lilacinum, and T. viride, against potato cyst nematodes under field conditions. The biocontrol agents were administered as seed treatment and soil drenching. Their efficacy was compared with that of carbofuran. The findings indicated that all three biocontrol agents effectively reduced cyst nematode eggs and density, root penetration, and egg numbers cyst-1, with varying degrees of efficacy based on their application method. P. fluorescens was the most effective in reducing egg density, multiplication rate, root penetration, and egg numbers cyst-1, followed by P. lilacinum. The potato plants treated with P. fluorescens and P. lilacinum were taller and had higher tuber yields than untreated plants. P. fluorescens showed higher root colonization in cyst nematodes than T. viride, while P. lilacinum showed the opposite trend. The biocontrol agent efficacy was comparable to that of carbofuran. In conclusion, P. fluorescens and P. lilacinum applied through seeds as liquid formulations were highly effective for managing cyst nematodes under naturally infested field conditions [11].
Eggplant is an important crop for many farmers and communities around the world. It is a high-value crop that can provide a source of income for small-scale farmers, and it is often grown in areas with limited water resources or poor soil quality, where other crops may struggle to thrive [18]. P. lilacinum may control several plant pathogens in eggplants, and some studies have shown that its effectiveness relies on the dose. An in vitro study using eggplants showed that P. lilacinum significantly reduced the rates of egg hatching and juvenile survival of M. incognita in a dose-dependent manner. Interestingly, P. lilacinum penetrated the eggs and promoted damage in the juvenile, indicating its high potential for controlling this parasite. The results indicated that P. lilacinum could be used as a biological control for reducing diseases caused by nematodes in eggplants [19]. An excellent strategy to improve the effectiveness of P. lilacinum to control nematodes is its inoculation mixed with other fungi. An interesting fungus that may be inoculated with P. lilacinum has been Trichoderma due to its synergistic potential to control nematodes and its compatibility with P. lilacinum. A study has shown that three Trichoderma strains and two P. lilacinum strains may be used against M. javanica, a plant-parasitic worm that reduces pineapple output. Pineapple is an important crop in agriculture due to its economic, nutritional, adaptability, environmental, and cultural benefits [20]. Two Trichoderma and two P. lilacinum strains substantially lowered nematode egg and egg mass production, reducing root galling harm and increasing plant root mass growth. Trichoderma asperullum and P. lilacinum strains reduced galls, egg mass, and ova. Trichoderma atroviride did not impact worm growth or root damage, unlike the control T. asperellum. and P. lilacinum strains from pineapple can, directly and indirectly, impact worm growth and host response, suggesting that they may inhibit M. javanica [21].
Soybean is an essential crop worldwide, and its grains are a primary source of protein and oil for both human and animal consumption and are used to make a variety of food products, including tofu, soy milk, soy sauce, and vegetable oil, which are consumed around the world. Soybean is also a key ingredient in livestock, poultry, and aquaculture animal feed. It is a cost-effective source of protein and essential amino acids for animal growth and health. Soybean is one of the most widely traded crops in the world, with major producers including the United States, Brazil, and Argentina. Soybean exports generate billions of dollars in revenue yearly, with China being the largest importer of soybeans [22]. However, it is susceptible to various types of nematodes that can cause significant harm and reduce its productivity, resulting in substantial financial losses. Mixtures of P. lilacinum, Claroideoglomus etunicatum, and Rhizophagus clarus were evaluated against M. incognita in soybean under greenhouse conditions. Twelve indigenous fungal and bacterial rhizosphere isolates were tested against the root-knot nematode M. graminicola. Research has shown that biocontrol agent culture filtrates impede egg hatching and kill nematode juveniles [10]. Another important crop is rice. Rice is a staple food for over half of the world's population, particularly in Asia and Africa. It provides a significant source of calories, carbohydrates, and nutrients, such as vitamins and minerals, essential for human health. Rice production and trade are significant contributors to the global economy. This cereal is one of the most important crops for small farmers in developing countries, providing income and employment opportunities. In addition, rice exports generate billions of dollars in revenue each year for major producers such as Thailand, Vietnam, and India. Rice cultivation can have positive environmental impacts, as it can help prevent soil erosion, promote biodiversity, and conserve water resources through efficient irrigation practices [23]. The mixture of microorganisms has also been evaluated in rice crops. The six most effective biocontrol agents have been tested regarding their potential for reducing nematode damage and boosting plant growth and yield. Pseudomonas putida controlled root-knot disease best of the biocontrol agents evaluated. T. harzianum colonized M. graminicola eggs, juveniles, and adult females the most, followed by P. lilacinum. Bacillus subtilis had limited impacts. P. putida, P. fluorescens, and T. harzianum had the highest rhizosphere population increase. In naturally infected areas, P. putida and T. harzianum controlled M. graminicola and improved rice tillering and yield. Pseudomonas putida and T. harzianum are potential biocontrol agents for controlling M. graminicola in rice crops [24]. Endophyte fungi have the potential to not only enhance plant growth but also act as natural pest control agents by reducing the survival rate of insect pests. Numerous endophytic fungi exhibit entomopathogenic abilities and are currently being utilized for insect control, such as P. lilacinum. However, limited information is available regarding the potential use of Beauveria bassiana with P. lilacinum, known for its insecticidal properties, which can reside within plants without causing any harm. The presence of these fungi in cotton plants positively impacted their growth and made it difficult for insects to survive on them. This indicates that these fungi could be a sustainable and effective alternative for protecting crops from insects. The results suggest that manipulating fungal endophytes could be valuable in integrated pest management strategies prioritizing sustainability [6].
Fungal endophytes can shield plants from a variety of stresses, including pests. Entomopathogenic fungal endophytes are the term given to this fungus, and the biological functions these fungi can perform are significant in farming systems. Entomopathogenic fungal strains have been found in several plants, and a single isolate can be introduced to develop as an endophyte across a range of plants [6]. According to [25], these entomopathogenic fungal endophytes are categorized as nonclavicipitaceous, which refers to fungal endophytes that are typically spread laterally. Many nonclavicipitaceous entomopathogens, such as Beauveria bassiana, Lecanicillium lecanii, Metarhizium anisopliae, and Isaria (Paecilomyces) spp., may combat plant diseases in planta and promote plant development. The impact of P. lilacinum and B. bassiana fungi that potentially benefit soil and plant health in sandy and clayey soils from the Brazilian Cerrado has been examined. In a greenhouse, assays were conducted using corn and soybean plants. The study findings suggest that applying these fungal isolates can benefit soil function and quality in the medium to long term. These positive impacts could lead to the development and adoption of more sustainable agricultural practices, which is of significant importance [26]. To evaluate the potential use of P. lilacinum against insects, this fungus was tested for its ability to manage the Thai orchid bug Thrips palmi. A study showed that P. lilacinum could biocontrol Thrips palmi in Thai flower farms. After four applications, both P. lilacinum therapies considerably reduced thrips. Other biocontrol drugs, imidacloprid, and the water + clay control did not significantly affect the thrips population. In the field test, the water + clay solution killed some thrips, probably due to clay particle tips wounding the insect skin [27].
The cotton crop is an important crop due to its economic, cultural, and social significance. Its fibres are used in the textile industry, it provides income for millions of farmers, and its byproducts are used for food and animal feed. Additionally, cotton production can support biodiversity and promote sustainable agriculture practices [28]. An important insect that promotes high production losses in cotton crops is Helicoverpa zea larvae. P. lilacinum has also been explored due to its potential to control insects when inoculated with other fungal endophytes, particularly Beauveria bassiana, to improve cotton growth and reduce the survival rate of Helicoverpa zea larvae. The researchers observed that these endophytes, once introduced into cotton plants, adversely impacted the survival of larvae that consumed them. Consequently, the authors proposed that manipulating fungal endophytes could significantly impact sustainable integrated pest management practices [6].
Most entomopathogenic fungi infect and ultimately destroy insects and other arthropods via conidia that adhere to the cuticle and hyphae that penetrate the cuticle following conidial germination. It is extremely uncommon for insects to become infected via the digestive tract after ingesting spores. Upon the demise of a host, new spores are typically produced on the corpse and disseminated into the adjacent environment [29]. P. lilacinum has been tested as a biological control agent of several insect pests. For example, it was demonstrated that this fungus was able to control leaf-cutter ants. The inoculation of this fungus significantly reduced the survival of ants from six Acromyrmex lundii field colonies, with a median lethal time of 6-7 days. The fungus was responsible for 85.6% of the mortality in inoculated ants and showed a good competitive capability against other entomopathogens that naturally infected ants. Horizontal transmission to noninoculated ants was also observed. The study proposes the use of P. lilacinum as a potential biological control agent of leaf-cutter ants in crops and plantations [30]. P. lilacinum can be used to control another important insect that causes great economic losses. For instance, the pathogenicity of P. lilacinum isolated from Egypt was evaluated against four pests, Bemisia tabaci, Tetranychus urticae, Thrips tabaci, and Diuraphis noxia. The fungal filtrate had a higher toxic effect on all tested pests than the spore suspension. The greater fungal filtrate concentration resulted in higher mortality rates seven days posttreatment, reaching 100% for T. tabaci and T. urticae and 97% for B. tabaci and D. noxia [31]. The fungus P. lilacinum has proven to be a highly effective biocontrol agent against a broad spectrum of insects, including the tomato leafminer pest. This is an interesting finding that highlights the potential of this fungus as a natural solution for pest management [32].
Purpureocillum lilacinum is a fungus that produces a variety of secondary metabolites with potentially important applications in medicine, agriculture, and industry. Some of these secondary metabolites have been found to possess antimicrobial, anticancer, and antiviral properties, making them promising candidates for developing new drugs [33]. Plants and microbes only create secondary metabolites during certain stages of development. Pigments, hormones, poisons, and antibiotics are all tiny molecules with complicated chemical structures that are not essential for normal development and reproduction [34]. Purpureocillum lilacinum has been identified as a great producer of leucinostatin D, acremoxanthone G, and acremonidin. Leucinostatin A, B, C, and D are considered biologically active secondary metabolites. They were found to be effective against a wide variety of yeasts, both pathogenic and nonpathogenic, as well as filamentous fungi, and to have moderate activity against gram-positive bacteria. In addition to secondary metabolites with potential use in biotechnology, the fungus also produces a substance of agricultural importance [7]. The effects of two secondary metabolites produced by P. lilacinum, indole-3-carboxaldehyde and indole-3-carboxylic acid, on regulating tobacco mosaic virus have been evaluated. In vitro and in vivo, these secondary metabolites demonstrated potent antiviral activity. They split virus virions and caused the fragmented particles to clump together. Virus application also caused immunological reactions or resilience to virus infection in tobacco, including the expression of hypersensitive reactions, increased defense-related enzymes, and overexpression of pathogenesis-related proteins. The increase in the salicylic acid biosynthesis of several genes showed that salicylic acid functioned as a defense-related signaling molecule in plants. As a result, indole compounds can activate tobacco defenses against mosaic viruses and other viruses and can be used for disease management [35]. Various food-grade antioxidants and natural polyphenols are compatible with P. lilacinum. The most suitable compounds with the 21 genotypes of P. lilacinum were found to be 2(3)-tert-butyl-4 hydroxyanisole (BHA), 2,6-di(tert-butyl)-p-cresol (BHT), 3-phenyl-2-propenoic acid (CA), and the mixture of BHA and BHT. Furthermore, certain types develop better in the presence of certain therapies. The effect of these compounds and entomopathogenic fungus mixtures on Aspergillus flavus growth metrics and toxin generation in vitro was also investigated. The study found that combining entomopathogenic fungus with BHA (1 mM) enhanced the latency period and reduced the Aspergillus flavus growth rate and aflatoxin B1 generation at three water activities evaluated. These results imply that combining food-grade antioxidants and natural polyphenols with entomopathogenic fungi could be a successful and long-term approach to managing aflatoxin-producing fungi [36].
Purpureocillium lilacinum as biofertilizer
Some P. lilacinum strains present several characteristics related to plant growth promotion. However, few studies have been carried out showing these abilities. This fungus is a promising microorganism that can be used for this purpose. A study with maize, bean and soybean evaluated the potential of P. lilacinum, Purpureocillium lavendulum, and Metarhizium marquandii as plant growth-promoters. The researchers tried the most promising fungi in greenhouse trials for their ability to solubilize phosphorus and make indoleacetic acid (IAA). Some breeds boosted phosphate and nitrogen supply and affected plant growth metrics differently for each crop. However, field changes did not always match in vitro changes. In conclusion, these fungi may be used as bioinoculants for agricultural plant growth [9]. The effects of five beneficial nonmycorrhizal fungi with P. lilacinum on soil and plants in sandy and clayey soils from the Brazilian Cerrado were investigated. The study was conducted under corn and soybean plants in greenhouse assays. The results indicate that applying these fungal isolates can positively impact soil function and quality in the medium and long term. This could ultimately contribute to developing and adopting more sustainable agricultural practices [12]. Another study investigated the effects of five fungal endophytes, including P. lilacinum, on the growth, biological, and chemical properties of rhizosphere soil under Phaseolus vulgaris L. The study aimed to identify the impact of fungal isolates on plant growth and soil fertility and to highlight the benefits of using these treatments in reducing the environmental impacts of mineral fertilizers and pesticides in sustainable agriculture. The report also discusses the role of plant growth-promoting fungi in regulating phytohormones and contributing to plant growth. The findings suggest that fungal isolates can enhance soil fertility and promote plant growth, making them valuable in sustainable agricultural management systems [4]. P. lilacinum can improve soil nutrients and plant growth with biofertilizers. Biofertilizer microorganisms that increase nutrient availability through biological processes were assessed for Capsicum chinense plants, focusing on P. lilacinum. P. lilacinum solubilized basic P sources and produced siderophores and indole-3-acetic acid in this study. P. lilacinum also affected greenhouse plant growth, crop quality, and yield. P. lilacinum solubilized Ca3(PO4)2 better than FePO4, creating siderophores and IAA. P. lilacinum boosted Capsicum chinense plant height, stem girth, and foliar P. Inoculated plants had heavier fruit and larger lengthwise diameters. Fruit yield remained unchanged. This work shows that P. lilacinum biofertilizer improves C. chinense plant growth and nutrient intake [3].
Purpureocillum lilacinum for bioremediation
Purpureocillum lilacinum can be used for bioremediation, reducing soil pollution. It is a good strategy to decrease environmental impacts and improve soil degradation. A mixture of P. lilacinum and Penicillium chrysogenum has been studied for the bioremediation of heavy oil waste. When fed carbon and nitrogen, these fungi biodegrade oil waste compounds. Fungal therapy altered hydrocarbon component abundances. P. lilacinum degraded 100% cycloalkanes, 21.2% n-alkanes, 15.1% aromatics, and 14.5% iso-alkanes. P. chrysogenum altered the hydrocarbon makeup differently. It decreased n-alkanes (1.3%) and increased cycloalkanes (82.6%), aromatics (10.2%), and iso-alkanes (11.9%). P. lilacinum and P. chrysogenum also cleared 15.3% and 7.6% of the changed oil sludge's petroleum compounds, respectively. These findings demonstrate that P. lilacinum and P. chrysogenum can bioremediate oil sludge-contaminated soils by degrading hydrocarbons [37]. Another study investigated the impact of copper addition to soil on the growth and physiology of Sedum alfredii, a type of plant. The findings reveal that adding copper to the soil led to increased copper accumulation in the plant, with variations in copper content observed between the aboveground and belowground parts of the plant. Additionally, the study assessed the chlorophyll content and water status to evaluate their physiological response to copper exposure [38].
Mechanisms of action
As previously mentioned, the fungus P. lilacinum has shown great potential as an endophyte and entomopathogenic fungus with numerous applications in biotechnology and agriculture. Various studies have highlighted its abilities, including biocontrol, biofertilization, and bioremediation, achieved through different modes of action. In the case of biocontrol of nematodes, the fungus produces various enzymes, including chitinases and proteases, which digest the chitin and proteins in the nematode's body, leading to its death [39]. Moreover, this fungus is capable of reducing the incidence of phytopathogenic fungi by damaging their cell membranes and components, ultimately leading to their demise [40]. Furthermore, during certain stages of fungal development and under appropriate environmental conditions, P. lilacinum produces secondary metabolites by expressing specific genes, which are then released into the environment. These molecules have various applications in biotechnology and medicine [7]. In Figure 1, a schematic representation of the isolation, identification, and various uses of P. lilacinum in agriculture is presented. Figure 2 shows the evaluations regarding the inoculation of P. lilacinum in the plants with two modes of application in the crops. Table 1 displays several abilities of P. lilacinum, including its ability to control phytopathogenic fungi, promote plant growth, and produce secondary metabolites.
Conclusions
Purpureocillum lilacinum is an endophytic, entomopathogenic microorganism with several important abilities that can be used for various purposes. It can be used for nematode and insect control, as well as a biofertilizer to make nutrients available to plants in the soil. Additionally, it can be used for phytopathogenic control to reduce the presence of pathogens and subsequently decrease the incidence of diseases. By harnessing these diverse skills, P. lilacinum presents promising applications in biotechnology and agriculture. Its use can help reduce the dependency on chemical fertilizers, pesticides, and nematicides and contribute to sustainable production practices.
Future Perspectives
As mentioned, the fungus Purpureocillum lilacinum presents many different skills, and many of them may be used for biotechnological and agricultural purposes. However, some gaps need to be solved, and information should be generated. Could this fungus make nitrogen available, releasing the nitrogen and phosphorus tapped in the organic matter? Could this fungus make the available phosphorus by solubilizing different sources of unavailable phosphorus? Which insects could be controlled by inoculating this fungus? Could this fungus be used to reduce production costs by reducing the use of chemical fertilizers and pesticides without reducing productivity? These are some gaps that more research needs to answer to make this fungus available to farmers.
Funding
The authors thank FAPESP for financial support Project Number 2021/10821-8.
References
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Frontiers in plant science 2018, 9, 1473. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Pataczek, L.; Hilger, T.H.; Zahir, Z.A.; Hussain, A.; Rasche, F.; Schafleitner, R.; Solberg, S.Ø. Perspectives of microbial inoculation for sustainable development and environmental management. Frontiers in microbiology 2018, 9, 2992. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Salazar, R.; Sánchez-García, I.; Chan-Cupul, W.; Ruiz-Sánchez, E.; Hernández-Ortega, H.A.; Pineda-Lucatero, J.; Figueroa-Chávez, D. Plant growth, foliar nutritional content and fruit yield of Capsicum chinense biofertilized with Purpureocillium lilacinum under greenhouse conditions. Scientia Horticulturae 2020, 261, 108950. [Google Scholar] [CrossRef]
- Alves, G.S.; Bertini, S.C.B.; Barbosa, B.B.; Pimentel, J.P.; Junior, V.A.R.; de Oliveira Mendes, G.; Azevedo, L.C.B. Fungal endophytes inoculation improves soil nutrient availability, arbuscular mycorrhizal colonization and common bean growth. Rhizosphere 2021, 18, 100330. [Google Scholar] [CrossRef]
- Dahlin, P.; Eder, R.; Consoli, E.; Krauss, J.; Kiewnick, S. Integrated control of Meloidogyne incognita in tomatoes using fluopyram and Purpureocillium lilacinum strain 251. Crop Protection 2019, 124, 104874. [Google Scholar] [CrossRef]
- Lopez, D.C.; Sword, G.A. The endophytic fungal entomopathogens Beauveria bassiana and Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum) and negatively affect survival of the cotton bollworm (Helicoverpa zea). Biological Control 2015, 89, 53–60. [Google Scholar] [CrossRef]
- Chen, W.; Hu, Q. Secondary metabolites of Purpureocillium lilacinum. Molecules 2021, 27, 18. [Google Scholar] [CrossRef]
- Spina, F.; Tummino, M.L.; Poli, A.; Prigione, V.; Ilieva, V.; Cocconcelli, P.; Puglisi, E.; Bracco, P.; Zanetti, M.; Varese, G.C. Low density polyethylene degradation by filamentous fungi. Environmental Pollution 2021, 274, 116548. [Google Scholar] [CrossRef]
- Baron, N.C.; de Souza Pollo, A.; Rigobelo, E.C. Purpureocillium lilacinum and Metarhizium marquandii as plant growth-promoting fungi. PeerJ 2020, 8, e9005. [Google Scholar] [CrossRef]
- Messa, V.R.; da Costa, A.C.T.; Kuhn, O.J.; Stroze, C.T. Nematophagous and endomycorrhizal fungi in the control of Meloidogyne incognita in soybean. Rhizosphere 2020, 15, 100222. [Google Scholar] [CrossRef]
- Nagachandrabose, S. Management of potato cyst nematodes using liquid bioformulations of Pseudomonas fluorescens, Purpureocillium lilacinum and Trichoderma viride. Potato Research 2020, 63, 479–496. [Google Scholar] [CrossRef]
- Barbosa, B.B.; Pimentel, J.P.; Rodovalho, N.S.; Bertini, S.C.B.; Kumar, A.; Ferreira, L.F.R.; Azevedo, L.C.B. Ascomycetous isolates promote soil biological and nutritional attributes in corn and soybeans in sandy and clayey soils. Rhizosphere 2022, 24, 100625. [Google Scholar] [CrossRef]
- El-Ashry, R.; Ali, M.A.; Elsobki, A.E.; Aioub, A.A. Integrated management of Meloidogyne incognita on tomato using combinations of abamectin, Purpureocillium lilacinum, rhizobacteria, and botanicals compared with nematicide. Egyptian Journal of Biological Pest Control 2021, 31, 1–10. [Google Scholar] [CrossRef]
- Giné, A.; Sorribas, F.J. Effect of plant resistance and BioAct WG (Purpureocillium lilacinum strain 251) on Meloidogyne incognita in a tomato–cucumber rotation in a greenhouse. Pest management science 2017, 73, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Girardi, N.S.; Sosa, A.L.; Etcheverry, M.G.; Passone, M.A. In vitro characterization bioassays of the nematophagous fungus Purpureocillium lilacinum: Evaluation on growth, extracellular enzymes, mycotoxins and survival in the surrounding agroecosystem of tomato. Fungal Biology 2022, 126, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sharma, S.; Sabir, N.; El-Sheikh, M.A.; Alyemeni, M. Impact assessment of Karanja deoiled cake and sundried biogas slurry as a mixed substrate on the nematicidal potential of Purpureocillium lilacinum. Journal of King Saud University-Science 2021, 33, 101399. [Google Scholar] [CrossRef]
- Silva, D.M.; de Souza, V.H.M.; Moral, R.d.A.; Delalibera Júnior, I.; Mascarin, G.M. Production of Purpureocillium lilacinum and Pochonia chlamydosporia by Submerged Liquid Fermentation and Bioactivity against Tetranychus urticae and Heterodera glycines through Seed Inoculation. Journal of Fungi 2022, 8, 511. [Google Scholar] [CrossRef]
- Chapman, M.A. Introduction: The importance of eggplant. The Eggplant Genome 2019, 1–10. [Google Scholar]
- Khan, M.; Tanaka, K. Purpureocillium lilacinum for plant growth promotion and biocontrol against root-knot nematodes infecting eggplant. Plos one 2023, 18, e0283550. [Google Scholar] [CrossRef]
- Hossain, M. World pineapple production: An overview. African Journal of Food, Agriculture, Nutrition and Development 2016, 16, 11443–11456. [Google Scholar] [CrossRef]
- Kiriga, A.W.; Haukeland, S.; Kariuki, G.M.; Coyne, D.L.; Beek, N.V. Effect of Trichoderma spp. and Purpureocillium lilacinum on Meloidogyne javanica in commercial pineapple production in Kenya. Biological Control 2018, 119, 27–32. [Google Scholar] [CrossRef]
- Pagano, M.C.; Miransari, M. The importance of soybean production worldwide. In Abiotic and biotic stresses in soybean production; Elsevier: 2016; pp. 1-26.
- Khush, G. Productivity improvements in rice. Nutrition reviews 2003, 61, S114–S116. [Google Scholar] [CrossRef] [PubMed]
- Haque, Z.; Khan, M.R.; Ahamad, F. Relative antagonistic potential of some rhizosphere biocontrol agents for the management of rice root-knot nematode, Meloidogyne graminicola. Biological control 2018, 126, 109–116. [Google Scholar] [CrossRef]
- Hernandez-Trejo, A.; Estrada-Drouaillet, B.; Lopez-Santillan, J.A.; Rios-Velasco, C.; Rodríguez-Herrera, R.; Osorio-Hernández, E. Effects of native entomopathogenic fungal strains and neem extract on Spodoptera frugiperda on Maize. Southwestern entomologist 2019, 44, 117–124. [Google Scholar] [CrossRef]
- Castillo Lopez, D.; Zhu-Salzman, K.; Ek-Ramos, M.J.; Sword, G.A. The entomopathogenic fungal endophytes Purpureocillium lilacinum (formerly Paecilomyces lilacinus) and Beauveria bassiana negatively affect cotton aphid reproduction under both greenhouse and field conditions. PloS one 2014, 9, e103891. [Google Scholar] [CrossRef] [PubMed]
- Hotaka, D.; Amnuaykanjanasin, A.; Maketon, C.; Siritutsoontorn, S.; Maketon, M. Efficacy of Purpureocillium lilacinum CKPL-053 in controlling Thrips palmi (Thysanoptera: Thripidae) in orchid farms in Thailand. Applied Entomology and Zoology 2015, 50, 317–329. [Google Scholar] [CrossRef]
- Vitale, J. Economic importance of cotton in Burkina Faso. Food and Agriculture Organisation, Rome, Italy. Retrieved April 2018, 20, 2018. [Google Scholar]
- Rodrigues, J.; Bergamini, C.; Montalva, C.; Humber, R.A.; Luz, C. Simple method to detect and to isolate entomopathogenic fungi (Hypocreales) from mosquito larvae. Journal of Invertebrate Pathology 2021, 182, 107581. [Google Scholar] [CrossRef]
- Goffré, D.; Folgarait, P.J. Purpureocillium lilacinum, potential agent for biological control of the leaf-cutting ant Acromyrmex lundii. Journal of Invertebrate Pathology 2015, 130, 107–115. [Google Scholar] [CrossRef]
- Desoky, S.M.A.; Abdelall, M.F.; Ahmed, Y. Isolation, Identification, Evaluation of Purpureocillium Lilacinum Egyptian Isolate Toxicity Test in Vitro and Analysis Its Bioactive Products. Journal of the Advances in Agricultural Researches 2022, 27, 602–617. [Google Scholar] [CrossRef]
- Bali, G.K.; Singh, S.K.; Maurya, D.K.; Wani, F.J.; Pandit, R.S. Morphological and molecular identification of the entomopathogenic fungus Purpureocillium lilacinum and its virulence against Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae) larvae and pupae. Egyptian Journal of Biological Pest Control 2022, 32, 86. [Google Scholar] [CrossRef]
- Rodriguez, R.; White Jr, J.; Arnold, A.; Redman, a.R.a. Fungal endophytes: diversity and functional roles. New phytologist 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Liu, Z.; Lin, R.; Li, E.; Mao, Z.; Ling, J.; Yang, Y.; Yin, W.-B.; Xie, B. Biosynthesis of antibiotic leucinostatins in bio-control fungus Purpureocillium lilacinum and their inhibition on Phytophthora revealed by genome mining. PLoS pathogens 2016, 12, e1005685. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wu, H.; Zhou, W.; Yuan, Z.; Hao, J.; Liu, X.; Han, L. Effects of indole derivatives from Purpureocillium lilacinum in controlling tobacco mosaic virus. Pesticide Biochemistry and Physiology 2022, 183, 105077. [Google Scholar] [CrossRef] [PubMed]
- Barra, P.; Nesci, A.; Etcheverry, M. In vitro compatibility of natural and food grade fungicide and insecticide substances with Purpureocillium lilacinum and their effect against Aspergillus flavus. Journal of stored products research 2013, 54, 67–73. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, J.; Liu, Y.; Feng, W. Biodegradation of hydrocarbons by Purpureocillium lilacinum and Penicillium chrysogenum from heavy oil sludge and their potential for bioremediation of contaminated soils. International Biodeterioration & Biodegradation 2023, 178, 105566. [Google Scholar]
- Gong, B.; Liu, G.; Liao, R.; Song, J.; Zhang, H. Endophytic fungus Purpureocillium sp. A5 protect mangrove plant Kandelia candel under copper stress. Brazilian journal of microbiology 2017, 48, 530–536. [Google Scholar] [CrossRef]
- Zhan, J.; Qin, Y.; Gao, K.; Fan, Z.; Wang, L.; Xing, R.; Liu, S.; Li, P. Efficacy of a chitin-based water-soluble derivative in inducing Purpureocillium lilacinum against nematode disease (Meloidogyne incognita). International Journal of Molecular Sciences 2021, 22, 6870. [Google Scholar] [CrossRef]
- Elsherbiny, E.; Taher, M.; Abd El-Aziz, M.; Mohamed, S. Action mechanisms and biocontrol of Purpureocillium lilacinum against green mould caused by Penicillium digitatum in orange fruit. Journal of Applied Microbiology 2021, 131, 1378–1390. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the steps in which the fungus was isolated from soil samples in the first phase. The isolates were removed from the pure culture (isolated) in the second phase. In the third phase, the fungus was identified as Purpureocillum lilacinum using molecular biology. The fungal strain was then cultured in liquid medium to test its capacity to create indol acetic acid and secondary metabolites, as well as its ability to detoxify soil and water through bioremediation.
Figure 1.
Schematic representation of the steps in which the fungus was isolated from soil samples in the first phase. The isolates were removed from the pure culture (isolated) in the second phase. In the third phase, the fungus was identified as Purpureocillum lilacinum using molecular biology. The fungal strain was then cultured in liquid medium to test its capacity to create indol acetic acid and secondary metabolites, as well as its ability to detoxify soil and water through bioremediation.
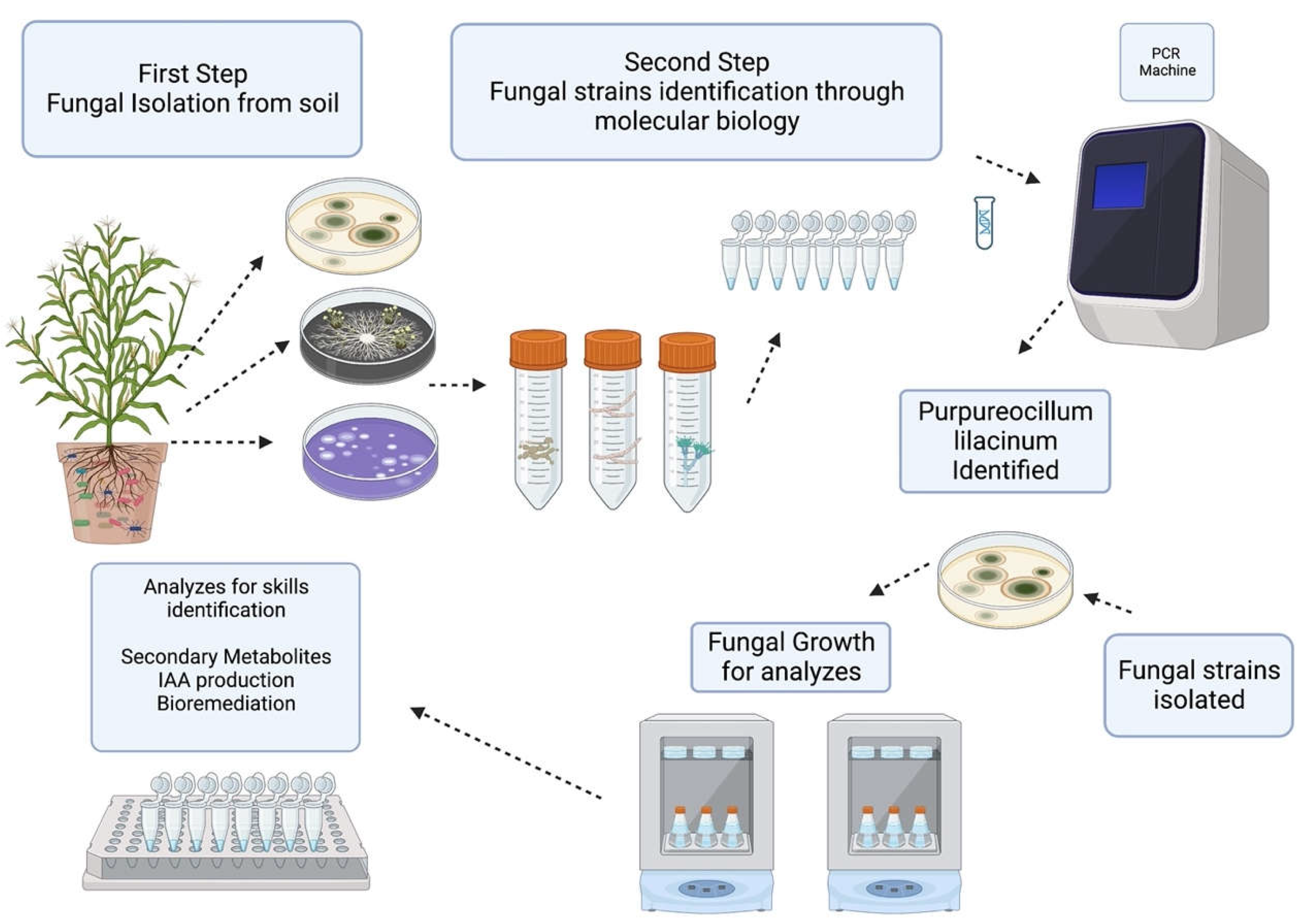
Figure 2.
The fungus was cultured in liquid media and then applied to plants to test and assess its influence on plant growth. The fungus was also sprayed on plants to assess its biological control impact in lowering the occurrence of nematodes in the soil. In addition, the fungus was administered to the plant to verify and assess pest control. Finally, the fungus was introduced to water and soil to test its bioremediation impact. The modes of application might be furrow straight into the soil or areal application on the leaves.
Figure 2.
The fungus was cultured in liquid media and then applied to plants to test and assess its influence on plant growth. The fungus was also sprayed on plants to assess its biological control impact in lowering the occurrence of nematodes in the soil. In addition, the fungus was administered to the plant to verify and assess pest control. Finally, the fungus was introduced to water and soil to test its bioremediation impact. The modes of application might be furrow straight into the soil or areal application on the leaves.
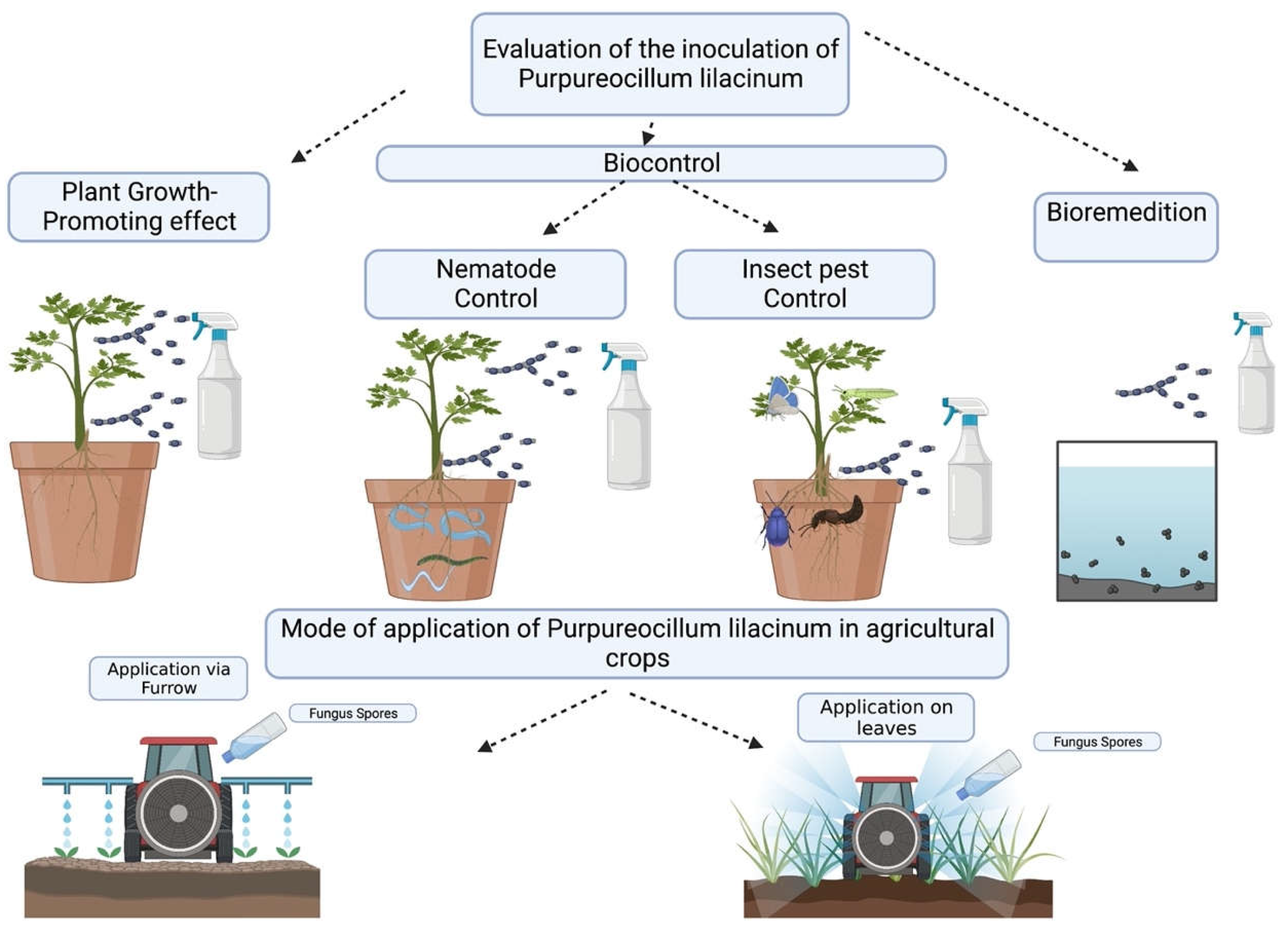

Table 1.
Purpureocillum lilacinum abilities to control phytopathogen fungi, secondary metabolites production and plant growth promoting.
Table 1.
Purpureocillum lilacinum abilities to control phytopathogen fungi, secondary metabolites production and plant growth promoting.
| Kind of Effect | Results | References |
|---|---|---|
|
Nematode Control |
Control of root-knot nematodes in tomatoes | Kepenekci et al., 2018 |
| Control of Meloidogyne javanica in commercial pineapple |
Kiriga et al., 2018 | |
| The fungus showed maximum egg mass inhibition of Meloidogyne incognita | Sharma et al., 2021 | |
| Reduction of Meloidogyne incognita in tomatoes | Dahlin et al., 2019 | |
| The fungus was effectively applied as biocontrol agents of phytoparasitic nematodes in tomatoes under variable agroecological conditions. |
Girardi et al., 2022 | |
| Suppressed nematode Meloidogyne incognita population in a tomato–cucumber rotation in a greenhouse |
Giné and Sorribas, 2017 | |
| The fungus controlled root-knot nematodes infecting eggplant | Khan and Tanaka, 2023 | |
|
Secondary Metabolites |
Production of indole-3-carboxaldehyde (A1) andindole-3-carboxylic acid for tobacco mosaic virus control | Sun et al., 2022 |
| Production of metabolites for biological control | Chen and Hu, 2021 | |
| Proteases production | Chen and Hu, 2021 | |
|
Biological Control |
Controlled the insect Thrips palmi in orchid farms | Hotaka et al., 2015 |
| Potential control against cotton aphids | Castillo Lopez et al., 2014 | |
|
Plant-Growth |
In soybean stimulated root growth and nutrient absorption | Barbosa et al., 2022 |
| Increased the availability of P and N and promoted the growth of maize, beans and soybean | Baron et al., 2020 | |
| Improved soil nutrient availability in common bean growth | Alves et al., 2021 | |
| Promoted significant increases in plant dry biomass in cotton crop | Lopez and Sword, 2015 | |
|
Bioremediation |
Reduced the amount of total petroleum hydrocarbons in contaminated soils | Yang et al., 2023 |
| Removed 44.55% of crude oil from contaminated soil | Benguenab and Chibani, 2021 | |
| Protected mangrove plant Kandelia candel under copper stress | Gong et al., 2017 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



