
Submitted:
26 May 2023
Posted:
29 May 2023
You are already at the latest version
Abstract
Prolonged exercise induces acute renal injury; however, the precise mechanism remains unclear. We investigated the effects of neutrophil depletion in male C57BL/6J mice. Male C57BL/6J mice were divided into four groups: sedentary with control antibody, sedentary with antineutrophil antibody, exhaustive exercise with control antibody, and exhaustive exercise with antineutrophil antibody. Antineutrophil (1A8) or control antibody was administered intraperitoneally to the mice prior to their running on a treadmill. Renal histology was assessed 24 h after exhaustive exercise, and the concentration of kidney injury molecule (KIM)-1 was measured using enzyme-linked immunosorbent assay. The expression levels of inflammatory cytokines were measured using quantitative reverse transcriptase-polymerase chain reaction. Furthermore, nicotinamide adenine dinucleotide phosphate oxidase activity and hydrogen peroxide concentration in the kidneys were measured. The pathologic changes were manifested as congested and swollen glomeruli, tubular dilatation, and nuclear infiltration after exhaustive exercise. These changes were suppressed by the administration of the 1A8 antibodies. KIM-1 concentration increased after exhaustive exercise but were reduced by the 1A8 antibodies. Treatment with the 1A8 antibody also decreased exhaustive exercise-induced inflammation and reactive oxygen species levels in the kidney. These results suggest that neutrophils contribute to exercise-induced acute renal injury by regulating inflammation and reactive oxygen species levels.
Keywords:
neutrophil
; exhaustive exercise
; inflammation
; cytokines
; reactive oxygen species
; acute kidney injury
1. Introduction
Moderate exercise is effective in maintaining or improving health [1], whereas prolonged intense physical activity, particularly during endurance exercise causes tissue injury, including muscle and acute renal injury [2,3]. In fact, with regard to acute renal injury, it has been reported that gross and microscopic hematuria was observed in 20% of the runners after marathon races [4]. We also have been shown that after duathlon racing in humans, acute renal injury was indicated by elevated serum creatinine (CRE) and urine protein levels, and tubular epithelial cells were also detected in urine sediments [5]. Recently, a systematic review of endurance exercise-induced acute renal injury was reported [6]. In 11 case reports, 27 cases of acute kidney injury requiring hospital treatment were reported, and in 21 studies, renal function were assessed before and after endurance exercise in 800 participants, with increased serum CRE levels. An animal model also showed that exhaustive exercise increased the levels of blood urea nitrogen (BUN) and CRE in the plasma [7]. In addition, we have reported that exhaustive exercise causes renal histological injury manifested as congested and swollen glomeruli, tubular dilatation, and inflammatory cell infiltration in the interstitium [8]. We also found an increase in kidney injury molecule (KIM)-1 and inflammation after exhaustive exercise. Nonetheless, the exact mechanisms responsible for acute renal injury following intense exercise remain unclear.
Several experimental models of acute kidney injury have demonstrated the infiltration of inflammatory cells, such as neutrophils and macrophages [9]. Inflammatory cells release proinflammatory cytokines (such as tumor necrosis factor (TNF)-α, interleukin (IL)-1β and IL-6) and reactive oxygen species (ROS) [10,11]. The damaging effects of inflammation and ROS on the kidney implies that inflammatory cells may play an important role in the mechanisms involved in exercise-induced acute renal injury. For example, several studies have reported that macrophages are involved in ischemia/reperfusion (I/R)-induced acute kidney injury [12,13,14]. One of the primary reasons of exercise-induced acute renal injury is that during exercise, the blood flow to the kidneys decreases as the body directs blood towards the working muscles, leading to I/R-induced reoxygenation after exercise [15]. In fact, during intense exercise, renal blood flow has been shown to be reduced to 25% of resting levels [16]. Furthermore, renal biopsy after endurance exercise showed features of acute tubular necrosis, suggesting an ischaemic aetiology [6]. Thus, exhaustive exercise can cause acute renal injury, similar to that induced by I/R. We have been reported that exhaustive exercise-induced renal injury is suppressed by the depletion of macrophages [8]. These findings indicate that macrophage infiltration of the kidneys contributes significantly to the development of acute kidney injury after exhaustive exercise.
In addition to macrophages, some studies suggest that neutrophils play an important role in the development of acute kidney injury through the inflammatory pathway. For example, neutrophil depletion by antineutrophil serum administration has been shown to ameliorate acute kidney injury induced by I/R [17,18]. Grenz et al. also showed that neutrophil depletion improved acute kidney injury induced by I/R and decreased the production of inflammatory cytokines, such as TNF-α [19]. I/R-induced renal injury is also prevented in models of neutrophil depletion by administration of antibodies to ICAM-1 and ICAM-1 knockout [20,21]. It has also been reported that exhaustive exercise-induced renal injury, including infiltration of immune cells, occurs 6 h after exercise, and that these are suppressed by administration of antioxidant supplements [22]. This infiltration of immune cells seen in the exhaustive exercise-induced renal injury in the early stage, such as 6 h after exercise, might have been caused by neutrophils. In addition, neutrophils are known to release macrophage chemoattractants, such as monocyte chemoattractant protein-1 (MCP-1), and regulate the infiltration of macrophages into local tissues to induce inflammation [23]. These results indicate that neutrophil infiltration might be a crucial mediator of the inflammatory pathways leading to acute kidney injury after exhaustive exercise. However, the specific mechanisms underlying neutrophil-mediated acute renal injury during exercise are currently unknown. Identifying the mechanisms underlying exercise-induced acute renal injury could facilitate the development of innovative prevention and treatment approaches for athletes engaged in strenuous training. Given the growing popularity of endurance exercise events such as marathon and ultramarathon races, it may also be possible to prevent post-race injuries in citizen runners. The aim of this study was to investigate whether the augmented infiltration of neutrophils following exhaustive exercise leads to inflammation and acute renal injury. To achieve this, we examined the effect of antineutrophil antibody injection on inflammation, macrophage infiltration, and acute renal injury after exhaustive exercise.
2. Materials and Methods
2.1. Animals
Male C57BL/6J mice were purchased from Kiwa Laboratory Animals (Wakayama, Japan) at 10 weeks of age and housed with four mice per cage in a controlled environment under a light/dark cycle (lights on at 9:00 a.m. and off at 9:00 p.m.). The experimental procedures complied with the guiding principles for the care and use of animals at the Waseda University, and were approved by the Institutional Animal Care and Use Committee of the university (2013-A110). Mice were randomly assigned to four groups: sedentary with control antibody (n = 20), sedentary with antineutrophil antibody (n = 20), exhaustive exercise with control antibody (n = 20), and exhaustive exercise with antineutrophil antibody (n = 20). All mice had free access to standard chow and water.
2.2. Injection of antineutrophil antibody
A neutrophil-specific antibody, anti-Ly-6G (clone 1A8) and isotype control antibody (clone 2A3) were purchased from Bio X Cell (Sunnyvale, CA, USA). The 1A8 (0.5 μg) and 2A3 (0.5 μg) antibodies were individually diluted in phosphate-buffered saline, and the mice were treated with 150 μL of either antibody solution intraperitoneally, according to their respective experimental groups.
2.3. Exercise protocol
Mice in the sedentary groups remained under resting conditions in the cage, whereas mice in the exercise groups were subjected to exhaustive exercise 48 h after injection. One week before exhaustive exercise, mice in all groups were familiarized with running on a motorized treadmill (Natsume, Tokyo, Japan). On the day of the experiment, the mice were forced via a shock grid to run on a treadmill with a 7% gradient at a speed of 10 m/min for 15 min, followed by 15 m/min for 15 min, 20 m/min for 15 min, and finally 24 m/min until exhaustion. Exhaustion was defined as the point at which the mice refused to run, despite touching the shock grid five times.
2.4. Blood and kidney sampling
Ten animals from each group were sacrificed immediately after exhaustive exercise, and 10 at 24 h after exhaustive exercise. Anesthesia was induced with 2% isoflurane inhalation at 0.8 L/min, maintained at 1% at 0.8 L/min. Blood samples were collected from the abdominal aorta using a 1-mL syringe mounted with a 20-gauge needle and coated with heparin (5,000 IU/mL, Nipro, Osaka, Japan). Blood samples were transferred to a tube coated with heparin and centrifuged at 2,600 × g for 10 min, and the plasma was stored at -80°C until analysis.
The kidneys were removed, and the right kidneys were immersed in liquid nitrogen, snap frozen, and stored at -80°C until analysis; the left kidneys were frozen in Tissue-tek Crymould (Sakura, Torrance, CA) filled with OCT compound (Sakura) with samples immersed in precooled isopentane at -80°C.
2.5. Assessment of renal function
Plasma CRE and BUN levels, as blood markers of renal injury, were analyzed using Oriental Yeast Co., Ltd. (Tokyo, Japan).
2.6. Hematoxylin and eosin (H&E) staining
The kidney specimens were fixed in 10% paraformaldehyde before being embedded in paraffin. The specimens were sectioned at 3 μm, deparaffinized, and stained with H&E for light microscopic analysis.
2.7. Immunohistochemical staining
Immunohistochemical staining was performed to evaluate the expression of Ly-6G, KIM-1, and F4/80. The specimens were sectioned at 3 μm, deparaffinized, and stained using the ImmunoCruz ABC Staining System (Santa Cruz Biotechnology, Dallas, TX, USA). Mouse antiLy-6G antibody (127601; BioLegend, San Diego, CA, USA), rabbit antiKIM-1 antibody (ab47635; Abcam, London, UK), and rabbit antimouse F4/80 monoclonal antibody (M4150, Spring Bioscience, Pleasanton, CA, USA) were used as primary antibodies. Four randomly selected images were recorded at 200× magnification and Ly-6G-positive cells were counted using BZ-2 software (Keyence, Osaka, Japan).
2.8. Terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick-end labeling (TUNEL) assay
The TUNEL assay for the detection of apoptotic cells was performed using an apoptosis in situ detection kit (Wako, Osaka, Japan), according to the manufacturer’s protocol. TUNEL-positive cells were counted in four random /200× fields using BZ-2 software (Keyence).
2.9. KIM-1 assay
The concentration of KIM-1 in the kidney was measured using an enzyme-linked immunosorbent assay (ELISA) kit (Abcam). The assay was performed in accordance with the ELISA kit instructions.
2.10. Quantitative reverse transcriptase-polymerase chain reaction (RT-PCR)
Total RNA was extracted from the kidneys using the RNeasy Mini Kit (Qiagen, Valencia, CA, USA), according to the manufacturer’s instructions. Total RNA purity was assessed using the NanoDrop system (NanoDrop Technologies, Wilmington, DE, USA). Total RNA was reverse-transcribed into complementary DNA (cDNA) using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Waltham, MA, USA). Quantitative RT-PCR was performed with the Fast 7500 real-time PCR system (Applied Biosystems) using the Fast SYBR Green PCR Master Mix (Applied Biosystems). The thermal profile consisted of denaturation at 95°C for 10 min, followed by 40 cycles of 95°C for 3 s, and annealing at 60°C for 15 s. 18S ribosomal RNA was used as a housekeeping gene control, and all data were normalized to the expression levels of 18S ribosomal RNA. The data were expressed as the number of fold changes relative to the values of the sedentary group with the control antibody group. Specific PCR primer pairs used for each gene are listed in Table 1.
2.11. Nicotinamide adenine dinucleotide phosphate (NADPH) oxidase activity
NADPH oxidase activity in total kidney was determined as the oxidation of NADPH measured at 340 nm in a reaction mixture containing 50 mM Tris, 50 mM 2-(N-morpholino) ethanesulfonic acid (pH 7.0), 1 mM KCN to inhibit low levels of mitochondrial oxidase activity, and 150 mM NADPH (Sigma) for 1 min at 37°C.
2.12. Hydrogen peroxide assay
To examine hydrogen peroxide levels in the kidney, total kidney tissue was homogenized in a tissue protein extraction reagent with a protease inhibitor (Thermo, Rockford, IL, USA). Protein concentration was measured using the BCA protein assay (Thermo). Hydrogen peroxide levels were measured using the SensoLyte ADHP Hydrogen Peroxide Assay Kit (Fremount, CA, USA).
2.13. Statistical analyses
All data are presented as mean ± standard error of the mean (SEM). All statistical analyses were performed using SAS software version 9.4. To evaluate the statistical significance of exhaustive exercise and antineutrophil antibody treatment, the data were analyzed using two-way repeated measures ANOVA or two-way ANOVA. If significant interactions were observed, further comparisons were performed using Tukey’s HSD post-hoc test.
3. Results
3.1. Running time
The mean running time until the mice became exhausted was 320.3 ± 50.2 min in the control antibody group and 331.4 ± 46.0 min in the 1A8 antibody group; these values were not statistically different.
3.2. Neutrophil infiltration into the kidney
To identify the effect of 1A8 antibody treatment on exhaustive exercise-induced neutrophil infiltration, we performed immunohistochemical staining and examined the mRNA expression of Ly-6G, a specific marker of neutrophils. Ly-6G immunohistochemical staining revealed that exhaustive exercise induced a substantial increase in neutrophil infiltration into the kidney, but this infiltration was markedly reduced by the injection of the 1A8 antibody (Figure 1A and B). Consistently, while exhaustive exercise increased Ly-6G mRNA in the kidney, injection of the 1A8 antibody reduced it (Figure 1C).
3.3. Renal function
Plasma BUN and CRE were measured immediately and 24 hours after exercise to assess renal function. Immediately after exercise, plasma BUN was significantly increased by exhaustive exercise and it was suppressed by 1A8 treatment. After 24 hours of exercise, they were not statistically different between the groups (Table 2). On the other hand, there was no group and time interaction for CRE (Table 2).
3.4. Renal histology
We performed H&E, KIM-1, and TUNEL staining to reveal the kidney injury. H&E staining showed significant pathological changes in the exhaustive exercise group, manifested as congested and swollen glomeruli, tubular dilatation, and inflammatory cell infiltration in the interstitium. Compared with the exhaustive exercise group, the 1A8 antibody treatment group showed fewer histological abnormalities (Figure 2A).
Immunohistochemical staining for KIM-1, a marker of kidney injury, showed that KIM-1 expression in the exhaustive exercise with control antibody group increased predominantly in the tubular cells compared with the sedentary with control antibody group (Figure 2A). Increased KIM-1 expression was suppressed after exercise in the 1A8 antibody group (Figure 2A). Consistently, KIM-1 concentration and mRNA expression were significantly higher in the exercise with control antibody group than in the sedentary group. These increases were significantly suppressed in the exercise group with the 1A8 antibody (Figure 2B, C).
TUNEL-positive cells, a marker of apoptotic cell death, increased substantially 24 h after exhaustive exercise; however, 1A8 antibody treatment significantly reduced them (Figure 2D).

Table 1.
Effects of exhaustive exercise and neutrophil depletion on renal function.
| Gene | Time | Sed | Sed + 1A8 | Ex | Ex + 1A8 |
|---|---|---|---|---|---|
| BUN (mg/dL) | 0 h | 27.2 ±. 0.9 | 25.4 ± 1.3 | 54.9 ± 1.3* | 45.0 ± 5.0§ |
| 24 h | 25.7 ± 1.0 | 22.6 ± 1.5 | 22.6 ± 1.5 | 19.0 ± 0.8 | |
| CRE (mol/L) | 0 h | 84.0 ± 5.2 | 80.0 ± 7.8 | 87.0 ± 4.2 | 80.0 ± 2.5 |
| 24 h | 76.0 ± 3.0 | 81.0 ± 3.4 | 65.0 ± 3.0 | 69.0 ± 3.4 |
Values represent means ± SEM. Blood urea nitrogen (BUN); creatinine (CRE); sedentary (Sed); exercise (Ex). Analyses were performed using two-way repeated measures ANOVA. *P<0.05 vs Sedentary, §P<0.05 vs Exercise at the same time point.
3.5. Inflammatory cytokines and ROS in the kidney
To identify the effect of 1A8 antibody treatment on exhaustive exercise-induced renal inflammation and ROS production, we examined the mRNA expression levels of TNF-, IL-6, IL-1, NADPH oxidase activity, and hydrogen peroxide concentration. TNF- and IL-6 mRNA expression levels were increased by exercise but ameliorated by the 1A8 antibody (Figure 3A, B). Changes in IL-1 mRNA expression were similar, and not significantly different (Figure 3C). NADPH oxidase activity and hydrogen peroxide concentration were significantly increased by exhaustive exercise but were suppressed by 1A8 antibody treatment (Figure 3D, F).
3.6. Macrophage infiltration into the kidney
The effect of exhaustive exercise and neutrophil depletion on macrophage infiltration in the kidney was evaluated using immunohistochemical staining and mRNA expression of F4/80, a specific marker of macrophages. F4/80 immunohistochemical staining revealed that exhaustive exercise induced a substantial increase in macrophage infiltration into the kidney, but this infiltration was markedly reduced by the treatment of the 1A8 antibody (Figure 4A and B). Similarly, although exhaustive exercise increased the F4/80 mRNA level, injection of the 1A8 antibody reduced it (Figure 4C). The mRNA expression of MCP-1, which recruits monocytes and macrophages to the sites of inflammation, was increased by exercise but ameliorated by the 1A8 antibody (Figure 4F).
To assess the effects of exhaustive running on the phenotypic switch in renal macrophage polarization, we examined the gene expression of M1 and M2 macrophage markers in the kidney. The expression of CD11c mRNA, which reflects the presence of M1 macrophages, was significantly increased in the exhaustive exercise group. This increase was significantly suppressed by the 1A8 treatment. In contrast, CD163 mRNA expression, reflecting the presence of M2 macrophages, was not significantly altered by exercise or 1A8 treatment.
4. Discussion
The incidence of exercise-induced acute renal injury has been reported to be 4-85% [24,25,26,27,28]. Given the increasing popularity of endurance exercise events, clarifying the mechanisms of exercise-induced renal injury would be useful information not only for athletes and citizen runners but also for exploring the countermeasures such as prevention and treatment. However, the underlying mechanisms are not well understood. This study was an attempt to suppress neutrophil infiltration in acute renal injury.
Prolonged exercise increases the concentration and migration of neutrophils in the blood [29,30]. Activated neutrophils then produce inflammatory mediators, ROS, and proteases that can promote tissue injury. Neutrophils infiltrate into tissues via chemokines and adhesion factors in I/R-induced kidney injury [9]. We showed increased blood IL-8, which is neutrophil chemokine and CD62L expression on blood neutrophils after prolonged exercise [31,32], suggesting increased infiltration capacity of neutrophils. In this study, Ly-6G immunohistochemical staining and mRNA expression showed an increase in neutrophil infiltration into the kidneys after exhaustive exercise. We used clone 1A8, an antiLy-6G antibody, to deplete neutrophils from the tissues. The 1A8 antibody was injected intraperitoneally 24 h before exercise and successfully reduced neutrophil infiltration in the kidney after exhaustive exercise (Figure 1). These results demonstrated the efficacy of our protocol in reducing neutrophil infiltration.
Immune cells, such as neutrophils and macrophages, trigger inflammatory responses in the kidney by activating the production of proinflammatory cytokines [19]. In addition, neutrophils undergo a burst of oxygen consumption via NADPH oxidase, leading to the generation of ROS, including hydrogen peroxide. Inflammatory cytokines and ROS produced by neutrophils further exacerbate acute kidney injury. In fact, it has been reported that the expression levels of inflammatory cytokines are elevated in the kidneys after exhaustive exercise8. In other studies, renal malondialdehyde increased 24 h after exhaustive exercise, suggesting increased oxidative stress [22]. In the present study, acute renal injury after exhaustive exercise was reversed by blocking neutrophil infiltration into the kidney with a preexercise injection of 1A8 antibody (Figure 2). Our findings support prior studies showing that acute kidney injury after I/R was significantly reduced in neutrophil-depleted mice. Furthermore, neutrophil depletion with 1A8 antibody prior to exercise was found to reduce mRNA expression of inflammatory cytokines such as TNF-α and IL-6 in the kidneys, as well as reactive oxygen species (ROS) levels. In a previous study, TNF-α neutralization inhibited ischemia-induced renal tubular cell apoptosis [33]. In addition, NADPH oxidase is activated during I/R and plays a potential role in the pathogenesis of progressive renal injury [34]. These results support a reduction in renal injury and apoptosis in this study. Taken together, the results of our study demonstrate that neutrophils play important roles in the production of inflammatory cytokines, and ROS, leading to acute renal injury.
A limitation of this study is the lack of evaluation over time. Because Wu et al. found that renal injury, inflammation, and oxidative stress responses peak 24 hours after exhaustive exercise [22], we evaluated at this time point. In addition, they showed that blood CRE and BUN levels increased immediately after exhaustive exercise and returned to baseline at 24 h [22]. We therefore also assessed blood CRE and BUN immediately after exercise. BUN was significantly increased by exhaustive exercise, in agreement with previous studies, while CRE was not affected by exhaustive exercise (Table 2). In previous studies, blood CRE peaked 6 hours after exhaustive exercise [22], and this study might not have captured the post-exercise increase. Recently, KIM-1 has attracted attention as a marker in exercise-induced acute renal injury [35,36]. In the present study, exhaustive exercise induced pathological changes in the kidneys and increased the KIM-1 concentration and mRNA expression (Figure 2). Previous studies have found elevated urinary KIM-1 after endurance exercise, suggesting early tubular dysfunction [37].Thus, although we did not find an increase in CRE in this study, tissue injury in the kidneys would have occurred.
Previous studies have shown that macrophage depletion markedly reduced I/R-induced acute kidney injury and inflammation [12,13,14]. We have also reported that macrophage depletion reduced acute renal injury and inflammation after exhaustive exercise in mice [8]. This evidence suggests that macrophage infiltration is the primary cause of acute renal injury following exhaustive exercise. In the present study, the 1A8 antibody treatment decreased macrophage infiltration in the kidneys. Interestingly, it has been reported that there are different types of macrophages [38]. Monocytes recruited in injured kidneys differentiate into functionally diverse macrophages. M1 macrophages produce ROS and inflammatory cytokines, which exacerbate renal injury. On the other hand, M2 macrophages produce anti-inflammatory cytokines such as IL-10 and IL-1 receptor antagonists, which moderate the inflammatory response to tissue injury [39]. In acute kidney injury induced by I/R, M1 macrophages are recruited to the kidney in the first 48 h, and M2 macrophages predominate at later time-points and contribute to tissue repair [40]. In our study, it was shown that exercise-induced increase in CD11c, a marker of M1 macrophages, was suppressed by neutrophil depletion, suggesting that neutrophils contribute to the infiltration of M1 macrophages but not M2 macrophages. However, the mechanisms by which neutrophils regulate macrophage infiltration into the kidneys remain unclear. Macrophages migrate to the kidney after neutrophils infiltrate it [41]. Signalling pathways triggered by CCR2 activation are involved in macrophage infiltration. In a previous study, CCR2-knockout mice showed reduced macrophage infiltration compared to wild-type mice [42]. In addition, I/R-induced acute kidney injury was suppressed by MCP-1 knockdown. Neutrophils have been shown to release MCP-1, which is a chemoattractant for macrophages, and thereby modulate the infiltration of macrophages into local tissues, resulting in the induction of inflammation [23]. In the present study, the treatment of 1A8 antibodies prevented the increase in MCP-1 levels after exhaustive exercise. Taken together, the presented evidence suggests that increased production of MCP-1 may be linked to the enhanced macrophages infiltration following exhaustive exercise. Furthermore, these findings suggest that neutrophils may be a source of kidney tissue-derived chemokines that contribute to macrophage infiltration in the kidney after exhaustive exercise.
I/R-induced renal injury involves not only neutrophils and macrophages but also other immune cells such as dendric cells, T lymphocytes, B lymphocytes [43]. In the future, the contribution of cells other than neutrophils and macrophages to exercise-induced renal injury should also be clarified.
5. Conclusions
We demonstrated that 1A8 antibody treatment substantially decreased the expression of proinflammatory cytokines, macrophage infiltration, and acute renal injury induced by exhaustive exercise. These findings may lead to new prevention and treatment strategies for athletes who undergo intense training.
Author Contributions
Conceptualization, K.S.; investigation, T.M.; writing—original draft preparation, T.M.; writing—review and editing, M.S. and K.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the MEXT JSPS KAKENHI, grant Number: 20H00574.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Animal Care and Use Committee in the university (2013-A110).
Data Availability Statement
The raw data supporting the conclusions of this manuscript will be made available by the authors, without undue reservation, on request to the corresponding author.
Acknowledgments
We thank Drs. Hiromi Miyazaki, and Saeko Okutsu for technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Suzuki, K.; Hayashida, H. Effect of Exercise Intensity on Cell-Mediated Immunity. Sports (Basel) 2021, 9. [Google Scholar] [CrossRef]
- Clarkson, P.M. Exertional rhabdomyolysis and acute renal failure in marathon runners. Sports Med 2007, 37, 361–363. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Nakaji, S.; Yamada, M.; Liu, Q.; Kurakake, S.; Okamura, N.; Kumae, T.; Umeda, T.; Sugawara, K. Impact of a competitive marathon race on systemic cytokine and neutrophil responses. Med Sci Sports Exerc 2003, 35, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Siegel, A.J.; Hennekens, C.H.; Solomon, H.S.; Van Boeckel, B. Exercise-related hematuria. Findings in a group of marathon runners. JAMA 1979, 241, 391–392. [Google Scholar] [CrossRef]
- Sugama, K.; Suzuki, K.; Yoshitani, K.; Shiraishi, K.; Miura, S.; Yoshioka, H.; Mori, Y.; Kometani, T. Changes of thioredoxin, oxidative stress markers, inflammation and muscle/renal damage following intensive endurance exercise. Exerc Immunol Rev 2015, 21, 130–142. [Google Scholar] [PubMed]
- Hodgson, L.E.; Walter, E.; Venn, R.M.; Galloway, R.; Pitsiladis, Y.; Sardat, F.; Forni, L.G. Acute kidney injury associated with endurance events-is it a cause for concern? A systematic review. BMJ Open Sport Exerc Med 2017, 3, e000093. [Google Scholar] [CrossRef]
- Lin, X.; Jiang, C.; Luo, Z.; Qu, S. Protective effect of erythropoietin on renal injury induced in rats by four weeks of exhaustive exercise. BMC Nephrol 2013, 14, 130. [Google Scholar] [CrossRef]
- Mizokami, T.; Shimada, M.; Suzuki, K. Macrophage Depletion Attenuates Acute Renal Damage after Exhaustive Exercise in Mice. Int J Sports Med 2022, 43, 964–970. [Google Scholar] [CrossRef]
- Zuk, A.; Bonventre, J.V. Acute Kidney Injury. Annu Rev Med 2016, 67, 293–307. [Google Scholar] [CrossRef]
- Bonventre, J.V.; Yang, L. Cellular pathophysiology of ischemic acute kidney injury. J Clin Invest 2011, 121, 4210–4221. [Google Scholar] [CrossRef]
- Friedewald, J.J.; Rabb, H. Inflammatory cells in ischemic acute renal failure. Kidney Int 2004, 66, 486–491. [Google Scholar] [CrossRef]
- Day, Y.J.; Huang, L.; Ye, H.; Linden, J.; Okusa, M.D. Renal ischemia-reperfusion injury and adenosine 2A receptor-mediated tissue protection: role of macrophages. Am J Physiol Renal Physiol 2005, 288, F722–F731. [Google Scholar] [CrossRef]
- Oh, D.J.; Dursun, B.; He, Z.; Lu, L.; Hoke, T.S.; Ljubanovic, D.; Faubel, S.; Edelstein, C.L. Fractalkine receptor (CX3CR1) inhibition is protective against ischemic acute renal failure in mice. Am J Physiol Renal Physiol 2008, 294, F264–F271. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.K.; Sung, S.A.; Cho, W.Y.; Go, K.J.; Kim, H.K. Macrophages contribute to the initiation of ischaemic acute renal failure in rats. Nephrol Dial Transplant 2006, 21, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Castenfors, J.; Mossfeldt, F.; Piscator, M. Effect of prolonged heavy exercise on renal function and urinary protein excretion. Acta Physiol Scand 1967, 70, 194–206. [Google Scholar] [CrossRef]
- Poortmans, J.R. Exercise and renal function. Sports Med 1984, 1, 125–153. [Google Scholar] [CrossRef]
- Hellberg, P.O.; Kallskog, T.O. Neutrophil-mediated post-ischemic tubular leakage in the rat kidney. Kidney Int 1989, 36, 555–561. [Google Scholar] [CrossRef]
- Klausner, J.M.; Paterson, I.S.; Goldman, G.; Kobzik, L.; Rodzen, C.; Lawrence, R.; Valeri, C.R.; Shepro, D.; Hechtman, H.B. Postischemic renal injury is mediated by neutrophils and leukotrienes. Am J Physiol 1989, 256, F794–F802. [Google Scholar] [CrossRef]
- Grenz, A.; Kim, J.H.; Bauerle, J.D.; Tak, E.; Eltzschig, H.K.; Clambey, E.T. Adora2b adenosine receptor signaling protects during acute kidney injury via inhibition of neutrophil-dependent TNF-alpha release. J Immunol 2012, 189, 4566–4573. [Google Scholar] [CrossRef]
- Kelly, K.J.; Williams, W.W., Jr.; Colvin, R.B.; Bonventre, J.V. Antibody to intercellular adhesion molecule 1 protects the kidney against ischemic injury. Proc Natl Acad Sci U S A 1994, 91, 812–816. [Google Scholar] [CrossRef]
- Kelly, K.J.; Williams, W.W., Jr.; Colvin, R.B.; Meehan, S.M.; Springer, T.A.; Gutierrez-Ramos, J.C.; Bonventre, J.V. Intercellular adhesion molecule-1-deficient mice are protected against ischemic renal injury. J Clin Invest 1996, 97, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.L.; Chen, Y.S.; Huang, X.D.; Zhang, L.X. Exhaustive swimming exercise related kidney injury in rats - protective effects of acetylbritannilactone. Int J Sports Med 2012, 33, 1–7. [Google Scholar] [CrossRef]
- Soehnlein, O.; Zernecke, A.; Eriksson, E.E.; Rothfuchs, A.G.; Pham, C.T.; Herwald, H.; Bidzhekov, K.; Rottenberg, M.E.; Weber, C.; Lindbom, L. Neutrophil secretion products pave the way for inflammatory monocytes. Blood 2008, 112, 1461–1471. [Google Scholar] [CrossRef]
- Hoffman, M.D.; Weiss, R.H. Does Acute Kidney Injury From an Ultramarathon Increase the Risk for Greater Subsequent Injury? Clin J Sport Med 2016, 26, 417–422. [Google Scholar] [CrossRef]
- Kao, W.F.; Hou, S.K.; Chiu, Y.H.; Chou, S.L.; Kuo, F.C.; Wang, S.H.; Chen, J.J. Effects of 100-km ultramarathon on acute kidney injury. Clin J Sport Med 2015, 25, 49–54. [Google Scholar] [CrossRef]
- Mansour, S.G.; Verma, G.; Pata, R.W.; Martin, T.G.; Perazella, M.A.; Parikh, C.R. Kidney Injury and Repair Biomarkers in Marathon Runners. Am J Kidney Dis 2017, 70, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Lipman, G.S.; Krabak, B.J.; Waite, B.L.; Logan, S.B.; Menon, A.; Chan, G.K. A prospective cohort study of acute kidney injury in multi-stage ultramarathon runners: the Biochemistry in Endurance Runner Study (BIERS). Res Sports Med 2014, 22, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Poussel, M.; Touze, C.; Allado, E.; Frimat, L.; Hily, O.; Thilly, N.; Rousseau, H.; Vauthier, J.C.; Chenuel, B. Ultramarathon and Renal Function: Does Exercise-Induced Acute Kidney Injury Really Exist in Common Conditions? Front Sports Act Living 2019, 1, 71. [Google Scholar] [CrossRef]
- Suzuki, K.; Totsuka, M.; Nakaji, S.; Yamada, M.; Kudoh, S.; Liu, Q.; Sugawara, K.; Yamaya, K.; Sato, K. Endurance exercise causes interaction among stress hormones, cytokines, neutrophil dynamics, and muscle damage. J Appl Physiol (1985) 1999, 87, 1360–1367. [Google Scholar] [CrossRef]
- Suzuki, K.; Yamada, M.; Kurakake, S.; Okamura, N.; Yamaya, K.; Liu, Q.; Kudoh, S.; Kowatari, K.; Nakaji, S.; Sugawara, K. Circulating cytokines and hormones with immunosuppressive but neutrophil-priming potentials rise after endurance exercise in humans. Eur J Appl Physiol 2000, 81, 281–287. [Google Scholar] [CrossRef]
- Sugama, K.; Suzuki, K.; Yoshitani, K.; Shiraishi, K.; Kometani, T. Urinary excretion of cytokines versus their plasma levels after endurance exercise. Exerc Immunol Rev 2013, 19, 29–48. [Google Scholar] [PubMed]
- Mizokami, T.; Suzuki, K. Neutrophil Depletion Attenuates Acute Liver Stress after Exhaustive Exercise in Mice. Med Sci Sports Exerc 2023, 55, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Meldrum, K.K.; Burnett, A.L.; Meng, X.; Misseri, R.; Shaw, M.B.; Gearhart, J.P.; Meldrum, D.R. Liposomal delivery of heat shock protein 72 into renal tubular cells blocks nuclear factor-kappaB activation, tumor necrosis factor-alpha production, and subsequent ischemia-induced apoptosis. Circ Res 2003, 92, 293–299. [Google Scholar] [CrossRef]
- Simone, S.; Rascio, F.; Castellano, G.; Divella, C.; Chieti, A.; Ditonno, P.; Battaglia, M.; Crovace, A.; Staffieri, F.; Oortwijn, B.; et al. Complement-dependent NADPH oxidase enzyme activation in renal ischemia/reperfusion injury. Free Radic Biol Med 2014, 74, 263–273. [Google Scholar] [CrossRef]
- Kashani, K.; Cheungpasitporn, W.; Ronco, C. Biomarkers of acute kidney injury: the pathway from discovery to clinical adoption. Clin Chem Lab Med 2017, 55, 1074–1089. [Google Scholar] [CrossRef] [PubMed]
- Juett, L.A.; James, L.J.; Mears, S.A. Effects of Exercise on Acute Kidney Injury Biomarkers and the Potential Influence of Fluid Intake. Ann Nutr Metab 2020, 76 Suppl 1, 53–59. [Google Scholar] [CrossRef]
- Hewing, B.; Schattke, S.; Spethmann, S.; Sanad, W.; Schroeckh, S.; Schimke, I.; Halleck, F.; Peters, H.; Brechtel, L.; Lock, J.; et al. Cardiac and renal function in a large cohort of amateur marathon runners. Cardiovasc Ultrasound 2015, 13, 13. [Google Scholar] [CrossRef]
- Gordon, S.; Taylor, P.R. Monocyte and macrophage heterogeneity. Nat Rev Immunol 2005, 5, 953–964. [Google Scholar] [CrossRef]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Lee, S.; Huen, S.; Nishio, H.; Nishio, S.; Lee, H.K.; Choi, B.S.; Ruhrberg, C.; Cantley, L.G. Distinct macrophage phenotypes contribute to kidney injury and repair. J Am Soc Nephrol 2011, 22, 317–326. [Google Scholar] [CrossRef]
- Kezic, A.; Stajic, N.; Thaiss, F. Innate Immune Response in Kidney Ischemia/Reperfusion Injury: Potential Target for Therapy. J Immunol Res 2017, 2017, 6305439. [Google Scholar] [CrossRef] [PubMed]
- Furuichi, K.; Wada, T.; Iwata, Y.; Kitagawa, K.; Kobayashi, K.; Hashimoto, H.; Ishiwata, Y.; Asano, M.; Wang, H.; Matsushima, K.; et al. CCR2 signaling contributes to ischemia-reperfusion injury in kidney. J Am Soc Nephrol 2003, 14, 2503–2515. [Google Scholar] [CrossRef] [PubMed]
- Bonavia, A.; Singbartl, K. A review of the role of immune cells in acute kidney injury. Pediatr Nephrol 2018, 33, 1629–1639. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of exhaustive exercise and 1A8 antibody on neutrophil infiltration in mice kidney. (A) Histochemistry analysis of Ly-6G (brown; Ly-6G positive cells, original magnification ×200). The Ly-6G-positive cells are marked by arrows. (B) Ly-6G-positive cells, (C) Ly-6G mRNA expression in the kidney. Values represent means ± standard error of the mean. Analyses were performed using two-way analysis of variance for multiple comparisons. *P < 0.05. Sedentary (Sed); exercise (Ex).
Figure 1.
Effects of exhaustive exercise and 1A8 antibody on neutrophil infiltration in mice kidney. (A) Histochemistry analysis of Ly-6G (brown; Ly-6G positive cells, original magnification ×200). The Ly-6G-positive cells are marked by arrows. (B) Ly-6G-positive cells, (C) Ly-6G mRNA expression in the kidney. Values represent means ± standard error of the mean. Analyses were performed using two-way analysis of variance for multiple comparisons. *P < 0.05. Sedentary (Sed); exercise (Ex).
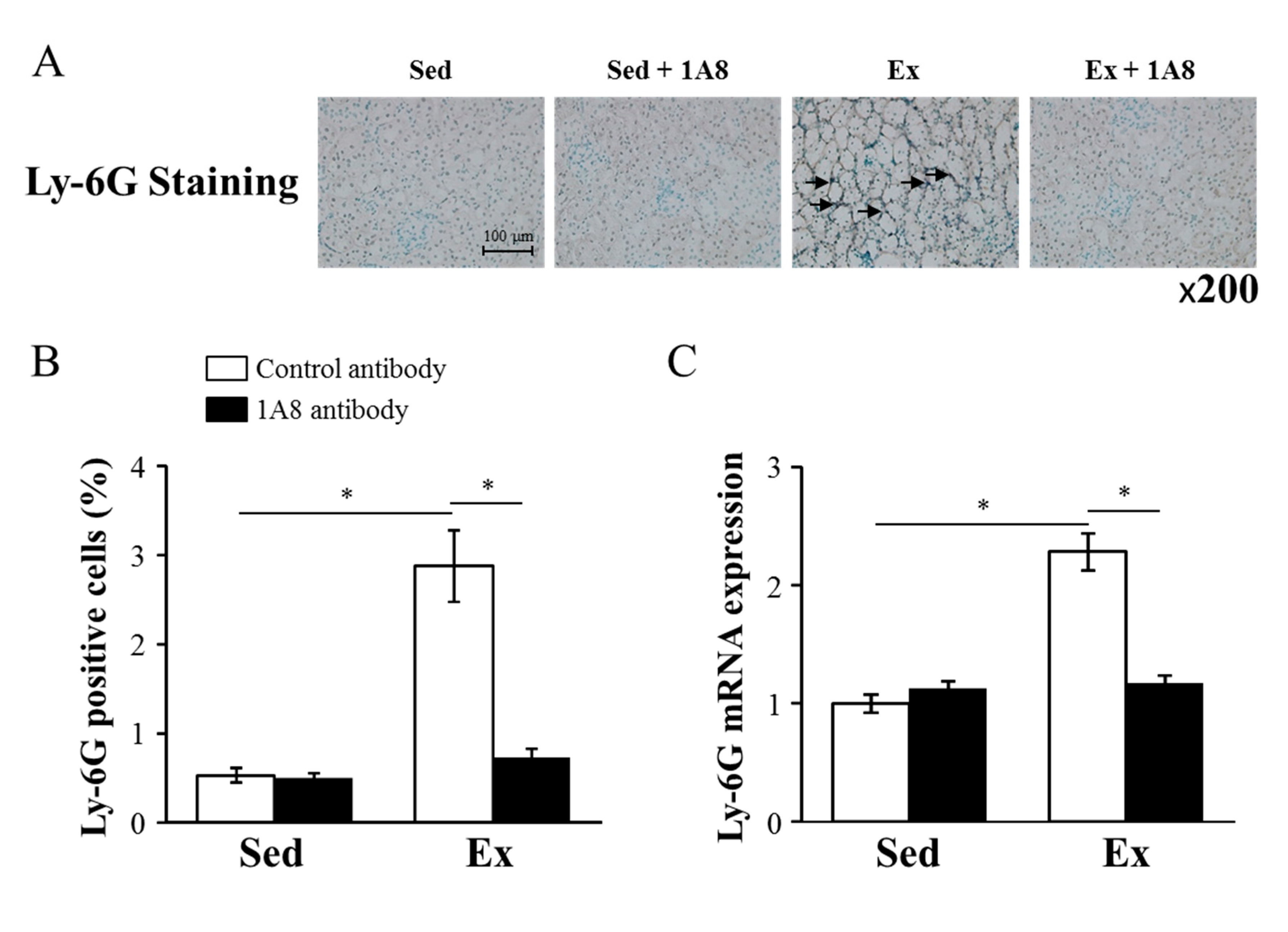
Figure 2.
Effects of exhaustive exercise and 1A8 antibody on renal damage in mice. (A) HE, KIM, and TUNEL staining of kidney sections (original magnification ×200), black arrow shows location of glomerular congestion and swelling, white arrow shows inflammatory cells infiltration and # shows tubular dilatation. A number of TUNEL-positive cells are marked by arrowheads. G shows the renal glomeruli, RT shows renal tubules. (B) (C) KIM-1 concentration and mRNA expression of kidney, (D) The percentage of TUNEL-positive cells. Values represent means ± standard error of the mean. Analyses were performed using two-way analysis of variance for multiple compar-isons. *P < 0.05. Kidney injury molecule (KIM); terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick-end labeling (TUNEL); sedentary (Sed); exercise (Ex).
Figure 2.
Effects of exhaustive exercise and 1A8 antibody on renal damage in mice. (A) HE, KIM, and TUNEL staining of kidney sections (original magnification ×200), black arrow shows location of glomerular congestion and swelling, white arrow shows inflammatory cells infiltration and # shows tubular dilatation. A number of TUNEL-positive cells are marked by arrowheads. G shows the renal glomeruli, RT shows renal tubules. (B) (C) KIM-1 concentration and mRNA expression of kidney, (D) The percentage of TUNEL-positive cells. Values represent means ± standard error of the mean. Analyses were performed using two-way analysis of variance for multiple compar-isons. *P < 0.05. Kidney injury molecule (KIM); terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick-end labeling (TUNEL); sedentary (Sed); exercise (Ex).
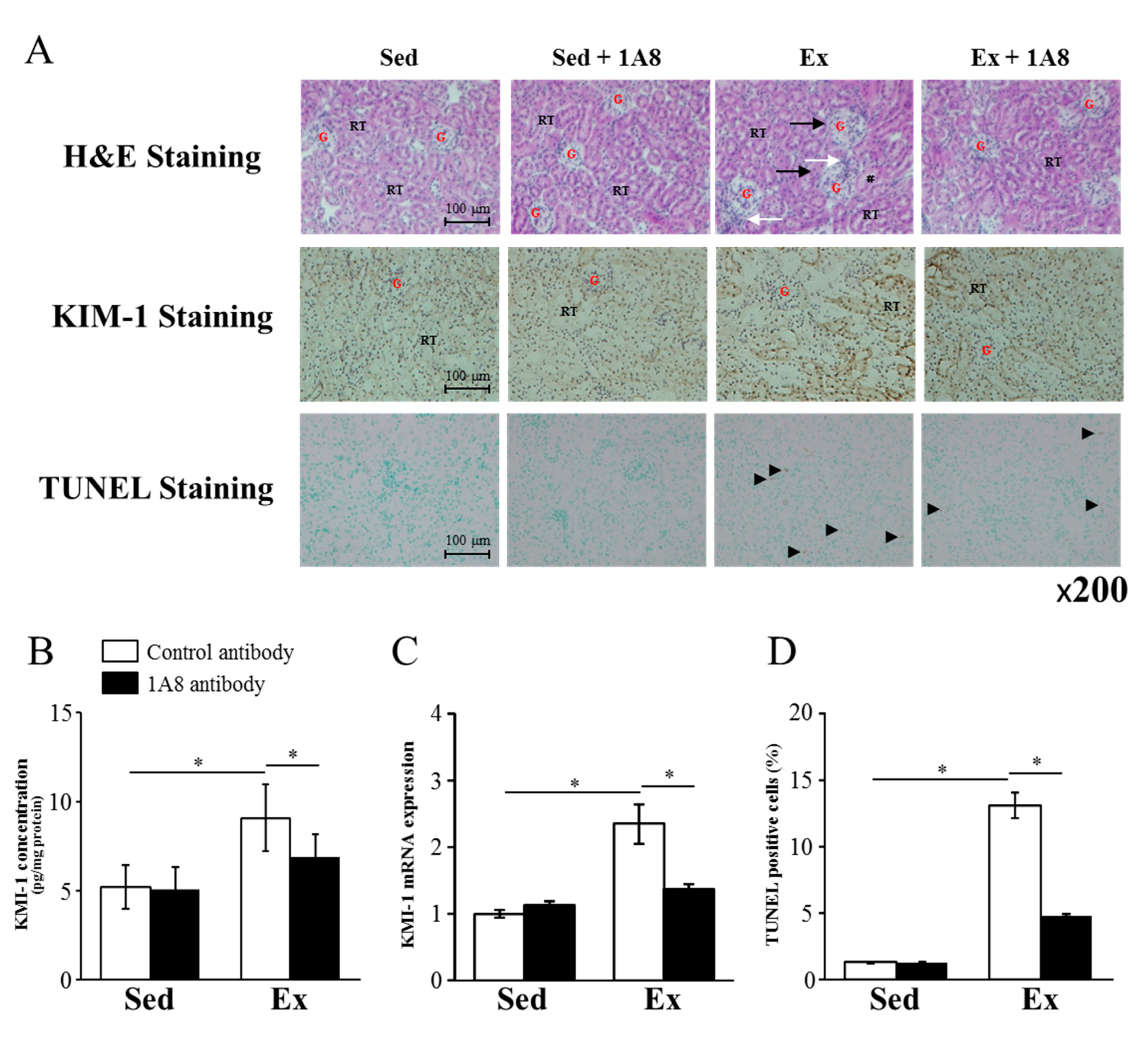
Figure 3.
Effects of exhaustive exercise and 1A8 antibody on the expressions of inflammatory cytokines and reactive oxygen species in the kidney. (A) TNF-, (B) IL-6, and (C) IL-1 mRNA expression in the kidney. (D) NADPH oxidase activity and (E) hydrogen peroxide concentration in kidney. Values represent means ± standard error of mean. Analyses were performed using two-way analysis of variance for multiple comparisons. *P< 0.05. Sedentary (Sed); exercise (Ex); tumor necrosis factor (TNF); interleukin (IL).
Figure 3.
Effects of exhaustive exercise and 1A8 antibody on the expressions of inflammatory cytokines and reactive oxygen species in the kidney. (A) TNF-, (B) IL-6, and (C) IL-1 mRNA expression in the kidney. (D) NADPH oxidase activity and (E) hydrogen peroxide concentration in kidney. Values represent means ± standard error of mean. Analyses were performed using two-way analysis of variance for multiple comparisons. *P< 0.05. Sedentary (Sed); exercise (Ex); tumor necrosis factor (TNF); interleukin (IL).
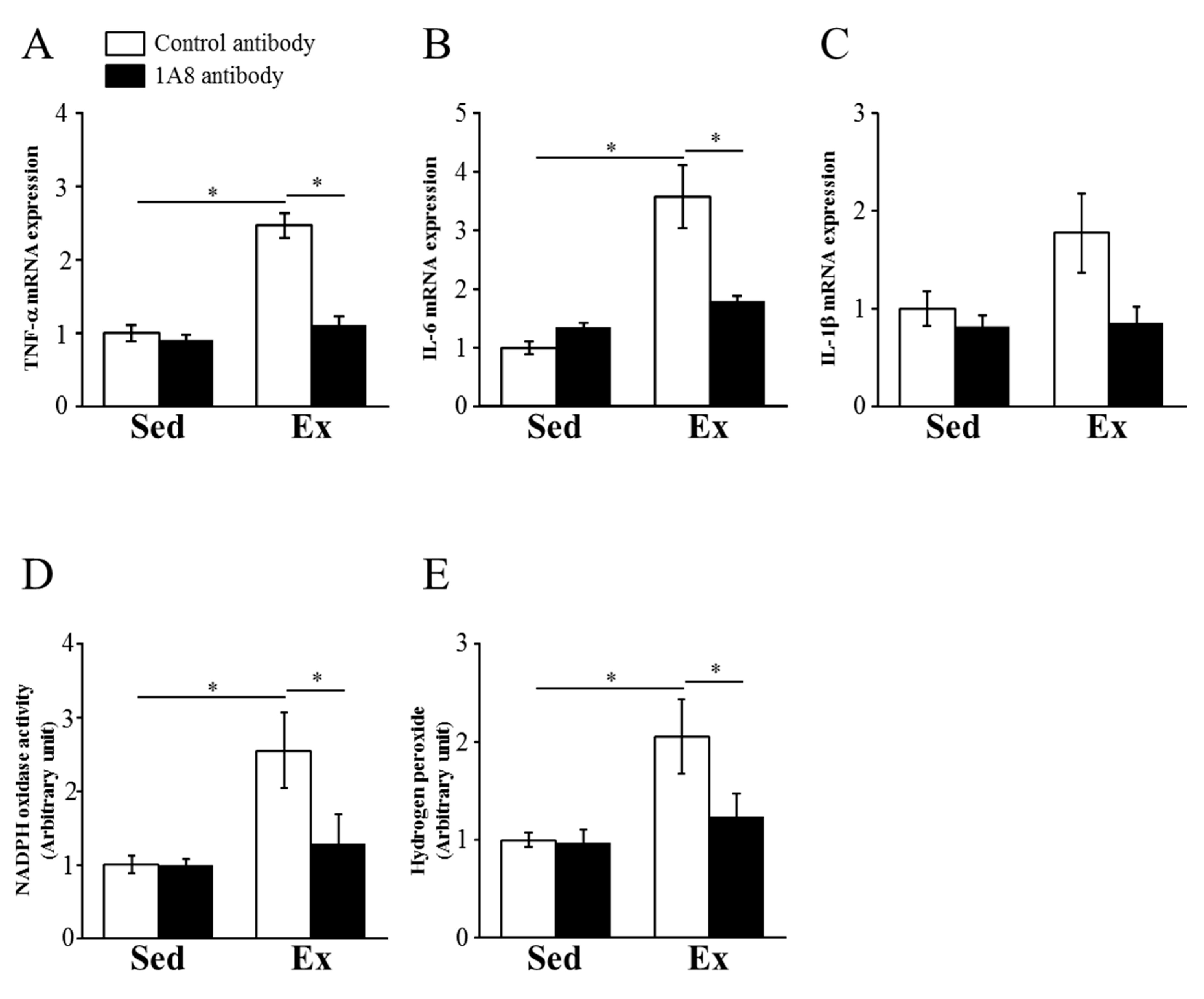
Figure 4.
Effects of exhaustive exercise and 1A8 antibody on macrophage infiltration in mice kidney. (A) Histochemistry analysis of F4/80 (brown; F4/80 positive cells, original magnification ×200). The F4/80 positive cells are marked by arrows. (B) F4/80-positive cells, (C) F4/80, (D) CD11c, (E) CD163, and (F) MCP-1 mRNA expression levels in the kidney. Values represent means ± standard error of mean. Analyses were performed using two-way analysis of variance for multiple comparisons. *P < 0.05. Monocyte chemoattractant protein (MCP); sedentary (Sed); exercise (Ex).
Figure 4.
Effects of exhaustive exercise and 1A8 antibody on macrophage infiltration in mice kidney. (A) Histochemistry analysis of F4/80 (brown; F4/80 positive cells, original magnification ×200). The F4/80 positive cells are marked by arrows. (B) F4/80-positive cells, (C) F4/80, (D) CD11c, (E) CD163, and (F) MCP-1 mRNA expression levels in the kidney. Values represent means ± standard error of mean. Analyses were performed using two-way analysis of variance for multiple comparisons. *P < 0.05. Monocyte chemoattractant protein (MCP); sedentary (Sed); exercise (Ex).
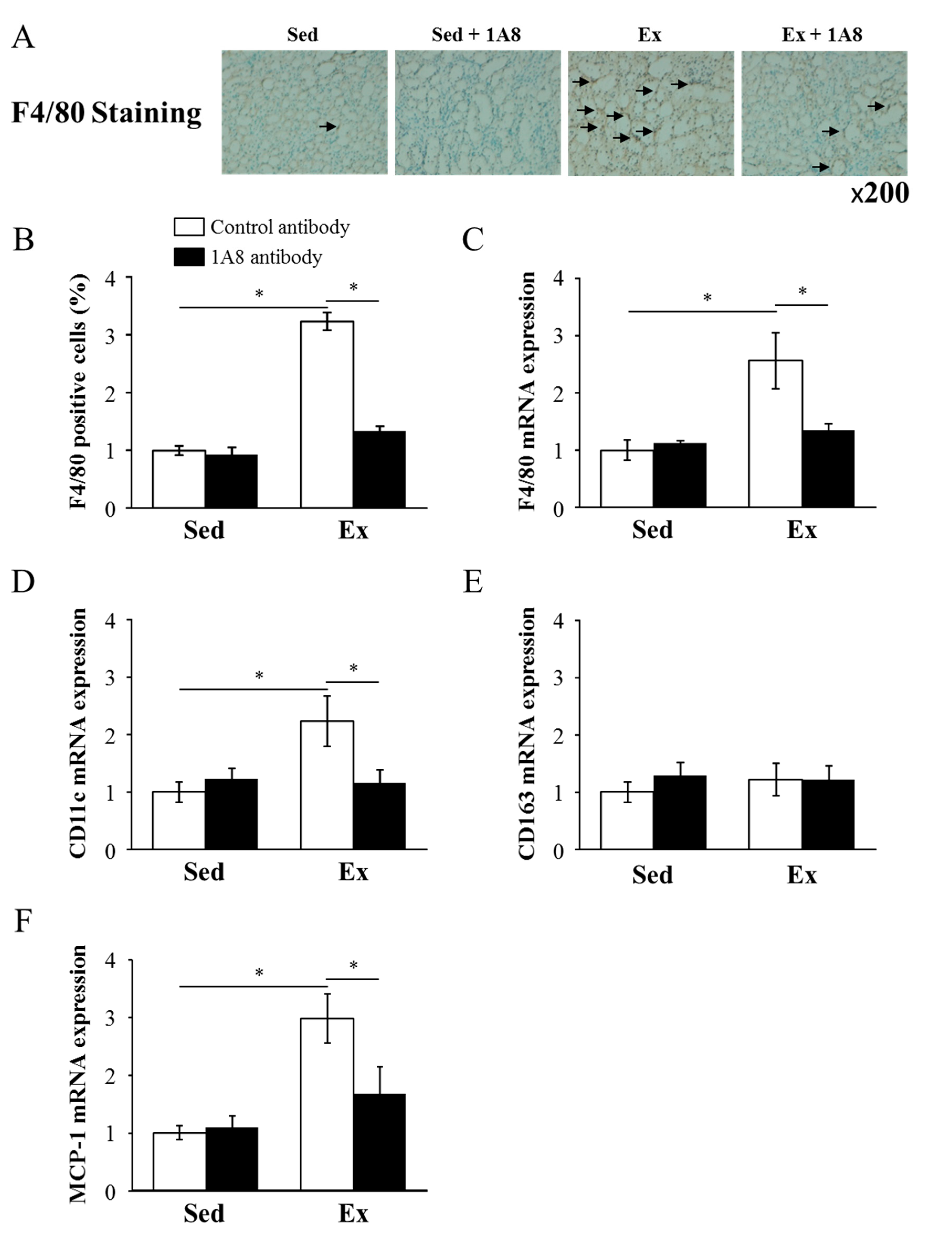
Table 1.
Primer sequences for RT-PCR analysis.
| Gene | Forward | Reverse |
|---|---|---|
| 18s ribosomal RNA | CGGCTACCACATCCAAGGA | AGCTGGAATTACCGCGGC |
| Ly-6G | TGGACTCTCACAGAAGCAAAG | GCAGAGGTCTTCCTTCCAACA |
| TNF- | TCTTCTCATTCCTGCTTGTGG | GAGGCCATTTGGGAACTTCT |
| IL-6 | AACGATGATGCACTTGCAGA | TGGTACTCCAGAAGACCAGAGG |
| IL-1 | GGGCCTCAAAGGAAAGAATC | TTGCTTGGGATCCACACTCT |
| F4/80 | CTTTGGCTATGGGCTTCCAGTC | GCAAGGAGGACAGAG-TTTATCGTG |
| MCP-1 | CTTCTGGGCCTGCTGTTCA | CCAGCCTACTCATTGGGATCA |
Tumor necrosis factor (TNF); interleukin (IL); monocyte chemoattractant protein (MCP).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



