
Submitted:
26 May 2023
Posted:
29 May 2023
You are already at the latest version
Abstract
As global climate change continues and the vegetation period lengthens, the importance of research into cyanobacterial recruitment biomass and associated toxin risks is growing. While most studies focus on planktonic cyanobacteria causing blooms, benthic cyanobacteria have been less explored. This study reports for the first time on the mass proliferation of benthic epilithic macrocolonies of potentially toxic cyanobacteria, Gloeotrichia cf. natans, in water bodies located in a region with continuous permafrost in Yakutia, North-East Russia. The study characterizes the environmental conditions of its habitat, including the chemical composition of the water, which expands our understanding of this species' ecology. Cyanotoxins (microcystins, cylindrospermopsin, saxitoxins, and anatoxin-a) were not detected in the biomass of Gloeotrichia cf. natans using liquid chromatography-mass spectrometry and PCR methods.
Keywords:
benthic
; macrocolonies
; cyanobacteria
; Gloeotrichia
; Nostocales
; cryolithozone
; Yakutia
1. Introduction
Cyanobacteria are an important component of aquatic ecosystems, especially as nitrogen-fixing organisms [1]. However, since many species of cyanobacteria are capable of producing toxins (including hepatotoxins, neurotoxins, cytotoxins, and dermatotoxins), their mass proliferation poses a serious threat to fishing and recreational water use [2,3]. The phenomenon of a rapid increase of planktonic cyanobacteria populations, known as 'blooms', has long been the focus of a large number of studies. While research on the mass proliferation of benthic cyanobacteria lags somewhat behind, the associated risks remain insufficiently studied. The number of publications dedicated to the investigation of toxic benthic cyanobacterial species has increased in recent decades, but despite this, benthic cyanobacteria remain much less studied than planktonic ones [4,5]. In recent decades, aquatic ecosystems around the world have become increasingly vulnerable to the effects of phenomena related to global climate change [6,7]. The problem of invasive species, which are colonizing new ecological spaces made available by changes in a number of environmental parameters, has become acute [8]. Invasive taxons, appearing where they were not previously observed, are able to rapid growth, and form dominate assemblages, displacing native species [9].
A population of tapering, heterocyte-bearing trichomes arranged radially inside hemispherical or spherical mucilage colonies investment was recently discovered on a bank of Buotama River, Yakutia Republic, Russia. It was completely consistent with the description of Gloeotrichia.
According to recent studies [5,10,11,12,13], Gloeotrichia species are known to produce microcystin-LR and microcystin-RR of a potent heptapeptide hepatotoxins group. Gloeotrichia genus (Aphanizomenonaceae, Nostocales) includes 26 species of filamentous, nitrogen-fixing cyanobacteria that form large colonies [14,15]. Overall, the research on Gloeotrichia toxin production is still limited, and more studies are needed to fully understand the types and concentrations of toxins produced by different Gloeotrichia species.
In this study, we report for the first time on the mass proliferation of macrocolonies of a potentially toxic cyanobacterium Gloeotrichia cf. natans Rabenh. ex Born. et Flah. in the continuous permafrost zone of Yakutia. The progress of development and distribution of potentially harmful species may have substantial effects on low-nutrient waterbodies used for drinking water handling and recreation.
The aim of this study was to characterize the ecological conditions of Gloeotrichia cf. natans vegetation in the northern river flowing through the continuous permafrost zone, to expand knowledge of the ecology of this species and to test its ability to produce toxins.
2. Materials and Methods
2.1. Study area
The study area is located in North-East Russia on the 61st parallel north in the continuous permafrost zone. The climate is extremely continental with long, severe winter and short, hot summer. The annual temperature range is of almost 100 degrees: from -60 °C in winter to +40 °C in summer [16]. The region also includes the pole of cold, located in the settlement of Oimyakon, where the lowest temperatures for the Northern Hemisphere have been recorded.
The Buotama River is a right tributary of the middle reaches of the Lena River. The length of the Buotama River is 418 km, and the basin area is 12,600 km² [17]. Mean annual discharge reaches 43 m³·s-1 [18]. The river is fed by snow and rain. It freezes from October to November, and the ice cover starts to break up from late April to early May [19]. The Buotama River flows through the Lena Pillars National Park, which is a territory under legal protection by an international convention administered by UNESCO and has been inscribed on the World Heritage List. Our research was conducted on the Buotama River, 4 km upstream from its mouth (Figure 1). The width of the river channel at this site is 200 m. The geographic coordinates of the sampling location are 61°13'39.55"N, 128°46'14.28"E (WGS 84). The depth of the river channel at a rapid river shoal is 0.3 m, at a reach section of a stream is 2.5 m, and the flow velocity is 0.8 m·s-1 and 0.3 m·s-1, respectively.
2.2. Sampling
Field material was sampled on August 16, 2021. Macrocolonies of Gloeotrichia cf. natans were collected in the lower reaches of the Buotama River on a shallow river shoal and along the riverbanks directly below the shoal. Visible colonies attached to rocks on the pebble at the bottom of the river shoal were detached from the substrate using a plastic spatula. Larger free-floating colonies, which had detached from the substrate and were carried by the current towards the shore, were collected in the immediate vicinity of this river shoal. Macrocolonies were collected in sufficient quantities in this area, from different rocks and various places along the shore, for study under a microscope. This material, in an unfixed, living state, was used for microscopic examination. Another part of this material was immediately frozen at -20°C for subsequent detection of toxins, PCR analysis and DNA extraction. Water sample for hydrochemical analysis was collected by scooping 2 L and sent to the laboratory for immediate analysis.
2.3. Algological analysis
Fragments of live macrocolonies were examined under an Olympus BH-2 (Olympus, Tokyo, Japan) light microscope at ×100 and ×400 magnifications. Photomicrographs were obtained using a CK-13 digital camera (Lomo-microsystems, Russia). Identification was carried out following J. Komárek [20].
2.4. Water chemistry analysis
Chemical analyses were performed on water sample using standard methods [21]. Water temperature was measured with an electronic thermometer Chektemp (Hanna Instruments, Woonsocket, USA). Water color was determined using a photometric method. pH was measured using a potentiometric method. Water salinity was calculated as the sum of ions using different methods: turbidimetry for sulphate anions, flame spectrophotometry for potassium and sodium cations, mercurimetry for chloride ions, and titration for calcium, magnesium, and bicarbonate ions. The hardness of water was determined by complexometric titrations using eriochrome black T as an indicator. A photometric method was applied to determine nutrients concentrations. Nessler’s reagent, Griess reagent, salicylic acid and sulfosalicylic acid were used for the measurement of ammonium ion, nitrite ion, nitrate ion and total iron, respectively; ammonium molybdate were used for the measurement of phosphate ion and silicon (Si-SiO2). A combined reagent composed of ammonium molybdate and ascorbic acid was used to determine total phosphorus content. A photometric method was applied to determine chemical oxygen demand (COD). A fluorimetric method with the Fluorat-02 device was used to measure the levels of petrochemicals and phenols. The content of manganese, nickel, copper and zinc was determined by atomic absorption spectrometry, using electrothermal vaporization.
To quality control of the analysis, the method provides repeatability limit coefficients (R), which correspond to the following values: R=2 (hardness), R=3 (Ca, HCO3), R=6 (SO4), R=8 (Cl), R=10 (Mg, Na, K), R=12 (P tot), R=13 (NH4), R=14 (NO3, PO4), R=17 (NO2), R=18 (Fe tot), R=21 (color), R=25 (COD), R =28 (Cu, Ni, Zn), R=31 (Si, Mn), R=48 (phenols), R=55 (petrochemicals) and R=0,2 (рН). The measurement result (Xmean) was taken as the arithmetic mean of two parallel detections (X1, X2), for which the following condition is satisfied: для pH; for hardness, calcium (Ca), bicarbonates (HCO3), sulfates (SO4), ammonium (NH4), nitrites (NO2), color; for magnesium (Mg), sodium (Na), potassium (K), chlorides (Cl), nitrates (NO3), silicon (SiO2), total phosphorus (P tot), phosphates (PО4), COD, iron total (Fe tot), petrochemicals, phenols, manganese (Mn), copper (Cu), nickel (Ni), zinc (Zn).
2.5. Detection of cyanotoxins by liquid chromatography–mass spectrometry
To detect cyanotoxins, the analytical-grade chemicals were used for the liquid chromatography-mass spectrometry analysis. Acetonitrile (HPLC-grade) and methanol (LiChrosolv hypergrade for LC-MS) were purchased from Merck (Darmstadt, Germany); Formic acid (98–100%) was obtained from Fluka Chemika (Buchs, Switzerland). High quality water (18.2 MΩ cm−1) was produced by the Millipore Direct-Q water purification system (Bedford, MA, USA). The MC-LR, MC-RR, MC-YR standards were purchased from Sigma Aldrich; the MC-LY, MC-LA, MC-LW, MCLF, [D-Asp3]MC-LR and [D-Asp3]MC-RR were from Enzo Life sciences, Inc. New York, USA, anatoxin-a fumarate was obtained from Tocris Bioscience (Bristol, UK). Сylindrospermopsin (CYN) and deoxy-CYN were purchased from National Research Council, Canada and Novakits, France.
The high-performance liquid chromatography – high-resolution mass-spectrometry (HPLC-HRMS) method (LTQ OrbiTrap (“Finnigan”), ESI+) was used to define the profile of toxins and quantify them.
The sample preparation procedures were performed following Chernova et al. [22], which involved the extraction of cyanotoxins from the lyophilized biomass samples through treatment with 1 ml of 75% methanol in an ultrasonic bath.
Analyses of extracts were performed using the LC-20 Prominence HPLC system (Shimadzu, Japan) coupled with a Hybrid Ion Trap-Orbitrap Mass Spectrometer – LTQ Orbitrap XL (Thermo Fisher Scientific, San Jose, USA) according to Chernova et al. [22]. Separation of the toxins was achieved on a Thermo Hypersil Gold RP C18 column (100 × 3 mm, 3 μm) with a Hypersil Gold drop-in guard column (Thermo Fisher Scientific) by gradient elution (0.2 mL min-1).
Mass spectrometric analysis was carried out under conditions of electrospray ionization in the positive ion detection mode. The identification of target compounds, whose standards were available, was based on the accurate mass measurement of [М+Н]+ or [М+2Н]2+ ions (resolution of 30000, accuracy within 5 ppm), the collected fragmentation spectrum of the ions and the retention times. The detection of AN-a was based on the presence of a peak with m/z at 166.12 and retention time corresponding to the AN-a standard. The structure of the neurotoxin was confirmed using fragmentation spectra (MS/MS). The concentrations of the toxins were calculated based on the peak area of standards analyzed on the same day and under the same conditions.
2.6. PCR analysis of cyanotoxin biosynthesis genes
DNA was isolated from the lyophilized biomass of Gloeotrichia cf. natans using the silica sorption method and a set of reagents Diatom DNA Prep 200 (Isogene Laboratory, Russia), following the manufacturer’s instructions. The study searched for four genes responsible for synthesizing microcystins, cylindrospermopsin, anatoxin-a, and saxitoxins, which are mcyE, cyrJ, anaC, and stxA, respectively. Polymerase chain reactions (PCRs) were performed with DreamTaq PCR Master Mix (Thermo Scientific, United States) in a CFX96 Touch thermal cycler (Bio-Rad, United States) using specific primers HEPF/HEPR (amplicon size 472 bp), cynsulF/cylnamR (584 bp), anaC–genF/anaC–genR (366 bp), and sxtaf/sxtar (600 bp) under amplification conditions described in the literature [23,24,25,26]. Reaction mixtures were separated by electrophoresis in 1.5% agarose gel, stained with ethidium bromide, and analyzed under UV illumination using a Gel Doc XR+ gel documenting system (Bio-Rad, United States).
2.7. DNA extraction, purification and PCR
DNA was extracted from the environmental sample with the DNeasy Plant Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol. Amplification of partial 16S rRNA gene and 16S-23S ITS rRNA region was performed with using cyanobacteria-specific primer pairs 1 suggested by [27] and 27F suggested by [28]. PCR was carried out in 20 µL volumes with MasDDTaqMIX (Dialat Ltd., Moscow, Russia), 10 mol of each oligonucleotide primer, and 1 ng of DNA with the following amplification cycles: 3 min at 94 °C, 40 cycles (30 s 94 °C, 40 s 56 °C, 60 s 72 °C) and 2 min of final extension time at 72 °C. Amplified fragments were visualized on 1% agarose TAE (mixture of Tris base, acetic acid and EDTA) gels by ethidium bromide staining, purified with the QIAquick Gel Extraction Kit (Qiagen, Hilden, Germany), and then used as a template in sequencing reactions with the ABI PRISM® BigDye™Terminator v. 3.1 (Applied Biosystems, Foster City, CA, USA) Sequencing Ready Reaction Kit following the standard protocol provided for Applied Biosystems 3730 DNA Analyzer (Applied Biosystems, Foster City, CA, USA) in “Genome” Center of EIMB (Moscow, Russia). The internal primer 2 by [29] was additionally used for sequencing.
2.8. Molecular Analyses
The single amplicon was obtained and the nucleotide sequence of partial 16S rRNA gene and 16S-23S ITS rRNA region were assembled and edited using BioEdit 7.0.1. [30]. The comprehensive set of sequences from Nostocales was collected from GenBank [31] and CyanoSeq [32]. A maximum likelihood (ML) tree was constructed using RaxML v. 1.0.0 [33]. The ModelGenerator [34] was used for the Maximum likelihood model determination, which appears to be GTR+I+G. The phylogenetic tree was graphically represented by FigTree v. 1.4.4 [35].
3. Results
3.1. Water chemistry
Water temperature during the sampling event was 17°С. Water pH was slightly alkaline (Table 1). The water of the river was fresh, with low salinity. In the major ionic composition, the predominance of bicarbonates was detected (32-36%-equiv). The concentration of chlorides and sulfates was negligible. A large proportion of calcium (45-48 %-equiv) was in the cation composition. According to major ionic constituents, the water belonged to hydrocarbonate class, calcium group, type 2-3, hardness was low. The detected concentrations of petrochemicals and phenols were negligible. Concentrations of total phosphorus and mineral phosphorus were low, below the detection limit of the analysis. The content of ammonium nitrogen, nitrate nitrogen, silica, total iron, and nickel were characterized by low values. Slightly elevated indices were noted for COD, water color index, nitrite nitrogen, manganese and zinc. The relatively high concentration of copper is of natural origin and has been often noted for the rivers of the studied region [36].
3.2. Field algological observations
For the first time, the occurrence of mass growing of cyanobacterial macrocolonies in the estuary of the Buotama River was discovered by us on August 5th, 2021. By the term "mass growing", we mean that cyanobacterial macrocolonies were found on all the stones randomly lifted from the river bottom. (Figure 2, c). Large olive-green-brown colonies of irregular spherical shape, hollow inside, with a diameter of 1 to 3 cm, were found lying freely in masses among the pebbles along the river banks (Figure 2 a,d). Further investigation showed that on the bottom of a nearby shallow (up to 0.7 m) river rapids shoal, attached to a pebble substrate, cyanobacterial colonies of the same color, irregular semi-spherical shape, and ranging in size from 1 to 10 mm, were mass growing (Figure 2 b). All colonies were dense, slimy to the touch, soft, mashed when squeezed. The colonies had a distinctive earthy odor.
Following the discovery, a field excursion was commenced on August 16th, 2021, to gather algological and hydrochemical samples from the field and transfer them to the laboratory.
3.3. Microscopic survey
The colonies of Gloeotrichia cf. natans are irregularly spherical and olive-green-brown, measuring up to 3 cm in diameter. Small young colonies are initially attached to pebbles underwater onto a river bottom, and later become free-floating and washed onto the river bank. Larger colonies are hollow. The colonies are composed of densely radially arranged filaments with bases oriented in the center of a colony and embedded in a mucilage matrix (Figure 2 e). The colonial slime is firm with periderm and delimited. Sheats are vase-like, widened, colorless to yellowish, thin, widened at the base, and distant from the trichomes. They are scarcely transversely constricted (Figure 2 f,g, denoted with blue arrows). Trichomes with constricted cross-walls at the base and are not constricted away from a base (Figure 2 h, denoted with green arrows). Thichomes are heteropolar with widened base and other ends of trichomes are elongated into long, cellular hairs. Young cells are cylindrical, longer than wide, later barrel-shaped at the base of trichomes, and slightly shorter than wide up to isodiametric. Cells are olive-green without aerotopes, 5-6 µm wide at the base. Heterocytes are basal, single, and sphaerical (Figure 2 f-h, denoted with black arrows). Akinetes are cylindrical with rounded ends (Figure 2 g, denoted with white arrow), straight or arcuated (Figure 2 e, denoted with red arrows), 30-50 x 6-8 µm. Although our specimen morphologically coincided with Gloeotrichia natans this species is a variable with numerous forms and its robust identification should be confirmed genetically.
Figure 2.
Macrocolonies of Gloeotrichia cf. natans observed with no magnification, lying freely among pebbles on the riverbank (a); colonies attached to rocks underwater (b), and on a rock lifted out of the water (c); olive-green-brown colonies, that are irregularly spherical and hollow, with diameters ranging from 1 to 3 cm, collected along the riverbank among the pebbles (d). Light microscopy of Gloeotrichia natans reveals a part of the colony consisting of densely radially arranged filaments, with straight or arcuated akinetes visible (e, denoted with red arrows); basal heterocytes that are single and spherical (f-h, denoted with black arrows), cylindrical akinetes with rounded ends (g, denoted with white arrow); vase-like widened sheaths that are colorless to yellowish, thin, widened at the base, distant from the trichomes, and scarcely transversely constricted (f,g, denoted with blue arrows); trichomes with constricted cross-walls at the base, and not constricted away from the base (h, denoted with green arrows); young cells that are cylindrical, longer than wide, and later barrel-shaped at the base of trichomes (h).
Figure 2.
Macrocolonies of Gloeotrichia cf. natans observed with no magnification, lying freely among pebbles on the riverbank (a); colonies attached to rocks underwater (b), and on a rock lifted out of the water (c); olive-green-brown colonies, that are irregularly spherical and hollow, with diameters ranging from 1 to 3 cm, collected along the riverbank among the pebbles (d). Light microscopy of Gloeotrichia natans reveals a part of the colony consisting of densely radially arranged filaments, with straight or arcuated akinetes visible (e, denoted with red arrows); basal heterocytes that are single and spherical (f-h, denoted with black arrows), cylindrical akinetes with rounded ends (g, denoted with white arrow); vase-like widened sheaths that are colorless to yellowish, thin, widened at the base, distant from the trichomes, and scarcely transversely constricted (f,g, denoted with blue arrows); trichomes with constricted cross-walls at the base, and not constricted away from the base (h, denoted with green arrows); young cells that are cylindrical, longer than wide, and later barrel-shaped at the base of trichomes (h).

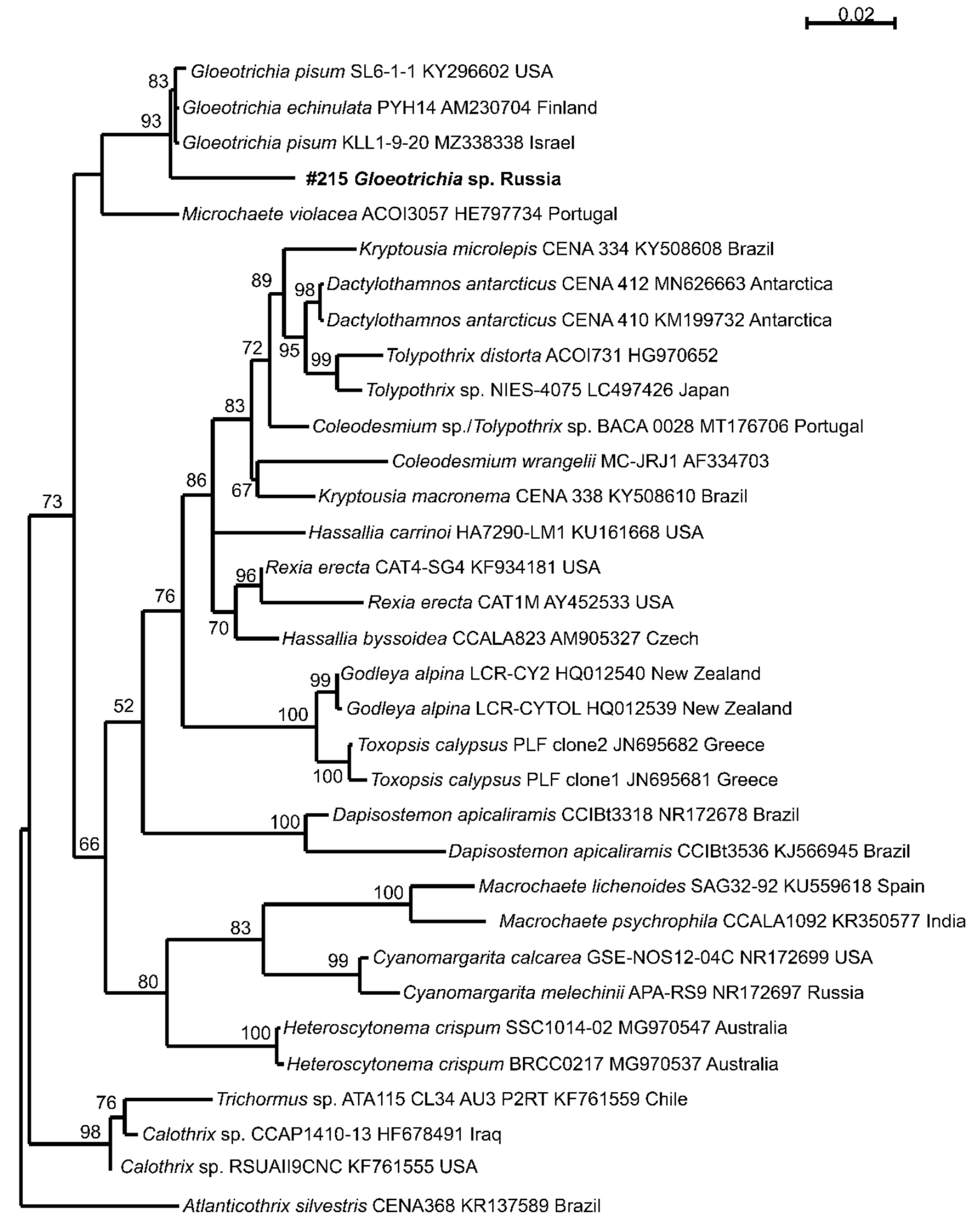
3.4. Phylogeny
The identification of cyanobacteria is challenging due to their high morphological variability. Obtained 16S rRNA gene of Buotama river environmental sample was assessment with Nostocales type strains of cyanobacterial CyanoSeq database sequences [32]. The studied sample is clustering with three Gloeotrichia-species, but forms a long separate branch. Therefore, our sample of Gloeotrichia cf. natans has low similarity in 16S rRNA gene sequence with G. pisum, and G. echinulata. Unfortunately, public databases are not containing a sequence of G. natans and we cannot evidence that our cyanobacterium belongs to this species.
In the future we plan to conduct a study aimed at definitively establishing the species classification of this particular sample. Perhaps we will able to get herbarium data of specifically to Gloeotrichia natans.
Figure 3.
Phylogram obtained under the maximum likelihood approach for 33 accession of Nostocales based on the 16S rRNA gene with Atlanticothrix silvestris as the outgroup. Bootstrap support values from the maximum likelihood more than 50% are indicated.
Figure 3.
Phylogram obtained under the maximum likelihood approach for 33 accession of Nostocales based on the 16S rRNA gene with Atlanticothrix silvestris as the outgroup. Bootstrap support values from the maximum likelihood more than 50% are indicated.
3.5. Cyanotoxins detection
No anatoxin-a, cylindrospermopsin, or any variants of microcystins were detected in the biomass of Gloeotrichia cf. natans above detection limit of 0.1mkg g-1. Furthermore, the absence of the ability of Gloeotrichia cf. natans to produce cyanobacterial hepatotoxins and neurotoxins was confirmed and supplemented by the negative results of PCR amplification of of mcyE, anaC, cyrJ and sxtA genes, which is consistent with the LC/MS data.
4. Discussion
Gloeotrichia natans is a freshwater cyanobacterium that is widely distributed across different regions of the world (Figure 4). The most of the occurrences are warm climate regions, and a temperate zone. The exceptions are locations in Komi Republic [37] and the mouth of Lena river [38].
The occurrences of Gloeotrichia natans have been record in Yakutia previously. The discovery of this species in Central Yakutia was reported by L. Ye. Komarenko and I. I. Vasilyeva [41] (Lake Tyungulu) and L. Kopyrina et al. [42] (Lake Dyiere) (Figure 3). V. I. Zakharova et al. [43] reported on the findings of Gloeotrichia natans in the basins of the Lena and Aldan rivers, without definitely location.
A few records this species from the Arctic and Subarctic area has been listed in the open information system on biodiversity “L.” [39,44]. It is an occurrence of Gloeotrichia natans in a lake on the Samoylovsky island in the Lena River Delta [38], and in an Unnamed peat lake in Kamchatka Peninsula [45]. On the territory of Asian Russia, the species is more common in southern areas and was noted in Primorye Region, Khabarovskyi Region, Amurskaya Oblast, Sakhalinskaya Oblast [46]. It is known that such a large river crossing the continent in the meridional direction, like the Lena River, is a transit corridor along which microalgae significantly expand their range to the North [47]. The records of G. natans in Yakutia are belonged to the area along the Lena River (Figure 4), which, in our opinion, confirms the hypothesis about the transfer of this species from the southern regions to the north along the course of this large river.
As far as we managed to find out, the mass growing of colonies of this species in the continuous permafrost zone was not noted earlier. The occurrence of Gloeotrichia natans in the Buotama River has not been reported previously, therefore information on when this species appeared here is absent.
To date, most research on cyanobacterial blooms has focused on high-nutrient lakes. Gloeotrichia natans is known to thrive in eutrophic waters also. We recorded the cyanobacterium Gloeotrichia cf. natans in the river with low concentrations of nitrogen and phosphorus.
The data we obtained indicate high water quality of Buotama River. According to the classification of stream trophic state suggested by W. K. Dodds [48] the Buotama River is classified as mesotrophic based on its total phosphorus (Ptot) content and oligotrophic based on its total nitrogen (Ntot) content. The main cause of high water quality might be that the river flows through a protected area where the influence of anthropogenic factors on the water ecosystem is excluded.
Despite the oligotrophic rate of the water course in low-nutrients systems occasionally exhibited high levels of cyanobacteria.
Over the past few decades, there has been a growing trend of abnormal development of filamentous cyanobacteria in oligotrophic water bodies in various parts of the world where such occurrences were once non-existent. Scientists have documented instances of extensive proliferation of benthic filamentous cyanobacteria in shallow and pristine water bodies with high water quality, despite the absence of eutrophication indicators [49]. Among the reasons for this phenomenon, the increase in water temperature and the lengthening of the vegetation season are cited, which are associated with modern global climate change [49]. It is known that an increase in water temperature stimulates the growth of filamentous cyanobacteria, which form mats and biofilms on the surfaces of submerged objects [50].
Global climate change has led to changes in the ice regime of water bodies in the studied region. From 1980 to 2014, there was an increase in the duration of the ice-free period at various water bodies in Asian Russia, with an average increase of 4.63 to 11 days per decade [51]. The shortening of the ice-free period leads to an extension of the vegetation season in the water bodies of the region. Therefore, the observed phenomenon of massive proliferation of benthic cyanobacteria may be associated with global climate change. In any case, we believe that this phenomenon requires further study and monitoring in the northern water bodies, particularly in the water bodies of Yakutia.
Gloeotrichia natans is commonly found in a variety of aquatic habitats, including lakes, ponds, rivers, and streams. It can grow in a range of environmental conditions. Its typical habitats are stagnant waters, where it usually grows as an epiphyte on water plants. [20,41,52,53]. We found Gloeotrichia cf. natans in a river where aquatic plants are absent, and it was developing epilithically. We did not find any published data on findings of this species in lotic environments or epilithic habitats. Thus, our discovery expands knowledge of the ecology of the species.
Cyanobacteria produce a wide range of metabolites. Among them non-toxic odorants and highly toxic metabolites – cyanotoxins. The production of geosmin – the odorant, causing a characteristic earthy smell – was noted organoleptically. According to F. Jüttner and S. B. Watson [54], the geosmin odor detection threshold in humans is very low, ranging from 0.006 to 0.01 micrograms per liter in water. Thus, the massive proliferation of the species could affect the quality of recreational waters. It is known that cyanotoxin-producing species are widely distributed in benthic habitats, with benthic anatoxins being recorded in 8 countries and benthic microcystins in 10 countries [5]. In the past two decades, have been published several studies confirming the distribution of cyanobacterial blooms and occurrences of toxic metabolites in benthic habitats of polar and subpolar regions [55,56,57,58,59,60]. Our recent studies of plankton in lakes of Yakutia made it possible, for the first time in the cryolithozone of northeastern Russia, to detect the presence of the mcyE gene responsible for the biosynthesis of microcystin cyanotoxins, as well as several structural variants of microcystins [61]. This confirms the possibility of cyanobacteria producing toxic metabolites in the harsh climatic conditions of the region, in water bodies located in the continuous permafrost zone. Therefore, we decided to study cyanotoxin production by Gloeotrichia cf. natans colonies.
The most well-studied toxic species of the genus Gloeotrichia is G. echinulata. This is the only species of this genus that vegetates in plankton, the ability of which to produce cyanotoxins has been known for a long time [10,11]. Its produces a low concentration of microcystin-LR (MC-LR) [10], which can have adverse effects on plankton, fish, as well as humans, livestock, and pets [11].
Cantoral Uriza et al. [13] report on the toxigenicity of G. natans. The authors conducted extensive sampling throughout Spain, under a range of conditions and in distinct aquatic and terrestrial habitats. Colonies of G. natans were found by the researchers in Lagoa Vixán, Corrubedo, where the species vegetated epiphytically. A strain of G. natans was isolated from the colonies, in which microcystins MC-LF and MC-RR were detected.
It is known that different strains of the same species can either exhibit toxicity or be non-toxic. Additionally, representatives of the same species can produce different types of toxins depending on their geographic locations [5].
Microcystins and anatoxins were not detected in the colonies of Gloeotrichia cf. natans found by us using LC/MS method. Genes responsible for the synthesis of hepatotoxins (microcystins and cylindrospermopsin) and neurotoxins (anatoxin-a and saxitoxins) were also not detected in the studied samples. Therefore, the mass proliferation of G. cf. natans in this region is unlikely to create risks for other aquatic inhabitants and does not pose a threat to human health. However, some difficulties in understanding toxic productivity in benthic habitats treat to detecting the environmental conditions that promote the proliferation of toxic cyanobacteria, the nature of their temporal dynamics and life cycle, and the effects of environmental variables [5]. We cannot rule out the possibility that other populations of this species will not exhibit toxicity, or that the studied population, through horizontal transfer, will not acquire toxin-producing genes in the future. So, in our opinion, in the context of global climate change and the lengthening of the growing season in Arctic water bodies, such occurrences of mass proliferation of potentially toxic cyanobacterial species should be constantly monitored.
5. Conclusions
This study, for the first time in the water bodies located in the zone of continuous permafrost, revealed the massive proliferation of macrocolonies of the potentially toxic cyanobacterium Gloeotrichia cf. natans. Molecular and analytical methods were used to establish that the studied species Gloeotrichia cf. natans did not produce hepatotoxins and neurotoxins, but we were able to expand knowledge about the ecology of the species and its ability to vegetate epilithically in lotic habitats. Our discovery of Gloeotrichia cf. natans is not the first for the region, but it has not been previously recorded in the studied Buotama River. The waters of the Buotama River are characterized by high quality and low trophic status. In recent decades, in different regions of the world, massive proliferation of benthic cyanobacteria has been observed in oligotrophic water bodies, which is partly attributed to global climate change and an increase in the vegetation period. For water bodies in the northern region we studied, the ice-free period has been decreasing over the past several decades, leading to an increase in the length of the vegetation period. Under these conditions, the occurrence of massive proliferation of benthic cyanobacteria should be closely monitored.
Supplementary Materials
Not applicable.
Author Contributions
Conceptualization, V.G. and D.D.; methodology, V.G. and D.D.; software, D.D. and A.V.; validation, S.B., E.Ch., S.S.; formal analysis, V.G., S.S.; investigation, V.G., O.G.; resources, O.G.; data curation, V.G. and D.D.; writing—original draft preparation, V.G., D.D. and A.V.; writing—review and editing, V.G., S.S, E.Ch. and D.D..; visualization, Z.Zh.; supervision, V.G.; project administration, V.G.; funding acquisition, V.G., D.D., S.S., E.Ch. All authors have read and agreed to the published version of the manuscript.
Funding
The research was carried out within the state assignment of the Ministry of Science and Higher Education of the Russian Federation (theme No. FWRS-2021-0023, reg. No. АААА-А21-121012190038-0), (theme No. 121051100099-5) and within the framework of state budgetary theme no. FFZF-2022-0012, RosRid no. 122041100086-5. The study of molecular identification of specimen was funded by the Russian Science Foundation, grant number 21-14-00029 (https://rscf.ru/project/21-14-00029/). Molecular genetic studies were performed by S.I. Sidelev in the scientific educational laboratory "Molecular genetics and biotechnology" under YarSU’s development program framework for 2030 (№ 123042800011-6).
Data Availability Statement
Not applicable.
Acknowledgments
This work was partly supported by the Israeli Ministry of Aliyah and Integration.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: risk management for health protection. Toxicology and Applied Pharmacology 2005, 203, 264–272. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environmental Microbiology 2009, 1, 27–37. [Google Scholar] [CrossRef]
- Neil, J.M.O.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 313–334. [Google Scholar] [CrossRef]
- Gaget, V.; Hobson, P.; Keulen, A.; Newton, K.; Monis, P.; Humpage, A.; Weyrich, L.; Brookes, J. Toolbox for the sampling and monitoring of benthic cyanobacteria. Water Research 2020, 169, 115222. [Google Scholar] [CrossRef]
- Wood, S.; Kelly, L.; Bouma-Gregson, K.; Humbert, J.; Laughinghouse, H.; Lazorchak, J.; McAllister, T.; McQueen, A.; Pokrzywinski, K.; Puddick, J.; Quiblier, C.; Reitz, L.; Ryan, K.; Vadeboncoeur, Y.; Zastepa, A.; Davis, T. Toxic benthic freshwater cyanobacterial proliferations: Challenges and solutions for enhancing knowledge and improving monitoring and mitigation. Freshwater Biology 2020, 65, 1824–1842. [Google Scholar] [CrossRef]
- Heino, J.; Virkkala, R.; Toivonen, H. Climate change and freshwater biodiversity: detected patterns, future trends and adaptations in northern regions. Biological Reviews 2009, 84, 39–54. [Google Scholar] [CrossRef]
- Winder, M.; Sommer, U. Phytoplankton response to a changing climate. Hydrobiologia 2012, 698, 5–16. [CrossRef]
- Korneva, L.G. Invasions of alien species of planktonic microalgae into the fresh waters of Holarctic (Review). Russian Journal of Biological Invasions 2014, 5, 65–81. [Google Scholar] [CrossRef]
- Lang-Yona, N.; Alster, A.; Cummings, D.; Freiman, Z.; Kaplan-Levy, R.; Lupu, A.; Malinsky-Rushansky, N.; Ninio, S.; Sukenik, A.; Viner-Mozzini, Y.; Zohary, T. Gloeotrichia pisum in Lake Kinneret: A successful epiphytic cyanobacterium. J. Phycol. 2023, 59, 97–110. [Google Scholar] [CrossRef]
- Carey, C.C.; Haney, J.F.; Cottingham, K.L. First report of microcystin-LR in the cyanobacterium Gloeotrichia echinulata 2007, 22, 337–339. [CrossRef]
- Carey, C.C.; Ewing, H.A.; Cottingham, K.L. Occurrence and toxicity of the cyanobacterium Gloeotrichia echinulata in low-nutrient lakes in the northeastern United States. Aquatic Ecology 2012, 46, 395–409. [Google Scholar] [CrossRef]
- Stamann, S. "Toxin Production and Population Dynamics of Gloeotrichia echinulata with Considerations of Global Climate Change". Masters Theses 2015, 775. http://scholarworks.gvsu.edu/theses/775.
- Cantoral Uriza, E.A.; Asencio, A.D.; Aboal, M. Are We Underestimating Benthic Cyanotoxins? Extensive Sampling Results from Spain. Toxins 2017, 9, 385. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. Available online: https://www.algaebase.org (accessed on 29 March 2023).
- Strunecky, O.; Ivanova, A. An updated classification of cyanobacterial orders and families based on phylogenomic and polyphasic analysis. Journal of Phycology 2023, 59, 12–51. [Google Scholar] [CrossRef]
- Izyumenko, S.A. Climate of the Yakut ASSR (atlas). Ed. Leningrad: Hydrometeorological Publishing House, Russia, 1968; 32 p. (In Russian).
- State Water Register. Available online: http://textual.ru/gvr/index.php?card=243859 (accessed on 23 March 2023). (In Russian).
- Surface water resources of the USSR. Lensko-Indigirsky district. Leningrad: Gidrometeoizdat, USSR, 1972; 651 p. (In Russian).
- Chistyakov G. Ye. Water resources of the rivers of Yakutia; Moscow: Nauka, Russia, 1964; 255 p. (In Russian).
- Komárek, J. Heterocytous Genera. Cyanoprokaryota; Springer Spektrum, Berlin, 2013; 3 (3), 3–1130.
- Semenov, A.D. (Ed.) Guidance on the chemical analysis of surface waters of the land; Gidrometeoizdat: Leningrad, USSR, 1977; 541 p. (In Russian) [Google Scholar]
- Chernova, E. Chernova, E., Russkikh, Ia., Voyakina, E., Zhakovskaya, Z. Occurrence of microcystins and anatoxin-a in eutrophic lakes of Saint Petersburg, Northwestern Russia. Oceanol. Hydrobiological Stud. 2016, 45 (4), 466–484. [CrossRef]
- Jungblut, A.D.; Neilan, B.A. Molecular identification and evolution of the cyclic peptide hepatotoxins, microcystin and nodularin, synthetase genes in three orders of cyanobacteria. Arch. Microbiol. 2006, 185, 107–114. [Google Scholar] [CrossRef]
- Mihali, T.K.; Kellmann, R.; Muenchhoff, J.; et al. Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis, Applied and Environmental. Microbiology 2008, 74, 716–722. [Google Scholar]
- Ballot, А.; Fastner, J.; Wiedner, C. Paralytic shellfish poisoning toxin-producing cyanobacterium Aphanizomenon gracile in Northeast Germany. Appl. Environ. Microbiol. 2010, 76, 1173–1180. [Google Scholar] [CrossRef]
- Rantala‒Ylinen, A.; Känä, S.; Wang, H.; Rouhiainen, L.; Wahlsten, M.; Rizzi, E.; Berg, K.; Gugger, M.; Sivonen, K. Anatoxin–a synthetase gene cluster of the cyanobacterium Anabaena sp. strain 37 and molecular methods to detect potential producers. Appl. Environ. Microbiol. 2011, 77, 7271–7278. [Google Scholar] [CrossRef]
- Wilmotte, A.; Van Der Auwera, C.; De Wachter, R. Structure of the 16S ribosomal RNA of the thermophilic cyanobacteria Chlorogloeopsis HTF (‘Mastigocladus laminosus HTF’) strain PCC7518 and phylogenetic analysis. FEBS Lett. 1993, 317, 96–100. [Google Scholar] [CrossRef]
- Neilan, B.A.; Jacobs, D.; Therese, D.D.; Blackall, L.L.; Hawkins, P.R.; Cox, P.T.; Goodman, A.E. rRNA Sequences and Evolutionary Relationships among Toxic and Nontoxic Cyanobacteria of the Genus Microcystis. Int. J. Syst. Bacteriol. 1997, 47, 693–697. [Google Scholar] [CrossRef]
- Nübel, U.; Garcia-Pichel, F.; Muyzer, G. PCR primers to amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symposium Series 1999, 41, 95–98. [Google Scholar]
- Benson, D.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.; Ostell, J. Sayers EW. GenBank. Nucleic Acids Res. 2013, 41, 36–42. [Google Scholar] [CrossRef]
- Lefler, F.; Berthold, D. Dail Laughinghouse H 4th. CyanoSeq: a database of cyanobacterial 16S rRNA sequences with curated taxonomy. J Phycol. [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable, and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef]
- Keane, T.M.; Creevey, C.J.; Pentony, M.M.; Naughton, T.J.; Mclnerney, J.O. Assessment of methods for amino acid matrix selection and their use on empirical data shows that ad hoc assumptions for choice of matrix are not justified. BMC Evolutionary Biology 2006, 6(1), 29. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree Version 1.4.4. 2020. (Default Version). Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 15 May 2023).
- Gabyshev, V.A. Gabyshev V.A., Gabysheva O.I. Phytoplankton of the largest rivers of Yakutia and adjacent territories of Eastern Siberia; Novosibirsk: SibAk, Russia, 2018; 414 p. (In Russian).
- Getzen, M.V. Algae of Pechora river basin. Leningrad: Nauka, USSR, 1973; 148 pp. (In Russian).
- Kiselev, G.A.; Balashova, N.B.; Ulanova, A.A. Ecological characteristics of algae from the lakes of Samoilovsky Island (Lena Delta Nature Reserve, Russia). In Proceedings of the IV International conference «Advances in modern phycology», Kyiv, Ukraine, 23–25 May 2012. (In Russian). [Google Scholar]
- Melekhin, A.V.; Davydov, D.A.; Borovichev, E.A.; Shalygin, S.S.; Konstantinova, N.A. CRIS – service for input, storage and analysis of the biodiversity data of the cryptogams. Folia Cryptogamica Estonica 2019, 56, 99–108. [Google Scholar] [CrossRef]
- GBIF.org. GBIF Occurrence (07 April 2023). [CrossRef]
- Komarenko, L.Ye.; Vasilyeva, I.I. Freshwater diatoms and blue-green algae of water bodies of Yakutia. Moscow: Nauka, Russia, 1975. 424 p. (In Russian).
- Kopyrina, L.; Pshennikova, P.; Barinova, S. Diversity and ecological characteristic of algae and cyanobacteria of thermokarst lakes in Yakutia (northeastern Russia). International Journal of Oceanography and Hydrobiology 2020, 49, 99–122. [Google Scholar] [CrossRef]
- Zakharova, V. I.; Kuznetsova, L. V.; Ivanova, Ye. I. et al. Diversity of the flora of Yakutia; Novosibirsk: Publishing house of the Siberian Branch of the Russian Academy of Sciences, Russia, 2005; 328 p. (In Russian).
- CYANOpro. Available online: http://kpabg.ru/cyanopro/?q=node/96073 (accessed on 23 March 2023).
- Elenkin, A.A. Kamchatka expedition of Fyodor Pavlovich Ryabushinsky. Botanical department. Issue. 2. Spore plants of Kamchatka: 1) Algae, 2) Mushrooms. Printing house P.P. Ryabushinsky: Moscow, Russia, 1914, 1–402. (In Russian).
- Medvedeva, L.A., Nikulina, T.V. Catalogue of freshwater algae of the southern part of the Russian Far East; Vladivostok: Dalnauka, Russia, 2014; 271 p. (In Russian).
- Gabyshev, V. A.; Tsarenko, P. M.; Ivanova A., P. Diversity and features of the spatial structure of algal communities of water bodies and watercourses in the Lena River estuary. Inland water biology 2019, 12(1), S1–S9. [Google Scholar] [CrossRef]
- Dodds, W. K. Trophic state, eutrophication and nutrient criteria in streams. Trends in Ecology & Evolution 2007, 22, 669–676. [Google Scholar] [CrossRef]
- Vadeboncoeur, Y.; Moore, M.; Stewart, S.; Chandra, S.; Atkins, K.; Baron, J.; Bouma-Gregson, K.; Brothers, S.; Francoeur, S.; Genzoli, L.; Higgins, S.; Hilt, S.; Katona, L.; Kelly, D.; Oleksy, I.; Ozersky, T.; Power, M.; Roberts, D.; Smits, A.; Timoshkin, O.; Tromboni, F.; Zanden, M.; Volkova, E.; Waters, S.; Wood, S.; Yamamuro, M. Blue Waters, Green Bottoms: Benthic Filamentous Algal Blooms Are an Emerging Threat to Clear Lakes Worldwide. Bioscience 2021, 71, 1011–1027, PMID:34616235; PMCID: PMC8490932. [Google Scholar] [CrossRef]
- DeNicola, D. M. Periphyton responses to temperature at different ecological levels. In Algal ecology: Freshwater benthic ecosystem; San Diego, USA: Academic press, USA, 1996. [Google Scholar]
- Vuglinsky, V.; Valatin, D. Changes in Ice Cover Duration and Maximum Ice Thickness for Rivers and Lakes in the Asian Part of Russia. Natural Resources 2018, 9, 73–87. [Google Scholar] [CrossRef]
- Gollerbakh, M.M.; Kosinskaya, Ye.K.; Polyanskiy, V.I. Blue green algae. Identification guide to freshwater algae of the USSR. Moscow: Nauka, USSR, 1953; 650 p. (In Russian).
- Starmach, K. Cyanophyta – Sinice. Glaucophyta – Glaukofity. Flora Słodkowodna Polski; Warszawa: PWN, Poland, 1966; 2, 808 p. (in Polish).
- Jüttner, F.; Watson, S. Biochemical and Ecological Control of Geosmin and 2-methylisoborneol in Source Waters. Applied and Environmental Microbiology 2007, 73(14), 4395–4706. [Google Scholar] [CrossRef]
- Hitzfeld, B. C.; Lampert, C. S.; Spaeth, N.; Mountfort, D.; Kaspar, H.; Dietrich, D. Toxin production in cyanobacterial mats from ponds on the McMurdo ice shelf, Antarctica. Toxicon 2000, 38, 1731–1748. [Google Scholar] [CrossRef]
- Jungblut, A. D.; Hoeger, S. J.; Mountfort, D.; Hitzfeld, B.; Dietrich, D.; Neilan, B. Characterization of microcystin production in an Antarctic cyanobacterial mat community. Toxicon 2006, 47, 271–278. [Google Scholar] [CrossRef]
- Wood, S.; Mountfort, D.; Selwood, A.; Holland, P.; Puddick, J.; Cary, S. Widespread distribution and identification of eight novel microcystins in Antarctic cyanobacterial mats. Appl. Environ. Microbiol. 2008, 74, 7243–7251. [Google Scholar] [CrossRef]
- Kleinteich, J.; Wood, S.; Küpper, F.; Camacho, A.; Quesada, A.; Frickey, T.; Dietrich, D. Temperature-related changes in polar cyanobacterial mat diversity and toxin production. Nat. Clim. Chang. 2012, 2, 356–360. [Google Scholar] [CrossRef]
- Kleinteich, J.; Hildebrand, F.; Wood, S.; Cirés, S.; Agha, R.; Quesada, A.; Pearce, D.; Convey, P.; Küpper, F.; Dietrich, D. Diversity of toxin and non-toxin containing cyanobacterial mats of meltwater ponds on the Antarctic Peninsula: A pyrosequencing approach. Antarct. Sci. 2014, 26, 521–532. [Google Scholar] [CrossRef]
- Chrapusta, E.; Wegrzyn, M.; Zabaglo, K.; Kaminski, A.; Adamski, M.; Wietrzyk, P.; Bialczyk, J. Microcystins and anatoxin-a in Arctic biocrust cyanobacterial communities. Toxicon 2015, 101, 35–40. [Google Scholar] [CrossRef]
- Gabyshev, V. A.; Sidelev, S. I.; Chernova, E. N.; Gabysheva, O. I.; Voronov, I. V.; Zhakovskaya Z., A. Limnological characterization and first data on the occurrence of toxigenic cyanobacteria and cyanotoxins in the plankton of some lakes in the permafrost zone (Yakutia, Russia). Contemporary Problems of Ecology 2023, 16, 89–102. [Google Scholar] [CrossRef]
Figure 1.
The study area. Location of the sampling point at the Buotama River section (a). Map of the study area with a red dot indicating the sampling point and a green line representing the border of the Lena Pillars National Park (b). Geographic location of the study area (red dot), with a red line showing the administrative border of Yakutia and a black dashed line indicating the polar circle (c).
Figure 1.
The study area. Location of the sampling point at the Buotama River section (a). Map of the study area with a red dot indicating the sampling point and a green line representing the border of the Lena Pillars National Park (b). Geographic location of the study area (red dot), with a red line showing the administrative border of Yakutia and a black dashed line indicating the polar circle (c).
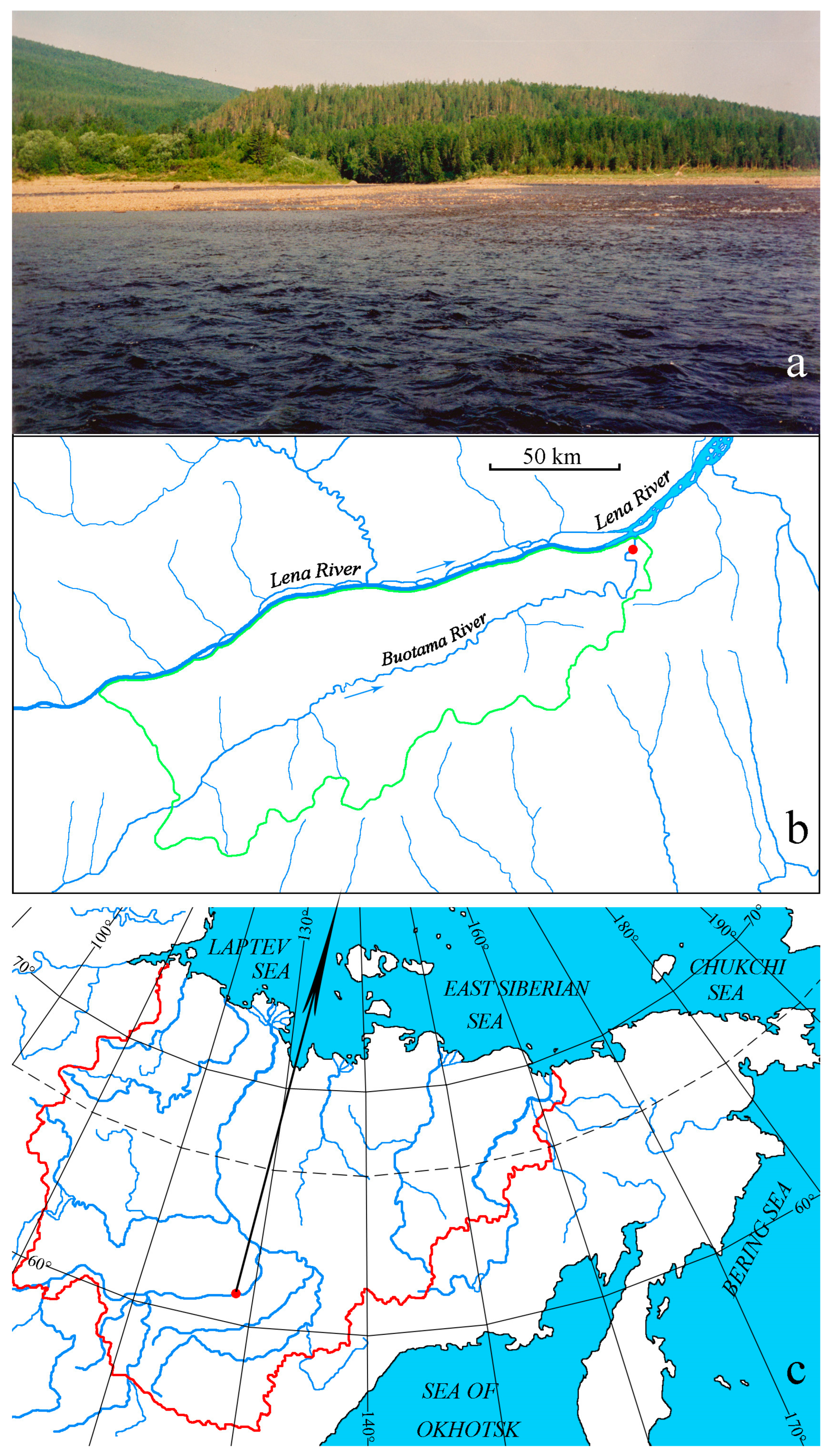
Figure 4.
Distribution of Gloeotrichia natans according to information system “L” [39] and GBIF.org [40] data.
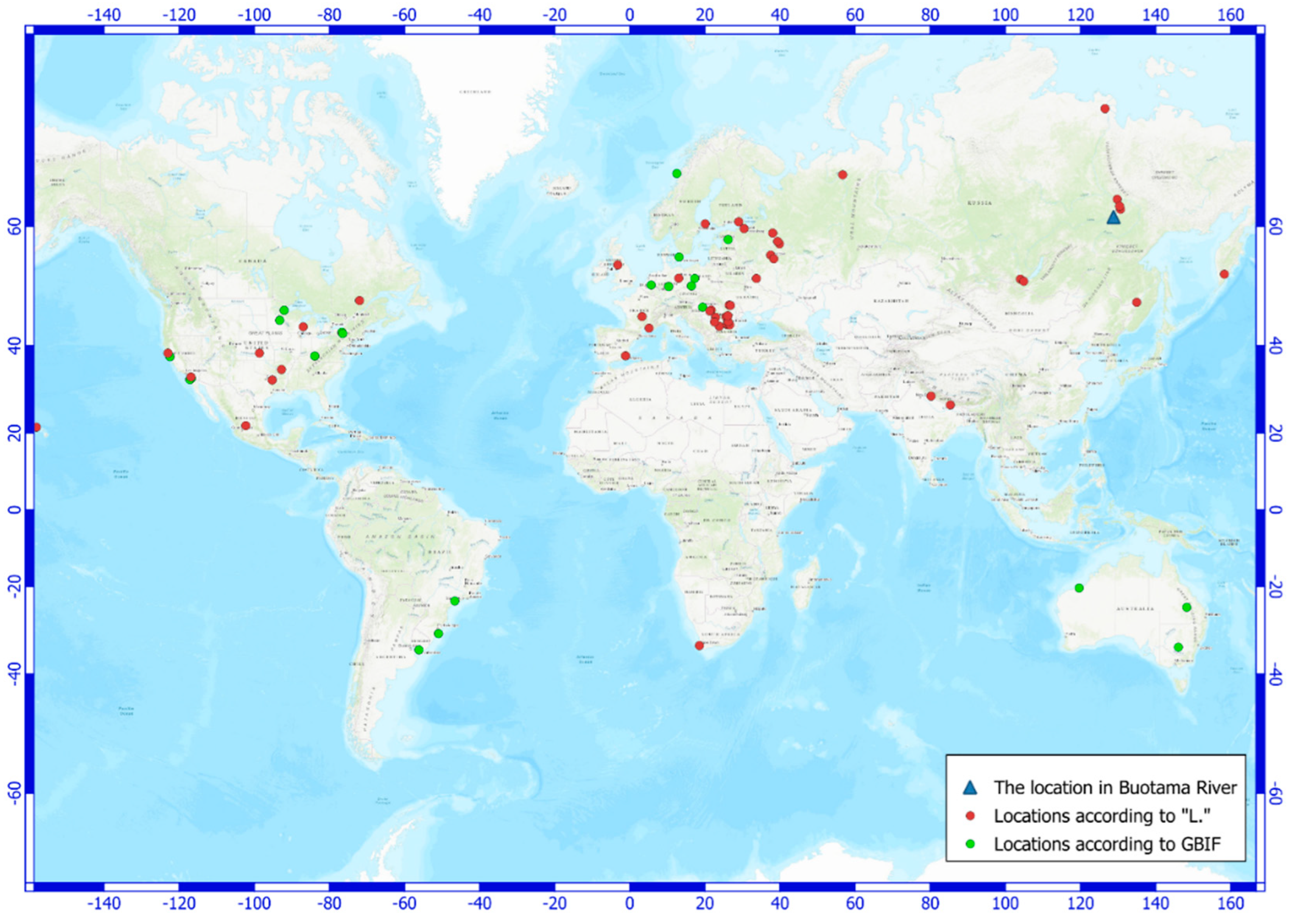

Table 1.
Averaged chemical variables with standard deviations of Buotama River water.
| Variable | Mean value | Standard deviation |
|---|---|---|
| pH | 8.47 | 0.0354 |
| TDS, mg L-1 | 185.76 | 1.8385 |
| Hardness, mmol. L-1 | 2.40 | 0.0141 |
| Ca, mg L-1 | 45.69 | 0.2687 |
| Mg, mg L-1 | 1.46 | 0.0566 |
| Na, mg L-1 | 1.19 | 0.0707 |
| K, mg L-1 | 0.58 | 0.0283 |
| HCO3, mg L-1 | 109.84 | 0.7071 |
| Cl, mg L-1 | 14.18 | 0.4243 |
| SO4, mg L-1 | 12.82 | 0.2828 |
| N-NH4, mg L-1 | 0.13 | 0.0071 |
| N-NO3, mg L-1 | 0.03 | 0.0071 |
| N-NO2, mg L-1 | 0.03 | 0.0 |
| Si-SiO2, mg L-1 | 2.04 | 0.2828 |
| P tot, mg L-1 | < 0.04 | 0.0028 |
| PО4, mg L-1 | < 0.02 | 0.0028 |
| Color, Pt/Co grad. | 44 | 4.2426 |
| COD, mg O L-1 | 20.20 | 2.1213 |
| Fe tot, mg L-1 | < 0.05 | 0.0028 |
| Petrochemicals, mg L−1 | 0.005 | 0.0014 |
| Phenols, mg L−1 | 0.0005 | 0.0001 |
| Mn, µg L-1 | 33.00 | 5.6569 |
| Cu, µg L-1 | 6.00 | 0.7071 |
| Ni, µg L-1 | 4.40 | 0.7071 |
| Zn, µg L-1 | 13.70 | 1.1314 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



