
Submitted:
26 May 2023
Posted:
29 May 2023
You are already at the latest version
Abstract
Tryptophan is an essential nutrient required to produce niacin (vitamin B3), which is needed for energy metabolism and DNA production. Alterations in tryptophan metabolism could have significant effects on aging and musculoskeletal health. The kynurenine pathway, the main catabolic route of tryptophan, is modulated by inflammatory factors, which are increased in older persons, a process known as inflammaging. Osteoporosis, sarcopenia, osteosarcopenia and frailty have also been linked with chronic increased levels of inflammatory factors. Due to the disruption of the kynurenine pathway by chronic inflammation and/or changes in the gut microbiota, serum levels of toxic kynurenines are increased and have been associated with osteoporosis, sarcopenia and frailty. In contrast, anabolic end products of this pathway, such as picolinic acid, have demonstrated a beneficial effect on bone and muscle. In addition, recent studies have shown that physical activity can modulate this pathway by promoting the secretion of anabolic kynurenines. According to the evidence collected, kynurenines could have a promising role as biomarkers for osteoporosis sarcopenia, osteosarcopenia and frailty in older persons. In addition, some of these metabolites could become important targets for developing new pharmacological treatments for these conditions.
Keywords:
Tryptophan
; Kynurenine pathway
; Aging
; older adults
; Inflammation
; Osteoporosis
; Sarcopenia
; Frailty
; Biomarkers
; Osteosarcopenia
1. Introduction
Among the 20 natural amino acids that play a fundamental role in animal nutrition and health, Tryptophan (Trp) must be highlighted. It is an apolar, essential, α-amino acid that must be provided by diet, serving as a precursor for several metabolites which play critical roles in body homeostasis [1,2]. Most Trp is metabolized in the liver, and the remaining Trp, as well as its liver degradation products, are either distributed to the peripheral circulation or transported to tissues such as skeletal muscle and the central nervous system. Trp not taken up by the upper gastrointestinal tract is metabolized by resident microbiota [3,4].
Concerning Trp metabolism, the best-known pathway is its conversion to 5-hydroxytryptamine (serotonin), a central nervous system neurotransmitter mainly involved in controlling adaptive, mood, and cognition [5]. Since serotonin can be converted to melatonin and N-acetyl serotonin, another role of Trp metabolites is maintaining circadian rhythmicity [6]. However, most free Trp is degraded through the kynurenine pathway (KP) [7]. The KP produces biologically active metabolites known as kynurenines, which have been related to immune response, inflammation and excitatory neurotransmission due to their relevant role in mitochondrial energy production and many enzymatic redox reactions.
According to recent evidence, disturbed tryptophan catabolism through KP could play an important role in the pathogenesis of several age-associated diseases, particularly neurodegenerative, autoimmune and cardiovascular diseases [8,9,10,11,12]. Recently, research efforts have been oriented to evaluate the relationship between KP and osteoporosis, sarcopenia, osteosarcopenia and frailty in older adults with promising results. This review aims to associate the complex interplay between them, gathering the most relevant evidence to date.
1.1. Kynurenine pathway, the main route for tryptophan metabolism
Kynurenine metabolism is the central catabolic route for ingested Trp. It has two major branches, ending in the production of NAD+ or the neuroactive metabolite kynurenic acid (KA), respectively [13]. Each branch is active in multiple cell types and tissues, with local and systemic implications. The KP starts with converting Trp into N-formyl kynurenine (NFK). This process is a redox reaction mediated by one of three enzymes: tryptophan 2,3-dioxygenase (TDO2), a tetramer, and indoleamine 2,3-dioxygenase 1 and 2 (IDO1,2), both monomers [14,15]. This process is considered the rate-limiting KP step. TDO mediates this process in the liver, being responsible for the majority of conversion in normal conditions [16]. Moreover, TDO is the clue determinant of Trp availability in extrahepatic tissues [17,18,19,20]. At this stage, the activity of IDO1 and IDO2 enzymes is crucial. IDO2 enzyme possesses a lower catalytic action, its primary function being a negative regulator of the function of IDO1 enzyme, which has the major catalytic activity between them (Figure 1) [21].
The next step is regulated by mArylformamidaseenzyme (AFMID). AFMID removes the NFK formyl group, which results in obtaining kynurenine (KYN), the main product in this pathway. KYN can be converted to kynurenic acid (KINA), to anthranilic acid (AA) or to 3-hydroxykynurenine (3HK) by one of the three isoforms of kynurenine aminotransferase (KYAT1–3) or glutamic-oxaloacetic transaminase 2 (GOT2); the kynureninase (KYNU); or the mitochondrial-associated enzyme kynurenine 3-monooxygenase (KMO), respectively [22]. KYNU and the KYAT enzymes can turn 3HK into 3-hydroxy anthranilic acid (3HAA) or xanthurenic acid (XA). 3HAA can also be produced via non-enzymatic reaction with AA as the substrate. After a few reactions, 3HAA can be turned into picolinic acid (PIC). As an alternative step, 3-HAA could be quickly converted into quinolinic acid (QUIN) by a non-enzymatic reaction. As the final step of this pathway, NAD+ is produced as the end product of KP with multiple biological functions associated with energy metabolism, calcium homeostasis and gene expression [22,23].
1.2. Does aging modulate the kynurenine pathway? IDO1, the key to this interplay
Several studies have revealed that the aging process remodels the immune system and triggers pro-inflammatory changes in senescent non-immune cells [24]. This age-related chronic low-grade inflammation, called inflammaging, can induce significant epigenetic and transcriptional changes in human tissues [25]. The most significant one is an increase in the expression of inflammatory mediators and the anomalous activation of several signaling pathways [26].
Growing evidence shows that changes in Trp metabolism with aging are involved in the pathogenesis of multiple age-related diseases [11,27]. Interestingly IDO1 activation plays a pivotal role in the pathogenesis of those diseases [26]. Several inflammatory factors can induce the expression of the IDO1 gene via the activation of the NF-κB and JAK-STAT signaling pathways. Due to the age-related chronic inflammatory stage, levels of KYN are increased due to high levels of IDO-1 activity, which is also associated with high serum levels of interleukin-6 (IL-6), interleukin-1β (IL-1β), and interferon-γ (IF-γ); all potent activators of this enzyme [11,27,28]. The depletion of Trp and the subsequent accumulation of KYN metabolites through the activation of IDO1 disturb tissue homeostasis and promote inflammatory pathologies. Moreover, depletion of L-Trp in chronic inflammation might reduce protein synthesis and promote atrophy in the host tissue (Figure 2) [20,29,30,31].
The aryl hydrocarbon receptor (AhR), a potent immunosuppressive transcription factor, has also been involved in this complex process. Increased activation of AhR signaling increases tissue degeneration and thus promotes the aging process. Due to the high activity of IDO1 in old persons, with aging, high levels of KIN and KYNA can activate the AhR. As a result, IDO1/KYN/AhR signaling is significantly increased, promoting an immunosuppressive state in the inflammatory microenvironment, and being a breeding ground for chronic age-related diseases [32,33,34,35,36,37,38,39]. In addition, decreased levels of KP metabolites PIC and QUIN have been described in older persons, partly due to the age-related changes in the microbiota, which affect its capacity to produce these anabolic products [40].
2. Kynurenine pathway: an unexplored mechanism for osteoporosis
2.1. The link between aging and osteoporosis.
Bone is an active and heterogeneous tissue composed of different cell populations as well as an extracellular matrix (ECM). Mesenchymal stem cells (MSCs), osteoblasts and osteocytes are vital in preserving bone homeostasis and keeping the balance between modeling and remodeling [41,42,43,44,45,46]. In this respect, the differentiation of the adipogenic and osteogenic bone marrow mesenchymal stem cells (BMSCs) also plays an essential role [47,48]. The loss of the normal bone microarchitecture, the MSCs damage with aging due to inflammaging, and the marked increase in oxidative stress are all mechanisms of age-related bone loss and osteoporosis. This implies a decline in MSCs number and a change in their normal function, shifting to more adipogenic differentiation. This complex process which affects all levels of bone homeostasis has resulted in bone loss and osteoporosis, where aging is the major risk factor [49,50,51,52].
Osteoporosis characterizes by structural bone deterioration and decreased bone mineral density (BMD), frequently associated with frailty, sarcopenia and falls. In this context, bone fracture risk is increased, feeding this complex circuit that unequivocally implies an increase in disability, loss of quality of life and increased mortality in older persons, making osteoporosis a global problem [53,54,55].
2.2. Kynurenine pathway as a regulator of bone metabolism
Catabolic Trp metabolites may be key factors in the development of osteoporosis in older persons. It has been described that KP targets metabolic pathways critical for osteogenic homeostasis, in particular autophagy and senescence in BMSCs, both mechanisms to fight cell damage. There is also a link between KP and BMSCs differentiation via autophagy, a dynamic process essential for maintaining bone homeostasis, helping protect cells by recycling damaged and generating new energy and molecular supporting structures. Some studies have reported that KYN decreases CXCL12 protein levels, a critical element in BMSC osteogenesis, via the AhR signaling pathway, thus altering bone homeostasis [56,57,58,59,60]. On the other hand, regarding cellular senescence, defined as an early expiry of cell function and cell division, KYN increases the level of P21, a cyclin-dependent kinases (CDKs) inhibitor which forms part of the P21/CDKN1A pathway, a senescence inducer at G1/S1 phase of the cell cycle. Because P53 activates P21, a probable interaction between this tumor-suppressing transcription factor and KYN has been postulated. The role of other kynurenines in this complex process is still unclear [61,62,63,64,65]. Another point of interest is the relation between KYN and BMSCs differentiation. KYN decreases Hdac3 and CXCL12 and increases miR-29b-1-5p levels, a micro RNA that downregulates Hdac3 and CXCL12, both osteogenic genes, and upregulates AhR mRNA levels. The overall effect of such interactions is shifting from osteogenic to adipogenic differentiation in BMSCs [66,67,68,69,70,71,72].
Human studies have reported that high levels of 3-HK, KYNA and AA have a detrimental effect on bone, decreasing BMD and increasing fracture risk [73]. KYNA products on BMSCs are still unknown, and its interaction with glutamate receptors, such as the NMDA receptor presented in bone cells, could give us the answers. On its part, 3-HK has a pro-oxidative nature, which can induce ROS leading to cell apoptosis. The paper of AA is still unknown. However, increased levels have been described in patients with osteoporosis, making it a promising biological marker [74,75,76]. In contrast, high serum levels of 3-HAA, PIC, QUIN, and NAD+ increase BMD and are associated with lower fracture risk [73]. 3-AA has been described to have both pro-oxidant and antioxidant effects depending on the context, and the low serum levels observed in patients with osteoporosis maybe answer to a lower conversion from AA to 3AA. Evidence on the role of PIC points to an osteogenic effect. As a nuance, some studies have associated this metal chelator with increased adipose tissue in the bone marrow, suggesting a probable dose-dependence effect on bone. QUIN, a metabolite known for its interaction with NMDA receptors, has also been associated with osteogenic properties, but the mechanism is still unknown. Finally, NAD+, the terminal oxidation product of Trp metabolism, has been suggested to increase bone density. However, further studies are still needed [74,77,78,79].
Some studies have measured serum levels of kynurenines in a cohort of older patients with osteoporosis observing low levels of 3-HAA and high levels of AA, with the restoring to control values after osteoporosis treatment. This makes sense if we compare these results with previous evidence, suggesting a promising paper on kynurenines in monitoring osteoporosis treatment [80,81]. Nevertheless, due to the large and diverse therapeutic arsenal in osteoporosis, we need higher quality studies investigating the clinical role of kynurenines in different scenarios to be able to affirm this assertion. Moreover, osteogenic metabolites of KP as could be PIC have been described as having significant anabolic potential, being considered a seductive therapeutic target. However, to date, we only have results on its benefits in cellular and animal studies [82].
Collectively, current evidence connects KP with the regulation of MSCs differentiation and bone homeostasis both involved in osteoporosis pathogenesis, making this pathway a potentially useful diagnostic and therapeutic target in clinical practice (Figure 3).
3. Kynurenines can modulate the risk of sarcopenia
3.1. The link between aging and sarcopenia.
Skeletal muscle (SM) is one of our largest organs, representing almost 30-40% of the human body weight. Its main functions include voluntary movement and regulation of glucose metabolism. SM comprises bundles of muscle fibers, each formed by the fusion of mononucleated myoblasts. Moreover, it has its stem cell population, known as muscle satellite (stem) cells (MuSCs) and fibro-adipogenic progenitors. Myoblast fusion and MuSCs play a critical role in skeletal muscle development and regeneration [83,84,85].
SM is a dynamic organ with plenty of modulators, aging being an important regulating factor. Aging has been associated with a less regenerative capacity of SM. Phenotypically this implies a reduced ability to tolerate muscle damage, resulting in muscle atrophy and lost function. Several groups have reported the loss of nuclei in fibers, reduced numbers of myofibers and hypertrophic myofibers, and infiltration with adipose and fibrotic tissue [86] all associated with age-induced loss of function of MuSCs. It responds to intrinsic and extrinsic changes, being remarkable the effect of inflammaging and the increase of oxidative stress age-related and their effect in those cells, producing DNA and proteins damage through modifying signaling pathways and altering their metabolism, with a lower anabolic capacity [87,88,89,90]. Within these pathways, the activation of WNT-pathways, TGF-b-, FGF2, JAK/STAT3, p38 andp16INK4a has been described as reducing MuSCs capacity of regeneration inducing them, mainly, into a pre-senescent site or an apoptosis state [91,92,93,94,95,96,97].
Aging has been associated with dysfunctional homeostasis of skeletal muscle. This leads to loss of strength and function, both components of sarcopenia, a disease whose knowledge is currently the focus of intense research. Sarcopenia is defined by low strength, muscle mass and physical performance. Its prevalence increases with age and is particularly linked with hospitalization risk in older adults, multi-morbidity and frailty [98,99,100,101,102]. An increased risk of fractures and falls, mortality and health care cost has been linked with sarcopenia, which when concomitantly occurs with osteoporosis configures a syndrome known as osteosarcopenia [103,104,105,106]. To date, there is not an international consensus about sarcopenia, especially in what respects the diagnosis and pharmacological treatment, making this entity a promising target for studies.
3.1. The role of kynurenines in sarcopenia: a promising interplay
A connection between sarcopenia and the KP has been reported. KP likely contributes to muscle atrophy and oxidative stress via KYN. Some studies have demonstrated that KYN treatment reduces muscle size and strength and increases muscle lipid peroxidation, increasing protein catabolism and levels of reactive oxygen species (ROS) independently of Ahr activation. KYN treatment has also been associated with the preservation of body fat. Similarly, inhibition of IDO1 activity has been linked to increased muscle strength and fiber size [107,108]. Chronically elevated ROS levels and increased lipid peroxidation products have been associated with sarcopenia [109,110].
Furthermore, physical exercise is another skeletal muscle modulator, which affects strength and function and regulates nutrient metabolism. Aerobic exercise increases human lean muscle levels of PGC-1α1, a transcriptional coactivator essential for adaptive tissue response [111]. Activated PGC-1α1 together with PPARα/δ increases skeletal muscle expression of the KAT gene, which enzymes reduce KYN levels increasing KYNA production [112,113].
Some studies have evaluated the relationship between Trp metabolites and physical performance in aging populations observing that serum KYNA levels, as well as KYNA/KYN, PIC/QUIN, KYNA/PIC and KYNA/QUIN ratios, were associated with a lower likelihood of sarcopenia parameters, mainly grip strength and gait speed. On the other hand, lower physical function resulted in increased levels of 3-HAA, QUIN/KYN ratio and QUIN. Moreover, elevated serum IL-6, an inflammation marker, was observed in participants with higher KYN, KYN/TRP, 3-HK, QUIN, and QUIN/KYN ratios (Figure 4) [114].
Overall, kynurenines can be considered biomarkers and potential therapeutic targets for sarcopenia in the future. However, and despite a clear biological plausibility, further investigating the underlying mechanisms is needed to allow the use of KP metabolites in developing new diagnostic and therapeutic strategies to tackle sarcopenia and osteosarcopenia.
4. Frailty, an expression of aging and poor physical activity: ¿Have the kynurenines something to tell?
Frailty is defined as a clinical syndrome characterized by diminished endurance, strength, and decreased physiologic functions, which is embodied in phenotypic signs. Frailty has an estimated prevalence of 4% to 59,1%, in older persons. It is diagnosed in clinical practice by using very common criteria including five clinical criteria proposed by Fried et al. or deficit accumulation proposed by Rockwood et al. Due to its multidimensional nature, frailty origin usually responds to the interaction of multiple causal factors being remarkable physical, biological, psychological, genetic, environmental and social factors [115,116,117,118]. Frailty increases body susceptibility to acute illness and stressors, raising disability, institutionalization, falls, fractures and mortality risk [119,120]. These features are somewhat reminiscent of osteosarcopenia, in fact, frailty and osteosarcopenia are entities that usually go hand in hand.
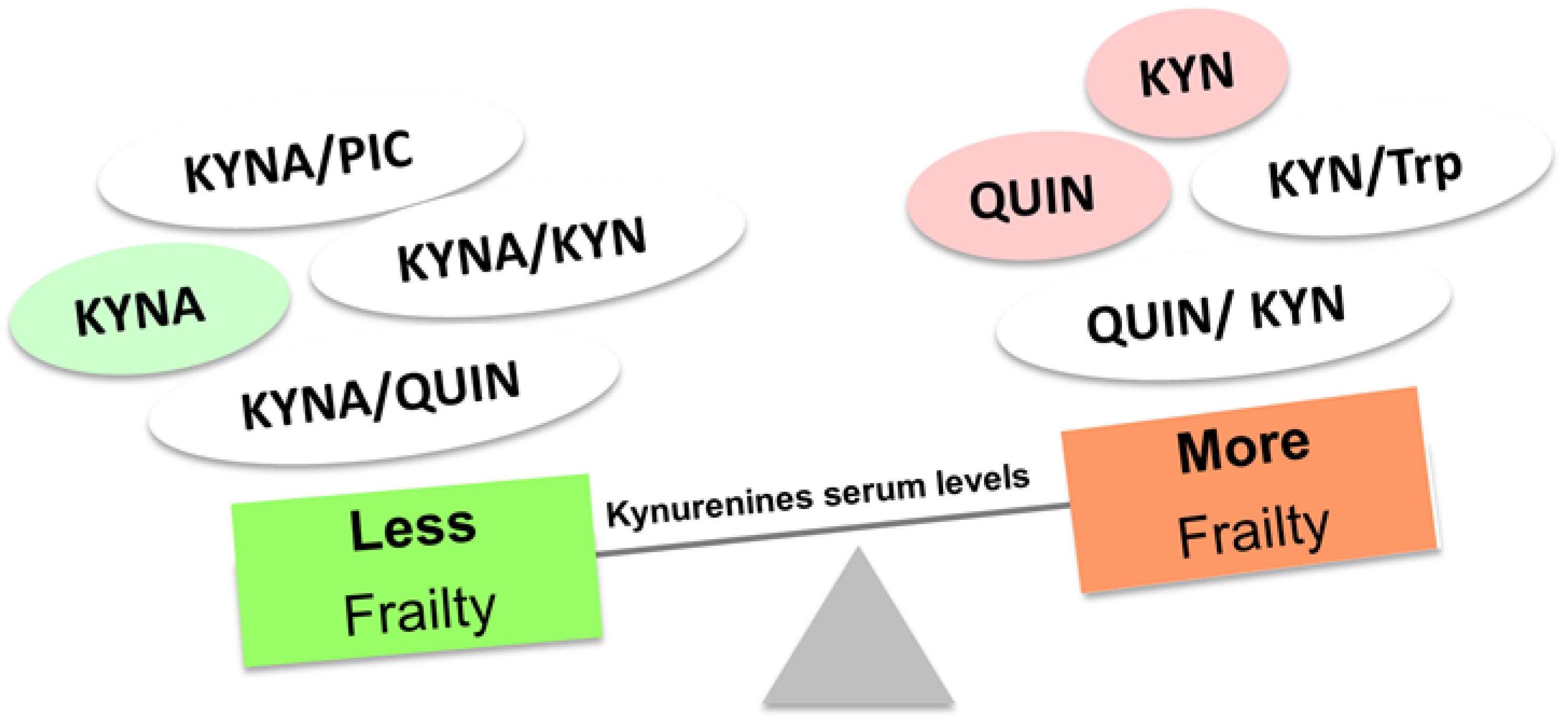
Frailty is a state in which physiologic reserve is reduced. This is explained mainly because of inflammaging. For this reason, it is difficult to understand these two entities as independent processes, particularly in the context of the KP. Indeed, several studies have tried to link physical frailty, frailty index and some physical parameters with KP. It has been reported that high KYN/Trp ratio levels in frail older adults diagnosed according to Fried´s criteria [119,121,122]. Along the same line, high levels of KYN have been seen in the serum of frail patients using both Fried et al. and Rockwood et al. criteria, also obtaining statistically significant differences by analyzing KYN interaction with physical parameters including time to complete 5 chair stands, grip strength or gait speed [123]. In another sample of frail people, KYNA, KYNA/KYN, KYNA/QUIN, and KYNA/PIC ratios were associated with a lower likelihood of frailty by the Rockwood index and Fried’s criteria. On the other hand, QUIN and QUIN/KYN ratios showed an association with a higher probability of frailty and increased IL-6. Regarding physical parameters, a reduction in gait speed was observed at high levels of 3-HAA and QUIN. High QUIN/KYN was also linked with less grip strength, while higher grip strength and gait speed were observed with an increased KYNA/QUIN ratio (Figure 5) [114].
Due to the lack of a homogeneous and unique definition for frailty and the complexity of this entity, the development of biomarkers has been extremely challenging. These collected data suggest that kynurenines could be an attractive biomarker for assessing frailty in humans. In addition, anabolic metabolites of the KP, such as picolinic acid, could have a therapeutic potential for frailty. However, further evidence is still needed.
4. Conclusions
Aging has been associated with changes in Trp metabolism mostly due to inflammaging and alterations in gut microbiota. IDO1, one of the main enzymes of extrahepatic Trp metabolism, is induced by inflammatory factors, which are increased in older persons because of inflammaging. This could explain why, with aging, serum levels of Trp are low and KYN are high. This finding could be associated with an increase individual´s predisposition to disease (i.e., neurodegenerative and musculoskeletal) by inducing significant epigenetic and transcriptional changes in human tissues. In addition, age-induced alterations in the gut microbiota have been associated with reduced capacity to metabolize Trp into its end products while favoring the production of the toxic KYN.
Osteoporosis, sarcopenia, osteosarcopenia, and frailty are very prevalent in old age. These entities are linked with chronically increased levels of inflammatory factors and changes in the microbiota. Serum levels of kynurenines are altered in older persons with osteoporosis, sarcopenia and frailty, where higher levels of catabolic kynurenines are observed. KYN can misbalance bone homeostasis, making it susceptible to osteoporosis by affecting senescence, autophagy and differentiation of bone marrow stem cells. KYN treatment also reduces muscle size and strength and increases muscle lipid peroxidation upgrading protein catabolism, leading to sarcopenia. Some studies have demonstrated that poor physical activity, one of the main factors contributing to frailty, osteoporosis and sarcopenia, can modulate KP, inducing changes in KYN levels that are very similar to those observed in frail older persons. In contrast, we and others have reported that some anabolic kynurenines may have anabolic properties in bone and skeletal muscle cells.
In conclusion, kynurenines could become potential biomarkers for osteoporosis, sarcopenia, osteosarcopenia and frailty in older persons. In addition, some of these metabolic products could be an interesting target in the development of new pharmacological treatments. Although current evidence is promising, more studies are needed to address these issues.
Author Contributions
Conceptualization, J.B. and G.D.; writing, all authors; reviewing and editing, all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This review received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liu, J.-J.; Movassat, J.; Portha, B. Emerging Role for Kynurenines in Metabolic Pathologies. Curr. Opin. Clin. Nutr. Metab. Care 2019, 22, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Bender, D.A. Effects of a Dietary Excess of Leucine on the Metabolism of Tryptophan in the Rat: A Mechanism for the Pellagragenic Action of Leucine. Br. J. Nutr. 1983, 50, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Boado, R.J.; Li, J.Y.; Nagaya, M.; Zhang, C.; Pardridge, W.M. Selective Expression of the Large Neutral Amino Acid Transporter at the Blood-Brain Barrier. Proc. Natl. Acad. Sci. U. S. A. 1999, 96, 12079–12084. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.S.; Davies, S.S. Microbial Metabolism of Dietary Components to Bioactive Metabolites: Opportunities for New Therapeutic Interventions. Genome Med. 2016, 8, 46. [Google Scholar] [CrossRef] [PubMed]
- Takikawa, O.; Yoshida, R.; Kido, R.; Hayaishi, O. Tryptophan Degradation in Mice Initiated by Indoleamine 2,3-Dioxygenase. J. Biol. Chem. 1986, 261, 3648–3653. [Google Scholar] [CrossRef]
- Canli, T.; Lesch, K.-P. Long Story Short: The Serotonin Transporter in Emotion Regulation and Social Cognition. Nat. Neurosci. 2007, 10, 1103–1109. [Google Scholar] [CrossRef]
- Yates, C.A.; Herbert, J. Differential Circadian Rhythms in Pineal and Hypothalamic 5-HT Induced by Artificial Photoperiods or Melatonin. Nature 1976, 262, 219–220. [Google Scholar] [CrossRef]
- Stone, T.W.; Stoy, N.; Darlington, L.G. An Expanding Range of Targets for Kynurenine Metabolites of Tryptophan. Trends Pharmacol. Sci. 2013, 34, 136–143. [Google Scholar] [CrossRef]
- Kim, B.-J.; Hamrick, M.W.; Yoo, H.J.; Lee, S.H.; Kim, S.J.; Koh, J.-M.; Isales, C.M. The Detrimental Effects of Kynurenine, a Tryptophan Metabolite, on Human Bone Metabolism. J. Clin. Endocrinol. Metab. 2019, 104, 2334–2342. [Google Scholar] [CrossRef]
- Sorgdrager, F.J.H.; Vermeiren, Y.; Van Faassen, M.; van der Ley, C.; Nollen, E.A.A.; Kema, I.P.; De Deyn, P.P. Age- and Disease-Specific Changes of the Kynurenine Pathway in Parkinson’s and Alzheimer’s Disease. J. Neurochem. 2019, 151, 656–668. [Google Scholar] [CrossRef]
- Baumgartner, R.; Forteza, M.J.; Ketelhuth, D.F.J. The Interplay between Cytokines and the Kynurenine Pathway in Inflammation and Atherosclerosis. Cytokine 2019, 122, 154148. [Google Scholar] [CrossRef] [PubMed]
- Lytton, S.D.; Osiecki, M.; MałgorzataWoźniak; Cukrowska, B.; Wierzbicka, A.; Goliszek, M.; Socha, P.; Janczyk, W.; Dayanakli, D.; Abendroth, D.; et al. Tryptophan-Kynurenine Profile in Pediatric Autoimmune Hepatitis. Immunol. Res. 2019, 67, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Ramprasath, T.; Wang, H.; Zou, M.-H. Abnormal Kynurenine Pathway of Tryptophan Catabolism in Cardiovascular Diseases. Cell. Mol. Life Sci. CMLS 2017, 74, 2899–2916. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Menzies, K.J.; Auwerx, J. NAD(+) Metabolism and the Control of Energy Homeostasis: A Balancing Act between Mitochondria and the Nucleus. Cell Metab. 2015, 22, 31–53. [Google Scholar] [CrossRef]
- Nelp, M.T.; Kates, P.A.; Hunt, J.T.; Newitt, J.A.; Balog, A.; Maley, D.; Zhu, X.; Abell, L.; Allentoff, A.; Borzilleri, R.; et al. Immune-Modulating Enzyme Indoleamine 2,3-Dioxygenase Is Effectively Inhibited by Targeting Its Apo-Form. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, 3249–3254. [Google Scholar] [CrossRef] [PubMed]
- Kanai, M.; Funakoshi, H.; Takahashi, H.; Hayakawa, T.; Mizuno, S.; Matsumoto, K.; Nakamura, T. Tryptophan 2,3-Dioxygenase Is a Key Modulator of Physiological Neurogenesis and Anxiety-Related Behavior in Mice. Mol. Brain 2009, 2, 8. [Google Scholar] [CrossRef]
- Civen, M.; Knox, W.E. The Independence of Hydrocortisone and Tryptophan Inductions of Tryptophan Pyrrolase. J. Biol. Chem. 1959, 234, 1787–1790. [Google Scholar] [CrossRef]
- Greengard, O.; Feigelson, P. A Difference between the Modes of Action of Substrate and Hormonal Inducers of Rat Liver Tryptophan Pyrrolase. Nature 1961, 190, 446–447. [Google Scholar] [CrossRef]
- Badawy, A.A.-B. Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. Int. J. Tryptophan Res. IJTR 2017, 10, 1178646917691938. [Google Scholar] [CrossRef]
- Pallotta, M.T.; Rossini, S.; Suvieri, C.; Coletti, A.; Orabona, C.; Macchiarulo, A.; Volpi, C.; Grohmann, U. Indoleamine 2,3-Dioxygenase 1 (IDO1): An up-to-Date Overview of an Eclectic Immunoregulatory Enzyme. FEBS J. 2022, 289, 6099–6118. [Google Scholar] [CrossRef]
- Lee, Y.-K.; Lee, H.B.; Shin, D.-M.; Kang, M.J.; Yi, E.C.; Noh, S.; Lee, J.; Lee, C.; Min, C.-K.; Choi, E.Y. Heme-Binding-Mediated Negative Regulation of the Tryptophan Metabolic Enzyme Indoleamine 2,3-Dioxygenase 1 (IDO1) by IDO2. Exp. Mol. Med. 2014, 46, e121. [Google Scholar] [CrossRef] [PubMed]
- Schwarcz, R.; Bruno, J.P.; Muchowski, P.J.; Wu, H.-Q. Kynurenines in the Mammalian Brain: When Physiology Meets Pathology. Nat. Rev. Neurosci. 2012, 13, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Nishizuka, Y.; Hayaishi, O. Enzymic Synthesis of Niacin Nucleotides from 3-Hydroxyanthranilic Acid in Mammalian Liver. J. Biol. Chem. 1963, 238, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Freund, A.; Orjalo, A.V.; Desprez, P.-Y.; Campisi, J. Inflammatory Networks during Cellular Senescence: Causes and Consequences. Trends Mol. Med. 2010, 16, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Valensin, S.; Bonafè, M.; Paolisso, G.; Yashin, A.I.; Monti, D.; De Benedictis, G. The Network and the Remodeling Theories of Aging: Historical Background and New Perspectives. Exp. Gerontol. 2000, 35, 879–896. [Google Scholar] [CrossRef]
- Benayoun, B.A.; Pollina, E.A.; Singh, P.P.; Mahmoudi, S.; Harel, I.; Casey, K.M.; Dulken, B.W.; Kundaje, A.; Brunet, A. Remodeling of Epigenome and Transcriptome Landscapes with Aging in Mice Reveals Widespread Induction of Inflammatory Responses. Genome Res. 2019, 29, 697–709. [Google Scholar] [CrossRef]
- Yu, J.; Wang, Y.; Yan, F.; Zhang, P.; Li, H.; Zhao, H.; Yan, C.; Yan, F.; Ren, X. Noncanonical NF-ΚB Activation Mediates STAT3-Stimulated IDO Upregulation in Myeloid-Derived Suppressor Cells in Breast Cancer. J. Immunol. Baltim. Md 1950 2014, 193, 2574–2586. [Google Scholar] [CrossRef]
- Mailankot, M.; Nagaraj, R.H. Induction of Indoleamine 2,3-Dioxygenase by Interferon-Gamma in Human Lens Epithelial Cells: Apoptosis through the Formation of 3-Hydroxykynurenine. Int. J. Biochem. Cell Biol. 2010, 42, 1446–1454. [Google Scholar] [CrossRef]
- Schmidt, S.V.; Schultze, J.L. New Insights into IDO Biology in Bacterial and Viral Infections. Front. Immunol. 2014, 5, 384. [Google Scholar] [CrossRef]
- Wang, X.-F.; Wang, H.-S.; Wang, H.; Zhang, F.; Wang, K.-F.; Guo, Q.; Zhang, G.; Cai, S.-H.; Du, J. The Role of Indoleamine 2,3-Dioxygenase (IDO) in Immune Tolerance: Focus on Macrophage Polarization of THP-1 Cells. Cell. Immunol. 2014, 289, 42–48. [Google Scholar] [CrossRef]
- Munn, D.H.; Mellor, A.L. IDO in the Tumor Microenvironment: Inflammation, Counter-Regulation, and Tolerance. Trends Immunol. 2016, 37, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Mezrich, J.D.; Fechner, J.H.; Zhang, X.; Johnson, B.P.; Burlingham, W.J.; Bradfield, C.A. An Interaction between Kynurenine and the Aryl Hydrocarbon Receptor Can Generate Regulatory T Cells. J. Immunol. Baltim. Md 1950 2010, 185, 3190–3198. [Google Scholar] [CrossRef] [PubMed]
- Rothhammer, V.; Quintana, F.J. The Aryl Hydrocarbon Receptor: An Environmental Sensor Integrating Immune Responses in Health and Disease. Nat. Rev. Immunol. 2019, 19, 184–197. [Google Scholar] [CrossRef] [PubMed]
- DiNatale, B.C.; Murray, I.A.; Schroeder, J.C.; Flaveny, C.A.; Lahoti, T.S.; Laurenzana, E.M.; Omiecinski, C.J.; Perdew, G.H. Kynurenic Acid Is a Potent Endogenous Aryl Hydrocarbon Receptor Ligand That Synergistically Induces Interleukin-6 in the Presence of Inflammatory Signaling. Toxicol. Sci. Off. J. Soc. Toxicol. 2010, 115, 89–97. [Google Scholar] [CrossRef]
- Sadik, A.; Somarribas Patterson, L.F.; Öztürk, S.; Mohapatra, S.R.; Panitz, V.; Secker, P.F.; Pfänder, P.; Loth, S.; Salem, H.; Prentzell, M.T.; et al. IL4I1 Is a Metabolic Immune Checkpoint That Activates the AHR and Promotes Tumor Progression. Cell 2020, 182, 1252–1270. [Google Scholar] [CrossRef]
- Zhu, J.; Luo, L.; Tian, L.; Yin, S.; Ma, X.; Cheng, S.; Tang, W.; Yu, J.; Ma, W.; Zhou, X.; et al. Aryl Hydrocarbon Receptor Promotes IL-10 Expression in Inflammatory Macrophages Through Src-STAT3 Signaling Pathway. Front. Immunol. 2018, 9, 2033. [Google Scholar] [CrossRef]
- Torti, M.F.; Giovannoni, F.; Quintana, F.J.; García, C.C. The Aryl Hydrocarbon Receptor as a Modulator of Anti-Viral Immunity. Front. Immunol. 2021, 12, 624293. [Google Scholar] [CrossRef]
- Piper, C.J.M.; Rosser, E.C.; Oleinika, K.; Nistala, K.; Krausgruber, T.; Rendeiro, A.F.; Banos, A.; Drozdov, I.; Villa, M.; Thomson, S.; et al. Aryl Hydrocarbon Receptor Contributes to the Transcriptional Program of IL-10-Producing Regulatory B Cells. Cell Rep. 2019, 29, 1878–1892. [Google Scholar] [CrossRef]
- Gandhi, R.; Kumar, D.; Burns, E.J.; Nadeau, M.; Dake, B.; Laroni, A.; Kozoriz, D.; Weiner, H.L.; Quintana, F.J. Activation of the Aryl Hydrocarbon Receptor Induces Human Type 1 Regulatory T Cell-like and Foxp3(+) Regulatory T Cells. Nat. Immunol. 2010, 11, 846–853. [Google Scholar] [CrossRef]
- Watson, M.D.; Cross, B.L.; Grosicki, G.J. Evidence for the Contribution of Gut Microbiota to Age-Related Anabolic Resistance. Nutrients 2021, 13, 706. [Google Scholar] [CrossRef]
- Manolagas, S.C.; Parfitt, A.M. What Old Means to Bone. Trends Endocrinol. Metab. TEM 2010, 21, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Schönherr, E.; Hausser, H.J. Extracellular Matrix and Cytokines: A Functional Unit. Dev. Immunol. 2000, 7, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Manolagas, S.C. Birth and Death of Bone Cells: Basic Regulatory Mechanisms and Implications for the Pathogenesis and Treatment of Osteoporosis. Endocr. Rev. 2000, 21, 115–137. [Google Scholar] [PubMed]
- Kim, S.W.; Lu, Y.; Williams, E.A.; Lai, F.; Lee, J.Y.; Enishi, T.; Balani, D.H.; Ominsky, M.S.; Ke, H.Z.; Kronenberg, H.M.; et al. Sclerostin Antibody Administration Converts Bone Lining Cells Into Active Osteoblasts. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2017, 32, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Schaffler, M.B.; Cheung, W.-Y.; Majeska, R.; Kennedy, O. Osteocytes: Master Orchestrators of Bone. Calcif. Tissue Int. 2014, 94, 5–24. [Google Scholar] [CrossRef] [PubMed]
- Matic, I.; Matthews, B.G.; Wang, X.; Dyment, N.A.; Worthley, D.L.; Rowe, D.W.; Grcevic, D.; Kalajzic, I. Quiescent Bone Lining Cells Are a Major Source of Osteoblasts During Adulthood. Stem Cells Dayt. Ohio 2016, 34, 2930–2942. [Google Scholar] [CrossRef]
- Tzeng, Y.-S.; Chung, N.-C.; Chen, Y.-R.; Huang, H.-Y.; Chuang, W.-P.; Lai, D.-M. Imbalanced Osteogenesis and Adipogenesis in Mice Deficient in the Chemokine Cxcl12/Sdf1 in the Bone Mesenchymal Stem/Progenitor Cells. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2018, 33, 679–690. [Google Scholar] [CrossRef]
- Ripoll, C.B.; Bunnell, B.A. Comparative Characterization of Mesenchymal Stem Cells from EGFP Transgenic and Non-Transgenic Mice. BMC Cell Biol. 2009, 10, 3. [Google Scholar] [CrossRef]
- Atashi, F.; Modarressi, A.; Pepper, M.S. The Role of Reactive Oxygen Species in Mesenchymal Stem Cell Adipogenic and Osteogenic Differentiation: A Review. Stem Cells Dev. 2015, 24, 1150–1163. [Google Scholar] [CrossRef]
- Chen, F.; Liu, Y.; Wong, N.-K.; Xiao, J.; So, K.-F. Oxidative Stress in Stem Cell Aging. Cell Transplant. 2017, 26, 1483–1495. [Google Scholar] [CrossRef]
- Coipeau, P.; Rosset, P.; Langonne, A.; Gaillard, J.; Delorme, B.; Rico, A.; Domenech, J.; Charbord, P.; Sensebe, L. Impaired Differentiation Potential of Human Trabecular Bone Mesenchymal Stromal Cells from Elderly Patients. Cytotherapy 2009, 11. [Google Scholar] [CrossRef] [PubMed]
- Infante, A.; Rodríguez, C.I. Osteogenesis and Aging: Lessons from Mesenchymal Stem Cells. Stem Cell Res. Ther. 2018, 9, 244. [Google Scholar] [CrossRef] [PubMed]
- Bonafede, M.; Espindle, D.; Bower, A.G. The Direct and Indirect Costs of Long Bone Fractures in a Working Age US Population. J. Med. Econ. 2013, 16, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Wright, N.C.; Looker, A.C.; Saag, K.G.; Curtis, J.R.; Delzell, E.S.; Randall, S.; Dawson-Hughes, B. The Recent Prevalence of Osteoporosis and Low Bone Mass in the United States Based on Bone Mineral Density at the Femoral Neck or Lumbar Spine. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2014, 29, 2520–2526. [Google Scholar] [CrossRef]
- Sözen, T.; Özışık, L.; Başaran, N.Ç. An Overview and Management of Osteoporosis. Eur. J. Rheumatol. 2017, 4, 46–56. [Google Scholar] [CrossRef]
- Elmansi, A.M.; Hussein, K.A.; Herrero, S.M.; Periyasamy-Thandavan, S.; Aguilar-Pérez, A.; Kondrikova, G.; Kondrikov, D.; Eisa, N.H.; Pierce, J.L.; Kaiser, H.; et al. Age-Related Increase of Kynurenine Enhances MiR29b-1-5p to Decrease Both CXCL12 Signaling and the Epigenetic Enzyme Hdac3 in Bone Marrow Stromal Cells. Bone Rep. 2020, 12, 100270. [Google Scholar] [CrossRef]
- Guan, J.-L.; Simon, A.K.; Prescott, M.; Menendez, J.A.; Liu, F.; Wang, F.; Wang, C.; Wolvetang, E.; Vazquez-Martin, A.; Zhang, J. Autophagy in Stem Cells. Autophagy 2013, 9, 830–849. [Google Scholar] [CrossRef]
- Qi, M.; Zhang, L.; Ma, Y.; Shuai, Y.; Li, L.; Luo, K.; Liu, W.; Jin, Y. Autophagy Maintains the Function of Bone Marrow Mesenchymal Stem Cells to Prevent Estrogen Deficiency-Induced Osteoporosis. Theranostics 2017, 7, 4498–4516. [Google Scholar] [CrossRef]
- Zhou, Z.; Shi, G.; Zheng, X.; Jiang, S.; Jiang, L. Autophagy Activation Facilitates Mechanical Stimulation-Promoted Osteoblast Differentiation and Ameliorates Hindlimb Unloading-Induced Bone Loss. Biochem. Biophys. Res. Commun. 2018, 498, 667–673. [Google Scholar] [CrossRef]
- Zhang, B.; Hou, R.; Zou, Z.; Luo, T.; Zhang, Y.; Wang, L.; Wang, B. Mechanically Induced Autophagy Is Associated with ATP Metabolism and Cellular Viability in Osteocytes in Vitro. Redox Biol. 2018, 14, 492–498. [Google Scholar] [CrossRef]
- Kondrikov, D.; Elmansi, A.; Bragg, R.T.; Mobley, T.; Barrett, T.; Eisa, N.; Kondrikova, G.; Schoeinlein, P.; Aguilar-Perez, A.; Shi, X.-M.; et al. Kynurenine Inhibits Autophagy and Promotes Senescence in Aged Bone Marrow Mesenchymal Stem Cells through the Aryl Hydrocarbon Receptor Pathway. Exp. Gerontol. 2020, 130, 110805. [Google Scholar] [CrossRef] [PubMed]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O. A Biomarker That Identifies Senescent Human Cells in Culture and in Aging Skin in Vivo. Proc. Natl. Acad. Sci. U. S. A. 1995, 92, 9363–9367. [Google Scholar] [CrossRef] [PubMed]
- Chilosi, M.; Carloni, A.; Rossi, A.; Poletti, V. Premature Lung Aging and Cellular Senescence in the Pathogenesis of Idiopathic Pulmonary Fibrosis and COPD/Emphysema. Transl. Res. J. Lab. Clin. Med. 2013, 162, 156–173. [Google Scholar] [CrossRef]
- Pignolo, R.J.; Samsonraj, R.M.; Law, S.F.; Wang, H.; Chandra, A. Targeting Cell Senescence for the Treatment of Age-Related Bone Loss. Curr. Osteoporos. Rep. 2019, 17, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.Y.; Jee, H.J.; Um, J.-H.; Kim, Y.M.; Bae, S.S.; Yun, J. Cooperation between P21 and Akt Is Required for P53-Dependent Cellular Senescence. Aging Cell 2017, 16, 1094–1103. [Google Scholar] [CrossRef] [PubMed]
- McGee-Lawrence, M.E.; Carpio, L.R.; Schulze, R.J.; Pierce, J.L.; McNiven, M.A.; Farr, J.N.; Khosla, S.; Oursler, M.J.; Westendorf, J.J. Hdac3 Deficiency Increases Marrow Adiposity and Induces Lipid Storage and Glucocorticoid Metabolism in Osteochondroprogenitor Cells. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2016, 31, 116–128. [Google Scholar] [CrossRef]
- Feichtinger, X.; Muschitz, C.; Heimel, P.; Baierl, A.; Fahrleitner-Pammer, A.; Redl, H.; Resch, H.; Geiger, E.; Skalicky, S.; Dormann, R.; et al. Bone-Related Circulating MicroRNAs MiR-29b-3p, MiR-550a-3p, and MiR-324-3p and Their Association to Bone Microstructure and Histomorphometry. Sci. Rep. 2018, 8, 4867. [Google Scholar] [CrossRef]
- Razidlo, D.F.; Whitney, T.J.; Casper, M.E.; McGee-Lawrence, M.E.; Stensgard, B.A.; Li, X.; Secreto, F.J.; Knutson, S.K.; Hiebert, S.W.; Westendorf, J.J. Histone Deacetylase 3 Depletion in Osteo/Chondroprogenitor Cells Decreases Bone Density and Increases Marrow Fat. PloS One 2010, 5, e11492. [Google Scholar] [CrossRef] [PubMed]
- Refaey, M.E.; McGee-Lawrence, M.E.; Fulzele, S.; Kennedy, E.J.; Bollag, W.B.; Elsalanty, M.; Zhong, Q.; Ding, K.-H.; Bendzunas, N.G.; Shi, X.-M.; et al. Kynurenine, a Tryptophan Metabolite That Accumulates With Age, Induces Bone Loss. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2017, 32, 2182–2193. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.A.R.; Stein, J.L.; Westendorf, J.J.; van Wijnen, A.J. Chromatin Modifiers and Histone Modifications in Bone Formation, Regeneration, and Therapeutic Intervention for Bone-Related Disease. Bone 2015, 81, 739–745. [Google Scholar] [CrossRef]
- Feigenson, M.; Shull, L.C.; Taylor, E.L.; Camilleri, E.T.; Riester, S.M.; van Wijnen, A.J.; Bradley, E.W.; Westendorf, J.J. Histone Deacetylase 3 Deletion in Mesenchymal Progenitor Cells Hinders Long Bone Development. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2017, 32, 2453–2465. [Google Scholar] [CrossRef]
- Carpio, L.R.; Bradley, E.W.; McGee-Lawrence, M.E.; Weivoda, M.M.; Poston, D.D.; Dudakovic, A.; Xu, M.; Tchkonia, T.; Kirkland, J.L.; van Wijnen, A.J.; et al. Histone Deacetylase 3 Supports Endochondral Bone Formation by Controlling Cytokine Signaling and Matrix Remodeling. Sci. Signal. 2016, 9, ra79. [Google Scholar] [CrossRef] [PubMed]
- Anaya, J.M.; Bollag, W.B.; Hamrick, M.W.; Isales, C.M. The Role of Tryptophan Metabolites in Musculoskeletal Stem Cell Aging. Int. J. Mol. Sci. 2020, 21, 6670. [Google Scholar] [CrossRef] [PubMed]
- Darlington, L.G.; Forrest, C.M.; Mackay, G.M.; Smith, R.A.; Smith, A.J.; Stoy, N.; Stone, T.W. On the Biological Importance of the 3-Hydroxyanthranilic Acid: Anthranilic Acid Ratio. Int. J. Tryptophan Res. IJTR 2010, 3, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Isales, C.; Ding, K.; Bollag, W.; McGee-Lawrence, M.; Hill, W.; Shi, X.; Elsalanty, M.; Hamrick, M. KYNURENIC ACID A TRYPTOPHAN METABOLITE INDUCES BONE LOSS IN MICE. Innov. Aging 2018, 2, 100–101. [Google Scholar] [CrossRef]
- Okuda, S.; Nishiyama, N.; Saito, H.; Katsuki, H. Hydrogen Peroxide-Mediated Neuronal Cell Death Induced by an Endogenous Neurotoxin, 3-Hydroxykynurenine. Proc. Natl. Acad. Sci. U. S. A. 1996, 93, 12553–12558. [Google Scholar] [CrossRef]
- Aman, Y.; Qiu, Y.; Tao, J.; Fang, E. Therapeutic Potential of Boosting NAD+ in Aging and Age-Related Diseases. Transl. Med. Aging 2018, 2. [Google Scholar] [CrossRef]
- Ding, K.; McGee-Lawrence, M.E.; Kaiser, H.; Sharma, A.K.; Pierce, J.L.; Irsik, D.L.; Bollag, W.B.; Xu, J.; Zhong, Q.; Hill, W.; et al. Picolinic Acid, a Tryptophan Oxidation Product, Does Not Impact Bone Mineral Density but Increases Marrow Adiposity. Exp. Gerontol. 2020, 133, 110885. [Google Scholar] [CrossRef]
- Vidal, C.; Li, W.; Santner-Nanan, B.; Lim, C.K.; Guillemin, G.J.; Ball, H.J.; Hunt, N.H.; Nanan, R.; Duque, G. The Kynurenine Pathway of Tryptophan Degradation Is Activated during Osteoblastogenesis. Stem Cells Dayt. Ohio 2015, 33, 111–121. [Google Scholar] [CrossRef]
- Forrest, C.M.; Mackay, G.M.; Oxford, L.; Stoy, N.; Stone, T.W.; Darlington, L.G. Kynurenine Pathway Metabolism in Patients with Osteoporosis after 2 Years of Drug Treatment. Clin. Exp. Pharmacol. Physiol. 2006, 33, 1078–1087. [Google Scholar] [CrossRef]
- Apalset, E.M.; Gjesdal, C.G.; Ueland, P.M.; Midttun, Ø.; Ulvik, A.; Eide, G.E.; Meyer, K.; Tell, G.S. Interferon (IFN)-γ-Mediated Inflammation and the Kynurenine Pathway in Relation to Bone Mineral Density: The Hordaland Health Study. Clin. Exp. Immunol. 2014, 176, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Duque, G.; Vidal, C.; Li, W.; Al Saedi, A.; Khalil, M.; Lim, C.K.; Myers, D.E.; Guillemin, G.J. Picolinic Acid, a Catabolite of Tryptophan, Has an Anabolic Effect on Bone In Vivo. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2020, 35, 2275–2288. [Google Scholar] [CrossRef] [PubMed]
- Janssen, I.; Heymsfield, S.B.; Wang, Z.M.; Ross, R. Skeletal Muscle Mass and Distribution in 468 Men and Women Aged 18-88 Yr. J. Appl. Physiol. Bethesda Md 1985 2000, 89, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jin, P.; Duan, R.; Chen, E.H. Mechanisms of Myoblast Fusion during Muscle Development. Curr. Opin. Genet. Dev. 2015, 32, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Shavlakadze, T.; McGeachie, J.; Grounds, M.D. Delayed but Excellent Myogenic Stem Cell Response of Regenerating Geriatric Skeletal Muscles in Mice. Biogerontology 2010, 11, 363–376. [Google Scholar] [CrossRef]
- Fry, C.S.; Lee, J.D.; Mula, J.; Kirby, T.J.; Jackson, J.R.; Liu, F.; Yang, L.; Mendias, C.L.; Dupont-Versteegden, E.E.; McCarthy, J.J.; et al. Inducible Depletion of Satellite Cells in Adult, Sedentary Mice Impairs Muscle Regenerative Capacity without Affecting Sarcopenia. Nat. Med. 2015, 21, 76–80. [Google Scholar] [CrossRef]
- Sousa-Victor, P.; Gutarra, S.; García-Prat, L.; Rodriguez-Ubreva, J.; Ortet, L.; Ruiz-Bonilla, V.; Jardí, M.; Ballestar, E.; González, S.; Serrano, A.L.; et al. Geriatric Muscle Stem Cells Switch Reversible Quiescence into Senescence. Nature 2014, 506, 316–321. [Google Scholar] [CrossRef]
- Carlson, M.E.; Hsu, M.; Conboy, I.M. Imbalance between PSmad3 and Notch Induces CDK Inhibitors in Old Muscle Stem Cells. Nature 2008, 454, 528–532. [Google Scholar] [CrossRef]
- Conboy, I.M.; Conboy, M.J.; Smythe, G.M.; Rando, T.A. Notch-Mediated Restoration of Regenerative Potential to Aged Muscle. Science 2003, 302, 1575–1577. [Google Scholar] [CrossRef]
- Brack, A.S.; Conboy, M.J.; Roy, S.; Lee, M.; Kuo, C.J.; Keller, C.; Rando, T.A. Increased Wnt Signaling during Aging Alters Muscle Stem Cell Fate and Increases Fibrosis. Science 2007, 317, 807–810. [Google Scholar] [CrossRef]
- Bernet, J.D.; Doles, J.D.; Hall, J.K.; Kelly Tanaka, K.; Carter, T.A.; Olwin, B.B. P38 MAPK Signaling Underlies a Cell-Autonomous Loss of Stem Cell Self-Renewal in Skeletal Muscle of Aged Mice. Nat. Med. 2014, 20, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Chakkalakal, J.V.; Jones, K.M.; Basson, M.A.; Brack, A.S. The Aged Niche Disrupts Muscle Stem Cell Quiescence. Nature 2012, 490, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Tierney, M.T.; Aydogdu, T.; Sala, D.; Malecova, B.; Gatto, S.; Puri, P.L.; Latella, L.; Sacco, A. STAT3 Signaling Controls Satellite Cell Expansion and Skeletal Muscle Repair. Nat. Med. 2014, 20, 1182–1186. [Google Scholar] [CrossRef] [PubMed]
- Price, F.D.; von Maltzahn, J.; Bentzinger, C.F.; Dumont, N.A.; Yin, H.; Chang, N.C.; Wilson, D.H.; Frenette, J.; Rudnicki, M.A. Inhibition of JAK-STAT Signaling Stimulates Adult Satellite Cell Function. Nat. Med. 2014, 20, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, B.D.; Gilbert, P.M.; Porpiglia, E.; Mourkioti, F.; Lee, S.P.; Corbel, S.Y.; Llewellyn, M.E.; Delp, S.L.; Blau, H.M. Rejuvenation of the Muscle Stem Cell Population Restores Strength to Injured Aged Muscles. Nat. Med. 2014, 20, 255–264. [Google Scholar] [CrossRef]
- Sousa-Victor, P.; Perdiguero, E.; Muñoz-Cánoves, P. Geroconversion of Aged Muscle Stem Cells under Regenerative Pressure. Cell Cycle Georget. Tex 2014, 13, 3183–3190. [Google Scholar] [CrossRef]
- Segalés, J.; Perdiguero, E.; Muñoz-Cánoves, P. Regulation of Muscle Stem Cell Functions: A Focus on the P38 MAPK Signaling Pathway. Front. Cell Dev. Biol. 2016, 4, 91. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Sarcopenia: Revised European Consensus on Definition and Diagnosis. Age Ageing 2019, 48, 601. [Google Scholar] [CrossRef]
- Dennison, E.M.; Sayer, A.A.; Cooper, C. Epidemiology of Sarcopenia and Insight into Possible Therapeutic Targets. Nat. Rev. Rheumatol. 2017, 13, 340–347. [Google Scholar] [CrossRef]
- Bruyère, O.; Beaudart, C.; Ethgen, O.; Reginster, J.-Y.; Locquet, M. The Health Economics Burden of Sarcopenia: A Systematic Review. Maturitas 2019, 119, 61–69. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; Sayer, A.A. Sarcopenia. Lancet Lond. Engl. 2019, 393, 2636–2646. [Google Scholar] [CrossRef] [PubMed]
- Sui, S.X.; Holloway-Kew, K.L.; Hyde, N.K.; Williams, L.J.; Tembo, M.C.; Leach, S.; Pasco, J.A. Prevalence of Sarcopenia Employing Population-Specific Cut-Points: Cross-Sectional Data from the Geelong Osteoporosis Study, Australia. J. Clin. Med. 2021, 10, 343. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.Q.; Yu, S.; Tucker, G.R.; Adams, R.J.; Cesari, M.; Theou, O.; Visvanathan, R. Frailty and Sarcopenia in Combination Are More Predictive of Mortality than Either Condition Alone. Maturitas 2021, 144, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, S.K.; Tsintavis, P.; Potsaki, P.; Papandreou, D. Differences in the Prevalence of Sarcopenia in Community-Dwelling, Nursing Home and Hospitalized Individuals. A Systematic Review and Meta-Analysis. J. Nutr. Health Aging 2020, 24, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, J.; Geerlings, M.A.J.; Reijnierse, E.M.; Phassouliotis, C.; Lim, W.K.; Maier, A.B. Prevalence of Sarcopenia as a Comorbid Disease: A Systematic Review and Meta-Analysis. Exp. Gerontol. 2020, 131, 110801. [Google Scholar] [CrossRef]
- Xu, J.; Wan, C.S.; Ktoris, K.; Reijnierse, E.M.; Maier, A.B. Sarcopenia Is Associated with Mortality in Adults: A Systematic Review and Meta-Analysis. Gerontology 2022, 68, 361–376. [Google Scholar] [CrossRef]
- Kaiser, H.; Yu, K.; Pandya, C.; Mendhe, B.; Isales, C.M.; McGee-Lawrence, M.E.; Johnson, M.; Fulzele, S.; Hamrick, M.W. Kynurenine, a Tryptophan Metabolite That Increases with Age, Induces Muscle Atrophy and Lipid Peroxidation. Oxid. Med. Cell. Longev. 2019, 2019, 9894238. [Google Scholar] [CrossRef]
- Dukes, A.; Davis, C.; El Refaey, M.; Upadhyay, S.; Mork, S.; Arounleut, P.; Johnson, M.H.; Hill, W.D.; Isales, C.M.; Hamrick, M.W. The Aromatic Amino Acid Tryptophan Stimulates Skeletal Muscle IGF1/P70s6k/MTor Signaling in Vivo and the Expression of Myogenic Genes in Vitro. Nutr. Burbank Los Angel. Cty. Calif 2015, 31, 1018–1024. [Google Scholar] [CrossRef]
- Bellanti, F.; Romano, A.D.; Lo Buglio, A.; Castriotta, V.; Guglielmi, G.; Greco, A.; Serviddio, G.; Vendemiale, G. Oxidative Stress Is Increased in Sarcopenia and Associated with Cardiovascular Disease Risk in Sarcopenic Obesity. Maturitas 2018. [Google Scholar] [CrossRef]
- Coto Montes, A.; Boga, J.A.; Bermejo Millo, C.; Rubio González, A.; Potes Ochoa, Y.; Vega Naredo, I.; Martínez Reig, M.; Romero Rizos, L.; Sánchez Jurado, P.M.; Solano, J.J.; et al. Potential Early Biomarkers of Sarcopenia among Independent Older Adults. Maturitas 2017, 104, 117–122. [Google Scholar] [CrossRef]
- Correia, J.C.; Ferreira, D.M.S.; Ruas, J.L. Intercellular: Local and Systemic Actions of Skeletal Muscle PGC-1s. Trends Endocrinol. Metab. TEM 2015, 26, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Agudelo, L.Z.; Femenía, T.; Orhan, F.; Porsmyr-Palmertz, M.; Goiny, M.; Martinez-Redondo, V.; Correia, J.C.; Izadi, M.; Bhat, M.; Schuppe-Koistinen, I.; et al. Skeletal Muscle PGC-1α1 Modulates Kynurenine Metabolism and Mediates Resilience to Stress-Induced Depression. Cell 2014, 159, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Schlittler, M.; Goiny, M.; Agudelo, L.Z.; Venckunas, T.; Brazaitis, M.; Skurvydas, A.; Kamandulis, S.; Ruas, J.L.; Erhardt, S.; Westerblad, H.; et al. Endurance Exercise Increases Skeletal Muscle Kynurenine Aminotransferases and Plasma Kynurenic Acid in Humans. Am. J. Physiol. Cell Physiol. 2016, 310, C836–840. [Google Scholar] [CrossRef] [PubMed]
- Al Saedi, A.; Chow, S.; Vogrin, S.; Guillemin, G.J.; Duque, G. Association Between Tryptophan Metabolites, Physical Performance, and Frailty in Older Persons. Int. J. Tryptophan Res. IJTR 2022, 15, 11786469211069952. [Google Scholar] [CrossRef]
- Ahmed, N.; Mandel, R.; Fain, M.J. Frailty: An Emerging Geriatric Syndrome. Am. J. Med. 2007, 120, 748–753. [Google Scholar] [CrossRef]
- Walston, J.; Hadley, E.C.; Ferrucci, L.; Guralnik, J.M.; Newman, A.B.; Studenski, S.A.; Ershler, W.B.; Harris, T.; Fried, L.P. Research Agenda for Frailty in Older Adults: Toward a Better Understanding of Physiology and Etiology: Summary from the American Geriatrics Society/National Institute on Aging Research Conference on Frailty in Older Adults. J. Am. Geriatr. Soc. 2006, 54, 991–1001. [Google Scholar] [CrossRef]
- Rockwood, K.; Mitnitski, A. Frailty in Relation to the Accumulation of Deficits. J. Gerontol. A. Biol. Sci. Med. Sci. 2007, 62, 722–727. [Google Scholar] [CrossRef]
- Morley, J.E.; Vellas, B.; van Kan, G.A.; Anker, S.D.; Bauer, J.M.; Bernabei, R.; Cesari, M.; Chumlea, W.C.; Doehner, W.; Evans, J.; et al. Frailty Consensus: A Call to Action. J. Am. Med. Dir. Assoc. 2013, 14, 392–397. [Google Scholar] [CrossRef]
- Fried, L.P.; Tangen, C.M.; Walston, J.; Newman, A.B.; Hirsch, C.; Gottdiener, J.; Seeman, T.; Tracy, R.; Kop, W.J.; Burke, G.; et al. Frailty in Older Adults: Evidence for a Phenotype. J. Gerontol. A. Biol. Sci. Med. Sci. 2001, 56, M146–156. [Google Scholar] [CrossRef]
- Clegg, A.; Young, J.; Iliffe, S.; Rikkert, M.O.; Rockwood, K. Frailty in Elderly People. Lancet Lond. Engl. 2013, 381, 752–762. [Google Scholar] [CrossRef]
- Valdiglesias, V.; Marcos-Pérez, D.; Lorenzi, M.; Onder, G.; Gostner, J.M.; Strasser, B.; Fuchs, D.; Bonassi, S. Immunological Alterations in Frail Older Adults: A Cross Sectional Study. Exp. Gerontol. 2018, 112, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Marcos-Pérez, D.; Sánchez-Flores, M.; Maseda, A.; Lorenzo-López, L.; Millán-Calenti, J.C.; Strasser, B.; Gostner, J.M.; Fuchs, D.; Pásaro, E.; Valdiglesias, V.; et al. Frailty Status in Older Adults Is Related to Alterations in Indoleamine 2,3-Dioxygenase 1 and Guanosine Triphosphate Cyclohydrolase I Enzymatic Pathways. J. Am. Med. Dir. Assoc. 2017, 18, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Jang, I.-Y.; Park, J.H.; Kim, J.H.; Lee, S.; Lee, E.; Lee, J.Y.; Park, S.J.; Kim, D.A.; Hamrick, M.W.; Kim, B.-J. The Association of Circulating Kynurenine, a Tryptophan Metabolite, with Frailty in Older Adults. Aging 2020, 12, 22253–22265. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Kynurenine pathway. One of the rate-limiting steps is mediated by IDO1, the enzyme responsible for obtaining extrahepatic kynurenine (KYN). NAD+ and kynurenic acid are the ending products.
Figure 1.
Kynurenine pathway. One of the rate-limiting steps is mediated by IDO1, the enzyme responsible for obtaining extrahepatic kynurenine (KYN). NAD+ and kynurenic acid are the ending products.
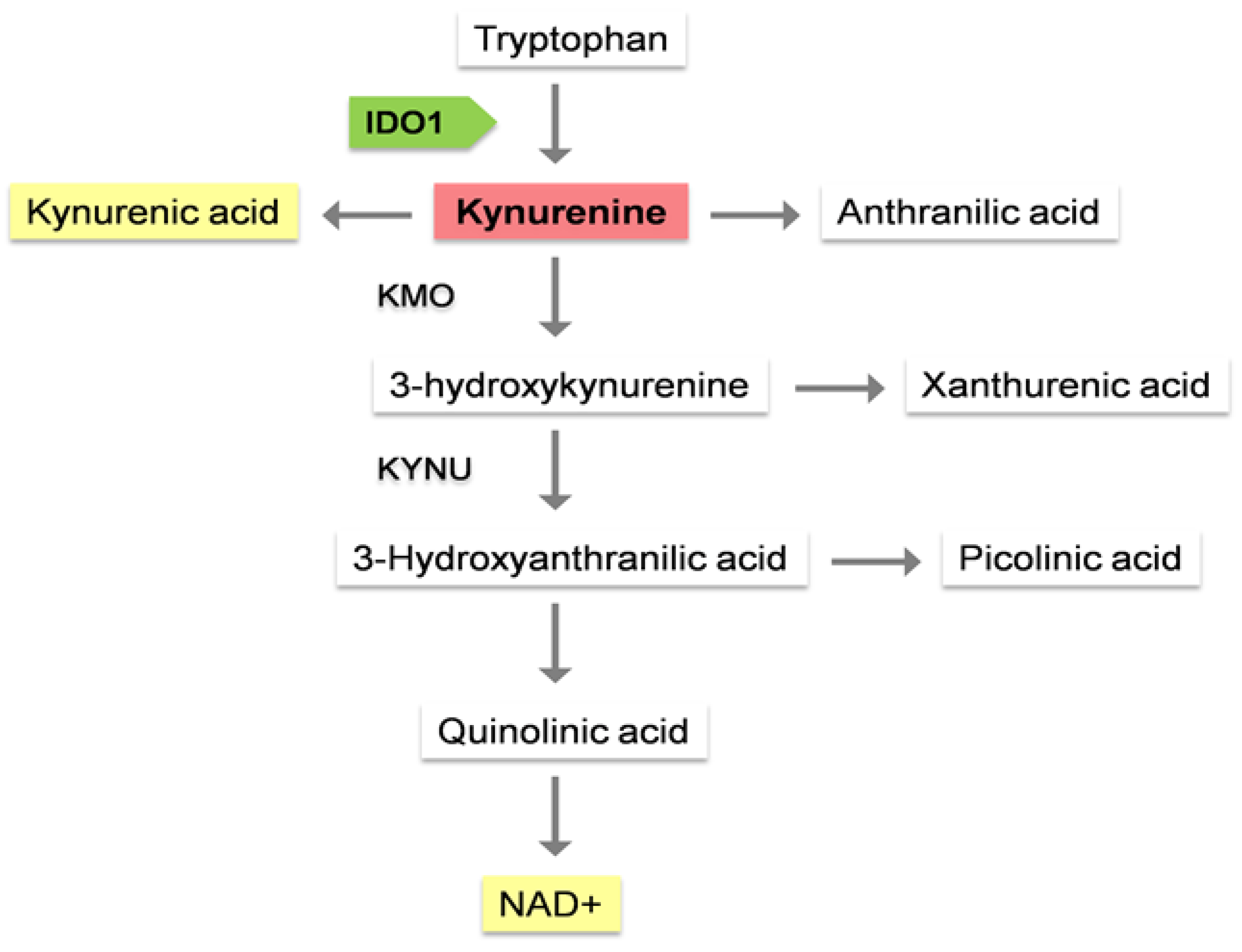
Figure 2.
Due to inflammaging, there is an increase of IL-6, IL-1β and IF-γ, which can induce IDO1 activity. As a result, low levels of Trp and high levels of KYN are observed. This would imply an alteration of metabolism with a greater tendency to catabolism as opposed to anabolism, which could explain part of the pathogenesis of some chronic diseases.
Figure 2.
Due to inflammaging, there is an increase of IL-6, IL-1β and IF-γ, which can induce IDO1 activity. As a result, low levels of Trp and high levels of KYN are observed. This would imply an alteration of metabolism with a greater tendency to catabolism as opposed to anabolism, which could explain part of the pathogenesis of some chronic diseases.
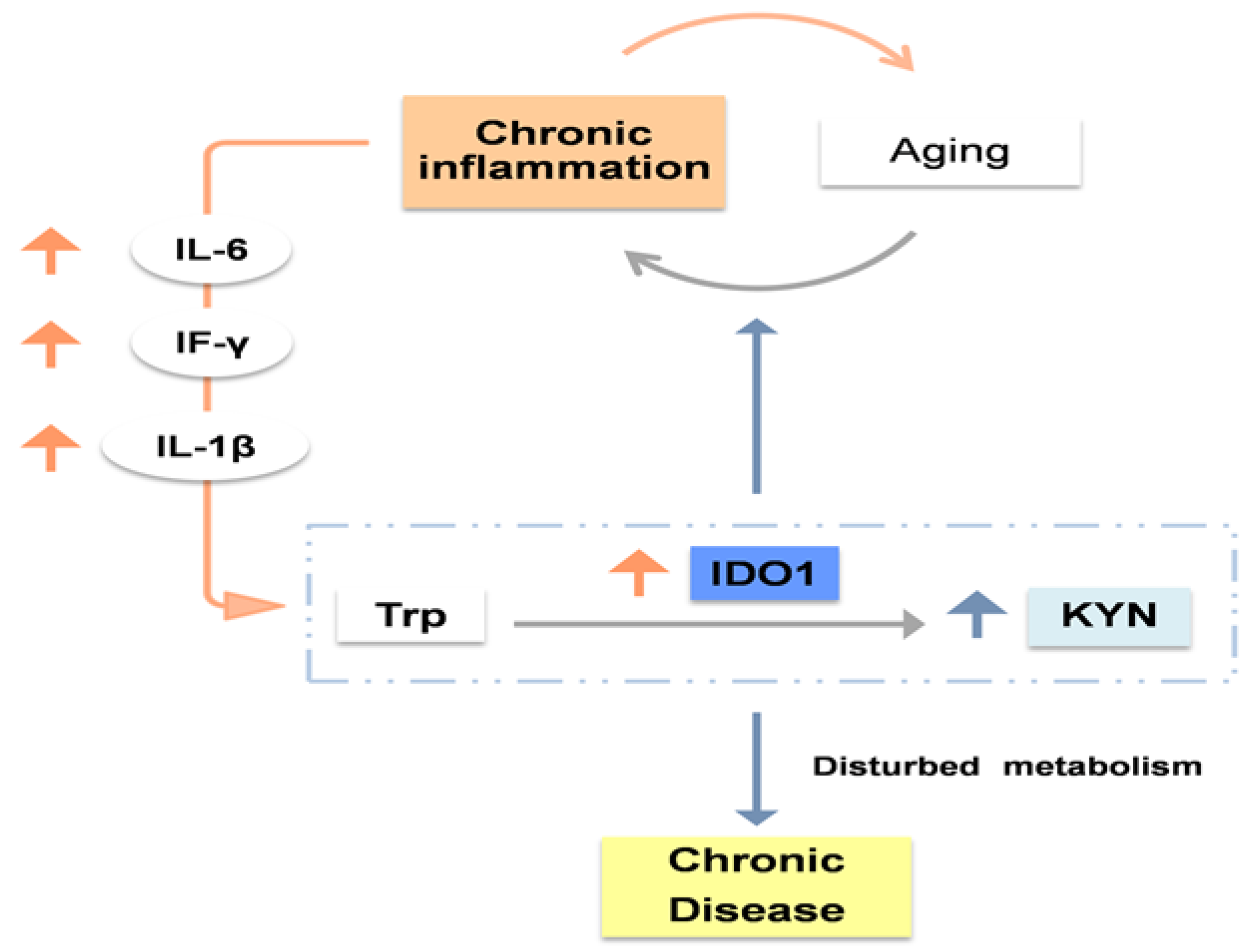
Figure 3.
High levels of KYNA, KYN, AA and 3-HK, as well as low levels of QUIN, PIC, 3-AA and NAD+ have been associated with osteoporosis, which increases fracture risk. This could be explained by the alteration in bone homeostasis. In this respect, low osteoblast activity, high osteoclast activity and disrupted bone marrow mesenchymal stem cells have been reported.
Figure 3.
High levels of KYNA, KYN, AA and 3-HK, as well as low levels of QUIN, PIC, 3-AA and NAD+ have been associated with osteoporosis, which increases fracture risk. This could be explained by the alteration in bone homeostasis. In this respect, low osteoblast activity, high osteoclast activity and disrupted bone marrow mesenchymal stem cells have been reported.
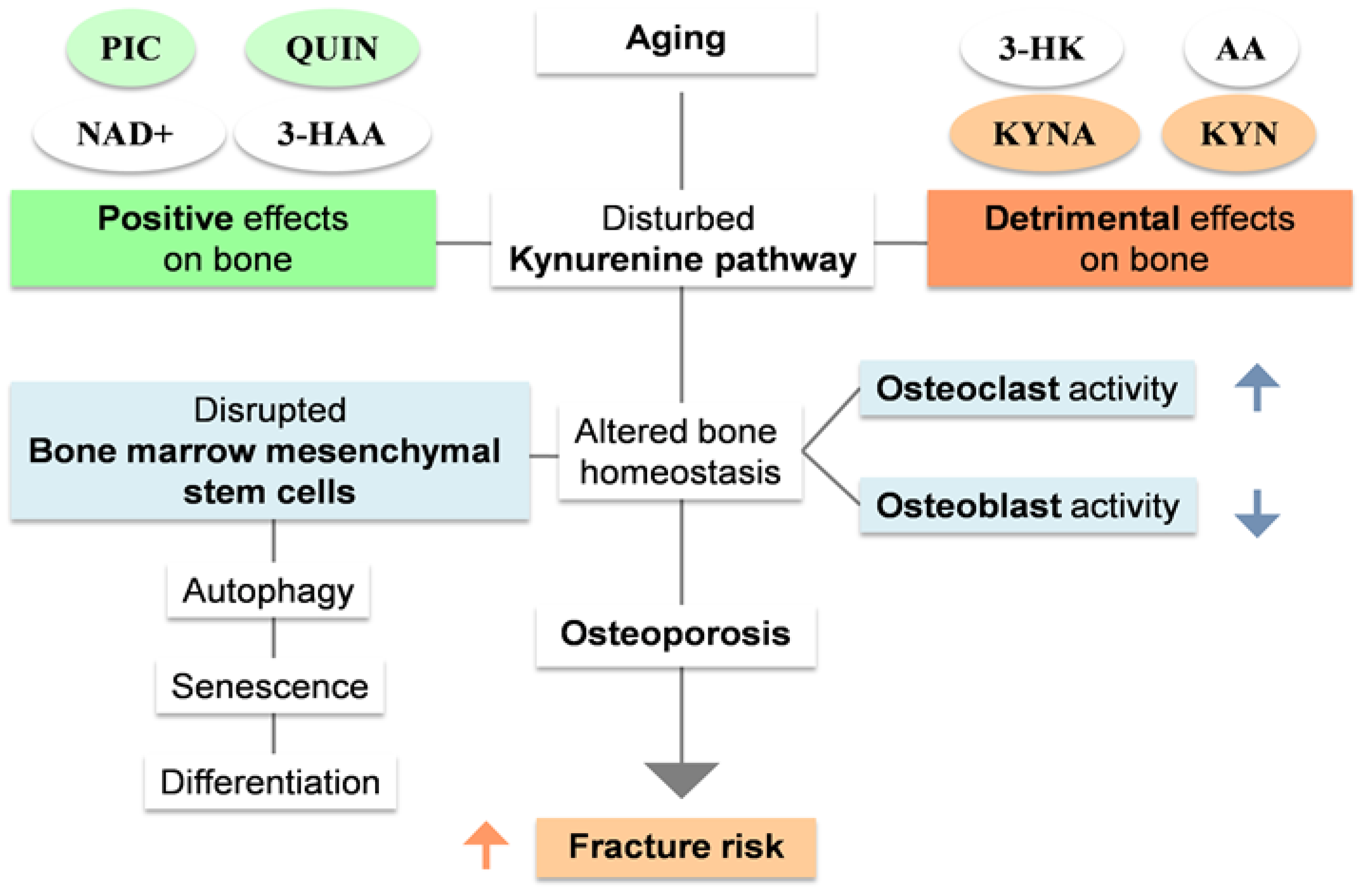
Figure 4.
High levels of 3-AA, QUIN, KYN and QUIN/KYN ratio as well as low levels of KYNA and KYNA/QUIN, KYNA/PIC, KYNA/KYN and PIC/QUIN ratios have been associated with sarcopenia. Low physical activity, one of its main components can modulate KP with less conversion of KYN to KYNA due low levels of PPARα/δ.
Figure 4.
High levels of 3-AA, QUIN, KYN and QUIN/KYN ratio as well as low levels of KYNA and KYNA/QUIN, KYNA/PIC, KYNA/KYN and PIC/QUIN ratios have been associated with sarcopenia. Low physical activity, one of its main components can modulate KP with less conversion of KYN to KYNA due low levels of PPARα/δ.
Figure 5.
Kynurenines serum levels have been shown to be altered in frail old persons.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



