
Submitted:
26 May 2023
Posted:
30 May 2023
You are already at the latest version
Abstract
The migratory life history of anadromous salmonids requires successful migration between nursery, feeding and spawning habitats. Smoltification is the major transformation anadromous salmonids undergo before migration to feeding areas and prepares juvenile fish for downstream migration and entry to seawater. We reviewed the effects of climate change on smolt ecology from growth of juveniles in freshwater to early post-smolts in sea. Shift in the suitable thermal conditions by climate change is causing Atlantic salmon to expand their range northwards, while in the southern edge of their distribution populations struggle with high temperatures and occasional droughts. Climatic conditions, particularly warmer temperatures, are affecting growth during freshwater phase in the river. Better growth in the northern latitudes leads to earlier smoltification. Thermal refuges, the areas of cooler water in the river, are especially important for salmonids impacted by climate change. Restoring and maintaining connectivity and suitable diverse mosaic habitat in rivers are important for survival and growth throughout the range. The start of the smolt migration has shifted earlier as a response to increasing water temperatures, which has led to concerns of mismatch with optimal conditions for post-smolts in the sea decreasing their survival. A wide smolt window allowing all migrating phenotypes from early to late migrant’s safe access to sea is important in changing environmental conditions. This is true also for regulated rivers, where flow regulation practices cause selection pressures on migrating salmonid phenotypes. Life history in freshwater affects also marine survival, and there is a need for better collaboration across life stages and habitats among researchers and managers to boost the smolt production in rivers.
Keywords:
Climate change
; salmonids
; Salmo
; rivers
; freshwater
; migration
1. Introduction
Anadromous and potamodromous salmonids migrate from their natal river to a feeding environment before returning for reproduction [1,2,3]. Migration enables fish to exploit many temporally productive and spatially discrete habitats with various fitness benefits (e.g., growth, reproduction, predator avoidance) [4]. Migratory life history requires unrestricted migration routes between nursery, feeding and spawning habitats [5]. During each life stage salmonids utilize the habitat that is advantageous for them, thus the migration between habitats clearly has adaptive value for the individuals [6]. Nursery and feeding habitats differ in environmental characteristics, and migrations precede adaptive physiological transformations and changes in the phenotype and behavior to be better suited in the new environment.
Smoltification is the major transformation that anadromous salmonids undergo before migration to feeding areas. Smolting prepares fish for downstream migration and entry to seawater. Atlantic salmon Salmo salar juveniles can stay in their natal river habitat for growth for 1–8 years before migrating [7,8]. Of the Pacific salmon, pink salmon Oncorhynchus gorbuscha and chum salmon Oncorhynchus keta can move almost directly after the emergence at fry-stage into sea water, while the others (masu salmon Oncorhynchus masou, O. tshawytscha, O. nerka and steelhead (rainbow) trout Oncorhynchus mykiss spend one or more years in fresh water before migrating to sea for feeding [7]. When smolting the phenotype of fish changes: coloration of smolts becomes silvery and body more streamlined [2]. This together with darkened fins, dark back and white abdomen helps fish to hide better in the pelagic environment [5]. Behavioral changes include loss of rheotaxis, and juveniles become more pelagic. Also, their tendency to group increases [9]. Several physiological changes take place, for example, increased salinity tolerance, increased metabolism, and the olfactory imprinting [10]. Environmental cues, photoperiod, temperature, and water flow, regulate physiological changes and initiate migration [2,5]. Smolting and seawater tolerance seem to develop for lake migrating populations also [11], but see [12], obviously due to genetic connections [13], although individuals spend their whole life cycle in freshwater.
Both Atlantic and Pacific salmon populations have been in decline throughout their range [13,14,15]. To turn this trend around, it is important to understand the role of different environmental and anthropogenic factors behind the decline [16]. Numerous effects have been found to influence the decline, and with the complex life history of migrating salmonids, the reasons for declines are obviously multiple and hard to unravel [17,18]. Anthropogenic activities have a long history of altering salmonid populations, thus also smolt development and smolt migration. Smolting as life stage is often more sensitive compared to other life stages [2]. Smolt development, behavior and survival during migration may be adversely affected by several anthropogenic activities, such as hydropower, land use, pollution, fish farming and parasites, like the sea lice Lepeophtheirus salmonis [2,5]. Climate change changes the environmental cues and interacts with the other anthropogenic pressures to affect smolting and smolt migration.
Historically climatic variability has affected patterns of abundance in Atlantic salmon and Pacific salmon populations [19,20,21,22]. Recently human exploitation, and especially climate change effects, have taken a larger impact on the declining stocks [13,17]. Although estuarine and marine mortality has been found to be an important determinant of survival, marine mortality is dependent on factors acting in freshwater and during smolt migration [23]. Thorstad et al. [24] argue that the best fundamental strategy to mitigate the changing environmental conditions should be to ensure that the highest number of wild smolts in the best condition migrate from rivers and coastal areas to feeding areas. Several factors, like the timing of migration and smolt size, must be in balance with the marine conditions for the successful fulfilment of the life cycle [18,23,25]. In addition to survival of smolts during migration and post-smolts during early arrival to sea, it is important to address the links between river habitat conditions and physiological requirements of salmonids during their juvenile life stages in freshwater habitats [26]. Climate change will continue to affect not only smolting and migration, but also instream habitats across all seasons [26].
In this paper we review climate change effects on 1) in-river habitat conditions preparatory for smolting, 2) smolting process, 3) smolt migration and 4) early post-smolt survival. Our focus is on Salmo spp., but when relevant we also refer to Pacific salmon (Onchorhynchus sp.).
2. Climate change and salmonids distribution
Human activities are estimated to have induced approximately 1.0°C of global warming above pre-industrial levels (from 1880 to 2017), with a likely range of 0.8°C to 1.2°C. Global warming is likely to reach 1.5°C at about 2030 if temperature continues to increase at the current rate [27]. For example, higher winter discharge, earlier snowmelt, and earlier onset of summer low flow periods are predicted throughout the range of Atlantic salmon [28,29].
Increasing global surface temperatures are very likely to lead to changes in precipitation and atmospheric moisture because of changes in atmospheric circulation, a more active hydrological cycle, and increases in the water-holding capacity throughout the atmosphere. Overall, global land precipitation has increased by about 2% since the beginning of the 20th century. There have been marked increases in precipitation in the latter part of the 20th century over northern Europe, however, with a general decrease southward to the Mediterranean. Dry wintertime conditions over southern Europe and the Mediterranean and wetter than normal conditions over many parts of northern Europe and Scandinavia [30] are linked to strong positive values of the North Atlantic Oscillation (NAO), with more anticyclonic conditions over southern Europe and stronger westerly winds over northern Europe reviewed by [31].
Northern Eurasia (north of approximately 40°N) showed widespread and statistically significant increases in winter precipitation during 1921–2015, with values exceeding 1.2–1.6 mm mo−1 per decade west of the Ural Mountains and along the east coast, while southern Europe exhibits coherent yet weaker amplitude drying trends that attain statistical significance over the eastern Mediterranean. These precipitation trends occur in the context of changes in the largescale atmospheric circulation, with negative SLP (Sea Level Pressure) trends over northern Eurasia and positive SLP trends over the central North Atlantic extending into southwestern Europe [32].
The magnitude of climate change is considered to depend on the atmospheric load of the two most important greenhouse gases, carbon dioxide (CO2) and methane (CH4). The terrestrial biosphere plays an important role in the global carbon balance. In boreal zones, forests and peatlands are an essential part of the global carbon cycle. Recent temperature increases have been associated with increasing forest fire activity in Canada since about 1970 and exceptionally warm summer conditions in Russia during the 2010 fire season reviewed by [33].
Atlantic salmon is distributed form northern Portugal (42°N) to River Kara in northern Russian in Europe [34], and West Atlantic salmon is distributed from Connecticut River to Ungava region of northern Quebec. Southern Atlantic salmon populations have declined dramatically and face the highest risk of extinction as global warming moves its thermal niche northwards [35]. Suitable thermal habitat for salmon is expected to extend northwards with the invasion of new spawning, nursery, and feeding areas north of the species’ present distributional range but with the loss of the most southern populations [36,37,38,39]. Indeed, salmon are already responding to warmer temperatures by expanding their range northwards into the Arctic Ocean [40,41] and disappearing from the southern edge of their distribution area [7,39,42,43]. The stock complex of Atlantic salmon in Europe has experienced a multidecadal decline in recruitment, resulting in the lowest stock abundances observed since 1970. Atlantic salmon abundance and productivity show similar patterns of decline across six widespread regions of North America [44]. Abundance declined in late 1980s and early 1990s after which it remained stable at low levels. Climate-driven environmental factors, as changes in plankton communities and prey availability in warmer ocean temperature were linked to low productivity of North Atlantic salmon populations [44]. Landlocked European populations of salmonids are found in Norway, Sweden, Finland and Russian Karelia [45,46,47,48]. The landlocked stocks of salmon have declined throughout their whole distribution range [48,49]. Brown trout (Salmo trutta) is native to Europe and Asia where anadromous populations are found from Portugal to the White Sea [7]. In future, the living conditions for trout will probably deteriorate in the southern part of the current distribution. In the northern part of their current distribution, global warming may improve feeding opportunities, growth and survival conditions [7]. According to Filipe et al. [50], brown trout distribution will become progressively and dramatically reduced in European watercourses in future. Their forecasts indicated that the greatest losses in suitable habitats will take place in the southern Europe.
3. In-river habitat conditions preparatory for smolting
Most important climate change -driven habitat changes that influence salmonid juveniles in rivers are changes in thermal and hydrological regimes [51,52,53]. These changes will affect how the juveniles use their physical habitat and affect growth and survival.
Water temperature has various effects on the biology of salmonids. Thermal optima allow salmon to maximize growth while temperatures above thermal optima can stress fish and finally lead to mortality [54,55]. Salmonids are cold water species; global warming will generally have a large impact on their success. On large-scale northern populations are being predicted to do better compared to southern populations under global warming [37,56,57], but even in the same river the effects on different populations can vary [55]. Some northern populations can have an increase in parr recruitment and smolt production [57]. However, also some Arctic salmonids are already experiencing warm (>21°C), physiologically challenging migratory river conditions [58], and an increase in river water temperatures has already been widely observed in several rivers [59,60,61]. In general, high latitude ecosystems are facing rapid warming, and cold-water fish will be eventually displaced by fish adapted to warmer water [62]. The range of temperatures in which fish survive or grow differs between development stages and salmonid species for review see [7]. For Atlantic salmon, for example, eggs survive around 0-16°C, and alevins can develop normally up to 22°C [63]. Growth takes place in temperatures between 6°C and 22.5°C, maximum growth around 16°C, and upper lethal temperature is 29.5°C for parr depending on the acclimation period. With the warming of surface waters, the risk of local extinctions increases [64,65].
Smolt characteristics are in many ways dependent on the factors acting in freshwater [23,66]. Temperature during the development in freshwater phase may have an impact on later behavior and life-history traits of salmonids. For example, incubation of eggs in higher temperatures have resulted in fry with reduced swimming performance or later returning adults [67,68]. According to Thompson & Beauchamp [69] survival of steelhead trout in the marine environment can be driven by an overall higher growth rate set early in life freshwater, which results in a larger size at smolt migration. Climate induced instream thermal conditions affect parr size and age of emigration from the river [70]. For salmonid populations facing increased water temperatures, thermal heterogeneity in the river plays an important role in survival and growth [52,71]. The density of juveniles in thermal refuges has been found to increase after high temperature effects [52]. Maintaining and restoring a diverse mixture of habitats and thermal refugia is important for salmonids impacted by climate change [72]. Thermal topology can also influence fish growth: fish in the least complex network grew faster and were ready to smolt earlier than fish in the more spatially complex temperature network [73]. Climate induced high-water temperatures can also interact with parr density: while on Chinook salmon at low parr density the effect of temperature on growth was positive, at high densities the relationship turned out to be negative [71].
Especially in the southern margins of the salmonid distribution ranges, the availability of suitable cold-water environments become more important as the temperatures rises [74]. The temporal variability of these cold refuges is high; the most stable ones typically are the groundwater seeps and cold-water tributaries [75]. For the cold-water species like salmonids, headwater streams may become more important structural and thermal refuges. In our experience, headwaters are in many cases less impacted by humans than the main streams. On the other hand, high-elevation streams, especially those above snowlines, can be especially vulnerable to climate change because they are likely to experience the greatest snow–rain transition [76]. Stream size is a limiting factor for some salmonid species, but for species like coho salmon differently sized streams can provide alternative rearing habitat [77]. For brown trout small streams are important spawning and nursery habitats [78,79]. Brown trout is well adapted and influenced by habitat variables associated with the size of small streams, especially with the flow variations [78,80], and the population traits of anadromous brown trout from a small stream differ from those in larger rivers [68]. Conservation of headwater stream habitats maintains and increases the variability in habitats and life history of salmonids to mitigate the effects from climate change.
Seasonal flow is another key element impacted by climate change contributing largely to the habitat quality of salmonid juveniles [81]. Climate change has already profoundly altered the hydrological regimes of rivers. The intense changes for Atlantic salmon and brown trout include frequent periods with extreme weather, i.e., low and high flow events, precipitation falling as rain and less as snow, and decrease in ice-covered period [7,82]. These changes can have a large negative impact on freshwater salmon instream habitat [76,83]. Extremes in water flow can decrease recruitment and survival. Generally, the early life stages, i.e., the eggs, emerging alevins, fry and young juveniles, experience the highest mortalities [84,85]. High flow events during the emergence of fry from the gravel can cause flushing of fry to unsuitable habitats. The preferences for physical habitat parameters, like water velocity and depth, vary seasonally [86]. Climate change induced high or low flows causes variation in this suitable habitat for salmonid parr. Low flow conditions are often associated also with extended duration of high water temperatures [83]. The minimum levels of river flow have been found as regulators for parr survival, and hence for smolt production in Atlantic salmon and brown trout [57,80]. Stream hydrology is also predicted to change when winters get warmer, and increased fluctuations in winter discharge and temperatures and may lead to repeated ice formation and breakup [87,88,89]. As winters get warmer, it means less snow, more rain and higher winter discharges. These changes can negatively affect growth and survival of juvenile salmonids during winter [90]. The ability of the young salmonids to swim against strong currents is poor at low temperatures [7,91], and salmonid parr prefer relatively slow flow rates in winter [92]. Increased rain on snow with high flow can lead to in ice scouring of the streambed, which results in higher egg mortality [93]. The mortality of salmonids eggs and fry can become higher along climate change in northern rivers.
Water temperature and flow variation, the two important aspects of climate change, are interacting with anthropogenic activities, such as the land-use in the catchment, to affect the fish community in rivers [94]. Anthropogenic activities have for a long time altered migratory fish by closing pathways and creating challenging migration conditions for smolts. Climate change can further strengthen these human caused effects. Climate change will intensify extreme precipitation and flood events over all climate regions [95], but difference at regional scale can be high [96]. Increased precipitation intensity enhances suspended solid and nutrient loadings in rivers especially in human altered catchments [97]. Increased rainfall with land use (i.e., forestry, agriculture) will intensify brownification of surface waters due to increased loading of dissolved organic carbon from the catchments [98,99]. This widespread phenomenon especially in boreal region will deteriorate habitat quality of salmonid juveniles habituated to good freshwater quality. Reduced freshwater habitat connectivity can decrease growth of juveniles and may have deleterious impacts on later marine life stages [100]. Flow regulation typically creates flow and temperature conditions for fish species that prefer warm- and slow-water habitats and can thus favor invasive species. Physically challenging migratory conditions caused by flow regulation together with large diurnal temperature fluctuations can restrict migration of salmonids by limiting their ability to recover from fatiguing exercise [58]. Rapid temperature raise also has negative effect on the osmoregulatory performance of the Atlantic salmon smolts [101]. Consequently, catchment scale conservation, including flow and connectivity restoration, is an important management priority to maintain and improve juvenile salmonid production, and thus smolt production. Conservating headwater stream habitats maintains and increases the variability in habitats and life history of salmonids to mitigate the effects from climate change.
4. Smolting
In salmonids, genetic diversity together with developmental flexibility leads numerous pathways to residency, migration or maturation [102] and, especially among Pacific salmon, there are also other phenotypes than smolts outmigrating rivers [103,104]. Anadromous salmonid juveniles transform from parr to smolts to prepare for downstream migration and entry to seawater. Physiological and behavioral changes take place in the spring when juvenile salmonids undergo smolting. Smolting and smolt migration are considered as a critical life-history stages essential for survival [5]. While still in fresh water, fish undergo a preparatory smolting process involving morphological changes as they become silvery and streamlined [2], (Figure 1). Behavioral changes include decrease in rheotactic and optomotor sensitivities and fishes station holding abilities decrease [9]. Photoperiod and temperature regulate physiological changes through their impact on the neuroendocrine system [2]. Thus, because the photoperiod remains the same at the same date and site each year, temperature will be critical in determining responses to future climate change. Within the same river system distance to the sea does not seem to play a role: populations closer or further from the sea smoltify the same time [105,106]. Water flow, and its variability as another major environmental factor, can act more as a timer, for example, to initiate migration [2,7].
Figure 1.
When smolting Atlantic salmon smolts become silvery and body streamlined (Photo: Ville Vähä).
Figure 1.
When smolting Atlantic salmon smolts become silvery and body streamlined (Photo: Ville Vähä).

Smoltification varies depending on several factors like temperature, latitude, age and size, growth rate, and a combination of these factors. Hence, climate change with raising water temperatures obviously has a substantial impact on the smoltification process. Size and growth potential affect the timing of migration, and also survival of smolts [25,107,108,109,110]. Smolt age depends on growth rate: fast-growing parr smoltify younger and smaller than slow-growing parr [111]. Warmer river temperatures increase growth of parr and increase the share of fish smolting at earlier age [70]. Temperature naturally correlates with latitude and is a strong predictor of migration timing in Atlantic salmon [112].
The migration decisions, whether to smoltify or not, are decided between internal and external factors [110,113]. The important external factors are the growth rate and the energetic status of individual fish [5]. Differences in smolting rate between naturally anadromous and more resident populations have an inherited component [113]. High growth rate in late summer and early autumn the year before migration predicts smoltification [5].
Growth and energetics do not solely depend on temperature, but also on other factors, like the food availability. For example, it appears that that smolting may be switched off by poor nutritional conditions preceding smoltification [113,114]. Better growth conditions caused by river temperature rise can increase the proportion of sexually mature male parr, which have lower probability to migrate [115].
How climate change will affect individual growth rates and energetics in salmonid populations will depend intimately on how it affects the ecological status of rivers, and particularly food availability. On a local scale controlling climate change drivers is not possible, therefore proactive measures are recommended against population declines [116]. These measures can include sustainable land use in the catchment and maintaining a diverse mosaic of habitats for salmonids [72,116]. Freshwater environment is especially vulnerable to climate change effects because it is already exposed to numerous anthropogenic pressures and water temperature and flow are highly climate-dependent [117,118]. Territorial salmonid parr also have limited capabilities to disperse as the environment changes.
Climate change can strengthen or weaken the effects of the anthropogenic activities on water quality important for smoltifying salmonids. Pollutants, acidity and sedimentation can adversely affect smolt development which can have negative consequences for their readiness for life at sea [10,23]. Especially in northern temperate coastal regions, which will receive higher winter rainfall, phosphorus loading from land to streams is expected to increase, whereas a decline in warm temperate and arid climates is expected [119]. In the northern region increasing precipitation increases nutrient leaching especially from areas affected by human alteration: agriculture, forestry and other land uses [120]. For example, acid leaks from the catchment are expected to increase. Increased acidity has a large impact on the fish community, especially to acid sensitive salmonids [121]. Even short moderate exposure of acidity may require more than two weeks for the recovery of Atlantic salmon smolts [122]. Freshwater ecosystems are sensitive to anthropogenic flow regime alteration, which may cause temperature fluctuations. Close to its southernmost distribution, warming together with low flows threatens coho salmon in California, and environmental flow protections is needed to support Pacific salmon in a changing climate [123]. Rapid temperature shifts have negative impact on hypo-osmoregulatory capacities of Atlantic salmon smolts [124]. There is an interaction of salinity and elevated temperature on osmoregulatory performance of salmon smolts, and rapid temperature fluctuations above the threshold temperature (20 °C) have been found to cause osmoregulatory failure.
5. Smolt migrations
Smolts start their downstream migration during a “period of readiness”, a smolt window when they are physiologically prepared to meet the conditions in their feeding area [2,125]. With the Atlantic salmon smolt, migration typically takes place during spring and early summer at length of 12–25 cm [126]. Temperature and flow are mostly mentioned environmental cues triggering the smolt migration. Migration times differ between years and rivers; temperature can be a good predictor for the timing [127,128,129]. Warmer temperatures result in earlier migrations [128]. Typically, a correlation between the onset of smolt emigration and the water temperature has been found [130]. Temperature experience, accumulated temperature, is the cue to initiate migration rather than any threshold temperature [128,131]. The initiation of the smolt migration was positively associated with freshwater temperatures up to about 10 °C and levelling off at higher values [18]. Another major environmental clue that plays an important role in initiating smolt migration is the river flow. During the smolt window increased water flow initiates smolt migration [126,130,132,133], but high flows have been found also to influence the opposite way by depressing migration [128,130]. Depending on the conditions, the relative influence of water temperature and flow in initiating migration can differ across the years [134]. Other environmental cues, like the photoperiod, have been found to control the initiation of downstream migration [135], but temperature and flow are the key environmental factors to be considered in response to climate change.
When ready, smolts lose their willingness to maintain station in a flow and start migrating downstream with the help of the current. The speed of the current has an influence on the downstream travel time, but smolts actively swim, typically following the mainstream in the surface water layer [7,115,136]. Smolts predominantly migrate at night, but later in the migration period this may change [2,5,137]. Smolts migrate downstream in schools of varying sizes. Relatively little is known about the formation of these groups, some results indicate solitary movement from natal streams followed by schooling further downstream [138]. A genetic component is involved as smolts in Atlantic salmon migrate more in kin-structured groups than with the unrelated individuals [139]. Some environmental factors, like light and dark variation can influence schooling [140].
The timing of migrations has been adapted by evolution to avoid unfavorable conditions and arriving when environmental conditions are suitable for survival and growth [4]. Mismatched timing would lead to decreased fitness: depleted food sources and/or increased predation. As described above the environment has an effect on migration timing, but it is somewhat determined by also genetics [141,142]. The relative contribution of genetic differences remains uncertain [143]. Under climate induced environmental changes different migrating phenological traits may be especially important to the future fitness of the species [126]. It is obvious that Atlantic salmon migration timing is already responding to warming temperatures: the initiation of the smolt seaward migration has occurred approximately 2.5 days earlier per decade throughout the basin of the North Atlantic [18].Accordingly, the analysis of long time series (1978–2008) on the timing of smolt migration of Atlantic salmon in the River Bush, Northern Ireland, revealed that earlier emigration periods are evident across the time series [144]. Kastl et al. [123] found out that an increase from 10.2 to 12.8C in mean seasonal water temperature accelerated the migration window by three weeks in coho salmon living near its southern distribution range in California, USA.
The earlier migration timing has given rise to growing concerns about smolts potentially missing the optimum environmental migration “window” [23]. Global warming affects also the receiving marine ecosystem by increasing surface sea water temperatures, and the results of this mismatch are difficult to predict. Climate change affects how and when species interact, potentially decoupling species interactions, and combining others, restructuring predator−prey interactions [145]. Some of these mismatches may lead to increased predation on smolts, or cause starvation, some may have no effect. Better understanding of how these interactions work is crucial to predict vulnerability to climate change effects.
Changed timing of smolt migration can lead to long-term changes to migratory phenotypes of salmonids e.g., [4]. A wide migration window with diversity of phenotypes can act as a safeguard against uncertainty in resource availability, buffering the variability in predator pressure or thermal mismatch. Survival of phenotypes can depend on the seasonally fluctuating conditions, such as thermal or hydrological circumstances affecting food availability, either directly or indirectly [146]. For example, Sturrock et al. [104] found that relative proportions of migrating phenotypes that contributed to the spawning population differed between the wet and dry year in Chinook salmon. In California’s Chinook salmon the late migrating phenotype dominated, but other strategies played an important role in many years [72]. Kennedy and Crozier [144] observed that marine survival of one sea winter Atlantic salmon were strongly influenced by the run timing, and during the observation period later emigrating cohorts demonstrated increased survival. In lake migrating sockeye salmon entering Lake Washington juveniles migrating later in the season encountered higher zooplankton abundance and warmer water but the optimal date for lake entry ranged across years by up to a month [146]. These examples show that the success of migratory phenotypes varies with environmental conditions. Warming of waters may highlight the importance of rare phenotypes in responding to climate change [72]. Therefore, the loss of phenotypic diversity can have a large impact on population persistence in a warming climate (Figure 2).
Figure 2.
Smolt trap in the River Tornionjoki, Finland, to catch the downstream migrating smolts and monitor their annual numbers and condition (Photo: Ville Vähä).
Figure 2.
Smolt trap in the River Tornionjoki, Finland, to catch the downstream migrating smolts and monitor their annual numbers and condition (Photo: Ville Vähä).

Anthropogenic use of freshwaters, especially flow regulation with dam construction, has resulted in large population declines and loss of salmon life history diversity [147,148]. Anthropogenic pressures, including climate change, affect the selection pressures in migratory salmonids, also on migrating smolt phenotypes. Hence it would be difficult to consider potential evolutionary responses to climate change without considering other human effects. For example, migration route selection at hydropower plant intake has been found to be consistent with phenotypes, and those traits selecting turbines could potentially be eliminated from the population due to high mortality [149]. To secure the adaptive variability of smolts safe downstream passage should be ensured at hydrodams, either by physical structures or long enough spill time windows. A long spill time is needed to protect the earliest and latest phenotypes migrating [129]. Low flows are expected to become more frequent especially in the southern distribution area of salmonids, and during low flows even small weirs can cause significant delays in smolt migration impacts [150]. River regulation practices are affected by the climate induced changes in temperature and precipitation depending on the region, and they may also change election pressures affecting salmonid populations. The examples above postulate that improved flow management is needed under climate change to avoid further loss of phenotypic diversity in salmonids.
Survival during migration and patterns of mortality has the potential to yield important insights into population bottlenecks [151]. Smolts are vulnerable to predators during their downstream migration in the surface layer. In the southern River Minho (Spain/Portugal) and in the River Endrick (Scotland) the mortality of Atlantic salmon smolts by avian and piscine predators was high, demonstrating that the amount of smolts lost in the river is likely to have strong constraint to these populations [151,152]. High in-river mortalities during downstream migration have been found also in Pacific salmon [153]. Climate change may create conditions that allow successful spreading of predators including invasive species [154,155,156]. For example, increasing predator populations of cormorants, (Phalacrocorax carbo sinensis) and an invasive terrestrial predator in Europe, American mink (Neovison vison) can cause elevated predation pressure on smolts [157,158,159]. Further studies quantifying the impact of invasive species and climate change on smolt migration in more detail are needed for future management considerations.
6. Early post-smolt survival
Most mortality between smolt and adult stages is generally considered to take place during the first year of life at sea when survival, maturation, and migration trajectories are being defined [39,160,161,162]. The first year of salmon at sea, known as the post-smolt year, is characterized by variable rates of mortality [163]. Mortality has been often considered to be highest during the first few months at sea [164,165]. Young salmonids are sensitive to variable climatic factors and to food availability [166,167,168]. Reduced marine survival is widely accepted as an important contributor to the observed salmon population declines in recent decades [24,39,169,170]. Ocean climate variability during the first springtime months of juvenile salmon migration to sea seems to be central to the survival of North American stocks, whereas summer climate variation appears to be important to adult recruitment variation for European stocks [163]. In the Baltic Sea, marine survival estimates of salmon post-smolts were negatively correlated with temperature [171]. The anticipated warming due to global climate change will impose thermal conditions on salmon populations outside historical context and challenge the ability of many populations to persist [163].
Salmon post-smolt survival rates in the sea can be influenced by the condition and quality of the smolts when they leave fresh water [23]. In such an effect, the impacts of a stressor experienced in riverine environment do not emerge until the fish has entered the post-smolt phase in the sea [24]. Over recent decades, juvenile salmon in many rivers have grown faster but left freshwater at a younger age, resulting in smaller smolts and increased mortality at sea [23]. In the Gulf of St. Lawrence, northwest Atlantic, the survival of sea entering small smolts was found inferior to large smolts [172]. Smolt size can also influence the subsequent growth rate of Atlantic salmon at sea, with larger smolts showing slower growth [173].
Timing of salmon smolt seaward migration and size of the smolts must be in balance with the marine conditions for the successful fulfilment of the life cycle [18,23,174]. The smolt seaward migration should coincide with optimal thermal conditions at sea to maximize survival [2,39,175], but climate change has advanced the timing of salmon smolt migration and created mismatch with optimal conditions for post-smolt growth and survival [18,144,176]. Higher temperature increases in rivers compared to marine habitats increase the risk of resource mismatches and high early post-smolt mortality [168,177], potentially causing smolts to miss the optimum environmental migration "window" [23]. Throughout the basin of the North Atlantic, salmon smolts are initiating their seaward migration 2.5 days earlier per decade in response to the climate change [18]. Observations on brown trout in the River Imsa, Norway, suggest that increased water temperature will induce seaward migration in early spring when sea growth and survival are poor [168].
Warmer temperatures in the North Atlantic have modified oceanic conditions, reducing the growth and survival of salmon by decreasing marine feeding opportunities [39,162,178,179]. Spring plankton blooms and therefore the peak of higher trophic resources available for salmon may be advanced in the season and may occur in different places [180,181,182], thus potentially creating a mismatch between salmon smolt migration and available resources [170,183]. A climate-driven shift in zooplankton community composition towards more temperature tolerant species with limited nutritional content may be associated with decreased marine survival and growth of salmon smolts [167].
7. Conclusions
Atlantic salmon are already experiencing and responding to climate change induced warmer water temperatures at different scales. On a large scale there are signs of salmon expanding their range northwards, as expected due to suitable thermal habitat shift, while the southern populations are struggling more due to high temperatures and periodic droughts. While increased temperature can have positive effects on the production of northern populations, by increasing growth leading to earlier smoltification, there are also concerns of increased water quality problems by water brownification and eutrophication particularly from human impacted land areas by the effects from increased rain especially during winter. The risk of local extinction of salmon populations has increased especially in the southern edge of salmon distribution. Especially the low flow events, typically associated with high water temperatures, are the population bottlenecks. Mitigating the effects of climate change on a local scale to increase smolt production are flow management and precautionary efforts to maintain and improve the ecological status of rivers. These measures are land use planning and restoration on a catchment scale to diminish loading from the catchment and conservation and restoration of instream habitats.
Smolt characteristics are in many ways dependent on the factors acting in freshwater, and these characteristics affect the post-smolt survival in the feeding area (Figure 3). Thermal heterogeneity in the river has a significant role in survival and growth, and we should have better knowledge of the magnitude and location of cold-water refuges in streams. Mapping of these areas with the modern technology would help in conservation work. Maintaining diverse mosaic of habitats and connectivity through conservation and restoration is crucial to mitigate the climate change effects in rivers.
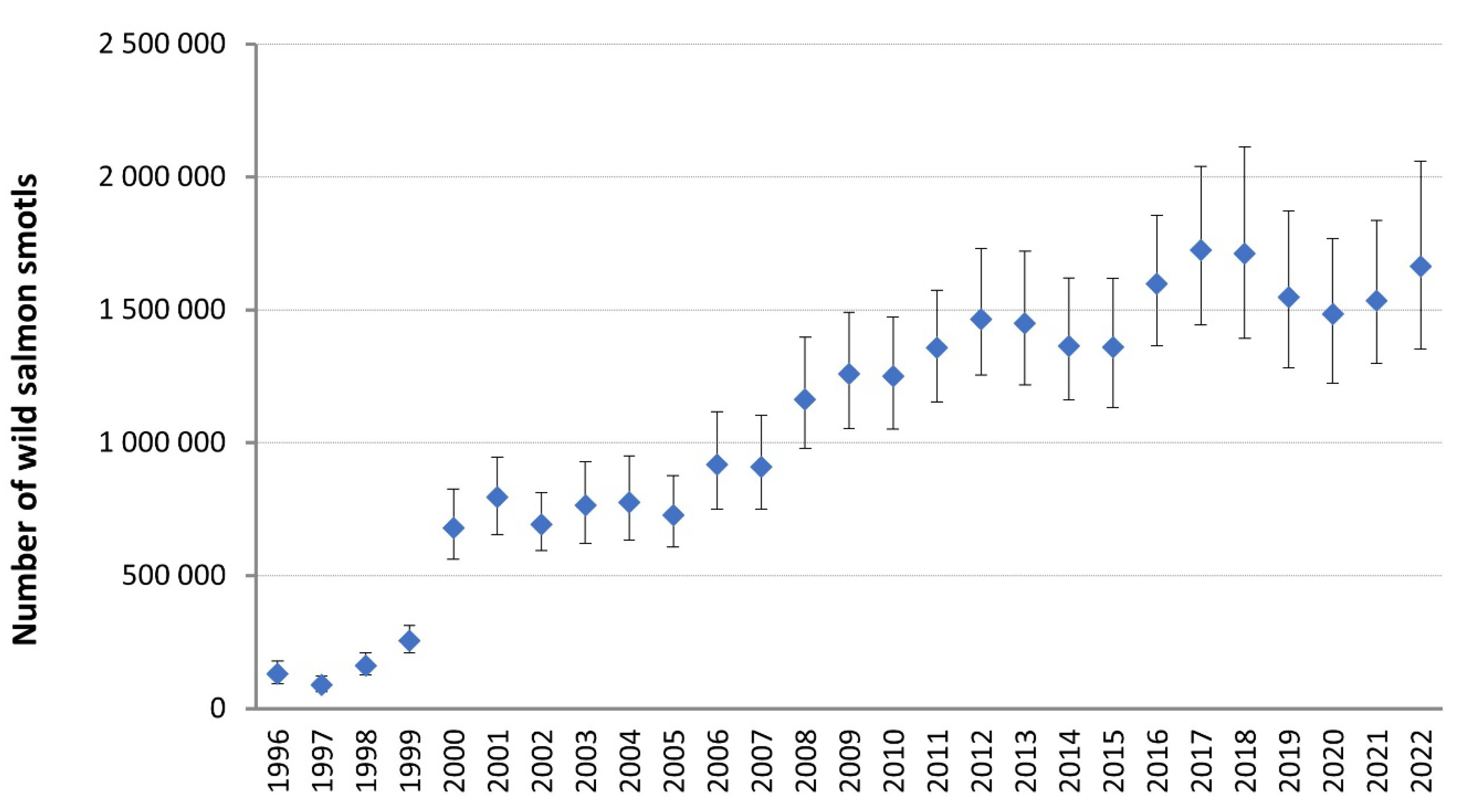
Figure 3.
Towards a better future: After successful management actions in the Baltic Sea and in the river itself the annual amount of smolts migrating from the River Tornionjoki has increased substantially [184].
Figure 3.
Towards a better future: After successful management actions in the Baltic Sea and in the river itself the annual amount of smolts migrating from the River Tornionjoki has increased substantially [184].
As a response to increasing temperatures an earlier migration timing of smolts is evident throughout the range of salmonids. This changes how and when species interact, and restructures also predator−prey interactions. To readjust to the changed, and still changing, conditions it is important to maintain as wide smolt-window as possible to allow all existing phenotypes, early or late migrants, to prevail. Under climate change different migrating phenological traits may be especially important to the future fitness of the species. This is especially important in regulated rivers where anthropogenic alteration of water flow creates not only increased mortality, but also artificial selection pressures on migrating smolts. This would mean, for example, longer spill water times or keeping the downstream routes open throughout the migration period.
Predation creates a substantial impact on migrating smolts and thus to the entire population. Climate change enhances the spreading of invasive species, including invasive predators, which can increase the total predation pressure on smolts. This emphasizes better control of invasive species, both prevention of their dispersal and better control of their populations.
Finally, we agree with earlier literature stating that there is urgent need for collaborations and research among scientists and managers across life cycle stages and ecosystems to address the research gaps [26], and that the basic strategy to protect salmonids against the effects of climate change should be to make sure that the maximum amount of wild smolts in the best condition leave from rivers e.g., [23,24].
References
- Dittman, A.H.; Quinn, T.P. Homing in Pacific Salmon: Mechanisms and Ecological Basis. J. Exp. Biol. 1996, 199, 83–91. [Google Scholar] [CrossRef]
- McCormick, S.D.; Hansen, L.P.; Quinn, T.P.; Saunders, R.L. 1998. Movement, migration, and smolting of Atlantic salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 1998, 55, 77–92. [Google Scholar] [CrossRef]
- Quinn, T.P.; Myers, K.W. Anadromy and the marine migrations of Pacific salmon and trout: Rounsefell revisited. Rev. Fish. Biol. Fish. 2004, 14, 421–442. [Google Scholar] [CrossRef]
- Lennox, R. J.; Chapman, J. M.; Souliere, C. M.; Tudorache, C.; Wikelski, M.; Metcalfe, J. D.; Cooke, S. J. Conservation physiology of animal migration. Conserv. Physio. 2016, 4, cov072. [Google Scholar] [CrossRef]
- Thorstad, E.B.; Whoriskey, F.; Uglem, I.; Moore, A.; Rikardsen, A.H.; Finstad, B. A critical life stage of the Atlantic salmon Salmo salar: behaviour and survival during the smolt and initial post-smolt migration. J. Fish Biol. 2012, 81, 500–542. [Google Scholar] [CrossRef]
- Lucas, M.C.; Baras, E.; Thom, T.J.; Duncan, D.; Slavík, O. Migration of Freshwater Fish. Blackwell Scientific Publishing. Oxford, UK, 2001, pp. 412.
- Jonsson, B.; Jonsson, N. A review of the likely effects of climate change on anadromous Atlantic salmon Salmo salar and brown trout Salmo trutta, with particular reference to water temperature and flow. J. Fish Biol. 2009, 75, 2381–447. [Google Scholar] [CrossRef]
- Erkinaro, J.; Czorlich, Y.; Orell, P.; Kuusela, J.; Falkegård, M.; Länsman, M.; Pulkkinen, H.; Primmer, C.R.; Niemelä, E. Life history variation across four decades in a diverse population complex of Atlantic salmon in a large subarctic river. Can. J. Fish. Aquat. Sci. 2018, 76, 42–55. [Google Scholar] [CrossRef]
- Veselov, A.E.; Kazakov, R.V.; Sysoyeva, M.I.; Bahmet, I.N. Ontogenesis of rheotactic and optomotor responses of juvenile Atlantic salmon. Aquaculture 1998, 168, 17–26. [Google Scholar] [CrossRef]
- Björnsson, B.T.; Stefansson, S.O.; McCormick, S.D. Environmental endocrinology of salmon smoltification. Gen. Comp. Endocrinol. 2011, 170, 290–298. [Google Scholar] [CrossRef]
- Piironen, J.; Kiiskinen, P.; Huuskonen, H.; Heikura-Ovaskainen, M.; Vornanen, M. Comparison of smoltification in Atlantic salmon (Salmo salar) from anadromous and landlocked populations under common garden conditions. Ann. Zool. Fenn. 2013, 50, 1–15. [Google Scholar] [CrossRef]
- Nilsen, T.O.; Ebbeson, L.O.E; Stefansson, S.O. Smolting in anadromous and landlocked strains of Atlantic salmon (Salmo salar). Aquaculture 2003, 222, 71–82. [Google Scholar] [CrossRef]
- Crozier, L.G.; Burke, B.J.; Chasco, B.E.; Widener, D.L.; Zabel, R.W. Climate change threatens Chinook salmon throughout their life cycle. Commun. Biol. 2021, 4, 222. [Google Scholar] [CrossRef]
- Olmos, M.; Massiot-Granier, F.; Prévost, E.; Chaput, G.; Bradbury, I.R.; Nevoux, M.; Rivot,E. Evidence for spatial coherence in time trends of marine life history traits of Atlantic salmon in the North Atlantic. Fish Fish. 2019, 20, 322–342. [Google Scholar] [CrossRef]
- Lehnert, S.J.; Kess, T.; Bentzen, P. M.; Kent, P.; Lien, S.; Gilbey, J.; Clément, M.; Jeffery, N.W.; Waples, R.S.; Bradbury, I.R. Genomic signatures and correlates of widespread population declines in salmon. Nat. Commun. 2019, 10, 2996. [Google Scholar] [CrossRef]
- Sobocinski, K.L.; Greene, C.M.; Anderson, J.H.; Kendall, N.W.; Schmidt, M.W.; Zimmerman, M.S.; Kemp, I.M.; Kim, S.; Ruff, C.P. A hypothesis-driven statistical approach for identifying ecosystem indicators of coho and Chinook salmon marine survival. Ecol. Indic. 2021, 124, 107403. [Google Scholar] [CrossRef]
- Dadswell, M.; Spares, A.; Reader, J.; McLean, M.; McDermott, T.; Samways, K.; Lilly, J. The Decline and Impending Collapse of the Atlantic Salmon (Salmo salar) Population in the North Atlantic Ocean: A Review of Possible Causes. Rev. Fish. Sci. Aquac. 2022, 30, 215–258. [Google Scholar] [CrossRef]
- Otero, J.; L'Abée-Lund, J.H.; Castro-Santos, T.; Leonardsson, K.; Storvik, G.O.; Jonsson, B.; Dempson, B.; Russell, I.C.; Jensen, A.J.; Baglinière, J.-L.; Dionne, M.; Armstrong, J.D.; Romakkaniemi, A.; Letcher, B.H.; Kocik, J.F.; Erkinaro, J.; Poole, R.; Rogan, G.; Lundqvist, H.; MacLean, J.C.; Jokikokko, E.; Arnekleiv, J.O.; Kennedy, R.J.; Niemelä, E.; Caballero, P.; Music, P.A.; Antonsson, T.; Gudjonsson, S.; Veselov, A.E.; Lamberg, A.; Groom, S.; Taylor, B.H.; Taberner, M.; Dillane, M.; Arnason, F.; Horton, G.; Hvidsten, N.A.; . Jonsson, I.R.; Jonsson, N.; McKelvey, S.; Næsje, T.F.; Skaala, O.; Smith, G.W.; Sægrov, H.; Stenseth, N.C.; Vøllestad, L.A. Basin-scale phenology and effects of climate variability on global timing of initial seaward migration of Atlantic salmon (Salmo salar). Glob. Chang. Biol. 2014, 20, 61–75. [Google Scholar] [CrossRef]
- Beamish, R.J.; Mahnken, C. A critical size and period hypothesis to explain natural regulation of salmon abundance and the linkage to climate and climate change. Prog. Oceanogr. 2001, 49, 423–437. [Google Scholar] [CrossRef]
- Huusko, A.; Hyvärinen, P. Atlantic salmon abundance and size track climate regimes in the Baltic Sea. Boreal Environ. Res. 2012, 17, 139–149. [Google Scholar]
- Sear, D.; Langdon, P.; Leng, M.; Edwards, M.; Heaton, T.; Langdon, C.; Leyland, J. Climate and human exploitation have regulated Atlantic salmon populations in the River Spey, Scotland, over the last 2000 years. The Holocene 2022, 32, 780–793. [Google Scholar] [CrossRef]
- Spanjer, A.R.; Gendaszek, A.S.; Wulfkuhle, E.J.; Black, R.W.; Jaeger, K.L. Assessing climate change impacts on Pacific salmon and trout using bioenergetics and spatiotemporal explicit river temperature predictions under varying riparian conditions. PLoS One 2022, 17, e0266871. [Google Scholar] [CrossRef]
- Russell, I.C.; Aprahamian, M.W.; Barry, J.; Davidson, I.C.; Fiske, P.; Ibbotson, A.T.; Kennedy, R.J.; Maclean, J.C.; Moore, A.; Otero, J.; Potter, E.C.E.; Todd, C.D. The influence of the freshwater environment and the biological characteristics of Atlantic salmon smolts on their subsequent marine survival. ICES J. Mar. Sci. 2012, 69, 1563–1573. [Google Scholar] [CrossRef]
- Thorstad, E.B.; Bliss, D.; Breau, C.; Damon-Randall, K.; Sundt-Hansen, L.E.; Hatfield, E.M.C.; Horsburgh, G.; Hansen, H.; Maoiléidigh, N. Ó,; Sheehan, T.; Sutton, S.G. 2021 Atlantic salmon in a rapidly changing environment—Facing the challenges of reduced marine survival and climate change. Aquatic Conserv.: Mar. Freshw. Ecosyst. 2021, 3, 2654–2665. [Google Scholar] [CrossRef]
- Jonsson, B.; Jonsson, M.; Jonsson, N. Optimal size at seaward migration in an anadromous salmonid. Mar. Ecol. Prog. Ser. 2016, 559, 193–200. [Google Scholar] [CrossRef]
- Gillis, C.-A.; Ouellet, V.; Breau, C.; Frechette, D.; Bergeron, N. Assessing climate change impacts on North American freshwater habitat of wild Atlantic salmon - urgent needs for collaborative research. Can. Water Resour. J. 2023. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2022: Impacts, Adaptation, and Vulnerability. In Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner H-O, Roberts D C, Tignor M, Poloczanska E S, Mintenbeck K, Alegría A, Craig M, Langsdorf S, Löschke S, Möller V, Okem A, Rama R, Eds; Cambridge University Press. Cambridge University Press, Cambridge, UK and New York, NY, USA, 2022, pp 3056.
- Thibeault, J.M.; Seth, A. Changing climate extremes in the Northeast United States: observations and projections from CMIP5. Clim. Change 2014, 127, 273–287. [Google Scholar] [CrossRef]
- Demaria, E. M. C.; Palmer, R.N.; Roundy, J.K. Regional Climate Change Projections of Streamflow in the Northeast and Midwest U.S. J. Hydrol. Reg. Stud. 2016, 5, 309–323. [Google Scholar] [CrossRef]
- Hanssen-Bauer, I.; Førland, E.J. Long-term trends in precipitation and temperature in the Norwegian Arctic: can they be explained by changes in the atmospheric circulation patterns. Clim. Res. 1998, 10, 143–153. [Google Scholar] [CrossRef]
- Dore, M.H.I. Climate change and changes in global precipitation patterns: What do we know? Environ. Int. 2005, 31, 1167–1181. [Google Scholar] [CrossRef]
- Guo, R.; Deser, C.; Terray, L.; Lehner, F. Human Influence on Winter Precipitation Trends (1921–2015) over North America and Eurasia Revealed by Dynamical Adjustment. Geophys. Res. Lett. 2019, 46, 3426–3434. [Google Scholar] [CrossRef]
- Terrier, A.; Martin, P.; Girardin, M.P.; Périé, C.; Legendre, P.; Bergeron, Y. Potential changes in forest composition could reduce impacts of climate change on boreal wildfires. Ecol. Appl. 2013, 23, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Kazakov, R. V. History and Atlantic salmon fishery state in Russia. In Atlanticheskiy losos; Kazakov R.V., Ed.; Nauka, Sankt-Petersburg, Soviet Union, 1998, pp. 335-380.
- Nicola, G.G.; Elvira, B.; Jonsson, B.; Ayllón, D.; Almodóvar, A. Local and global climatic drivers of Atlantic salmon decline in southern Europe. Fish. Res. 2018, 198, 78–85. [Google Scholar] [CrossRef]
- Todd, C.D.; Friedland, K.D.; MacLean, J.C.; Hazon, N.; Jensen, A.J. Getting into hot water? Atlantic salmon responses to climate change in freshwater and marine environments. In Atlantic salmon ecology; Aas, Ø., Klemetsen, A., Einum, S., Skurdal, J. Eds; Blackwell Publishing Ltd, Oxford, UK, 2011, pp 409-443.
- Hedger, R.D.; Sundt-Hansen, L.E.; Forseth, T.; Ugedal, O.; Diserud, O.H.; Kvambekk, Å.S.; Finstad, A.G. Predicting climate change effects on subarctic–Arctic populations of Atlantic salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 2013, 70, 159–168. [Google Scholar] [CrossRef]
- Hastings, R.A.; Rutterford, L.A.; Freer, J.J.; Collin, R.A.; Simpson, S.D.; Genner, M.J. Climate change drives poleward increases and equatorward declines in marine species. Curr. Biol. 2020, 30, 1572–1577. [Google Scholar] [CrossRef] [PubMed]
- Gillson, J.P.; Bašić,T. ; Davison, P.I.; Riley, W.D.; Talks, L.; Walker, A.M.; Russell, I.C. A review of marine stressors impacting Atlantic salmon Salmo salar, with an assessment of the major threats to English stocks. Rev. Fish. Biol. Fisheries 2022, 32, 879–919. [Google Scholar] [CrossRef]
- Jensen, A.J.; Karlsson, S.; Fiske, P.; Hansen, L.P.; Østborg, G.M.; Hindar, K. Origin and life history of Atlantic salmon (Salmo salar) near their northernmost oceanic limit. Can. J. Fish. Aquat. Sci. 2014, 71, 1740–1746. [Google Scholar] [CrossRef]
- Bilous, M.; Dunmall, K. Atlantic salmon in the Canadian Arctic: potential dispersal, establishment, and interaction with Arctic char. Rev. Fish Biol. Fish. 2020, 30, 463–483. [Google Scholar] [CrossRef]
- Parrish, D.L.; Behnke, R.J.; Gephard, S.R.; McCormick, S.D.; Reeves, G.H. Why aren’t there more Atlantic salmon (Salmo salar)? Can. J. Fish. Aquat. Sci. 1998, 55, 281–287. [Google Scholar] [CrossRef]
- Almodóvar, A.; Ayllón, D.; Nicola, G.G.; Jonsson, B.; Elvira, B. Climate-driven biophysical changes in feeding and breeding environments explain the decline of southernmost European Atlantic salmon populations. Can. J. Fish. Aquat. Sci. 2021, 76, 1581–1595. [Google Scholar] [CrossRef]
- Mills, K.E.; Pershing, A.J.; Sheehan, T.F.; Mountain, D. Climate and ecosystem linkages explain widespread declines in North American Atlantic salmon populations. Glob. Chang. Biol. 2013, 19, 3046–3061. [Google Scholar] [CrossRef]
- Berg, O.K. The formation of non-anadromous populations of Atlantic salmon, Salmo salar L., in Europe. J. Fish Biol. 1985; 27, 805–811. [Google Scholar]
- Ozerov, M.Y.; Veselov, A.J. , Lumme, J.; Primmer, C.R. Genetic structure of freshwater Atlantic salmon (Salmo salar L.) populations from the lakes Onega and Ladoga of northwest Russia and implications for conservation. Conserv. Genet. 2010, 11, 1711–1724. [Google Scholar] [CrossRef]
- Lumme, J.; Ozerov, M.Y.; Veselov, A.E.; Primmer, C.R. The Formation of Landlocked Populations of Atlantic Salmon. In Evolutionary Biology of the Atlantic Salmon; Vladic, T., Pettersson, E., Eds.; CRC Press, Boca Raton, USA, 2015, pp. 297.
- Kazakov, R.V. Distribution of Atlantic salmon, Salmo salar L., in freshwater bodies of Europe. Aquaculture Fish. Managem. 1992, 23, 461–475. [Google Scholar] [CrossRef]
- Leinonen, T.; Piironen, J.; Koljonen, M.-L.; Koskiniemi, J.; Kause, A. Restored river habitat provides a natural spawning area for a critically endangered landlocked Atlantic salmon population. PLoS One 2020, 15, e0232723. [Google Scholar] [CrossRef] [PubMed]
- Filipe, A.F.; Markovic, D.; Pletterbauer, F.; Tisseuil, C.; De Wever, A.; Schmutz, S.; Bonada, N.; Freyhof, J. Forecasting fish distribution along stream networks: brown trout (Salmo trutta) in Europe. Divers. Distrib. 2013, 19, 1059–1071. [Google Scholar] [CrossRef]
- Beaupré, J.; Boudreault, J.; Bergeron, N.E.; St-Hilaire, A. Inclusion of water temperature in a fuzzy logic Atlantic salmon (Salmo salar) parr habitat model. J. Therm. Biol. 2020, 87, 102471. [Google Scholar] [CrossRef] [PubMed]
- Corey, E.; Linnansaari, T.; Cunjak, R.A. High temperature events shape the broadscale distribution of juvenile Atlantic salmon (Salmo salar). Freshw. Biol. 2023, 63, 534–545. [Google Scholar] [CrossRef]
- Wilbur, N. M.; O’Sullivan, A.M.; MacQuarrie, K.T.B.; Linnansaari, T.; Curry, R.A. Characterizing physical habitat preferences and thermal refuge occupancy of brook trout (Salvelinus fontinalis) and Atlantic salmon (Salmo salar) at high river temperatures. Riv. Res. Appl. 2020, 36, 769–783. [Google Scholar] [CrossRef]
- Schreck, C.B.; Tort, L. The Concept of Stress in Fish. Fish Physiol. 2016, 35, 1–34. [Google Scholar]
- Zhang, X.; Li, H.-Y.; Deng, Z.D.; Leung, L.R.; Skalski, J.R.; Cooke, S.J. On the variable effects of climate change on Pacific salmon. Ecol. Modell. 2019, 397, 95–106. [Google Scholar] [CrossRef]
- Cunningham, C.J.; Westley, P.A.H.; Adkison, M.D. Signals of large scale climate drivers, hatchery enhancement, and marine factors in Yukon River Chinook salmon survival revealed with a Bayesian life history model. Glob. Chang. Biol. 2018, 24, 4399–4416. [Google Scholar] [CrossRef]
- Hvidsten, N.A.; Diserud, O.H.; Jensen, A.J.; Jensås, J.G.; Johnsen, B.O.; Ugedal, O. Water discharge affects Atlantic salmon Salmo salar smolt production: a 27 year study in the River Orkla, Norway. J. Fish Biol. 2015, 86, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.J.H.; Tierney, K.B. Warm northern river temperatures increase post-exercise fatigue in an Arctic migratory salmonid but not in a temperate relative. Func. Ecol. 2018, 32, 687–700. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Likens, G.E.; Jaworski, N.A.; Pace, M.L.; Sides, A.M.; Seekell, D.; Belt, K.T.; Secor, D.H.; Wingate, R.L. Rising stream and river temperatures in the United States. Front. Ecol. Environ. 2010, 8, 461–466. [Google Scholar] [CrossRef]
- Isaak, D.J.; Wollrab, S.; Horan, D.; Chandler, G. Climate change effects on stream and river temperatures across the northwest U.S. from 1980–2009 and implications for salmonid fishes. Clim. Change 2012, 113, 499–524. [Google Scholar] [CrossRef]
- Arevalo, E.; Maire, A.; Tétard, S.; Prévost, E.; Lange, F.; Marchand, F.; Josset, Q.; Drouineau, H. Does global change increase the risk of maladaptation of Atlantic salmon migration through joint modifications of river temperature and discharge? Proc. R. Soc. B. 2021, 288, 20211882. [Google Scholar] [CrossRef] [PubMed]
- Smalås, A.; Primicerio, R. ; Kahilainen, Terentyev, P.; Kashulin, N.; Zubova, E.; Amundsen, P-A. Increased importance of cool-water fish at high latitudes emerges from individual level responses to warming. Authorea. 29 January 2023. [Google Scholar] [CrossRef]
- Solomon, D.J.; Lightfoot, G.W. The Thermal Biology of Brown Trout and Atlantic Salmon; Science Reports, Environment Agency, Rio House, Waterside Drive, Aztec West, Almondsbury, Bristol, BS32 4UD, 2008, pp. 42.
- McGinnity, P.; Jennings, E.; deEyto, E.; Allott, N.; Samuelsson, P.; Rogan, G.; Whelan, K.; Cross. T. Impact of naturally spawning captive-bred atlantic salmon on wild populations: depressed recruitment and increased risk of climate-mediated extinction. Proc. R. Soc. B. 2009, 276, 3601–3610. [Google Scholar] [CrossRef] [PubMed]
- Piou, C.; Prevost, E. Contrasting effects of climate change in continental vs. oceanic environments on population persistence and microevolution of Atlantic salmon. Glob. Chang. Biol. 2013, 19, 711–723. [Google Scholar] [CrossRef]
- Burton, T.; McKelvey, S.; Stewart, D.C.; Armstrong, J.D.; Metcalfe, N.B. Offspring investment in wild Atlantic salmon (Salmo salar): relationships with smolt age and spawning condition. Ecol. Freshw. Fish 2013, 22, 317–332. [Google Scholar] [CrossRef]
- Burt, J.M.; Hinch, S.G.; Patterson, D.A. The importance of parentage in assessing temperature effects on fish early life history: A review of the experimental literature. Rev. Fish Biol. Fish. 2011, 21, 377–406. [Google Scholar] [CrossRef]
- Jonsson, B.; Jonsson, N. Egg incubation temperature affects the timing of the Atlantic salmon Salmo salar homing migration. J. Fish Biol. 2018, 93, 1016–1020. [Google Scholar] [CrossRef]
- Thompson, J.N.; Beauchamp, D. Size-Selective Mortality of Steelhead during Freshwater and Marine Life Stages Related to Freshwater Growth in the Skagit River, Washington. Trans. Am. Fish. Soc. 2014, 143, 910–925. [Google Scholar] [CrossRef]
- Jonsson, N.; Jonsson, B.; Hansen, L.P. Does climate during embryonic development influence parr growth and age of seaward migration in Atlantic salmon (Salmo salar)? Can. J. Fish. Aquat. Sci. 2005, 62, 2502–2508. [Google Scholar] [CrossRef]
- Crozier, L.G.; Zabel, R.W.; Hockersmith, E.E.; Achord, S. Interacting effects of density and temperature on body size in multiple populations of Chinook salmon. J. Anim. Ecol. 2010, 79, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Cordoleani, F.; Phillis, C.C.; Sturrock, A.M. FitzGerald, A.M.; Malkassian, A.; Whitman, G.E.; Weber, P.K.; Johnson, R.C. Threatened salmon rely on a rare life history strategy in a warming landscape. Nat. Clim. Chang. 2021, 11, 982–988. [Google Scholar] [CrossRef]
- Fullerton, A. H.; Burke, B.J.; Lawler, J.J.; Torgersen, C.E.; Ebersole, J.L.; Leibowitz, S.G. Simulated juvenile salmon growth and phenology respond to altered thermal regimes and stream network shape. Ecosphere 2017, 8, e02052. [Google Scholar] [CrossRef]
- Isaak, D.J.; Young. M.K. Cold-water habitats, climate refugia, and their utility for conserving salmonid fishes. Can. J. Fish. Aquat. Sci. 2023, e–First. [Google Scholar] [CrossRef]
- Dugdale, S.J.; Bergeron, N.E.; St-Hilaire, A. Temporal variability of thermal refuges and water temperature patterns in Atlantic salmon rivers. Remote Sensing of Environment 2013, 136, 358–373. [Google Scholar] [CrossRef]
- Battin, J.; Wiley, M.W.; Ruckelshaus, M.H.; Imaki, H. Projected impacts of climate change on salmon habitat restoration Proc. Natl. Acad. Sci. U.S.A. 2007, 104, 6720–6725. [Google Scholar] [CrossRef]
- Foley, K.M.; Rosenberger, S.A.; Mueter, F.J. Longitudinal Patterns of Juvenile Coho Salmon Distribution and Densities in Headwater Streams of the Little Susitna River, Alaska. Trans. Am. Fish. Soc. 2018, 147, 247–264. [Google Scholar] [CrossRef]
- Jonsson, B.; Jonsson, N.; Brodtkorb, E.; Ingebrigtsen, P.-J. Life-history traits of Brown Trout vary with the size of small streams. Func. Ecol. 2001, 15, 310–317. [Google Scholar] [CrossRef]
- Sutela, T.; Vehanen, T.; Jounela, P. Longitudinal patterns of fish assemblages in European boreal streams. Hydrobiologia 2020, 847, 3277–3290. [Google Scholar] [CrossRef]
- Jespersen, H.; Rasmussen, G.; Pedersen, S. Severity of summer drought as predictor for smolt recruitment in migratory brown trout (Salmo trutta). Ecol. Freshw. Fish 2021, 30, 115–124. [Google Scholar] [CrossRef]
- Mantua, N.; Tohver, I.; Hamlet, A. Climate change impacts on streamflow extremes and summertime stream temperature and their possible consequences for freshwater salmon habitat in Washington State. Clim. Change 2010, 102, 187–223. [Google Scholar] [CrossRef]
- Kang, D.H.; Gao, H.; Shi, X.; ul Islam, S.; Déry, S. J. Impacts of a rapidly declining mountain snowpack on streamflow timing in Canada’s Fraser River basin. Sci. Rep. 2016, 6, 19299. [Google Scholar] [CrossRef]
- Caissie, D. C.; Breau, J. H. ; Cameron,P. Water Temperature Characteristics within the Miramichi and Restigouche Rivers, Miramichi, New Brunswick, Canada. Canadian Science Advisory Secretariat, Research Document 2012/165.
- Hayes, D. S.; Moreira, M.; Boavida, I.; Haslauer, M.; Unfer, G.; Zeiringer, B.; Greimel, F.; Auer, S.; Ferreira, T.; Schmutz, S. Life stage-specific hydropeaking flow rules. Sustainability 2019, 11, 1547. [Google Scholar] [CrossRef]
- Huusko, A.; Greenberg, L.; Stickler, M.; Linnansaari, T.; Nykänen, M.; Vehanen, T.; Koljonen, S.; Louhi,P. ; Alfredsen, K. Life in the Ice Lane: A Review of the Ecology of Salmonids during Winter. River Res. Appl. 2007, 23, 469–491. [Google Scholar] [CrossRef]
- Mäki-Petäys, A.; Muotka, T.; Huusko, A.; Tikkanen, P.; Kreivi, P. Seasonal changes in habitat use and preference by juvenile brown trout, Salmo trutta, in a northern boreal river. Can. J. Fish. Aquat. Sci. 1997, 54, 520–530. [Google Scholar]
- Magnuson, J. J.; Robertson, D.M.; Benson, B.J.; Wynne, R.H.; Livingstone, D.M.; Arai, T.; Assel, R.A.; Barry, R.G.; Card, V.; Kuusisto, K.; Granin, N.G.; Prowse, T.D.; Stewart, K.M.; Vuglinski, V.S. Historical trends in lake and river ice cover in the Northern Hemisphere. Science 2000, 289, 1743–1746. [Google Scholar] [CrossRef] [PubMed]
- Pörtner, H. O.; Peck, M. A. Climate change effects on fishes and fisheries: Towards a cause-and-effect understanding. J. Fish Biol. 2010, 77, 1745–1779. [Google Scholar] [CrossRef] [PubMed]
- van Vliet, M. T. H.; Franssen, W. H. P.; Yearsley, J. R.; Ludwig, F.; Haddeland, I.; Lettenmaier, D. P.; Kabat, P. Global river discharge and water temperature under climate change. Glob. Environ. Change 2013, 23, 450–464. [Google Scholar] [CrossRef]
- Härkönen, L.; Louhi, P.; Huusko, R.; Huusko, A. Wintertime growth in Atlantic salmon under changing climate: the importance of ice cover for individual growth dynamics. Can. J. Fish. Aquat. Sci. 2021, 78, 1479–1485. [Google Scholar] [CrossRef]
- et al. 1996 Graham, W.D.; Thorpe, J.E. ; Metcalfe, N.B. Seasonal current holding performance of juvenile Atlantic salmon in relation to temperature and smolting. Can. J. Fish. Aquat. Sci. 1996, 53, 80–86. [Google Scholar]
- Enders, E.C.; Stickler, M.; Pennell, C.J.; Cote, D.; Alfredsen, K.; Scruton, D.A. Habitat use of Atlantic salmon parr (Salmo salar L. ) during winter. In Proceedings of 14th Workshop on the Hydraulics of Ice Covered Rivers, Quebec City, Canada, 2007; CGU HS Committee on River Ice Processes and the Environment, Canada, 2007, June 19 - 22; pp. 20–22.
- Cunjak, R.A.; Prowse, T.D.; Parrish, D.L. Atlantic salmon (Salmo salar) in winter: “the season of parr discontent”? Can. J. Fish. Aquat. Sci. 1999, 55, 161–180. [Google Scholar] [CrossRef]
- Comte, L.; Olden, J.D.; Tedesco, P.A.; Ruhi, A.; Giam, X. Climate and land-use changes interact to drive long-term reorganization of riverine fish communities globally. Proc. Natl. Acad. Sci. U.S.A. 2021, 118, e2011639118. [Google Scholar] [CrossRef]
- Tabari, H. Climate change impact on flood and extreme precipitation increases with water availability. Sci. Rep. 2020, 10, 13768. [Google Scholar] [CrossRef]
- Harp, R.D.; Horton, D.E. Observed Changes in Daily Precipitation Intensity in the United States. Geophys. Res. Lett. 2022, 49, e2022GL099955. [Google Scholar] [CrossRef]
- Cronin, L.; Regan, F.; Lucy, F.E. Detection of transient pollution events in an Irish river catchment in the context of increasing frequency and intensity of rainfall events due to climate change. EGU General Assembly 2023, EGU23, 15993. [Google Scholar]
- Weyhenmeyer, G.; Prairie, Y.T.; Tranvik, L.J. Browning of Boreal Freshwaters Coupled to Carbon-Iron Interactions along the Aquatic Continuum. PLoS One 2014, 9, e88104. [Google Scholar] [CrossRef]
- Kritzberg, E.S.; Hasselquist, E.M.; Škerlep, M. Stefan Löfgren, S.; Olsson, O.; Stadmark, J.; Valinia, S.; Hansson, L.-A.; Laudon, H. Browning of freshwaters: Consequences to ecosystem services, underlying drivers, and potential mitigation measures. Ambio 2020, 49, 375–390. [Google Scholar] [CrossRef]
- Sethi, S.A.; Carey, M.P.; Gerken, J.; Harris, B.P.; Wolf, N.; Cunningham, C.; Restrepo, F.; Ashline, J. Juvenile salmon habitat use drives variation in growth and highlights vulnerability to river fragmentation. Ecosphere 2022, 13, e4192. [Google Scholar] [CrossRef]
- Vargas-Chacoff, L.; Regish, A.M.; Weinstock, A.; McCormick, S.D. Effects of elevated temperature on osmoregulation and stress responses in Atlantic salmon Salmo salar smolts in fresh water and seawater. J. Fish Biol. 2018, 93, 550–559. [Google Scholar] [CrossRef]
- Thorpe, J.E. Maturation responses of salmonids to changing developmental opportunities. Mar. Ecol. Prog. Ser. 2007, 335, 285–288. [Google Scholar] [CrossRef]
- Copeland, T.; Venditti, D.A. Contribution of three life history types to smolt production in a Chinook salmon (Oncorhynchus tshawytscha) population. Can. J. Fish. Aquat. Sci. 2009, 66, 1658–1665. [Google Scholar] [CrossRef]
- Sturrock, A.M.; Wikert, J.D.; Heyne, T.; Mesick, C.; Hubbard, A.E.; Hinkelman, T.M.; Weber, P.K.; Whitman, G.E.; Glessner, J.G.; Johnson, R.C. Reconstructing the Migratory Behavior and Long-Term Survivorship of Juvenile Chinook Salmon under Contrasting Hydrologic Regimes. PLoS One 2015, 10, e0122380. [Google Scholar] [CrossRef]
- Bassett, M.C.; Patterson, P.A.; Shrimpton, J.M. Temporal and spatial differences in smolting among Oncorhynchus nerka populations throughout fresh and seawater migration J. Fish Biol. 2018, 93, 510–518. [Google Scholar] [CrossRef]
- Elsner, R.A.; Shrimpton, J.M. Is the duration of the smolt window related to migration distance in coho salmon Oncorhynchus kisutch? J. Fish Biol. 2018, 93, 501–509. [Google Scholar] [CrossRef]
- Bohlin, T.; Dellefors, C.; Faremo, U. Optimal Time and Size for Smolt Migration in Wild Sea Trout (Salmo trutta). Can. J. Fish. Aquat. Sci. 2011, 50, 224–232. [Google Scholar] [CrossRef]
- Jutila, E.; Jokikokko, E.; Julkunen. M. Long-term changes in the smolt size and age of Atlantic salmon, Salmo salar L., in a northern Baltic river related to parr density, growth opportunity and postsmolt survival. Ecol. Freshw. Fish 2006, 15, 321–330. [Google Scholar] [CrossRef]
- Gregory, S.D.; Ibbotson, A.T.; Riley, W.D.; Nevoux, M.; Lauridsen, R.B.; Russell, I.A.; Britton, J.R.; Gillingham, P.K.; Simmons, O.M.; Rivot, E. Atlantic salmon return rate increases with smolt length. ICES J. Mar. Sci. 2019, 76, 1702–1712. [Google Scholar] [CrossRef]
- Debes, P.V.; Piavchenko, N.; Erkinaro, J.; Primmer, C.R. Genetic growth potential, rather than phenotypic size, predicts migration phenotype in Atlantic salmon. Proc. R. Soc. B 2020, 287, 20200867. [Google Scholar] [CrossRef]
- Økland, F.; Jonsson, B.; Jensen, A.J.; Hansen, L.P. Is there a threshold size regulating seaward migration of brown trout and Atlantic salmon? J. Fish Biol. 1993, 42, 541–550. [Google Scholar] [CrossRef]
- Vollset, K.W.; Lennox, R.J.; Lamberg, A.; Skaala, Ø.; Sandvik, A.D.; Sægrov, H.; Kvingedal, E.; Kristensen, T.; Jensen, A.J.; Haraldstad, T.; Barlaup, B.T.; Ugedal, O. Predicting the nationwide outmigration timing of Atlantic salmon (Salmo salar) smolts along 12 degrees of latitude in Norway. Divers. Distrib. 2021, 27, 1383–1392. [Google Scholar] [CrossRef]
- Archer, L.C.; Hutton, S.A.; Harman, L.; Michael, N.; O'Grady, M.N.; Kerry, J.P.; Poole, W.R.; Gargan, P.; McGinnity, P.; Reed, T.E. The Interplay Between Extrinsic and Intrinsic Factors in Determining Migration Decisions in Brown Trout (Salmo trutta): An Experimental Study. Front. Ecol. Evol. 2019, 7, 222. [Google Scholar] [CrossRef]
- Jones, D.A.; Bergman, E.; Greenberg, L. Food availability in spring affects smolting in brown trout (Salmo trutta). Can. J. Fish. Aquat. Sci. 2015, 72, 1694–1699. [Google Scholar] [CrossRef]
- Fängstam, H.; Berglund, I.; Sjöberg, M.; Lundqvist, H. Effects of size and early sexual maturity on downstream migration during smolting in Baltic samon (Salmo salar). J. Fish Biol. 1993, 43, 517–529. [Google Scholar] [CrossRef]
- Nelson, K.C.; Palmer, M.A.; Pizzuto, J.E.; Moglen, G.E.; Angermeier, P.L.; Hilderbrand, R.H.; Dettinger, M.; Katharine Hayhoe, K. Forecasting the combined effects of urbanization and climate change on stream ecosystems: from impacts to management options J. Appl. Ecol. 2009, 46, 154–163. [Google Scholar] [CrossRef]
- Capon, S.J.; Stewart-Koster,B. ; Bunn, S.E. Future of Freshwater Ecosystems in a 1.5°C Warmer World. Front. Environ. Sci. 2021, 9, 784642. [Google Scholar] [CrossRef]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: impacts across multiple levels of organization. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2093–2106. [Google Scholar] [CrossRef]
- Jeppesen, E.; Kronvang, B.; Meerhoff, M.; Søndergaard, M.; Hansen, K.M.; Andersen, H.E.; Lauridsen, T.L.; Liboriussen, L.; Beklioglu, M.; Ozen, A.; Olesen, J.E. Climate change effects on runoff, catchment phosphorus loading and lake ecological state, and potential adaptations. J. Environ. Qual. 2009, 38, 1930–41. [Google Scholar] [CrossRef]
- Pihlainen, P.; Zandersen, M.; Hyytiäinen, K.; Andersen, H.E.; Bartosova, A.; Gustafsson, B.; Jabloun, M.; McCrackin, M.; Meier, H.E.M.; Olesen, J.E.; Saraiva, S.; Swaney, D.; Thodsen, H. Impacts of changing society and climate on nutrient loading to the Baltic Sea. Sci. Total Environ. 2020, 731, 138935. [Google Scholar] [CrossRef]
- Vehanen, T.; Sutela, T.; Aroviita, J.; Karjalainen, S.M.; Riihimäki, J.; Larsson, A.; Vuori, K.-M. Land use in acid sulphate soils degrades river water quality -Do the biological quality metrics respond? Ecol. Indic. 2022, 141, 109085. [Google Scholar] [CrossRef]
- Nilsen, T.O.; Ebbesson, L.O.; Handeland, S.O.; Kroglund, F.; Finstad, B.; Angotzi, A.R.; Stefansson. S.O. Atlantic salmon (Salmo salar L.) smolts require more than two weeks to recover from acidic water and aluminium exposure. Aquat. Toxicol. 2013, 142-143, 33–44. [Google Scholar] [CrossRef]
- Kastl, B.; Obedzinski, M.; Carlson, S. M.; Boucher, W.T.; Grantham, T.E. Migration in drought: Receding streams contract the seaward migration window of endangered salmon. Ecosphere 2022, 13, e4295. [Google Scholar] [CrossRef]
- Bernard, B.; Mandiki, S.N.M.; Duchatel, V.; Rollin, X.; Kestemont, P. A temperature shift on the migratory route similarly impairs hypo-osmoregulatory capacities in two strains of Atlantic salmon (Salmo salar L.) smolts. Fish Physiol. Biochem. 2019, 45, 1245–1260. [Google Scholar] [CrossRef]
- Hansen, L.P.; Jonsson, B. Salmon ranching experiments in the River Imsa: Effect of timing of Atlantic salmon (Salmo salar) smolt migration on survival to adults. Aquaculture 1989, 82, 367–373. [Google Scholar] [CrossRef]
- Bjerck, H.B.; Urke, H.A.; Haugen, T.O.; Alfredsen, J.A.; Ulvund, J.B.; Kristensen, T. Synchrony and multimodality in the timing of Atlantic salmon smolt migration in two Norwegian fjords. Sci. Rep. 2021, 11, 6504. [Google Scholar] [CrossRef]
- Achord, S.; Zabel, R.W.; Sandford, B.P. Migration Timing, Growth, and Estimated Parr-to-Smolt Survival Rates of Wild Snake River Spring–Summer Chinook Salmon from the Salmon River Basin, Idaho, to the Lower Snake River. Trans. Am. Fish. Soc. 2007, 136, 142–154. [Google Scholar] [CrossRef]
- Sykes, G.E.; Johnson, C.J.; Shrimpton, J.M. Temperature and Flow Effects on Migration Timing of Chinook Salmon Smolts. Trans. Am. Fish. Soc. 2009, 138, 1252–1265. [Google Scholar] [CrossRef]
- Frechette, D.M.; Hawkes, J.P.; Kocik, J.F. Managing for Atlantic Salmon Smolt Run Timing Variability in a Changing Climate. N. Am. J. Fish. Manag. 2023, 43, 517–538. [Google Scholar] [CrossRef]
- Antonsson, T.; Gudjonsson, S. Variability in Timing and Characteristics of Atlantic Salmon Smolt in Icelandic Rivers. Trans. Am. Fish. Soc. 2002, 131, 643–655. [Google Scholar] [CrossRef]
- Zydlewski, G.B.; Haro, A.; McCormick, S.D. Evidence for cumulative temperature as an initiating and terminating factor in downstream migratory behavior of Atlantic salmon (Salmo salar) smolts. Can. J. Fish. Aquat. Sci. 2005, 62, 68–78. [Google Scholar] [CrossRef]
- Hvidsten, N. A.; Jensen, A. J.; Vivås, H.; Bakke, Ø. Downstream migration of Atlantic salmon smolts in relation to water flow, water temperature, moon phase and social interaction. Nord. J. Freshw. Res. 1995, 70, 38–48. [Google Scholar]
- Persson, L.; Kagervall, A.; Leonardsson, K.; Royan, M.; Alanärä, A. The effect of physiological and environmental conditions on smolt migration in Atlantic salmon Salmo salar. Ecol. Freshw. Fish 2019, 28, 190–199. [Google Scholar] [CrossRef]
- Harvey, A.C.; Glover, K.A.; Wennevik, V.; Skaala, Ø. Atlantic salmon and sea trout display synchronised smolt migration relative to linked environmental cues. Sci. Rep. 2020, 10, 3529. [Google Scholar] [CrossRef]
- Zydlewski, G.B.; Stich, D.S.; McCormick, S.D. Photoperiod control of downstream movements of Atlantic salmon Salmo salar smolts. J. Fish Biol. 2014, 85, 1023–1041. [Google Scholar] [CrossRef]
- Karppinen, P.; Hynninen, M.; Vehanen, T.; Vähä, J.-P. Variations in migration behaviour and mortality of Atlantic salmon smolts in four different hydroelectric facilities. Fish. Manag. Ecol. 2021, 28, 253–267. [Google Scholar] [CrossRef]
- Aarestrup, K.; Nielsen, C.; Koed, A. Net ground speed of downstream migrating radio-tagged Atlantic salmon (Salmo salar L.) and brown trout (Salmo trutta L.) smolts in relation to environmental factors. Hydrobiologia 2002, 483, 95–102. [Google Scholar] [CrossRef]
- Riley, W.D. Seasonal downstream movements of juvenile Atlantic salmon, Salmo salar L., with evidence of solitary migration of smolts. Aquaculture 2007, 273, 194–199. [Google Scholar] [CrossRef]
- Olsén, K.H.; Petersson, E.; Ragnarsson, B.; Lundqvist, H.; Järvi, T. Downstream migration in Atlantic salmon (Salmo salar) smolt sibling groups. Can. J. Fish. Aquat. Sci. 2011, 61, 328–331. [Google Scholar] [CrossRef]
- Kemp, P.S.; Williams, J.G. Illumination influences the ability of migrating juvenile salmonids to pass a submerged experimental weir. Ecol. Freshw. Fish 2009, 18, 297–304. [Google Scholar] [CrossRef]
- Nielsen, C.; Holdensgaard, G.; Petersen, H.C.; Björnsson, B. T.; Madsen, S.S. Genetic differences in physiology, growth hormone levels and migratory behaviour of Atlantic salmon smolts. J. Fish Biol. 2001, 59, 28–44. [Google Scholar] [CrossRef]
- Stewart, D. C.; Middlemas, S.J.; Youngson, A.F. Population structuring in Atlantic salmon (Salmo salar): Evidence of genetic influence on the timing of smolt migration in sub-catchment stocks. Ecol. Freshw. Fish 2006, 15, 552–558. [Google Scholar] [CrossRef]
- Crozier, L.G.; Hendry, A.P.; Lawson, P.W.; Quinn, T.P; Mantua, N.J.; Battin, J.; Shaw, R.G.; Huey, R.B. Potential responses to climate change in organisms with complex life histories: evolution and plasticity in Pacific salmon. Evol. Appl. 2008, 1, 252–270. [Google Scholar] [CrossRef]
- Kennedy, R.J.; Crozier, W.W. Evidence of changing migratory patterns of wild Atlantic salmon Salmo salar smolts in the River Bush, Northern Ireland, and possible associations with climate change. J. Fish Biol. 2010, 76, 1786–1805. [Google Scholar] [CrossRef]
- Wilson, S.M.; Patterson, D.A.; Moore, J.M. Intra- and inter-population variation in sensitivity of migratory sockeye salmon smolts to phenological mismatch. Mar. Ecol. Prog. Ser. 2022, 692, 119–136. [Google Scholar] [CrossRef]
- Hovel, R.A.; Fresh, K.L.; Schroder, S.L.; Litt, A.H.; Quinn, T.P. A wide window of migration phenology captures inter-annual variability of favourable conditions: Results of a whole-lake experiment with juvenile Pacific salmon. Freshw. Biol. 2019, 64, 46–55. [Google Scholar] [CrossRef]
- Beechie, T.; Buhle, E.; Ruckelshaus, M.; Fullerton, A.; Holsinger, L. Hydrologic regime and the conservation of salmon life history diversity. Biol. Conserv. 2006, 130, 560–572. [Google Scholar] [CrossRef]
- McClure, M. M.; Carlson, S.M.; Beechie, T.J.; Pess, G.R.; Jorgensen, J.C. ; Sogard,, S. M.; Sultan, S.E.; Holzer, D.M.; Travis, J.; Sanderson, B.L.; Power, M.E.; Carmichael, R.W. Evolutionary consequences of habitat loss for Pacific anadromous salmonids: Salmonid habitat loss and evolution. Evol. Appl. 2008; 1, 300–318. [Google Scholar]
- Haraldstad, T.; Haugen, T.O.; Olsen, E.M.; Forseth, T.; Höglund, E. Hydropower-induced selection of behavioural traits in Atlantic salmon (Salmo salar). Sci. Rep. 2021, 11, 16444. [Google Scholar] [CrossRef]
- Gauld, N.R.; Campbell, R.N.; Lucas, M.C. Reduced flow impacts salmonid smolt emigration in a river with low-head weirs. Sci. Total Environ. 2013, 458–460, 435–443. [Google Scholar] [CrossRef]
- Chavarie, L. , Honkanen, M.; Newton, M.; Lilly, J.M.; Greetham, H.R.; Adams, C.A. The benefits of merging passive and active tracking approaches: New insights into riverine migration by salmonid smolts. Ecosphere 2022, 13, e4045. [Google Scholar] [CrossRef]
- Flávio, H.; Kennedy, R.; Ensing, D.; Jepsen, N.; Aarestrup, K. Marine mortality in the river? Atlantic salmon smolts under high predation pressure in the last kilometres of a river monitored for stock assessment. Fish. Manag. Ecol. 2020, 27, 92–101. [Google Scholar] [CrossRef]
- Welch, D.W.; Porter, A.D.; Rechisky, E.L. A synthesis of the coast-wide decline in survival of West Coast Chinook Salmon (Oncorhynchus tshawytscha, Salmonidae). Fish Fish. 2021, 22, 194–211. [Google Scholar] [CrossRef]
- Dukes, J.S.; Mooney, H.A. Does global change increase the success of biological invaders? Trends Ecol. Evol. 1999, 14, 135–139. [Google Scholar] [CrossRef]
- Rahel, F.J.; Olden, J.D. Assessing the Effects of Climate Change on Aquatic Invasive Species. Conserv. Biol. 2008, 22, 521–533. [Google Scholar] [CrossRef]
- Mainka, S.A.; Howard, G.W. Climate change and invasive species: double jeopardy. Integr. Zool. 2010, 5, 102–111. [Google Scholar] [CrossRef]
- Koed, A.; Baktoft, H.; Bak, B. D. Causes of mortality of Atlantic salmon (Salmo salar) and brown trout (Salmo trutta) smolts in a restored river and its estuary. River Res. Appl. 2006, 22, 69–78. [Google Scholar] [CrossRef]
- Jepsen, N. : Flávio, H.; Koed, A. The impact of Cormorant predation on Atlantic salmon and Sea trout smolt survival. Fish. Manag. Ecol. 2019, 26, 183–186. [Google Scholar] [CrossRef]
- Vehanen, T.; Huusko, A.; Bergman, E.; Enefalk, Å.; Louhi, P.; Sutela, T. American mink (Neovison vison) preying on hatchery and wild brown trout (Salmo trutta) juveniles in semi-natural streams. Freshw. Biol. 2022, 67, 433–444. [Google Scholar] [CrossRef]
- Hansen, L.P.; Quinn, T.P. The marine phase of the Atlantic salmon (Salmo salar) life cycle, with comparisons to Pacifc salmon. Can. J. Fish. Aquat. Sci. 1998, 55, 104–118. [Google Scholar] [CrossRef]
- Potter, E.C.E. ; Crozier. W.W. A perspective on the marine survival of Atlantic salmon. In The ocean life of Atlantic salmon: environmental and biological factors influencing survival; Mills, D., Ed.; Fishing News Books, Oxford, UK, 2000, pp 19–36.
- Friedland, K.D.; MacLean, J.C.; Hansen, L.P.; Peyronnet, A.J.; Karlsson, L.; Reddin, D.G.; Maoiléidigh, Ó.N.; McCarthy, J.L. The recruitment of Atlantic salmon in Europe. ICES J. Mar. Sci. 2009, 66, 289–304. [Google Scholar] [CrossRef]
- Friedland, K.D.; Shank, B.V.; Todd, C.D.; McGinnity, P.; Nye, J.A. Diferential response of continental stock complexes of Atlantic salmon (Salmo salar) to the Atlantic Multidecadal Oscillation. J. Mar. Syst. 2014, 133, 77–87. [Google Scholar] [CrossRef]
- Salminen, M.; Kuikka, S.; Erkamo, E. Annual variability in survival of sea-ranched Baltic salmon, Salmo salar L.: significance of smolt size and marine conditions. Fish. Manag. Ecol. 1995, 2, 171–184. [Google Scholar] [CrossRef]
- Friedland, K.D.; Reddin, D.G.; McMenemy, J.R.; Drinkwater, K.F. Multidecadal trends in North American Atlantic salmon (Salmo salar) stocks and climate trends relevant to juvenile survival. Can. J. Fish. Aquat. Sci. 2003, 60, 563–583. [Google Scholar] [CrossRef]
- Friedland, K.D.; Hansen, L.P.; Dunkley, D.A.; MacLean, J.C. Linkage between ocean climate, post-smolt growth, and survival of Atlantic salmon (Salmo salar L.) in the North Sea area. ICES J. Mar. Sci. 2000, 57, 419–429. [Google Scholar] [CrossRef]
- Beaugrand, G.; Reid, P.C. Long-term changes in phytoplankton, zooplankton and salmon related to climate. Glob. Chang. Biol. 2003, 9, 801–817. [Google Scholar] [CrossRef]
- Jonsson, B.; Jonsson, N. Migratory timing, marine survival and growth of anadromous brown trout in the River Imsa, Norway. J. Fish Biol. 2009, 74, 621–638. [Google Scholar] [CrossRef]
- Chaput, G. Overview of the status of Atlantic salmon (Salmo salar) in the North Atlantic and trends in marine mortality. ICES J. Mar. Sci. 2012, 69, 1538–1548. [Google Scholar] [CrossRef]
- Olmos, M.; Payne, M.R.; Nevoux, M.; Prévost, E; Chaput, G. ; Du Pontavice, H.; Guitton, J.; Sheehan, T.; Mills, K.; Rivot, E. Spatial synchrony in the response of a long range migratory species (Salmo salar) to climate change in the North Atlantic Ocean. Glob. Chang. Biol. 2020, 26, 1319–1337. [Google Scholar] [CrossRef]
- Friedland, K.D.; Dannewitz, J.; Romakkaniemi, A.; Palm, S.; Pulkkinen, H.; Pakarinen, T.; Oeberst, R. Post-smolt survival of Baltic salmon in context to changing environmental conditions and predators. ICES J. Mar. Sci. 2017, 74, 1344–1355. [Google Scholar] [CrossRef]
- Chaput, G.; Carr, J.; Daniels, J.; Tinker, S.; Jonsen, I.; Whoriskey, F. Atlantic salmon (Salmo salar) smolt and early post-smolt migration and survival inferred from multi-year and multi-stock acoustic telemetry studies in the Gulf of St. Lawrence, northwest Atlantic. ICES J. Mar. Sci. 2019, 76, 1107–1121. [Google Scholar] [CrossRef]
- Jonsson, N.; Jonsson, B. Growth and sexual maturation in Atlantic salmon Salmo salar L. J. Fish Biol. 2007, 71, 245–252. [Google Scholar] [CrossRef]
- Jonsson, B.; Jonsson, N.; Albretsen, J. Environmental change influences the life history of salmon Salmo salar in the North Atlantic Ocean. J. Fish Biol. 2016, 88, 618–637. [Google Scholar] [CrossRef] [PubMed]
- Hvidsten, N.A.; Heggberget, T.G.; Jensen, A.J. Sea water temperatures at Atlantic salmon smolt entrance. Nord. J. Freshw. Res. 1998, 74, 79–86. [Google Scholar]
- Simmons, O.M.; Gregory, S.D.; Gillingham, P.K.; Riley, W.D.; Scott, L.J.; Britton, J.R. Biological and environmental infuences on the migration phenology of Atlantic salmon Salmo salar smolts in a chalk stream in southern England. Freshw. Biol. 2021, 66, 1581–1594. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2007: The Physical Science Basis. In Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, 2007, pp. 996.
- Peyronnet, A.; Friedland, K.D.; Maoileidigh, N.O.; Manning, M.; Poole, W.R. Links between patterns of marine growth and survival of Atlantic salmon Salmo salar, L. J. Fish Biol. 2007, 71, 684–700. [Google Scholar] [CrossRef]
- Todd, C. D.; Hughes, S. L.; Marshall, T.; MacLean, J. C.; Lonergan, M. E.; Biow, E. M. Detrimental effects of recent ocean surface warming on growth condition of Atlantic salmon. Glob. Chang. Biol. 2008, 14, 1–13. [Google Scholar] [CrossRef]
- Edwards, M.; Beaugrand, G.; Hays, G.C.; Koslow, J.A.; Richardson, A.J. Multi-decadal oceanic ecological datasets and their application in marine policy and management. Trends Ecol. Evol. 2010, 25, 602–610. [Google Scholar] [CrossRef]
- Malick, M.J.; Cox, S.P.; Mueter, F.J.; Peterman, R.M.; Bradford, M. Linking phytoplankton phenology to salmon productivity along a north–south gradient in the Northeast Pacific Ocean. Can. J. Fish. Aquat. Sci. 2015, 72, 697–708. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Cushing, D.H. Plankton Production and Year-class Strength in Fish Populations: An Update of the Match/Mismatch Hypothesis. Adv. Mar. Biol. 1990, 26, 249–293. [Google Scholar]
- Palm, S.; Romakkaniemi, A.; Dannewitz, J.; Pakarinen, T.; Veneranta, L.; Huusko, R.; Isometsä, K.; Broman, A.; Miettinen, A. Tornionjoen lohi-, meritaimen- ja vaellussiikakannat – yhteinen ruotsalais-suomalainen biologinen selvitys sopivien kalastussääntöjen arvioimiseksi vuodelle 2023. Sveriges lantbruksuniversitet and Luonnonvarakeskus. Helsinki, Finland, 2023, pp. 62.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



