
Submitted:
29 May 2023
Posted:
30 May 2023
You are already at the latest version
Abstract
Type 2 Diabetes (T2D) is a growing global health problem that requires new and effective prevention and management strategies. Recent research has highlighted the role of epigenetic changes in the development and progression of T2D, and the potential of functional foods as complementary therapy for the disease. This review aims to provide an overview of the current state of knowledge on the preventive epigenetic mechanisms of functional foods in T2D. We provide background information on T2D and its current treatment approaches, an explanation of the concept of epigenetics, and an overview of the different functional foods with demonstrated preventive epigenetic effects in T2D. We also discuss the epigenetic mechanisms by which these functional foods prevent or manage T2D and the studies that have investigated their preventive epigenetic effects. In addition, we revisit works on the beneficial influence of functional foods against the programming and complications of parentally-triggered offspring diabetes. We also postulate, albeit based on the scarce data, that epigenetic inheritance mechanistically mediates the impacts of functional nutrition against the metabolic risk of diabetes in the offspring. Finally, our review highlights the importance of considering the preventive epigenetic mechanisms of functional foods as a potential avenue for the development of new prevention and management strategies for T2D.
Keywords:
diabetes
; epigenetic inheritance
; functional foods.
; transgenerational
1. Introduction
1.1. Background on Type 2 Diabetes and Its Treatment Approaches
Diabetes is the seventh leading cause of global mortality and has surpassed pandemic proportions. From less than 1 million deaths in the year 2000, the disease is responsible for almost 7 million deaths in 2021 [1]. Type 2 Diabetes (T2D) is specifically characterized by high levels of glucose in the blood, resulting from a combination of factors including insulin resistance, a decrease in insulin secretion, and an increase in glucose production by the liver [2]. T2D causes significant morbidity and mortality worldwide, and is associated with a number of serious health complications such as cardiovascular disease, kidney disease, and blindness [3,4].
Owing to the fact that high consumption of unhealthy diets and sedentary lifestyles have been much implicated, the current treatment approaches for T2D include lifestyle modifications (e.g., healthy diet and physical activity), oral medications, and insulin therapy [5,6]. Lifestyle modifications are considered the cornerstone of T2D management, and are often the first line of treatment for individuals with prediabetes or newly diagnosed T2D. In addition to lifestyle changes, oral medications such as metformin, sulfonylureas, and dipeptidyl peptidase-4 (DPP-4) inhibitors are commonly used to improve insulin sensitivity, increase insulin secretion, and decrease glucose production [7,8]. For individuals who require more intensive therapy, insulin therapy may be recommended.
While these traditional approaches have been effective in managing T2D, they have limitations and may not be effective for all individuals. As such, there is a growing interest in developing alternative strategies for preventing and managing T2D, including the use of functional foods with demonstrated preventive epigenetic effects [9].
Even though, the disease has no known cure, a combination of lifestyle modification and drug therapy in diagnosed persons has shown great success in reducing mortality and increasing quality of life. More importantly, as advocated by the World Health Organization, is the preventive narrative in reducing the global burden of the disease. Diet has been a cornerstone in the prevention of diabetes since the era of classical Greece. This is as enumerated in the tenet “Let food be thy medicine and medicine be thy food” as advocated by Hippocrates.
1.2. Dietary Lifestyle as a Risk Factor for Type 2 Diabetes
The risk to developing type 2 diabetes has been largely secondary to either environmental, lifestyle or genetic factors [10]. Concrete data has linked lifestyle factors like diet [11], heavy smoking [12], opioid use [13], physical inactivity (sedentarism), obesity [14,15] and alcoholism [16,17] to type 2 diabetes. Individual diets (food) and some dietary combinations have potential to differentially impact the risk of developing type 2 diabetes [18,19]. Numerous studies have shown that the type of food people consume could be associated with the development of diabetes. In a cross-sectional study, Shu et al. demonstrated an association between consuming western diet and the risk of type 2 diabetes mellitus among middle aged Chinese adults [20]. Also, several cohort and preclinical studies have highlighted diet as a contributory factor in predisposition to type 2 diabetes mellitus [11,21,22,23]. High fibre diet, low glycaemic index diet, and high amylose diet have been associated with less predisposition to type 2 diabetes when compared to their counterparts [24,25,26]. Nuts have also been shown to be beneficial in preclusion to type 2 diabetes. On the other hand, refined grains and sugar-sweetened beverages tend to promote predisposition to type 2 diabetes [27]. For instance, clinical trials of short- and medium-term durations on postprandial glucose excursion in normal patients, insulin sensitivity in obese patients, and glucose handling in diabetic patients confirmed the superiority of whole grain diet over refined grains in maintaining a near euglycemic state [28]. Studies have also shown that people who consume fermented dairy products are less likely to develop type 2 diabetes than those who consume non-fermented ones [29,30].
1.3. The Concept of Epigenetics and Its Role in Chronic Diseases
Epigenetics is the study of changes in gene function that occur without a change in the underlying DNA sequence [31]. These changes are caused by modifications to DNA and its associated proteins, such as histones, which play a crucial role in regulating gene expression. There are several different types of epigenetic modifications, including DNA methylation, histone modification, and non-coding RNA-mediated regulation, all of which can have a significant impact on gene expression [32]. Epigenetic changes can occur in response to a variety of environmental and lifestyle factors, such as diet, physical activity, and exposure to toxins. These changes can be transmitted from one generation to the next, and may play a key role in the development and progression of chronic diseases, including T2D [33].
Studies have demonstrated that epigenetic changes can lead to alterations in the expression of genes involved in glucose metabolism, insulin secretion, and inflammation, which are all important factors in the development and progression of T2D [34]. Additionally, epigenetic changes can increase the risk of T2D by altering the expression of genes involved in fat metabolism, which can lead to obesity and insulin resistance [35]. Epigenome-wide association studies have substantially linked DNA methylation to diabetes [36]. Using human pancreatic islets, Daneshpajooh et al. also demonstrated that diabetes and impaired insulin secretion could result from epigenetic modifications that alter gene expression [37]. In addition, epidemiological findings in offspring born to Dutch Hunger Winter mothers during World War II has provided some evidence of the predisposition to diabetes due to intra-uterine perturbations [38]. On the other hand, several evidence also highlight the mechanistic role of epigenetics in the dietary mitigation of type 2 diabetes. For instance, Li et al. show that diet high in folates reduce blood glucose levels and improve insulin sensitivity by altering DNA methylation patterns in adipose tissues of genes associated with type 2 diabetes [39]. The role of epigenetics in chronic diseases like T2D highlights the importance of environmental and lifestyle factors like diet in disease prevention and management. Therefore, this review sought to summarize the current knowledge on the preventive epigenetic mechanisms of functional foods in T2D.
2. The Role of Epigenetics in T2D
2.1. Overview of Epigenetic Changes Associated with T2D
T2D is associated with epigenetic changes that can impact the expression of genes involved in glucose metabolism, insulin signaling, and inflammation. These epigenetic changes include the following:
i. DNA methylation: DNA methylation is a chemical modification that involves the addition of a methyl group to the DNA molecule. Methylated DNA in mammals are usually in form of 5-methylcytosine. In lower organisms (Drosophila melanogaster, fungi and bacteria), majority of the methylated DNA are in the form of N6-methyladenine [40]. While N6-methyladenine has been associated with over-expression, 5-methylcytosine has been linked with gene repression [41,42]. In T2D, DNA methylation changes have been observed in genes involved in insulin secretion and glucose metabolism, leading to alterations in their expression and function [43]. Maternal high fat diet consumption in mice during gestation and lactation has been demonstrated to cause insulin resistance and glucose intolerance in their F1 offspring through promotion of Irs2 DNA methylation and decrease in Map2k DNA methylation [44]. DNA methylation has been hypothesized to act by disrupting the binding of transcription factors to recognition elements (containing a CG nucleotide) leading to silencing of gene expression [45]. These aberrations in DNA methylation (whether hypo- or hyper-methylations) are oftentimes secondary to DNA methyltransferases gene (DNMT1, DNMT3A, and DNMT3B) dysregulation/mutations.
ii. Histone modification: Histones are proteins that interact with DNA to help package it into a compact structure. Histone modifications include phosphorylation, methylation, acetylation and ubiquitination of histone molecules. This leads to conformational changes that remodels chromatin thereby altering the activity and interactions of transcription factors, nuclear protein and histone molecules, this in turn affects gene transcription and its sequelae [46]. In T2D, changes in histone modification have been observed in genes involved in insulin sensitivity, glucose metabolism, and inflammation [47].
iii. Non-coding RNA-mediated regulation: Non-coding RNAs, such as microRNAs, are short RNA molecules that do not encode for proteins but play a crucial role in regulating gene expression. For instance, the parental intake of five functional food oils have been shown to modulate miRNA expression in both parent and offspring [48]. Also, functional foods have also been shown to modulate the expression of lncRNA in parents and offspring [39,49,50,51]. Essentially in T2D, changes in the expression of non-coding RNAs have been observed in genes involved in insulin secretion and glucose metabolism, leading to alterations in their function [52,53].
These epigenetic changes associated with T2D can also interact with other factors, such as obesity and physical inactivity, which are known to increase the risk of T2D [35]. The complex interplay between these factors highlights the importance of a holistic approach to the prevention and management of T2D that takes into account both genetic and environmental factors.
Furthermore, epigenetic changes in genes involved in insulin secretion and sensitivity, glucose metabolism, inflammation, and adipocyte differentiation and function can have a profound impact on the development of type 2 diabetes (T2D). Insulin secretion and sensitivity can be decreased due to these epigenetic changes, leading to insulin resistance, which is a defining characteristic of T2D. Additionally, alterations in glucose metabolism can result in an increase in glucose production by the liver and elevated blood glucose levels, due to changes in glucose uptake, utilization, and storage in peripheral tissues. Inflammation is also impacted by these epigenetic changes, resulting in a persistent state of low-grade inflammation that is associated with the progression of T2D. Moreover, changes in the differentiation and function of adipocytes can lead to altered lipid storage and increased risk of obesity and insulin resistance. These various changes accumulate over time, resulting in a progressive decline in glucose metabolism and insulin sensitivity, which can increase the likelihood of developing T2D and its associated health complications.
2.2. Epigenetic Inheritance of T2D
Based on the sustainability of a conferred metabolic disease along subsequent generations of offspring, an epigenetic transfer may be termed transgenerational, intergenerational or multigenerational [54]. Such epigenetic change(s) could be important therapeutic target(s) for reducing diabetes because of the typical epigenetic signature of being modifiable and reversible. Numerous clinical observations and in vivo laboratory experiments involving animals have proved beyond conjecture that there is a generational link of type 2 diabetes between parents and their subsequent descendants. Compelling animal studies involving embryo transfer experiments [55] and discordant human siblings born to parent before and after the development of maternal diabetes [56] have given credence to the association between gestational diabetes and development of a later life type 2 diabetes. Later generational phenotypic expressions (e.g. type 2 diabetes) consequent on “developmental programming” due to early life environmental insults especially over- and under-nutrition of specific nutrients have demonstrated the risk of a generational diabetic link between offspring and their parents [57,58,59]. Crudo et al demonstrated a transgenerational association between late gestation glucocorticoid intervention in F1 pregnant guinea pigs and global DNA methylation and expression of crucial metabolic genes in subsequent generations of offspring [60]. In an earlier study, maternal under-nutrition had been associated with transgenerational increased neonatal adiposity in offspring of exposed women [61]. Furthermore, using mouse model, Pavlinkova and colleagues have demonstrated the association between paternal diabetes and sperm quality, and expression patterns in offspring of later generations [58]. With the foregoing observations and many more reports in the literature, it is almost impossible rule out the transmission of type 2 diabetes from parents to their subsequent generational offspring.
Accordingly, the underlying mechanisms responsible for transmission of type 2 diabetes have been explored since the early 90s. Diets have been shown to programme type 2 diabetes by altering structural and functional metabolic systems in adipose tissues, muscles, pancreas and liver [62,63,64]. Epigenetics have evolved as the underlying molecular mechanisms through which these dietary disturbances during intra-uterine life are registered and “reminisced” during later generations as metabolic dysfunctions like type 2 diabetes. These metabolic dysfunctions occur through upstream modifications in epigenetic signatures which influence gene expression without tampering with the DNA sequence. For example, studies have shown that diet influences the expression of intestinal fatty acid binding protein (IFABP) mRNA which will in turn affect all downstream processes affected by its protein [65]. IFABP is known to be an expressed product of the FABP2 gene which is responsible for a myriad of fatty acid trafficking across several metabolic pathways. This report indicates that diet can modify epigenetic memory of specific genes which in turn affect expression of proteins responsible for handling of specific metabolic pathways involving homeostasis of blood glucose and body weight. This in the long run impact the development of heritable type 2 diabetes mellitus.
Therefore, understanding the specifics of these changes is crucial for the development of new strategies for preventing and managing the disease, including the use of functional foods with preventive epigenetic effects.
3. Functional Foods and Their Epigenetic Effects
Since the 1970s, there has been an increased interest in diets that provide additional health benefits beyond the traditional macro- and micronutrients [66]. Diets with such properties have been termed “functional foods”. Functional foods can be regarded as processed foods that are taken whole or enhanced (enriched or fortified) to provide some tertiary benefit on the body’s physiology over and above the nutrition (primary) and sensory (secondary) benefits provided by conventional foods when imbibed at efficacious levels as part of a varied diet on a regular basis [29,66]. In his perspective article, Temple opines that “Functional foods are novel foods that have been formulated so that they contain substances or live microorganisms that have a possible health-enhancing or disease-preventing value, and at a concentration that is both safe and sufficiently high to achieve the intended benefit” [67]. These foods are designed to deliver specific health benefits and are often enriched with bioactive components such as vitamins, minerals, and phytochemicals.
Health benefits of functional foods can vary widely depending on the type of food and the specific bioactive components it contains. Functional foods have been extensively studied in recent years for their potential in promoting overall health and preventing various diseases. These foods have been shown to play a significant role in disease prevention by exerting a variety of positive effects on the body. One such effect is improving the nutrient status of the body. By providing essential vitamins and minerals that may be lacking in the average diet, functional foods can help improve the overall nutrient status of the individual [68]. Another important effect of functional foods is their ability to modulate inflammation. Some functional foods contain bioactive components that can help regulate the immune system and reduce chronic inflammation, which is associated with the development of many chronic diseases, including T2D [69].
Another key benefit of functional foods is their ability to regulate glucose metabolism. Some functional foods contain bioactive components that can help improve insulin sensitivity and regulate glucose metabolism, which can be especially beneficial for individuals who are at risk of developing T2D [9]. Additionally, some functional foods contain components that can modify epigenetic marks, such as DNA methylation and histone modification. For instance, a recent study by Kusuyama et al. demonstrates the potential of parental exercise & diet to determine the development of offspring diabetes by altering offspring histone methylation (an epigenetic change) [25]. Such epigenetic modifications may be responsible for the multigenerational transmission of impaired islet function [26,70].
3.1. Health Benefits of Functional Foods against Parentally Triggered Offspring Type 2 Diabetes
A number of functional foods have been proven beneficial to offspring defence against inherited predisposition to diabetes (see Table 1). These functional foods are categorised below into whole foods or enhanced foods:
i. Whole food
Whole foods defined here include diets that are largely unmodified and re-discovered to confer tertiary health benefits (i.e., against parentally triggered offspring diabetes) beyond the primary nutrition and secondary physicochemical benefits. These include milk fat, germinated brown rice and camel milk. Milk fat is an important component of mammalian breast milk that serves as good source of Milk Fat Globule Membrane (MFGM) – a fraction rich in bioactive lipids and proteins [71]. Ye et al. demonstrated the ability of MFGM to mitigate glucose intolerance, insulin resistance in both male and female offspring of mice challenged with HFD during pregnancy and lactation [72]. Of note, this instance of neonatal protection was associated with sexually specific remodulation of the HFD triggered gut dysbiosis. Similarly, studies by Mahmoud et al., [73] demonstrated that whole camel milk rich in camel whey protein prevents the reconstruct of diabetes and its related complications in offspring of diabetic mothers. In addition, whole grain - germinated brown rice, rich in gamma oryzanol, has been shown to raise adiponectin levels. This elevation in adiponectin promotes insulin sensitivity and suppresses glucose output [74]. The anti-diabetic effect of the whole brown rice was conferred to susceptible offspring via modifications of DNA methyl marks and post-translational acetylation of histone H3 & H4 [75]. These represent good examples of the epigenetic impact of functional foods across generation.
ii. Enhanced foods
Enhanced functional foods may be enriched or fortified with functional ingredients or may be formulations that exclude substances such as allergens, gluten or lactose [29]. The high glucose levels predominant in diabetes has been reported to cause a surge in the levels of zinc in urine by affecting urinary zinc absorption [76]. Investigations demonstrate that such significant excretion of endogenous zinc by the offspring is prevented following administration of poultry eggs enriched with Vitamin E and omega-3 fatty acids [77]. Another study demonstrates that the intake of poultry egg enriched with macronutrients (vitamin C and E) and micronutrients (Cu, Zn and Mn) by offspring of zinc triggered diabetic parents reverses the risk of inheriting type 2 diabetes and its complications such as oxidative damage [78]. This is particularly important because hyperglycaemia (of diabetes) propels the formation of free radicals of the oxygen species through glucose auto-oxidation and non-enzymatic glycations. Otherwise, an excess of these free radicals overcomes antioxidant defences and results in damages such as lipid peroxidation [79]. Thus, food enrichment mitigates the lipid peroxidation status of offspring born to diabetic parents. Sometimes, the nutrients implicated (for enrichment) are essential components of enzymes, some of which play significant roles in the antioxidant defence system. Consequently, such functional foods may serve as dietary supplements to normalize the mineral status in offspring of diabetic parents and may thus prevent the oxidative stress amongst populations consuming foods having high Zn levels.Furthermore, the length of chromosomal telomeres has been reported to shorten in diabetes and is negatively correlated with glucose concentrations and glycated hemoglobin (HbA1c) levels [80,81]. A study revealed that foods fortified with omega 3 polyunsaturated fatty acids (n-3 PUFA) depress the hepatic telomere shortening observed in offspring of gestationally diabetic (GDM) rats [82]. The n-3 PUFA administered to the offspring of GDM rats significantly raised insulin levels and reversed an impaired glucose tolerance that was observed to worsen with offspring age. In another instance, low birth weight common to most famine is associated with the development of type 2 diabetes [83]. Studies shows that modified poultry egg enriched with vitamin E and omega-3 fatty acids improve birth weight and enhance offspring survival [77]. Though these enhanced functional foods wane the risk of developing type 2 diabetes and its complications, demonstrations of the mechanistic bases behind their inherited benefits remains scarce.
3.2. Functional Foods with Preventive Epigenetic Effects in T2D
Several functional foods have been shown to have preventive epigenetic effects in T2D, including berries, cruciferous vegetables, green tea, omega-3 fatty acids, and spices.
i. Berries: Berries such as blueberries, blackberries, and strawberries contain bioactive compounds like anthocyanins and ellagitannins, which have been linked to the prevention of T2D by improving insulin sensitivity, regulating glucose metabolism, and reducing inflammation. In a study by Ma et al. [84] blueberry supplementation was found to alter DNA methylation patterns of genes involved in insulin signaling and inflammation in individuals with T2D, suggesting the potential role of blueberries in preventing T2D through epigenetic mechanisms.
ii. Cruciferous Vegetables: Cruciferous vegetables, such as broccoli, cauliflower, and cabbage, contain bioactive compounds such as sulforaphane and indole-3-carbinol, which have been found to reduce inflammation and improve insulin sensitivity, hence preventing T2D [85]. Cruciferous vegetable has also been shown to alter DNA methylation patterns of genes implicated in insulin signalling and inflammation in patients with T2D, implicating a possible role in T2D prevention via epigenetic processes [86].
iii. Green Tea: Green tea is rich in catechins, including epicatechin and epicatechin gallate, which have been shown to have preventive epigenetic effects in T2D, and referred to as epi-drugs [87]. These compounds have been shown to regulate glucose metabolism and improve insulin sensitivity through their interaction with specific epigenetic marks, such as histone modifications and DNA methylations [88,89]. Moreover, it has been shown that Epigallocatechin gallate inhibits the neural tube abnormalities caused by maternal diabetes and limits the increased expression and activity of DNA methyltransferases, resulting in the suppression of DNA hypermethylation and the restoration of neural tube closure required gene expression [90]. These findings suggest a preventive and management role for green tea in T2D through epigenetic mechanisms.
iv. Omega-3 Fatty Acids: Omega-3 fatty acids found in fatty fish like salmon and flaxseeds have been linked to the prevention of T2D through reducing inflammation and improving insulin sensitivity [91]. Moreover, studies have shown the possible link between omega-3 fatty acids supplementation and the methylation of inflammatory genes in T2D [92]. This implies the potential role of omega-3 fatty acids in preventing T2D through epigenetic mechanisms.
v. Spices: Spices such as turmeric and cinnamon contain bioactive compounds like curcumin and cinnamaldehyde, which have been linked to the prevention of T2D through improving insulin sensitivity and regulating glucose metabolism and inflammation. For instance, curcumin has been shown to regulate histone deacetylases, histone acetyltransferases, DNA methyltransferase I, and miRNAs [93]. Moreover, the study by Yun et al. [94] showed the potential of curcumin in T2D through histone modification. They showed that high glucose levels trigger the NF-B signalling pathway, resulting in the activation of pro-inflammatory genes. However, curcumin treatment of high glucose-induced cells stimulates HDAC activity, particularly HDAC2, while reducing HAT activity, particularly p300, resulting in deacetylation of p65 NF-B and, as a result, suppression of proinflammatory cytokine production. Therefore, the role of curcumin supplementation in the regulation of DNA methylation, histone acetylation, and miRNAs of genes involved in insulin signaling and inflammation in individuals with T2D suggests a potential role for curcumin in preventing T2D through epigenetic mechanisms [95].
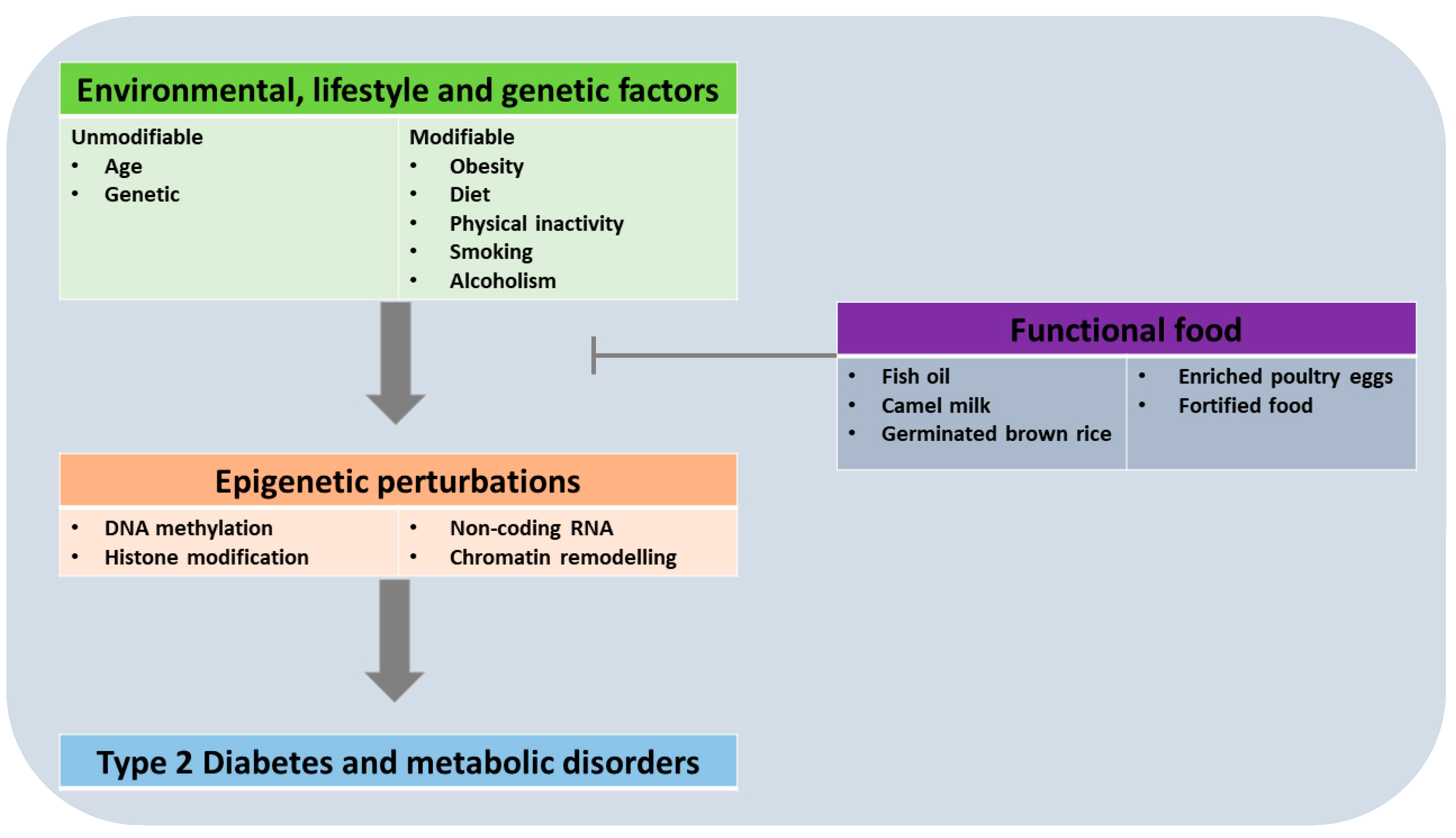
Since perinatal perturbations (malnutrition and other metabolic insults) in parents (F0) have been demonstrated to predispose offspring (F1) to the development of later-life risk factors to diabetes, then an appropriate functional food could prevent intergenerational epigenetic change(s) that could worsen the burden of diabetes (see Figure 1). This hypothesis is strengthened by the fact that supplementation of the nutritional factor in question during intra-uterine life prevents development of diabetic risk factors in subsequent generations [96,97,98,99]. This scenario is underscored in low- and middle-income countries where malnutrition due to poverty and illiteracy, and other perinatal perturbations are endemic. A plethora of data in literature have highlighted a number of diets with “functional” capability [66,100]. Germinated brown rice, fortified margarines, soya bean/soy, bitter melon, sea grape, and psyllium are among diets termed as functional foods based on animal studies and clinical trials [21,29,101,102,103,104]. The scientific evidence supporting these effects is growing, but more research is needed to fully understand the mechanisms underlying these effects and to determine the optimal dose and duration of consumption.

Table 1.
Functional foods demonstrating defence against offspring diabetes.
| Functional Food | Nature of functional food | Bioactive phytochemical/ zoochemical | Study Model | Efficacious levels | Methods | Nature of Antidiabetic Functionality | Mechanistic Basis | Conclusion | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Omega-3 enriched diet | Enriched | Omega-3 (ɷ3) | Offspring of Wistar rat | Efficacy of ɷ3-rich diet extends up to PND90 | Dam; gestation and lactation [control (C: 19% of lipids and ɷ6: ɷ3 = 12), HF (HF: 33% lipids and ɷ6: ɷ3 = 21), or HF enriched with ɷ3 (HFω3: 33% lipids and ɷ6: ɷ3 = 9) diet] | ɷ3 improved glycemic profile (insulin sensitivity, fasting glucose levels) | - | ɷ3-rich diet attenuates or prevents the short-term metabolic disruption elicited by HFD in offspring | [105] |
| Fat diet | Factor out formulation | Perinatal fat restriction | Offspring of mice | Low-fat dietary intervention across gestation and lactation | Dam; gestation or lactation (low and/or high fat diet) Offspring; Glucose tolerance and insulin sensitivity test at 12- and 70-week. |
Perinatal fat restriction provided adequate efficacy to restore insulin sensitivity in aging female progeny. | - | Fat restriction ameliorate glucose dysmetabolism and prevents diabetes | [106] |
| Milk fat | Whole | Milk Fat Globule Membrane (MFGM) | Offspring of C57BL/6 mice | 1,000 mg/kg BW/day MFGM | Dam; 3 weeks pre-gestation + gestation + lactation (high-fat diet (HFD) or a control diet) Pup; lactation (with or without 1,000 mg/kg BW/day MFGM supplementation) |
MFGM ameliorated metabolic disorder and improved glucose tolerance in offspring exposed to maternal HFD in a sex-specific manner. | Sex specific microbiota enrichment in offspring | MFGM is protective against transmitted glucose dysmetabolism | [72] |
| Fish oil food |
Fortified food | n-3 PUFA | Offspring of C57BL6J mice | 30 g FO/kg diet (equivalent to ~ 85–90 mg of FO per day for mice and 10 g per day for humans) | Dam; pre-gestation to lactation [HF diet (45% fat), HF + fish oil (FO - 30 g/kg of diet) and low fat (LF; 10% fat)] F1: after weaning (HF or FO). LF weaned onto LF as control |
FO lowers insulin resistance, reduces glucose intolerance and improves insulin sensitivity. | - | Fish oil improves glucose clearance and insulin sensitivity | [107] |
| Fish oil food |
Fortified food | n-3 PUFA | Offspring of C57BL/6 mice | 12.58 g/kg diet [Fish oil containing 70% (FA) DHA and 10% (FA) EPA in the form of triglyceride (DHA + EPA, 0.68% w/w)] | F0; pregnancy and lactation [control diet (AIN93G), n-3 PUFA-deficient diet and fish oil-contained n-3 PUFA rich diet (DHA + EPA, 0.68%, w/w)] F1; PD21 (high-fat diet or low-fat diet) |
n-3 PUFA deficiency elicits glucose intolerance and insulin resistance in offspring. n-3 PUFA helps ameliorates glucose dysmetabolism. | - | n-3 PUFA could potentially alleviate glucose metabolism disorders by increasing insulin sensitivity, inhibiting gluconeogenesis and promoting glycogenesis | [108] |
| Fish oil food | Fortified food | n-3 PUFA | Offspring of STZ induced GDM Wistar rats | 3rd month to 11th month of age | Dam; 6th day of gestation (STZ 30mk/kg) Offspring of GDM rats: First phase -3 months; standard diet (AIN-93, soybean oil). 2nd phase, 3-11 month; Group1- GDM offspring + AIN-93, soybean oil), Group2- GDM offspring + fish oil (60% n-3 PUFA), and Group3; GDM offspring + safflower oil (n-3 PUFA deficient) all versus control |
n-3 PUFA group had decreased oxidative stress, delayed hepatic telomere lengthening, inflammation with reduced levels of diabetes related metabolites compared to GDM group. | - | n-3 PUFA decreased the long-term risk of developing diabetes | [82] |
| Germinated brown rice | Whole germinated brown rice and oryzanol-rich extract | Oryzanol | Offspring of high-fat diet induced female Sprague Dawley rats | 50% GBR and 100 and 200 mg/kg of oryzanol-rich extract | Pregnant female Sprague Dawley rats were fed with high fat diet (HFD) alone, HFD + 50% germinated brown rice or HFD + oryzanol rich extract (100 or 200 mg/kg/day) throughout pregnancy and lactation. Their offspring were weaned at 4 weeks post-delivery and were followed up until 8 weeks. | GBR and oryzanol produced metabolic outcomes (adiponectin, 8-Iso prostaglandin) that favoured insulin sensitivity better than HFD feeding in the dams and offspring. | Altered global DNA methylation and modulated H3 and H4 acetylation | GBR and oryzanol can ameliorate HFD-induced epigenetically-mediated insulin resistance | [75] |
| Margarine | Fortified | Vitamin A (retinol and β-carotene) | Human offspring (49years) | Increased by 25 % from; 4.2 µg/g of retinol and 3.6 µg/g of β-carotene (equivalent to 0.6 % of the current RDA) to 6 µg/g of retinol and 3 µg/g of β-carotene (equivalent to 0.8 % of the current RDA). β-carotene may be a precursor of retinol. | Longitudinal follow-up on offspring of mothers who had been exposed to the extra vitamin A from margarine fortification (during pregnancy). As aftermath of a mandatory vitamin A fortification (of 25 %) issued by the Denmark government in 1962. | Significant more cases of T2DM in offspring of less-exposed compared to exposed. | - | Foetal exposure to extra vitamin A from fortified margarine may have lowered the risk of developing T2DM in adulthood | [109] |
| Camel milk | Whole | Undenatured camel whey protein (CWP) | Diabetic offspring of STZ diabetic mouse dams (BALB/c) | 100mg/kg for month of parturition | Dam; preconceptionally diabetic (2wks), gestational/ foetal exposure to CWP | Offspring at 3rd month postpartum; CWP restored the expression of ATF-3 and the levels of ROS, pro-inflammatory cytokines. Also normalized glucose and insulin levels compared to diabetic control. Improves survival | - | Mitigates tendency of the offspring to develop diabetes and related complications. | [73] |
| Enriched normo-caloric diets (flaxseed) | Enriched | Alpha-linolenic acid | C57Bl6/J mice | 94g % fat-free chow diet + 6% ALA enrichment (47.9 mole % 18:2n-6) | Dam; 2 weeks pre-conception and during gestation and lactation [diet of 94g % fat-free chow + 6% fat enriched in essential fatty acids (EFA): alpha-linolenic (ALA-18:3, n-3), linoleic (LA-18:2, n-6), or saturated fatty acids (SFA)]. HFD diet = 61.18 g% fat-free chow diet with 38 g% lard. Offspring; a 2-month post-weaning washing-out with normo-caloric diet (regular chow). |
ALA lowers glucose, insulin, HOMA index and stearoyl-CoA desaturase (SCD1) activity. | - | ALA-enriched maternal normo-caloric diets potentially attenuates insulin resistance in adult offspring | [110] |
| Modified Poultry egg | Enriched | Optimized nutrients compared to conventional eggs (Lower; protein, CHO, total lipids, cholesterol, TG, Zn. Higher; Cu, Mg, Vitamin E, C and omega-3 fatty acid (linolenic acid) | Diabetic offspring of diabetic rats induced by synthetic diet rich in high zinc (80mg/kg), high fat and refined sugar | For 30 days before mating | Three parental groups (both males and females); 1) control diet-105D, 2) diabetic diet-105D and 3) diabetic-egg mixed diet -75D+30D (105days before mating). Subset of groups 1&3; offspring (120D of modified egg diet from 9th day after weaning) | Parental intake of modified egg protected from malformations and improved offspring survival Offspring (F1) of diabetic parents fed modified egg had reduced blood glucose, blood pressure and lipid peroxidation. Normal lipid profile and improved enzyme activity. |
- | Though offspring of diabetic had high tendency to become diabetic. Modified egg reverses this risk and ameliorated complications like oxidative stress by normalizing mineral status. | [78] |
| Modified poultry egg | Enriched | Vitamin E and omega-3 fatty acids | Diabetic offspring of NIDDM Wistar rats induced by synthetic diet rich in high zinc (40 or 80mg/kg) | Parental groups (75days before mating); 1) control 20mg Zinc 2) 40mg/kg Zinc 3) 80mg/kg Zinc. Administered EM (4 liquid eggs /kg) to parents (1month before mating -weaning date PND21). Terminal assessment 18days post-weaning. | Enrichment enhanced survival and body weight, restored mineral basal mineral levels, reduced presence of trace elements in urine, prevented offspring abnormalities, | - | Vitamin E and omega-3 fatty acids protect offspring from damaging effects of diabetes | [77] |
ATF-3, activating transcription factor-3; CWP, Camel whey protein; EM, egg mixed diet; GDM, Gestational diabetes mellitus; NIDDM, non-insulin dependent diabetes mellitus; PND, postnatal day; PUFA, polyunsaturated fatty acids; RDA, recommended dietary allowance; STZ, streptozotocin; T2DM, type 2 diabetes, TG, triglyceride; ZN, zinc.
4. Implication for the Development of New Prevention and Management Strategies for T2D
The understanding of the preventive epigenetic mechanisms of functional foods in T2D has important implications for the development of new prevention and management strategies for this disease. Some of the key implications are:
i. Personalized Nutrition: By using functional foods that target specific epigenetic changes associated with T2D, it may be possible to develop personalized nutrition plans for individuals with this disease, or those at risk for developing it.
ii. Complementary Therapy: Functional foods have the potential to complement conventional treatments for T2D, such as lifestyle changes and pharmaceuticals, by modifying the epigenetic landscape and improving treatment outcomes.
iii. Disease Prevention: By targeting the epigenetic changes that contribute to the development and progression of T2D, functional foods have the potential to prevent the disease from developing in the first place.
iv. Reduced Dependence on Pharmaceuticals: By improving the efficacy of existing treatments and reducing the need for higher doses of medication, functional foods may also reduce the financial burden of T2D on both patients and the healthcare system.
In essence, the preventive epigenetic mechanisms of functional foods in T2D are a promising area of research with important implications for the development of new, complementary therapies for this disease. The existing evidence suggests that functional foods have the potential to improve treatment outcomes and prevent the disease from developing in the first place, but more research is needed to fully understand these mechanisms and to confirm these effects in human-based trials.
5. Conclusions and Future Outlook
The problem of T2D has been a leading health concern for a long time. To address the epidemic, we propose incorporating functional foods into our diets to not only prevent the onset of diabetes in offspring, but also improve the health of the parents, at minimal cost and for less medical consultation. We also noted that functional foods, taken at key developmental periods, may act via epigenetic means to positively impact offspring metabolic health, although the specific mechanisms and the impact of maternal versus paternal intake remain unclear and require further research. The existing evidence provides promising results for the preventive epigenetic effects of functional foods in T2D. However, more rigorous and long-term clinical studies are needed to fully understand the potential of functional foods in the prevention and management of this disease. Further research is needed to confirm the preventive epigenetic effects of functional foods in T2D through further clinical trials to determine optimal dosing and duration, understand specific epigenetic mechanisms through mechanistic studies, develop personalized nutrition plans based on individual's epigenetic profiles and dietary habits through research, determine efficacy of functional foods in reducing T2D incidence through interventional studies, and investigate the epigenetic effects of functional foods on other diabetes-related complications such as cardiovascular disease and neuropathy. The overall aim of future research in this field should be to build upon the existing evidence and provide a comprehensive understanding of the preventive epigenetic effects of functional foods in T2D for the development of new, evidence-based prevention and management strategies.
Acknowledgments
DU and KOS acknowledge the postgraduate sponsorship given by CAMRET (CAMRET/ 2019/MSc/003 & CAMRET/2019/PhD/001 respectively).
Conflict of Interest
None declared.
References
- World Health Organization WHO - The Top 10 Causes of Death. 24 Maggio 2018, 1–7.
- Hameed, I.; Masoodi, S.R.; Mir, S.A.; Nabi, M.; Ghazanfar, K.; Ganai, B.A. Type 2 Diabetes Mellitus: From a Metabolic Disorder to an Inflammatory Condition. World J. Diabetes 2015, 6, 598. [Google Scholar] [CrossRef]
- Deshpande, A.D.; Harris-Hayes, M.; Schootman, M. Epidemiology of Diabetes and Diabetes-Related Complications. Phys. Ther. 2008, 88, 1254–1264. [Google Scholar] [CrossRef] [PubMed]
- Petrie, J.R.; Guzik, T.J.; Touyz, R.M. Diabetes, Hypertension, and Cardiovascular Disease: Clinical Insights and Vascular Mechanisms. Can. J. Cardiol. 2018, 34, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Magkos, F.; Yannakoulia, M.; Chan, J.L.; Mantzoros, C.S. Management of the Metabolic Syndrome and Type 2 Diabetes through Lifestyle Modification. Annu. Rev. Nutr. 2009, 29, 223–256. [Google Scholar] [CrossRef] [PubMed]
- Asif, M. The Prevention and Control the Type-2 Diabetes by Changing Lifestyle and Dietary Pattern. J. Educ. Health Promot. 2014, 3, 1. [Google Scholar] [CrossRef]
- Chaudhury, A.; Duvoor, C.; Reddy Dendi, V.S.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, N.S.; Montales, M.T.; Kuriakose, K.; et al. Clinical Review of Antidiabetic Drugs: Implications for Type 2 Diabetes Mellitus Management. Front. Endocrinol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Sola, D.; Rossi, L.; Schianca, G.P.C.; Maffioli, P.; Bigliocca, M.; Mella, R.; Corlianò, F.; Paolo Fra, G.; Bartoli, E.; Derosa, G. Sulfonylureas and Their Use in Clinical Practice. Arch. Med. Sci. 2015, 11, 840–848. [Google Scholar] [CrossRef]
- Tuomilehto, J. Functional Foods and Lifestyle Approaches for Diabetes Prevention and Management. Nutrients 2017, 9, 1310. [Google Scholar]
- Kolb, H.; Martin, S. Environmental/lifestyle Factors in the Pathogenesis and Prevention of Type 2 Diabetes. BMC Med. 2017, 15, 131. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, B.; Zawawi, N.; Omar, A.R.; Ismail, M. Predisposition to Insulin Resistance and Obesity due to Staple Consumption of Rice: Amylose Content versus Germination Status. PLoS ONE 2017, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.A. Smoking and Type 2 Diabetes Mellitus. Diabetes Metab. J. 2012, 36, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Toorie, A.M.; Vassoler, F.M.; Qu, F.; Schonhoff, C.M.; Bradburn, S.; Murgatroyd, C.A.; Slonim, D.K.; Byrnes, E.M. A History of Opioid Exposure in Females Increases the Risk of Metabolic Disorders in Their Future Male Offspring. Addict Biol 2021, 26, e12856. [Google Scholar] [CrossRef]
- Uusitupa, M. Lifestyles Matter in the Prevention of Type 2 Diabetes. Diabetes Care 2002, 25, 1650–1651. [Google Scholar] [CrossRef]
- Silva, D.A.S.; Naghavi, M.; Duncan, B.B.; Schmidt, M.I.; de Souza, M. de F.M.; Malta, D.C. Physical Inactivity as Risk Factor for Mortality by Diabetes Mellitus in Brazil in 1990, 2006, and 2016. Diabetol. Metab. Syndr. 2019, 11, 23. [Google Scholar] [CrossRef]
- Al-Yasari, A.; Jabbar, S.; Cabrera, M.A.; Rousseau, B.; Sarkar, D.K. Preconception Alcohol Exposure Increases the Susceptibility to Diabetes in the Offspring. Endocrinology 2021, 162. [Google Scholar] [CrossRef]
- Holst, C.; Becker, U.; Jørgensen, M.E.; Grønbæk, M.; Tolstrup, J.S. Alcohol Drinking Patterns and Risk of Diabetes: A Cohort Study of 70,551 Men and Women from the General Danish Population. Diabetologia 2017, 60, 1941–1950. [Google Scholar] [CrossRef] [PubMed]
- Da Porto, A.; Cavarape, A.; Colussi, G.; Casarsa, V.; Catena, C.; Sechi, L.A. Polyphenols Rich Diets and Risk of Type 2 Diabetes. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Tertsunen, H.M.; Hantunen, S.; Tuomainen, T.P.; Virtanen, J.K. Adherence to a Healthy Nordic Diet and Risk of Type 2 Diabetes among Men: The Kuopio Ischaemic Heart Disease Risk Factor Study. Eur J Nutr 2021, 60, 3927–3934. [Google Scholar] [CrossRef]
- Shu, L.; Shen, X.M.; Li, C.; Zhang, X.Y.; Zheng, P.F. Dietary Patterns Are Associated with Type 2 Diabetes Mellitus among Middle-Aged Adults in Zhejiang Province, China. Nutr J 2017, 16, 81. [Google Scholar] [CrossRef]
- Imam, M.U.; Ishaka, A.; Ooi, D.J.; Zamri, N.D.M.; Sarega, N.; Ismail, M.; Esa, N.M. Germinated Brown Rice Regulates Hepatic Cholesterol Metabolism and Cardiovascular Disease Risk in Hypercholesterolaemic Rats. J. Funct. Foods 2014, 8, 193–203. [Google Scholar] [CrossRef]
- Stegemann, R.; Buchner, D.A. Transgenerational Inheritance of Metabolic Disease. Semin Cell Dev Biol 2015, 43, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Steyn, N.P.; Mann, J.; Bennett, P.H.; Temple, N.; Zimmet, P.; Tuomilehto, J.; Lindström, J.; Louheranta, A. Diet, Nutrition and the Prevention of Type 2 Diabetes. Public Health Nutr 2004, 7, 147–165. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, B.; Yakasai, H.M.; Zawawi, N.; Ismail, M. Compositional Analyses of White, Brown and Germinated Forms of Popular Malaysian Rice to Offer Insight into the Growing Diet-Related Diseases. J. Food Drug Anal. 2018, 26, 706–715. [Google Scholar] [CrossRef]
- Kusuyama, J.; Makarewicz, N.S.; Albertson, B.G.; Alves-Wagner, A.B.; Conlin, R.H.; Prince, N.B.; Alves, C.R.R.; Ramachandran, K.; Kozuka, C.; Xiudong, Y.; et al. Maternal Exercise-Induced SOD3 Reverses the Deleterious Effects of Maternal High-Fat Diet on Offspring Metabolism Through Stabilization of H3K4me3 and Protection Against WDR82 Carbonylation. Diabetes 2022, 71, 1170–1181. [Google Scholar] [CrossRef]
- Liu, S.; Manson, J.E.; Stampfer, M.J.; Hu, F.B.; Giovannucci, E.; Colditz, G.A.; Hennekens, C.H.; Willett, W.C. A Prospective Study of Whole-Grain Intake and Risk of Type 2 Diabetes Mellitus in US Women. Am J Public Health 2000, 90, 1409–1415. [Google Scholar] [CrossRef]
- Maki, K.C.; Phillips, A.K. Dietary Substitutions for Refined Carbohydrate That Show Promise for Reducing Risk of Type 2 Diabetes in Men and Women. J Nutr 2015, 145, 159s–163s. [Google Scholar] [CrossRef]
- Samra, R.A.; Anderson, G.H. Insoluble Cereal Fiber Reduces Appetite and Short-Term Food Intake and Glycemic Response to Food Consumed 75 Min Later by Healthy Men. Am. J. Clin. Nutr. 2007, 86, 972–979. [Google Scholar] [CrossRef]
- Arihara, K. Functional Foods. Encycl. Meat Sci. 2014, 2, 32–36. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Hoffmann, G.; Lampousi, A.M.; Knüppel, S.; Iqbal, K.; Schwedhelm, C.; Bechthold, A.; Schlesinger, S.; Boeing, H. Food Groups and Risk of Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis of Prospective Studies. Eur J Epidemiol 2017, 32, 363–375. [Google Scholar] [CrossRef]
- Wu, C.; Morris, J.R. Genes, Genetics, and Epigenetics: A Correspondence. Science 2001, 293, 1103–1105. [Google Scholar] [CrossRef] [PubMed]
- Gibney, E.R.; Nolan, C.M. Epigenetics and Gene Expression. Heredity ( 2010, 105, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Sanusi, K.O.; Uthman, Y.A.; Ooi, D.J.; Ismail, M.; Imam, M.U. Lifestyle and Preventive Medical Epigenetics. Med. Epigenetics 2021, 33–50. [Google Scholar] [CrossRef]
- Ahmed, S.A.H.; Ansari, S.A.; Mensah-Brown, E.P.K.; Emerald, B.S. The Role of DNA Methylation in the Pathogenesis of Type 2 Diabetes Mellitus. Clin. Epigenetics 2020, 12. [Google Scholar] [CrossRef]
- Ling, C.; Rönn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef]
- Ling, C.; Rönn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef]
- Daneshpajooh, M.; Bacos, K.; Bysani, M.; Bagge, A.; Ottosson Laakso, E.; Vikman, P.; Eliasson, L.; Mulder, H.; Ling, C. HDAC7 Is Overexpressed in Human Diabetic Islets and Impairs Insulin Secretion in Rat Islets and Clonal Beta Cells. Diabetologia 2017, 60, 116–125. [Google Scholar] [CrossRef]
- Tobi, E.W.; Slieker, R.C.; Luijk, R.; Dekkers, K.F.; Stein, A.D.; Xu, K.M.; Slagboom, P.E.; van Zwet, E.W.; Lumey, L.H.; Heijmans, B.T. DNA Methylation as a Mediator of the Association between Prenatal Adversity and Risk Factors for Metabolic Disease in Adulthood. Sci. Adv. 2018, 4, eaao4364. [Google Scholar] [CrossRef]
- Li, J.; Li, K.; Gao, J.; Guo, X.; Lu, M.; Li, Z.; Li, D. Maternal Exposure to an N-3 Polyunsaturated Fatty Acid Diet Decreases Mammary Cancer Risk of Female Offspring in Adulthood. Food Funct 2018, 9, 5768–5777. [Google Scholar] [CrossRef]
- Sun, Q.; Huang, S.; Wang, X.; Zhu, Y.; Chen, Z.; Chen, D. N6-Methyladenine Functions as a Potential Epigenetic Mark in Eukaryotes. Bioessays 2015, 37, 1155–1162. [Google Scholar] [CrossRef]
- Deaton, A.M.; Bird, A. CpG Islands and the Regulation of Transcription. Genes Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Huang, H.; Liu, D.; Cheng, Y.; Liu, X.; Zhang, W.; Yin, R.; Zhang, D.; Zhang, P.; Liu, J.; et al. N6-Methyladenine DNA Modification in Drosophila. Cell 2015, 161, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Pinney, S.E.; Pinney, S.E.O. DNA Methylation and Its Role in the Pathogenesis of Diabetes. Pediatr. Diabetes 2017, 18, 167–177. [Google Scholar]
- Zhang, Q.; Xiao, X.; Zheng, J.; Li, M.; Yu, M.; Ping, F.; Wang, T.; Wang, X. A Maternal High-Fat Diet Induces DNA Methylation Changes That Contribute to Glucose Intolerance in Offspring. Front. Endocrinol. 2019, 10, 871. [Google Scholar] [CrossRef] [PubMed]
- Blake, G.E.T.; Watson, E.D. Unravelling the Complex Mechanisms of Transgenerational Epigenetic Inheritance. Curr. Opin. Chem. Biol. 2016, 33, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Alaskhar Alhamwe, B.; Khalaila, R.; Wolf, J.; von Bülow, V.; Harb, H.; Alhamdan, F.; Hii, C.S.; Prescott, S.L.; Ferrante, A.; Renz, H.; et al. Histone Modifications and Their Role in Epigenetics of Atopy and Allergic Diseases. Allergy Asthma Clin Immunol 2018, 14, 39. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Luan, Y.; Feng, Q.; Chen, X.; Qin, B.; Ren, K. Di; Luan, Y. Epigenetics and Beyond: Targeting Histone Methylation to Treat Type 2 Diabetes Mellitus. Front. Pharmacol. 2022, 12. [Google Scholar] [CrossRef]
- Casas-Agustench, P.; Fernandes, F.S.; Tavares do Carmo, M.G.; Visioli, F.; Herrera, E.; Dávalos, A. Consumption of Distinct Dietary Lipids during Early Pregnancy Differentially Modulates the Expression of microRNAs in Mothers and Offspring. PLoS ONE 2015, 10, e0117858. [Google Scholar] [CrossRef]
- Mantilla-Escalante, D.C.; López de las Hazas, M.-C.; Crespo, M.C.; Martín-Hernández, R.; Tomé-Carneiro, J.; del Pozo-Acebo, L.; Salas-Salvadó, J.; Bulló, M.; Dávalos, A. Mediterranean Diet Enriched in Extra-Virgin Olive Oil or Nuts Modulates Circulating Exosomal Non-Coding RNAs. Eur J Nutr 2021, 60, 4279–4293. [Google Scholar] [CrossRef]
- Núñez-Acuña, G.; Détrée, C.; Gallardo-Escárate, C.; Gonçalves, A.T. Functional Diets Modulate lncRNA-Coding RNAs and Gene Interactions in the Intestine of Rainbow Trout Oncorhynchus Mykiss. Mar. Biotechnol. 2017, 19, 287–300. [Google Scholar] [CrossRef]
- Zhang, Q.; Xiao, X.; Zheng, J.; Li, M.; Yu, M.; Ping, F.; Wang, T.; Wang, X. Improvement in Glucose Metabolism in Adult Male Offspring of Maternal Mice Fed Diets Supplemented with Inulin via Regulation of the Hepatic Long Noncoding RNA Profile. Faseb J 2021, 35, e22003. [Google Scholar] [CrossRef] [PubMed]
- Formichi, C.; Nigi, L.; Grieco, G.E.; Maccora, C.; Fignani, D.; Brusco, N.; Licata, G.; Sebastiani, G.; Dotta, F. Non-Coding Rnas: Novel Players in Insulin Resistance and Related Diseases. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Chi, T.; Lin, J.; Wang, M.; Zhao, Y.; Liao, Z.; Wei, P. Non-Coding RNA as Biomarkers for Type 2 Diabetes Development and Clinical Management. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.F.; Lehner, B. Intergenerational and Transgenerational Epigenetic Inheritance in Animals. Nat Cell Biol 2019, 21, 143–151. [Google Scholar] [CrossRef]
- Gill-Randall, R.; Adams, D.; Ollerton, R.L.; Lewis, M.; Alcolado, J.C. Type 2 Diabetes Mellitus--Genes or Intrauterine Environment? An Embryo Transfer Paradigm in Rats. Diabetologia 2004, 47, 1354–1359. [Google Scholar] [CrossRef]
- Dabelea, D.; Hanson, R.L.; Lindsay, R.S.; Pettitt, D.J.; Imperatore, G.; Gabir, M.M.; Roumain, J.; Bennett, P.H.; Knowler, W.C. Intrauterine Exposure to Diabetes Conveys Risks for Type 2 Diabetes and Obesity: A Study of Discordant Sibships. Diabetes 2000, 49, 2208–2211. [Google Scholar] [CrossRef]
- Jimenez-Chillaron, J.C.; Isganaitis, E.; Charalambous, M.; Gesta, S.; Pentinat-Pelegrin, T.; Faucette, R.R.; Otis, J.P.; Chow, A.; Diaz, R.; Ferguson-Smith, A.; et al. Intergenerational Transmission of Glucose Intolerance and Obesity by in Utero Undernutrition in Mice. Diabetes 2009, 58, 460–468. [Google Scholar] [CrossRef]
- Pavlinkova, G.; Margaryan, H.; Zatecka, E.; Valaskova, E.; Elzeinova, F.; Kubatova, A.; Bohuslavova, R.; Peknicova, J. Transgenerational Inheritance of Susceptibility to Diabetes-Induced Male Subfertility. Sci. Rep. 2017, 7, 4940. [Google Scholar] [CrossRef]
- Sánchez-Soriano, C.; Pearson, E.R.; Reynolds, R.M. Associations between Parental Type 2 Diabetes Risk and Offspring Birthweight and Placental Weight: A Survival Analysis Using the Walker Cohort. Diabetologia 2022. [Google Scholar] [CrossRef]
- Crudo, A.; Petropoulos, S.; Moisiadis, V.G.; Iqbal, M.; Kostaki, A.; Machnes, Z.; Szyf, M.; Matthews, S.G. Prenatal Synthetic Glucocorticoid Treatment Changes DNA Methylation States in Male Organ Systems: Multigenerational Effects. Endocrinology 2012, 153, 3269–3283. [Google Scholar] [CrossRef]
- Painter, R.C.; Osmond, C.; Gluckman, P.; Hanson, M.; Phillips, D.I.; Roseboom, T.J. Transgenerational Effects of Prenatal Exposure to the Dutch Famine on Neonatal Adiposity and Health in Later Life. Bjog 2008, 115, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Barella, L.F.; de Oliveira, J.C.; Mathias, P.C. Pancreatic Islets and Their Roles in Metabolic Programming. Nutrition 2014, 30, 373–379. [Google Scholar] [CrossRef]
- Bouret, S.; Levin, B.E.; Ozanne, S.E. Gene-Environment Interactions Controlling Energy and Glucose Homeostasis and the Developmental Origins of Obesity. Physiol. Rev. 2015, 95, 47–82. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.H.; Haase, T.N.; Jaksch, C.; Nalla, A.; Søstrup, B.; Nalla, A.A.; Larsen, L.; Rasmussen, M.; Dalgaard, L.T.; Gaarn, L.W.; et al. Impact of Fetal and Neonatal Environment on Beta Cell Function and Development of Diabetes. Acta Obs. Gynecol Scand 2014, 93, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Ockner, R.K.; Manning, J.A. Fatty Acid-Binding Protein in Small Intestine. Identification, Isolation, and Evidence for Its Role in Cellular Fatty Acid Transport. J Clin Invest 1974, 54, 326–338. [Google Scholar] [CrossRef]
- Hasler, C.M. Functional Foods: Benefits, Concerns and Challenges - A Position Paper from the American Council on Science and Health. J. Nutr. 2002, 132, 3772–3781. [Google Scholar] [CrossRef]
- Temple, N.J. A Rational Definition for Functional Foods: A Perspective. Front. Nutr. 2022, 9. [Google Scholar] [CrossRef]
- Gul, K.; Singh, A.K.; Jabeen, R. Nutraceuticals and Functional Foods: The Foods for the Future World. Crit. Rev. Food Sci. Nutr. 2016, 56, 2617–2627. [Google Scholar] [CrossRef]
- Magrone, T.; de Heredia, F.P.; Jirillo, E.; Morabito, G.; Marcos, A.; Serafini, M. Functional Foods and Nutraceuticals as Therapeutic Tools for the Treatment of Diet-Related Diseases. Can. J. Physiol. Pharmacol. 2013, 91, 387–396. [Google Scholar] [CrossRef]
- Bansal, A.; Rashid, C.; Xin, F.; Li, C.; Polyak, E.; Duemler, A.; van der Meer, T.; Stefaniak, M.; Wajid, S.; Doliba, N.; et al. Sex- and Dose-Specific Effects of Maternal Bisphenol A Exposure on Pancreatic Islets of First- and Second-Generation Adult Mice Offspring. Env. Health Perspect 2017, 125, 97022. [Google Scholar] [CrossRef]
- Silva, R.C. da; Colleran, H.L.; Ibrahim, S.A. Milk Fat Globule Membrane in Infant Nutrition: A Dairy Industry Perspective. J. Dairy Res. 2021, 88, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, Q.; Xin, F.; Cao, B.; Qian, L.; Dong, Y. Neonatal Milk Fat Globule Membrane Supplementation During Breastfeeding Ameliorates the Deleterious Effects of Maternal High-Fat Diet on Metabolism and Modulates Gut Microbiota in Adult Mice Offspring in a Sex-Specific Way. Front Cell Infect Microbiol 2021, 11, 621957. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.H.; Badr, G.; El Shinnawy, N.A. Camel Whey Protein Improves Lymphocyte Function and Protects against Diabetes in the Offspring of Diabetic Mouse Dams. Int J Immunopathol Pharmacol 2016, 29, 632–646. [Google Scholar] [CrossRef] [PubMed]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Adamu, H.A.; Imam, M.U.; Ooi, D.J.; Esa, N.M.; Rosli, R.; Ismail, M. In Utero Exposure to Germinated Brown Rice and Its Oryzanol-Rich Extract Attenuated High Fat Diet-Induced Insulin Resistance in F1 Generation of Rats. BMC Complement Altern Med 2017, 17, 67. [Google Scholar] [CrossRef] [PubMed]
- Dubey, P.; Thakur, V.; Chattopadhyay, M. Role of Minerals and Trace Elements in Diabetes and Insulin Resistance. Nutrients 2020, 12, 1864. [Google Scholar] [CrossRef] [PubMed]
- Taneja, S.K.; Mandal, R. Effect of Modified Egg* on Developmental Defects in Neonates of NIDDM Induced Wistar Rats. Indian J Exp Biol 2006, 44, 863–874. [Google Scholar]
- Taneja, S.K.; Singh, K.B. Beneficial Effects of Modified Egg* on Oxidative Stress in F1- Generation of Metabolic Syndrome-X Induced Wistar Rat. Indian J Exp Biol 2009, 47, 104–112. [Google Scholar]
- Asmat, U.; Abad, K.; Ismail, K. Diabetes Mellitus and Oxidative stress—A Concise Review. Saudi Pharm. J. 2016, 24. [Google Scholar] [CrossRef] [PubMed]
- Grunnet, L.G.; Pilgaard, K.; Alibegovic, A.; Jensen, C.B.; Hjort, L.; Ozanne, S.E.; Bennett, M.; Vaag, A.; Brøns, C. Leukocyte Telomere Length Is Associated with Elevated Plasma Glucose and HbA1c in Young Healthy Men Independent of Birth Weight. Sci. Rep. 2019, 9, 7639. [Google Scholar] [CrossRef]
- Wang, J.; Dong, X.; Cao, L.; Sun, Y.; Qiu, Y.; Zhang, Y.; Cao, R.; Covasa, M.; Zhong, L. Association between Telomere Length and Diabetes Mellitus: A Meta-Analysis. J. Int. Med. Res. 2016, 44, 1156–1173. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xiao, H.; Li, J.; Guo, X.; Cai, W.; Li, D. N-3 Polyunsaturated Fatty Acids Decrease Long-Term Diabetic Risk of Offspring of Gestational Diabetes Rats by Postponing Shortening of Hepatic Telomeres and Modulating Liver Metabolism. Nutrients 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Bhupathiraju, S.N.; Hu, F.B. Epidemiology of Obesity and Diabetes and Their Cardiovascular Complications. Circ. Res. 2016, 118, 1723–1735. [Google Scholar] [CrossRef]
- Ma, L.; Sun, Z.; Zeng, Y.; Luo, M.; Yang, J. Molecular Mechanism and Health Role of Functional Ingredients in Blueberry for Chronic Disease in Human Beings. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef]
- Sulforaphane Protects against Cytokine- and Streptozotocin-Induced β-Cell Damage by Suppressing the NF-κB Pathway. Toxicol. Appl. Pharmacol. 2009, 235, 57–67. [CrossRef] [PubMed]
- Shanak, S.; Saad, B.; Zaid, H.; Carvalho, J.C.T. Metabolic and Epigenetic Action Mechanisms of Antidiabetic Medicinal Plants. Evidence-based Complement. Altern. Med. 2019, 2019. [Google Scholar] [CrossRef]
- Kaur, K.K.; Allahbadia, G.; Singh, M. Therapeutic Potential and Epigenetic Alterations of Plant Phytochemicals (as Epi-Drugs) for the Treatment of Type 2 Diabetes Mellitus: A Systematic Review. Adv Obes Weight Manag. 2021. [Google Scholar]
- Mousavi, A.; Vafa, M.; Neyestani, T.; Khamseh, M.; Hoseini, F. The Effects of Green Tea Consumption on Metabolic and Anthropometric Indices in Patients with Type 2 Diabetes. J. Res. Med. Sci. 2013, 18, 1080–1086. [Google Scholar]
- Ramírez-Alarcón, K.; Victoriano, M.; Mardones, L.; Villagran, M.; Al-Harrasi, A.; Al-Rawahi, A.; Cruz-Martins, N.; Sharifi-Rad, J.; Martorell, M. Phytochemicals as Potential Epidrugs in Type 2 Diabetes Mellitus. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef]
- Zhong, J.; Xu, C.; Reece, E.A.; Yang, P. The Green Tea Polyphenol EGCG Alleviates Maternal Diabetes–induced Neural Tube Defects by Inhibiting DNA Hypermethylation. Am. J. Obstet. Gynecol. 2016, 215, 368–e1. [Google Scholar] [CrossRef]
- K. , G.-B.; O., R.-L.; E., B.-C.; J.I., R.-B.; F.I., M.; E., M.-L.; J.A., M. Fatty Acids, Epigenetic Mechanisms and Chronic Diseases: A Systematic Review. Lipids Health Dis. 2019, 18, 178. [Google Scholar] [CrossRef]
- Hussey, B.; Lindley, M.R.; Mastana, S.S.; Mastana, S.S. Omega 3 Fatty Acids, Inflammation and DNA Methylation: An Overview. Clin. Lipidol. 2017, 12, 24–39. [Google Scholar]
- S. , R.; S.C., G.; B., P.; A., G.; B.B., A. Epigenetic Changes Induced by Curcumin and Other Natural Compounds. Genes Nutr. 2011, 6, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.M.; Jialal, I.; Devaraj, S. Epigenetic Regulation of High Glucose-Induced Proinflammatory Cytokine Production in Monocytes by Curcumin. J. Nutr. Biochem. 2011, 22, 450–458. [Google Scholar] [CrossRef]
- Tang, C.; Liu, Y.; Liu, S.; Yang, C.; Chen, L.; Tang, F.; Wang, F.; Zhan, L.; Deng, H.; Zhou, W.; et al. Curcumin and Its Analogs as Potential Epigenetic Modulators: Prevention of Diabetes and Its Complications. Pharmacology 2022, 107, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.C.W.; Popkin, B.M. Intergenerational Diabetes and obesity—A Cycle to Break? PLOS Med. 2017, 14, e1002415. [Google Scholar] [CrossRef]
- Marangoni, F.; Cetin, I.; Verduci, E.; Canzone, G.; Giovannini, M.; Scollo, P.; Corsello, G.; Poli, A. Maternal Diet and Nutrient Requirements in Pregnancy and Breastfeeding. An Italian Consensus Document. Nutrients 2016, 8, 629. [Google Scholar] [CrossRef]
- Marciniak, A.; Patro-Małysza, J.; Kimber-Trojnar, Ż.; Marciniak, B.; Oleszczuk, J.; Leszczyńska-Gorzelak, B. Fetal Programming of the Metabolic Syndrome. Taiwan. J. Obstet. Gynecol. 2017, 56, 133–138. [Google Scholar] [CrossRef]
- Sinclair, K.D.; Allegrucci, C.; Singh, R.; Gardner, D.S.; Sebastian, S.; Bispham, J.; Thurston, A.; Huntley, J.F.; Rees, W.D.; Maloney, C.A.; et al. DNA Methylation, Insulin Resistance, and Blood Pressure in Offspring Determined by Maternal Periconceptional B Vitamin and Methionine Status. Proc Natl Acad Sci U S A 2007, 104, 19351–19356. [Google Scholar] [CrossRef]
- Kazeem, M.I.; Davies, T.C. Anti-Diabetic Functional Foods as Sources of Insulin Secreting, Insulin Sensitizing and Insulin Mimetic Agents. J. Funct. Foods 2016, 20, 122–138. [Google Scholar] [CrossRef]
- Neacsu, M.; Raikos, V.; Benavides-Paz, Y.; Duncan, S.H.; Duncan, G.J.; Christie, J.S.; Johnstone, A.M.; Russell, W.R. Sapogenol Is a Major Microbial Metabolite in Human Plasma Associated with High Protein Soy-Based Diets: The Relevance for Functional Food Formulations. Foods 2020, 9. [Google Scholar] [CrossRef]
- Plat, J.; Baumgartner, S.; Vanmierlo, T.; Lütjohann, D.; Calkins, K.L.; Burrin, D.G.; Guthrie, G.; Thijs, C.; Te Velde, A.A.; Vreugdenhil, A.C.E.; et al. Plant-Based Sterols and Stanols in Health & disease: “Consequences of Human Development in a Plant-Based Environment? ” Prog Lipid Res 2019, 74, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Chu, Y. Whole Grain Oats, More than Just a Fiber: Role of Unique Phytochemicals. Mol Nutr Food Res 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Tessari, P.; Lante, A. A Multifunctional Bread Rich in Beta Glucans and Low in Starch Improves Metabolic Control in Type 2 Diabetes: A Controlled Trial. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Alves-de-Oliveira, D.S.; Bloise, A.; Silva, L.M.L.; Rocha-Junior, R.L.; Lima-Júnior, N.C.; Menezes, L.G.S.; Silva, E.G.S.; De Oliveira, Y.; Wanderley, A.G.; de-Brito-Alves, J.L.; et al. Maternal Consumption of ɷ3 Attenuates Metabolic Disruption Elicited by Saturated Fatty Acids-Enriched Diet in Offspring Rats. Nutr Metab Cardiovasc Dis 2022, 32, 279–289. [Google Scholar] [CrossRef]
- Ueno, M.; Liu, S.; Kiyoi, T.; Sugiyama, T.; Mogi, M. Perinatal Low-Fat Dietary Intervention Affects Glucose Metabolism in Female Adult and Aging Offspring. Geriatr Gerontol Int 2022, 22, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, L.; Menikdiwela, K.R.; Spainhour, S.; Eboh, T.; Moustaid-Moussa, N. Sex Differences in Early Programming by Maternal High Fat Diet Induced-Obesity and Fish Oil Supplementation in Mice. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Wang, D.D.; Wu, F.; Zhang, L.Y.; Zhao, Y.C.; Wang, C.C.; Xue, C.H.; Yanagita, T.; Zhang, T.T.; Wang, Y.M. Effects of Dietary N-3 PUFA Levels in Early Life on Susceptibility to High-Fat-Diet-Induced Metabolic Syndrome in Adult Mice. J Nutr Biochem 2021, 89, 108578. [Google Scholar] [CrossRef]
- Keller, A.; Ängquist, L.; Jacobsen, R.; Vaag, A.; Heitmann, B.L. A Retrospective Analysis of a Societal Experiment among the Danish Population Suggests That Exposure to Extra Doses of Vitamin A during Fetal Development May Lower Type 2 Diabetes Mellitus (T2DM) Risk Later in Life. Br J Nutr 2017, 117, 731–736. [Google Scholar] [CrossRef]
- Hollander, K.S.; Tempel Brami, C.; Konikoff, F.M.; Fainaru, M.; Leikin-Frenkel, A. Dietary Enrichment with Alpha-Linolenic Acid during Pregnancy Attenuates Insulin Resistance in Adult Offspring in Mice. Arch Physiol Biochem 2014, 120, 99–111. [Google Scholar] [CrossRef]
Figure 1.
Modifiable and unmodifiable factors leading to epigenetic perturbations in type 2 diabetes and metabolic disorders.
Figure 1.
Modifiable and unmodifiable factors leading to epigenetic perturbations in type 2 diabetes and metabolic disorders.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



