Submitted:
26 May 2023
Posted:
30 May 2023
You are already at the latest version
Abstract
Affem Boussou community forest (AFC) abounds in important biological resources. This study, which contributes to its better management, examines the spatiotemporal dynamics of the vegetation and its ecological and structural characteristics to propose a zoning plan for the said forest. The analysis of the spatiotemporal dynamics of land use in the AFC from Google Earth images of 2015, 2018, and 2021 revealed a regressive trend of formations: crops and fallows (-33.98%), dense dry forests (-7.92%), gallery forests (-3.46%), plantations (-100%), grassy savannahs and meadows (-18.84%) except for tree/shrub savannahs (484.23%). The floristic inventory identified 163 species divided into 129 genera and 55 families. Fabaceae (14.02%), and Combretaceae (10.55%) are the most represented families. Anogeissus leiocarpa (5.19%) and Vitellaria paradoxa (4.72%) are the most frequent species. We note the dominance of individuals of small diameters. The regeneration potential of the AFC is 64 feet/ha due to 21 feet/ha of suckers, 29 feet/ha of seedlings, and 14 feet/ha of shoots. As a zoning plan, the AFC was subdivided into four series: the agroforestry zone (18.80%), the sustainable production forest zone (42.22%), the buffer zone (11%), and the biological conservation zone (28%). These results constitute a scientific basis for testing ecological indicators of sustainable management of community forests in Togo.
Keywords:
land use changes
; zoning
; community forest
; resilience
; Togo
1. Introduction
Ecosystem degradation evolves with bioclimatic conditions and anthropogenic action [1,2]. A direct consequence of this degradation is the fragmentation of plant formations [3]. This is compounded by a certain lack of institutional willingness and the absence of adequate management tools. As a result, forest density may decrease significantly, and some species may be extinguished [4]. From 1990 to 2015, the world’s total forest area decreased from 31.6% to 30.6% [5]. This forest cover has continued to decline since the beginning of the century in the intertropical zone. Concomitantly with this reduction in forest cover, cultivated areas have increased considerably [6,7], leading to a degradation characterized by an increasing loss of their floristic diversity procession.
In West Africa, numerous studies [8,9,10,11] have shown that human activities (slash-and-burn agriculture, ranches and other rangelands, mining, urbanization) negatively impact the structure, the floristic composition, and the dynamics of natural forests. The most dominant forms of disturbance that contribute to shaping the structure and physiognomy of the vegetation [7,12,13] include wildfires, anarchic exploitation of woody resources, shortening of the duration of fallows, and land clearing coupled with hydrological stress related to irregular rainfall.
For decades, village communities in Togo have preserved portions of their land for hunting, biodiversity conservation, land reserve, cultural, and spiritual purposes. Some of these areas, which today constitute community forests, are managed according to traditional rules. The capacity of these areas to conserve fauna and flora, the effectiveness of their management, and interactions with riparian communities are also hotly debated [7,14]. The main products derived from these forest ecosystems contribute to food security and are sources of income for the populations [15]. Therefore knowledge of all ecological and physiognomic aspects of the vegetation is essential to assess the resources of these areas.
In recent years, the central region of Togo has faced significant degradation of forest ecosystems, with negative repercussions for the population. This situation has caused a substantial loss of vegetation cover, leading to rapid land degradation and a significant loss of biodiversity. Between 1986 and 2003, open and dry forests and forest-savannah mosaics saw a 26.52% regression versus an increase against an increase in anthropogenic formations of around 26.48% [16]. This degradation not only adds to ecosystem depletion but also jeopardizes the quality of life and potentially the survival of vulnerable populations that rely heavily on ecological services. Being one of the leading Wood energy suppliers in Togo, with more than 100 localities with high wood energy productivity [17], the resources of this region are highly coveted. The Affem Boussou community forest in the Tchamba prefecture is not spared from this situation. For the best management of these ecosystems, it is necessary to question the spatial translation of this ecosystem degradation. What is its extent to date? The community forest plays a crucial role in the conservation and management of biodiversity hence the interest in analyzing the spatiotemporal dynamics of the vegetation. The overall goal of this research is to contribute to advancing information on forest dynamics in Togo better to direct the country’s forest management strategies and policies. More specifically, it is about analyzing land use changes between 2015, 2018, and 2021, characterizing the structural diversity of the ecosystem, and providing a zoning plan for the mentioned community forest.
2. Materials and Methods
2.1. Study Area
Affem community forest is located in the prefecture of Tchamba, more specifically in the canton of Affem (Figure 1). It is situated between 9°11′ and 9°13′ North latitude and 1°55′ and 1°56′ East latitude. It covers an area of 82,19 ha. It belongs to ecological zone III [18], in the Benin-Togolese Peneplain, whose altitude varies between 200 and 400 m. and rests on a granite-gneissic crystalline base of the Benin-Togolese Plain structural unit [19]. Plant forms of Affem can be found on a wide range of soils, the most common of which are leached tropical ferruginous soils, soils with limited erosion, ferralitic soils, and vertisols. It has a Sudanian-type climate with a wet season from April to October and a dry season from November to March [20,21]. The annual rainfall ranges between 1200 and 1300 mm. The relative humidity ranges from 60 to 80%, while the evaporation rate is approximately 1600 mm/year. Monthly temperatures range from 25 to 38 °C (with a monthly average of 26,5 °C). The vegetation in the study site is essentially characterized by Guinean wooded savannahs [19]. Vitellaria paradoxa, Tectona grandis, Terminalia spp, Lonchocarpus sericeus, Piliostigma thonningii, and Pterocarpus erinaceus are the most common woody species encountered. The fauna is dominated by rodents (rats, squirrels, agoutis, wild mice, hares), reptiles (green mambas, lizards), and birds (coliforms, gruiforms, columbines, cuculiforms, partridges, parrots).
Tchamba Prefecture is home to a diverse range of ethnic groups. Among them, Tchamba, Kamboli, Koussountou, Bago, Kotokoli, Kabyè etc. Tchamba had 34626 residents at the time of the first census of 1970 [22]. It climbed to 46,000 people in the second census in 1981, 74,000 people in 2004, and 131674 people in 2010. It has a young population, around 20% living in the city core, and is distinguished by rapid growth. The population of the villages Adjeidè (Kri-Kri), Larini, and Affem, considered the AFC riparian population, is estimated to be 25000 people. Agriculture and commerce are the primary activities in Affem. The township provides cereals, tubers, cashew nuts, and charcoal to the nearby towns, mainly Tchamba [23].
2.2. Data Collection
2.2.1. Images Acquisition
High-resolution images covering three (3) different years (2015, 2018, and 2021) were downloaded from several online geospatial data sources. Thus, using the add-on “open layer plugging” and shapefile of the study area, the ESRI image of 2021 followed by the digital globe image of 2018 and an aerial image of 2015 were respectively downloaded with ESRI map, google satellite and Bing aerial plugging. The choice of these images and their temporality is justified by their availability and their free access [24,25]. The relatively small size area of the studied zone has also guided the use of high-resolution images with the need to map the significant spatial entities’ features significantly [26,27].
The land classification system was derived from the national one adapted to FAO LCS [28]. The riparian forest, dense dry forest, plantations, open forest, shrub/tree savannah, and fallow/cropland are the important land cover type identified after visual interpretation of the images and exploitation of auxiliaries’ earth observation data [29,30]. The digitization from the screen was carried out. After digitalization, post-classification processing was performed, such as thematic vector analysis, land cover type area computation [31,32], and layout of the time series map.
Finally, the land cover type dynamic was analyzed to assess changes in land use type between the three different periods [33,34]. Differences among a given land cover type between two periods were determined by the relationship below
Where ΔU represents the annual rate of change, U2 land cover type for the upper year, U1 land cover type for the lower year, and n the number of years between the two dates.
2.2.2. Sampling
From the Affem Boussou Community Forest (AFC) vector file and QGIS 3.83 software (Folega et al. 2021), 34 regular gird points space from each other of 100 m were generated as forest inventory plot’s location (Figure 1). This choice is explained by the willingness to systematically sample plant species in all the different forms of land use in the AFC.
2.2.3. Affem Boussou Community Forest (AFC) Plant Resource Inventories
The floristic inventory consisted of recording all woody species in plots and herbaceous species in 10 m x 10 m subplots. All species were assigned their abundance/dominance coefficient according to the Braun-Blanquet scale (Braun Blanquet, 1932). The choice of the technique is motivated by its successful use in Togo by previous authors with the same research topic [37,38].
The forest inventory consisted of measuring the height of the bole, the total tree height of woody species, and the diameter at breast height (DBH) ≥ 10 cm [39]. All tree diameters were measured using a measuring tape at 1.30 m from the ground. Bole and total tree height were estimated by visual notation.
The regeneration of the species was inventoried in 3 sub-plots of 2 m x 2 m installed diagonally in the middle of each plot. It was done on individuals with a diameter between 5 and 10 cm, considered as potential regeneration [4]. The regeneration mode (natural seedlings, stump sprouts, suckers) was also considered [40].
The ecological inventory was carried out in 50 m x 20 m plots. Ecological variables such as topography, overall vegetation cover, and evidence of human activities were recorded within each plot. Formation types were also identified. Coordinates of all inventory plots were recorded with a GPS (Global Positioning System) receiver.
2.3.2. Analysis of Floristic Diversity
The analysis of the floristic diversity allowed us to draw up a list of the species recorded and to group them by family. A matrix “surveys x species” was elaborated based on the presence and absence of species: in rows the species and in columns the surveys. The elaborated matrix was submitted to multivariate analysis techniques to highlight the major ecological gradients as well as the identification and the typology of major plant communities that emerge from them [41]. Thus the matrix was subjected to DECORANA® (DEtrended CORrespondence ANAlysis) method for plant community identification purposes [42,43] with CANOCO (CANOnial Community Ordination) software [44,45]. For each identified plant grouping, the list of flora affected, followed by its corresponding family, life forms [46], and chorology [47] was assigned. The alpha diversity, such as specific richness, specific frequencies, and abundances, was determined [48]. It is followed by Shannon Index (Ish) and Pielou Equitability Index (Eq) [49] for both plant communities. The formula for each of these indices is shown in the table below.

Table 1.
Components of alpha diversity.
| Parameters | Formula | References |
|---|---|---|
| Shannon Index (Ish) | Ish = -∑(ni/n) log2 (ni/n) | [49] |
| Pielou Equitability Index (E) | Eq = isℎ /log2S | [41,48] |
| S = total number of species in a biotope; ni = number of individuals of species i, n = total number of individuals | ||
The density of the feet, average total height, average diameter, basal area, and regeneration rate were estimated as dendrometric parameters.
Table 2.
Different statistical parameters.
| Parameters | Formula | Source |
|---|---|---|
| Total density (D) of woody | D=n/s | [37,50] |
| Average diameter (Dm) | Dm = 1/n ∑di | [51] |
| Basal area (G | G = π/4s ∑0,0001d2 | [52] |
| Average heightRate of regeneration | Hm = 1/n ∑ hiTr = n/(n+N) x 100 | [51](Bowers, 1986) |
| n = number of plants per survey and S = survey area in hectare di= diameter at 1.30 m of tree i and hi= height of tree n= number of species at dbh < 10 cm; N = number of plants at dbh ≥ 10 cm. | ||
2.3.3. Zoning Plan
Knowing how the AFC land use changes over the time series, including its flora and ecosystem potentialities [53]. Thus, it is easier to suggest a management plan design map base zoning through buffer geoprocessing analysis. To design the zoning management map of AFC, the land use map of 2021 output associated with ecosystems assessment results was processed to define major management zones by taking into consideration both ecosystems met in AFC. In overall, under QGIS software [54], buffer design followed by asymmetric difference geoprocessing analysis was performed [55,56]. Finally, the key role or management vocation was defined for each designed zone [57,58].
3. Results
3.1. Spatial Characterization of Land Use
The Affem Boussou Community Forest (ACF) has 82,19 ha in size. In 2015, six (6) land cover categories (Figure 2) were found within this area: dense dry forest, gallery forest, plantations, tree savannah/shrub savannah, grassland savannah/meadow, and crop/fallow. These different vegetation formations have undergone significant changes between 2015 and 2018. The extent of dense dry forests has grown. This area, which was 35,89 ha or 43,66% of the total area, rose by 1.78% i.e., 36,53 ha. The same is true for tree savannahs/shrub savannahs, which expanded from 2,41 ha (2,93% of total area) in 2015 to 12.84 ha (15,62% of total forest area). This land use unit recorded an increase of 10.42 ha or 432.40% (Table 3). The area of gallery forests decreased by -0.76%, from 10.95 ha in 2015 to 10.86 ha in 2018, i.e., a loss of 0.08 ha. Crops/fallow land has dropped from 10.82 ha (13.17% of total) to 8.37 ha (-22.63% regression rate). The grassy savannahs/meadows decreased from 2.4 hectares in 2015 to 13.59 ha in 2018, a 36.41% decrease. In addition, 0.75 hectares of plantation acreage was converted to crops/fallow in 2015 (Figure 3).
Five (05) land cover types were identified in 2021 (Figure 4): dense dry forests, gallery forests, tree savannahs/shrub savannahs, grassy savannahs/meadows, and crops/fallow. Dry dense forest, gallery forest, and camp/fallow decreased in the area. The area of crops/fallow land decreased from 8.37 ha to 7.15 ha, a regression rate of -14.66%, while that of gallery forests decreased from 10.86 ha to 10.57 ha, a regression rate of -2.72%. The area of dry forests has reduced from 36.57 ha in 2018 (44.44% of the total area) to 33.05 ha in 2020, a regression rate of -9.53%. On the other hand, tree savannahs/shrubs and grassy savannahs/meadows have experienced a significant increase in their extent. Tree/shrub savannahs increased from 12.84 ha to 14.08 ha, an increase of 9.73%. The emergence of grassy savannas/grasslands is the most significant. They grew from 13.59 ha to 17.34 ha, i.e., a growth rate of 27.63% (Table 3).
In 6 years, the areas of crops/fallow, plantations, gallery forests, and dense dry forests, grassy savannahs/meadows have decreased. Crops/fallow have lost 3.68 ha of their area, a regression rate of -33.98%. Gallery forests regressed by -3.46% with a loss of 0.38 ha. The dry forests have declined by 2.84 ha, i.e., -7.92%. The plantations, present in 2015 were completely transformed into “other land use units. Grassland savannahs/grasslands lost 4.03 ha, a regression rate of -18.84%. The wooded savannahs/shrub savannahs have evolved with a progression rate of 4884.23% with a gain of 11.67 ha.
3.2. Floristic Assessment
The flora consists of 163 species divided into 129 genera and 55 families. Fabaceae (14.02%), Combretaceae (10.55%), Rubiaceae (9.13%), Sapotaceae (6.30%), and the other families (60%). (See Figure 5). Anogeissus leiocarpa (5.19%), Vitellaria paradoxa (4.72%), Daniella oliveri (3.15%), and Khaya senegalensis (3.15%) dominate the vegetation.
3.3. Plant Formation of ACF
Hierarchical ascending Classification were performed, using CANOCO (Canonical Community Ordination) software allowed to discriminate the 32 plots into five groups: Tree/shrub savannahs (G1), dense dry forests (G2), agrosystems (Crops/fallow) (G3), grassy savannahs/meadows (G4) and gallery forests (G5) (Figure 6)
3.3.1. Tree/Shrub Savannahs (G1)
Group G1 is composed of five (5) plots with 56 species classified in 51 genera and 33 families. Anogeissus leiocarpa (6.76%), Albizia zygia (4.05%), Cola gigantea (4.05%), Diospyros mombouttensis (4.05%), and Khaya senegalensis (4.05%) are the most common species. Fabaceae (16.22%), Combretaceae (9.46%), Malvaceae (8.11%), and Meliaceae (5.41%) are the most represented families. This group has a plant density of 74 plants/ha. The average diameter is 22.5 ± 11.14 cm, the average height is 16.13 ± 4.28 m, and the bole height is 8.39 ± 3.32 m, while the basal area is 2.98 m2/ha. Shannon’s diversity index is 3.90, and Piélou’s Equitability is 0.97 with a sandy-clay soil type. Regarding regeneration, group G1 had a density of 78 plants/ha, with 20 plants/ha of shoots, 54 plants/ha of seedlings, and 24 plants/ha of suckers.
The distribution of woody individuals by diameter class shows an “L” shape, with individuals in the 10-18 cm diameter class dominating (Figure 7A), implying that small-diameter individuals are dominant, which translates into the dominance of small-diameter individuals. On the other hand, the distribution of height classes indicates a “bell” structure. This structure is adjusted by the value of the Weibull shape coefficient, which is c = 2.135, reflecting a predominance of medium-height individuals (Figure 7B). Thus, the dominant individuals are between 7 and 17 m in height.
3.3.2. Dense Dry Forests (G2)
This group consists of five (5) plots, with 64 species divided into 52 genera and 29 families. Vitellaria paradoxa (7.95%), Anogeissus leiocarpa (6.82%), Cola gigantea (2.27%), and Khaya senegalensis (2.27%) are the most common species. However, Combretaceae (14.77%), Fabaceae (14.77%), Sapotaceae (10.23%), Rubiaceae (9.09%), and Malvaceae (4.55%) are the most common families. This group has a density of 136 plants/ha. The average diameter is 17.2 ± 5.42 cm, the average height is 16.37 ± 4.11 m, and the bole height is 3.73±2.35 m. The basal area is 1865 m2/ha. Shannon’s diversity index is 3.63 bits, and Piélou’s Equitability is 0.87 with a sandy-clay soil type. Group G2 has a regeneration density of 64 feet/ha due to 26 feet/ha of seedlings, 20 feet/ha of suckers, and 18 feet/ha of shoots.
The distribution of woody individuals by diameter class shows an “L” structure centered on a 10-14 cm diameter class (Figure 8A), indicating the dominance of individuals with small diameters. A bell-shaped design represents the height classes. A predominance of low and medium-height individuals characterizes this distribution. The Weibull coefficient “c” ranges from 1 to 3.6. Thus, the dominant individuals are between 6-10 m. (Figure 8B).
3.3.3. Crops and Fallows (Agrosystem) (G3)
The G3 group consists of 7 plots, with 94 species listed in 79 genera and 36 families. The most represented species are Anogeissus leiocarpa (5,89%), Vitellaria paradoxa (4.89%), Daniella oliveri (4.35%), Crossopteryx febrifuga (2.18%) et Borassus aethiopum (1.63%). The most represented families are Fabaceae (14.13%), Combretaceae (13.04%), Rubiaceae (9.78%), and Moraceae (5.98%). This group has a density of 36 plants/ha. The average diameter is 13±3.78 cm, the average total height is 12.35±4.48 m, and the bole height is 3.51±2.01 m. The basal area is 4.38 m2/ha. Shannon’s diversity index is 4.34, and Piélou’s Equitability is 0.96, with a clay-silt soil type. Regeneration is dominated by seedlings with a density of 23 feet/ha, followed by suckers with 18 feet/ha and shoots with 6 feet/ha.
The distribution of woody individuals by diameter class shows an “L” structure marked by a dominance of individuals in the 10-12 cm diameter class (Figure 9A), thus reflecting a dominance of small-diameter individuals. The representation of the height classes shows a bell-shaped structure. A predominance of low and medium-height individuals characterizes this distribution. The Weibull coefficient “c” ranges from 1 to 3.6. As a result, the dominating individuals range in height from 6 to 8 m (Figure 9B).
3.3.4. Grassy Savannahs and Meadows (G4)
Group G4 is composed of 7 records in which 83 species are listed in 75 genera and 35 families. The most represented species are Anogeissus leiocarpa (4.11%), Vitellaria paradoxa (4.11%), Ficuse sur forssk (3.42%), Parkia biglobosa (3.42%) Phoenix reclinata (2.74%). The most presented families are Fabaceae (12.33%), Combretaceae (10.96%), Rubiaceae (10.96%), and Asteraceae (6.85%). This group has a density of 36 plants/ha. The average diameter is 22.5 ± 17.18 cm, the average height is 9.62 ± 5.47 m, and the bole height is 5.16 ± 2.73 m. The basal area is 16.43 m2/ha. Shannon’s diversity index is 4.04 bits, and Piélou’s Equitability is 0.92 with a sandy-clay soil type. In terms of regeneration, group G4 has a density of 32 feet/ha due to 10 feet/ha of seedlings, 16 feet/ha of suckers, and 6 feet/ha of shoots.
The distribution of woody individuals by diameter class shows an “L” structure centered on a 10-16 cm diameter class (Figure 10A), thus representing a dominance of small-diameter individuals. The representation of height classes shows a bell-shaped structure. A predominance of low and medium-height individuals characterizes this distribution. The Weibull coefficient “c” ranges from 1 to 3.6. Thus, the dominant individuals are between 6-10 m. (Figure 10B).
3.3.5. Gallery Forests (G5)
The G5 group is composed of 8 plots in which 81 species are listed in 67 genera and 36 families. The most represented species are Daniella oliverie (4.90%), Vitellaria paradoxa (4.90%), Anogeissus leiocarpa (3.50%), Chassalia kolly (2.80%) Piliostigma thonningii (1.40). The families most presented are Fabaceae (14.69%), Combretaceae (9.10%), Rubiaceae (9.10%), and Sapotaceae (5.60%). This group has a density of 94 plants/ha. The average diameter is 30±15.30 cm, the average height is 14,84±5.16 m, and the bole height is 6.01±3.63 m. The basal area is 71.23 m2/ha. Shannon’s diversity index is 3.97 bits, and Piélou’s Equitability is 0.90 with a sandy-silty soil type. In terms of regeneration, group G5 has a density of 78 feet/ha due to 35 feet/ha of seedlings, 25 feet/ha of suckers, and 18 feet/ha of shoots.
The distribution of woody individuals by diameter class shows an “L” structure centered on a 10-14 cm diameter class (Figure 11A), thus representing a dominance of small-diameter individuals. The representation of height classes shows a bell-shaped structure. A predominance of low and medium-height individuals characterizes this distribution. Weibuul’s coefficient “c” is between 1 ≤ c ≤ 3.6. Thus, the dominant individuals are between 4-8 m. (Figure 12).
3.4. Regeneration
AFC has a regeneration potential of is 64 ft/ha due to 21 ft/ha of suckers, 29 ft/ha of seedlings, and 14 ft/ha of sprouts. Pavetta capensis (13.85%), Anogeissus leiocarpa (12.31%), Khaya senegalensis (10.76%), Vitellaria paradoxa (9.23%), and Daneilla oliveri (7.69%) have the highest regeneration density for suckers (Figure 13). For seedlings, there are Anogeissus leiocarpa s (19.57%), Lonchocarpus seruceus (6.52%), Acacia polyacantha (5.43%), Daniella oliveri (5.43%), Piliostigma thoningii (4.43%) and for the rejects we have Anogeissus leiocarpa (20.93) Lonchocarpus seruceus (13.95%), Khaya senegalensis (11.62%), Daneilla oliveri (4.65%) and Vitellaria paradoxa (4.65%).
3.5. Proposition of Zoning Plan
AFC is an area where many actors carry out several activities. This can be a source of mismanagement due to the open access status of the resources. Participatory management is necessary to preserve the interests of the various actors and to allow the forest to play its role. It is therefore required to fragment the space, i.e., to subdivide it into zones according to the nature of the activities and/or interventions to be carried out there. AFC management must meet three objectives (1) to conserve ecological functions and biological diversity; (2) to contribute to the sustainable supply of agro-sylva-pastoral products for AFC’s residents; (3) to ensure the protection and conservation of natural resources. The overall objective is to contribute to the development of community forestry, By providing the provision of products and services to meet the needs and expectations of the population and the authorities in terms of social, economic, and environmental well-being.
The division into series and management plan of ACF
A serie, by definition, is a unit of objective and treatment. As a result, it clusters together areas and portions that will receive the same treatment. The AFC is divided into four series to meet the management plan’s objectives:
- The agroforestry series (series 1)
The agroforestry series consists of crops, fallow land, and pasture. The series covers 15.45 ha or 18.80% of the total area of AFC. It is primarily made up of initially forested regions that have been destroyed and cultivated in the past. Agro-sylva-pastoral activities will be carried out, an association of perennial crops with forest plants and animals (pastures). Reforestation activities will be carried out with agroforestry species such as Samanea samane, Senna siamea, and Albizia lebbeck.
Table 5.
Management Series and vocation use.
| Series (zone) | Area (ha) | Primary use | Secondary use |
|---|---|---|---|
| Agroforestry vocation | 15.45 | Agricultural and forage product | Soil fertilization |
| Sustainable forestry production | 34.70 | Energy wood, timber, non-timber forest products | Environment protection |
| Buffer | 9.03 | Separation of the protection and production area | |
| Biological Conservation | 23.01 | Conservation of biodiversity and genetic resources | Tourism, carbon sequestration, and recreation |
- The forest series of sustainable production (series 2)
It accounts 42.22% of the total area of the forest or 34.70 ha. The riparian communities will be authorized to collect non-wood products and some stem in a customary and non-destructive manner. This should allow for controlled harvesting, the preservation of the area, and the conservation of biological diversity. The absence of penetration road building and industrial exploitation are assets in achieving these objectives. The tagging of the trees becomes compulsory for the felling and thus allows more precise and more reasoned forest utilization. The production of timber and the reconstitution of the floristic diversity justify the implementation of this series. Full plantations are reserved for young and very degraded fallows. The main species used are Anogeissus leiocarpa, Khaya spp, Daneilla, and Terminalia spp. Other methods adapted to the state of the stands can be used, such as enrichment in layers (with the following species: Terminalia spp, Afzelia spp, Pterocarpus, Khaya spp), and planting in densely spaced plots, as they allow the conservation of significant biodiversity by maintaining the inter-band floristic composition. To ensure the success of these enhancements, maintenance must be carried out over time, which entails significant recurrent costs. Planting under cover might also contribute to the diversification of this composition.
- The buffer series (series 3)
The buffer zone is an area that separates the sustainable production zone from the biological conservation zone. It covers an area of 9.03 ha or 11% of the total area of AFC. Some human activities are allowed in this area, and we try to limit their impacts.
- The biological conservation series (series 4)
This series represents 23.01 ha or 28% of the total area of AFC. The value of this series in terms of biological diversity justifies its conservation in its current state, and it covers areas sensitive to significant erosion processes. Industrial operators are excluded from any logging activity in this area. They serve as a vital part of the ecological reservoir’s protection zone; hence entry is restricted.
4. Discussion
The spatiotemporal dynamics of AFC vegetation were assessed, and five major land-use units were identified: dense dry forest, crops and fallow, gallery forest, shrub/shrub savannah, and grassland savannah/meadows. Plantations are only observed in 2015 due to protection measures initiated by the village community. The study of [59] identified the same number of land use units, by national and international classification systems. An anthropization of vegetation formations is observed over the years. These phenomena are mainly due to human action as reported by [60,61] in a study on village green belts in the prefecture of Avé in Togo. Several studies have confirmed the socio-economic dependence of riparian populations on forest massifs [62,63,64,65]. The steady diminution of farmed lands is beginning to bear fruit as a result of riparian people’s awareness of the need of plant resource protection and sustainable management. Crops/fallow have regressed considerably, losing 14.66% of their surface area between 2018 and 2021. However, savannah formations have increased, with increases of 27.63% and 9.73% for grassland savannahs/meadows and tree/shrub savannahs, respectively. These findings contradict those of [66] in a study on changes in vegetation cover in the watershed of Zio in southwestern Togo by remote sensing, where regression of grassy savannahs to the profile of crops/ fallows and built-up areas/bare ground is recorded. This is due mainly to the extension of the habitat in the Zio watershed, which is primarily linked to population growth and peri-urbanization [66]. In the context of declining and unevenly distributed rainfall recorded in recent years, [67,68] the watershed is also suitable for agricultural activities. On the other hand, the increase in ruminant livestock in the central region, which has resulted in an increasing need for grazing lands, could explain the observed regression of cultivated areas in favor of savanna formations in the AFC [69] notes the same decrease in cultivated areas in northern Senegal.
The AFC’s floristic analysis reveals significant potential for woody and non-woody species. There are 163 species, which are classified 129 genera and 55 families. This diversity is more important than that found by [70] in the Abdoulaye reserve and by [71] in the Kaodji community forest in Benin, with 69 and 83 woody species, respectively. The variance is attributed not only to differences in sample methodology and sampling region but also to anthropogenic forces that cause the degradation of the various formations. The species richness varies from one group to another. It is more important in the crops and fallow lands (94 species) and low in the tree/shrub savannahs (56 species). These results are consistent with those of [71] where the lowest diversity is observed in the savannas. The five identified groups all have an L-shaped distribution of diameter classes. From an ecological point of view, AFC would have all the assets for its reconstitution of the high density of juvenile individuals according to the conclusions of [72,73] in a study within the community forest of Edouwossi-Copé in southern Togo. It is in this dynamic that this forest benefits from the program of restoration of forest landscapes and good governance initiated by the GIZ in the framework of the mechanism forests and farmers “Forest For Future” (F4F), which aims to contribute to the fight against climate change through the restoration and management of forests. The non-negligible regeneration rate (64 feet/ha) is much lower than the 1713 feet/ha regeneration rate of the Agbandi community forest in central Togo [74]. This large discrepancy could be explained by the dominance of open forests in the Agbandi community forest.
The requirement to reconcile sustainable production, enhanced community revenues, and excellent ecosystem services would lend validity to the AFC zoning plan [75]. The zoning of forest domains has often been considered the result of a process of capitalization of the results of diagnosing the state of the resource encountered in the forests. In addition to being forward-thinking, these plans would be part of the programmatic processes for formalizing community forests. The optimal use of the study’s findings can lead to preliminary steps toward orienting future AFC actions toward a sustainable management mechanism consistent with similar activities undertaken in the region.
5. Conclusion
The study of the spatiotemporal dynamics of land use in the AFC revealed a regressive trend of formations. The floristic inventory allowed a list of 163 species divided into 129 genera and 55 families. Fabaceae, Combretaceae, Rubiaceae, and Sapotaceae are the most represented families. Anogeissus leiocarpa, Vitellaria paradoxa, Daniella oliveri, and Khaya senegalensis are the most represented species. The predominance of young individuals is noted in all formations of the community forest. The regeneration density of the AFC is 64 feet/ha due to 21 feet/ha of suckers, 29 feet/ha of seedlings, and 14 feet/ha of shoots. Considering these results, it is important to strengthen management measures to conserve the biodiversity of ACF. The zoning of forest domains has often been considered the result of a process of capitalizing on the results of the diagnosis on the state of forest conservation.
Author Contributions
F.F. has conceptualized, analyzed data and wrote the first draft, D.B. W.A. H.P and I.P.N. contributed to the manuscript editing. All authors have read and validated the final manuscript.
Funding
This study was financed by Forest Farms Facility (FFF) of FAO (Togo).
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
The Authors thank Forest Farms Facility (FFF) of FAO and the Laboratory of Botany and Plant Ecology (LBEV) of University of Lomé (Togo) for their unconditional support.
Conflicts of Interest
Authors declare that there is no competing interest.
References
- Akognongbe, A.; Abdoulaye, D.; Vissin, E.W.; Boko, M.J.A.S.R.I.d.S.e.T. Dynamique de l’occupation du sol dans le bassin versant de l’Oueme à l’exutoire de Bétérou (Bénin). 2014, 10.
- Hlovor, A.K.D.; Adjonou, K.; Dangbo, F.A.; Abotsi, K.E.; Afelu, B.; Kokou, K. Dynamique du couvert forestier dans la partie méridionale des monts Togo, Afrique de l’ouest.
- Mama, A.; Sinsin, B.; De Cannière, C.; Bogaert, J.J.T. Anthropisation et dynamique des paysages en zone soudanienne au nord du Bénin. 2013, 31.
- Dourma, M.; Batawila, K.; Wala, K.; Kokou, K.; Guelly, K.A.; Bellefontaine, R.; De Foucault, B.; Akpagana, K. Régénération naturelle des peuplements à Isoberlinia spp. en zone soudanienne au Togo. Acta botanica gallica 2009, 156, 415–425. [Google Scholar] [CrossRef]
- FAO. La situation des forêts du monde. Les forêts au service du développement durable. Licence: CC BY-NC-SA 3.0 IGO: 158p. 2018.
- Bergonzini, J.-C. Changements climatiques, désertification, diversité biologique, et forêts; SILVA: 2004.
- Folega, F.; Pereki, H.; Woegan, A.; Dourma, M.; Atakpama, W.; Maza, M.; Akpagana, K.J.J.R.S.U.L. Caractérisation écologique de la Forêt Communautaire d’Edouwossi-Cope (Région des Plateaux-Togo). 2017, 19, 47-61.
- Lykke, A.M.J.J.o.E.M. Local perceptions of vegetation change and priorities for conservation of woody-savanna vegetation in Senegal. 2000, 59, 107-120. [CrossRef]
- Tente, B.; Sinsin, B.J.É.s.l.F.e.l.V.d.B.F. Diversité et structure des formations arborescentes du secteur Perma-Toucountouna dans la chaîne de l’Atacora (Bénin). 2002, 6, 31-42.
- UICN/PACO. Evaluation de l’efficacité de la gestion des aires protégées : aires protégées du Togo. 2008.
- Badjare, B.; Woegan, Y.; Folega, F.; Atakpama, W.; Wala, K.; Akpagana, K. Vulnérabilité des ressources ligneuses en lien avec les différentes formes d’usages au Togo: Cas du paysage des aires protégées Doungh-fosse aux lions (Région des Savanes). Revue Agrobiologia 2021, 11, 2552–2565. [Google Scholar]
- Hien, P.; Bac, V.; Tham, H.; Nhan, D.; Vinh, L.J.A.E. Influence of meteorological conditions on PM2. 5 and PM2. 5− 10 concentrations during the monsoon season in Hanoi, Vietnam. 2002, 36, 3473–3484. [Google Scholar]
- Grégoire, J.-M.; Simonetti, D.J.R.S. Interannual changes of fire activity in the protected areas of the SUN network and other parks and reserves of the West and Central Africa region derived from MODIS observations. 2010, 2, 446-463. [CrossRef]
- Nguinguiri, J.-C. Les approches participatives dans la gestion des écosystèmes forestiers d’Afrique centrale: revue des initiatives existantes; CIFOR: 1999.
- Morou, B.; Lawali, S.; Oumani, A.A.; Ounani, H.; Guero, C.; Mahamane, A.J.A.S. Ressources forestières ligneuses: diversité et usages dans le terroir villageois de Dan Saga. 2016, 12, 228-239.
- Koumoi, Z.; Alassane, A.; Djangbedja, M.; Boukpessi, T.; Kouya, A.-E. Dynamique spatio-temporelle de l’occupation du sol dans le Centre-Togo. AHOHO-Revue de Géographie du LARDYMES 2013, 7, 163–172. [Google Scholar]
- Kaina, A.; Dourma, M.; Folega, F.; Diwediga, B.; Kpérkouma, W.; Akpagana, K. Localisation des bassins de production de bois énergie et typologie des acteurs de la filière dans la région centrale du Togo. 2021.
- Ern, H. Die Vegetation Togos. Gliederung, Gefährdung, Erhaltung. Willdenowia 1979, 295-312.
- Koumoi, Z.; Boukpessi, T.; Kpedenou, K.D. Principaux facteurs explicatifs de la dynamique de l’occupation du sol dans le Centre-Togo: apport des SIG et des statistiques spatiales. Revue Ivoirienne de Géographie des Savanes (RIGES) 2017, 252-273.
- Banito, A.; Kpemoua, E.K.; Dayiwo, R.K.; Tedihou, E.; Sikirou, R. Inventaire des maladies de l’anacardier (Anacardium occidentale L.) dans la préfecture de Tchamba au Togo. International Journal of Biological and Chemical Sciences 2021, 15, 2514–2525. [Google Scholar] [CrossRef]
- METEO-TOGO. Données de la Météo, Région centrale. https://www.meteocity.com 2020.
- Adougnon, K. Togo: le recensement général de la population du Togo de 1970. 1980.
- Roudié, P. Aspects du développement récent de l’économie togolaise. Les Cahiers d’Outre-Mer 1978, 31, 359–374. [Google Scholar] [CrossRef]
- Rand, J.T.; Kramer, L.A.; Garrity, C.P.; Hoen, B.D.; Diffendorfer, J.E.; Hunt, H.E.; Spears, M. A continuously updated, geospatially rectified database of utility-scale wind turbines in the United States. Scientific data 2020, 7, 15. [Google Scholar] [CrossRef]
- Kolarić, S.; Shelden, D. Toward an Open IoT Implementation for Urban Environments: The Architecture of the DBL SmartCity Platform. Smart Cities and Smart Governance: Towards the 22nd Century Sustainable City 2021, 345-372.
- Blaschke, T.; Burnett, C.; Pekkarinen, A. Image segmentation methods for object-based analysis and classification. Remote sensing image analysis: Including the spatial domain 2004, 211-236.
- Parivallal, R.; Nagarajan, B. Supervised Classification Methods For Object Identification Using Google Map Image. International J. Eng. Sci. Manag. Res 2014, 1, 71. [Google Scholar]
- Di Gregorio, A. Land cover classification system: classification concepts and user manual: LCCS; Food & Agriculture Org.: 2005; Volume 2.
- Li, X.; Shen, H.; Li, H.; Zhang, L. Patch matching-based multitemporal group sparse representation for the missing information reconstruction of remote-sensing images. IEEE Journal of Selected Topics in Applied Earth Observations and Remote Sensing 2016, 9, 3629–3641. [Google Scholar] [CrossRef]
- Schmitt, M.; Zhu, X.X. Data fusion and remote sensing: An ever-growing relationship. IEEE Geoscience and Remote Sensing Magazine 2016, 4, 6–23. [Google Scholar] [CrossRef]
- Yang, L.; Xian, G.; Klaver, J.M.; Deal, B. Urban land-cover change detection through sub-pixel imperviousness mapping using remotely sensed data. Photogrammetric Engineering & Remote Sensing 2003, 69, 1003–1010. [Google Scholar] [CrossRef]
- Stefanov, W.L.; Ramsey, M.S.; Christensen, P.R. Monitoring urban land cover change: An expert system approach to land cover classification of semiarid to arid urban centers. Remote sensing of Environment 2001, 77, 173–185. [Google Scholar] [CrossRef]
- Ahlqvist, O. Extending post-classification change detection using semantic similarity metrics to overcome class heterogeneity: A study of 1992 and 2001 US National Land Cover Database changes. Remote Sensing of Environment 2008, 112, 1226–1241. [Google Scholar] [CrossRef]
- Arouna, O.; Etene, C.G.; Issiako, D. Dynamique de l’occupation des terres et état de la flore et de la végétation dans le bassin supérieur de l’Alibori au Benin. Journal of Applied Biosciences 2016, 108, 10543–10552. [Google Scholar] [CrossRef]
- Ouedraogo, O.; Thiombiano, A.; Hahn-Hadjali, K.; Guinko, S. Diversité et structure des groupements ligneux du parc national d’Arly (Est du Burkina Faso). Flora et Vegetatio Sudano-Sambesica 2008, 11, 5–16. [Google Scholar] [CrossRef]
- Aboubacar, K.; Douma, S.; Mamoudou, B.M.; Seyni, R.S.D. Structure des populations naturelles de Neocarya macrophylla (Sabine) Prance, ligneux d’intérêt alimentaire, dans le Dallol Bosso, Niger. BOIS & FORETS DES TROPIQUES 2018, 337, 67–78. [Google Scholar] [CrossRef]
- Folega, F.; Zhang, C.; Woegan, Y.; Wala, K.; Dourma, M.; Batawila, K.; Seburanga, J.; Zhao, X.; Akpagana, K. Structure and ecology of forest plant community in Togo. Journal of Tropical Forest Science 2014, 225–239. [Google Scholar]
- FOLEGA, F.; BIMARE, K.; KONATE, D.; KPERKOUMA, K.M.W.; KOFFI, A. Inventaire et séquestration de carbone de la végétation de l’emprise urbaine de la ville de Dapaong, Togo. Espace Géographique et Société Marocaine 2020.
- Pereki, H.; Wala, K.; Thiel-Clemen, T.; Bessike, M.P.B.; Zida, M.; Dourma, M.; Batawila, K.; Akpagana, K. Woody species diversity and important value indices in dense dry forests in Abdoulaye Wildlife Reserve (Togo, West Africa). International Journal of Biodiversity and Conservation 2013, 5, 358–366. [Google Scholar]
- Sokpon, N.; Biaou, S.H.; Ouinsavi, C.; Hunhyet, O. Bases techniques pour une gestion durable des forêts claires du Nord-Bénin: rotation, diamètre minimal d’exploitabilité et régénération. BOIS & FORETS DES TROPIQUES 2006, 287, 45–57. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical ecology; Elsevier: 2012.
- Kim, J.-H.; Park, J.-M.; Jung, K.-S.; Ri, C.-U. DECORANA and TWINSPAN Aided Analysis of Koelreuteria paniculata Community Formation. Korean Journal of Environment and Ecology 2005, 19, 9–18. [Google Scholar]
- Hill, M. Decorana. A FORTRAN program for detrended correspondence analysis and reciprocal averaging 1979.
- raak, T. CANOCO reference manual and CanoDraw for Windows user’s guide: software for canonical community ordination,(version 45). (No Title) 2002, 500 pp.
- Ter Braak, C.t.; Smilauer, P. CANOCO reference manual and user’s guide to Canoco for Windows: software for canonical community ordination (version 4). 1998.
- Raunkiaer, C. The life forms of plants and statistical plant geography; being the collected papers of C. Raunkiaer. The life forms of plants and statistical plant geography; being the collected papers of C. Raunkiaer. 1934.
- White, F. La végétation de l’Afrique: mémoire accompagnant la carte de végétation de l’Afrique Unesco/AETFAT/UNSO; IRD Editions: 1986; Volume 20.
- Magurran, A.E. Diversity indices and species abundance models. In Ecological diversity and its measurement; Springer: 1988; pp. 7-45.
- Chao, A.; Shen, T.-J. Nonparametric estimation of Shannon’s index of diversity when there are unseen species in sample. Environmental and ecological statistics 2003, 10, 429–443. [Google Scholar] [CrossRef]
- Sickle, J.V.; Gregory, S.V. Modeling inputs of large woody debris to streams from falling trees. Canadian Journal of Forest Research 1990, 20, 1593–1601. [Google Scholar] [CrossRef]
- Bégin, J.; Raulier, F. Comparaison de différentes approches, modèles et tailles d’échantillons pour l’établissement de relations hauteur-diamètre locales. Canadian Journal of Forest Research 1995, 25, 1303–1312. [Google Scholar] [CrossRef]
- Décourt, N. Utilisation de la photographie pour mesurer les surfaces terrières. Revue forestière française 1956, 505-507. [CrossRef]
- Folega, F.; Atakpama, W.; Wala, K.; Mukete, B.; Shozo, S.; Akira, O.; Zhao, X.-h.; Akpagana, K. Land use patterns and tree species diversity in the Volta Geological Unit, Togo. Journal of Mountain Science 2019, 16, 1869–1882. [Google Scholar] [CrossRef]
- Team, Q.G.D. Quantum GIS Geographic Information System 2009. 2021.
- Eiselt, H.; Marianov, V.; Bhadury, J. Mathematical and Geospatial Tools. In Multicriteria Location Analysis; Springer: 2023; pp. 107-142.
- Rial-Berriel, C.; Acosta-Dacal, A.; Pérez, M.Á.C.; Suárez-Pérez, A.; Melián, A.M.; Zumbado, M.; Hernández, L.A.H.; Ruiz-Suárez, N.; Hernández, Á.R.; Boada, L.D. Intensive livestock farming as a major determinant of the exposure to anticoagulant rodenticides in raptors of the Canary Islands (Spain). Science of the Total Environment 2021, 768, 144386. [Google Scholar] [CrossRef]
- Nurrochmat, D.R.; Pribadi, R.; Siregar, H.; Justianto, A.; Park, M.S. Transformation of agro-forest management policy under the dynamic circumstances of a two-decade regional autonomy in Indonesia. Forests 2021, 12, 419. [Google Scholar] [CrossRef]
- Hajjar, R.; Oldekop, J.A.; Cronkleton, P.; Newton, P.; Russell, A.J.; Zhou, W. A global analysis of the social and environmental outcomes of community forests. Nature Sustainability 2021, 4, 216–224. [Google Scholar] [CrossRef]
- Mensah, K.; Folega, F.; Woegan, Y.A.; Atakpama, W.; Pereki, H.; Wala, K.; Akpagana, K. État et dynamique spatio-temporelle de la forêt communautaire d’EdouwossiCopé, Région des Plateaux-Togo. Rev Écosystèmes et Paysages (Togo) 2022, Vol 2, 12-26 pp.
- Bigma, B.; Woegan, Y.A.; Bawa, A.; Diwediga, B.; Koumantiga, D.; Wala, K.; Akpagana, K. Ceintures vertes villageoises de la préfecture de l’Avé au Togo (Afrique de l’ouest) : diversité floristique, menaces et modes de gestion. Rev Écosystèmes et Paysages (Togo), 2022, 01, vol 02.
- Fousseni, F.; Marra, D.; Wala, K.; Batawila, K.; Zhang, C.-y.; Zhao, X.-h.; Akpagana, K. Assessment and impact of anthropogenic disturbances in protected areas of northern Togo. For. Stud. China 2012, 14, 216–223. [Google Scholar] [CrossRef]
- Issifou, A.; Folega, F.; Kombate, B.; Atakpama, W.; BATAWILA, K.; Ketoh, G.K.; Akpagana, K. Cartographie participative des terroirs riverains de la réserve de faune d’Abdoulaye au Tog. Rev Écosystèmes et Paysages (Togo), 2022, N°01, vol 02.
- Zanh, G.G.; Barima, Y.S.S.; Kouakou, K.A.; Sangne, Y.C. Usages des produits forestiers non-ligneux selon les communautés riveraines de la forêt classée du Haut-Sassandra (Centre-Ouest de la Côte d’Ivoire). Int. J. Pure App. Biosci 2016, 4, 212–225. [Google Scholar]
- Boulogne, M. Vulnérabilité des paysages forestiers dans le parc de Ranomafana (Madagascar): dynamiques environnementales et trajectoires agroforestières; theses.fr: 2016.
- Ouattara, B.; Sanou, L.; Koala, J.; Hien, M. Utilisations locales et vulnérabilité des espèces ligneuses dans les forêts classées de Oualou et de Tissé au Burkina Faso, Afrique de l’Ouest; afriquescience.net: 2021.
- Seou, E.; Takou, P.W.; Boukpessi, T. Analyse des changements par télédétection de la couverture végétale du bassin de Zio (Sud-Ouest Togo). Rev Écosystèmes et Paysages (Togo) 2022, Vol 2, 126-139pp. 2.
- Abalo, M.; Badabate, D.; Fousseni, F.; Kpérkouma, W.; Koffi, A. Landscape-based Analysis of wetlands patterns in the Ogou River basin in Togo (West Africa). Environmental Challenges 2021, 2, 100013. [Google Scholar] [CrossRef]
- Folega, F.; Kanda, M.; Fandjinou, K.; Bohnett, E.; Wala, K.; Batawila, K.; Akpagana, K. Flora and Typology of Wetlands of Haho River Watershed, Togo. Sustainability 2023, 15, 2814. [Google Scholar] [CrossRef]
- Sarr, M.A. Cartographie des changements de l’occupation du sol entre 1990 et 2002 dans le nord du Sénégal (Ferlo) à partir des images Landsat. Cybergeo: European Journal of Geography 2009.
- Adjonou, K.; Djiwa, O.; Kombate, Y.; Kokutse, A.D.; Kokou, K. Etude de la dynamique spatiale et structure des forêts denses sèches reliques du Togo: implications pour une gestion durable des aires protégées. International Journal of Biological and Chemical Sciences 2010, 4. [Google Scholar] [CrossRef]
- Djego, J.; Gibigaye, M.; Tente, B.; Sinsin, B. Analyses écologique et structurale de la forêt communautaire de Kaodji au Bénin. International Journal of Biological and Chemical Sciences 2012, 6, 705–713. [Google Scholar] [CrossRef]
- Folega, F.; Pereki, H.; Woegan, A.Y.; Dourma, M.; Atakpama, W.; Maza, M.S.; Akpagana, K. Caractérisation écologique de la Forêt Communautaire d’Edouwossi-Cope (Région des Plateaux-Togo). J. Rech. Sci. Univ. Lomé (Togo) 2017, 19, 47–61. [Google Scholar]
- Folega, F.; Atakpama, W.; Pereki, H.; Djiwa, O.; Dourma, M.; Kombate, B.; Abreni, K.; Wala, K.; Akpagana, K. Potentialites ecologiques et socio-economiques de la foret communautaire d’agbedougbe (region des Plateaux-Togo). Journal de la Recherche Scientifique de l’Université de Lomé 2017, 19, 31–49. [Google Scholar]
- Bawa, D.M.-e.; Folega, F.; Atato, A.; Diwediga, B.; Wala, K.; Akpagana, K. Caractéristiques floristiques et structurales de la forêt communautaire d’Agbandi au centre du Togo (Afrique de l’ouest). Rev Écosystèmes et Paysages (Togo) 2022, Vol 2, 55-74 pp. 2.
- Folega, F.; Datche-Danha, K.E.; akim FOLEGA, A.; Woegan, A.Y.; Kperkouma, W.; Akpagana, K. Diversité des services écosystémiques et utilisation des terres dans le paysage du socle Eburnéen au Togo: Diversity of ecosystem services and land use in the Eburnean basement landscape of Togo. Revue Nature et Technologie 2022, 14, 61–75. [Google Scholar]
Figure 1.
Location of Affem Community Forest.
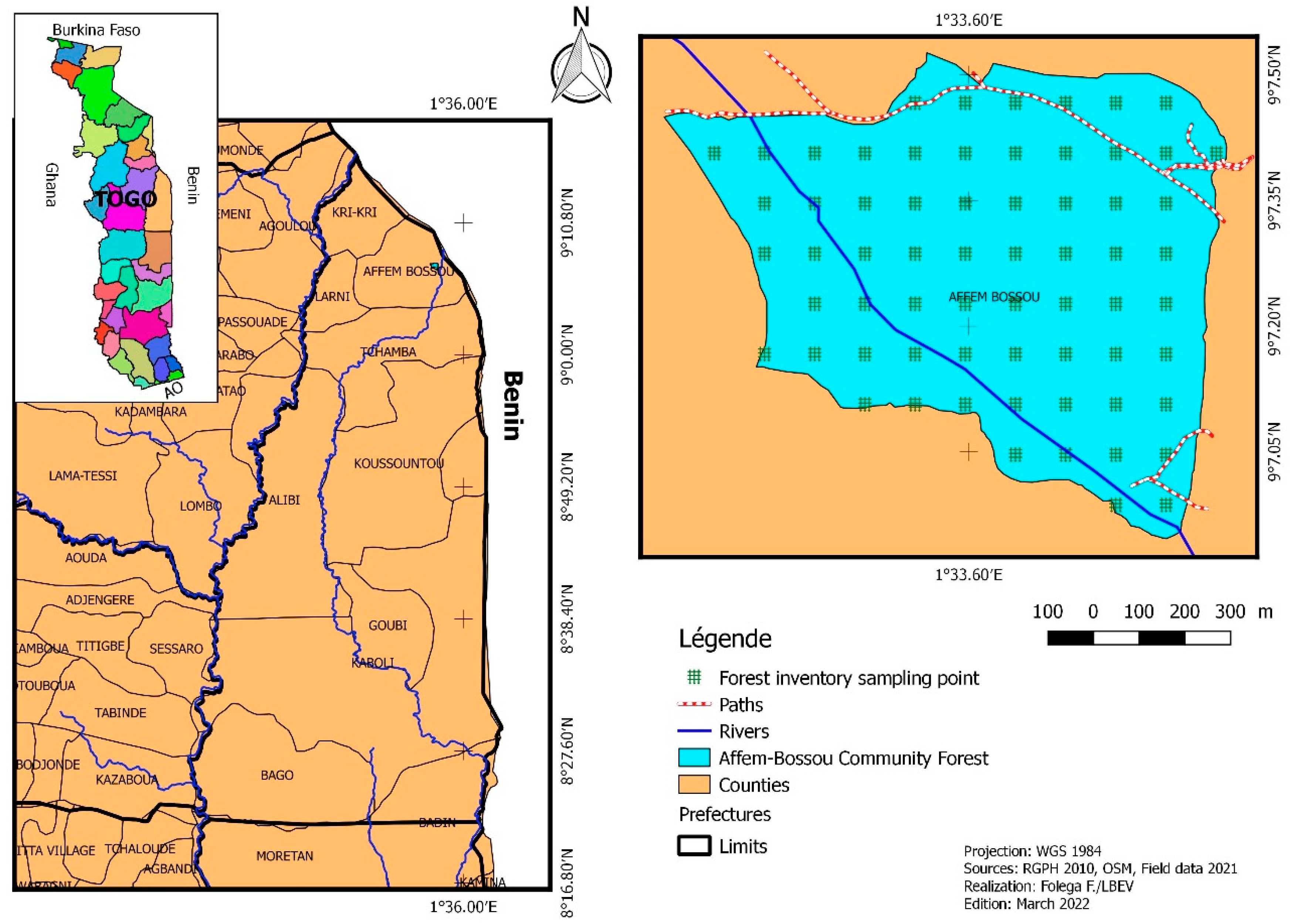
Figure 2.
Land use in 2015.
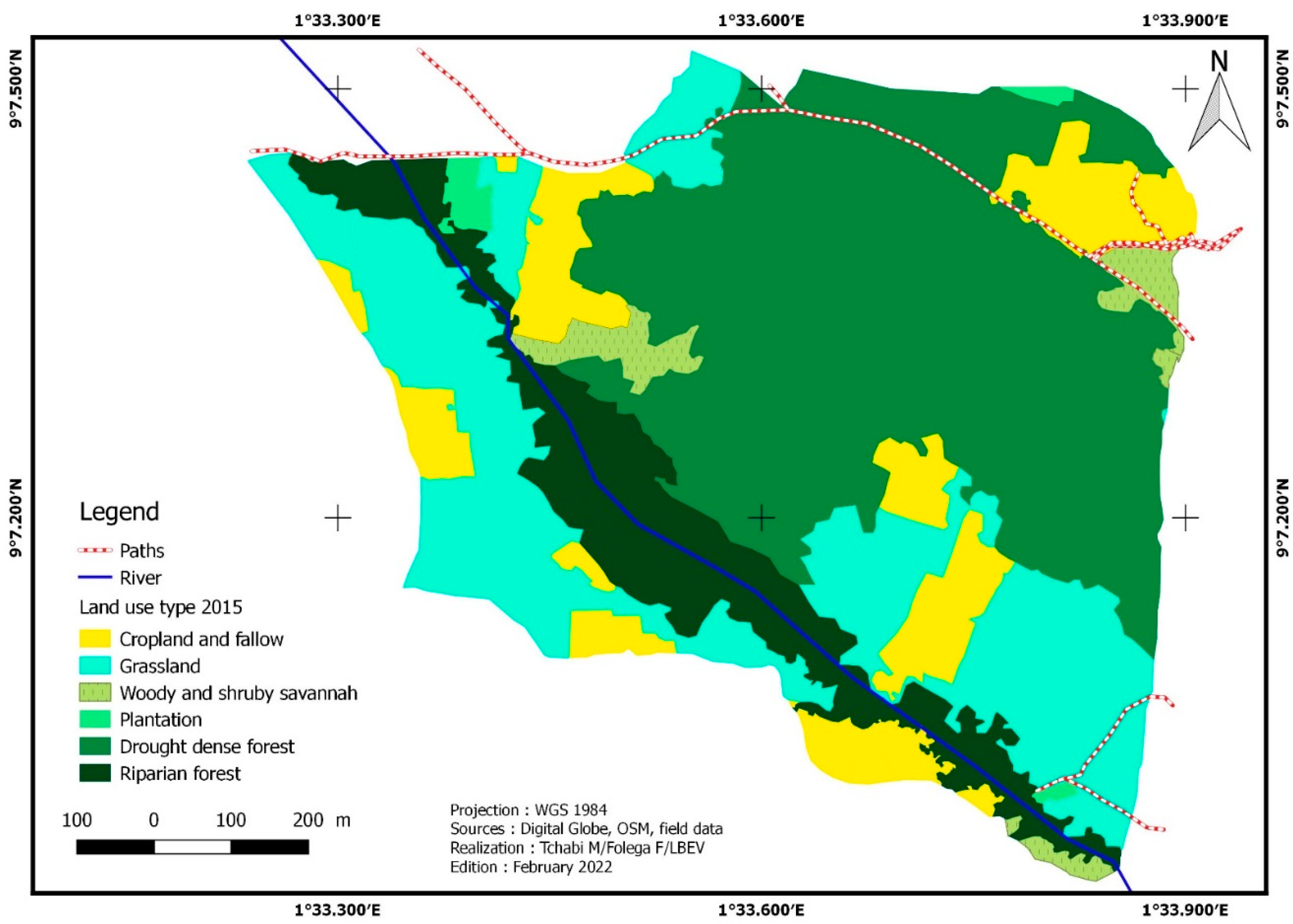
Figure 3.
Land use in 2018.
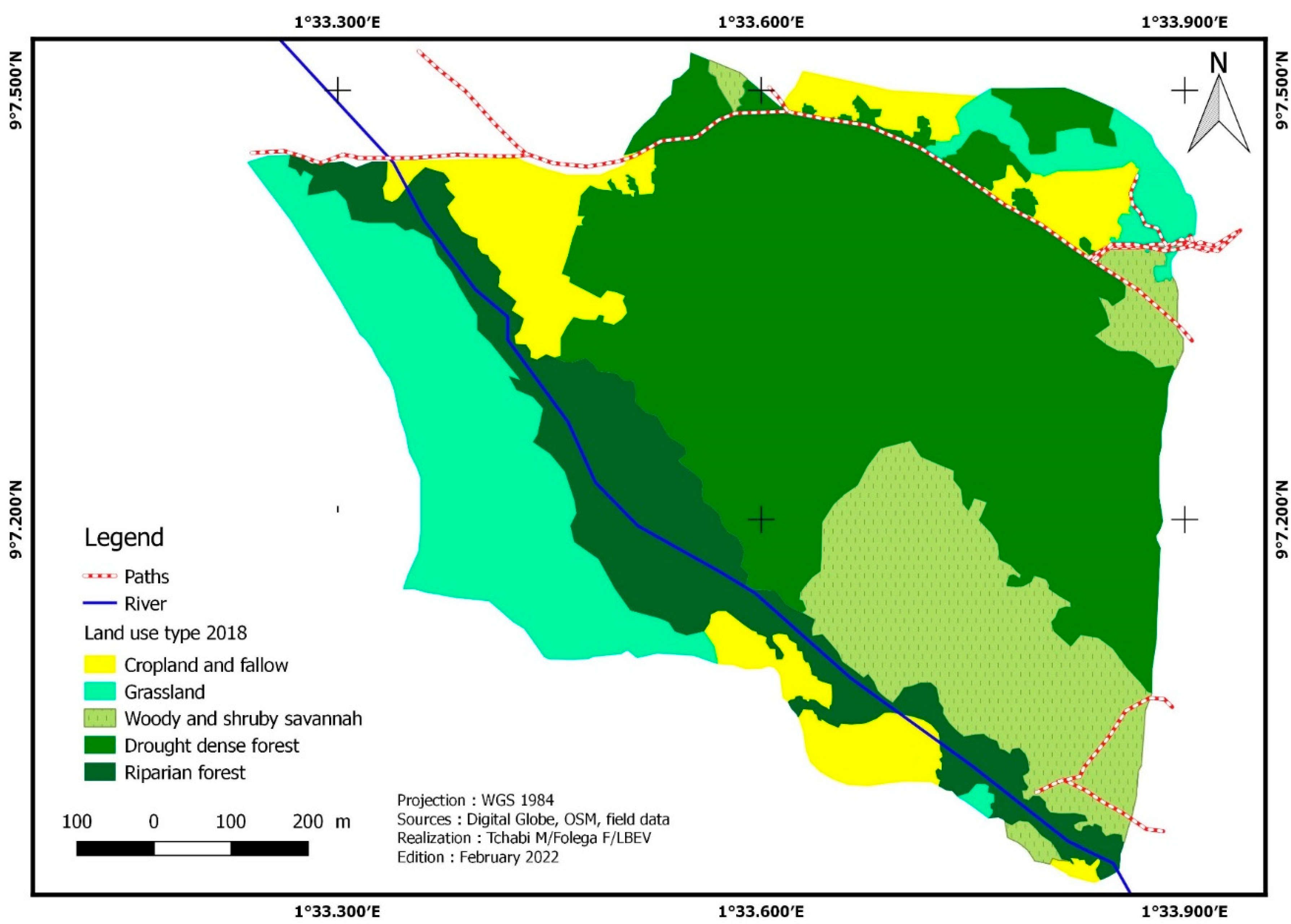
Figure 4.
Land use in 2021.
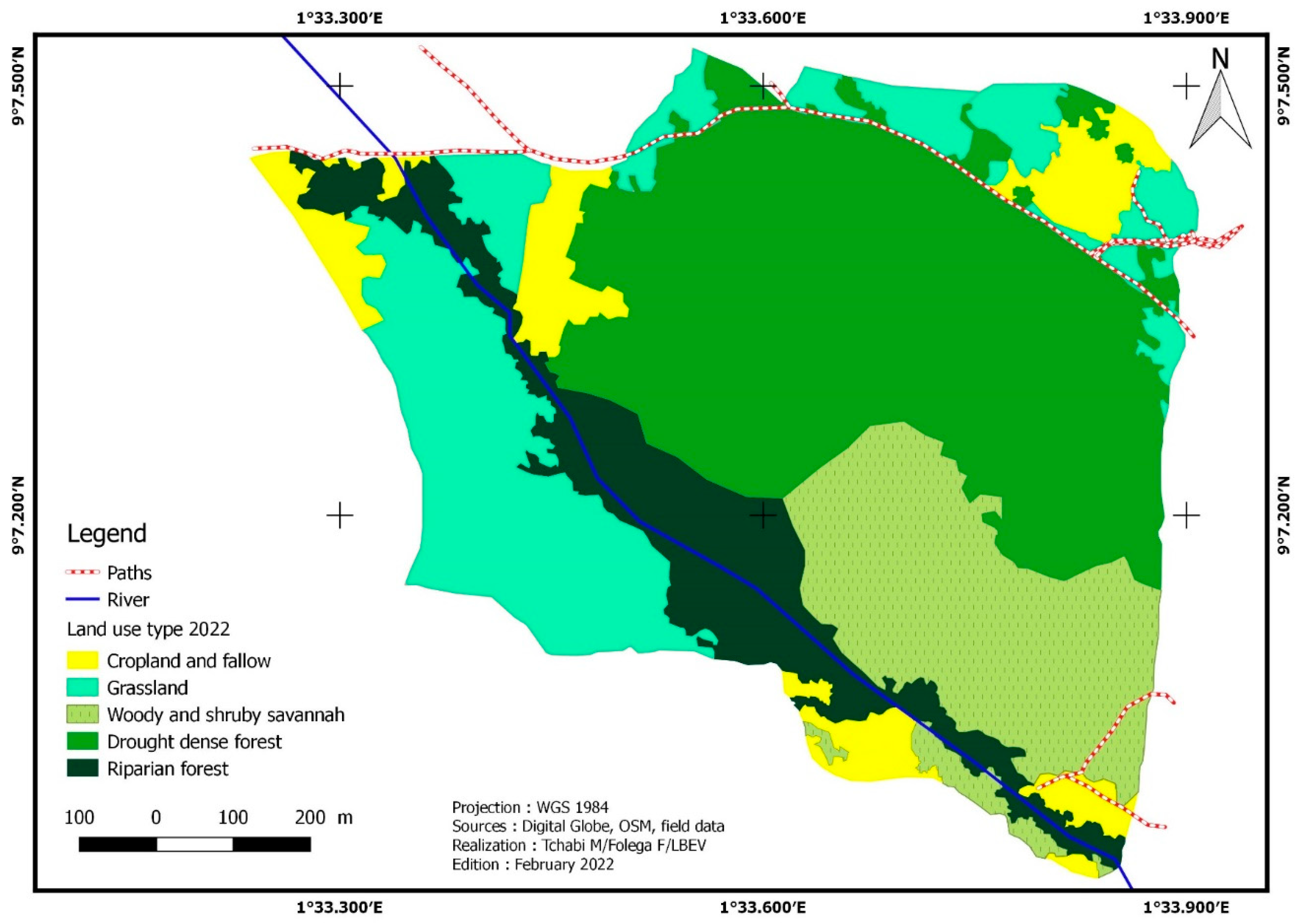
Figure 5.
Raw spectrum of Affem forest families.
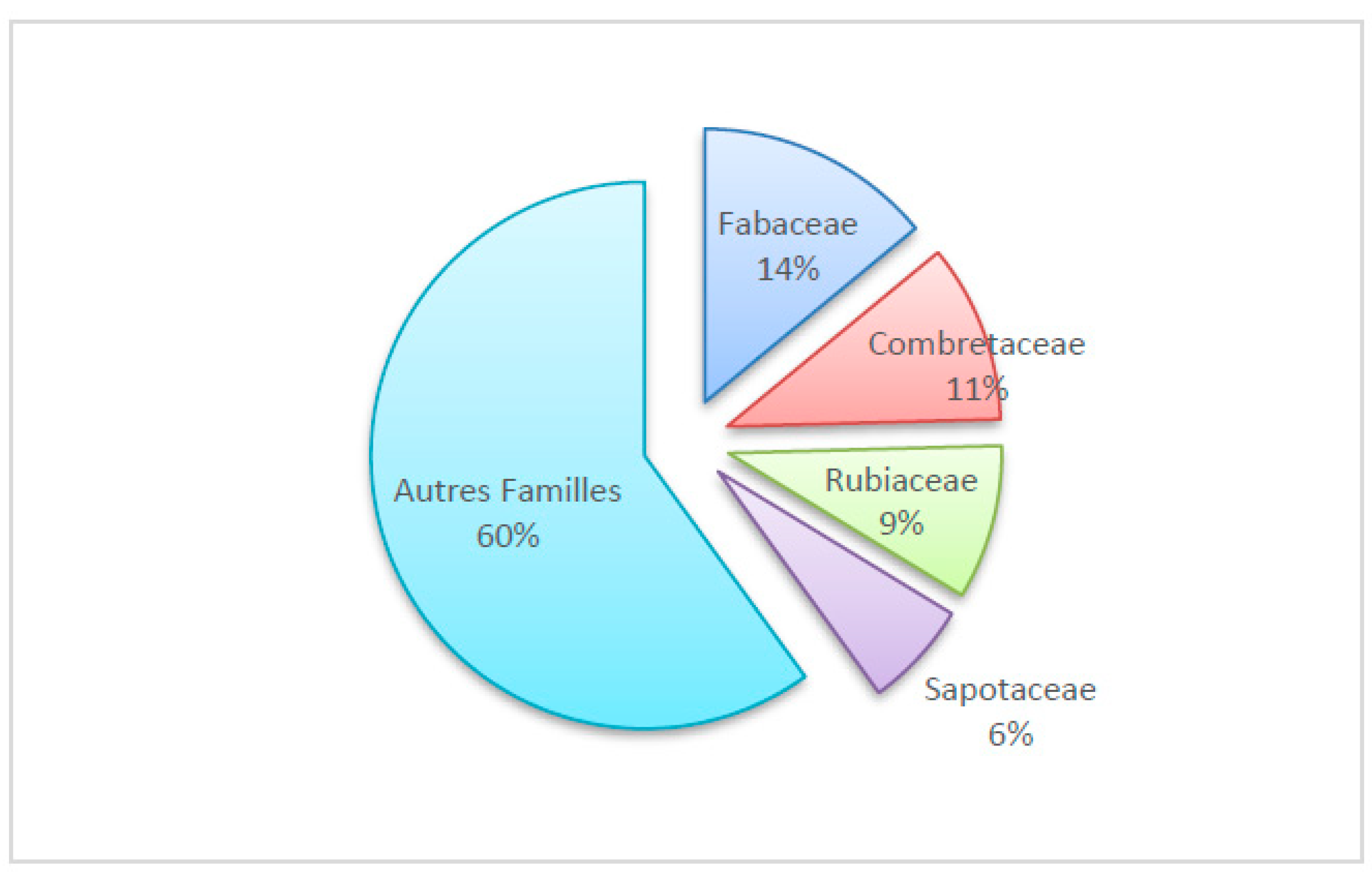
Figure 6.
Hierarchical ascending classification of AFC records.
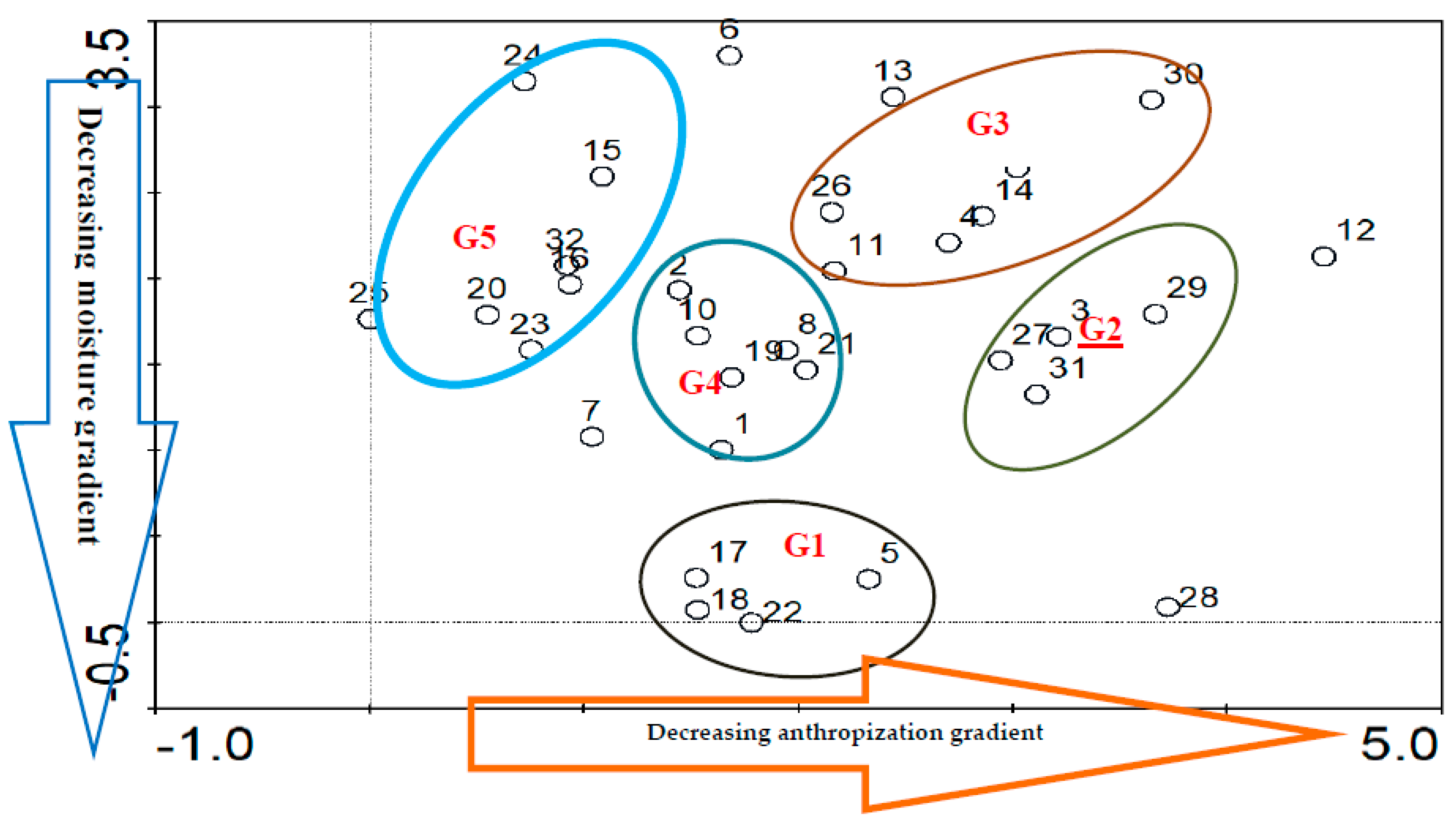
Figure 7.
Distribution of diameter (A) and height (B) classes of group G1.
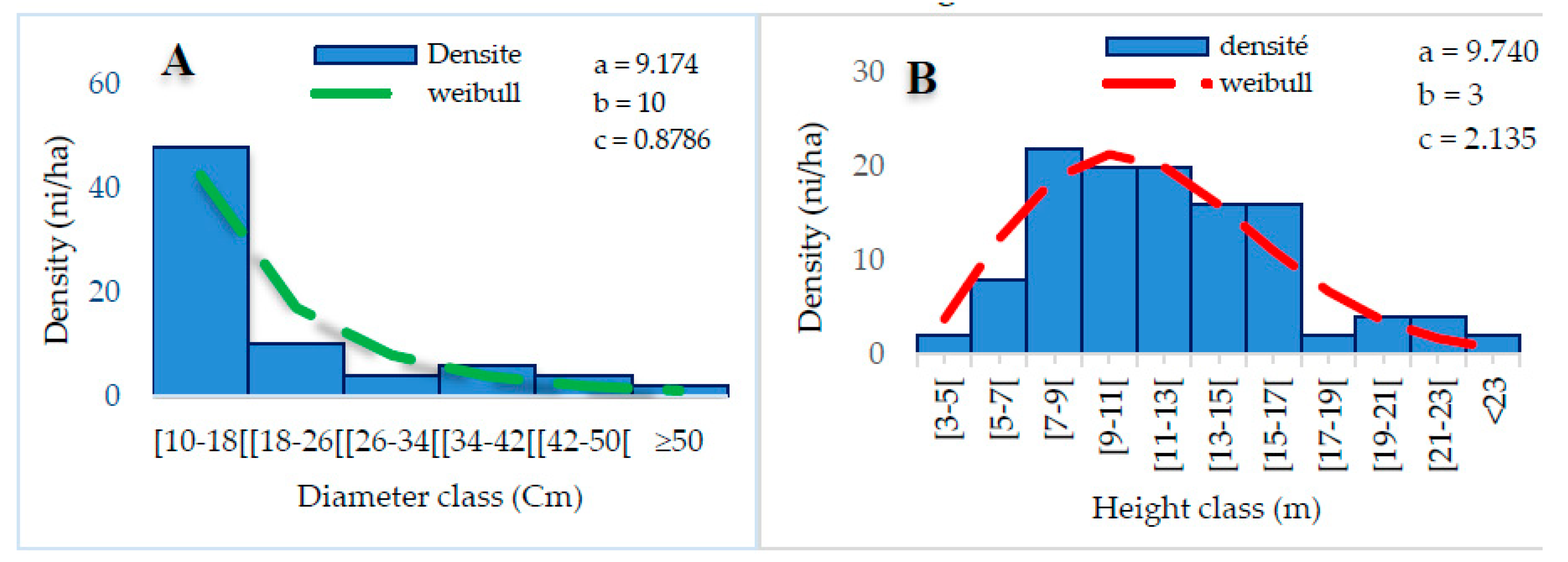
Figure 8.
Distribution of diameter (A) and height (B) classes of group G2.
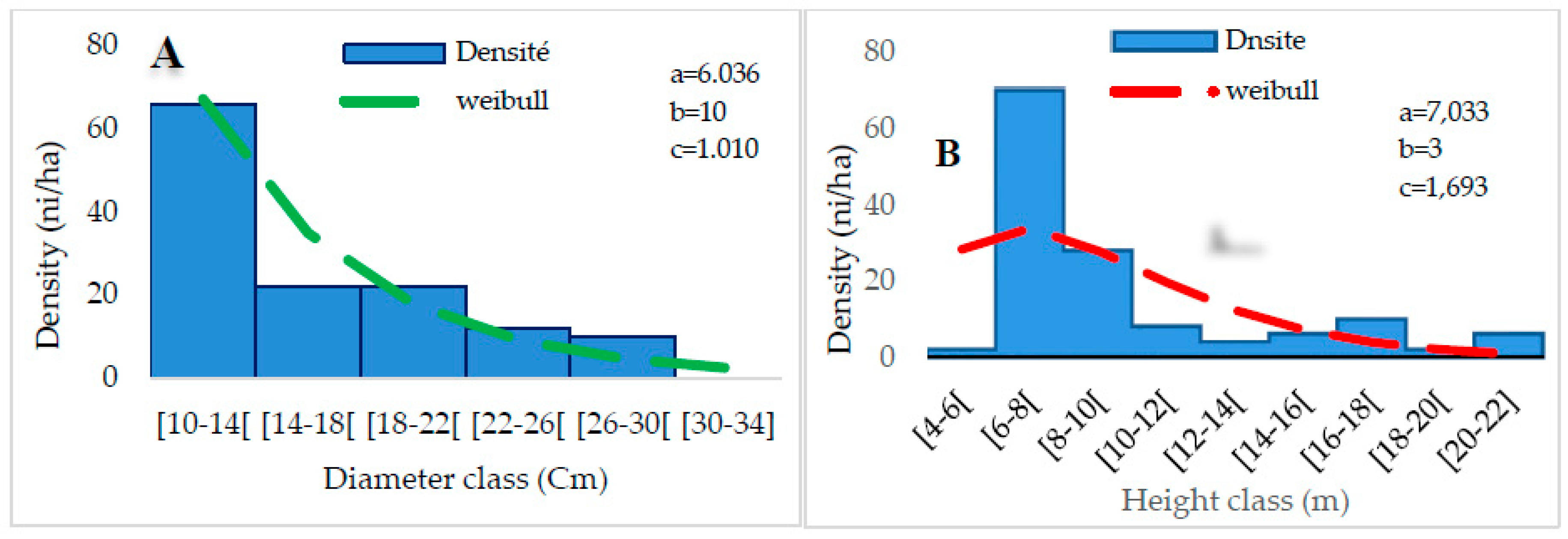
Figure 9.
Distribution of diameter (A) and height (B) classes of group G3.
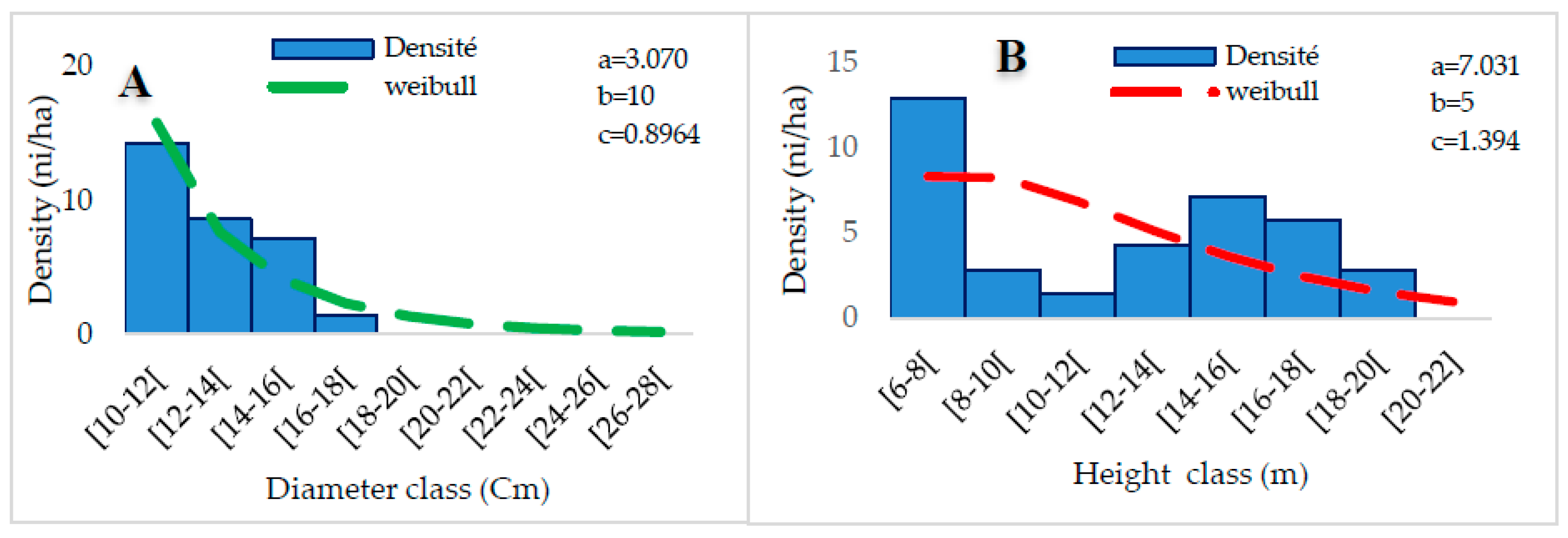
Figure 10.
Distribution of diameter (A) and height (B) classes of group G4.
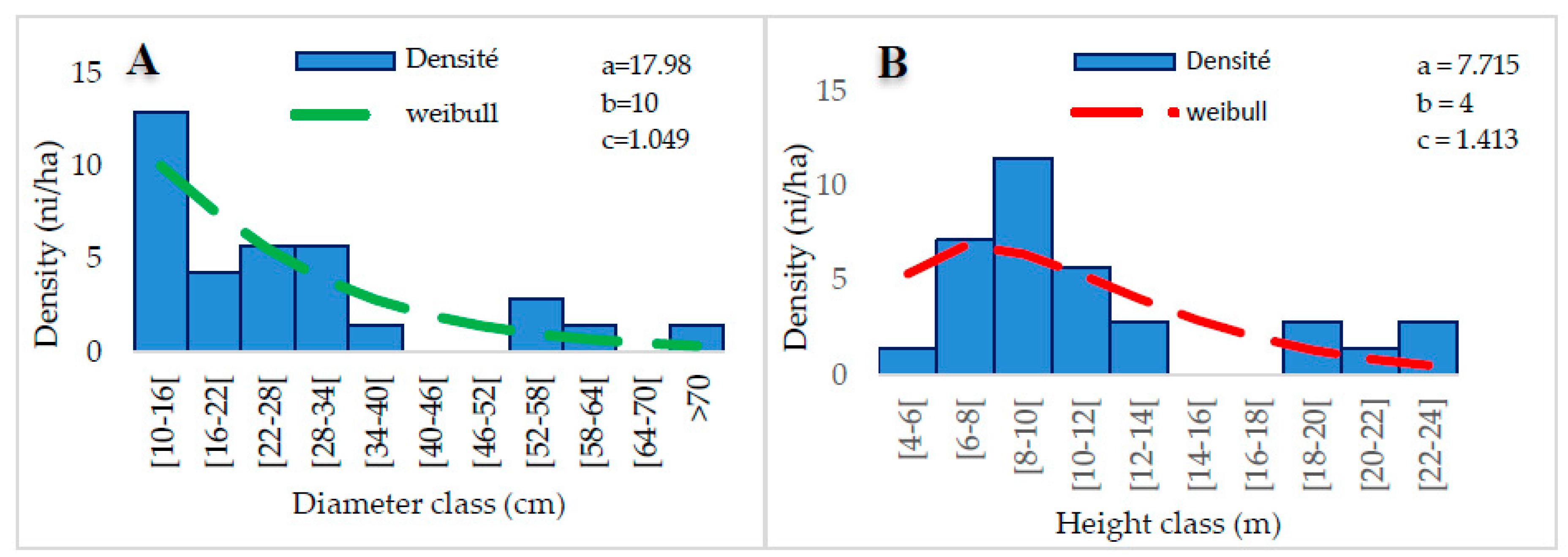
Figure 11.
Distribution of diameter (A) and height (B) classes of group G5.
Figure 12.
Map of the zoning plan.
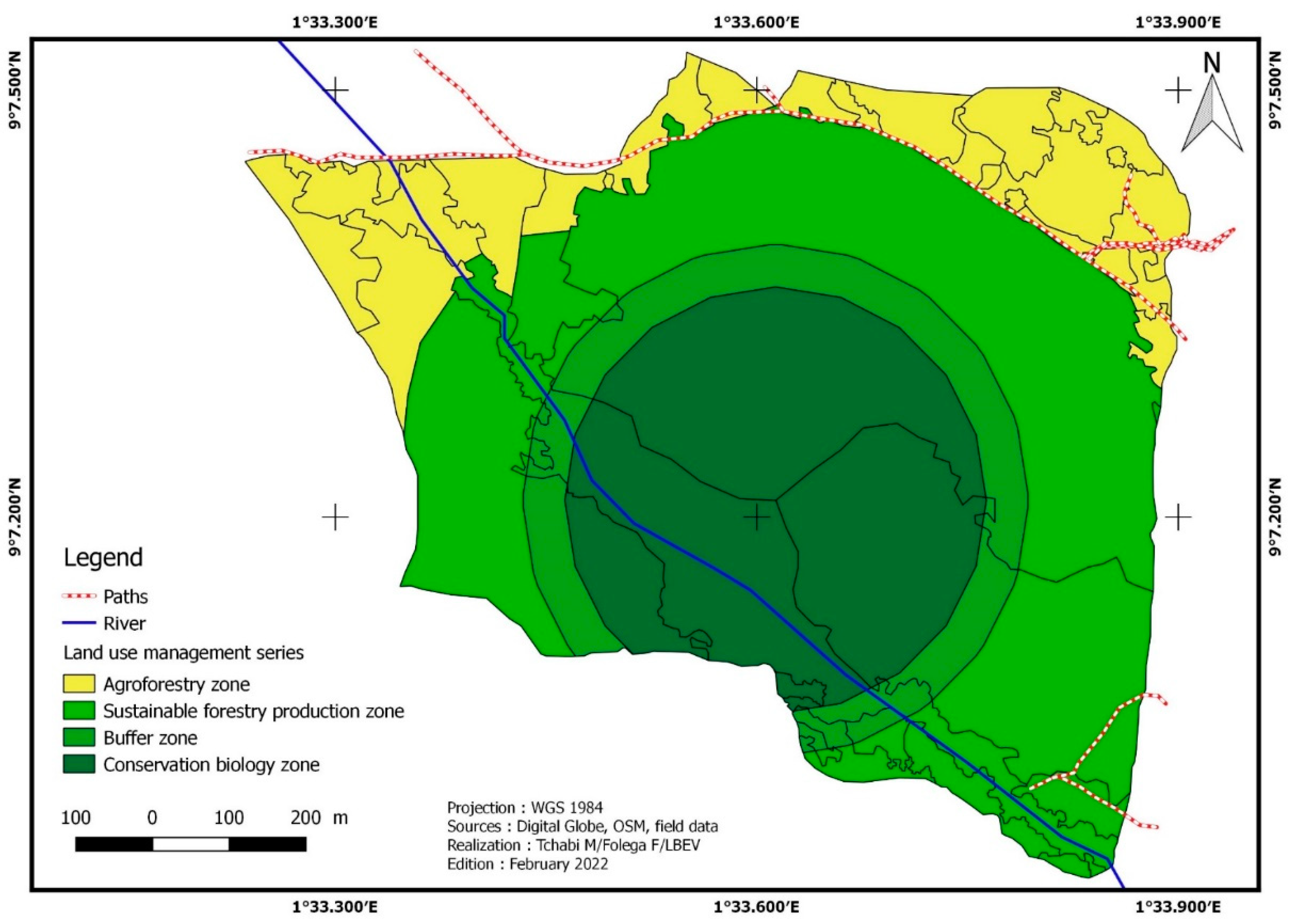
Table 3.
Land Use Unit Assessment Rate.
| Surface area | Assessment Rate % | ||||||||
| 2015 | 2018 | 2021 | |||||||
| Land use Units | Ha | % | Ha | % | Ha | % | 2015-2018 | 2018-2021 | 2015-2021 |
| Crop and fallow | 10.82 | 13.17 | 8.37 | 10.19 | 7.15 | 8.69 | -22.63 | -14.66 | -33.98 |
| Dense dry forest | 35.89 | 43.66 | 36.53 | 44.44 | 33.05 | 40.21 | 1.78 | -9.53 | -7.92 |
| Gallery forest | 10.95 | 13.32 | 10.86 | 13.22 | 10.57 | 12.86 | -0.76 | -2.72 | -3.46 |
| Plantation | 0.75 | 0.91 | - | - | - | - | -100 | - | -100 |
| Tree/Shrub savannahs | 2.41 | 2.93 | 12.84 | 15.62 | 14.08 | 17.14 | 432.40 | 9.73 | 484.23 |
| Grassy Savannah and meadow | 21.37 | 26.00 | 13.59 | 16.53 | 17.34 | 21.10 | -36.41 | 27.63 | -18.84 |
Table 4.
Structural characteristics and diversity index of groups.
| Group | Density (ft/ha) | Average diameter (cm) | Average height (m) | Bole height (m) | Basal area G (m2/ha) | Shannon Index | Pielou Equitability Index |
|---|---|---|---|---|---|---|---|
| G1 | 74 | 22.5 ± 11.14 | 16.1 ± 4.28 | 8.39 ± 3.32 | 2.98 | 3.9 | 0.97 |
| G2 | 136 | 17.2 ± 5.42 | 9.62 ± 4.11 | 3.73 ± 2.35 | 18.65 | 3.63 | 0.87 |
| G3 | 38 | 13 ± 3.78 | 12.35 ± 4.48 | 3.51 ± 2.01 | 4.38 | 4.34 | 0.96 |
| G4 | 36 | 22.5 ± 17.18 | 16.37 ± 5.47 | 5.16 ± 2.73 | 16.43 | 4.04 | 0.92 |
| G5 | 94 | 30 ± 15.30 | 14.84 ± 5.16 | 6.01 ± 3.63 | 7.12 | 3.97 | 0.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



