
Submitted:
29 May 2023
Posted:
30 May 2023
You are already at the latest version
Abstract
Urinary extracellular vesicles (uEV) hold non-invasive RNA biomarkers for genitourinary tract diseases. However, missing knowledge about reference genes and effects of pre-analytical choices hinder biomarker studies. We aimed to assess how pre-analytical variables (urine storage temperature, isolation workflow) affect diabetic kidney disease (DKD) -linked miRNAs or kidney -linked miRNAs and mRNAs (kidney-RNAs) in uEV isolates and to discover stable reference mRNAs across diverse uEV datasets. We studied nine raw and normalized sequencing datasets including healthy controls and individuals with prostate cancer or type 1 diabetes with or without albuminuria. We focused on kidney-RNAs reviewing literature for DKD-linked miRNAs from kidney tissue, cell culture and uEV/urine experiments. RNAs were analyzed by expression heatmaps, hierarchical clustering and selecting stable mRNAs with normalized counts (>200) and minimal coefficient of variation. Kidney-RNAs were decreased after urine storage at -20°C vs -80°C. Isolation workflows captured kidney-RNAs with different efficiencies. Ultracentrifugation captured DKD -linked miRNAs that separated healthy and diabetic macroalbuminuria groups. Eleven mRNAs were stably expressed across the datasets. Hence, preanalytical choices had variable effects on kidney-RNAs – analyzing kidney-RNAs complemented global correlation, which could fade differences in some relevant RNAs. Replicating prior DKD-marker results and discovery of candidate reference mRNAs encourages further uEV biomarker studies.
Keywords:
urinary extracellular vesicles
; exosomes
; urine
; diabetic kidney disease
; reference genes
; miRNA
; mRNA
; sequencing
1. Introduction
Extracellular vesicles (EV) are nowadays a hot topic in the biomarker research field (Chorley et al. 2021; Zhao et al. 2020; de Freitas et al. 2021; Xu et al. 2020). Urinary EV (uEV) are of particular interest for pathologies of the genitourinary tract (Erdbrugger and Le 2016; Puhka et al. 2022; Miranda et al. 2010; Margolis et al. 2022). Specifically for diabetic kidney disease (DKD), a microvascular complication of diabetes, uEV are a promising source of non-invasive biomarkers (Dwivedi et al. 2023; Zubiri et al. 2014; Ghai et al. 2018) that might complement, reduce the need for or eventually even replace kidney biopsies and facilitate early diagnostics and prognosis.
Currently, effort is put on research and to set forth recommendations for uEV work e.g. in sample handling, storage, uEV isolation and reporting (Erdbrügger et al. 2021; Barreiro et al. 2021; Barreiro et al. 2020; Mussack, Wittmann, and Pfaffl 2019; Dong et al. 2020; van Royen et al. 2023; López-Guerrero et al. 2023). This is highly important because there are vast differences in the preanalytical, analytical and reporting procedures. For example, a recent survey by the Spanish Society for Research and Innovation in Extracellular Vesicles (GEIVEX) found that the variability of pre-analytical procedures can be as high as 94% (López-Guerrero et al. 2023). Without some level of standardization, the biomarker discovery results are seldom highly robust or reproducible (Nieuwland et al. 2022).
More specifically, one of the most pressing problem in the pre-analytical part is that many collections in laboratories and biobanks may not be handled and stored optimally for uEV research. Moreover, only few studies have characterized the effect of preanalytical variables on the uEV, especially regarding the end-point biomolecular level used in biomarker studies, e.g. the transcriptome (Barreiro et al. 2021). Equally, only few studies have comprehensively characterized the effect of EV isolation methods on transcriptomics (Mussack, Wittmann, and Pfaffl 2019; Barreiro et al. 2020; Dong et al. 2020; García-Flores et al. 2021; Park et al. 2020; Srinivasan et al. 2019). Thus, it is difficult to compare results between dissimilar studies.
Urinary EV capture kidney transcriptome and proteome (Miranda et al. 2014; Blijdorp et al. 2022; Miranda et al. 2010; Bazzell et al. 2018; Zhang et al. 2016; Dwivedi et al. 2023). We have previously shown by RNA sequencing technologies that some pre-analytical variables such as urine storage temperature and isolation methods affect the uEV RNA yield and global miRNA and mRNA profiles (Barreiro et al. 2021; Dwivedi et al. 2023). However, for kidney research, it would be important to understand how exactly the preanalytical choices affect the uEV as a “liquid kidney biopsy”. Are all the uEV miRNAs and mRNAs - highly or specifically expressed by the kidney - affected by the different pre-analytical variables and to which extent? Are the kidney derived RNAs for example missing completely or just downregulated and therefore still available as biomarkers?
Urinary EV reference genes represent another unmet need in the EV field. Both research on EV reference genes and recommendations on how to select the reference genes are increasing (Dai et al. 2021; Gouin et al. 2017). However, only a moderate number of sequencing datasets are currently available for rigorous search of robust reference genes that would be stable across studies, at least for uEV. Again, the effect of pre-analytical variables, or demographic and disease status variables, on the stability of reference genes is not clear. This represents a problem for qPCR validation experiments. GAPDH is commonly utilized to normalize gene expression but does not work equally fine for all tissues, biofluids, or disease status (Kozera and Rapacz 2013; Singh et al. 2022; Gorji-Bahri et al. 2021). In conclusion, it is unclear how candidate markers reported by different studies could be replicated under different experimental conditions.
In this study, we assessed the effect of storage temperature and uEV isolation workflows on uEV transcriptome by focusing on highly expressed miRNAs and enriched genes of the kidney. We assessed the replicability of previously described candidate miRNA markers of DKD and explored the existence of reference genes across diverse uEV sequencing datasets.
2. Methods
2.1. miRNA and mRNA sequencing datasets
The datasets included in this study were retrieved from previous publications from our group (Barreiro et al. 2020; Barreiro et al. 2021; Dwivedi et al. 2023). Details for each dataset are included in Table 1. Of note, except for the isolation workflow dataset, the rest of the samples were processed by ultracentrifugation.
2.2. Kidney top expressed miRNAs and kidney enriched miRNAs in uEV
Kidney enriched genes (“At least four-fold higher mRNA level in kidney compared to the average level in all other tissues”) were retrieved from The Human Protein Atlas (Habuka et al. 2014; Uhlén et al. 2015) (www.proteinatlas.org) (v20, accessed on 19.11.20). For miRNAs, we used top kidney expressed miRNAs which were retrieved from miRNATissueAtlas2 (Keller et al. 2022) (https://ccb-web.cs.uni-saarland.de/tissueatlas2) (accessed on 17.06.22) and For these analysis, raw sequencing counts were normalized as described in the original publications by using TMM (trimmed mean of M values) (Robinson and Oshlack 2010) in edgeR (Robinson, McCarthy, and Smyth 2010) or DEseq2 normalization (Anders and Huber 2010; Love, Huber, and Anders 2014) .
2.3. Literature review of miRNAs associated with DKD
We did a literature review of miRNAs associated with DKD based on evidence from tissue (human or animal models) or in vitro models and for miRNAs based on evidence form human urine, urinary sediments or uEV. For the later, some studies reported miRNAs with nominal p-values, in such cases we considered only the miRNAs that had been also validated with another quantification method or by using in-vitro or in-vivo models.
2.4. Stable mRNAs across datasets
All datasets were normalized using TMM normalization. Of note, samples from overnight (ON) and 24 h collections (24h), with and without preclearing and technical replicas were normalized together and we refer to this dataset as “technical dataset”. Genes with normalized counts of CPM > 200 in all samples were filtered to calculate the coefficient of variation (CV). The top 100 genes with the lowest CVs were selected from each experimental dataset and the gene lists were compared to identify shared genes across datasets. To assess to which pathways the stable genes contribute to, we used Uniprot knowledge base (UniProt Consortium 2023) (https://www.uniprot.org/). Protein interaction was assessed using STRING V11.5 (Szklarczyk et al. 2019) (https://string-db.org/).
2.5. Data visualization
For data visualization, built-in R functions or packages ggplot2 (Wickham 2016), pheatmap (Kolde 2019), and reshape2 (Wickham 2007), were used. Values are represented as mean ± SEM (standard error of the mean). Figure panels were prepared using corelDRAW 2022 v24.1.0360 (Corel Corporation, ON, Canada).
3. Results
Our study focused on the kidney-linked and putative reference RNAs in uEV isolates targeting applicability for biomarker discovery. The uEV isolates used to generate the eleven sequencing datasets analyzed in this study were comprehensively characterized in our original publications (Table 1) by electron microscopy, Western blotting, and nanoparticle tracking, RNA fragment length and protein analysis. Briefly, this quality control indicted that the main population of uEV and RNA was small in size and length (<300 nm and <300 nt, respectively) and that the presence of e.g. remnant Tamm-Horsfall protein varied, but was not extensive.
3.1. Dysregulated miRNAs in samples stored at suboptimal temperature: significance for kidney disease biomarker discovery
Previously, we reported different miRNA profiles from uEV isolated from urines stored at -20°C vs -80°C (Barreiro et al. 2021) (from now on, for simplicity, we will refer to these samples as “urines stored at -20°C or -80°C”). Specifically, we found 29 downregulated and 4 upregulated uEV miRNAs in urines stored at -20°C compared to the ones stored at -80°C. To assess the biological relevance of the dysregulated miRNAs, we performed a literature review and found that 25/33 miRNAs were associated with kidney diseases (Table 2). In addition, a careful comparison of the raw and normalized counts revealed that most of the downregulated miRNAs in urines stored at -20°C failed to be detected (raw counts=0), while the 4 downregulated miRNAs in urines stored at -80°C were stably expressed across samples and had high raw counts (Table 2, Table S1). Thus, in urines stored at -20°C significant number of potential kidney disease markers were lost and the upregulated genes’ raw counts were actually lower than in urines stored at -20°C. -80°C samples.
3.2. Effect of pre-analytical variables on kidney transcriptome in uEV isolates
In previous studies we determined that some pre-analytical variables such as storage temperature affect the global uEV transcriptome (Barreiro et al. 2020; Barreiro et al. 2021). As the uEV have shown potential as “liquid kidney biopsy” (Dwivedi et al., 2023), we now assessed whether these pre-analytical variables impact the kidney transcriptome in uEV isolates. Here we analyzed the expression level of “kidney-RNAs” i.e. top kidney expressed miRNAs and kidney enriched mRNAs.
3.2.1. Effect of storage temperature
To analyze the effect of urine storage temperature on miRNAs that have high expression in the kidney, we focused on the top 40 kidney expressed miRNAs. In our dataset (n=8 samples), we found 29 out of the 40 miRNAs and for 22 of those, the normalized expression level was lower in urines stored at -20°C than in urines stored at -80°C (Figure 1). Of note, two of the miRNAs were not detected at all in the -20°C samples (Table S2).
Out of 56 kidney enriched mRNAs we found 33 in our dataset. Analysis of the expression levels showed that 15 mRNAs were poorly represented in urines stored at -20°C compared to the ones stored at -80°C (Figure 1 A). Importantly, 10 of the mRNAs were not detected at all in the -20°C samples (raw counts=0) (Table S2).
3.2.2. Effect of isolation workflows
We next analyzed the effect of the EV isolation workflows on the uEV expression of kidney-RNAs. Out of 40 highly expressed miRNAs of the kidney, we found 36 in our datasets (n=26 samples). All the miRNAs were stably expressed across the different isolation workflows but the expression of 18 miRNAs was lower in samples from HFD workflow (Figure 2 A). We then analyzed differences in the normalized counts of these 18 miRNAs between HFD and UC samples (samples that showed low expression in HFD 4,5,6,8,9,10) and we observed that the normalized counts were systematically lower in HFD samples compared to UC, with differences ranging between 3-58%. MiRNAs with highest differences (>35%) were hsa-miR-101-3p, hsa-miR-26a-5p, hsa-miR-26b-5p, hsa-miR-27a-3p, hsa-miR-29c-3p. Regarding the kidney enriched genes, we found 31 out of the 56 and all of them had lower expression in samples from Norgen urine Exosome Purification and RNA Isolation Midi Kit (NG) (Figure 2 B). Five of the mRNA were not detected in any of the NG samples (raw counts=0) and generally, many of the samples had raw count 0 (Table S3).
Both temperature and isolation workflow impact the kidney transcriptome in uEV isolates and these differences are better captured by analyzing kidney-RNAs than analyzing global expression.
3.3. Replication of DKD –associated miRNA by UC –based uEV isolation and sequencing workflow
Prior research has reported many miRNAs that associate with DKD in T1D and/or T2D. Thus, we carried out a literature search to generate a list of these DKD -linked miRNAs (p adj <0.05 or p<0.05 and other evidence of association, see methods) and used it for studying their expression in the UC –isolated uEV from DKD patients vs heathy controls (n=10 samples). We found i) 107 miRNAs based on evidence from tissue (human or disease models) or in vitro models and ii) 63 miRNAs based on evidence form human urine, urinary sediments or uEV (Table 3 and 4). We analysed their expression levels in our uEV data using expression heatmaps and checked whether the miRNAs could cluster the healthy control and macroalbuminuria groups separately by hierarchical clustering. Our uEV set showed expression of 39 out of 107 miRNAs (36%) dysregulated in DKD with evidence from tissue and in vitro studies, but they did not separate the groups (Figure 3 A). However, our uEV set showed expression of a higher proportion of miRNAs - 31 out of 54 (57%) – that were dysregulated in DKD with evidence from urine, urine sediment or uEV. Importantly, this set of miRNAs could divide the DKD and healthy control groups into separate clusters (Figure 3 B). We focused on the first and last cluster of miRNAs because on those clusters the expression differences were better observed. From those miRNAs, we compared the direction of change between the literature review and our dataset. For the first cluster, miR-30b-5p, miR-221-3p, let-7f-1-3p, and miR-30c-5p followed the same direction of change in both i.e. downregulated in DKD. In contrast, miR-15b-5p was upregulated in the literature with evidence from uEV/urine or urine sediments but downregulated in our uEV dataset. For the second cluster, all miRNAs (miR-424-5p, miR-486-3p, miR-335-5p, miR-126-3p) had the same direction of change than what was found in the literature i.e. upregulated in DKD. Moreover, miRNAs had evidence of association with DKD in vitro or in vivo and/or association with DKD pathways (Table 5).
To assess whether these 31 miRNAs would show some specificity for DKD, we carried a similar analysis using our uEV PCa dataset. Supporting DKD specificity, the analysis did not separate the PCa patients from healthy controls (Figure 3 C).
Taken together, despite variability between experimental setups, some uEV/urine/urine sediment miRNAs candidate markers associated with DKD in the literature were found in our uEV dataset and expression level change between experimental groups was concordant.

Table 4.
MiRNAs associated with DKD development and/or progression with evidence from urine, urinary sediments or uEV. Chronic kidney disease (CKD), diabetic kidney disease (DKD), intermittent microalbuminuria (IMA), persistent microalbuminuria (PMA), microalbuminuria (MA), type 1 diabetes (T1D), type 2 diabetes (T2D). * Validated with an independent cohort, ** validated with another detection method, in kidney biopsies, in vitro or in a model organism.
Table 4.
MiRNAs associated with DKD development and/or progression with evidence from urine, urinary sediments or uEV. Chronic kidney disease (CKD), diabetic kidney disease (DKD), intermittent microalbuminuria (IMA), persistent microalbuminuria (PMA), microalbuminuria (MA), type 1 diabetes (T1D), type 2 diabetes (T2D). * Validated with an independent cohort, ** validated with another detection method, in kidney biopsies, in vitro or in a model organism.
| Sample | Groups | Upregulated miRNAs | Downregulated miRNAs | Reference |
|---|---|---|---|---|
| Urine | Urine from T1D (Normal, overt nephropathy, intermittent microalbuminuria, persistent microalbuminuria) | DKD vs non DKD: miR-619, miR-486-3p, miR-335-5p, miR-552, miR-1912, miR-1124-3p, miR-424-5p, miR-141-3p, miR-29b-1-5p | DKD vs non-DKD: miR-221-3p | (Argyropoulos et al. 2013) |
| MA vs baseline: miR-214-3p, miR-92b-5p, miR-765, miR-429, miR-373-5p, miR-1913, miR-638 | MA vs baseline: miR-323b-5p, miR-221-3p, miR-524-5p, miR-188-3p | |||
| PMA vs IMA: miR323b-5p, miR-433, miR-17-5p, miR-222-3p, 628-5p | PMA vs IMA: miR-589-5p, miR-373-5p, miR-92a-3p | |||
| Urinary sediments | Diabetic glomerulosclerosis, minimal change nephropathy or focal glomerulosclerosis, membranous nephropathy, and healthy donors | miR-200c | miR-638, miR-192 | (Wang et al. 2013) |
| uEV** | T1D with normoalbuminuria and microalbuminuria and non-diabetic controls | miR-130a, miR-145 | miR-155, miR-424 | (Barutta et al. 2013) |
| Urine | T2D DKD, T2D, and healthy donors | miR-126 (T2D DKD > T2D) | (Liu et al. 2014) | |
| uEV | T2D normoalbuminuric, microalbuminuric, or macroalbuminuric | microalbuminuria vs normoalbuminuria and controls: miR-192, miR-194, and miR-215. | macroalbuminuria vs microalbuminuria: miR-192, miR215 | (Jia et al. 2016) |
| uEV* | T2D DKD, T2D, and healthy donors | miR-320c, miR-6068 | (Delić et al. 2016) | |
| urine pellets and uEV * | T2D albuminuric, normoalbuminuric, and healthy controls | miR-15b, miR-34a, miR-636 | (Eissa et al. 2016) | |
| uEV* | T2D normoalbuminuria and microalbuminuria | miR-877-3p | (Xie et al. 2017) | |
| uEV** | T1D normoalbuminuria, intermittent macroalbuminuria, persistent macroalbuminuria, and overt macroalbuminuria | Overt vs normal: miR-26a-1-5p, miR-30-5p PMA vs IMA/ non microalbuminuria: miR-200c-3p | Overt vs normal: miR144-3p | (Ghai et al. 2018) |
| Urine** | PMA vs IMA: miR-10a-5p, miR-200a-3p | |||
| Urine* | Diabetic, DKD and healthy donors | miR-126-3p, miR-155-5p, and miR-29b-3p | (Beltrami et al. 2018) | |
| Urine* | Urine and plasma from T1D and DKD | miR-30e-5p | (Dieter et al. 2019) | |
| uEV* | T2D DKD, T2D normal renal function, and non-T2D CKD | miR-21-5p | miR-30b-5p | (Zang et al. 2019) |
| Urine** | DKD and non-diabetic renal disease | T2D vs non-diabetic renal disease: miR-27-3p, miR-1228 | (Conserva et al. 2019) | |
| uEV* | T2D and normoalbuminuria, microalbuminuria or macroalbuminuria and healthy donors | miR-15b-5p | (Tsai et al. 2020) | |
| uEV* | TD2 DKD and healthy donors | miR-30e-3p, miR-30c-5p, miR-190a-5p, miR-98-3p, let-7a-3p, miR-30b-5p, and let-7f-1-3p | (Park et al. 2022) |
Table 5.
miRNAs dysregulated in uEV/urine/urinary sediments. Cluster 1 and 4 miRNAs and association with diabetic kidney disease or diabetic kidney disease mechanisms. Acute kidney injury (AKI), diabetic kidney disease (DKD), type 2 diabetes (T2D), urinary extracellular vesicle (uEV).
Table 5.
miRNAs dysregulated in uEV/urine/urinary sediments. Cluster 1 and 4 miRNAs and association with diabetic kidney disease or diabetic kidney disease mechanisms. Acute kidney injury (AKI), diabetic kidney disease (DKD), type 2 diabetes (T2D), urinary extracellular vesicle (uEV).
| Regulation | ||||
| DKD in UC dataset |
uEV/ urine/ Urine sediments literature |
Examples of association with diabetic Kidney disease or kidney diseases, or pathways associated with DKD (e.g. fibrosis, inflammation, autophagy, and oxidative stress) | ||
| Cluster 1 | miR-30b-5p | down | down | In hyperglycemic conditions, expression levels reduced in HK-2 cells and epithelial-mesenchyma increased (Wang et al. 2021) |
| miR-221-3p | down | down | Associated with tumorigenesis and prognosis of kidney renal clear cell carcinoma (Zhou et al. 2022). In HUVEC cells, hyperglycemia induced this miRNA and was associated with impairment of endothelial cell migration and homing mediated (Li et al. 2009). | |
| miR-15b-5p | down | up | Up regulated in urine from db/db mouse and T2D patients. In mesangial cell lines hyperglycemia upregulates this miRNA and targets BCL-2 inducing apoptosis (Tsai et al. 2020). | |
| let-7f-1-3p | down | down | Upregulated in the kidney of a rat model of AKI (Liu et al. 2019). By regulating negatively STAT3, induces autophagy in glioma cells (Yang et al. 2020). | |
| miR-30c-5p | down | down | Downregulate in kidney tissue from diabetic patients and high glucose-induced HK-2 cells. Downregulation causes epithelial to mesenchymal transition mediated by ROCK2 (Cui, Yu, and Cui 2020). | |
| Cluster 4 | miR-424-5p | up | up | Upregulated in high fat diet induced mouse and hepatocytes treated with palmitate. miR-424-5p suppressed insulin receptor expression in hepatocytes i.e. role in insulin resistance(Min, Yang, and Lee 2018). |
| miR-486-3p | up | up | Downregulated in biopsies from patients with diabetic nephropathy (Baker et al. 2017) | |
| miR-335-5p | up | up | In childhood asthma, the upregulation of this miRNA is associated with reduced inflammation, fibrosis and autophagy by regulating ATG5 (Liang et al. 2022). | |
| miR-126-3p | up | up | Increased in kidney biopsies from patients with DKD (Beltrami et al. 2018) | |
3.4. Exploratory analysis of reference mRNA in uEV
To select the most stable mRNAs that could serve as candidate reference genes we first focused on uEV samples from our DKD studies that included men only. This choice was due to expected and higher sample-linked (Dwivedi et al., 2023) and also biological heterogeneity in the women´s cohorts. Datasets were analyzed separately to avoid batch effects i.e. isolation workflows (n=20 samples), in column DNAse treatment (n=19 samples), technical dataset (type of collection, pre-clearing, and replicability, see methods) (n=39 samples), and DKD cohort 1 (T1D, men) (n=72 samples). Of note, NG isolation workflow data and storage temperature dataset were excluded from the analysisdue to the low expression level of many mRNAs (please see raw counts in Table S3). The top 100 uEV genes with lowest CV were selected from each dataset and the genes overlapping between all of them were selected for further analysis. We found 32 uEV genes in common between the datasets (Figure 4 A).
We next expanded our reference gene analysis to check the stability of expression including women´s uEV samples. Here we searched genes in common between the DKD male (32 uEV stable mRNAs from first search) and DKD cohort 2 (T2D, women) (n=30 samples) using again the top 100 uEV RNA with low CV (in cohort 2), which showed 18 mRNAs in common (Figure 4 B). Finally, we assessed whether some of these 18 mRNAs could also be found from the PCa dataset (n=8 samples) listing the top 100 uEV mRNAs with low CV. This analysis showed 11 mRNAs in common (HSPD1, SRSF3, VAPA, RAB1A, MORF4L1, PGK1, RHOA, UBE2D3, DAZAP2, UBC, ACTG1) with low CV (Figure 4 C, Table 6).
We analyzed the stable genes counts per million (CPM) across samples. In addition we included GAPDH (a commonly used reference genes) and a gene with high CV (UPK1A) to facilitate the comparison (Table 5). CPM analysis showed that ACTG1 presents similar variation than GAPDH across samples but in both cases the variation was low compared to the gene with highest CV (UPK1A) in all datasets (Figures 5,6 and Figures S1 and S2). For visualization of CPM values across samples, the candidate reference genes were sorted by decreasing standard deviation (SD) value. The 5 genes with lowest SD value are plotted in Figure 5 and Figure 6 (A-D) and the remaining 6 genes are plotted in Figures S1 and S2 (A-B). We also summarized the data in boxplots to visualize the CPM dispersion per gene (Figure 5 E-H and Figure 6 C,D).
We next examined to which pathways the stable genes contribute and as shown in Table 6, they participate in several different prominent pathways. However, UBE and UBE2D3 both ubiquitylate proteins. Moreover, an analysis of protein interaction using STRING showed interaction (with experimental evidence) of UBC with UBE2D3 and DAZAP2 and of MORF4L1 with ACTG1 (Figure 7).
Further we analyzed the stability of the candidate reference genes in datasets from samples that did not perform well in mRNA sequencing i.e. urines stored at -20°C and uEV isolated with NG isolation workflow. We found that all genes were less stable in samples stored at -20°C and in NG isolates (Figure S3). Of note, in many NG samples the candidate reference genes were not detected. Despite of this, HSPD1, SRSF3, VAPA, RAB1A, MORF4L1, PGK1, RHOA, UBE2D3, DAZAP2, UBC, ACTG1, showed to be stable in all other diverse experimental conditions and across disease groups. Thus, they may serve as reference genes for uEV mRNA related research.
4. Discussion
Urinary EV have been regarded as a promising source of biomarkers (Erdbrugger and Le 2016) and this idea is getting support from an increasing number of studies reporting candidate markers for diseases of diverse etiology (Margolis et al. 2022; Agudiez et al. 2020; Zhu et al. 2022; Zang et al. 2019; Dwivedi et al. 2023). However, many obstacles prevent replication of biomarker results and, as a consequence, clinical translation. In this study, we approached three of these obstacles: urine storage, uEV isolation and reference genes in kidney disease transcriptomic research.
The first obstacle is the lack of guidelines to handle and store urine. Urine storage temperature (-20°C vs -80°C) has been shown to affect the size and concentration of uEV (Oosthuyzen et al. 2013) and recovery of uEV protein markers but the latter could be sorted out by vortexing samples after thawing (Zhou et al. 2006). In addition, qPCR-based research has been done on uEV RNA by comparing storage temperatures – including -80°C, 4°C, room temperature and 37°C with variable results (Hogan et al. 2015; Armstrong et al. 2018; Vago et al. 2022). Our group showed that the global uEV miRNA and mRNA profiles were affected when urines were stored at -20°C vs -80°C (Barreiro et al. 2021) and we found sets of downregulated and upregulated genes. As particularly the -20°C downregulated genes were involved e.g. in carbohydrate or lipid metabolism, the result suggested that -20°C stored samples are less useful for studying kidney diseases. Here, analyzing further the data, we found that a striking 75% of the -20°C downregulated miRNAs were associated with various kidney diseases (Table 2). Thus, the result reinforces the idea of avoiding urine samples stored at suboptimal temperatures (Barreiro et al. 2021), because such samples might not contain putative valuable disease markers anymore.
We also observed that despite the normalized differential expression, miRNAs that were up-regulated in samples stored at -20°C had still lower raw counts than the same miRNAs in -80°C stored samples. Thus, the result was the opposite than what the normalized counts showed. TMM normalization is a method based on library size that uses scaling of raw reads to render library sizes comparable (Abbas-Aghababazadeh, Li, and Fridley 2018). Considering that the library size of the -20°C samples was smaller (higher number of 0 raw counts and lower expression in general) than that of the -80°C samples, the upregulation of miRNAs in -20°C samples may be an artifact of the data analysis. Further, we showed that kidney-RNAs were detected in small quantities after storage at -20°C (Figure 1). In particular, kidney enriched mRNAs in uEV isolates were highly affected since almost one third (30%) of them were not detected at all in samples stored at -20°C. Our results agree with and provide further support to a set of urine storage guidelines that has been published recently (van Royen et al. 2023).
The second obstacle is the lack of standardization of uEV isolation methods. Currently, many isolation principles and workflows are available (Royo et al. 2020) and it is well known that they typically produce different results (Vago et al. 2022; Barreiro et al. 2020; Sáenz-Cuesta et al. 2015; Mussack, Wittmann, and Pfaffl 2019; Dong et al. 2020). Obviously, this represents a problem for study comparisons, even if reporting guidelines now help to identify differences, facilitate replication and/or explain the lack of it (Van Deun et al. 2017; Erdbrügger et al. 2021). Prior studies have explored the effect of uEV isolation workflows on uEV RNA sequencing profiles focusing on miRNA sequencing (Mussack, Wittmann, and Pfaffl 2019; Srinivasan et al. 2019; Park et al. 2020; García-Flores et al. 2021). We have previously demonstrated that the uEV isolation workflow (UC, HFD and NG) has a surprisingly variable impact on the miRNA and mRNA profiles (Barreiro et al. 2021). Specifically, global miRNA profile analysis suggested that the three workflows were similar overall or – at least – did not differ systematically. This was in contrast to the global mRNA results, where UC and HFD were similar while NG clustered separately (Barreiro et al. 2021). Here, by analyzing the top expressed miRNAs of the kidney, we found that the expression of 18 miRNAs was lower for a set of HFD samples compared to UC and NG samples (Figure 2). While for 13 miRNAs the differences in the expression levels between UC and HFD were small (3-35%) and could be related to technical bias, for 5 miRNAs differences were in the range of 35-55% and could represent real differences. Of note, miRNA hsa-miR-101-3p (a top kidney expressed miRNA) was significantly downregulated in HFD relative to UC samples (Barreiro et al. 2020). The observation that methods capture slightly different kidney enriched miRNAs could be explained, at least partly, by differences in the uEV populations and/or non-EV components captured by the isolation workflows (Barreiro et al. 2020). On the other hand, analysis of kidney enriched mRNAs was consistent with the global analysis i.e. NG samples could not capture these genes as well as UC and HFD (Figure 2). Thus, this shows that for a specific research topic like kidney research, it is best to evaluate the differences between methods using specific end-point targets (kidney-RNAs) in addition to a global level analysis.
In addition to urine storage temperature and uEV isolation workflows, many other pre-analytical experimental conditions impact the analytical endpoints as well (Erdbrügger et al. 2021). As experimental set-ups can differ greatly between studies (López-Guerrero et al. 2023), biomarker results cannot be replicated hindering translation of findings to clinic (Yekula et al. 2020). Considering all the variability, we were positively surprised that our UC workflow replicated some of the previous results from DKD miRNA studies using uEV/urine or urine sediments (see Table 4) i.e. a set of the miRNAs separated experimental groups (healthy controls vs T1D with macroalbuminuria) (Figure 3). Further, specifically eight miRNAs followed the same regulation direction in both the literature and our dataset. Six of the eight miRNAs had also evidence of DKD-linked dysregulation in kidney tissue and/or in vitro models of DKD (in addition to evidence in urine/uEV) and all of them related to pathological mechanisms in DKD such as fibrosis and impaired autophagy (Tuttle et al. 2022; DeFronzo, Reeves, and Awad 2021; Toth-Manikowski and Atta 2015) (Table S4). However, our dataset had a low number of samples and thus replication of findings in bigger cohorts is still needed.
The third obstacle jeopardizing the biomarker replication is the lack of normalizers - in the EV transcriptomics field, this means lack of stable reference genes across e.g. many pre-analytical workflows and disease conditions. Urinary EV reference genes are a poorly explored topic. While some recommendations exist on how to select reference genes or normalize gene expression data (Gouin et al. 2017; Dai et al. 2021; Mateescu et al. 2017), there are only few studies on this topic in urine. GAPDH, a commonly used reference gene, and UBC were the most stable in EV derived from liver and breast cancer cell lines (Gorji-Bahri et al. 2021). In contrast, Singh et al. (2022) tested five common reference genes (including GAPDH) and found that B2M and RPL13 were the most stable in uEV isolated using PEG from patients with renal graft dysfunction. Thus, the stability of GAPDH appears to be dependent on the disease, biofluid and/or EV isolation method. In this study, using datasets available in our original publications (Barreiro et al. 2020; Barreiro et al. 2021; Dwivedi et al. 2023), we discovered 11 mRNAs (HSPD1,SRSF3, VAPA, RAB1A, MORF4L1, PGK1, RHOA, UBE2D3, DAZAP2, UBC, ACTG1) that were stable across datasets including different preanalytical conditions, men and women, healthy controls, T1D and T2D patients with different albuminuria status; and prostate cancer patients (Figures 4,5,6, Figures S1 and S2). However, in poor quality sequencing datasets (urine stored at -20°C and NG isolation workflow), the candidate genes showed poor stability i.e. high CV (Figure S3). Of note, our finding regarding UBC stability in uEV is concordant with findings of Gorgi Bahri (2021) in cell culture media derived EV. Further, even though GAPDH was not one of the most stable mRNAs, it was less variable as UPK1A (a highly variable mRNA selected to compare our candidate reference mRNAs).
One of the reasons that prevents the study of uEV reference genes is the lack of data. Many studies have focused on miRNA/small RNA sequencing, but only few on RNA sequencing. Moreover the few studies with a good amount of uEV samples from patients (Ghai et al. 2018; Chen et al. 2021) do not have the associated raw sequencing data and/or raw sequencing counts freely available (to date). Local regulations could hinder the publication of sequencing data but raw count data describing all the pre-processing and alignment procedures is also helpful for the research community. Such practicalities should be considered for the informed consent and ethical permissions. Given more available datasets in future, the stability of our 11 candidate mRNAs could be further tested and a combination of selected genes used as reference genes e.g. by calculating the geometrical mean (Vandesompele et al. 2002). As it is recommended that reference genes should belong to different biological pathways and that expression is regulated independently for each reference gene, caution should be taken if using UBC and UBED3 and/or DAZAP2 or MORF4L1 and ACTG1 together since their corresponding proteins show experimentally validated interactions (Figure 7) and all are involved in ubiquitylation. In addition, UBC and UBE2D3 are co-expressed (https://string-db.org/) and form a protein complex (Pruneda et al. 2014).
We acknowledge that a full understanding of the effect of all preanalytical choices and pathophysiological conditions for transcriptomic applications calls for big testing resources. Ideally, cross-laboratory testing should be performed, and laboratories could implement reference materials, a gold standard isolation protocol, and housekeeping normalizers. Our results here help towards this goal by providing new insights for the three key obstacles hindering uEV biomarker validation. For the first two, urine storage and uEV isolation, we found that it is important to study the raw counts in addition to the normalized counts and kidney-RNAs in addition to the global transcriptome – they offer different although complementary results. For the third, the reference genes, we provide 11 mRNAs that could be tested for qPCR normalization in the context of DKD and prostate cancer. Finally, despite the known and hereby addressed variability between uEV studies, we successfully replicated many previously found urine/uEV/urinary pellet miRNAs associated with DKD in our UC DKD dataset. We regard this as an encouraging result for the reproducibility of uEV biomarker research.
Supplementary Materials
Figure S1: The mRNA sequencing read counts of six of the eleven candidate reference genes across samples in preanalytical and DKD uEV datasets from men. Figure S2: The mRNA sequencing read counts of six of the eleven candidate reference genes across samples in uEV datasets from DKD study of women and from prostate cancer patients. Figure S3: The mRNA sequencing read counts of candidate reference genes in storage temperature and NG datasets. Table S1: Dysregulated miRNAs in samples stored at -20°C, raw and normalized counts. Table S2: Kidney RNAs raw and normalized counts for storage temperature dataset. Table S3: Kidney RNAs raw and normalized counts for Isolation workflows.
Authors’ Contributions
Made substantial contributions to conceptualization: Barreiro K, Puhka M; investigation: Barreiro K, Dwivedi O, Rannikko A, Holthöfer H, Tuomi T, Groop PH, Puhka M; formal analysis and visualization: Barreiro K, Puhka M; supervision: Puhka M; writing-original draft: Barreiro K, Puhka M; writing-review and editing: all authors contributed.
Financial Support and Sponsorship
This project has received funding from the Innovative Medicines Initiative 2 Joint Undertaking under grant agreement No 115974. The JU receives support from the European Union’s Horizon 2020 research and innovation programme and EFPIA and JDRF. Any dissemination of results reflects only the author’s view; the JU is not responsible for any use that may be made of the information it contains.
Availability of Data and Materials
Datasets used in this publication are available in the original publications mentioned in Table 1.
Acknowledgments
We acknowledge the ISEV Urine Task force and BEAt-DKD project partners for discussions and inspiration.
Conflicts of Interest
K.B., O.D.P, H.H., T.T., and M.P. declare no conflict of interest. P-H.G. has received research grants from Eli Lilly and Roche; is an advisory board member for AbbVie, AstraZeneca, Boehringer Ingelheim, Cebix, Eli Lilly, Jansen, MSD, Novartis, NovoNordisk and Sanofi; and has received lecture fees from Boehringer Ingelheim, Eli Lilly, Elo Water, Genzyme, MSD, Novartis, Novo Nordisk and Sanofi. K.B., O.D.P, H.H., T.T., and M.P. declare no conflict of interest.
Declarations
Some of the results presented here are part of Karina Barrreiro’s dissertation which is accessible in the Digital Repository of the University of Helsinki (HELDA).
References
- Abbas-Aghababazadeh, F.; Li, Q.; Fridley, B.L. Comparison of normalization approaches for gene expression studies completed with high-throughput sequencing. PLOS ONE 2018, 13, e0206312. [Google Scholar] [CrossRef]
- Agudiez, M.; Martinez, P.J.; Martin-Lorenzo, M.; Heredero, A.; Santiago-Hernandez, A.; Molero, D.; Garcia-Segura, J.M.; Aldamiz-Echevarria, G.; Alvarez-Llamas, G. Analysis of urinary exosomal metabolites identifies cardiovascular risk signatures with added value to urine analysis. BMC Biol. 2020, 18, 1–9. [Google Scholar] [CrossRef]
- Alvarez, M. L. M. Khosroheidari, E. Eddy, and J. Kiefer. 2013. ’Role of microRNA 1207-5P and its host gene, the long non-coding RNA Pvt1, as mediators of extracellular matrix accumulation in the kidney: implications for diabetic nephropathy’, PLoS One, 8: e77468.
- Anders, S. , and W. Huber. 2010. ’Differential expression analysis for sequence count data’, Genome Biol, 11: R106.
- Argyropoulos, C. K. Wang, S. McClarty, D. Huang, J. Bernardo, D. Ellis, T. Orchard, D. Galas, and J. Johnson. 2013. ’Urinary microRNA profiling in the nephropathy of type 1 diabetes’, PLoS One, 8: e54662.
- Armstrong, D.A.; Dessaint, J.A.; Ringelberg, C.S.; Hazlett, H.F.; Howard, L.; Abdalla, M.A.; Barnaby, R.L.; Stanton, B.A.; Cervinski, M.A.; Ashare, A. Pre-Analytical Handling Conditions and Small RNA Recovery from Urine for miRNA Profiling. J. Mol. Diagn. 2018, 20, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Geng, J.; Zhou, Z.; Tian, J.; Li, X. MicroRNA-130b improves renal tubulointerstitial fibrosis via repression of Snail-induced epithelial-mesenchymal transition in diabetic nephropathy. Sci. Rep. 2016, 6, 20475. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.A.; Davis, S.J.; Liu, P.; Pan, X.; Williams, A.M.; Iczkowski, K.A.; Gallagher, S.T.; Bishop, K.; Regner, K.R.; Liu, Y.; et al. Tissue-Specific MicroRNA Expression Patterns in Four Types of Kidney Disease. J. Am. Soc. Nephrol. 2017, 28, 2985–2992. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, K.; Dwivedi, O.P.; Leparc, G.; Rolser, M.; Delic, D.; Forsblom, C.; Groop, P.; Groop, L.; Huber, T.B.; Puhka, M.; et al. Comparison of urinary extracellular vesicle isolation methods for transcriptomic biomarker research in diabetic kidney disease. J. Extracell. Vesicles 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, K. O. P. Dwivedi, S. Valkonen, P. H. Groop, T. Tuomi, H. Holthofer, A. Rannikko, M. Yliperttula, P. Siljander, S. Laitinen, E. Serkkola, T. Af Hällström, C. Forsblom, L. Groop, and M. Puhka. 2021. ’Urinary extracellular vesicles: Assessment of pre-analytical variables and development of a quality control with focus on transcriptomic biomarker research’, J Extracell Vesicles, 10: e12158.
- Barutta, F.; Tricarico, M.; Corbelli, A.; Annaratone, L.; Pinach, S.; Grimaldi, S.; Bruno, G.; Cimino, D.; Taverna, D.; Deregibus, M.C.; et al. Urinary Exosomal MicroRNAs in Incipient Diabetic Nephropathy. PLOS ONE 2013, 8, e73798. [Google Scholar] [CrossRef] [PubMed]
- Bazzell, B.G.; Rainey, W.E.; Auchus, R.J.; Zocco, D.; Bruttini, M.; Hummel, S.L.; Byrd, J.B. Human Urinary mRNA as a Biomarker of Cardiovascular Disease. Circ. Genom. Precis. Med. 2018, 11, e002213–e002213. [Google Scholar] [CrossRef]
- Beltrami, C. K. Simpson, M. Jesky, A. Wonnacott, C. Carrington, P. Holmans, L. Newbury, R. Jenkins, T. Ashdown, C. Dayan, S. Satchell, P. Corish, P. Cockwell, D. Fraser, and T. Bowen. 2018. ’Association of Elevated Urinary miR-126, miR-155, and miR-29b with Diabetic Kidney Disease’, Am J Pathol, 188: 1982-92.
- Bera, A.; Das, F.; Ghosh-Choudhury, N.; Mariappan, M.M.; Kasinath, B.S.; Choudhury, G.G. Reciprocal regulation of miR-214 and PTEN by high glucose regulates renal glomerular mesangial and proximal tubular epithelial cell hypertrophy and matrix expansion. Am. J. Physiol. Physiol. 2017, 313, C430–C447. [Google Scholar] [CrossRef]
- Bhatt, K.; Lanting, L.L.; Jia, Y.; Yadav, S.; Reddy, M.A.; Magilnick, N.; Boldin, M.; Natarajan, R. Anti-Inflammatory Role of MicroRNA-146a in the Pathogenesis of Diabetic Nephropathy. J. Am. Soc. Nephrol. 2016, 27, 2277–2288. [Google Scholar] [CrossRef]
- Bijkerk, R. R. G. de Bruin, C. van Solingen, J. M. van Gils, J. M. Duijs, E. P. van der Veer, T. J. Rabelink, B. D. Humphreys, and A. J. van Zonneveld. 2016. ’Silencing of microRNA-132 reduces renal fibrosis by selectively inhibiting myofibroblast proliferation’, Kidney Int, 89: 1268-80.
- Blijdorp, C.J.; Hartjes, T.A.; Wei, K.; van Heugten, M.H.; Bovée, D.M.; Budde, R.P.; van de Wetering, J.; Hoenderop, J.G.; van Royen, M.E.; Zietse, R.; et al. Nephron mass determines the excretion rate of urinary extracellular vesicles. J. Extracell. Vesicles 2022, 11, e12181. [Google Scholar] [CrossRef]
- Chen, H.-Y.; Zhong, X.; Huang, X.R.; Meng, X.-M.; You, Y.; Chung, A.C.; Lan, H.Y. MicroRNA-29b Inhibits Diabetic Nephropathy in db/db Mice. Mol. Ther. 2014, 22, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhao, L.; Xing, Y.; Lin, B. RETRACTED: Down-regulation of microRNA-21 reduces inflammation and podocyte apoptosis in diabetic nephropathy by relieving the repression of TIMP3 expression. Biomed. Pharmacother. 2018, 108, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Q.; Wang, X.X.; Yao, X.M.; Zhang, D.L.; Yang, X.F.; Tian, S.F.; Wang, N.S. Abated microRNA-195 expression protected mesangial cells from apoptosis in early diabetic renal injury in mice. J. Nephrol. 2011, 25, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhu, Q.; Cheng, L.; Wang, Y.; Li, M.; Yang, Q.; Hu, L.; Lou, D.; Li, J.; Dong, X.; et al. Exosome detection via the ultrafast-isolation system: EXODUS. Nat. Methods 2021, 18, 212–218. [Google Scholar] [CrossRef]
- Chorley, B. N. H. Ellinger-Ziegelbauer, M. Tackett, F. J. Simutis, A. H. Harrill, J. McDuffie, E. Atabakhsh, R. Nassirpour, L. O. Whiteley, J. F. Léonard, G. K. Carswell, E. Harpur, C. L. Chen, and J. C. Gautier. 2021. ’Urinary miRNA Biomarkers of Drug-Induced Kidney Injury and Their Site Specificity Within the Nephron’, Toxicol Sci, 180: 1-16.
- Conserva, F.; Barozzino, M.; Pesce, F.; Divella, C.; Oranger, A.; Papale, M.; Sallustio, F.; Simone, S.; Laviola, L.; Giorgino, F.; et al. Urinary miRNA-27b-3p and miRNA-1228-3p correlate with the progression of Kidney Fibrosis in Diabetic Nephropathy. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cui, L. M. Yu, and X. Cui. 2020. ’MiR-30c-5p/ROCK2 axis regulates cell proliferation, apoptosis and EMT via the PI3K/AKT signaling pathway in HG-induced HK-2 cells’, Open Life Sci, 15: 959-70.
- Dai, Y.; Cao, Y.; Köhler, J.; Lu, A.; Xu, S.; Wang, H. Unbiased RNA-Seq-driven identification and validation of reference genes for quantitative RT-PCR analyses of pooled cancer exosomes. BMC Genom. 2021, 22, 1–13. [Google Scholar] [CrossRef]
- De Freitas, R.C.C.; Hirata, R.D.C.; Hirata, M.H.; Aikawa, E. Circulating Extracellular Vesicles As Biomarkers and Drug Delivery Vehicles in Cardiovascular Diseases. Biomolecules 2021, 11, 388. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Reeves, W.B.; Awad, A.S. Pathophysiology of diabetic kidney disease: impact of SGLT2 inhibitors. Nat. Rev. Nephrol. 2021, 17, 319–334. [Google Scholar] [CrossRef]
- Delić, D.; Eisele, C.; Schmid, R.; Baum, P.; Wiech, F.; Gerl, M.; Zimdahl, H.; Pullen, S.S.; Urquhart, R. Urinary Exosomal miRNA Signature in Type II Diabetic Nephropathy Patients. PLOS ONE 2016, 11, e0150154. [Google Scholar] [CrossRef]
- Deshpande, S.D.; Putta, S.; Wang, M.; Lai, J.Y.; Bitzer, M.; Nelson, R.G.; Lanting, L.L.; Kato, M.; Natarajan, R. Transforming Growth Factor-β–Induced Cross Talk Between p53 and a MicroRNA in the Pathogenesis of Diabetic Nephropathy. Diabetes 2013, 62, 3151–3162. [Google Scholar] [CrossRef] [PubMed]
- Dey, N. F. Das, M. M. Mariappan, C. C. Mandal, N. Ghosh-Choudhury, B. S. Kasinath, and G. G. Choudhury. 2011. ’MicroRNA-21 orchestrates high glucose-induced signals to TOR complex 1, resulting in renal cell pathology in diabetes’, J Biol Chem, 286: 25586-603.
- Dieter, C.; Assmann, T.S.; Costa, A.R.; Canani, L.H.; de Souza, B.M.; Bauer, A.C.; Crispim, D. MiR-30e-5p and MiR-15a-5p Expressions in Plasma and Urine of Type 1 Diabetic Patients With Diabetic Kidney Disease. Front. Genet. 2019, 10, 563. [Google Scholar] [CrossRef] [PubMed]
- Dong, L. R. C. Zieren, K. Horie, C. J. Kim, E. Mallick, Y. Jing, M. Feng, M. D. Kuczler, J. Green, S. R. Amend, K. W. Witwer, T. M. de Reijke, Y. K. Cho, K. J. Pienta, and W. Xue. 2020. ’Comprehensive evaluation of methods for small extracellular vesicles separation from human plasma, urine and cell culture medium’, J Extracell Vesicles, 10: e12044.
- Duan, Y.; Chen, B.; Chen, F.; Yang, S.; Zhu, C.; Ma, Y.; Li, Y.; Shi, J. Exosomal microRNA-16-5p from human urine-derived stem cells ameliorates diabetic nephropathy through protection of podocyte. J. Cell. Mol. Med. 2019, 25, 10798–10813. [Google Scholar] [CrossRef]
- Dwivedi, O.P.; Barreiro, K.; Käräjämäki, A.; Valo, E.; Giri, A.K.; Prasad, R.B.; Das Roy, R.; Thorn, L.M.; Rannikko, A.; Holthöfer, H.; et al. Genome-wide mRNA profiling in urinary extracellular vesicles reveals stress gene signature for diabetic kidney disease. iScience 2023, 26, 106686. [Google Scholar] [CrossRef]
- Eissa, S.; Matboli, M.; Aboushahba, R.; Bekhet, M.M.; Soliman, Y. Urinary exosomal microRNA panel unravels novel biomarkers for diagnosis of type 2 diabetic kidney disease. J. Diabetes its Complicat. 2016, 30, 1585–1592. [Google Scholar] [CrossRef] [PubMed]
- Erdbrugger, U. and T. H. Le. 2016. ’Extracellular Vesicles in Renal Diseases: More than Novel Biomarkers?’, J Am Soc Nephrol, 27: 12-26.
- Erdbrügger, U. C. J. Blijdorp, I. V. Bijnsdorp, F. E. Borràs, D. Burger, B. Bussolati, J. B. Byrd, A. Clayton, J. W. Dear, J. M. Falcón-Pérez, C. Grange, A. F. Hill, H. Holthöfer, E. J. Hoorn, G. Jenster, C. R. Jimenez, K. Junker, J. Klein, M. A. Knepper, E. H. Koritzinsky, J. M. Luther, M. Lenassi, J. Leivo, I. Mertens, L. Musante, E. Oeyen, M. Puhka, M. E. van Royen, C. Sánchez, C. Soekmadji, V. Thongboonkerd, V. van Steijn, G. Verhaegh, J. P. Webber, K. Witwer, P. S. T. Yuen, L. Zheng, A. Llorente, and E. S. Martens-Uzunova. 2021. ’Urinary extracellular vesicles: A position paper by the Urine Task Force of the International Society for Extracellular Vesicles’, J Extracell Vesicles, 10: e12093.
- Fu, Y.; Zhang, Y.; Wang, Z.; Wang, L.; Wei, X.; Zhang, B.; Wen, Z.; Fang, H.; Pang, Q.; Yi, F. Regulation of NADPH Oxidase Activity Is Associated with miRNA-25-Mediated NOX4 Expression in Experimental Diabetic Nephropathy. Am. J. Nephrol. 2010, 32, 581–589. [Google Scholar] [CrossRef]
- Gao, Y. W. Xu, C. Guo, and T. Huang. 2022. ’GATA1 regulates the microRNA-328-3p/PIM1 axis via circular RNA ITGB1 to promote renal ischemia/reperfusion injury in HK-2 cells’, Int J Mol Med, 50.
- García-Flores, M. C. M. Sánchez-López, M. Ramírez-Calvo, A. Fernández-Serra, A. Marcilla, and J. A. López-Guerrero. 2021. ’Isolation and characterization of urine microvesicles from prostate cancer patients: different approaches, different visions’, BMC Urol, 21: 137.
- Ghai, V.; Wu, X.; Bheda-Malge, A.; Argyropoulos, C.P.; Bernardo, J.F.; Orchard, T.; Galas, D.; Wang, K. Genome-wide Profiling of Urinary Extracellular Vesicle microRNAs Associated With Diabetic Nephropathy in Type 1 Diabetes. Kidney Int. Rep. 2017, 3, 555–572. [Google Scholar] [CrossRef] [PubMed]
- Gorji-Bahri, G. N. Moradtabrizi, F. Vakhshiteh, and A. Hashemi. 2021. ’Validation of common reference genes stability in exosomal mRNA-isolated from liver and breast cancer cell lines’, Cell Biol Int, 45: 1098-110.
- Gouin, K. K. Peck, T. Antes, J. L. Johnson, C. Li, S. D. Vaturi, R. Middleton, G. de Couto, A. S. Walravens, L. Rodriguez-Borlado, R. R. Smith, L. Marbán, E. Marbán, and A. G. Ibrahim. 2017. ’A comprehensive method for identification of suitable reference genes in extracellular vesicles’, J Extracell Vesicles, 6: 1347019.
- Habuka, M. L. Fagerberg, B. M. Hallström, C. Kampf, K. Edlund, Å Sivertsson, T. Yamamoto, F. Pontén, M. Uhlén, and J. Odeberg. 2014. ’The kidney transcriptome and proteome defined by transcriptomics and antibody-based profiling’, PLoS One, 9: e116125.
- Han, X.; Li, Q.; Wang, C.; Li, Y. MicroRNA-204-3p Attenuates High Glucose-Induced MPC5 Podocytes Apoptosis by Targeting Braykinin B2 Receptor. Exp. Clin. Endocrinol. Diabetes 2018, 127, 387–395. [Google Scholar] [CrossRef]
- He, F.; Peng, F.; Xia, X.; Zhao, C.; Luo, Q.; Guan, W.; Li, Z.; Yu, X.; Huang, F. MiR-135a promotes renal fibrosis in diabetic nephropathy by regulating TRPC1. Diabetologia 2014, 57, 1726–1736. [Google Scholar] [CrossRef]
- He, X.; Zeng, X. LncRNA SNHG16 Aggravates High Glucose-Induced Podocytes Injury in Diabetic Nephropathy Through Targeting miR-106a and Thereby Up-Regulating KLF9. Diabetes, Metab. Syndr. Obesity: Targets Ther. 2020, 13, 3551–3560. [Google Scholar] [CrossRef]
- Hogan, M.C.; Lieske, J.C.; Lienczewski, C.C.; Nesbitt, L.L.; Wickman, L.T.; Heyer, C.M.; Harris, P.C.; Ward, C.J.; Sundsbak, J.L.; Manganelli, L.; et al. Strategy and rationale for urine collection protocols employed in the NEPTUNE study. BMC Nephrol. 2015, 16, 190. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y. Y. Li, Y. Wang, W. Li, and Z. Xiao. 2021. ’Screening and Analysis of Key Genes in miRNA-mRNA Regulatory Network of Membranous Nephropathy’, J Healthc Eng, 2021: 5331948.
- Hsu, Y.-C.; Chang, P.-J.; Ho, C.; Huang, Y.-T.; Shih, Y.-H.; Wang, C.-J.; Lin, C.-L. Protective effects of miR-29a on diabetic glomerular dysfunction by modulation of DKK1/Wnt/β-catenin signaling. Sci. Rep. 2016, 6, 30575. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liu, Y.; Li, L.; Su, B.; Yang, L.; Fan, W.; Yin, Q.; Chen, L.; Cui, T.; Zhang, J.; et al. Involvement of inflammation-related miR-155 and miR-146a in diabetic nephropathy: implications for glomerular endothelial injury. BMC Nephrol. 2014, 15, 142–142. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Guan, M.; Zheng, Z.; Zhang, Q.; Tang, C.; Xu, W.; Xiao, Z.; Wang, L.; Xue, Y. miRNAs in Urine Extracellular Vesicles as Predictors of Early-Stage Diabetic Nephropathy. J. Diabetes Res. 2016, 2016, 1–10. [Google Scholar] [CrossRef]
- Jiang, Y. W. Wang, Z. Y. Liu, Y. Xie, Y. Qian, and X. N. Cai. 2018. ’Overexpression of miR-130a-3p/301a-3p attenuates high glucose-induced MPC5 podocyte dysfunction through suppression of TNF-α signaling’, Exp Ther Med, 15: 1021-28.
- Jiang, Z.; Tang, Y.; Song, H.; Yang, M.; Li, B.; Ni, C. miRNA-342 suppresses renal interstitial fibrosis in diabetic nephropathy by targeting SOX6. Int. J. Mol. Med. 2019, 45, 45–52. [Google Scholar] [CrossRef]
- Jones, T.F.; Bekele, S.; O’dwyer, M.J.; Prowle, J.R. MicroRNAs in Acute Kidney Injury. Nephron 2018, 140, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Arce, L.; Wang, M.; Putta, S.; Lanting, L.; Natarajan, R. A microRNA circuit mediates transforming growth factor-β1 autoregulation in renal glomerular mesangial cells. Kidney Int. 2011, 80, 358–368. [Google Scholar] [CrossRef]
- Kato, M. S. Putta, M. Wang, H. Yuan, L. Lanting, I. Nair, A. Gunn, Y. Nakagawa, H. Shimano, I. Todorov, J. J. Rossi, and R. Natarajan. 2009. ’TGF-beta activates Akt kinase through a microRNA-dependent amplifying circuit targeting PTEN’, Nat Cell Biol, 11: 881-9.
- Kato, M. L. Wang, S. Putta, M. Wang, H. Yuan, G. Sun, L. Lanting, I. Todorov, J. J. Rossi, and R. Natarajan. 2010. ’Post-transcriptional up-regulation of Tsc-22 by Ybx1, a target of miR-216a, mediates TGF-{beta}-induced collagen expression in kidney cells’, J Biol Chem, 285: 34004-15.
- Kato, M.; Wang, M.; Chen, Z.; Bhatt, K.; Oh, H.J.; Lanting, L.; Deshpande, S.; Jia, Y.; Lai, J.Y.; O’connor, C.L.; et al. An endoplasmic reticulum stress-regulated lncRNA hosting a microRNA megacluster induces early features of diabetic nephropathy. Nat. Commun. 2016, 7, 12864. [Google Scholar] [CrossRef]
- Kato, M. J. Zhang, M. Wang, L. Lanting, H. Yuan, J. J. Rossi, and R. Natarajan. 2007. ’MicroRNA-192 in diabetic kidney glomeruli and its function in TGF-beta-induced collagen expression via inhibition of E-box repressors’, Proc Natl Acad Sci U S A, 104: 3432-7.
- Keller, A.; Gröger, L.; Tschernig, T.; Solomon, J.; Laham, O.; Schaum, N.; Wagner, V.; Kern, F.; Schmartz, G.P.; Li, Y.; et al. miRNATissueAtlas2: an update to the human miRNA tissue atlas. Nucleic Acids Res. 2021, 50, D211–D221. [Google Scholar] [CrossRef]
- Koga, K.; Yokoi, H.; Mori, K.; Kasahara, M.; Kuwabara, T.; Imamaki, H.; Ishii, A.; Mori, K.P.; Kato, Y.; Ohno, S.; et al. MicroRNA-26a inhibits TGF-β-induced extracellular matrix protein expression in podocytes by targeting CTGF and is downregulated in diabetic nephropathy. Diabetologia 2015, 58, 2169–2180. [Google Scholar] [CrossRef]
- Kolde, Raivo. 2019. ’pheatmap: Pretty Heatmaps. ’, R package version 1.0.12. Available online: https://CRAN.R-project.org/package=pheatmap.
- Kozera, B. , and M. Rapacz. 2013. ’Reference genes in real-time PCR’, J Appl Genet, 54: 391-406.
- Krupa, A.; Jenkins, R.; Luo, D.D.; Lewis, A.; Phillips, A.; Fraser, D. Loss of MicroRNA-192 Promotes Fibrogenesis in Diabetic Nephropathy. J. Am. Soc. Nephrol. 2010, 21, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Kölling, M.; Kaucsar, T.; Schauerte, C.; Hübner, A.; Dettling, A.; Park, J.-K.; Busch, M.; Wulff, X.; Meier, M.; Scherf, K.; et al. Therapeutic miR-21 Silencing Ameliorates Diabetic Kidney Disease in Mice. Mol. Ther. 2016, 25, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Lai, J. Y. J. Luo, C. O’Connor, X. Jing, V. Nair, W. Ju, A. Randolph, I. Z. Ben-Dov, R. N. Matar, D. Briskin, J. Zavadil, R. G. Nelson, T. Tuschl, F. C. Brosius, 3rd, M. Kretzler, and M. Bitzer. 2015. ’MicroRNA-21 in glomerular injury’, J Am Soc Nephrol, 26: 805-16.
- Lee, H.W.; Khan, S.Q.; Khaliqdina, S.; Altintas, M.M.; Grahammer, F.; Zhao, J.L.; Koh, K.H.; Tardi, N.J.; Faridi, M.H.; Geraghty, T.; et al. Absence of miR-146a in Podocytes Increases Risk of Diabetic Glomerulopathy via Up-regulation of ErbB4 and Notch-1. J. Biol. Chem. 2017, 292, 732–747. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Lu, Z.; Jia, J.; Zheng, Z.; Lin, S. MiR-124 is Related to Podocytic Adhesive Capacity Damage in STZ-Induced Uninephrectomized Diabetic Rats. Kidney Blood Press. Res. 2013, 37, 422–431. [Google Scholar] [CrossRef]
- Li, H.; Zhu, X.; Zhang, J.; Shi, J. MicroRNA-25 inhibits high glucose-induced apoptosis in renal tubular epithelial cells via PTEN/AKT pathway. Biomed. Pharmacother. 2017, 96, 471–479. [Google Scholar] [CrossRef]
- Li, N. L. J. Wang, W. L. Xu, S. Liu, and J. Y. Yu. 2019. ’MicroRNA-379-5p suppresses renal fibrosis by regulating the LIN28/let-7 axis in diabetic nephropathy’, Int J Mol Med, 44: 1619-28.
- Li, X.; Dong, Z.-Q.; Chang, H.; Zhou, H.-B.; Wang, J.; Yang, Z.-J.; Qiu, M.; Bai, W.-F.; Shi, S.-L. Screening and identification of key microRNAs and regulatory pathways associated with the renal fibrosis process. Mol. Omics 2022, 18, 520–533. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, Y.-H.; Li, F.; Yang, T.; Lu, Y.W.; Geng, Y.-J. microRNA-221 regulates high glucose-induced endothelial dysfunction. Biochem. Biophys. Res. Commun. 2009, 381, 81–83. [Google Scholar] [CrossRef]
- Liang, Q. J. He, Q. Yang, Q. Zhang, and Y. Xu. 2022. ’MicroRNA-335-5p alleviates inflammatory response, airway fibrosis, and autophagy in childhood asthma through targeted regulation of autophagy related 5’, Bioengineered, 13: 1791-801.
- Lin, C.-L.; Lee, P.-H.; Hsu, Y.-C.; Lei, C.-C.; Ko, J.-Y.; Chuang, P.-C.; Huang, Y.-T.; Wang, S.-Y.; Wu, S.-L.; Chen, Y.-S.; et al. MicroRNA-29a Promotion of Nephrin Acetylation Ameliorates Hyperglycemia-Induced Podocyte Dysfunction. J. Am. Soc. Nephrol. 2014, 25, 1698–1709. [Google Scholar] [CrossRef]
- Liu, H.; Wang, X.; Liu, S.; Li, H.; Yuan, X.; Feng, B.; Bai, H.; Zhao, B.; Chu, Y.; Li, H. Effects and mechanism of miR-23b on glucose-mediated epithelial-to-mesenchymal transition in diabetic nephropathy. Int. J. Biochem. Cell Biol. 2016, 70, 149–160. [Google Scholar] [CrossRef]
- Liu, W.-T.; Peng, F.-F.; Li, H.-Y.; Chen, X.-W.; Gong, W.-Q.; Chen, W.-J.; Chen, Y.-H.; Li, P.-L.; Li, S.-T.; Xu, Z.-Z.; et al. Metadherin facilitates podocyte apoptosis in diabetic nephropathy. Cell Death Dis. 2016, 7, e2477–e2477. [Google Scholar] [CrossRef]
- Liu, X.-D.; Zhang, L.-Y.; Zhu, T.-C.; Zhang, R.-F.; Wang, S.-L.; Bao, Y. Overexpression of miR-34c inhibits high glucose-induced apoptosis in podocytes by targeting Notch signaling pathways. Int J Clin Exp Pathol 2015, 8, 4525–34. [Google Scholar] [PubMed]
- Liu, Y.; Gao, G.; Yang, C.; Zhou, K.; Shen, B.; Liang, H.; Jiang, X. Stability of miR-126 in Urine and Its Potential as a Biomarker for Renal Endothelial Injury with Diabetic Nephropathy. Int. J. Endocrinol. 2014, 2014, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, H.; Liu, J.; Han, P.; Li, X.; Bai, H.; Zhang, C.; Sun, X.; Teng, Y.; Zhang, Y.; et al. Variations in MicroRNA-25 Expression Influence the Severity of Diabetic Kidney Disease. J. Am. Soc. Nephrol. 2017, 28, 3627–3638. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, B.; Liu, Y.; Chen, S.; Yang, J.; Liu, J.; Sun, G.; Bei, W.-J.; Wang, K.; Chen, Z.; et al. MicroRNA expression profile by next-generation sequencing in a novel rat model of contrast-induced acute kidney injury. Ann. Transl. Med. 2019, 7, 178–178. [Google Scholar] [CrossRef]
- Long, J.; Wang, Y.; Wang, W.; Chang, B.H.J.; Danesh, F.R. Identification of MicroRNA-93 as a Novel Regulator of Vascular Endothelial Growth Factor in Hyperglycemic Conditions. J. Biol. Chem. 2010, 285, 23457–23465. [Google Scholar] [CrossRef] [PubMed]
- MicroRNA-29c is a signature microRNA under high glucose conditions that targets Sprouty homolog 1, and its in vivo knockdown prevents progression of diabetic nephropathy’, J Biol Chem, 286: 11837-48.
- López-Guerrero, José A., Mar Valés-Gómez, Francesc E. Borrás, Juan Manuel Falcón-Pérez, María J. Vicent, and María Yáñez-Mó. 2023. ’Standardising the preanalytical reporting of biospecimens to improve reproducibility in extracellular vesicle research – A GEIVEX study’, Journal of Extracellular Biology, 2: e76.
- Love, M. I. W. Huber, and S. Anders. 2014. ’Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2’, Genome Biol, 15: 550.
- Margolis, E.; Brown, G.; Partin, A.; Carter, B.; McKiernan, J.; Tutrone, R.; Torkler, P.; Fischer, C.; Tadigotla, V.; Noerholm, M.; et al. Predicting high-grade prostate cancer at initial biopsy: clinical performance of the ExoDx (EPI) Prostate Intelliscore test in three independent prospective studies. Prostate Cancer Prostatic Dis. 2021, 25, 296–301. [Google Scholar] [CrossRef]
- Mateescu, B. E. J. Kowal, B. W. van Balkom, S. Bartel, S. N. Bhattacharyya, E. I. Buzás, A. H. Buck, P. de Candia, F. W. Chow, S. Das, T. A. Driedonks, L. Fernández-Messina, F. Haderk, A. F. Hill, J. C. Jones, K. R. Van Keuren-Jensen, C. P. Lai, C. Lässer, I. D. Liegro, T. R. Lunavat, M. J. Lorenowicz, S. L. Maas, I. Mäger, M. Mittelbrunn, S. Momma, K. Mukherjee, M. Nawaz, D. M. Pegtel, M. W. Pfaffl, R. M. Schiffelers, H. Tahara, C. Théry, J. P. Tosar, M. H. Wauben, K. W. Witwer, and E. N. Nolte-’t Hoen. 2017. ’Obstacles and opportunities in the functional analysis of extracellular vesicle RNA - an ISEV position paper’, J Extracell Vesicles, 6: 1286095.
- McClelland, A.D.; Herman-Edelstein, M.; Komers, R.; Jha, J.C.; Winbanks, C.E.; Hagiwara, S.; Gregorevic, P.; Kantharidis, P.; Cooper, M.E. miR-21 promotes renal fibrosis in diabetic nephropathy by targeting PTEN and SMAD7. Clin. Sci. 2015, 129, 1237–1249. [Google Scholar] [CrossRef]
- Miller, D.; Eagle-Hemming, B.; Sheikh, S.; Joel-David, L.; Adebayo, A.; Lai, F.Y.; Roman, M.; Kumar, T.; Aujla, H.; Murphy, G.J.; et al. Urinary extracellular vesicles and micro-RNA as markers of acute kidney injury after cardiac surgery. Sci. Rep. 2022, 12, 1–11. [Google Scholar] [CrossRef]
- Min, K.-H.; Yang, W.-M.; Lee, W. Saturated fatty acids-induced miR-424–5p aggravates insulin resistance via targeting insulin receptor in hepatocytes. Biochem. Biophys. Res. Commun. 2018, 503, 1587–1593. [Google Scholar] [CrossRef]
- Miranda, K.C.; Bond, D.T.; Levin, J.Z.; Adiconis, X.; Sivachenko, A.; Russ, C.; Brown, D.; Nusbaum, C.; Russo, L.M. Massively Parallel Sequencing of Human Urinary Exosome/Microvesicle RNA Reveals a Predominance of Non-Coding RNA. PLOS ONE 2014, 9, e96094. [Google Scholar] [CrossRef]
- Miranda, K. C. D. T. Bond, M. McKee, J. Skog, T. G. Păunescu, N. Da Silva, D. Brown, and L. M. Russo. 2010. ’Nucleic acids within urinary exosomes/microvesicles are potential biomarkers for renal disease’, Kidney Int, 78: 191-9.
- Mishra, A.; Ayasolla, K.; Kumar, V.; Lan, X.; Vashistha, H.; Aslam, R.; Hussain, A.; Chowdhary, S.; Shoshtari, S.M.; Paliwal, N.; et al. Modulation of apolipoprotein L1-microRNA-193a axis prevents podocyte dedifferentiation in high-glucose milieu. Am. J. Physiol. Physiol. 2018, 314, F832–F843. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Singh, R.S.; Kumari, M.; Garg, D.; Upadhyay, A.; Ecelbarger, C.M.; Tripathy, S.; Tiwari, S. Urinary Exosomal microRNA-451-5p Is a Potential Early Biomarker of Diabetic Nephropathy in Rats. PLOS ONE 2016, 11, e0154055–e0154055. [Google Scholar] [CrossRef] [PubMed]
- Mussack, V.; Wittmann, G.; Pfaffl, M.W. Comparing small urinary extracellular vesicle purification methods with a view to RNA sequencing—Enabling robust and non-invasive biomarker research. Biomol. Detect. Quantif. 2019, 17, 100089. [Google Scholar] [CrossRef] [PubMed]
- Müller-Deile, J.; Dannenberg, J.; Schroder, P.; Lin, M.-H.; Miner, J.H.; Chen, R.; Bräsen, J.-H.; Thum, T.; Nyström, J.; Staggs, L.B.; et al. Podocytes regulate the glomerular basement membrane protein nephronectin by means of miR-378a-3p in glomerular diseases. Kidney Int. 2017, 92, 836–849. [Google Scholar] [CrossRef]
- Nieuwland, R.; Siljander, P.R.-M.; Falcón-Pérez, J.M.; Witwer, K.W. Reproducibility of extracellular vesicle research. Eur. J. Cell Biol. 2022, 101, 151226. [Google Scholar] [CrossRef]
- Oh, H. J. M. Kato, S. Deshpande, E. Zhang, S. Das, L. Lanting, M. Wang, and R. Natarajan. 2016. ’Inhibition of the processing of miR-25 by HIPK2-Phosphorylated-MeCP2 induces NOX4 in early diabetic nephropathy’, Sci Rep, 6: 38789.
- Oosthuyzen, W.; Sime, N.E.L.; Ivy, J.R.; Turtle, E.J.; Street, J.M.; Pound, J.; Bath, L.E.; Webb, D.J.; Gregory, C.D.; Bailey, M.; et al. Quantification of human urinary exosomes by nanoparticle tracking analysis. J. Physiol. 2013, 591, 5833–5842. [Google Scholar] [CrossRef]
- Park, J.T.; Kato, M.; Yuan, H.; Castro, N.; Lanting, L.; Wang, M.; Natarajan, R. FOG2 Protein Down-regulation by Transforming Growth Factor-β1-induced MicroRNA-200b/c Leads to Akt Kinase Activation and Glomerular Mesangial Hypertrophy Related to Diabetic Nephropathy. J. Biol. Chem. 2013, 288, 22469–22480. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, O.-H.; Lee, K.; Park, I.B.; Kim, N.H.; Moon, S.; Im, J.; Sharma, S.P.; Oh, B.-C.; Nam, S.; et al. Plasma and urinary extracellular vesicle microRNAs and their related pathways in diabetic kidney disease. Genomics 2022, 114, 110407. [Google Scholar] [CrossRef]
- Park, S.; Lee, K.; Park, I.B.; Kim, N.H.; Cho, S.; Rhee, W.J.; Oh, Y.; Choi, J.; Nam, S.; Lee, D.H. The profiles of microRNAs from urinary extracellular vesicles (EVs) prepared by various isolation methods and their correlation with serum EV microRNAs. Diabetes Res. Clin. Pr. 2020, 160, 108010. [Google Scholar] [CrossRef]
- Pavkovic, M.; Vaidya, V.S. MicroRNAs and drug-induced kidney injury. Pharmacol. Ther. 2016, 163, 48–57. [Google Scholar] [CrossRef]
- Pruneda, J. N. F. D. Smith, A. Daurie, D. L. Swaney, J. Villén, J. D. Scott, A. W. Stadnyk, I. Le Trong, R. E. Stenkamp, R. E. Klevit, J. R. Rohde, and P. S. Brzovic. 2014. ’E2~Ub conjugates regulate the kinase activity of Shigella effector OspG during pathogenesis’, Embo j, 33: 437-49.
- Puhka, M. L. Thierens, D. Nicorici, T. Forsman, T. Mirtti, T. Af Hällström, E. Serkkola, and A. Rannikko. 2022. ’Exploration of Extracellular Vesicle miRNAs, Targeted mRNAs and Pathways in Prostate Cancer: Relation to Disease Status and Progression’, Cancers (Basel), 14.
- Qian, X.; Tan, J.; Liu, L.; Chen, S.; You, N.; Yong, H.; Pan, M.; You, Q.; Ding, D.; Lu, Y. MicroRNA-134-5p promotes high glucose-induced podocyte apoptosis by targeting bcl-2. Am J Transl Res 2018, 10, 989–997. [Google Scholar] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [PubMed]
- Royo, F.; Théry, C.; Falcón-Pérez, J.M.; Nieuwland, R.; Witwer, K.W. Methods for Separation and Characterization of Extracellular Vesicles: Results of a Worldwide Survey Performed by the ISEV Rigor and Standardization Subcommittee. Cells 2020, 9, 1955. [Google Scholar] [CrossRef] [PubMed]
- Sáenz-Cuesta, M.; Arbelaiz, A.; Oregi, A.; Irizar, H.; Osorio-Querejeta, I.; Muñoz-Culla, M.; Banales, J.M.; Falcón-Pérez, J.M.; Olascoaga, J.; Otaegui, D. Methods for extracellular vesicles isolation in a hospital setting. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef]
- Scian, M.J.; Maluf, D.G.; David, K.G.; Archer, K.J.; Suh, J.L.; Wolen, A.R.; Mba, M.U.; Massey, H.D.; King, A.L.; Gehr, T.; et al. MicroRNA Profiles in Allograft Tissues and Paired Urines Associate With Chronic Allograft Dysfunction With IF/TA. Am. J. Transplant. 2011, 11, 2110–2122. [Google Scholar] [CrossRef]
- Sessa, F.; Salerno, M.; Bertozzi, G.; Cipolloni, L.; Messina, G.; Aromatario, M.; Polo, L.; Turillazzi, E.; Pomara, C. miRNAs as Novel Biomarkers of Chronic Kidney Injury in Anabolic-Androgenic Steroid Users: An Experimental Study. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef]
- Shin, Y. D. Y. Kim, J. Y. Ko, Y. M. Woo, and J. H. Park. 2018. ’Regulation of KLF12 by microRNA-20b and microRNA-106a in cystogenesis’, Faseb j, 32: 3574-82.
- Singh, A. D. S. Patnam, R. Koyyada, R. Samal, S. B. Alvi, G. Satyanaryana, R. Andrews, A. K. Panigrahi, A. K. Rengan, S. S. Mudigonda, S. Maitra, and M. V. Sasidhar. 2022. ’Identifying stable reference genes in polyethene glycol precipitated urinary extracellular vesicles for RT-qPCR-based gene expression studies in renal graft dysfunction patients’, Transpl Immunol, 75: 101715.
- Song, L.; Feng, S.; Yu, H.; Shi, S. Dexmedetomidine Protects Against Kidney Fibrosis in Diabetic Mice by Targeting miR-101-3p-Mediated EndMT. Dose-Response 2022, 20. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Yeri, A.; Cheah, P.S.; Chung, A.; Danielson, K.; De Hoff, P.; Filant, J.; Laurent, C.D.; Laurent, L.D.; Magee, R.; et al. Small RNA Sequencing across Diverse Biofluids Identifies Optimal Methods for exRNA Isolation. Cell 2019, 177, 446–462. [Google Scholar] [CrossRef]
- Su, J. J. Ren, H. Chen, and B. Liu. 2020. ’MicroRNA-140-5p ameliorates the high glucose-induced apoptosis and inflammation through suppressing TLR4/NF-κB signaling pathway in human renal tubular epithelial cells’, Biosci Rep, 40.
- Sun, J.; Li, Z.P.; Zhang, R.Q.; Zhang, H.M. Repression of miR-217 protects against high glucose-induced podocyte injury and insulin resistance by restoring PTEN-mediated autophagy pathway. Biochem. Biophys. Res. Commun. 2017, 483, 318–324. [Google Scholar] [CrossRef]
- Sun, Y. R. Peng, H. Peng, H. Liu, L. Wen, T. Wu, H. Yi, A. Li, and Z. Zhang. 2016. ’miR-451 suppresses the NF-kappaB-mediated proinflammatory molecules expression through inhibiting LMP7 in diabetic nephropathy’, Mol Cell Endocrinol, 433: 75-86.
- Sun, Z.; Ma, Y.; Chen, F.; Wang, S.; Chen, B.; Shi, J. miR-133b and miR-199b knockdown attenuate TGF-β1-induced epithelial to mesenchymal transition and renal fibrosis by targeting SIRT1 in diabetic nephropathy. Eur. J. Pharmacol. 2018, 837, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D. A. L. Gable, D. Lyon, A. Junge, S. Wyder, J. Huerta-Cepas, M. Simonovic, N. T. Doncheva, J. H. Morris, P. Bork, L. J. Jensen, and C. V. Mering. 2019. ’STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets’, Nucleic Acids Res, 47: D607-d13.
- Toth-Manikowski, S.; Atta, M.G. Diabetic Kidney Disease: Pathophysiology and Therapeutic Targets. J. Diabetes Res. 2015, 2015, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Trevisani, F.; Ghidini, M.; Larcher, A.; Lampis, A.; Lote, H.; Manunta, P.; Alibrandi, M.T.S.; Zagato, L.; Citterio, L.; Dell’Antonio, G.; et al. MicroRNA 193b-3p as a predictive biomarker of chronic kidney disease in patients undergoing radical nephrectomy for renal cell carcinoma. Br. J. Cancer 2016, 115, 1343–1350. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.-C.; Kuo, M.-C.; Hung, W.-W.; Wu, L.-Y.; Wu, P.-H.; Chang, W.-A.; Kuo, P.-L.; Hsu, Y.-L. High Glucose Induces Mesangial Cell Apoptosis through miR-15b-5p and Promotes Diabetic Nephropathy by Extracellular Vesicle Delivery. Mol. Ther. 2020, 28, 963–974. [Google Scholar] [CrossRef]
- Tuttle, K.R.; Agarwal, R.; Alpers, C.E.; Bakris, G.L.; Brosius, F.C.; Kolkhof, P.; Uribarri, J. Molecular mechanisms and therapeutic targets for diabetic kidney disease. Kidney Int. 2022, 102, 248–260. [Google Scholar] [CrossRef] [PubMed]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Martin, M.-J.; Orchard, S.; Magrane, M.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bye-A-Jee, H.; et al.; The UniProt Consortium UniProt: the Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2022, 51, D523–D531. [Google Scholar] [CrossRef]
- Vago, R.; Radano, G.; Zocco, D.; Zarovni, N. Urine stabilization and normalization strategies favor unbiased analysis of urinary EV content. Sci. Rep. 2022, 12, 1–20. [Google Scholar] [CrossRef]
- EV-TRACK Consortium; Van Deun, J.; Mestdagh, P.; Agostinis, P.; Akay, Ö.; Anand, S.; Anckaert, J.; Martinez, Z.A.; Baetens, T.; Beghein, E.; et al. EV-TRACK: Transparent reporting and centralizing knowledge in extracellular vesicle research. Nat. Methods 2017, 14, 228–232. [Google Scholar] [CrossRef]
- van Royen, M. E. C. Soekmadji, C. Grange, J. P. Webber, T. Tertel, M. Droste, A. Buescher, B. Giebel, G. W. Jenster, A. Llorente, C. J. Blijdorp, D. Burger, U. Erdbrügger, and E. S. Martens-Uzunova. 2023. ’The quick reference card "Storage of urinary EVs" - A practical guideline tool for research and clinical laboratories’, J Extracell Vesicles, 12: e12286.
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef]
- Wan, R.J.; Li, Y.H. MicroRNA-146a/NAPDH oxidase4 decreases reactive oxygen species generation and inflammation in a diabetic nephropathy model. Mol. Med. Rep. 2018, 17, 4759–4766. [Google Scholar] [CrossRef] [PubMed]
- Wang, B. M. Herman-Edelstein, P. Koh, W. Burns, K. Jandeleit-Dahm, A. Watson, M. Saleem, G. J. Goodall, S. M. Twigg, M. E. Cooper, and P. Kantharidis. 2010. ’E-cadherin expression is regulated by miR-192/215 by a mechanism that is independent of the profibrotic effects of transforming growth factor-beta’, Diabetes, 59: 1794-802.
- Wang, B.; Jha, J.C.; Hagiwara, S.; McClelland, A.D.; Jandeleit-Dahm, K.; Thomas, M.C.; Cooper, M.E.; Kantharidis, P. Transforming growth factor-β1-mediated renal fibrosis is dependent on the regulation of transforming growth factor receptor 1 expression by let-7b. Kidney Int. 2014, 85, 352–361. [Google Scholar] [CrossRef]
- Wang, B.; Koh, P.; Winbanks, C.; Coughlan, M.T.; McClelland, A.; Watson, A.; Jandeleit-Dahm, K.; Burns, W.C.; Thomas, M.C.; Cooper, M.E.; et al. miR-200a Prevents Renal Fibrogenesis Through Repression of TGF-β2 Expression. Diabetes 2010, 60, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Komers, R.; Carew, R.; Winbanks, C.E.; Xu, B.; Herman-Edelstein, M.; Koh, P.; Thomas, M.; Jandeleit-Dahm, K.; Gregorevic, P.; et al. Suppression of microRNA-29 Expression by TGF-β1 Promotes Collagen Expression and Renal Fibrosis. J. Am. Soc. Nephrol. 2012, 23, 252–265. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, F.; Tian, Q.; Sheng, K. CircVMA21 ameliorates lipopolysaccharide (LPS)-induced HK-2 cell injury depending on the regulation of miR-7-5p/PPARA. Autoimmunity 2021, 55, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Kwan, B.C.-H.; Lai, F.M.-M.; Chow, K.-M.; Li, P.K.-T.; Szeto, C.-C. Urinary sediment miRNA levels in adult nephrotic syndrome. Clin. Chim. Acta 2013, 418, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Duan, L.; Tian, L.; Liu, J.; Wang, S.; Gao, Y.; Yang, J. Serum miR-21 may be a Potential Diagnostic Biomarker for Diabetic Nephropathy. Exp. Clin. Endocrinol. Diabetes 2015, 124, 417–423. [Google Scholar] [CrossRef]
- Wang, J.-Y.; Gao, Y.-B.; Zhang, N.; Zou, D.-W.; Wang, P.; Zhu, Z.-Y.; Li, J.-Y.; Zhou, S.-N.; Wang, S.-C.; Wang, Y.-Y.; et al. miR-21 overexpression enhances TGF-β1-induced epithelial-to-mesenchymal transition by target smad7 and aggravates renal damage in diabetic nephropathy. Mol. Cell. Endocrinol. 2014, 392, 163–172. [Google Scholar] [CrossRef]
- Wang, L.; Li, H. MiR-770-5p facilitates podocyte apoptosis and inflammation in diabetic nephropathy by targeting TIMP3. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Y.; Minto, A.W.; Wang, J.; Shi, Q.; Li, X.; Quigg, R.J. MicroRNA-377 is up-regulated and can lead to increased fibronectin production in diabetic nephropathy. FASEB J. 2008, 22, 4126–4135. [Google Scholar] [CrossRef]
- Wang, S. X. Wen, X. R. Han, Y. J. Wang, M. Shen, S. H. Fan, J. Zhuang, Z. F. Zhang, Q. Shan, M. Q. Li, B. Hu, C. H. Sun, D. M. Wu, J. Lu, and Y. L. Zheng. 2018. ’Repression of microRNA-382 inhibits glomerular mesangial cell proliferation and extracellular matrix accumulation via FoxO1 in mice with diabetic nephropathy’, Cell Prolif, 51: e12462.
- Wang, X.; Lin, B.; Nie, L.; Li, P. microRNA-20b contributes to high glucose-induced podocyte apoptosis by targeting SIRT7. Mol. Med. Rep. 2017, 16, 5667–5674. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, E.; Wang, Y.; Jiang, Z.; Gui, D.; Cheng, D.; Chen, T.; Wang, N. MiR-196a Regulates High Glucose-Induced Mesangial Cell Hypertrophy by Targeting p27kip1. JALA: J. Assoc. Lab. Autom. 2015, 20, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, E.; Wang, Y.; Li, J.; Cheng, D.; Chen, Y.; Gui, D.; Wang, N. Cross talk between miR-214 and PTEN attenuates glomerular hypertrophy under diabetic conditions. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Zhang, L.; Bai, L.; Chen, S.; Wu, H.; Sun, L.; Wang, X. miR-30b-5p modulate renal epithelial-mesenchymal transition in diabetic nephropathy by directly targeting SNAI1. Biochem. Biophys. Res. Commun. 2021, 535, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. Z. J. Zheng, Y. J. Jia, Y. L. Yang, and Y. M. Xue. 2018. ’Role of p53/miR-155-5p/sirt1 loop in renal tubular injury of diabetic kidney disease’, J Transl Med, 16: 146.
- Wei, B. Y. S. Liu, and H. X. Guan. 2020. ’MicroRNA-145-5p attenuates high glucose-induced apoptosis by targeting the Notch signaling pathway in podocytes’, Exp Ther Med, 19: 1915-24.
- Wickham, H. 2016. ggplot2: Elegant Graphics for Data Analysis (Springer-Verlag New York).
- Wickham, H. Reshaping Data with thereshapePackage. J. Stat. Softw. 2007, 21, 1–20. [Google Scholar] [CrossRef]
- Wu, L. Q. Wang, F. Guo, X. Ma, H. Ji, F. Liu, Y. Zhao, and G. Qin. 2016. ’MicroRNA-27a Induces Mesangial Cell Injury by Targeting of PPARγ, and its In Vivo Knockdown Prevents Progression of Diabetic Nephropathy’, Sci Rep, 6: 26072.
- Xie, Y.; Jia, Y.; Cuihua, X.; Hu, F.; Xue, M.; Xue, Y. Urinary Exosomal MicroRNA Profiling in Incipient Type 2 Diabetic Kidney Disease. J. Diabetes Res. 2017, 2017, 1–10. [Google Scholar] [CrossRef]
- Xu, H.; Sun, F.; Li, X.; Sun, L. Down-regulation of miR-23a inhibits high glucose-induced EMT and renal fibrogenesis by up-regulation of SnoN. Hum. Cell 2018, 31, 22–32. [Google Scholar] [CrossRef]
- Xu, K.; Liu, Q.; Wu, K.; Liu, L.; Zhao, M.; Yang, H.; Wang, X.; Wang, W. Extracellular vesicles as potential biomarkers and therapeutic approaches in autoimmune diseases. J. Transl. Med. 2020, 18, 1–8. [Google Scholar] [CrossRef]
- Xu, P.; Guan, M.-P.; Bi, J.-G.; Wang, D.; Zheng, Z.-J.; Xue, Y.-M. High glucose down-regulates microRNA-181a-5p to increase pro-fibrotic gene expression by targeting early growth response factor 1 in HK-2 cells. Cell. Signal. 2017, 31, 96–104. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, J.; Fan, L.; He, X. miR-423-5p suppresses high-glucose-induced podocyte injury by targeting Nox4. Biochem. Biophys. Res. Commun. 2018, 505, 339–345. [Google Scholar] [CrossRef]
- Xue, M.; Li, Y.; Hu, F.; Jia, Y.-J.; Zheng, Z.-J.; Wang, L.; Xue, Y.-M. High glucose up-regulates microRNA-34a-5p to aggravate fibrosis by targeting SIRT1 in HK-2 cells. Biochem. Biophys. Res. Commun. 2018, 498, 38–44. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Q.; Li, S. MicroRNA-218 promotes high glucose-induced apoptosis in podocytes by targeting heme oxygenase-1. Biochem. Biophys. Res. Commun. 2016, 471, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Guo, Z.; Dong, J.; Sheng, S.; Wang, Y.; Yu, L.; Wang, H.; Tang, L. miR-374a Regulates Inflammatory Response in Diabetic Nephropathy by Targeting MCP-1 Expression. Front. Pharmacol. 2018, 9, 900. [Google Scholar] [CrossRef]
- Yang, Z. Y. Y. Wang, Q. Liu, and M. Wu. 2020. ’microRNA cluster MC-let-7a-1~let-7d promotes autophagy and apoptosis of glioma cells by down-regulating STAT3’, CNS Neurosci Ther, 26: 319-31.
- Yao, T.; Zha, D.; Gao, P.; Shui, H.; Wu, X. MiR-874 alleviates renal injury and inflammatory response in diabetic nephropathy through targeting toll-like receptor-4. J. Cell. Physiol. 2018, 234, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Yekula, A.; Muralidharan, K.; Kang, K.M.; Wang, L.; Balaj, L.; Carter, B.S. From laboratory to clinic: Translation of extracellular vesicle based cancer biomarkers. Methods 2020, 177, 58–66. [Google Scholar] [CrossRef]
- Yu, L.; Gu, T.; Shi, E.; Wang, Y.; Fang, Q.; Wang, C. Dysregulation of renal microRNA expression after deep hypothermic circulatory arrest in rats. Eur. J. Cardio-Thoracic Surg. 2016, 49, 1725–1731. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Jia, Y.-Y.; Wang, M.; Mu, L.; Li, H.-J. PTGER3 and MMP-2 play potential roles in diabetic nephropathy via competing endogenous RNA mechanisms. BMC Nephrol. 2021, 22, 1–11. [Google Scholar] [CrossRef]
- Zanchi, C.; Macconi, D.; Trionfini, P.; Tomasoni, S.; Rottoli, D.; Locatelli, M.; Rudnicki, M.; Vandesompele, J.; Mestdagh, P.; Remuzzi, G.; et al. MicroRNA-184 is a downstream effector of albuminuria driving renal fibrosis in rats with diabetic nephropathy. Diabetologia 2017, 60, 1114–1125. [Google Scholar] [CrossRef]
- Zang, J.; Maxwell, A.P.; Simpson, D.A.; McKay, G.J. Differential Expression of Urinary Exosomal MicroRNAs miR-21-5p and miR-30b-5p in Individuals with Diabetic Kidney Disease. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Zha, F.; Bai, L.; Tang, B.; Li, J.; Wang, Y.; Zheng, P.; Ji, T.; Bai, S. MicroRNA-503 contributes to podocyte injury via targeting E2F3 in diabetic nephropathy. J. Cell. Biochem. 2019, 120, 12574–12581. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, Y.; Cai, R.; Jin, J.; He, Q. Differential Expression of Urinary Exosomal Small RNAs in Idiopathic Membranous Nephropathy. BioMed Res. Int. 2020, 2020, 1–12. [Google Scholar] [CrossRef]
- Zhang, L.; He, S.; Guo, S.; Xie, W.; Xin, R.; Yu, H.; Yang, F.; Qiu, J.; Zhang, D.; Zhou, S.; et al. Down-regulation of miR-34a alleviates mesangial proliferation in vitro and glomerular hypertrophy in early diabetic nephropathy mice by targeting GAS1. J. Diabetes Complicat. 2014, 28, 259–264. [Google Scholar] [CrossRef]
- Zhang, R. L. Qin, and J. Shi. 2020. ’MicroRNA-199a-3p suppresses high glucose-induced apoptosis and inflammation by regulating the IKKβ/NF-κB signaling pathway in renal tubular epithelial cells’, Int J Mol Med, 46: 2161-71.
- Zhang, W.; Zhou, X.; Zhang, H.; Yao, Q.; Liu, Y.; Dong, Z. Extracellular vesicles in diagnosis and therapy of kidney diseases. Am. J. Physiol. Physiol. 2016, 311, F844–F851. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, S.; Wu, D.; Liu, X.; Shi, M.; Wang, Y.; Zhang, F.; Ding, J.; Xiao, Y.; Guo, B. MicroRNA-22 Promotes Renal Tubulointerstitial Fibrosis by Targeting PTEN and Suppressing Autophagy in Diabetic Nephropathy. J. Diabetes Res. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Peng, H.; Chen, J.; Chen, X.; Han, F.; Xu, X.; He, X.; Yan, N. MicroRNA-21 protects from mesangial cell proliferation induced by diabetic nephropathy in db/db mice. FEBS Lett. 2009, 583, 2009–2014. [Google Scholar] [CrossRef]
- Zhao, B.; Li, H.; Liu, J.; Han, P.; Zhang, C.; Bai, H.; Yuan, X.; Wang, X.; Li, L.; Ma, H.; et al. MicroRNA-23b Targets Ras GTPase-Activating Protein SH3 Domain-Binding Protein 2 to Alleviate Fibrosis and Albuminuria in Diabetic Nephropathy. J. Am. Soc. Nephrol. 2016, 27, 2597–2608. [Google Scholar] [CrossRef]
- Zhao, D.; Jia, J.; Shao, H. miR-30e targets GLIPR-2 to modulate diabetic nephropathy: in vitro and in vivo experiments. J. Mol. Endocrinol. 2017, 59, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Shen, A.; Guo, F.; Song, Y.; Jing, N.; Ding, X.; Pan, M.; Zhang, H.; Wang, J.; Wu, L.; et al. Urinary Exosomal MiRNA-4534 as a Novel Diagnostic Biomarker for Diabetic Kidney Disease. Front. Endocrinol. 2020, 11. [Google Scholar] [CrossRef]
- Zhong, X.; Chung, A.C.K.; Chen, H.Y.; Dong, Y.; Meng, X.M.; Li, R.; Yang, W.; Hou, F.F.; Lan, H.Y. miR-21 is a key therapeutic target for renal injury in a mouse model of type 2 diabetes. Diabetologia 2013, 56, 663–674. [Google Scholar] [CrossRef]
- Zhou, H. P. S. Yuen, T. Pisitkun, P. A. Gonzales, H. Yasuda, J. W. Dear, P. Gross, M. A. Knepper, and R. A. Star. 2006. ’Collection, storage, preservation, and normalization of human urinary exosomes for biomarker discovery’, Kidney Int, 69: 1471-6.
- Zhou, X. G. Liu, M. Xu, X. Ying, B. Li, F. Cao, S. Cheng, B. Xiao, M. Cheng, L. Liang, M. Jia, W. Li, J. Liu, and Z. Li. 2022. ’Comprehensive analysis of PTEN-related ceRNA network revealing the key pathways WDFY3-AS2 - miR-21-5p/miR-221-3p/miR-222-3p - TIMP3 as potential biomarker in tumorigenesis and prognosis of kidney renal clear cell carcinoma’, Mol Carcinog, 61: 508-23.
- Zhou, X.; Qu, Z.; Zhu, C.; Lin, Z.; Huo, Y.; Wang, X.; Wang, J.; Li, B. Identification of urinary microRNA biomarkers for detection of gentamicin-induced acute kidney injury in rats. Regul. Toxicol. Pharmacol. 2016, 78, 78–84. [Google Scholar] [CrossRef]
- Zhou, Z.; Wan, J.; Hou, X.; Geng, J.; Li, X.; Bai, X. MicroRNA-27a promotes podocyte injury via PPARγ-mediated β-catenin activation in diabetic nephropathy. Cell Death Dis. 2017, 8, e2658–e2658. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Li, H.; Ao, Z.; Xu, H.; Luo, J.; Kaurich, C.; Yang, R.; Zhu, P.-W.; Chen, S.-D.; Wang, X.-D.; et al. Lipidomic identification of urinary extracellular vesicles for non-alcoholic steatohepatitis diagnosis. J. Nanobiotechnology 2022, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zubiri, I.; Posada-Ayala, M.; Sanz-Maroto, A.; Calvo, E.; Martin-Lorenzo, M.; Gonzalez-Calero, L.; de la Cuesta, F.; Lopez, J.A.; Fernandez-Fernandez, B.; Ortiz, A.; et al. Diabetic nephropathy induces changes in the proteome of human urinary exosomes as revealed by label-free comparative analysis. J. Proteom. 2014, 96, 92–102. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of storage temperature on kidney top expressed miRNAs and kidney enriched genes in uEV isolates. Urine EV were isolated by UC from urine stored at -20◦C vs -80◦C. A: Heatmap depicts the expression level of 29 out of 40 top miRNAs expressed in kidney (miRNATissueAtlas2) and found in the uEV isolates (miRNAs with ≥ 1 raw count in at least 50% of the samples). B. Heatmap depicts the expression of 33 out of 53 kidney enriched genes (human protein atlas) found in the uEV isolates (genes with ≥ 5 raw counts in at least 50% of the samples). Micro RNA (miRNA), messenger RNA (mRNA), ultracentrifugation (UC), urinary extracellular vesicles (uEV).
Figure 1.
Effect of storage temperature on kidney top expressed miRNAs and kidney enriched genes in uEV isolates. Urine EV were isolated by UC from urine stored at -20◦C vs -80◦C. A: Heatmap depicts the expression level of 29 out of 40 top miRNAs expressed in kidney (miRNATissueAtlas2) and found in the uEV isolates (miRNAs with ≥ 1 raw count in at least 50% of the samples). B. Heatmap depicts the expression of 33 out of 53 kidney enriched genes (human protein atlas) found in the uEV isolates (genes with ≥ 5 raw counts in at least 50% of the samples). Micro RNA (miRNA), messenger RNA (mRNA), ultracentrifugation (UC), urinary extracellular vesicles (uEV).
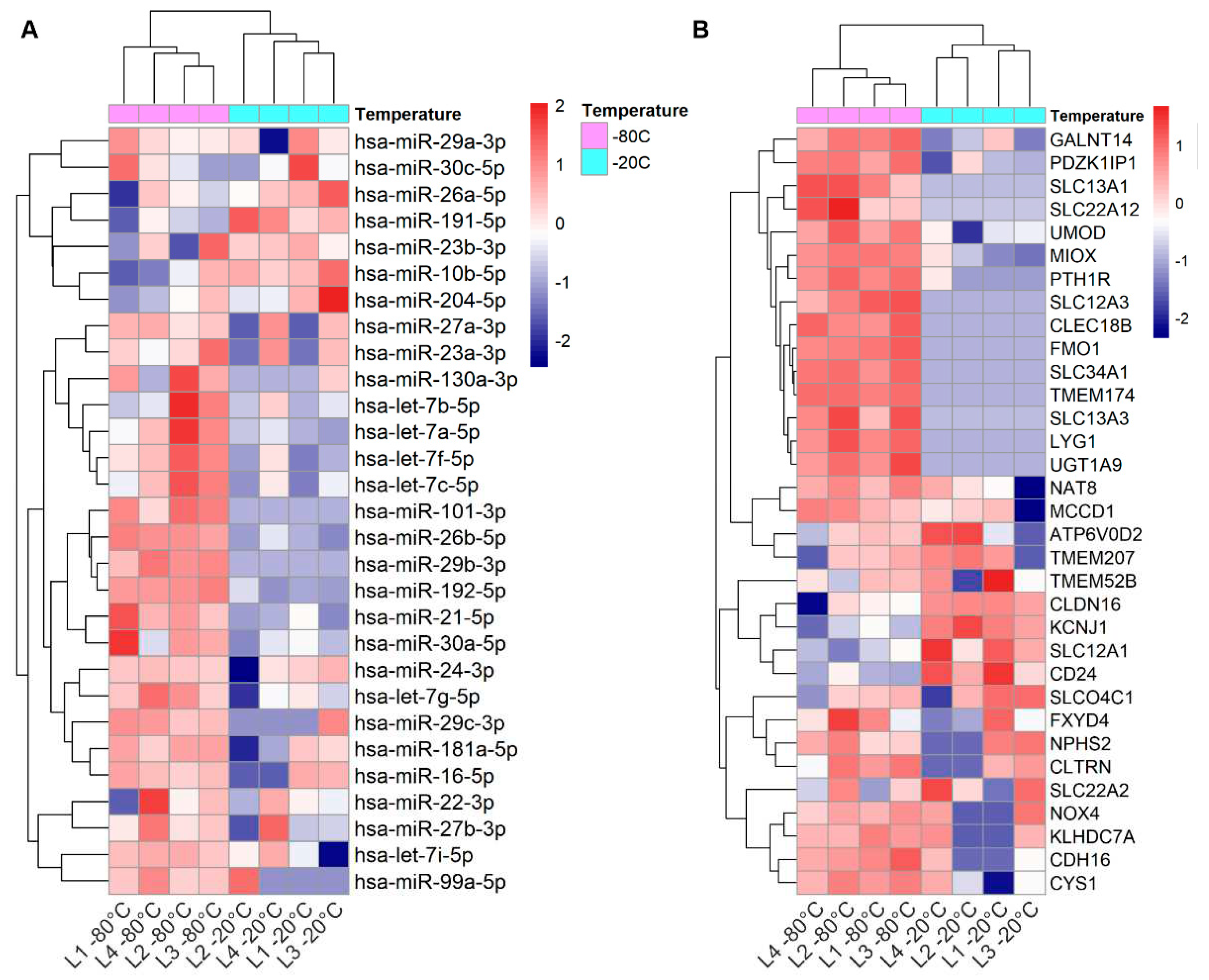
Figure 2.
Effect of EV isolation workflows on kidney top expressed miRNAs and kidney enriched genes in uEV isolates. rine EV were isolated by HFD, NG and UC workflows from urine samples of healthy controls (n=5) and T1D patients with macroalbuminuria (n=5). A. Heatmap depicts the expression level of 36 out of 40 top miRNAs expressed in kidney (miRNATissueAtlas2) and found in the uEV isolates (miRNAs with ≥ 5 raw counts in at least 50% of the samples). B. Heatmap depicts the expression level of 31 out of 56 kidney enriched genes (Human protein atlas) found in the uEV isolates (genes with ≥ 5 raw counts in at least 50% of the samples).
Figure 2.
Effect of EV isolation workflows on kidney top expressed miRNAs and kidney enriched genes in uEV isolates. rine EV were isolated by HFD, NG and UC workflows from urine samples of healthy controls (n=5) and T1D patients with macroalbuminuria (n=5). A. Heatmap depicts the expression level of 36 out of 40 top miRNAs expressed in kidney (miRNATissueAtlas2) and found in the uEV isolates (miRNAs with ≥ 5 raw counts in at least 50% of the samples). B. Heatmap depicts the expression level of 31 out of 56 kidney enriched genes (Human protein atlas) found in the uEV isolates (genes with ≥ 5 raw counts in at least 50% of the samples).
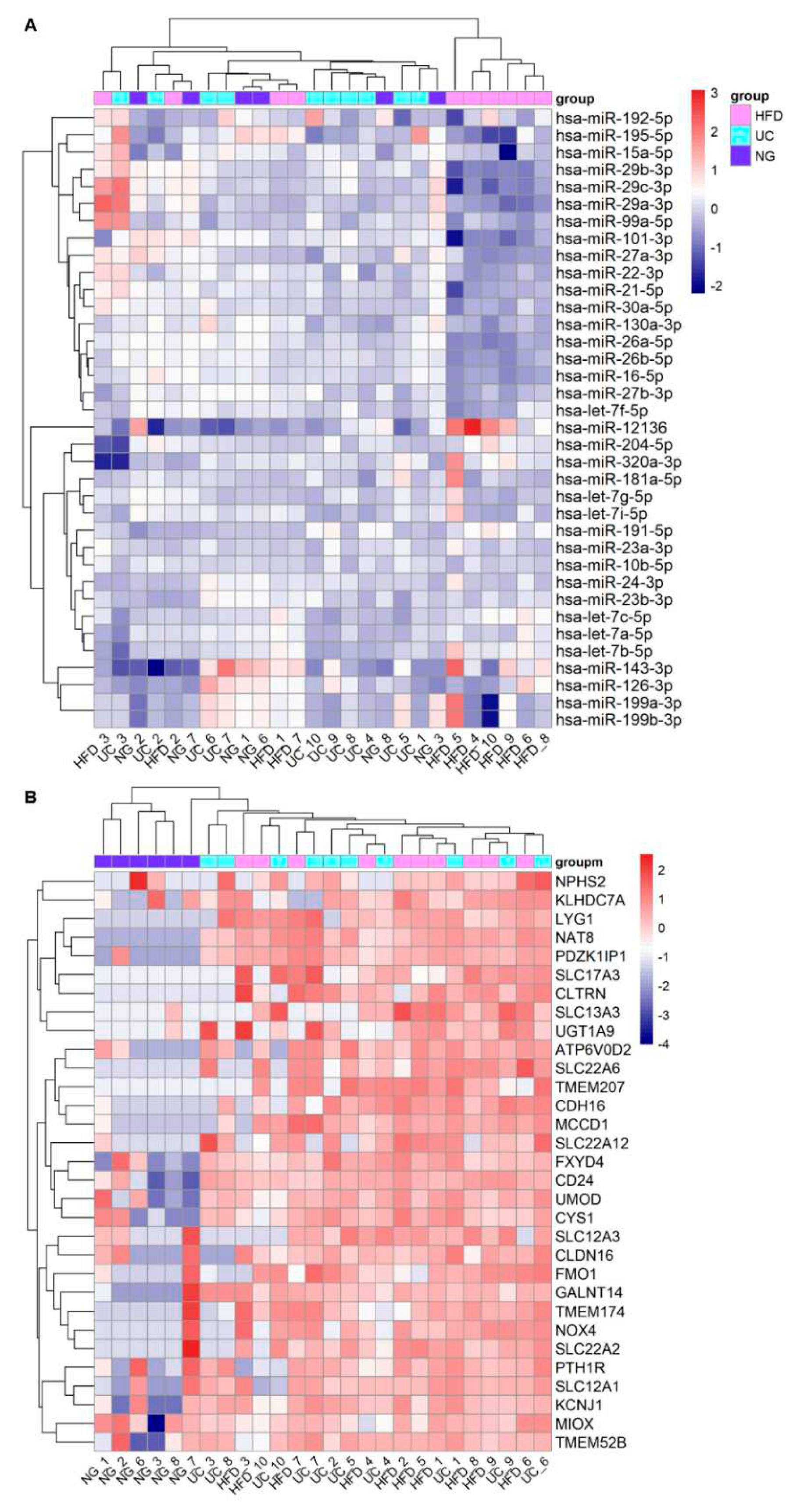
Figure 3.
Urinary EV capture miRNAs associated with DKD. Expression Heatmaps depict the expression of miRNAs associated with DKD and expressed in our uEV datasets. A. MiRNAs with evidence of dysregulation in kidney tissue and or cell lines and B. miRNAs dysregulated in uEV/urine/urinary sediments. The uEV expression data used in A and B corresponds to the UC isolation workflow dataset comprising healthy control and T1D macroalbuminuria groups (Barreiro et al., 2020, part of our UC miRNA dataset in tables 3 and 4). C. The 31 miRNAs that could separate individuals with DKD and macroalbuminuria (as shown in B) were analyzed in the PCa uEV miRNA dataset (Puhka et al. 2022). Diabetic kidney disease (DKD), Prostate cancer (PCa), ultracentrifugation (UC), urinary extracellular vesicles (uEV).
Figure 3.
Urinary EV capture miRNAs associated with DKD. Expression Heatmaps depict the expression of miRNAs associated with DKD and expressed in our uEV datasets. A. MiRNAs with evidence of dysregulation in kidney tissue and or cell lines and B. miRNAs dysregulated in uEV/urine/urinary sediments. The uEV expression data used in A and B corresponds to the UC isolation workflow dataset comprising healthy control and T1D macroalbuminuria groups (Barreiro et al., 2020, part of our UC miRNA dataset in tables 3 and 4). C. The 31 miRNAs that could separate individuals with DKD and macroalbuminuria (as shown in B) were analyzed in the PCa uEV miRNA dataset (Puhka et al. 2022). Diabetic kidney disease (DKD), Prostate cancer (PCa), ultracentrifugation (UC), urinary extracellular vesicles (uEV).
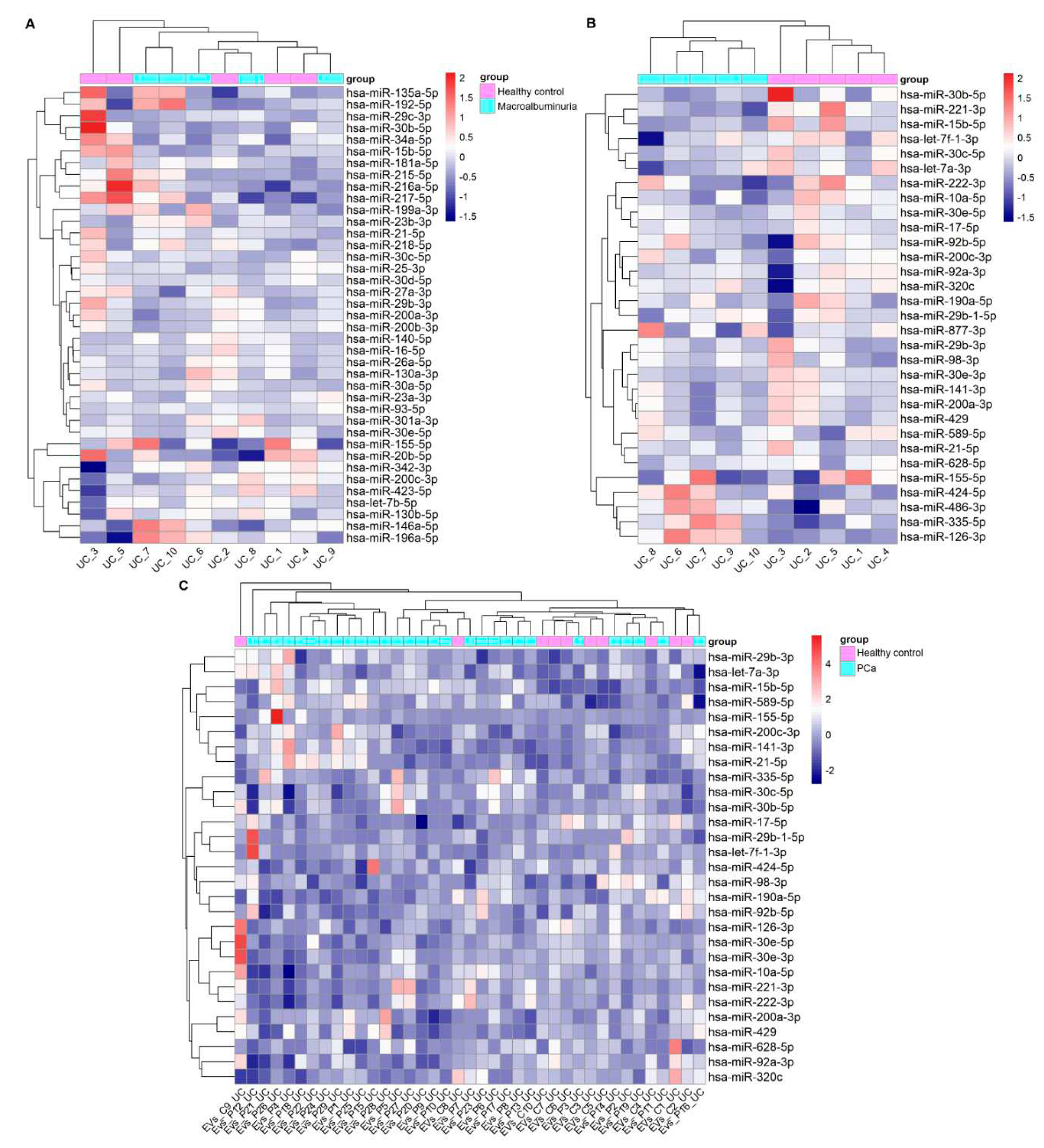
Figure 4.
Stable mRNA in common across diverse uEV datasets. Venn diagrams depict elements in common between the different datasets. A-C. A 3-step search for the stable uEV mRNA using the top 100 genes with the lowest CV from each dataset. Diabetic kidney disease (DKD), prostate cancer (PCa), urinary extracellular vesicles (uEV).
Figure 4.
Stable mRNA in common across diverse uEV datasets. Venn diagrams depict elements in common between the different datasets. A-C. A 3-step search for the stable uEV mRNA using the top 100 genes with the lowest CV from each dataset. Diabetic kidney disease (DKD), prostate cancer (PCa), urinary extracellular vesicles (uEV).
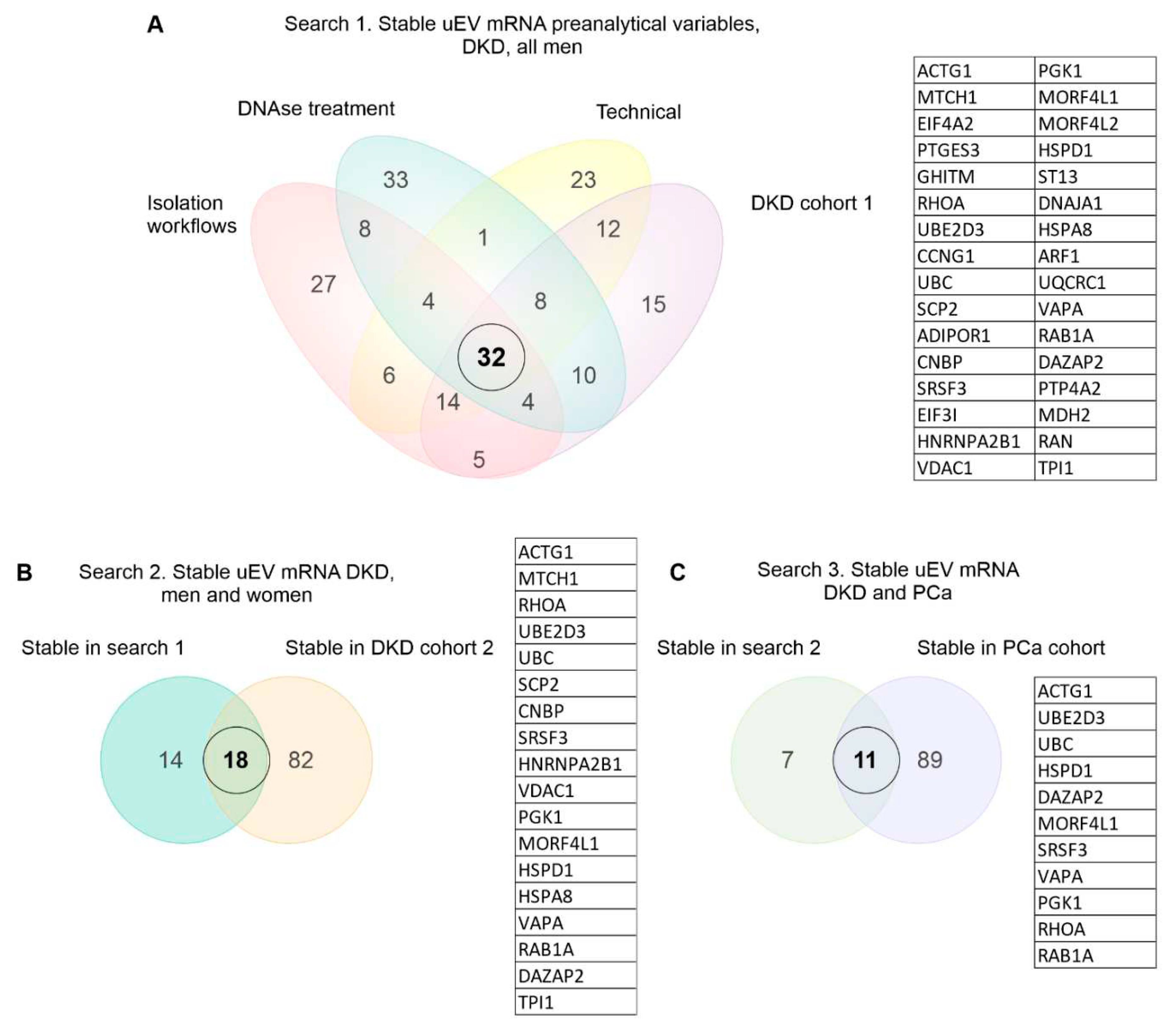
Figure 5.
The mRNA sequencing read counts of candidate reference in preanalytical and DKD uEV datasets from men. The uEV datasets included healthy controls and individuals with type 1 diabetes and different stages of albuminuria as well as comparisons of pre-analytical variables (all male). A-D. Line graphs depicts CPM of HSPD1, SRSF3, VAPA, RAB1A and MORF4L1 across samples and E-F. Boxplots depict CPM per candidate reference genes. A reference gene used commonly for normalization (GAPDH) and a gene with high CV in all datasets (UPK1A) were included. A,E. EV isolation workflows, B,F. in column DNAse treatment during uEV RNA extraction, C,G. a technical dataset (type of urine collection, pre-clearing the urine before freezing, and technical replicates) and D,H. DKD cohort 1. Sample pairs or triplicates are named similarly apart from the abbreviation of the tested variable. Centrifuged (C), Coefficient of variation (CV), counts per millon (CPM), hydrostatic filtration dialysis (HFD), macroalbuminuria (Macro), microalbuminuria (Micro), normoalbuminuria (Normo), ultracentrifugation (UC).
Figure 5.
The mRNA sequencing read counts of candidate reference in preanalytical and DKD uEV datasets from men. The uEV datasets included healthy controls and individuals with type 1 diabetes and different stages of albuminuria as well as comparisons of pre-analytical variables (all male). A-D. Line graphs depicts CPM of HSPD1, SRSF3, VAPA, RAB1A and MORF4L1 across samples and E-F. Boxplots depict CPM per candidate reference genes. A reference gene used commonly for normalization (GAPDH) and a gene with high CV in all datasets (UPK1A) were included. A,E. EV isolation workflows, B,F. in column DNAse treatment during uEV RNA extraction, C,G. a technical dataset (type of urine collection, pre-clearing the urine before freezing, and technical replicates) and D,H. DKD cohort 1. Sample pairs or triplicates are named similarly apart from the abbreviation of the tested variable. Centrifuged (C), Coefficient of variation (CV), counts per millon (CPM), hydrostatic filtration dialysis (HFD), macroalbuminuria (Macro), microalbuminuria (Micro), normoalbuminuria (Normo), ultracentrifugation (UC).
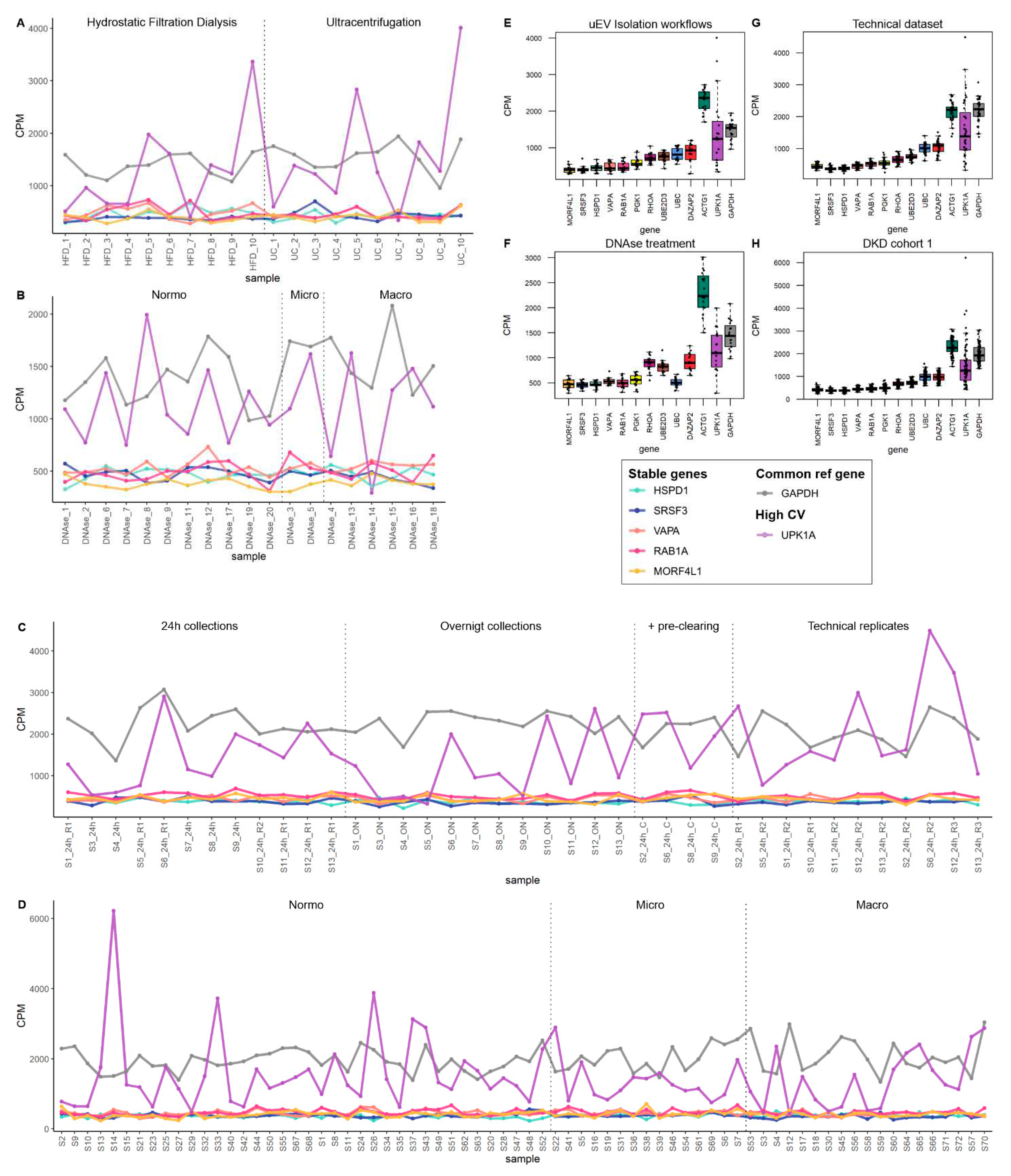
Figure 6.
The mRNA sequencing read counts of the candidate reference genes in uEV datasets from DKD study of women and from prostate cancer patients. A,B. Line graphs depicts CPM of HSPD1, SRSF3, VAPA, RAB1A and MORF4L1 across samples and C,D. Boxplots depict CPM per candidate reference genes. A reference gene used commonly for normalization (GAPDH) and a gene with high CV in all datasets (UPK1A) were included. The uEV datasets included A. DKD cohort 2 (women with type 1 diabetes and different stages of albuminuria) and B. PCa patients and healthy controls (technical replicates, R1-3). Samples PCa1, 3 and 4 were obtained before prostatectomy. Sample PCa2 was obtained after protectomy from the same donor as PCa1. Coefficient of variation (CV), counts per millon (CPM), macroalbuminuria (Macro), microalbuminuria (Micro), normoalbuminuria (Normo), prostate cancer (PCa).
Figure 6.
The mRNA sequencing read counts of the candidate reference genes in uEV datasets from DKD study of women and from prostate cancer patients. A,B. Line graphs depicts CPM of HSPD1, SRSF3, VAPA, RAB1A and MORF4L1 across samples and C,D. Boxplots depict CPM per candidate reference genes. A reference gene used commonly for normalization (GAPDH) and a gene with high CV in all datasets (UPK1A) were included. The uEV datasets included A. DKD cohort 2 (women with type 1 diabetes and different stages of albuminuria) and B. PCa patients and healthy controls (technical replicates, R1-3). Samples PCa1, 3 and 4 were obtained before prostatectomy. Sample PCa2 was obtained after protectomy from the same donor as PCa1. Coefficient of variation (CV), counts per millon (CPM), macroalbuminuria (Macro), microalbuminuria (Micro), normoalbuminuria (Normo), prostate cancer (PCa).
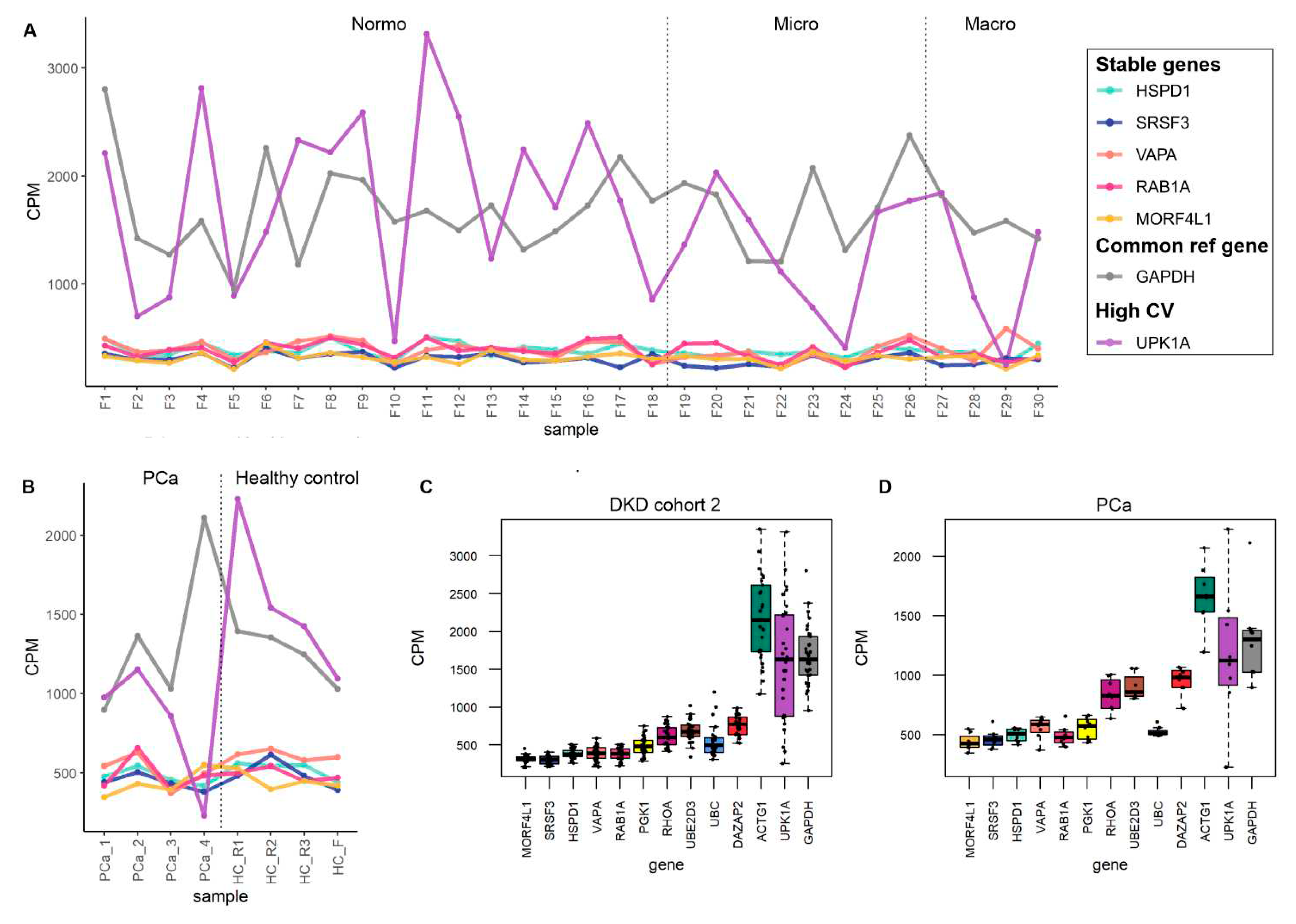
Figure 7.
Experimentally validated protein-protein interactions between the 11 stable genes. Network was generated using STRING (https://string-db.org/) and reproduced under Creative Commons BY 4.0 license (https://creativecommons.org/licenses/by/4.0/).
Figure 7.
Experimentally validated protein-protein interactions between the 11 stable genes. Network was generated using STRING (https://string-db.org/) and reproduced under Creative Commons BY 4.0 license (https://creativecommons.org/licenses/by/4.0/).
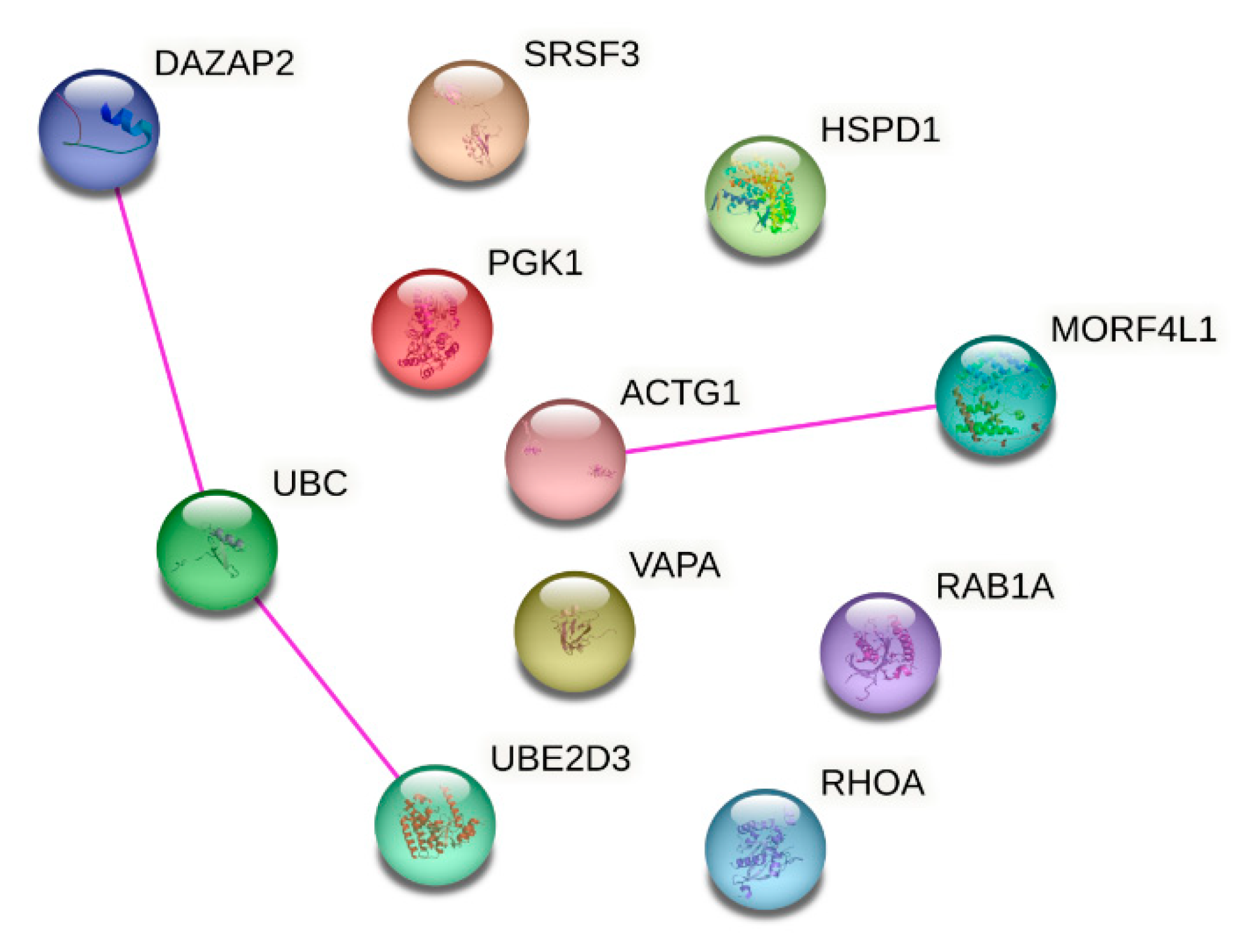
Table 1.
Datasets included in this study. Diabetic kidney disease (DKD), hydrostatic filtration dialysis (HFD), overnight (ON), prostate cancer (PCa), ultracentrifugation (UC), urinary extracellular vesicles (uEV), Urine Exosome Purification and RNA Isolation Midi Kit (NG). *NG was not included in the analysis due to the poor performance on RNA sequencing.
Table 1.
Datasets included in this study. Diabetic kidney disease (DKD), hydrostatic filtration dialysis (HFD), overnight (ON), prostate cancer (PCa), ultracentrifugation (UC), urinary extracellular vesicles (uEV), Urine Exosome Purification and RNA Isolation Midi Kit (NG). *NG was not included in the analysis due to the poor performance on RNA sequencing.
| Study | Description | storage temp | PI* | pre-clearing | isolation method | Urine sample type and disease | n (donors) | Analysis type | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Isolation workflows | uEV were isolated from urines | -80°C | yes | no | UC, HFD, and NG | 24h urine samples from healthy controls and T1D patients with macroalbuminuria. All men (n=20). | healthy controls=5macroalbuminuria=5 | miRNA and mRNA sequencing | (Barreiro et al. 2020) |
| Storage temperature | uEV were isolated from paired urine aliquots stored at -20°C or -80°C. | -80°C/-20°C | yes | no | UC | 24h urine samples from T1D patients with normoalbuminuria, microalbuminuria or macroalbuminuria. All men (n=19). | normoalbuminuria=2macroalbuminuria=2 | miRNA and mRNA sequencing | (Barreiro et al. 2021) |
| DNAse treatment | uEV RNA was isolated adding an in-column DNAse digestion step. | -80°C | yes | no | UC | 24h urine samples from T1D patients with normoalbuminuria, microalbuminuria or macroalbuminuria. All men. | normoalbuminuria=11microalbuminuria=2macroalbuminuria=6 | mRNA sequencing | (Barreiro et al. 2021) |
| ON/24h | uEV were isolated from urines derived from donors that provided on the same day 24h urine (full void during 24 hours) or ON urine (full first void). | -80°C | Yes | no | UC | ON and 24h urine samples from healthy controls and T1D patients with normoalbuminuria or macroalbuminuria. All men. | ON/24 pairs=12 | mRNA sequencing. | (Dwivedi et al. 2023) |
| Pre-clearing | uEV isolated from paired urine aliquots processed +/- pre-clearing before storage. | -80°C | Yes | yes/no | UC | 24h urine samples from T1D patients with microalbuminuria or macroalbuminuria. All men. | pre-clearing pairs=4 | mRNA sequencing | (Dwivedi et al. 2023) |
| Replicability of UC workflow | Pairs of urine aliquots were stored and processed at different time points (up to 5 months) | -80°C | yes | no | UC | 24h urine samples from healthy controls and T1D patients. All men. | duplicates=6triplicates=2 | mRNA sequencing | (Dwivedi et al. 2023) |
| DKD cohort 1 | uEV isolated from urines to | -80°C | yes | yes-no** | UC | 24h or ON urine samples from T1D patients with normoalbuminuria, microalbuminuria or macroalbuminuria. All men. | normoalbuminuria=38microalbuminuria=15macroalbuminuria=19 | mRNA sequencing | (Dwivedi et al. 2023) |
| DKD cohort 2 | uEV isolated from urines to validate candidate markers. | -80°C | yes | yes | UC | 24h urine samples from T1D patients with normoalbuminuria, microalbuminuria or macroalbuminuria. All women. | normoalbuminuria=18microalbuminuria=8macroalbuminuria=4 | mRNA sequencing | unpublished raw count data |
| PCa cohort | uEV isolated from urine samples from PCa patients | -80°C | no | yes | UC | Spot urine samples from healthy technical controls and PCa patients before and after radical prostatectomy. All men | PCa=3healthy controls= 2 (1 men with 3 technical replicates and 1 woman) | mRNA sequencing | (Puhka et al. 2022) |
Table 2.
Down- and up- regulated miRNAs in uEV derived from urines stored at -20°C for up to 1 year. Acute kidney injury (AKI), chronic kidney disease (CKD), diabetic kidney disease (DKD), lipopolysaccharide (LPS), streptozotocin (STZ), urinary extracellular vesicle (uEV).
Table 2.
Down- and up- regulated miRNAs in uEV derived from urines stored at -20°C for up to 1 year. Acute kidney injury (AKI), chronic kidney disease (CKD), diabetic kidney disease (DKD), lipopolysaccharide (LPS), streptozotocin (STZ), urinary extracellular vesicle (uEV).
| raw counts | ||||
|---|---|---|---|---|
| ID | -80°C | -20°C | Association with kidney diseases | |
| Downregulated | hsa-miR-21-5p | 42760 ± 22321 | 230 ± 115 | Downregulation in kidneys from CKD donors. (Sessa et al. 2020) |
| hsa-miR-375 | 11496 ± 4880 | 26 ± 13 | Pro-apoptotic in an in vitro model of AKI (renal tubular cells) (Jones et al. 2018) . | |
| hsa-miR-192-5p | 10651 ± 5178 | 15 ± 8 | Downregulation in DKD. Associated with fibrosis (Krupa et al. 2010). | |
| hsa-miR-378a-3p | 1445 ± 740 | 10 ± 5 | Upregulation seen in biopsies from donors with glomerular diseases (Müller-Deile et al. 2017). | |
| hsa-miR-101-3p | 971 ± 483 | 0 | Downregulated in kidneys from a mouse diabetic nephropathy model (STZ). Antifibrotic (Song et al. 2022). | |
| hsa-miR-107 | 700 ± 270 | 1 ± 0.3 | Downregulated in kidney biopsies from allograft dysfunction (Scian et al. 2011). | |
| hsa-miR-320b | 466 ± 262 | 2 ± 1 | ||
| hsa-miR-345-5p | 236 ± 121 | 0 | Upregulated in urine from a chemical model of AKI in rats (Zhou et al. 2016). | |
| hsa-miR-328-3p | 203 ± 88 | 0 | Downregulated in proximal tubule cells that underwent ischemia/reperfusion (Gao et al. 2022). | |
| hsa-miR-204-3p | 202 ± 157 | 0 | Upregulation protected podocytes exposed to high glucose from apoptosis (Han et al. 2019). | |
| hsa-miR-7-5p | 174 ± 119 | 0 | Downregulation protected proximal tubule cells from LPS in vitro (Wang et al. 2022). | |
| hsa-miR-197-3p | 154 ± 92 | 0 | Downregulated in urine from donors with intermittent MA (Ghai et al. 2018). | |
| hsa-miR-20b-5p | 151 ± 80 | 0 | Downregulated in kidneys and cell lines from mouse models of polycystic kidney disease (Shin et al. 2018). | |
| hsa-miR-148a-5p | 133 ± 60 | 0 | Increased in urine from donors with persistent macroalbuminuria (Ghai et al. 2018). | |
| hsa-miR-10a-3p | 114 ± 79 | 0 | Downregulated in kidneys from a mouse model of AKI (Yu et al. 2016). | |
| hsa-miR-629-5p | 109 ± 81 | 0 | Upregulated in kidney biopsies from donors with acute tubular necrosis (Pavkovic and Vaidya 2016). | |
| hsa-miR-92a-1-5p | 101 ± 59 | 0 | ||
| hsa-miR-193b-3p | 100 ± 62 | 0 | Upregulated in kidney from chronic kidney disease biopsies (Trevisani et al. 2016). | |
| hsa-miR-340-5p | 99 ± 36 | 0 | ||
| hsa-miR-3065-5p | 98 ± 43 | 0 | Upregulated in a mouse model of renal fibrosis (Li et al. 2022). | |
| hsa-miR-106a-5p | 92 ± 50 | 0 | Downregulation associated with podocyte injury induced by high glucose (He and Zeng 2020). | |
| hsa-miR-7704 | 87 ± 37 | 0 | ||
| hsa-miR-324-5p | 74 ± 42 | 0 | ||
| hsa-miR-374b-5p | 60 ± 24 | 0 | Downregulated in diabetic kidney biopsies (Yu et al. 2021). | |
| hsa-miR-99b-3p | 59 ± 19 | 0 | ||
| hsa-miR-4728-3p | 59 ± 20 | 0 | ||
| hsa-miR-132-3p | 57 ± 12 | 0 | Upregulation increases fibrosis in mouse and in vitro (Bijkerk et al. 2016). | |
| hsa-miR-361-5p | 55 ± 15 | 0 | ||
| hsa-miR-664a-5p | 54 ± 15 | 0 | Upregulated in uEV from donors with Idiopathic Membranous Nephropathy (Zhang et al. 2020). | |
| upregulated | hsa-miR-10a-5p | 47380 ± 33187 | 5272 ± 2636 | Downregulated in urine of individuals with AKI (Miller et al. 2022). |
| hsa-miR-125a-5p | 1017 ± 399 | 103 ± 51 | Downregulated in urine from donors with membranous nephropathy (Hou et al. 2021) | |
| hsa-miR-92b-3p | 864 ± 413 | 58 ± 29 | Upregulated in urine from donors with persistent macroalbuminuria (Ghai et al. 2018). | |
| hsa-miR-3960 | 77 ± 44 | 15 ± 7 | Upregulated in kidney biopsies from donors with acute tubular necrosis (Pavkovic and Vaidya 2016) | |
Table 3.
MiRNAs associated with DKD development and/or progression with evidence in kidney tissue and/or cell lines.
Table 3.
MiRNAs associated with DKD development and/or progression with evidence in kidney tissue and/or cell lines.
| miRNA (human) | Evidence | miRNA Regulation in disease group | Reference |
|---|---|---|---|
| let-7b-5p | in vitro and in vivo | Down | (Wang, Jha, et al. 2014) |
| miR-15b-5p | in vitro and in vivo | Up | (Tsai et al. 2020) |
| miR-16-5p | in vitro | Down | (Duan et al. 2021) |
| miR-20b-5p | in vitro | Up | (Wang et al. 2017) |
| miR-21-5p | in vitro and in vivo | Down | (Zhang et al. 2009) |
| miR-21-5p | in vitro and in vivo | Up | (Dey et al. 2011) |
| miR-21-5p | in vitro and in vivo | Up | (Zhong et al. 2013) |
| miR-21-5p | in vitro and in vivo | Up | (Chen et al. 2018) |
| miR-21-5p | in vitro and in vivo | - | (Wang, Gao, et al. 2014) |
| miR-21-5p | in vitro and in vivo | Up | (Lai et al. 2015) |
| miR-21-5p | in vitro and in vivo | Up | (Wang, Duan, et al. 2016) |
| miR-21-5p | in vitro and in vivo | Up | (Kölling et al. 2017) |
| miR-21 | in vivo | Up | (McClelland et al. 2015) |
| miR-22 | in vitro and in vivo | Up | (Zhang et al. 2018) |
| miR-23a-3p | in vitro | Up | (Xu, Sun, et al. 2018) |
| miR-23b-3p | in vitro and in vivo | Down | (Liu, Wang, et al. 2016) |
| miR-23b | in vitro and in vivo | Down | (Zhao et al. 2016) |
| miR-25-3p | in vitro and in vivo | Down | (Fu et al. 2010) |
| miR-25-3p | in vitro and in vivo | Down | (Oh et al. 2016) |
| miR-25-3p | in vitro and in vivo | Down | (Li et al. 2017) |
| miR-25 | in vitro and in vivo | Down | (Liu et al. 2017) |
| miR-26a-5p | in vitro and in vivo | Down | (Koga et al. 2015) |
| miR-27a-3p | in vitro and in vivo | Up | (Wu et al. 2016) |
| miR-27a | in vitro and in vivo | Up | (Zhou et al. 2017) |
| miR-29b-3p | in vitro and in vivo | Up | (Beltrami et al. 2018) |
| miR-29b-3p | in vitro and in vivo | Down | (Chen et al. 2014) |
| miR-29c-3p | in vitro and in vivo | Up | (Long et al. 2011) |
| miR-29a | in vitro and in vivo | Down | (Lin et al. 2014) |
| miR-29a | in vitro and in vivo | Down | (Hsu et al. 2016) |
| miR-29a/b/c family | in vitro and in vivo | Down | (Wang et al. 2012) |
| miR-30e-5p | in vitro and in vivo | Down | (Zhao, Jia, and Shao 2017) |
| miR-30s (family) | in vitro and in vivo | Down | (Liu, Peng, et al. 2016) |
| miR-30b-5p | in vitro | Down | (Wang et al. 2021) |
| miR-30c-5p | in vitro and in vivo | Down | (Cui, Yu, and Cui 2020) |
| miR-34a-5p | in vitro and in vivo | Up | (Zhang et al. 2014) |
| miR-34a-5p | in vitro and in vivo | Up | (Xue et al. 2018) |
| miR-34c-5p | in vitro | Down | (Liu et al. 2015) |
| miR-93-5p | in vitro and in vivo | Down | (Long et al. 2010) |
| miR-124-5p | in vivo | Up | (Li et al. 2013) |
| hsa-miR-126-3p | in vitro and in vivo | Up | (Beltrami et al. 2018) |
| miR-130a-3p | in vitro | Down | (Jiang et al. 2018) |
| miR-130b-5p | in vitro and in vivo | Down | (Bai et al. 2016) |
| miR-133b | in vitro and in vivo | Up | (Sun et al. 2018) |
| miR-134-5p | in vitro and in vivo | Up | (Qian et al. 2018) |
| miR-135a-5p | in vitro and in vivo | Up | (He et al. 2014) |
| miR-140-5p | in vitro and in vivo | Down | (Su et al. 2020) |
| miR-145-5p | in vitro and in vivo | Up | (Barutta et al. 2013) |
| miR-145-5p | in vitro | Down | (Wei, Liu, and Guan 2020) |
| miR-146a-5p | in vitro and in vivo | Up | (Huang et al. 2014) |
| miR-146a-5p | in vitro and in vivo | Down | (Lee et al. 2017) |
| miR-146a | in vivo | Down | (Bhatt et al. 2016) |
| miR-146a | in vitro and in vivo | Down | (Wan and Li 2018) |
| miR-155-5p | in vitro and in vivo | Up | (Huang et al. 2014) |
| miR-155-5p | in vitro and in vivo | Up | (Beltrami et al. 2018) |
| miR-155-5p | in vitro | Up | (Wang, Zheng, et al. 2018) |
| miR-181a-5p | in vitro and in vivo | Down | (Xu et al. 2017) |
| miR-192-5p | in vitro and in vivo | Up | (Kato et al. 2007) |
| miR-192-5p | in vitro and in vivo | Down | (Wang et al. 2010) |
| miR-192-5p | in vitro and in vivo | Up | (Kato et al. 2011) |
| miR-192 | in vitro and in vivo | Up in microalbuminuria, Down in macroalbuminuria | (Jia et al. 2016) |
| miR-192 | in vitro and in vivo | Down | (Krupa et al. 2010) |
| miR-192 | in vitro and in vivo | Up | (Deshpande et al. 2013) |
| miR-193a | in vitro | Up | (Mishra et al. 2018) |
| miR-195 | in vitro and in vivo | Down | (Chen et al. 2012) |
| miR-196a-5p | in vitro and in vivo | Down | (Wang et al. 2015) |
| miR-199a-3p | in vitro and in vivo | Down | (Zhang, Qin, and Shi 2020) |
| miR-199b-5p | in vitro and in vivo | Up | (Sun et al. 2018) |
| miR-200a-3p | in vitro and in vivo | Down | (Wang et al. 2011) |
| miR-200 b/c-3p | in vitro and in vivo | Up | (Kato et al. 2011) |
| miR-200 b/c | in vitro and in vivo | Up | (Park et al. 2013) |
| miR-214-3p | in vitro and in vivo | Up | (Wang, Shen, et al. 2016) |
| miR-214-3p | in vitro and in vivo | Up | (Bera et al. 2017) |
| miR-215-5p | in vitro and in vivo | Down | (Wang et al. 2010) |
| miR-216a-5p | in vitro and in vivo | Up | (Kato et al. 2009) |
| miR-216a-5p | in vivo | UP | (Kato et al. 2010) |
| miR-217-5p | in vitro | Up | (Sun et al. 2017) |
| miR-217-5p | in vitro and in vivo | Up | (Kato et al. 2009) |
| miR-218-5p | in vitro | Up | (Yang, Wang, and Li 2016) |
| miR-301a-3p | in vitro | Down | (Jiang et al. 2018) |
| miR-342-3p | in vitro and in vivo | Down | (Jiang et al. 2020) |
| miR-374a | in vitro and in vivo | Down | (Yang et al. 2018) |
| miR-377-3p | in vitro and in vivo | Up | (Wang et al. 2008) |
| miR-379-5p | in vitro and in vivo | Down | (Li et al. 2019) |
| miR-379 megacluster | in vitro and in vivo | Up | (Kato et al. 2016) |
| miR-382 | in vitro and in vivo | Up | (Wang, Wen, et al. 2018) |
| miR-423-5p | in vitro | Down | (Xu, Zhang, et al. 2018) |
| miR-451a | in vitro and in vivo | Down | (Sun et al. 2016) |
| miR-451a | in vivo | Up/Down | (Mohan et al. 2016) |
| miR-503 | in vitro and in vivo | Up | (Zha et al. 2019) |
| miR-770-5p | in vitro and in vivo | Up | (Wang and Li 2020) |
| miR-874 | in vitro and in vivo | Up (overt nephropathy) | (Zanchi et al. 2017) |
| miR-874 | in vitro and in vivo | Down | (Yao et al. 2018) |
| miR-1207-5p | in vitro | Up | (Alvarez et al. 2013) |
Table 6.
Coefficient of variation (CV) of the stable genes across datasets. Diabetic kidney disease (DKD), prostate cancer (PCa), type 1 diabetes (T1D). * Commonly used reference gene for normalization of qPCR data. ** Gene with high CV in all datasets.
Table 6.
Coefficient of variation (CV) of the stable genes across datasets. Diabetic kidney disease (DKD), prostate cancer (PCa), type 1 diabetes (T1D). * Commonly used reference gene for normalization of qPCR data. ** Gene with high CV in all datasets.
| CV | ||||||
|---|---|---|---|---|---|---|
| Isolation workflows | DNAse treatment | Technical datasets | DKD cohort 1 (T1D, men) | DKD cohort 2 (T1D, women) | PCa | |
| HSPD1 | 0.23 | 0.13 | 0.15 | 0.14 | 0.16 | 0.12 |
| SRSF3 | 0.21 | 0.13 | 0.17 | 0.15 | 0.18 | 0.16 |
| VAPA | 0.26 | 0.13 | 0.16 | 0.16 | 0.23 | 0.16 |
| RAB1A | 0.26 | 0.19 | 0.15 | 0.18 | 0.21 | 0.17 |
| MORF4L1 | 0.22 | 0.13 | 0.17 | 0.21 | 0.16 | 0.16 |
| PGK1 | 0.22 | 0.20 | 0.21 | 0.19 | 0.24 | 0.16 |
| RHOA | 0.17 | 0.16 | 0.19 | 0.15 | 0.22 | 0.16 |
| UBE2D3 | 0.18 | 0.14 | 0.13 | 0.15 | 0.20 | 0.11 |
| DAZAP2 | 0.26 | 0.19 | 0.19 | 0.20 | 0.17 | 0.16 |
| UBC | 0.19 | 0.16 | 0.16 | 0.20 | 0.38 | 0.11 |
| ACTG1 | 0.13 | 0.19 | 0.14 | 0.15 | 0.25 | 0.08 |
| GAPDH* | 0.18 | 0.20 | 0.17 | 0.19 | 0.24 | 0.29 |
| UPK1A** | 0.70 | 0.36 | 0.59 | 0.63 | 0.49 | 0.49 |
Table 6.
Gene ontology biological processes associated with the stable genes (Uniprot, https://www.uniprot.org/).
Table 6.
Gene ontology biological processes associated with the stable genes (Uniprot, https://www.uniprot.org/).
| Gene name | Entry | Protein names | Gene Ontology (biological process) |
| PGK1 | P00558 | Phosphoglycerate kinase 1 | canonical glycolysis [GO:0061621]; cellular response to hypoxia [GO:0071456]; epithelial cell differentiation [GO:0030855]; gluconeogenesis [GO:0006094]; glycolytic process [GO:0006096]; negative regulation of angiogenesis [GO:0016525]; phosphorylation [GO:0016310]; plasminogen activation [GO:0031639] |
| UBC | P0CG48 | Polyubiquitin-C | modification-dependent protein catabolic process [GO:0019941]; protein ubiquitination [GO:0016567] |
| HSPD1 | P10809 | 60 kDa heat shock protein, mitochondrial | ’de novo’ protein folding [GO:0006458]; activation of cysteine-type endopeptidase activity involved in apoptotic process [GO:0006919]; apoptotic mitochondrial changes [GO:0008637]; B cell activation [GO:0042113]; B cell proliferation [GO:0042100]; biological process involved in interaction with symbiont [GO:0051702]; cellular response to interleukin-7 [GO:0098761]; chaperone-mediated protein complex assembly [GO:0051131]; isotype switching to IgG isotypes [GO:0048291]; mitochondrial unfolded protein response [GO:0034514]; MyD88-dependent toll-like receptor signaling pathway [GO:0002755]; negative regulation of apoptotic process [GO:0043066]; positive regulation of apoptotic process [GO:0043065]; positive regulation of interferon-alpha production [GO:0032727]; positive regulation of interleukin-10 production [GO:0032733]; positive regulation of interleukin-12 production [GO:0032735]; positive regulation of interleukin-6 production [GO:0032755]; positive regulation of macrophage activation [GO:0043032]; positive regulation of T cell activation [GO:0050870]; positive regulation of T cell mediated immune response to tumor cell [GO:0002842]; positive regulation of type II interferon production [GO:0032729]; protein folding [GO:0006457]; protein import into mitochondrial intermembrane space [GO:0045041]; protein maturation [GO:0051604]; protein refolding [GO:0042026]; protein stabilization [GO:0050821]; response to cold [GO:0009409]; response to unfolded protein [GO:0006986]; T cell activation [GO:0042110] |
| UBE2D3 | P61077 | Ubiquitin-conjugating enzyme E2 D3 | apoptotic process [GO:0006915]; DNA repair [GO:0006281]; negative regulation of BMP signaling pathway [GO:0030514]; negative regulation of transcription by RNA polymerase II [GO:0000122]; positive regulation of protein targeting to mitochondrion [GO:1903955]; proteasome-mediated ubiquitin-dependent protein catabolic process [GO:0043161]; protein autoubiquitination [GO:0051865]; protein K11-linked ubiquitination [GO:0070979]; protein K48-linked ubiquitination [GO:0070936]; protein modification process [GO:0036211]; protein monoubiquitination [GO:0006513]; protein polyubiquitination [GO:0000209]; protein ubiquitination [GO:0016567] |
| RHOA | P61586 | Transforming protein RhoA | actin cytoskeleton organization [GO:0030036]; actin cytoskeleton reorganization [GO:0031532]; actin filament organization [GO:0007015]; alpha-beta T cell lineage commitment [GO:0002363]; androgen receptor signaling pathway [GO:0030521]; angiotensin-mediated vasoconstriction involved in regulation of systemic arterial blood pressure [GO:0001998]; aortic valve formation [GO:0003189]; apical junction assembly [GO:0043297]; apolipoprotein A-I-mediated signaling pathway [GO:0038027]; beta selection [GO:0043366]; cell junction assembly [GO:0034329]; cell migration [GO:0016477]; cell-matrix adhesion [GO:0007160]; cellular response to chemokine [GO:1990869]; cellular response to cytokine stimulus [GO:0071345]; cellular response to lipopolysaccharide [GO:0071222]; cerebral cortex cell migration [GO:0021795]; cleavage furrow formation [GO:0036089]; cortical cytoskeleton organization [GO:0030865]; cytoplasmic microtubule organization [GO:0031122]; endothelial cell migration [GO:0043542]; endothelial tube lumen extension [GO:0097498]; establishment of epithelial cell apical/basal polarity [GO:0045198]; establishment or maintenance of cell polarity [GO:0007163]; forebrain radial glial cell differentiation [GO:0021861]; GTP metabolic process [GO:0046039]; kidney development [GO:0001822]; mitotic cleavage furrow formation [GO:1903673]; mitotic spindle assembly [GO:0090307]; motor neuron apoptotic process [GO:0097049]; negative chemotaxis [GO:0050919]; negative regulation of cell migration involved in sprouting angiogenesis [GO:0090051]; negative regulation of cell size [GO:0045792]; negative regulation of cell-substrate adhesion [GO:0010812]; negative regulation of I-kappaB kinase/NF-kappaB signaling [GO:0043124]; negative regulation of intracellular steroid hormone receptor signaling pathway [GO:0033144]; negative regulation of motor neuron apoptotic process [GO:2000672]; negative regulation of neuron differentiation [GO:0045665]; negative regulation of neuron projection development [GO:0010977]; negative regulation of oxidative phosphorylation [GO:0090324]; negative regulation of reactive oxygen species biosynthetic process [GO:1903427]; negative regulation of vascular associated smooth muscle cell migration [GO:1904753]; negative regulation of vascular associated smooth muscle cell proliferation [GO:1904706]; neuron migration [GO:0001764]; neuron projection morphogenesis [GO:0048812]; odontogenesis [GO:0042476]; ossification involved in bone maturation [GO:0043931]; positive regulation of actin filament polymerization [GO:0030838]; positive regulation of alpha-beta T cell differentiation [GO:0046638]; positive regulation of cell growth [GO:0030307]; positive regulation of cysteine-type endopeptidase activity involved in apoptotic process [GO:0043280]; positive regulation of cytokinesis [GO:0032467]; positive regulation of I-kappaB kinase/NF-kappaB signaling [GO:0043123]; positive regulation of leukocyte adhesion to vascular endothelial cell [GO:1904996]; positive regulation of lipase activity [GO:0060193]; positive regulation of neuron apoptotic process [GO:0043525]; positive regulation of neuron differentiation [GO:0045666]; positive regulation of NIK/NF-kappaB signaling [GO:1901224]; positive regulation of podosome assembly [GO:0071803]; positive regulation of protein serine/threonine kinase activity [GO:0071902]; positive regulation of stress fiber assembly [GO:0051496]; positive regulation of T cell migration [GO:2000406]; positive regulation of translation [GO:0045727]; positive regulation of vascular associated smooth muscle contraction [GO:1904695]; regulation of actin cytoskeleton organization [GO:0032956]; regulation of calcium ion transport [GO:0051924]; regulation of cell migration [GO:0030334]; regulation of cell shape [GO:0008360]; regulation of dendrite development [GO:0050773]; regulation of focal adhesion assembly [GO:0051893]; regulation of microtubule cytoskeleton organization [GO:0070507]; regulation of modification of postsynaptic actin cytoskeleton [GO:1905274]; regulation of modification of postsynaptic structure [GO:0099159]; regulation of neural precursor cell proliferation [GO:2000177]; regulation of osteoblast proliferation [GO:0033688]; regulation of systemic arterial blood pressure by endothelin [GO:0003100]; regulation of transcription by RNA polymerase II [GO:0006357]; response to amino acid [GO:0043200]; response to ethanol [GO:0045471]; response to glucocorticoid [GO:0051384]; response to glucose [GO:0009749]; response to hypoxia [GO:0001666]; response to mechanical stimulus [GO:0009612]; response to xenobiotic stimulus [GO:0009410]; Rho protein signal transduction [GO:0007266]; Roundabout signaling pathway [GO:0035385]; skeletal muscle satellite cell migration [GO:1902766]; skeletal muscle tissue development [GO:0007519]; stress fiber assembly [GO:0043149]; stress-activated protein kinase signaling cascade [GO:0031098]; substantia nigra development [GO:0021762]; substrate adhesion-dependent cell spreading [GO:0034446]; trabecula morphogenesis [GO:0061383]; Wnt signaling pathway, planar cell polarity pathway [GO:0060071]; wound healing, spreading of cells [GO:0044319] |
| RAB1A | P62820 | Ras-related protein Rab-1A | autophagosome assembly [GO:0000045]; autophagy [GO:0006914]; cell migration [GO:0016477]; COPII-coated vesicle cargo loading [GO:0090110]; defense response to bacterium [GO:0042742]; endocytosis [GO:0006897]; endoplasmic reticulum to Golgi vesicle-mediated transport [GO:0006888]; Golgi organization [GO:0007030]; growth hormone secretion [GO:0030252]; intracellular protein transport [GO:0006886]; melanosome transport [GO:0032402]; positive regulation of glycoprotein metabolic process [GO:1903020]; positive regulation of interleukin-8 production [GO:0032757]; substrate adhesion-dependent cell spreading [GO:0034446]; vesicle transport along microtubule [GO:0047496]; vesicle-mediated transport [GO:0016192]; virion assembly [GO:0019068] |
| ACTG1 | P63261 | Actin, cytoplasmic 2 | angiogenesis [GO:0001525]; cellular response to type II interferon [GO:0071346]; maintenance of blood-brain barrier [GO:0035633]; morphogenesis of a polarized epithelium [GO:0001738]; platelet aggregation [GO:0070527]; positive regulation of cell migration [GO:0030335]; positive regulation of gene expression [GO:0010628]; positive regulation of wound healing [GO:0090303]; protein localization to bicellular tight junction [GO:1902396]; regulation of focal adhesion assembly [GO:0051893]; regulation of stress fiber assembly [GO:0051492]; regulation of synaptic vesicle endocytosis [GO:1900242]; regulation of transepithelial transport [GO:0150111]; response to calcium ion [GO:0051592]; response to mechanical stimulus [GO:0009612]; retina homeostasis [GO:0001895]; sarcomere organization [GO:0045214]; tight junction assembly [GO:0120192] |
| SRSF3 | P84103 | Serine/arginine-rich splicing factor 3 (Pre-mRNA-splicing factor SRP20) | cellular response to leukemia inhibitory factor [GO:1990830]; mRNA export from nucleus [GO:0006406]; mRNA splicing, via spliceosome [GO:0000398]; primary miRNA processing [GO:0031053]; regulation of mRNA splicing, via spliceosome [GO:0048024] |
| DAZAP2 | Q15038 | DAZ-associated protein 2 (Deleted in azoospermia-associated protein 2) | positive regulation of protein serine/threonine kinase activity [GO:0071902]; positive regulation of RNA polymerase II regulatory region sequence-specific DNA binding [GO:1905636]; protein destabilization [GO:0031648]; stress granule assembly [GO:0034063] |
| VAPA | Q9P0L0 | Vesicle-associated membrane protein-associated protein A (VAMP-A) | cell death [GO:0008219]; ceramide transport [GO:0035627]; COPII-coated vesicle budding [GO:0090114]; endoplasmic reticulum to Golgi vesicle-mediated transport [GO:0006888]; membrane fusion [GO:0061025]; negative regulation by host of viral genome replication [GO:0044828]; neuron projection development [GO:0031175]; phospholipid transport [GO:0015914]; positive regulation by host of viral genome replication [GO:0044829]; positive regulation of I-kappaB kinase/NF-kappaB signaling [GO:0043123]; protein localization to endoplasmic reticulum [GO:0070972]; sphingomyelin biosynthetic process [GO:0006686]; sterol transport [GO:0015918]; viral release from host cell [GO:0019076] |
| MORF4L1 | Q9UBU8 | Mortality factor 4-like protein 1 (MORF-related gene 15 protein) | chromatin organization [GO:0006325]; double-strand break repair via homologous recombination [GO:0000724]; fibroblast proliferation [GO:0048144]; histone acetylation [GO:0016573]; histone deacetylation [GO:0016575]; histone H2A acetylation [GO:0043968]; histone H4 acetylation [GO:0043967]; positive regulation of DNA-templated transcription [GO:0045893]; positive regulation of double-strand break repair via homologous recombination [GO:1905168]; regulation of apoptotic process [GO:0042981]; regulation of cell cycle [GO:0051726]; regulation of double-strand break repair [GO:2000779] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



