
Submitted:
29 May 2023
Posted:
31 May 2023
You are already at the latest version
Abstract
Pharmaceuticals including antibiotics are among hazardous micropollutants (HMP) of the environment. Incomplete degradation of the HMP leads to their persistence in water bodies causing a plethora of deleterious effects. Conventional wastewater treatment cannot remove HMP completely, and a promising alternative is comprised by biotechnologies based on microalgae. The use of immobilized microalgae in environmental biotechnology is advantageous since immobilized cultures allow recycling of the microalgal cells, support higher cell densities, and boost tolerance of microalgae to stresses including HMP. Here we report on a comparative study of HMP (exemplified by the antibiotic ceftriaxone) removal by suspended and chitosan-immobilized cells of Lobosphaera sp. IPPAS C-2047 unialgal culture. We also monitored the dynamics of photosynthetic pigments content and the physiological condition of the cells as reflected by the activity of their photosynthetic apparatus. Special attention was paid to the changes in the accumulation of valuable arachidonic acid (C20:4). In addition to this, we followed the changes in the culture microbiome induced the antibiotic exposure. Collectively, our results showed that both suspended and immobilized cultures took up ceftriaxone from the culture medium. In the case of immobilized culture, a significant amount of the antibiotic was adsorbed on the chitosan carrier itself. The dynamics of the taxonomic composition of the microbiome of the microalgae was more shifted by the immobilization on the chitosan than by exposure to 20 mg/L of ceftriaxone. The possibility and limitations of the using of chitosan-immobilized Lobosphaera sp. IPPAS C-2047 for HMP removal coupled with the production of valuable long-chain polyunsaturated fatty acids is discussed.
Keywords:
Lobosphaera
; attached cultivation
; antibiotics
; bioremoval
; arachidonic acid
1. Introduction
Immobilization of microalgal cells on various synthetic and natural carriers attracted increasing interest in the field of wastewater treatment [1,2]. The main advantages of immobilized (attached) microalgal cultures are higher resistance to toxicants and overall robustness, increased rate of biomass and value-added molecule accumulation as compared to suspended cultures [3]. Moreover, attached cultivation eliminates the need for costly cell-harvesting procedures such as centrifugation; attached cultures can be also used through several production cycles without significant loss of their activity [1,4]. Immobilized microalgae cultivation was shown to be a promising technology for removal of antibiotics from wastewater [3,5,6]. Microalgae can effectively remove antibiotics from wastewaters (see [5,7] and references therein). For example, a high-rate algal pond (HRAP) system dominated by Coelastrum sp. removed 33 antibiotics (with an average concentration of 223 µg/L) from municipal wastewater with average antibiotic removal rates being up to 50% higher than those of the conventional activated sludge process [8]. The immobilized microalgal biomass grown in suitable wastewater and enriched with nutrients, lipids, and carotenoids can be used as biofertilizer, for production of biofuel and value-added compounds [9].
The successful application of immobilized microalgae in wastewater treatment depends on the informed choice of the optimal carrier and immobilization technique [2]. Current progress in material science made available a broad spectrum of natural and synthetic materials for microalgae immobilization [1]. Among them, synthetic polymers (acrylamide resins, polyurethanes, polyvinyl alcohol, polypropylene) feature a high mechanical and chemical stability, but using of synthetic polymers entails the problem of the utilization of used cell carriers [2]. The key advantages of the natural carriers (loofa, sphagnum, turf, cellulose, carrageenan, alginate) are hydrophilicity, biocompatibility, and safety for the environment should be balanced against their low porosity and mechanical stability in wastewater [2,4]. Particularly, the cross-linked chitosan-based polymeric materials as biocompatible, cost-effective, and non-toxic carriers are prospective for immobilization of microalgae [2,10]. These polymers have a variety of potential applications as biosorbents for water biotreatment applications due to a high potential for sorption of heavy metal ions, dyes, phenol, polychlorinated biphenyls [11].
A serious concern is that of leakage of antibiotics into the environment from wastewater treatment plants [5,12]. Most antibiotics are hardly removed by the bacteria of activated sewage sludge since the antibiotics are purposedly designed to inhibit bacterial growth and metabolism [13]. Incomplete, slow degradation of the antibiotics leads to their increased persistence in the environment and enrichment of natural communities and wastewater treatment plants with pathogenic and opportunistic microorganisms resistant to antibiotics and hence dangerous to human health [12,14]. Although concentrations of hazardous micropollutants including antibiotics currently encountered in waste streams are low in comparison with their total organics content, these micropollutants impose a serious threat to the environment due to their high biotoxicity. Although environmental concentrations of antibiotics are seldom reported to have acute toxicity to aquatic organisms, long-term negative effects on individual organisms and populations have been documented, including the impact on microbial consortia [6].
As noted above, microalgae-based technology has been reported as an effective method to remove and degrade antibiotics with added benefits of CO2 fixation, accumulation of value-added products, nutrients, and other organic pollutants removal [6,15]. At the same time, little is known about the influence of antibiotics on the biology of immobilized microalgal cells. To bridge this gap, at least in part, we studied the influence of a widespread antibiotic ceftriaxone (CTA) on the accumulation of arachidonic by suspended and immobilized on the cross-linked chitosan carrier microalgae Lobosphaera sp. strain IPPAS C-2047. Representatives of the genus Lobosphaera including the studied strain are the richest plant source of the polyunsaturated ω-6 arachidonic acid (ARA) [16]. Arachidonic acid is an essential, structural, and functional constituent of cell membranes, it is used as a nutraceutical food and feed additive [17,18].
The removal of the antibiotic by different cultures of the Lobosphaera sp. was also studied. Results of this study would augment the development of the technologies with chitosan-immobilized microalgae intended for antibiotics removal from wastewater coupled with the production of valuable polyunsaturated fatty acids by attached microalgal cultures.
2. Results
The experiments in our study have been conducted in two phases. The first phase was comprised by pilot experiments carried out in flasks (see Methods) aimed mainly at the suitability of the initial concentration of ceftriaxone and other cultivation parameters for monitoring of the possible effects of the antibiotic on fatty acid production. The second phase was, including nitrogen starvation for the induction of lipid accumulation in the cells, carried out in glass column photobioreactor (Figure 1).
2.1. Accumulation of Chlorophyll by and Growth of the Cultures under N-Sufficient Conditions
At the pilot experiment phase (carried out in flasks), both the suspended and the chitosan-immobilized cultures demonstrated, after a 3-4 d lag, a steady accumulation of chlorophyll indicative of culture growth (Figure 2a). The suspended culture accumulated chlorophyll faster than the immobilized one until the 7th day of the experiment but later the rates of chlorophyll accumulation by the cultures were commensurate to each other. The presence of CTA in the studied concentration did not exert a measurable effect on the culture growth at the pilot experiment phase in either suspended or immobilized culture.
Surprisingly, the cultures exhibited a drastically different behavior in the second phase experiments carried out in the glass column photobioreactor in the presence of CTA (Figure 2b). Thus, the kinetics of chlorophyll accumulation by the suspended culture was close to that documented at the first phase (cf. the curves with closed squares in Figure 2a,b). By contrast, the chitosan-immobilized culture accumulated chlorophyll at a much slower rate than the suspended culture under the same experimental conditions or the immobilized culture incubated with CTA in flasks (cf. the curves with closed circles in Figure 2a,b). Notably, the decline showed by the immobilized cultures can hardly be explained by the effect of immobilization per se since it was not observed in our previous experiments with the same culture under the same experimental conditions (Figure S1).
2.2. Functioning of the Photosynthetic Apparatus of the Microalgal Cells
In view of the observed kinetics of chlorophyll accumulation, it was essential to compare the functioning of the photosynthetic apparatus in the experimental cultures (Figure 3 and Figure 4). At the first (pilot) phase, the cultures exhibited a characteristic pattern of changes in the magnitude of potential maximal photochemical quantum yield of photosystem II (Fv/Fm) which depended on the immobilization state and was only slightly affected by the presence of CTA (Figure 3a). Regardless of the cultivation conditions, the Fv/Fm did not change or even declined slightly during the first day after beginning of the experiment remaining at the level of ca. 0.60, likely due to the mild stress caused by the culture handling during the inoculation of the flasks and/or cell carriers. Later, Fv/Fm in the immobilized cultures increased to 0.65–0.70, likely due to accumulation of chlorophyll and an (Figure 2a) increase in cell self-shading resulting from it. Notably, the lag in Fv/Fm attributable to acclimation of the photosynthetic apparatus of the cells after inoculation was considerably longer in the suspended cultures (around 8 days, circles in Figure 3a). This effect cab be tentatively attributed to lower self-shading in the suspended culture and hence to higher effective per-cell light intensity in this case.
Expectedly, the magnitude of the NPQ parameter indicative of the degree of engagement of the photoprotective mechanisms in the cells followed the opposite trend as compared with that of Fv/Fm (cf. Figure 3a,b). Thus, the initial decline in Fv/Fm corresponded to an increase in NPQ suggesting that the thermal dissipation of the light energy absorbed in excess under the stress condition was engaged by the cells under our experimental conditions. Later, after successful acclimation of the cells, relaxation of NPQ was observed. As with Fv/Fm, the immobilized cells showed a faster decline in NPQ than the suspended cells.
None of the parameters was profoundly affected by the presence of 20 mg/L CTA. The only exception was comprised by the immobilized culture around the 4th day of the experiment when Fv/Fm of the immobilized culture with CTA was significantly lower (and NPQ was correspondingly higher) than that of the immobilized culture incubated without the antibiotic (cf. curves with squares in Figure 3a,b).
Similar trends of changes in Fv/Fm and NPQ was observed in the second phase (in the cultures incubated in the photobioreactor; Figure 4). The synchronous spike of NPQ observed during the first 1-2 days of incubation in the photobioreactor of both suspended and immobilized cultures (Figure 4b) was likely associated with the acclimation of the microalgal cells to a different illumination condition but not with the presence of antibiotic (see below).
Overall, CTA in the studied concentration did not exert dramatic effects of the functioning of photosynthetic apparatus of the studied strain of Lobosphaera sp. Notably, there was no sizeable effect of the antibiotic on the capacity of the cells to photochemically convert and/or thermally dissipate the absorbed light energy even in the case where severe retardation of chlorophyll accumulation took place (the immobilized culture incubated with CTA in the photobioreactor).
2.3. Effects of the Immobilization and CTA on the Culture Microbiome
At the first phase of the experiments, DNA metabarcoding data have been obtained to reveal possible effects of the treatments on the culture microbiome. The results revealed the presence of 120 genera of prokaryotic organisms the cultures. Comparison of the taxonomic composition at the level of the most represented genera (Figure 5a) showed that the microbiomes of these samples changed significantly both in time (7 and 14 d) and depending on the treatment. At the end of the first week, the genus-level diversity of the sample was relatively narrow. By the end of the second week, the diversity of represented genera increased, but microbiomes of the same culture types (i.e., suspended or immobilized) were similar regarding of CTA presence. The microbial diversity of the cell-free samples of chitosan incubated with CTA did not change considerably during the two-week incubation.
The results of DNA metabarcoding were further analyzed in terms of biodiversity indices (Figure 5b). At the end of the first week of incubation, the lowest biodiversity was observed in the immobilized culture incubated with CTA, and the highest biodiversity of the microbiome was exhibited by the suspended cultures without the antibiotic, at the biodiversity of the suspended cultures with CTA was on an intermediate level. This trend largely remained by the end of the experiment.
Along with the assessment of inter-variant biodiversity, the uniformity of the distribution of taxonomic groups was assessed with the Simpson’s index. The highest value of this index was found in the immobilized culture incubated with CTA for 7 d evident of the domination of certain groups in the microbiome. The lowest value of the Simpson’s index was observed in the suspended cultures with no CTA added after 14 d manifesting the even contributions of different taxonomic groups into the microbiome of these cultures.
Of special note are inverse trends of the Shannon-Weaver and Simpson indices during the experiment arising likely due to immobilization and/or the presence of CTA. The incubation with CTA resulted in a narrowing of the microbial diversity within the first week. It is possible to think that the microbiome in this case was overtaken by the organisms with resistance to CTA or those gaining a competitive advantage after displacement of the microbiome composition by the antibiotic. A lower biodiversity of the immobilized cultures’ microbiome may indicate that a selection of the microorganisms regarding their capacity of immobilization on the carrier takes place. One cannot exclude the potentiating effect of CTA due to its concentrating by the carrier (see section 2.4).
To visualize the degree of similarity of the culture microbiome in different experimental treatments, the Morishita β-diversity index values were calculated and visualized as heat map (Figure 5c) showing three spectacular patterns. The first included the samples incubated for 7 d, where suspended and immobilized culture possessed peculiar microbiome compositions. The second pattern emerged from comparing of the samples obtained after 7 d and 14 d of cultivation, in which only suspended cultures possessed a similar microbiome composition regardless of CTA presence. The third pattern was represented by the increase in overall similarity of the microbiomes from different cultures regardless of immobilization and CTA.
2.4. Kinetics of Changes in the Residual CTA Concentration in the Medium
For a more confident interpretation of the results reflecting microalgal culture condition, we monitored (by UPLC-ESI-MS) the actual changes of the antibiotic amount added to the medium in the beginning of the experiment (Figure 6). Since CTA is known to degrade in the presence of oxygen even without participation of biological agents, a decline in its concentration in the medium lacking cells and chitosan carrier was monitoring under conditions in which the cultures were incubated (light, bubbling, etc.). Regardless of the cultivation conditions, the abiotic degradation of CTA accounted for ca. 20% of the initially added amount of the antibiotic (see curves with open circles in Figure 6).
Our preliminary studies showed that CTA is readily adsorbed on the chitosan-based cell carriers. Therefore, the adsorption of CTA was separately monitored in un-inoculated flasks or columns containing the same amount of the carrier as was used in the experimental variant with the immobilized microalgae. Our estimations showed that cell carrier itself can adsorb up to 45% or 60% of the added CTA during incubation in the flasks or in the columns, respectively; see curves with open squares in Figure 6a and 6b (these estimations also include abiotic oxidation of CTA). The most rapid decline of the CTA concentration in the medium took place during the first 1-2 d of the experiment, later the trend of CTA removal from the medium slowed down or levelled off.
Obviously, abiotic oxidation and the adsorption on the carrier were the main processes contributing to the observed removal of CTA from the medium. Relatively small a proportion of the removed CTA (several percents of the amount initially added to culture) can be attributed to the microalgal cells. However, it is difficult to say was it due to internalization of CTA with its subsequent metabolization or just from adsorption on the cell surface. Indirect evidence of the predominance of the latter process can be seen in Figure 5: the suspended cultures which showed rapid growth (see Figure 2) also continued to remove CTA from the medium until the end of the observation period (closed circles in Figure 5a,b). The immobilized cultures showed a limited capacity for CTA removal after initial rapid decline of the antibiotic concentration (closed squares in Figure 6a,b). Nevertheless, the bulk removal of CTA added at the concentration of 20 mg/L was, under our experimental conditions, as high as 65% (in the flasks) or 85% (in the bubbled columns).
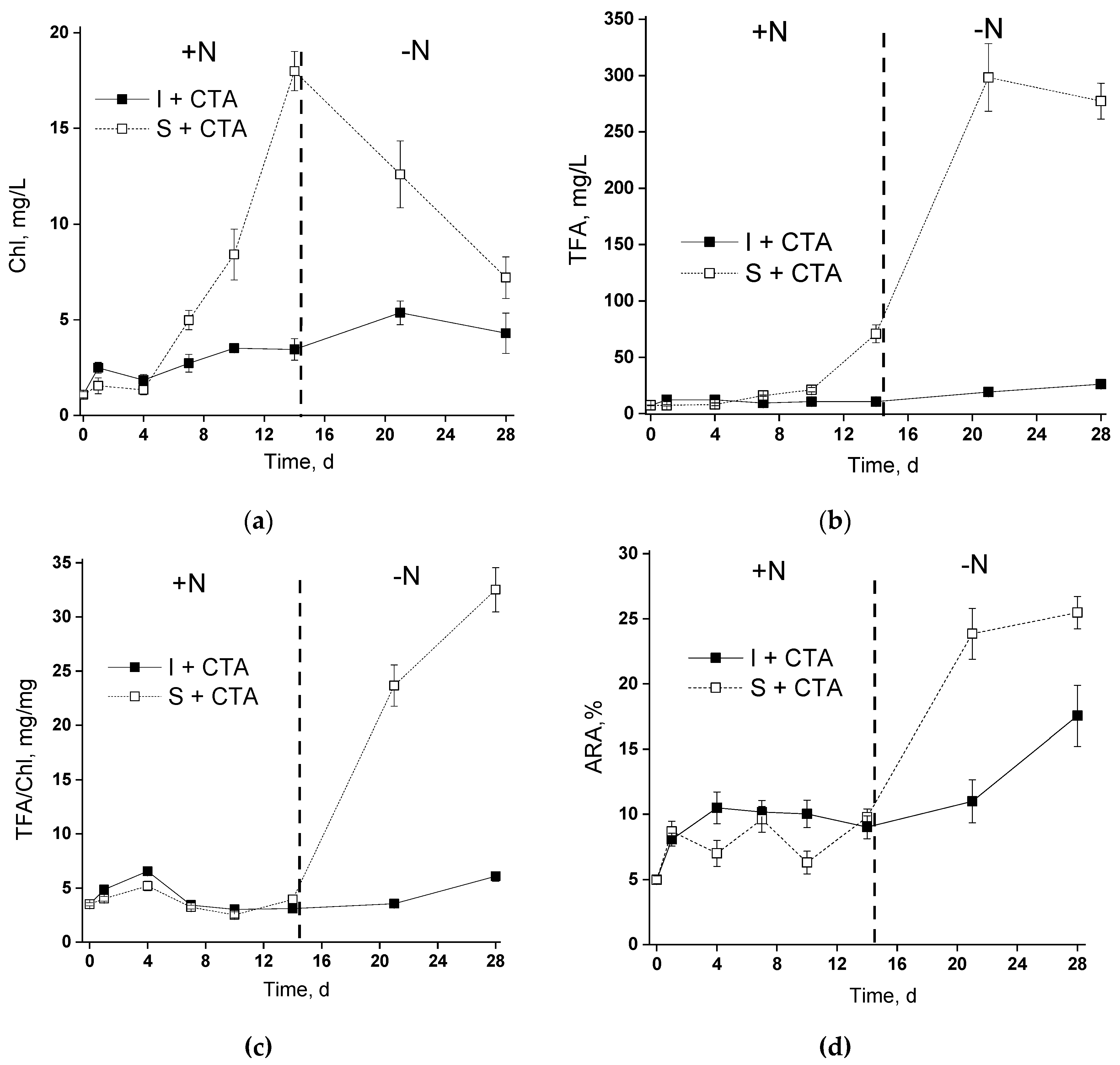
2.5. Induction of Lipid Accumulation in Microalgal Cells by Their Nitrogen Deprivation
The induction of lipid accumulation was accomplished by depriving of the microalgal cells grown in the photobioreactor (see Figure 1), either suspended or immobilized on the chitosan-based carrier (Figure 7). The nitrogen starvation was induced by changing the complete BG-11M medium with CTA for nitrogen-free BG-11M0 medium. The antibiotic was not added at the stage of nitrogen starvation since it can be a potential nitrogen source complicating the interpretation of the results.
In the suspended culture, N deprivation induced a sharp decline in chlorophyll content, a typical response of microalgal cells to nitrogen deprivation (Figure 7a). Interestingly, the immobilized culture featuring a slow accumulation of chlorophyll under nitrogen-sufficient conditions continued to accumulate the pigment for ca. 5 days after nitrogen deprivation (closed squares in Figure 7a).
In the nitrogen-sufficient cultures, the changes in total fatty acids (TFA) of the cell lipids (Figure 7a,b) closely followed the dynamics of chlorophyll content. As a result, volumetric TFA content increased along with chlorophyll accumulation, but TFA expressed per unit chlorophyll remained nearly constant in the N-sufficient cultures both in the immobilized and suspended N-sufficient cultures. Expectedly, the nitrogen deprivation triggered an increase in volumetric TFA and even higher increase in TFA per unit chlorophyll in the suspended culture. At the same time, only a modest increase in TFA was observed in the chitosan-immobilized microalgae. Neither nitrogen starvation nor the immobilization or CTA addition affected the qualitative profile of FA in the cell lipids of the Lobosphaera sp. (Tables S1 and S2).
The trend of changes in arachidonic acid (ARA) TFA percentage followed that of TFA (Figure 7b,c). Again, the increase in ARA percentage of TFA was more spectacular in the suspended culture than in the immobilized one (25% vs. 17%, respectively).
3. Discussion
Immobilization of the cells on biocompatible cell carriers is frequently employed in microalgal biotechnology to facilitate the biomass harvesting as well as to increase the stress resilience of microalgae and hence the culture robustness [1,4]. Chitosan-based biopolymers are highly promising cell carriers for immobilization of microalgal cells [11]. However, they can also be a source of nutrients interfering with the culture manipulation by controlled stress such as nitrogen deprivation for induction of lipids harboring valuable fatty acids. The using of attached cultivation and increased stress resilience ensued by is especially relevant for growing of microalgae in waste- and sidestreams as a source of nutrients. Many types of wastewaters (e.g., farming and municipal waste streams) contain sizeable amounts of hazardous micropollutant such as antibiotics [2,11,19]. At the same time, the information on possible effects of the presence of antibiotics on the biology of immobilized microalgal cell is scarce. In view of this, we attempted to reveal interactive effects of immobilization of the cells and a widespread antibiotic CTA on the production of a valuable polyunsaturated fatty acid ARA by its producer Lobosphaera sp. IPPAS C-2047. As a provocative test, we deliberately employed a high (20 mg/L) CTA concentration since it was shown that the Lobosphaera sp. is tolerant of CTA in the environmentally relevant concentrations around 1–3 mg/L [20]. Therefore, the first (pilot) phase of the study has been dedicated to testing the potential acute toxicity of CTA to the selected microalgal strain. Judging from the obtained results (Figures 2a, and 3), 20 mg/L CTA did not induce a profound deteriorative effect on the cells of Lobosphaera sp. cultivated in flasks regardless of the immobilization, only a slight inhibition of photosynthetic activity has been recorded in the immobilized cells at day 4 (Figure 3).
An unexpected result was represented by a severe retardation of the growth (as manifested by chlorophyll accumulation) of the chitosan-immobilized culture in the presence of CTA. Remarkably, this effect was not accompanied by severe disorders of the photosynthetic apparatus (Figure 4). A plausible explanation can be related with an increase in the concentration of CTA due to its sorption on chitosan. This effect may be not so pronounced during incubation in flasks in the absence of vigorous mixing while in the bubbled columns the rate of CTA adsorption (and hence the rate of the increase in its effective concentration in the vicinity of microalgal cells) was ca. 30% higher (open squares in Figures 6a and 6b). It seems that a sub-population of Lobosphaera sp. cells which was more resilient to increased CTA concentration was effectively selected on the chitosan-based cell carrier. These cells might have retained a functional photosynthetic apparatus and the essential responses of their lipid metabolism to nitrogen deprivation manifested as an induction of lipid accumulation.
Taking into account the results of our previous experiments on immobilization of Lobosphaera sp. on chitosan-based cell carriers one can conclude that the immobilization itself does not affect the FA profile (Tables S1 and S2), degree of lipid and, specifically, ARA induction profoundly (Figure S1, see also [19]). Typical ARA percentage of TFA of immobilized was in the range 20%–25%. Therefore, the decline in the amount TFA and ARA observed in the presence of CTA is attributable mostly to the effect of the antibiotic and not to the effect of the attachment to the carrier.
An intriguing question to be answered by this study was that about the capacity of suspended and chitosan-immobilized cultures of green microalgae such as Lobosphaera sp. to remove CTA from the medium. Comparison of the abiotic degradation of CTA and its adsorption to the chitosan-based carrier with the bulk CTA removal (Figure 6) made it clear that the removal of CTA in our system occurred mainly due to these two processes and contribution of the microalgal cells to CTA removal was minor. It is difficult to distinguish between the adsorption of CTA on the cell surface and uptake of the antibiotic by the cells. Still, one cannot rule out at least partial metabolization of CTA. Importantly, the cultures at the stage of nitrogen starvation were also affected by CTA despite replacement of the medium (see Methods) since a considerable amount of the antibiotic was adsorbed to the cell carrier (see Figure 6); likely, there was also some adsorption of CTA to the microalgal cell surface as well.
Diverse processes such as adsorption, advanced oxidation, and photocatalysis have been studied to remove antibiotics. As was summarized recently [5], the antibiotic removal efficiency by adsorption is highly adsorbent-dependent, and most of the adsorbents are expensive. Advanced oxidation and photocatalysis-based processes may be effective, but they need expensive catalysts; these processes can generate dangerous secondary pollutants. Advantages of the microalgae-based wastewater treatment process as a biological process are the absence of the need for expensive reagents and a low risk of secondary pollution [21]. Immobilization of the microalgal cells on biopolymer carriers increases efficiency of the process, also due to the sorption capacity of the carrier itself. In our experiments the CTA level was taken own to 6 mg/L or 4 mg/L.
To elucidate possible trends of changes of the Lobosphaera sp. culture microbiome, it was essential to evaluate the most represented microbial taxa and the effect of immobilization and CTA on them (Figure 5a). Notably, that all the bacterial genera discovered are typical for microalgal cultures under different cultivation conditions. Many representatives of these genera belong to the PGPB (plant growth promoting bacteria) group, such as Variovorax, Stenotrophomonas, Shinella, Devosia, Sphingomonas, etc. Representatives of microalgal parasites were not discovered. Importantly, some representatives of Variovorax, Stenotrophomonas, Flavobacterium are known to be resistant to a wide range of antibiotics, including CTA ceftriaxone, having the ability to degrade them [22,23].
Ample representation of the genera Variovorax and Stenotrophomonas in the chitosan-immobilized cultures can be due to the fact that the presence of the polymer creates favorable conditions for the development of these microorganisms. Thus, representatives of the genus Variovorax are known to be able to actively form biofilms, also on the surface of carriers [24]. Representatives of the genus Stenotrophomonas are known for the presence of chitinase and the ability to degrade chitosan [25]. At the same time, it was shown that some forms chitosan also have antimicrobial activity, particularly suppressing the growth of certain representatives of the genus Flavobacterium [26].
The decline of the difference between the microbiome in different experimental treatments can be explained by (i) decline in the CTA residual content and (ii) by acclimation and selection of the microorganisms favored by the presence of chitosan and CTA. This hypothesis is supported by gradual proliferation of the microorganisms with modest resistance to ceftriaxone (Pseudoxanthomonas, Devosia, Bosea) in the samples containing CTA. The potential inhibitory effects of chitosan for certain bacteria (e.g. from the genus Flavobacterium) was also gradually overcame, likely via formation of biofilm frequently observed on the surface of the carrier [19].
To conclude, the cultures of Lobosphaera sp., especially those immobilized on the chitosan-based cell carriers, turned to be suitable for removal of the antibiotics, especially taking into account that the CTA concentration in this study (20 mg/L) is higher than that typically encountered in the environment and even in the wastewater treatment plants. However, the attempts to couple the removal of hazardous pollutants such as antibiotics with production of valuable microalgal metabolites should use caution since toxic effects can emerge unexpectedly due to concentrating of the pollutants on the carrier. One should also be aware of the possible enrichment of the microbiome of the attached microalgal cultures with antibiotic-resistant bacteria.
4. Materials and Methods
4.1. Synthesis of the Cross-Linked Chitosan-Based Polymer
The cross-linked chitosan polymers were synthesized in the Laboratory of Polymer Materials of the Kurchatov Institute Research Center. Two mg of chitosan ChitoClear HQG 800 (Primex, Island; molecular weight of 600 kDa) were dissolved in 98 mL of 2% aqueous solution of acetic acid (LLC “Component-Reactiv”, Russia). The mixture was homogenized using a magnetic stirrer (Heidolph MR Hei-Tec, USA) for 24 h at 23 °C. The chitosan-based carrier polymers were synthesized from the solution of chitosan and glutaraldehyde as cross-linking agent using lyophilization technique. An aliquot of 0.76 mL of 2.5% aqueous solution of glutaraldehyde was added to the chitosan mixture. After 15-min stirring with a magnetic stirrer the mixture was placed in 12-well plates (2.7 mL per well), incubated in a freezer (24 h at –25 °C), and lyophilized using ALPHA 1-4LSC lyophilizer (Martin Christ, Germany) at 0.250 mbar for 24 h [27].
The resulting carriers in the form of porous discs (14 mm diameter, 10 mm thickness) were used in the experiments.
4.2. Strain and Cultivation Conditions
Lobosphaera sp. IPPAS C-2047 used in the presented investigation as the model microalga was obtained from the Culture Collection of K.A. Timiryazev Institute of Plant Physiology (IPPAS, Russian Academy of Science).
Preculture of Lobosphaera sp. IPPAS C-2047 was grown at 20 °C or 23 °C in a shaker incubator (New Brunswick, Innova-44R, N.Y., USA) at 120 rpm in 0.75 L flasks with 0.3 L of P-enriched modified BG-11 [28] medium, BG-11M (g/L: NaNO3 = 0.74, KNO3 = 0.9, K2HPO4 = 0.181, KH2PO4 = 0.089, MgSO4·7H2O = 0.075, CaCl2·2H2O = 0.036, citric acid = 0.006, ferric ammonium citrate = 0.006, Na2EDTA·2H2O = 0.001, Na2CO3 = 0.02, BG–11 trace metal solution) at 40 μmol PAR (photons/m2/s) provided by daylight fluorescent tubes (Philips TL-D 36W/54-765) as measured with a LiCor 850 quantum sensor (LiCor, Lincoln, NE, USA) and the atmospheric CO2 level. Culture pH was measured aseptically with a bench-top pH-meter (Hanna Instruments, Ann Arbor MI, USA).
To obtain the suspended cultures for the first phase (pilot) experiments, equal aliquots of the Lobosphaera sp. pre-culture were pelleted by centrifugation (5 min, 1000× g) and resuspended to ca. 1.5 mg chlorophyll/L in BG-11M medium or in the same medium supplemented with CTA (LLC “Biosintez,” Penza, Russia) to the final concentration of 20 mg/L and placed in 100-mL vented cultivation flasks (TPP, Switzerland).
In the second phase (bioreactor tests), the microalgal cell suspension was placed into the glass columns (600 mL; 4 cm i.d.; Figure 1) containing 400 mL of the cell suspension. The columns were incubated in a temperature-controlled water bath at 27 °C with constant bubbling with air passed through a 0.22 μm bacterial filter (Merck-Millipore, Billerica, MA, USA) and delivered at a rate of 300 mL/min (STP) at 40 μmol PAR (photons/m2/s).
Immobilization of the cells has been carried out as follows. A group of the pre-culture aliquots was gently re-suspended in BG-11M and pipetted evenly on the surface of the chitosan discs to carry out the immobilization of the cells (the final concentration of chlorophyll was ca. 24 µg/disc). For the pilot experiments, the chitosan discs with firmly attached Lobosphaera sp. cells were placed into the 100-mL vented-cap cultivation flasks (TPP, Switzerland) containing BG-11M medium supplemented with 20 mg/L of CTA or BG-11M lacking the antibiotic. Fir the second phase experiments (photobioreactor tests), the chitosan discs with attached Lobosphaera sp. cells were fixed on the in-house made stainless steel spikes within the bioreactor column (Figure 1) containing BG-11M medium pr the same medium supplemented with CTA to the final concentration of 20 mg/L.
At the first phase (pilot experiments), the flasks with the suspended or immobilized Lobosphaera sp. cells were cultured for 14 days on the Innova-44R incubator shaker at 120 rpm under the condition specified above. At the second phase, the columns were incubated in the bioreactor under the conditions described above.
To induce nitrogen starvation for lipid accumulation, the complete BG-11M medium in the columns of the photobioreactor was replaced with nitrogen-free medium (BG-11M0) after 14 d of cultivation. Specifically, the suspended cells were gently harvested by centrifugation (2 min, 1000× g), washed with BG-11M–N medium and re-suspended in the same medium. The immobilized cultures were transferred into the identical columns with BG-11M–N medium. All the cultures were incubated in the photobioreactor for another 14 days at 200 μmol PAR (photons/m2/s); all the other conditions were as specified above.
4.3. Chlorophyll and Nutrients Assay
Growth estimation was based on the chlorophyll content measurements. For chlorophyll assay determination, an aliquot of the cell suspension was sampled and harvested by centrifugation for 5 min at 3000 g. Total Chl were extracted by heating the cell pellet with 2 mL of dimethyl sulfoxide (DMSO) for 10 min at 70 °C. From the cells attached to the carrier, chlorophyll was extracted by adding 2 mL of DMSO to the carrier with the cells and heating for 10 min at 70 °C. Concentration of chlorophyll was determined in the DMSO extracts with an Agilent Cary 300 spectrophotometer (Walnut Creek, CA, USA) using equations reported in [29].
The residual phosphate and nitrate contents were checked using Thermo Dionex ICS 1600 HPLC (Sunnyvale, CA, USA) with a conductivity detector and IonPac AS12A anionic analytical column (5μm; 2 × 50 mm). The column temperature was maintained at 30 °C. The ions were eluted isocratically with 2.7 mM sodium carbonate/0.3 mM sodium bicarbonate buffer (flow rate of 0.3 mL/min).
4.4. Photosynthetic Activity and Photoprotective Mechanism Probing
Viability of the immobilized and suspended Lobosphaera cells was assumed to be reflected by photosynthetic activity of the cells. were monitored with a FluorCam FC 800-C (PSI, Drasov, Czech Republic) kinetic fluorescence imager. The chitosan carriers with immobilized cells or 5-mL aliquots of suspended culture were transferred into well plates and sealed aseptically. Estimations of the photosynthetic activity of the microalgal cells were obtained by recording Chl a fluorescence induction curves The recorded curves were processed by the built-in software of the fluorometer, and the following parameters indicative of the functional condition of the photosynthetic apparatus of the microalgal cells were calculated: potential maximum photochemical quantum yield of photosystem II, Qy = (Fm – Fo)/Fm = Fv/Fm, and non–photochemical quenching of the electron excitation energy in the light-harvesting antenna (parameter NPQ, NPQ = Fm/Fm' – 1) [30]. The PAM measurements were carried out after 10-min dark adaptation. Chlorophyll fluorescence was excited at 650 nm and recorded in the red region of the spectrum (λ > 680 nm). After the measurements, the samples were returned to the corresponding cultivation flacks or the bioreactor column under aseptic conditions. This procedure allowed repeated assessments of the attached and suspended cells under our experimental conditions.
4.5. Assay of Cell Lipid Fatty Acid Composition
Total cell lipids were extracted from these samples according to Folch [31]: the cell pellets or the cells on the carrier were homogenized in a mixture of chloroform and methanol (2:1, by volume). Distilled water was added to the homogenate in the amount of 20% of the homogenate volume, the mixture was dark incubated overnight at 4 °C, and centrifuged (10 min, 3000 g) until phase separation. The chloroform phase of the extract was evaporated to dryness on a Heidolph Laborota 4000 rotary evaporator (Heidolph, Schwabach, Germany) at 30 °C. As an internal standard, 50 micrograms of margaric acid (C17:0) were added to the samples. The samples were transmethylated by incubation with 2% sulfuric acid in methanol for 1.5 h at 80 °C. Fatty acid (FA) methyl esters were extracted with 2 ml of n-hexane. In the case of immobilized cells, the carriers with the cells were ground and extracted as described above.
The fatty acid (FA) profile of total cell lipids was analyzed using an Agilent 7890A gas chromatograph (Agilent Technologies, Santa Clara, CA, USA) equipped with a 30-meter HP-5MS UI capillary column (30 m × 0.25 mm × 0.25 microns; Agilent, USA). Helium with a flow rate of 1 mL/min was used as a carrier gas (for more details, see [32]).
The proportion of individual FA in the total FA of the Lobosphaera sp. cell lipids was inferred from the corre-sponding peak area. The absolute FA contents were calculated relative to the internal standard (C17:0) peak area.
4.6. UPLC-ESI-MS Assay of Ceftriaxone
The lyophilized culture broth samples were dissolved in methanol-water mixture (1:1, by vol.) and centrifuged (15 min, 15 000 rpm; centrifuge ELMI CM-50, Latvia). The samples were chromatographed on an ACQUITY UPLC H-Class PLUS (Waters, USA) equipped with a time-of-flight mass-selective detector Xevo G2-XS TOF (Waters, USA). An 0.5 µL sample aliquot was injected on a Titan C18 (100 × 2.1 mm, 1.9 µm; Supelco, Bellefonte, CA, USA) column maintained at 50°С and eluted with a gradient of 10 mM solution of ammonium acetate in deionized (Simplicity UV, Millipore, France) water (solvent A) and LC-MS grade (Panreac, Spain) acetonitrile (solvent B) delivered at a rate of 0.5 mL/min. The following gradient program was used (vol. % of the solvent B): 0–1 min, from 5% to 15%; 1–2 min, from 15% to 65%; 2–3 min, from 65% to 85%; 3–5 min, from 85% to 95% (see also [33]). The analysis was carried out in the negative ion detection mode (m/z range of 100–1900), the ion source parameters were: ion source temperature—150 °С, desolvation temperature—650°С; capillary voltage—3.0 kV; sample injection cone voltage—30 V, nitrogen (desolvation gas) flow rate—1101 L/h. The recorded data were processed with MassLynx software (Waters, USA) and CTA was quantified using external calibration method.
4.7. Taxonomical Profiling of the Culture Microbiome
Cell pellet was collected from 2–5 mL of each freshly taken sample by centrifugation (8000 × g) for 10 min and immediately frozen at –80°C and stored until analysis. For metagenomic DNA (mgDNA) extraction, the cell pellets were frozen in liquid nitrogen and ground to a fine powder using homogenizing pestles (SSIBio Corp., Lodi, CA, USA) in a 1.5 ml microcentrifuge tube. The freezing-homogenization procedure was repeated three times for each sample. mgDNA was isolated from the obtained homogenates using the DNeasy Plant Pro kit (QIAGEN, Hilden, Germany) according to the manufacturer's instructions, replacing the sample mixing procedure on the Vortex with manual mixing by shaking the tube.
Amplification of the 16S rRNA gene fragment with the V4 hypervariable region and preparation of libraries for sequencing was carried out as described previously [34] using oligonucleotide primers F515 (5'-gtgccagcmgccgcggtaa-3') and R806 (5'-ggactacvsgggtatctaat-3 ′) [35]. Sequencing was performed on a MiSeq instrument (Illumina, USA) using a MiSeq Reagent Kit v3 (600-cycle) (part number MS-102-3003, Illumina, USA) for paired-end read (2 × 300 bp). Initial data processing—sample demultiplexing and removal of adapters—was carried out using the Illumina software (Illumina, USA). Further denoising, sequence merging, restoration of original phylotypes (ASV, Amplicon sequence variant), deletion of chimeric reads, and taxonomic classification of the resulting ASVs were performed in the R software environment using the dada2 [36], phyloseq [37], and DECIPHER [38] packages, as well as the SSU 16s rRNA SILVA database (release 132) [39]. When analyzing the results of DNA metabarcoding, unclassified readings were removed from the samples, as well as those taxonomic groups that corresponded to one unpaired reading. For the number of readings, the average value was calculated for two biological replicates, for each of which two technical replicates were made. The Shannon-Weaver a-diversity (H) and Morishita β-diversity indices were calculated [40]. Morishita index values were visualized using an algorithm in the Python programming language (version 3.7.1) using the Matplotlib library.
4.8. Statistical Treatment
Under the specified conditions, three independent experiments were carried out for each treatment repeated in triplicate. The average values (n = 9) and corresponding standard deviation are shown unless stated otherwise. The significance of difference if the average values were analyzed using Student’s t test.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1 demonstrating that the accumulation of arachidonic acid in the immobilized and suspended cultures Lobosphaera sp. IPPAS C-2047 is similar ranging from 17 to 30% of the total fatty acids; Tables S1 and S2 with fatty acid profile of the suspended or immobilized cells of Lobosphaera sp. IPPAS C-2047 during their incubation in complete and nitrogen-free BG-11M medium.
Author Contributions
Conceptualization, S.V. and A.S.; methodology, T.G., C.A., and S.V.; validation, O.C. and O.G.; formal analysis, S.V., D.K., and P.Z.; investigation, S.V., A.L., P.Z., O.G., C.A., T.F., P.S., O.C., and A.S.; writing—original draft preparation, A.S. and S.V.; writing—review and editing, A.S.; visualization, S.V. and P.Z.; supervision, E.L.; project administration, A.S. and E.L.; funding acquisition, A.S. and S.V. All authors have read and agreed to the published version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by the Russian Science Foundation (grant 21-74-20004, cultivation of the microalgae in the presence of the antibiotic in the medium). UPLC-ESI-MS analysis was supported by the Ministry of Science and Higher Education of the Russian Federation themes No. 122042600086-7 and No. 122042700045-3. The rest of the investigations were supported by the Ministry of Science and Higher Education of the Russian Federation (grant 075-15-2021-1396).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are available from the corresponding author upon a reasonable request.
Acknowledgments
The photosynthetic activity of microalgae was estimated using the Shared Research Facility “Phototrophic Organism Phenotyping.”
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Moreno-Garrido, I. Microalgae immobilization: current techniques and uses. Bioresource technology 2008, 99, 3949–3964. [Google Scholar] [CrossRef] [PubMed]
- Vasilieva, S.; Lobakova, E.; Solovchenko, A. Biotechnological Applications of Immobilized Microalgae. Environmental Biotechnology 2021, 3, 193–220. [Google Scholar]
- Martins, S.C.S.; Martins, C.M.; Fiúza, L.M.C.G.; Santaella, S.T. Immobilization of microbial cells: A promising tool for treatment of toxic pollutants in industrial wastewater. African journal of biotechnology 2013, 12. [Google Scholar]
- de-Bashan, L.E.; Bashan, Y. Immobilized microalgae for removing pollutants: review of practical aspects. Bioresource Technology 2010, 101, 1611–1627. [Google Scholar] [CrossRef] [PubMed]
- Leng, L.; Wei, L.; Xiong, Q.; Xu, S.; Li, W.; Lv, S.; Lu, Q.; Wan, L.; Wen, Z.; Zhou, W. Use of microalgae based technology for the removal of antibiotics from wastewater: A review. Chemosphere 2020, 238, 124680. [Google Scholar] [CrossRef]
- Simon, M.; Joshi, H. A review on green technologies for the rejuvenation of polluted surface water bodies: Field-scale feasibility, challenges, and future perspectives. Journal of Environmental Chemical Engineering 2021, 9. [Google Scholar] [CrossRef]
- Xiong, J.-Q.; Kurade, M.B.; Jeon, B.-H. Can microalgae remove pharmaceutical contaminants from water? Trends in biotechnology 2018, 36, 30–44. [Google Scholar] [CrossRef]
- Villar-Navarro, E.; Baena-Nogueras, R.M.; Paniw, M.; Perales, J.A.; Lara-Martín, P.A. Removal of pharmaceuticals in urban wastewater: High rate algae pond (HRAP) based technologies as an alternative to activated sludge based processes. Water research 2018, 139, 19–29. [Google Scholar] [CrossRef]
- Guo, J.; Selby, K.; Boxall, A.B. Effects of Antibiotics on the Growth and Physiology of Chlorophytes, Cyanobacteria, and a Diatom. Archives of environmental contamination and toxicology 2016, 71, 589–602. [Google Scholar] [CrossRef]
- Vasilieva, S.; Lobakova, E.; Grigoriev, T.; Selyakh, I.; Semenova, L.; Chivkunova, O.; Gotovtsev, P.; Antipova, C.; Zagoskin, Y.; Scherbakov, P. , et al. Bio-inspired materials for nutrient biocapture from wastewater: Microalgal cells immobilized on chitosan-based carriers. Journal of Water Process Engineering, 2020. [Google Scholar] [CrossRef]
- Muniyappan Gandhi, R. Recent advances in chitosan based biosorbent for environmental clean-up. J Bioremediat Biodegrad 2016, 7, e173. [Google Scholar]
- Hiller, C.X.; Hübner, U.; Fajnorova, S.; Schwartz, T.; Drewes, J.E. Antibiotic microbial resistance (AMR) removal efficiencies by conventional and advanced wastewater treatment processes: A review. Science of The Total Environment 2019, 685, 596–608. [Google Scholar] [CrossRef] [PubMed]
- Manaia, C.M.; Rocha, J.; Scaccia, N.; Marano, R.; Radu, E.; Biancullo, F.; Cerqueira, F.; Fortunato, G.; Iakovides, I.C.; Zammit, I. , et al. Antibiotic resistance in wastewater treatment plants: Tackling the black box. Environ Int 2018, 115, 312–324. [Google Scholar] [CrossRef] [PubMed]
- Bouki, C.; Venieri, D.; Diamadopoulos, E. Detection and fate of antibiotic resistant bacteria in wastewater treatment plants: a review. Ecotoxicol Environ Saf 2013, 91, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Yoon, Y.; Ngo, H.H.; Jang, A. The application of microalgae in removing organic micropollutants in wastewater. Critical Reviews in Environmental Science and Technology 2021, 51, 1187–1220. [Google Scholar] [CrossRef]
- Bigogno, C.; Khozin-Goldberg, I.; Cohen, Z. Accumulation of arachidonic acid-rich triacylglycerols in the microalga Parietochloris incisa (Trebuxiophyceae, Chlorophyta). Phytochemistry 2002, 60, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Crawford, M.; Golfetto, I.; Ghebremeskel, K.; Min, Y.; Moodley, T.; Poston, L.; Phylactos, A.; Cunnane, S.; Schmidt, W. The potential role for arachidonic and docosahexaenoic acids in protection against some central nervous system injuries in preterm infants. Lipids 2003, 38, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.G.; Sargent, J.R. Arachidonic acid in aquaculture feeds: current status and future opportunities. Aquaculture 2003, 218, 491–499. [Google Scholar] [CrossRef]
- Vasilieva, S.; Shibzukhova, K.; Solovchenko, A.; Chivkunova, O.; Antipova, C.; Morozov, A.; Lobakova, E. Immobilization on polyethylenimine and chitosan sorbents modulates the production of valuable fatty acids by the chlorophyte Lobosphaera sp. IPPAS C-2047. Journal of Marine Science and Engineering 2023, 11, 865. [Google Scholar] [CrossRef]
- Solovchenko, A.E.; Vasilieva, S.G.; Zaitsev, P.; Lukyanov, A.A.; Skripnikova, E.V.; Antal, T.K. Approaches to rapid screening of pharmaceutical xenobiotic effects on microalgae via monitoring of photosynthetic apparatus condition. Journal of Applied Phycology 2022, 34, 353–361. [Google Scholar] [CrossRef]
- Encarnação, T.; Palito, C.; Pais, A.A.; Valente, A.J.; Burrows, H.D. Removal of pharmaceuticals from water by free and imobilised microalgae. Molecules 2020, 25, 3639. [Google Scholar] [CrossRef]
- Carlson, J.M.; Leonard, A.B.; Hyde, E.R.; Petrosino, J.F.; Primm, T.P. Microbiome disruption and recovery in the fish Gambusia affinis following exposure to broad-spectrum antibiotic. Infect Drug Resist 2017, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.-C.; Hsueh, P.-R.; Wu, J.-J.; Ho, S.-W.; Hsieh, W.-C.; Luh, K.-T. Antimicrobial susceptibility of flavobacteria as determined by agar dilution and disk diffusion methods. Antimicrobial agents and chemotherapy 1997, 41, 1301–1306. [Google Scholar] [CrossRef] [PubMed]
- Pehl, M.J.; Jamieson, W.D.; Kong, K.; Forbester, J.L.; Fredendall, R.J.; Gregory, G.A.; McFarland, J.E.; Healy, J.M.; Orwin, P.M. Genes that influence swarming motility and biofilm formation in Variovorax paradoxus EPS. PloS one 2012, 7, e31832. [Google Scholar] [CrossRef]
- Jankiewicz, U.; Brzezinska, M.S. Purification, characterization, and gene cloning of a chitinase from Stenotrophomonas maltophilia N4. Journal of Basic Microbiology 2015, 55, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.N.; Al-Saadi, T.M.; AL-Faragi, J.K. Effect of Chitosan nanoparticles loaded oxytetracycline hydrochloride on health status of common carp (Cyprinuscarpio L. ) infected with columnaris disease. In Proceedings of Journal of Physics: Conference Series; p. 032075.
- Romanova, O.; Grigor’ev, T.; Goncharov, M.; Rudyak, S.; Solov’yova, E.; Krasheninnikov, S.; Saprykin, V.; Sytina, E.; Chvalun, S.; Pal’tsev, M. Chitosan as a modifying component of artificial scaffold for human skin tissue engineering. Bulletin of experimental biology and medicine 2015, 159, 557–566. [Google Scholar] [CrossRef]
- Stanier, R.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Microbiology and Molecular Biology Reviews 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Solovchenko, A.; Merzlyak, M.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. Coordinated carotenoid and lipid syntheses induced in Parietochloris incisa (Chlorophyta, Trebouxiophyceae) mutant deficient in Δ5 desaturase by nitrogen starvation and high light. Journal of Phycology 2010, 46, 763–772. [Google Scholar] [CrossRef]
- Strasser, R.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a fluorescence: a signature of photosynthesis, Papageorgiou, G., Govindjee, Eds. Springer: 2004; pp. 321–362.
- Folch, J.; Lees, M.; Sloane-Stanley, G. A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Solovchenko, A.; Khozin-Goldberg, I.; Recht, L.; Boussiba, S. Stress-Induced Changes in Optical Properties, Pigment and Fatty Acid Content of Nannochloropsis sp.: Implications for Non-destructive Assay of Total Fatty Acids. Marine Biotechnology 2011, 13, 527–535. [Google Scholar] [CrossRef]
- Wongchang, T.; Winterberg, M.; Tarning, J.; Sriboonvorakul, N.; Muangnoicharoen, S.; Blessborn, D. Determination of ceftriaxone in human plasma using liquid chromatography–tandem mass spectrometry. Wellcome Open Research 2022, 4, 47. [Google Scholar] [CrossRef]
- Kublanovskaya, A.; Chekanov, K.; Solovchenko, A.; Lobakova, E. Cyanobacterial diversity in the algal–bacterial consortia from Subarctic regions: new insights from the rock baths at White Sea Coast. Hydrobiologia 2019, 830, 17–31. [Google Scholar] [CrossRef]
- Bates, S.T.; Berg-Lyons, D.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. Examining the global distribution of dominant archaeal populations in soil. The ISME journal 2011, 5, 908–917. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLOS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Wright, E.S. Using DECIPHER v2. 0 to analyze big biological sequence data in R. R J. 2016, 8, 352. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nature methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Horn, H.S. Measurement of" overlap" in comparative ecological studies. The American Naturalist 1966, 100, 419–424. [Google Scholar] [CrossRef]
Figure 1.
The suspended (left) and immobilized (right) cultures of Lobosphaera incisa IPPAS C-2047 incubated in the photobioreactor (see Methods).
Figure 1.
The suspended (left) and immobilized (right) cultures of Lobosphaera incisa IPPAS C-2047 incubated in the photobioreactor (see Methods).

Figure 2.
Changes in volumetric content of chlorophyll in the suspended (S) and immobilized (I) Lobosphaera sp. IPPAS C-2047 cultures during (a) pilot experiment in the flasks and (b) in the column photobioreactor during their incubation with (I+CTA or S+CTA, respectively) or without 20 mg/L ceftriaxone (I or S, respectively).
Figure 2.
Changes in volumetric content of chlorophyll in the suspended (S) and immobilized (I) Lobosphaera sp. IPPAS C-2047 cultures during (a) pilot experiment in the flasks and (b) in the column photobioreactor during their incubation with (I+CTA or S+CTA, respectively) or without 20 mg/L ceftriaxone (I or S, respectively).
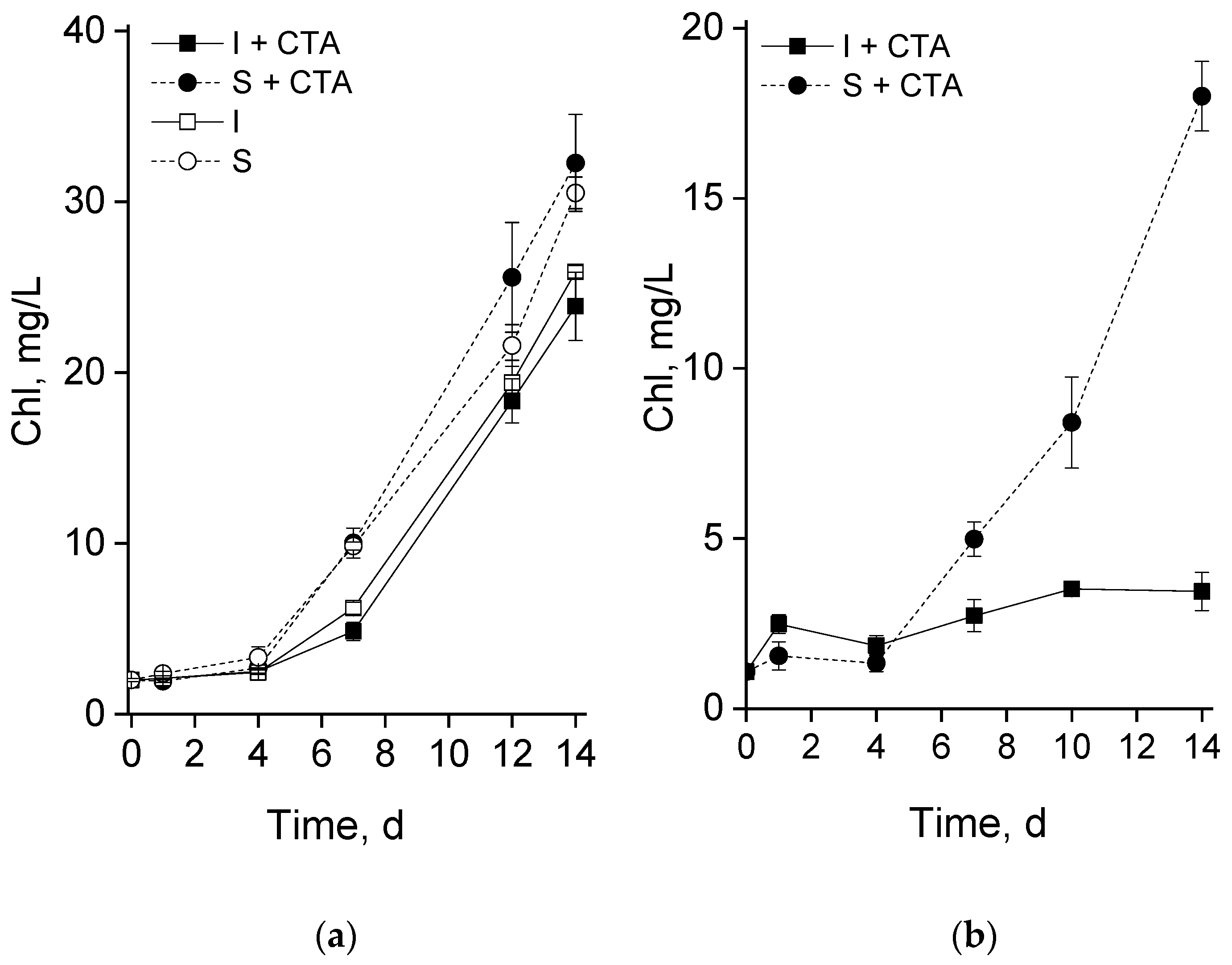
Figure 3.
Changes in (a) potential maximal photochemical quantum yield of photosystem II (Fv/Fm) and (b) Stern-Volmer non-photochemical quenching of chlorophyll excitation in the suspended and immobilized Lobosphaera sp. IPPAS C-2047 cultures during pilot experiment in the flasks during their incubation with or without 20 mg/L ceftriaxone. For curve designations, see the legend to Figure 2.
Figure 3.
Changes in (a) potential maximal photochemical quantum yield of photosystem II (Fv/Fm) and (b) Stern-Volmer non-photochemical quenching of chlorophyll excitation in the suspended and immobilized Lobosphaera sp. IPPAS C-2047 cultures during pilot experiment in the flasks during their incubation with or without 20 mg/L ceftriaxone. For curve designations, see the legend to Figure 2.
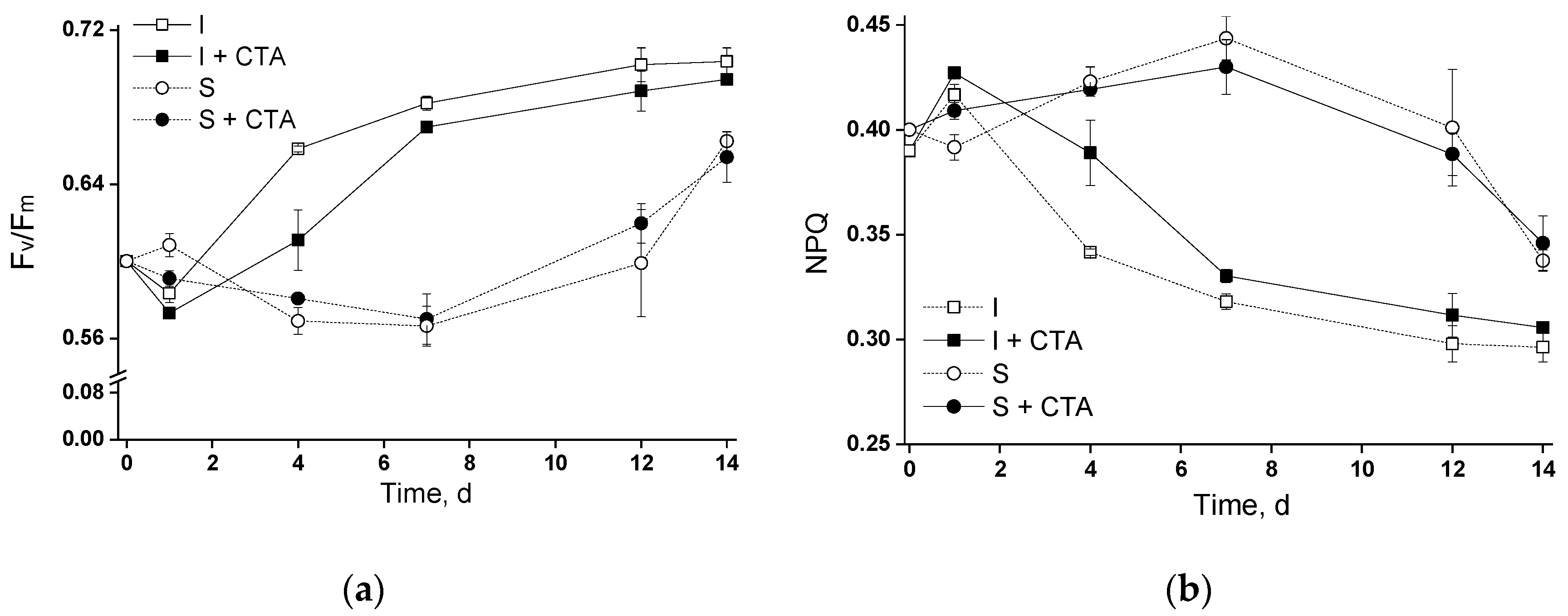
Figure 4.
Changes in (a) potential maximal photochemical quantum yield of photosystem II (Fv/Fm) and (b) Stern-Volmer non-photochemical quenching of chlorophyll excitation in the suspended and immobilized Lobosphaera sp. IPPAS C-2047 cultures during the experiment in the column photobioreactor during their incubation with or without ceftriaxone. For curve designations, see the legend to Figure 2.
Figure 4.
Changes in (a) potential maximal photochemical quantum yield of photosystem II (Fv/Fm) and (b) Stern-Volmer non-photochemical quenching of chlorophyll excitation in the suspended and immobilized Lobosphaera sp. IPPAS C-2047 cultures during the experiment in the column photobioreactor during their incubation with or without ceftriaxone. For curve designations, see the legend to Figure 2.
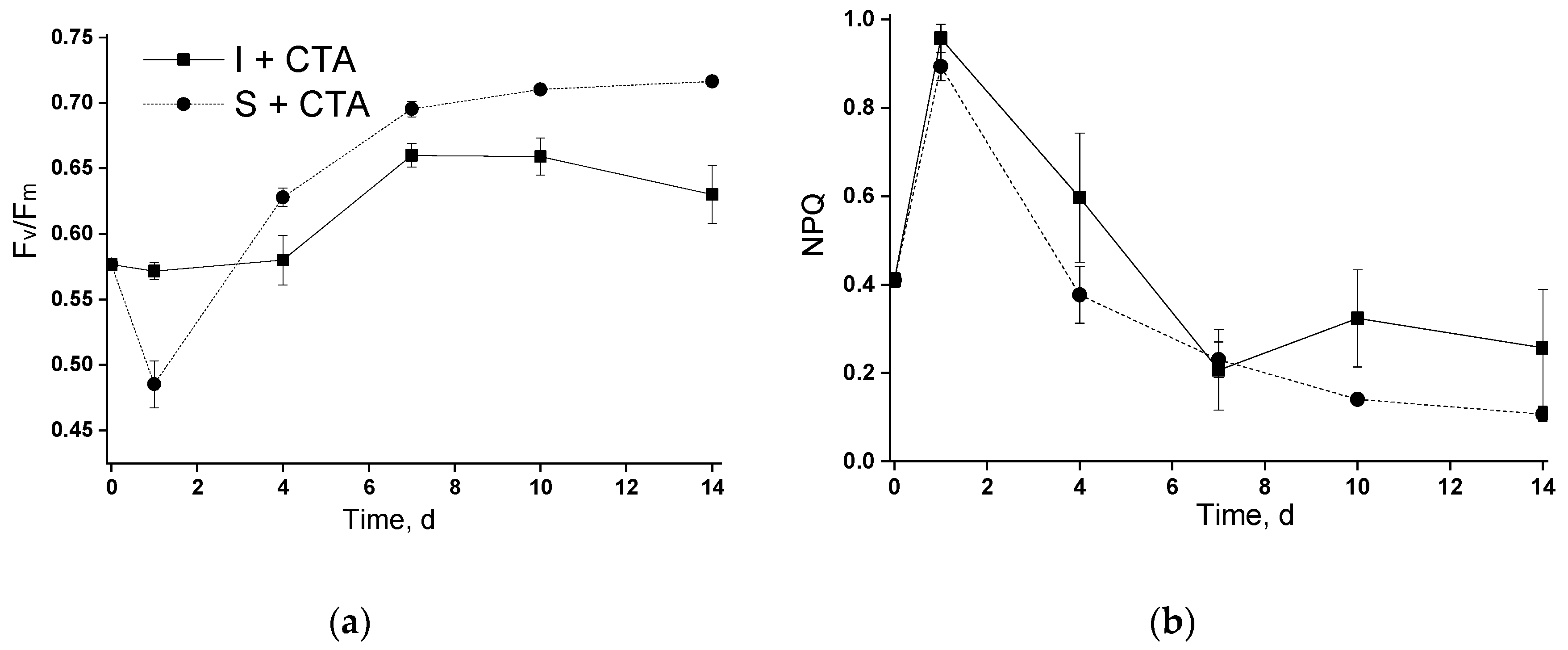
Figure 5.
Metagenomic analysis of the suspended (S) and immobilized (I) Lobosphaera sp. IPPAS C-2047 culture with (S+CTA or I+CTA, respectively) or without (S or I, respectively) addition of 20 mg/L ceftriaxone, in comparison to the preculture and blank chitosan with CTA, at the end of the 1 and 2 week of incubation. (a) Genus level taxonomic structure; (b) α-diversity index of Shannon (blue) and Simpson (orange); (с) β-diversity index of Morishita.
Figure 5.
Metagenomic analysis of the suspended (S) and immobilized (I) Lobosphaera sp. IPPAS C-2047 culture with (S+CTA or I+CTA, respectively) or without (S or I, respectively) addition of 20 mg/L ceftriaxone, in comparison to the preculture and blank chitosan with CTA, at the end of the 1 and 2 week of incubation. (a) Genus level taxonomic structure; (b) α-diversity index of Shannon (blue) and Simpson (orange); (с) β-diversity index of Morishita.
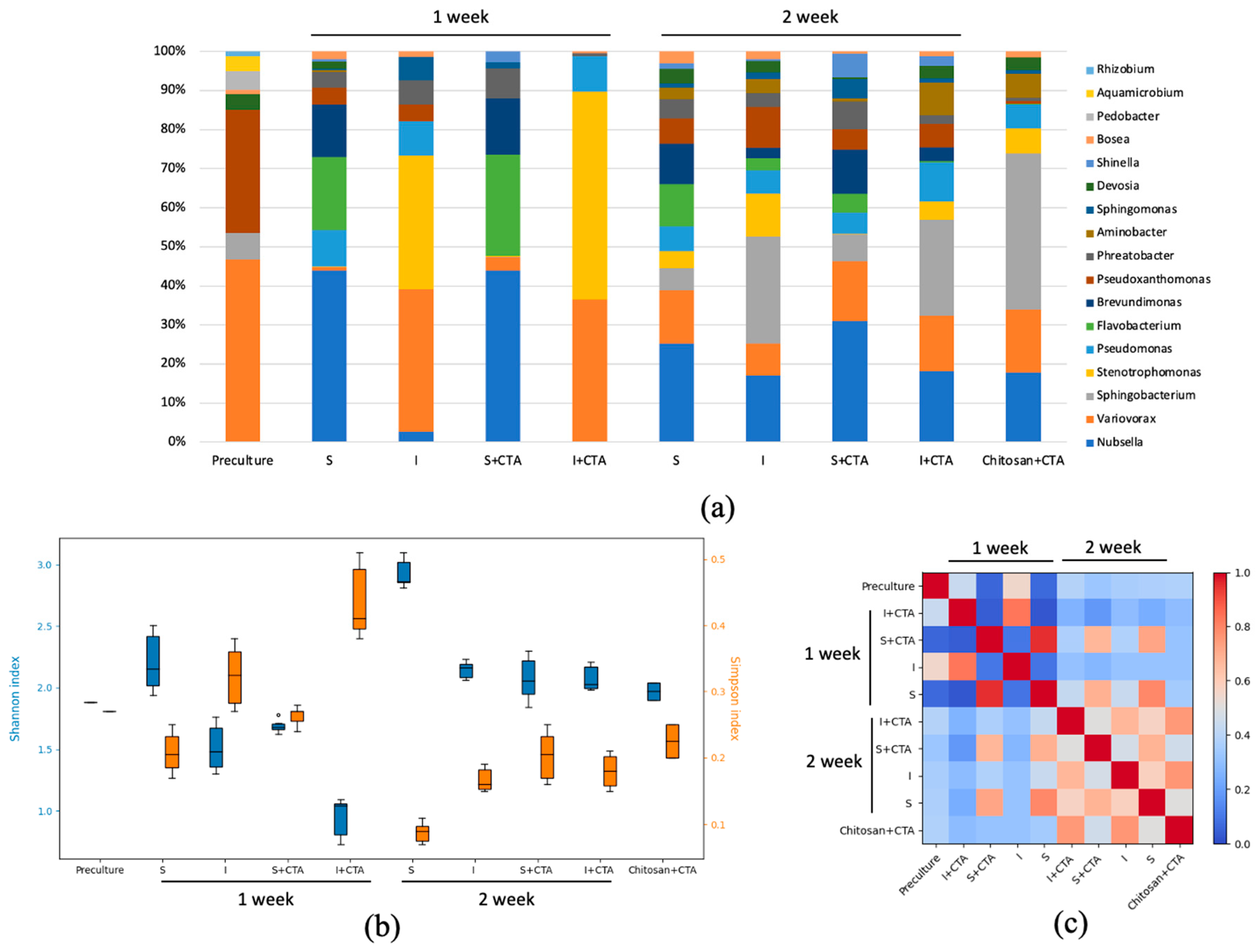
Figure 6.
Changes in the residual concentration of ceftriaxone in the cell-free BG-11M medium (CTA; open circles), cell-free BG-11M medium with chitosan (chitosan; open squares), suspended (S+CTA) and immobilized (I+CTA) Lobosphaera sp. IPPAS C-2047 cultures during (a) pilot experiment in the flasks and (b) in the column photobioreactor.
Figure 6.
Changes in the residual concentration of ceftriaxone in the cell-free BG-11M medium (CTA; open circles), cell-free BG-11M medium with chitosan (chitosan; open squares), suspended (S+CTA) and immobilized (I+CTA) Lobosphaera sp. IPPAS C-2047 cultures during (a) pilot experiment in the flasks and (b) in the column photobioreactor.
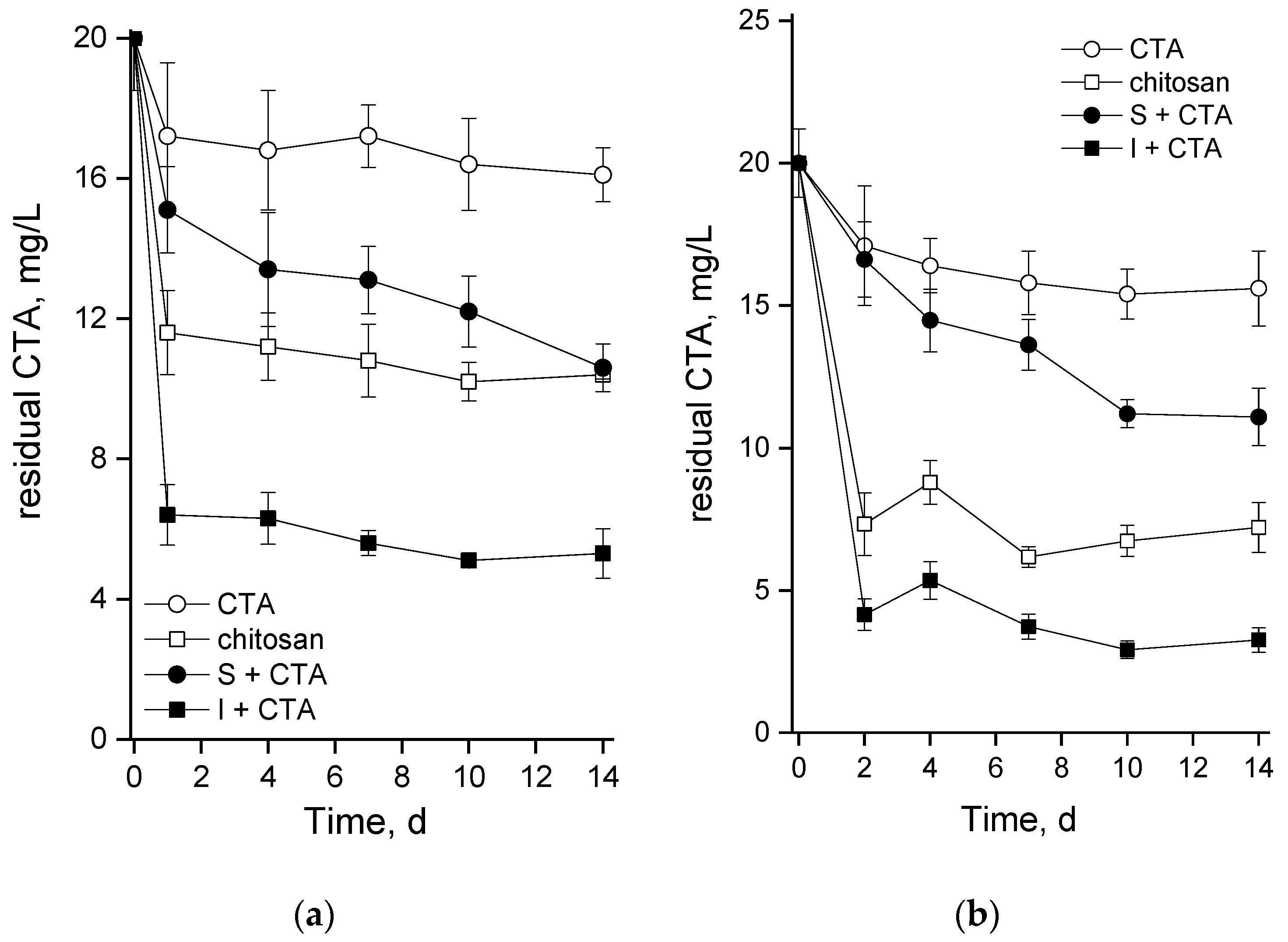
Figure 7.
Induction of fatty acid accumulation in the cells of the suspended (S) and immobilized (I) Lobosphaera sp. IPPAS C-2047 cultures during their incubation in complete BG-11M medium containing ceftriaxone (+N) and nitrogen-free, BG-11M0 medium lacking the antibiotic (–N). The changes in (a) chlorophyll, (b) volumetric total fatty acid content, (c) total fatty acid per unit chlorophyll, and (c) arachidonic acid percentage of total fatty acids are shown.
Figure 7.
Induction of fatty acid accumulation in the cells of the suspended (S) and immobilized (I) Lobosphaera sp. IPPAS C-2047 cultures during their incubation in complete BG-11M medium containing ceftriaxone (+N) and nitrogen-free, BG-11M0 medium lacking the antibiotic (–N). The changes in (a) chlorophyll, (b) volumetric total fatty acid content, (c) total fatty acid per unit chlorophyll, and (c) arachidonic acid percentage of total fatty acids are shown.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



