
Submitted:
30 May 2023
Posted:
31 May 2023
You are already at the latest version
Abstract
A simple and rapid method for the extraction of D-series resolvins (RvD1, RvD2, RvD3, RvD4, RvD5) released into Leibovitz's L-15 complete medium by head kidney cells from Atlantic salmon and further liquid chromatography triple quadrupole mass spectrometry determination is proposed. A three-level factorial design was proposed to select the optimal concentrations of internal standards that were used in the evaluation of the performance parameters such as linear range (0.1-50 ng/mL), limits of detection and quantification (0.05 and 0.1 ng/mL, respectively) and recovery values (98.99±0.00). The optimized method was used to determine the stimulated production of resolvins by head kidney cells exposed to docosahexaenoic acid, and the results indicated that it is possible that the production was controlled by circadian responses.
Keywords:
Resolvins
; Liquid chromatography quadrupole mass spectrometry
; Atlantic salmon
; Head kidney cells
; Liquid-liquid extraction
1. Introduction
Resolvins from the D-series are bioactive oxygenated metabolites of docosahexaenoic acid (22:6n-3; DHA) that were discovered in mice exudate cells treated with aspirin and DHA and were termed resolvins due to their role in dampening and promoting resolution of inflammation processes [1]. Resolvins from the D-series (RvD) are biosynthesized by the action of 15-LOX on DHA to produce 17S-hydroperoxydocosahexaenoic acid that is converted to various types of resolvin D (RvD): RvD1, RvD2, RvD3, RvD4, RvD5 and RvD6 by the action of 5-LOX [1,2]. RvD1 has therapeutic effects such as slowing the progression of osteoarthritis in joints, preventing neuronal dysfunction in Parkinson's disease, and increasing efferocytosis in the elderly [3]. RvD2 promotes subcellular localized healing, regenerative, and protective effects in burn wounds, such as keratinocyte restoration, muscle regeneration, tissue necrosis restriction, tumor growth inhibition and clearing cellular debris in mice [3,4]. RvD3 has powerful anti-inflammatory effects on leukocytes, decreases the levels of pro-inflammatory cytokines (MCP-1, IL-6, and keratinocyte chemoattractant protein), and eicosanoids (LTB4, PGD2 and TxB2), inhibits neutrophil transmigration and enhances macrophage absorption of microbial particles [5]. RvD4 protects organs in cases of ischemia kidney injury [3,6]. RvD5 improves phagocytosis by increasing neutrophil and macrophage movement, modulates TNF-α and NF-κB in blood, synovial fluid and exudates discharged after a hemorrhage and in the late stages of coagulation prolongs the agglutination process and reduces the likelihood of bleeding [7]. RvD6 increases nerve regeneration, stimulates hepatocyte growth factor genes specifically as upstream regulators and a gene network involved in axon growth and suppression of neuropathic pain, indicating a novel function of this lipid mediator to maintain cornea integrity and homeostasis after injury [2,8].
In general, resolvins produced from DHA have demonstrated promising therapeutic advantages in terms of cell damage reduction, oxidative stress inhibition, and tumor growth suppression [9]. In mammals, there is growing evidence that resolvins may assist in the resolution of acute inflammation and potently suppress inflammatory and neuropathic pain. Although it is unknown whether this is the case in fish, a study conducted by Ruyter and colleagues showed that greater dietary DHA levels lead to higher concentrations of resolvins in plasma, which may have health benefits in fish [10]. Neuronal deficiencies and developmental issues in larvae have been reported in fish fed with a diet scarce in DHA [11], hence the possible link between resolvins in neuronal function of fish.
Different approaches have been reported for the determination of resolvins in different kind of samples. Enzyme immunoassay (EIA) kits has been used for determining the production of RvD1 in human and fish plasma [10,12] with high sensitivity. However, it is also well known that EIA is prone to cross-reactivity, which in turn cause an overestimation of the levels of specific resolvins. In addition, EIA are limited to just one type of resolvin per commercial kit (e.g., either RvD1 or RvD2) which makes the technique remarkably expensive when different resolvins are considered.
High performance liquid chromatography with UV diode array detection (HPLC-DAD) and gas chromatography coupled to mass spectrometry (GC-MS) have been used as alternative techniques to validate the results of liquid chromatography mass spectrometry in tandem mode (LC-MS/MS). For instance, enzymatically generated RvD1 was determined by LC-MS/MS analysis and further evidence of its positive identification was obtained using HPLC-DAD to confirm the presence of a conjugated tetraene structure within RvD1 that is responsible for its characteristic triplet chromophore at a λmax = 301 nm. A subsequent GC-MS analysis was performed after derivatizing RvD1 with diazomethane to its corresponding trimethylsilyl derivative [9,13]. In a similar way, actively phagocytosing poly-morphonuclear neutrophils were converted to RvD2 and determined by LC-MS/MS analysis, followed by a subsequent GC-MS analysis of derivatized RvD2 to validate the LC-MS/MS determination [9]. The main drawback associated with HPLC-DAD is the potential coelution of isomeric resolvins with similar spectrum which may hinder their discrimination, while the most evident disadvantage GC-MS is that is restricted to thermally stable volatile compounds, generally prepared by time-consuming derivatization process [14,15,16].
Resolvins are commonly found in biological fluids and organs at extremely low concentrations, including peripheral blood, cerebral fluid, placenta, synovial fluids, urine, sputum, spleen, lymph nodes, cell cultures and others [17]. As a result, successful extraction is essential to wash and clean up the sample followed by drying up and reconstitution in a small volume to improve the concentration of analyte. An overview of the literature indicated that solid phase extraction (SPE) is arguably the most popular extraction method for the analysis of resolvins in different kind of samples and LC-MS/MS the preferred quantitative technique for its outstanding sensitivity. A recent article has proposed a cumbersome methodology that combines liquid-liquid extraction (LLE) with chloroform and acetate-water buffered at pH 4 followed by µ-SPE with methanol-water buffered at pH 4 and final LC-MS/MS analysis for the quantification of resolvins in human keratinocyte cell lysates to obtain recoveries around 42 and 64% [18]. Unfortunately, SPE alone (not to mention combined with LLE) is a time consuming and complicated method that requires multiple steps and different solvents prior to LC-MS/MS analysis. Our current SPE protocol for the analysis of resolvins (RvD1, RvD2) in cell culture requires a total of seven different solvents (six solvents for SPE and one solvent for final reconstitution) [14]. Poor recovery, reproducibility, and insufficient cleaning of sample extracts are some of the drawbacks commonly associated to SPE [19]. Different SPE adsorbent materials and LLE have been compared to propose a suitable method for the extraction of RvD1 from human endothelial cells and further determination by LC-MS, and the best results were obtained using LLE with a solution of methanol containing the internal standard [20]. Although it seems a promising approach, this LLE protocol was validated incorrectly by using spiked plasma samples instead of human endothelial cells. Furthermore, methanol is generally used for protein removal, therefore LLE with methanol for biological samples requires multiple and time-consuming centrifugation steps to ensure complete precipitation. For instance, centrifugation times of 45 min for metabolome [21] and 70 min for RvD1 analysis [20] followed by drying methanol with a stream of nitrogen that is a lengthy operation prior to final reconstitution and LC-MS/MS analysis.
Overall, sample preparation for the analysis of multiple compounds (e.g., resolvins) is the major bottleneck in analytical laboratories. The aim of this study is to propose a simple and rapid LLE procedure to quantify the temporal production of released resolvins (RvD1, RvD2, RvD3, RvD4, RvD5) into Leibovitz's L-15 complete medium by head kidney cells from Atlantic salmon exposed to DHA and further liquid chromatography triple quadrupole mass spectrometry (LC-MS/MS). To our knowledge, this is the first validated LLE procedure for quantifying biosynthesized D-series resolvins by cell cultures.
2. Results and discussion
2.1. Optimal Concentration of Internal Standards
A 32-factorial design, where the base 3 represents the number of concentration levels (low, medium, high) and the exponent 2 the number of factors (analyte and internal standard), was used to investigate whether the response factor (RF) remains constant at different internal standard (IS) concentrations over a fixed analytical range. The nine combinations of analytes and internal standards (32) suggested by the factorial design (Table 1) were prepared by dissolving the analytical resolvins and internal standards in the L-15 medium. For instance, experiment 4 represents a L-15 media solution containing all the analytes (RvD1, RvD2, RvD3, RvD4, RvD5) at a concentration level of 30 ng/mL and submitted to the extraction protocol by using a mixture of of deuterated internal standards (RvD1-d5 RvD2-d5 RvD3-d5) at 15 ng/mL each. Four blanks consisting of L-15 medium containing 0, 15, 30 and 45 ng/mL of internal standards were also prepared. The experiments in Table 1 were were prepared in triplicate and run in random order.
The RF for every resolvin at the different concentrations of IS were calculated at every experimental point in Table 1 by the expression RF=[IS]/[A]×(yA/yIS), where [A] and [IS] represent the analyte and IS concentrations and yA and yIS their corresponding signals, respectively. The results are shown in Table 2.
A multiple range test, applied to determine significant differences (p<0.05) between the calculated RF values, revealed that apart from RvD5, the RF remains constant for the concentrations 15 and 30 ng/mL of IS with coefficients of variations around 2%. Hence, 15 ng/mL of RvD1-d5 (for RvD1, RvD4 and RvD5), RvD2-d5 (for RvD2) and RvD3-d5 (for RvD3) that yields averages RF of 2.90±0.05, 14.47± 0.33, 34.33±0.61, 0.84± 0.01 and 1.01±0.04 for RvD1, RvD2, RvD3, RvD4, RvD5, respectively, was selected as the optimal concentration level to be used in connection with the proposed extraction protocol and further quantitative analysis by LC-MS/MS. Although, the RF values for RvD5 (1.10± 0.04, 1.01±0.02, 1.19±0.02 at 15, 30 and 45 ng/mL of IS, respectively) exhibited statistically significant differences, the RF for 15 ng/mL lied between 30 and 45 ng/mL with a coefficient of variation (3.2%) similar to those reported elsewhere (4-13% at 1 ng/mL IS) for cell experiments by LCMS 18.
2.2. Analytical Validation
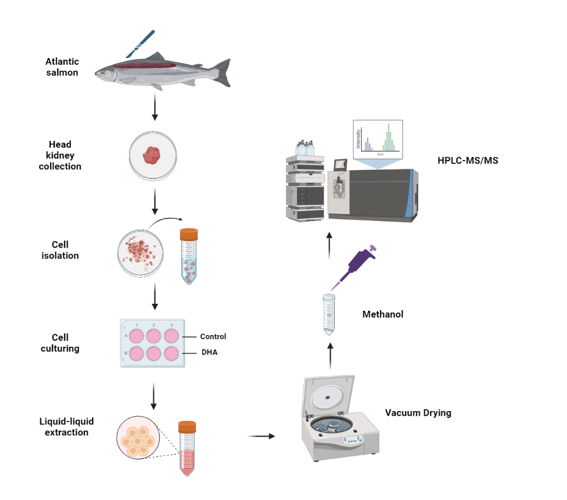
The deuterated internal standards, RvD1-d5, RvD2-d5 and RvD3-d5 were dissolved in acetonitrile (15 ng/mL each) and used to extract the resolvins from seven L-15 media preparations containing a mixture of 5 analytical resolvins (RvD1 to RvD5) at seven different concentrations (0, 1, 5, 15, 30, 45, 50 ng/mL). The characteristic mass fragments of the five resolvins and three internal standards were extracted from the precursor ions in both, unspiked (0 ng/mL) and spiked (1-50 ng/mL) L-15 media. The five resolvins were separated chromatographically according to their retention times, including RvD1 and RvD2 that have the same precursor (m/z 375) and product (m/z 141) ions. The chromatographic elution order was RvD3 (2.55 min), RvD2 (2.59 min) RvD1 (2.77 min), RvD4 (3.12 min), RvD5 (3.55 min). The extracted ion chromatograms (EIC) revealed that the proposed extraction protocol allow detecting unequivocally the resolvins with negligible background interferences (Figure 1), therefore the analyses were regarded as highly selective towards the five resolvins.
The chromatographic peak area ratios RvD1/RvD1-d5, RvD2/RvD2-d5, RvD3/RvD3-d5, RvD4/RvD1-d5 and RvD5/RvD1-d5 were calculated and plotted against the analytical concentrations to compute the function and obtain the various calibration parameters displayed in Table 3.
The degree of linearity of the calibrations was provided by both, the regression coefficients (R2) and the Fisher test defined as the quotient between the lack-of-fit to pure error variances (Fexp in Table 3). In general, the five resolvins were linear over the studied range of concentrations, as reflected in Table 3, where the R2 values (between 0.994-0.999) indicate that a high proportion of the variance of the calculated yA/yIS signals are explained by the analytical concentrations [A] in the proposed regression models. This conclusion is also supported by the Fexperimental values for the five calibration models that were lower than the critical value of 2.958 for 5 and 14 degrees of freedom at the 95% confidence level (Table 3). The LOD (0.028-0.059 ng/mL) and LOQ (0.074-0.180 ng/mL) were within the range of previously reported values for cell cultures [22] and considered appropriate for all resolvin species. In the present work, the LOQ for resolvins in completed L-15 media by using an Agilent 6495 triple quadrupole are similar (and in some cases better) to those reported in pure standards by using Sciex QTRAP 6500 [23,24]. For instance, the referred Quadrupole/QTRAP (present/[24]) values for RvD1, RvD2, RvD3, RvD4 and RvD5 are 0.127/0.05, 0.5/0.086, 0.097/0.05, 0.074/0.1 and 0.180/0.1, respectively. Based on the matrix complexity, namely the present L-15 medium versus pure standard [24] the LOQ values of the present research can be regarded as remarkable. In addition, the proposed extraction protocol in conjunction with triple quadrupole spectrometer is an outstanding strategy considering the widespread consensus that QTRAP delivers better data than quadrupole systems [25]. The recovery of the method, expressed as the ratio between found and nominal concentrations (100×[A]found/[A]nominal) was higher than 95% in all the cases as described in Table 3.
2.3. Analysis of released resolvins in L-15 media by head kidney cells
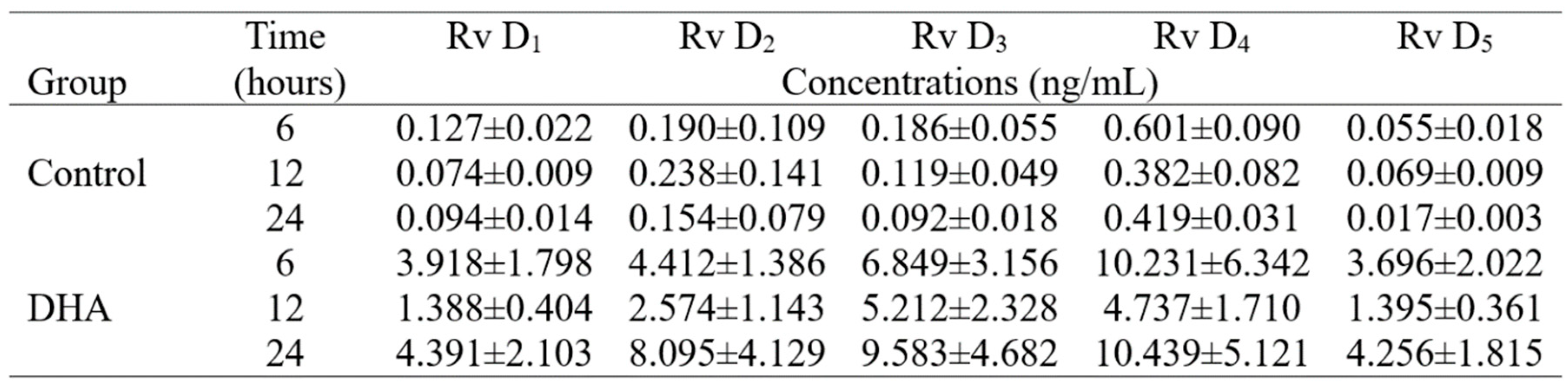
The proposed LLE protocol and further LC-MS/MS quantification was implemented to study the induced production of resolvins by salmon head kidney cells with and without exposure to exogenous DHA. The production of resolvins was expressed in ng/mL and measured at 6, 12 and 24 hours (Table 4). The levels of resolvins in decreasing order of concentrations were RvD4>RvD2>RvD3>RvD1>RvD5 and RvD4>RvD3>RvD2>RvD1>RvD5 in control and DHA, respectively. These levels agree with previously reported results that were obtained by using SPE and LC-MS/MS to estimate the production of RvD1 and RvD2 by salmon liver cells, and where it was suggested that the production of RvD4 was preferred over RvD1 and RvD2 after exposing the cells to different polyunsaturated fatty acids, including DHA [14].
A principal component analysis (PCA) indicated that 76.00% of the total data variability was explained by the concentrations of resolvins in control and DHA group, and 16.24% was explained by the different times (Figure 2).
The control and DHA groups were clearly discriminated along the PC1 axis and characterized by negative and positive scores along this axis, respectively. The degree of overlapping within the control and DHA group was higher in the former than in the latter group. For instance, the scores for the control at 6 or 12 or 24 hours appeared as three separated clusters along PC2 (red squares), while for the DHA the time clusters were separated along PC2 (green squares), but they were widely spread along PC1, indicating a higher dispersion of the DHA data. The PCA also showed that the highest concentrations of resolvins were associated with the DHA group suggesting that exogenous DHA promoted the production of resolvins. The vectors time and concentrations of resolvins were orthogonal, therefore changes in concentrations of RvD1, RvD2, RvD3, RvD4 and RvD5 were independent of the time. This lack of correlation between time and concentration was confirmed by studying the within and between variances at the three selected times for every resolvin. The estimated p-values were not significant (p>0.05) for every type of resolvin either in the control or in the DHA group.
The concentration/time relationship was computed from Table 4 and the results revealed a continuous decrease in production over the time for the five analyzed resolvins in the control group (Figure 3). In contrast, the DHA group showed higher production at subjective dawn (6 h) and subjective midnight (24 h) than subjective midday (12 h) for all resolvins (Figure 2), suggesting the presence of a circadian clock that may impose a 24-h rhythmicity to the head kidney cells to process the production of resolvins from the added 50 μM of DHA.
The direct influence of circadian rhythms on resolvin production remains unexplored. However, considering the present findings, it is plausible the existence of an underlying resolvin/15-LOX immune regulating clock that controls the production of resolvins from endogenous DHA. This remark is supported by the results from different studies that have observed higher levels of COX-1 and inflammatory prostaglandins (PGE2 and PGF2α) at midnight than at midday, and concluded that some immunological functions are controlled by circadian responses from a prostaglandin/COX-1 system [26]. Similarly, anti-inflammatory prostaglandin 15d-PGJ2, has been identified as an entrainment factor aligned with circadian oscillations [27].
3. Materials and Methods
3.1. Reagents
Resolvin D1 (RvD1, 95%), resolvin D2 (RvD2, 95%), resolvin D3 (RvD3, 95%), resolvin D4 (RvD4, 95%), resolvin D5 (RvD5, 95%), deuterated resolvin D1 (RvD1-d5, 95%), deuterated resolvin D2 (RvD2-d5, 95%) and deuterated resolvin D3 (RvD3-d5, 95%). Acetonitrile (99.8 %) and formic acid (98 %) were purchased from SigmaAldrich (St. Louis, MO, USA). 2-propanol (HPLC grade, 99.9 %) from Merck (Darmstadt, Germany). Chloroform (HPLC grade, 99.8%) was obtained from Merck (Darmstadt, Germany). A Millipore Milli-Q system was used to produce ultra-pure water 18 MΩ (Millipore, Milford, USA). Cis-4,7,10,13,16,19-docosahexaenoic acid (DHA, ≥98%) were purchased from Sigma–Aldrich (Oslo, Norway). Leibovitz`s L-15 medium from Sigma-Aldrich (St. Louis, MO, USA). Fetal bovine serum (FBS, cat# 14-801F) was from BioWhittaker (Petit Rechain, Belgium). The glutaMaxTM 100× (Gibco-BRL, cat# 35056) was from Gibco-BRL (CergyPontoise, France).
3.2. Head Kidney Cells
For each fish, the head kidneys were directly sampled and added PBS at 5⁰C and then cut with a scissor and squeezed through a 40 µM Falcon cell strainer. The cells were transferred to tubes and centrifuged in a Hettich Zentrifugen, 320 R, at 400 × g for 5 min at 4⁰C. Cell pellets were resuspended in PBS and layered carefully on top of equal amounts of diluted Percoll in density 1.08 g/mL. The tubes were centrifuged at 800 × g for 30 min at 4⁰C. The cell layer in the interface containing the head kidney leukocytes was collected and the cells were pelleted by centrifugation, 400 × g for 5 min at 4◦C. An additional washing step in PBS was performed. The cells were counted using a Bürker chamber and 0.4% trypan blue solution and the viability was above 85%.
3.3. Cell Cultures
A complete L-15 (cL-15) medium was supplemented with 10% foetal bovine serum (FBS), 2% pen/strep, 2% glutamaxTM100× and used to prepare cL-15 solutions containing DHA that was diluted to a concentration 50 μM and a control solution containing ethanol (the solvent used to dissolve the DHA). Approximately 1×107 salmon head kidney cells were cultured into each well (control and DHA). The cell culture plates were incubated in a normal atmosphere incubator (Sanyo Electric Company Ltd. Osaka, Japan) at 9⁰C for 6, 12 and 24 h under dark conditions. The two suspensions of cells (control and DHA) were prepared in pentaplicate. The head kidney cells were centrifuged at 50 × g for 5 min at 4◦C, the medium collected and stored at -80⁰C until extraction followed by LC-MS/MS analysis.
3.4. Optimal Concentrations of the Internal Standards
A 3k factorial design, where 3 represents the number of concentration levels (low, medium, high) and k the number of factors (analyte and internal standard), was used to study variations in the response factor (RF) when the concentrations of both, the internal standards (IS) and the analytical resolvins are varied between 15-45 ng/mL. The optimal IS concentrations should yield a stable RF over the explored analytical range.
3.5. Extraction protocol
Two successive aliquots of acetonitrile (500 μL) containing the mixture of internal standards at the concentration levels indicated in Table 1 and chloroform (500 μL) were added successively into an Eppendorf tube containing 200 μL of the mixture of resolvins at the concentration levels of every experiment in Table 1. The Eppendorf tube was vortex-mixed for 30 sec (Bandelin RK 100 ultra mixer, Berlin, Germany), centrifuged at 1620×g for 3 min (Eppendorf AG centrifuge, Hamburg, Germany), the top phase was removed, and the extraction procedure repeated in the remaining phase using acetonitrile without internal standards and chloroform. After removing the chloroform phase, the remaining solution was vacuum-dried at room temperature (Labconco vacuum drier system, Kansas, MO), diluted to 50 μL with methanol, transferred to an autosampler vial and submitted to LC-MS/MS analysis.
3.6. Analytical Performance
The parameters used to assess the analytical performance of the extraction method in conjunction with LC-MS/MS were selectivity, limit of detection (LOD), limit of quantification (LOQ), calibration range and recovery. The selectivity of the method was evaluated by comparing the chromatograms obtained after injection of L-15 medium samples with and without the analytes. The calibration curves for the resolvins in L-15 medium were prepared between 0-50 ng/mL and extracted as described above by using the optimal concentration of internal standards suggested by the 3k factorial design. The linearity was judged by computing both, variance ratio lack-of-fit to pure error and the coefficient of regression, as suggested by the Analytical Method Committee [28] and the International Council for Harmonisation guidance for validation of analytical procedures [29]. The ratio standard deviation (σ) to slope (φ) of the regression curves for every resolvin was used to determine the LOD (3.3×σ/φ) and LOQ (10×σ/φ) as described elsewhere [30]. The percentage of recovery was assessed by comparing the degree of agreement between experimental and nominal concentrations as acknowledged by the ICH [29,30].
3.7. Liquid Chromatography Mass Spectrometry
An Agilent ultra-high performance liquid chromatography (UHPLC) coupled to a 6495 QQQ triple quadrupole (Agilent Technologies, Waldbronn, Germany) with an electrospray ionization (ESI) interface and iFunnel ionization was used to quantify the eicosanoids. The UHPLC system was equipped with a Zorbax RRHD Eclipse Plus C18, 95Å, 2.1 × 50 mm, 1.8 µm chromatographic column. The mobile phase delivered at 0.4 mL/min in gradient mode consisted of ultra-pure water with 0.1 % formic acid (solution A) and an equal volume mixture of acetonitrile and methanol with 0.1 % formic acid (solution B). The solvent gradient was as follows: solution A was reduced from 60 to 5 % from 0.00 to 4.00 min, kept at 5 % between 4.00 and 5.50 min, increased to 60 % between 5.50 and 5.51 min and kept at 60 % between 5.51 and 10.00-min. Mass spectrometric detection was performed by multiple reactions monitoring (MRM) in negative mode. The monitored transitions in percentage of ion counts (%) were: m/z 375→ 141 for RvD1 and RvD2; m/z 375→147 for RvD3; m/z 375→101 for RvD4; m/z 359→199 for RvD5; m/z 380→ 141 for RvD1-d5; m/z 380 → 141 for RvD2-d5; m/z 380→147 for RvD3-d5. The ESI parameters were gas temperature (120◦C), gas flow rate (19 L/min), nebulizer pressure (20 psi), sheath gas temperature (300◦C), sheath gas flow (10 L/min), capillary voltage (3500V) and nozzle voltage (2000V). The integration of the chromatograms was performed using the MassHunter Qualitative Navigator software (version 10.0). The levels of resolvins were estimated by means of the internal standards and expressed in ng/ mL units.
3.8. Statistics
Statgraphics Centurion XV Version 15.2.11 (StatPoint Technologies, Inc. Warrenton, VA, USA) was used for the statistical analyses.
4. Conclusions
This is the first validated liquid-liquid extraction method for resolvins released by head kidney cells from Atlantic salmon in cL-15 media and further quantification by LC-MS/MS using the internal standard calibration method. The small amount of sample (200 μL), the low solvent consumption, the fast extraction times, the high sample throughput (40-50 samples/day) and LOD and LOQ similar to those reported by using SPE and pure standards are important features that make the present approach highly attractive to evaluate the production of resolvins by cell cultures challenged to polyunsaturated fatty acids, such as DHA.
In light of the present findings, it is plausible that production of resolvins by head kidney cells from endogenous DHA is controlled in part by circadian responses.
Acknowledgments
The European Commission in the context of the Erasmus Mundus Program is gratefully acknowledged for financial support of S.I.
References
- Serhan, C. N.; Dalli, J.; Colas, R. A.; Winkler, J. W.; Chiang, N. Protectins and Maresins: New pro-Resolving Families of Mediators in Acute Inflammation and Resolution Bioactive Metabolome. Biochim. Biophys. Acta - Mol. Cell Biol. Lipids 2015, 1851(4), 397–413. [Google Scholar] [CrossRef] [PubMed]
- Pham, T. L.; Kakazu, A. H.; He, J.; Nshimiyimana, R.; Petasis, N. A.; Jun, B.; Bazan, N. G.; Bazan, H. E. P. Elucidating the Structure and Functions of Resolvin D6 Isomers on Nerve Regeneration with a Distinctive Trigeminal Transcriptome. FASEB J. 2021, 35(8), 1–14. [Google Scholar] [CrossRef] [PubMed]
- Chiang, N.; Serha, C. N. Specialized Pro-Resolving Mediator Network: An Update on Production and Actions. Essays Biochem. 2020, 64(3), 443–462. [Google Scholar] [CrossRef]
- Dyall, S. C.; Balas, L.; Bazan, N. G.; Brenna, J. T.; Chiang, N.; da Costa Souza, F.; Dalli, J.; Durand, T.; Galano, J. M.; Lein, P. J.; Serhan, C. N.; Taha, A. Y. Polyunsaturated Fatty Acids and Fatty Acid-Derived Lipid Mediators: Recent Advances in the Understanding of Their Biosynthesis, Structures, and Functions. Prog. Lipid Res. 2022, 86, 101165. [Google Scholar] [CrossRef] [PubMed]
- Dalli, J.; Winkler, J. W.; Colas, R. A.; Arnardottir, H.; Cheng, C. Y. C.; Chiang, N.; Petasis, N. A.; Serhan, C. N. Resolvin D3 and Aspirin-Triggered Resolvin D3 Are Potent Immunoresolvents. Chem. Biol. 2013, 20(2), 188. [Google Scholar] [CrossRef]
- Winkler, J. W.; Orr, S. K.; Dalli, J.; Cheng, C.-Y. C.; Sanger, J. M.; Chiang, N.; Petasis, N. A.; Serhan, C. N. Resolvin D4 Stereoassignment and Its Novel Actions in Host Protection and Bacterial Clearance OPEN. 2015. [CrossRef]
- Kalyanaraman, C.; Tourdot, B. E.; Conrad, W. S.; Akinkugbe, O.; Freedman, J. C.; Holinstat, M.; Jacobson, M. P.; Holman, T. R.; Perry, S. C.; Kalyanaraman, C.; Tourdot, B. E.; Conrad, W. S.; Akinkugbe, O.; Cody Freedman, J.; Holinstat, M.; Jacobson, M. P.; Holman, T. R. 15-Lipoxygenase-1 Biosynthesis of 7S,14S-DiHDHA Implicates 15-Lipoxygenase-2 in Biosynthesis of Resolvin D5[S]. J. Lipid Res. 2020, 61(S). [Google Scholar] [CrossRef]
- Pham, T. L.; Kakazu, A. H.; He, J.; Jun, B.; Bazan, N. G.; Bazan, H. E. P. Novel RvD6 Stereoisomer Induces Corneal Nerve Regeneration and Wound Healing Post-Injury by Modulating Trigeminal Transcriptomic Signature. [CrossRef]
- N. Serhan, C.; Krishnamoorthy, S.; Recchiuti, A.; Chiang, N. Novel Anti-Inflammatory-Pro-Resolving Mediators and Their Receptors. Curr. Top. Med. Chem. 2012, 11(6), 629–647. [CrossRef]
- Ruyter, B.; Bou, M.; Berge, G. M.; Mørkøre, T.; Sissener, N. H.; Sanden, M.; Lutfi, E.; Romarheim, O. H.; Krasnov, A.; Østbye, T. K. K. A Dose-Response Study with Omega-3 Rich Canola Oil as a Novel Source of Docosahexaenoic Acid (DHA) in Feed for Atlantic Salmon (Salmo Salar) in Seawater; Effects on Performance, Tissue Fatty Acid Composition, and Fillet Quality. Aquaculture 2022, 561, 738733. [Google Scholar] [CrossRef]
- Shields, R. J. Larviculture of Marine Finfish in Europe. Aquaculture 2001, 200(1-2), 55–88. [Google Scholar] [CrossRef]
- Fedirko, V.; McKeown-Eyssen, G.; Serhan, C. N.; Barry, E. L.; Sandler, R. S.; Figueiredo, J. C.; Ahnen, D. J.; Bresalier, R. S.; Robertson, D. J.; Anderson, C. W.; Baron, J. A. Plasma Lipoxin A4 and Resolvin D1 Are Not Associated with Reduced Adenoma Risk in a Randomized Trial of Aspirin to Prevent Colon Adenomas. Mol. Carcinog. 2017, 56(8), 1977–1983. [Google Scholar] [CrossRef]
- Serhan, C. N.; Hong, S.; Gronert, K.; Colgan, S. P.; Devchand, P. R.; Mirick, G.; Moussignac, R. L. Resolvins: A Family of Bioactive Products of Omega-3 Fatty Acid Transformation Circuits Initiated by Aspirin Treatment That Counter Proinflammation Signals. J. Exp. Med. 2002, 196(8), 1025. [Google Scholar] [CrossRef]
- Lucena, E.; Yang, Y.; Mendez, C.; Holen, E.; Araujo, P. Extraction of Pro-and Anti-Inflammatory Biomarkers from Fish Cells Exposed to Polyunsaturated Fatty Acids and Quantification by Liquid Chromatography Tandem Mass Spectrometry. Current Analysis on Chemistry, vol. 1, pp. 1–09, 2018. available at http://www.mesfordpublisher.com/ journals/cac/articles/article-1/.
- Bordet, J. C.; Guichardant, M.; Lagarde, M. Arachidonic Acid Strongly Stimulates Prostaglandin I3 (PGI3) Production from Eicosapentaenoic Acid in Human Endothelial Cells. Biochem. Biophys. Res. Commun. 1986, 135(2), 403–410. [Google Scholar] [CrossRef] [PubMed]
- Tsikas, D.; Schwedhelm, E.; Gutzki, F. M.; Frölich, J. C. Gas Chromatographic-Mass Spectrometric Discrimination between 8-Iso-Prostaglandin E2 and Prostaglandin E2 through Derivatization by O-(2,3,4,5,6-Pentafluorobenzyl)Hydroxyl Amine. Anal. Biochem. 1998, 261(2), 230–232. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wu, X.; Liu, S.; Shen, D.; Zhu, J.; Liu, K. Role of Resolvins in the Inflammatory Resolution of Neurological Diseases. Front. Pharmacol. 2020, 11(May), 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fanti, F.; Oliva, E.; Tortolani, D.; Di Meo, C.; Fava, M.; Leuti, A.; Rapino, C.; Sergi, M.; Maccarrone, M.; Compagnone, D. ΜSPE Followed by HPLC–MS/MS for the Determination of Series D and E Resolvins in Biological Matrices. J. Pharm. Biomed. Anal. 2021, 203, 114181. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Eiroa, A.; Canle, M.; Leroy-Cancellieri, V.; Cerdà, V. Solid-Phase Extraction of Organic Compounds: A Critical Review. Part Ii. TrAC Trends Anal. Chem. 2016, 80, 655–667. [Google Scholar] [CrossRef]
- Dufour, D.; Khalil, A.; Nuyens, V.; Rousseau, A.; Delporte, C.; Noyon, C.; Cortese, M.; Reyé, F.; Pireaux, V.; Nève, J.; Vanhamme, L.; Robaye, B.; Lelubre, C.; Desmet, J. M.; Raes, M.; Boudjeltia, K. Z.; Van Antwerpen, P. Native and Myeloperoxidase-Oxidized Low-Density Lipoproteins Act in Synergy to Induce Release of Resolvin-D1 from Endothelial Cells. Atherosclerosis 2018, 272, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cruickshank, C.; Armstrong, M.; Mahaffey, S.; Reisdorph, R.; Reisdorph, N. New Sample Preparation Approach for Mass Spectrometry-Based Profiling of Plasma Results in Improved Coverage of Metabolome. J. Chromatogr. A 2013, 1300, 217–226. [Google Scholar] [CrossRef]
- Masoodi, M.; Mir, A. A.; Petasis, N. A.; Serhan, C. N.; Nicolaou, A. Simultaneous Lipidomic Analysis of Three Families of Bioactive Lipid Mediators Leukotrienes, Resolvins, Protectins and Related Hydroxy-Fatty Acids by Liquid Chromatography/Electrospray Ionisation Tandem Mass Spectrometry. Rapid Commun. Mass Spectrom. 2008, 22(2), 75–83. [Google Scholar] [CrossRef]
- Kutzner, L.; Rund, K. M.; Ostermann, A. I.; Hartung, N. M.; Galano, J. M.; Balas, L.; Durand, T.; Balzer, M. S.; David, S.; Schebb, N. H. Development of an Optimized LC-MS Method for the Detection of Specialized Pro-Resolving Mediators in Biological Samples. Frontiers in Pharmacology. 2019. [CrossRef]
- Norris, P.; Kapil, S.; Gorti, K. Targeted Profiling of Lipid Mediators. SCIEX Present. 2017, 1–6. [Google Scholar]
- Agilent 6470B or Sciex 7500 Triple Quad - Chromatography Forum. https://www.chromforum.org/viewtopic.php?t=110834 (accessed 2023-05-19).
- De Zavalía, N.; Fernandez, D. C.; Sande, P. H.; Keller Sarmiento, M. I.; Golombek, D. A.; Rosenstein, R. E.; Silberman, D. M. Circadian Variations of Prostaglandin E2 and F2 α Release in the Golden Hamster Retina. J. Neurochem. 2010, 112(4), 972–979. [Google Scholar] [CrossRef] [PubMed]
- Nakahata, Y.; Akashi, M.; Trcka, D.; Yasuda, A.; Takumi, T. The in Vitro Real-Time Oscillation Monitoring System Identifies Potential Entrainment Factors for Circadian Clocks. BMC Mol. Biol. 2006, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Analytical Method Committee. Is My Calibration Linear? Analyst 1994, 119(11), 2363–2366. [Google Scholar] [CrossRef]
- Stefanini-Oresic, L. Validation of Analytical Procedures: ICH Guidelines Q2(R2). Farm. Glas. 2022, 2(0), 1–34. [Google Scholar]
- Araujo, P. Key Aspects of Analytical Method Validation and Linearity Evaluation. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2009, 877(23), 2224–2234. [Google Scholar] [CrossRef]
Figure 1.
Extracted ion chromatograms to indicate the selectivity of the analysis towards the resolvins after implementing the proposed LLE protocol.
Figure 1.
Extracted ion chromatograms to indicate the selectivity of the analysis towards the resolvins after implementing the proposed LLE protocol.
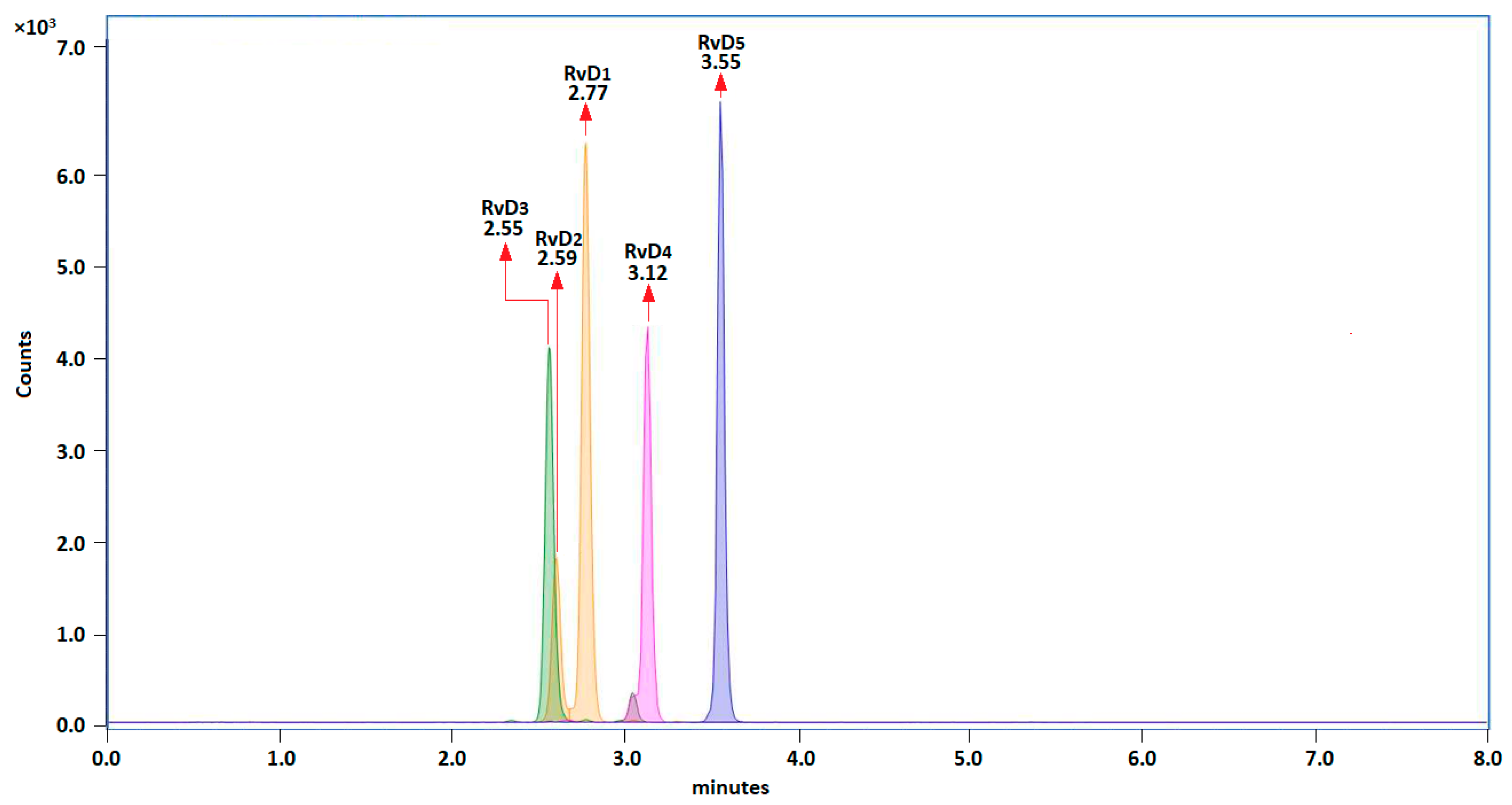
Figure 2.
Principal component analysis of the released resolvins in L-15 media by head kidney cells with and without exposure to DHA, after implementing the proposed LLE protocol.
Figure 2.
Principal component analysis of the released resolvins in L-15 media by head kidney cells with and without exposure to DHA, after implementing the proposed LLE protocol.
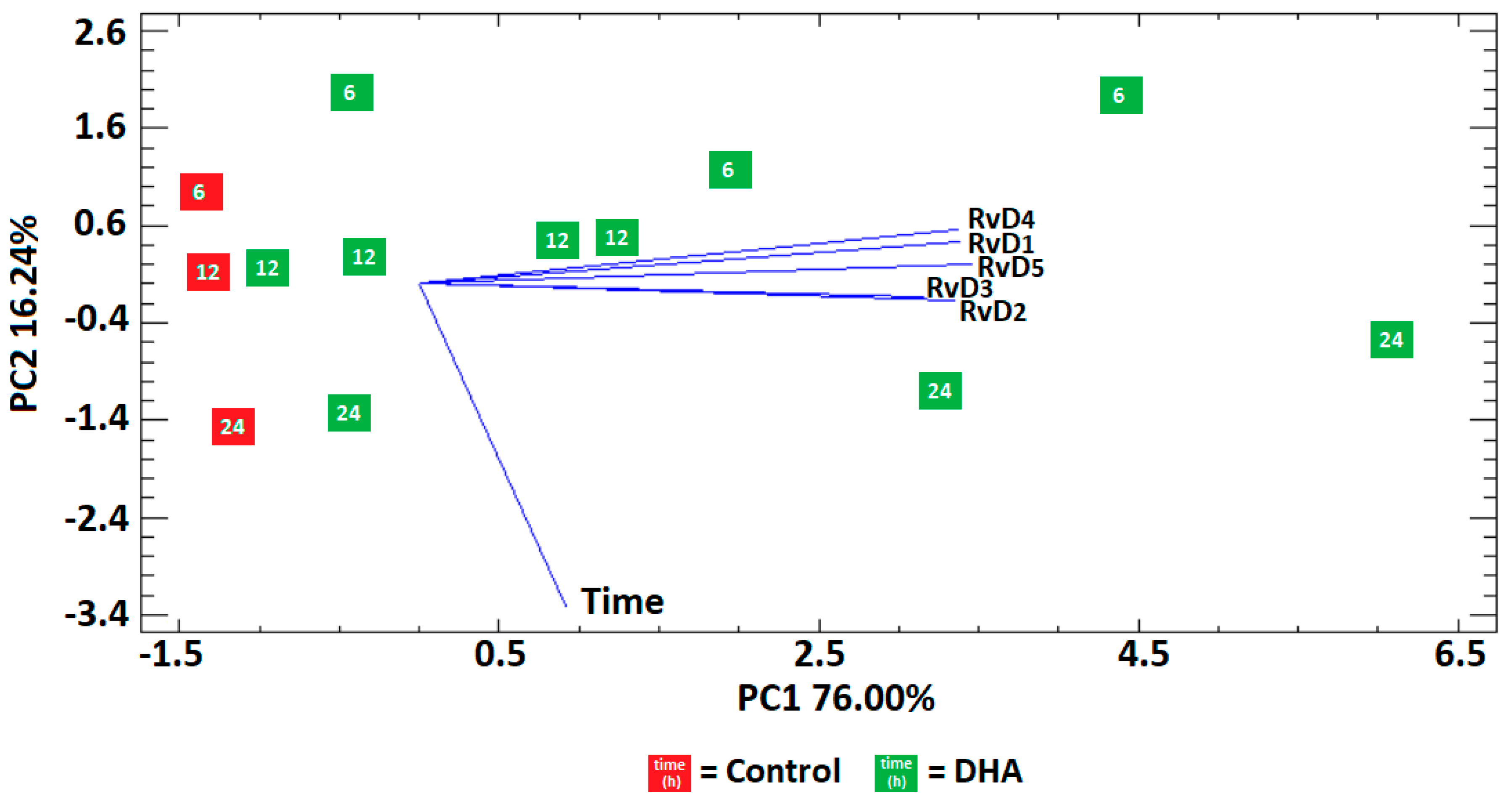
Figure 3.
Concentration/time ratio for the different resolvins in the control and DHA group. The former group shows a continuous decrease in production and the latter a plausible regulated production by a circadian clock.
Figure 3.
Concentration/time ratio for the different resolvins in the control and DHA group. The former group shows a continuous decrease in production and the latter a plausible regulated production by a circadian clock.

Table 1.
Proposed 32-factorial design to select the optimal concentrations of internal standards.
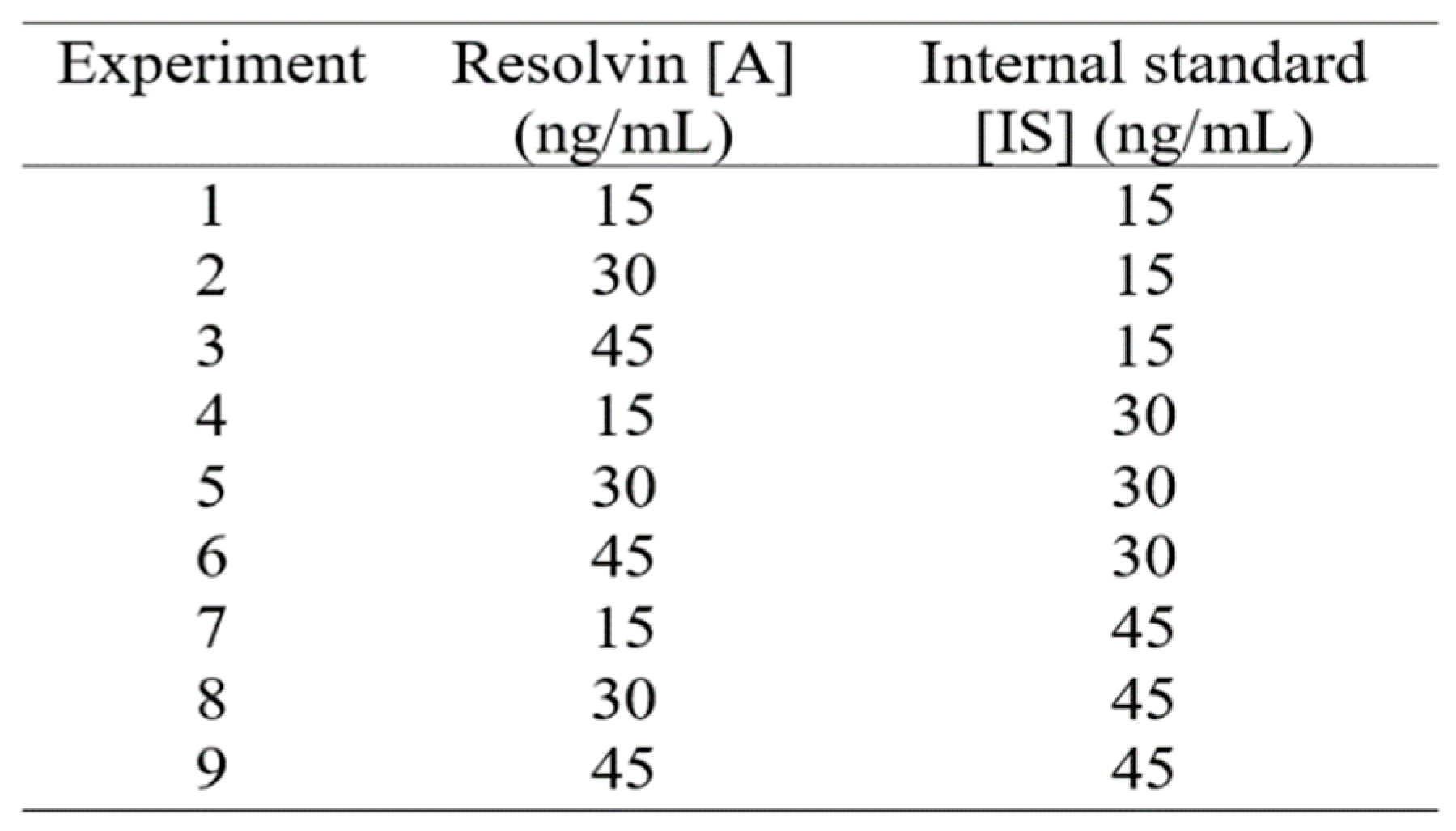
Table 2.
Calculated response factors (RF) after implementing the 32-factorial design to select the optimal amount of deuterated internal standard for the liquid-liquid extraction of resolvins from L-15 cell culture media. The RF values are expressed as average ± standard error.
Table 2.
Calculated response factors (RF) after implementing the 32-factorial design to select the optimal amount of deuterated internal standard for the liquid-liquid extraction of resolvins from L-15 cell culture media. The RF values are expressed as average ± standard error.
Table 3.
Analytical performance parameters. The linearity is judged by considering simultaneously the closeness of R2 to the unity and the comparison of Fexperimental against the tabulated Fcritical = 2.958 for 5 and 14 degrees of freedom at the 95% confidence level.
Table 3.
Analytical performance parameters. The linearity is judged by considering simultaneously the closeness of R2 to the unity and the comparison of Fexperimental against the tabulated Fcritical = 2.958 for 5 and 14 degrees of freedom at the 95% confidence level.
| Resolvin (0-50 ng/mL) |
R2 | Fexperimental | LOD | LOQ | Recovery (%) | ||
|---|---|---|---|---|---|---|---|
| RvD1 | 0.197 | -0.063 | 0.997 | 0.526 | 0.042 | 0.127 | 98.2±1.3 |
| RvD2 | 0.982 | -0.235 | 0.999 | 0.342 | 0.028 | 0.086 | 99.5±1.1 |
| RvD3 | 2.304 | -0.318 | 0.998 | 0.142 | 0.032 | 0.097 | 99.5±0.9 |
| RvD4 | 0.055 | 0.002 | 0.999 | 0.123 | 0.024 | 0.074 | 99.8±1.2 |
| RvD5 | 0.077 | -0.048 | 0.994 | 0.624 | 0.059 | 0.180 | 96.9±2.3 |
Table 4.
Temporal production of resolvins by head kidney cells from Atlantic salmon (Salmon salar)
exposed to docosahexaenoic acid.
Table 4.
Temporal production of resolvins by head kidney cells from Atlantic salmon (Salmon salar)
exposed to docosahexaenoic acid.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



